divalent cation hinder the solubilization of a tubulin kinase activity from trypanosoma cruzi...
TRANSCRIPT

367
Corresponding Author: José Bubis. Laboratorio de Química de Proteínas, Departamento de Biología Celular, División deCiencias Biológicas, Universidad Simón Bolívar, Apartado 89.000, Caracas 1081-A, Venezuela. Telephone: (58-212) 906-4219 - Fax: (58-212) 906-3064 - e-mail: [email protected]
Received: October 10, 2002. In revised form: May 12, 2003. Accepted: June 25, 2003
INTRODUCTION
Chagas’ disease is a major endemia in LatinAmerica (WHO, 1996). Trypanosoma cruzi,i ts et io logical agent, is a f lagel lateprotozoan that undergoes complexmorphological changes throughout its lifecycle in both the insect vector and thevertebrate host (De Souza, 1984; Andrewset al., 1987). The differentiation processappears to be highly regulated, and includess ign i f icant changes in b iochemicalpathways (Urbina et al., 1993), whichpermi ts i ts adaptat ion to var iousenvironments (Burleigh & Andrews, 1995).Addi t ional ly, drast ic changes in thecytoskeleton of the parasite are observedduring its differentiation.
Divalent cation hinder the solubilization of a tubulinkinase activity from Trypanosoma cruzi epimastigotes
GRACIELA UZCANGA, JOSÉ MANUEL GALÁN-CARIDAD,* KAREM NORIS SUAREZand JOSÉ BUBIS
Departamento de Biología Celular, Universidad Simón Bolívar, Caracas, Venezuela* Strang Cancer Research Laboratory, The Rockefeller University, Box 231, 1230 York Ave., NewYork, NY 10021, U.S.A.
ABSTRACT
Trypanosoma cruzi epimastigotes were extracted under various conditions in order to examine the role of divalentcations in the solubilization of microtubule proteins. When epimastigotes were homogenized in the presence of 5mM Mg+2 and 5 mM Ca+2, a protein kinase responsible for phosphorylating tubulin, as well as the tubulin thatbecame phosphorylated, remained tightly associated with the parasite particulate and detergent-resistant fractions.On the contrary, tubulin kinase and its substrate were predominantly released into the parasite cytosolic anddetergent-soluble fractions, when epimastigotes were extracted in the presence of 5 mM EDTA and 5 mM EGTA.These evidences demonstrated a divalent cation-dependent solubilization of the enzyme responsible for thephosphorylation of tubulin in T. cruzi epimastigotes and suggested a tight association between tubulin and thiskinase. Under all conditions tested, tubulin kinase activity in epimastigote extracts was lower than the addition ofthe corresponding value in the parasite cytosolic and membranous fractions, suggesting the presence of a kinaseinhibitor or regulatory subunit which also seemed to be modulated by divalent cations. Additionally, inhibitionexperiments in the presence of heparin, 2,3-bisphosphoglycerate and GTP established that the parasite tubulinkinase corresponded to a protein kinase CK2.
Key terms: microtubules, post-translational modification of tubulin, protein kinase CK2, protein phosphorylation-dephosphorylation, signal transduction, Trypanosoma cruzi, tubulin.
In most eukaryotic cells, the cytoskeletonconsists of three major filament systems:microtubules, intermediate filaments, andact in microf i laments, which forminterwoven scaffolding that permeates thecytoplasmic space of the cell. In contrast,the cytoskeleton of t rypanosomes iscomposed almost entirely of microtubulesand microtubule-associated proteins and isfurther distinguished by the conspicuousabsence of transcellular filaments (Gull,1999). A complex array of microtubulesthat includes the subpellicular network, theaxoneme, the basal body, the paraflagelarrod, the flagellum attachment zone and thef i laments responsib le for paras i teattachment conforms the trypanosome
Biol Res 36: 367-379, 2003

368 UZCANGA ET AL. Biol Res 36, 2003, 367-379
cytoskeleton (Robinson et al ., 1991;Scherével, 1994).
Similar to other eukaryotes, trypanosomemicrotubules are composed of two mainstructural subunits, α and ß tubulin, thatinteract with a number of less abundantmicrotubule-associated prote ins.Trypanosome α and ß tubulins have beenwell characterized and are similar insequence to those of other eukaryotes (Kohl& Gull, 1998). Despite these similarities,trypanosome microtubules exhibit a numberof unusual features that make them anattractive target for the development ofanti-trypanosomal drugs (Hill et al., 2000):a highly-ordered and uniformly-polarizedorganization; extensive intermicrotubuleand membrane-microtubule cross-links; andhigh stability at low temperatures.
Protein kinases CK1 and CK2, previouslyknown as casein kinases, belong to theser ine/ threonine prote in k inasessuperfamily and appear to be ubiquitous ineukaryotes. These enzymes have beenpostulated to regulate multiple pathwaysof cellular metabolism and gene expression(Guerra et al., 1999; Guerra & Issinger,1999) and seem to play an important role inin t racel lu lar s ignal ing dur ing ce l lproliferation and differentiation (Carroll etal., 1988; Pinna & Meggio, 1997; Guerra &Issinger, 1999). Protein kinases CK1 areactive monomers of 25-55 kDa (Tuazon &Traugh, 1991; Gross & Anderson, 1998).In contrast, protein kinases CK2 are α2
ß2,
α’2ß
2, or αα’ß
2 tetramers, where α and α’
(35-44 kDa) correspond to their catalyticsubunits and the ß subunits (24-30 kDa)have regulatory properties (Tuazon &Traugh, 1991; Issinger, 1993; Pinna, 1994,1997; Dobrowolska et al., 1999).
Recently, Casas et al . (2002) havedemonstrated that tubul in was thepredominant phosphoprote in inhomogenates of three d i f ferentd i f ferent ia t ion s tages of T. cruz i:ep imast igotes, t rypomast igotes andspheromastigotes. The phosphorylation oftubulin was inhibited with heparin, 2,3-bisphosphoglycerate (2,3-BPG), and GTPin a dose dependent manner, indicating thattubulin kinase corresponded to a proteinkinase CK2-like activity (Casas et al.,
2002). Interestingly, CK1-like and CK2-like activities have been previously shownin T. cruzi epimastigotes (Calabokis et al.,2002). Since divalent cations have beenrepor ted to s t rongly in f luence thedistribution of protein kinases and theirsubstrates in other systems (Ikeda et al.,1996; Nikula et al ., 1987; Clement-Chomienne & Walsh, 1996), we havestudied here the effect of Ca2+ and Mg2+ onthe solubilization of the CK2-like activityresponsible for the phosphorylation oftubulin in T. cruzi.
MATERIALS AND METHODS
Materials
Reagents and materials were purchased fromthe following sources: [γ-32P] ATP (3000Ci/mmol), New England Nuclear; P81phosphocellulose chromatography paper,Whatman; OptiPhase Hisafe II (scintillationliquid), LKB; polyvinylidene difluoride(PVDF) microporous membranes, Millipore;dephosphorylated casein, leupept in,benzamidine, phenyl methyl sulfonylfluoride (PMSF), L-trans-epoxysuccinyl-leucylamido(4-guanidino) butane (E-64),monoclonal anti-α tubulin (clone DM 1A),monoclonal anti-ß tubulin (clone TUB 2.1),heparin, 2,3-BPG, cAMP-dependent proteinkinase inhibitor PKI (6-22), 1-[N,O-bis(5-isoquinolinesulfonyl)-N-methyl-L-tyrosyl]-4-phenylpiperazine (KN-62), GTP, cAMP,bestat in, Sigma; N-(2-aminoethyl)-5-chloroisoquinoline-8-sulfonamide (CKI-7),Seikagaku America. Dr. Susan S. Taylor(University of California, San Diego,U.S.A.) generously donated synthetic Pep1(RRKDLHDDEEDEAMSITA) and Pep2(RRRADDSDDDDD) peptides. All otherchemicals were of the highest-quality gradeavailable.
Parasite culture
Trypanosoma cruzi epimastigote forms (EPstrain) were grown at 28ºC, in LIT-medium(liver infusion tryptose), as described(Rangel-Aldao et al., 1983). Epimastigotes

369UZCANGA ET AL. Biol Res 36, 2003, 367-379
at stationary phase were collected bycentrifugation at 3500x g, at 4 ºC for 15min, and washed three times with phosphatebuffered saline (PBS) solution (0.1 M, pH7.4) containing 10 mM NaCl and 42 mMsucrose. During the final wash, an aliquotof the resuspended epimastigotes was usedto count the number of parasites using aNeubauer chamber. The final cell pelletwas kept frozen at -80 ºC until further use.
Homogenizat ion of paras i tes andpreparat ion of so lub le, par t icu la te,detergent-soluble and detergent-resistantfractions
All steps were performed at 4 ºC. T. cruziepimastigotes (~ 5 x 109) were suspendedin 10 ml of various buffers containing thefollowing cocktail of protease inhibitors:50 µM PMSF, 10 µM leupeptin, 10 µM E-64 and 1 mM benzamidine.
Initially, parasite homogenates wereprepared using three different bufferconditions: 1) 50 mM Pipes (pH 6.9), 2 mMEGTA, 1 mM MgSO
4, 0.1 mM EDTA, 100
µM GTP, 5% glycerol (Buffer A); 2) 50mM Tris-HCl (pH 8.0), 1mM MgCl
2, 1 mM
CaCl2 (Buffer B); and 3) 50 mM Tris-HCl
(pH 8.0), 2 mM EDTA, 5% glycerol (BufferC). The various parasite suspensions werehomogenized on ice by sonication (35 W),using four cycles of 30 seconds each,separated by 2 min resting periods. Thehomogenates were then centrifuged at100000x g for 45 min producingsupernatants and sedimented fractions. Foreach buffer condition, the supernatant orsoluble fraction (S1) was conserved, andthe resulting pellet (P1) was resuspended,sonicated and centrifuged twice more, asdescr ibed above, obta in ing thecorresponding fractions S2, P2, S3 and P3.Then, the P3 fraction was resuspended inthe corresponding buffer containing 2%Triton X-100, and three consecut iveextractions were carried out. The resultingsupernatants and pellets were successivelynumbered. Resuspended pellet volumeswere maintained throughout the experimentin order to keep the relative protein
concentrat ion wi th respect to thecorresponding homogenate in each step.
A different set of experiments wasperformed in order to confirm that divalentions affected the solubilization of tubulinand i ts kinase. Brief ly, epimastigotehomogenates were prepared using thefollowing buffers: 1) 50 mM Tris-HCl pH(8.0), 5 mM MgCl
2, 5 mM CaCl
2 (Buffer
D), and 2) 50 mM Tris-HCl (pH 8.0), 5 mMEGTA, 5 mM EDTA (Buffer E). In eachcase, homogenates were sonicated andcentr i fuged as previously descr ibed.Following three extraction cycles withbuffer alone, three additional extractionswith the corresponding buffer containing2% Triton X-100 were also conducted.
Protein kinase assay
Protein kinase assays were performedaccording to Calabokis et al. (2002). Thereaction mixtures were carried out in afinal volume of 70 µl and contained 50mM Tris-HCl (pH 8.0), 12 mM MgCl
2, 20
mM KF, 100 µM [ γ32P] ATP (specificactivity ≈ 4500 cpm/pmol), and 50 µl ofthe parasite fraction. The reactions wereincubated for 20 min at room temperature,and an aliquot (50 µl) was spotted on P81phosphocellulose filter papers (2 cm x 2cm). The papers were washed with 50 mMphosphoric acid three times (15 min perwash), dried, and analyzed for radioactivityby liquid-scintillation counting. One unitof activity was defined as the amount ofenzyme that catalyzed the incorporation of1 pmol of inorganic phosphate (Pi) from [γ-32P] ATP per min. Since the volumes werekept constant throughout the extractionexperiments, relative enzymatic activitieswere reported as total pmoles [γ-32P] Piincorporated/min.
The remainder of the samples (20 µl)was boiled at 100oC, for 5 min, in thepresence of sample buffer for sodiumdodecy l su l fa te -po lyacry lamide ge lelectrophoresis (SDS-PAGE) (Laemmli,1970) and completely loaded on 12%polyacrylamide slab gels. The resulting[ 32P]-labeled phosphopolypeptides wereseparated by electrophoresis (Laemmli,

370
1970) and electrotransferred to PVDFmembranes (Towbin et al., 1979). Then,the membranes were exposed to Kodak X-Omat X-ray film at -80 ºC using intensifyingscreens, and the phosphorylated bands werequalitatively analyzed by autoradiography.For protein staining, the radioactively-labeled membranes were colored withCoomassie blue R-250.
Casein kinase activity was measured byincluding dephosphorylated casein (1 mg/ml) in the reaction mixtures. A similarprocedure was employed when the syntheticpeptides Pep1 (200 µM) and Pep2 (40 µM)were used as exogenous substrates, exceptthat 20 µM of the peptidase inhibitorbestatin was added to the reaction mixture.Specific casein, Pep1 and Pep2 kinaseactivities were assessed by subtracting theincorporation of [32P] ATP in the absenceof substrates. To determine the specifictubul in k inase act iv i ty , [32P] -phosphorylated tubulin was excised fromthe Coomassie b lue s ta ined-PVDFmembranes, transferred into scintillationvials, and counted.
Phosphorylation assays in the presence ofwell-known inhibitors for mammalianprotein kinases
The fractions obtained from epimastigoteshomogenized with Buffer D and E wereassayed for kinase activity in the presenceof increasing concentrations of heparin (0-133 µg/ml), 2,3-BPG (0-1.33 mM), GTP(0-133 µM), CKI-7 (0-1 µM), PKI (6-22)(0-1 µM), or KN-62 (0-2 µM). The assaywas performed with and without cAMP (50µM) when the effect of PKI (6-22) wasevaluated. Parallel experiments were alsocarr ied out us ing 1 mg/ml o fdephosphorylated casein as an exogenoussubstrate.
RESULTS
Epimastigote samples were homogenizedin the presence of various buffers (A, B, orC) in order to examine the role of divalentcat ions in the extract ion of parasite
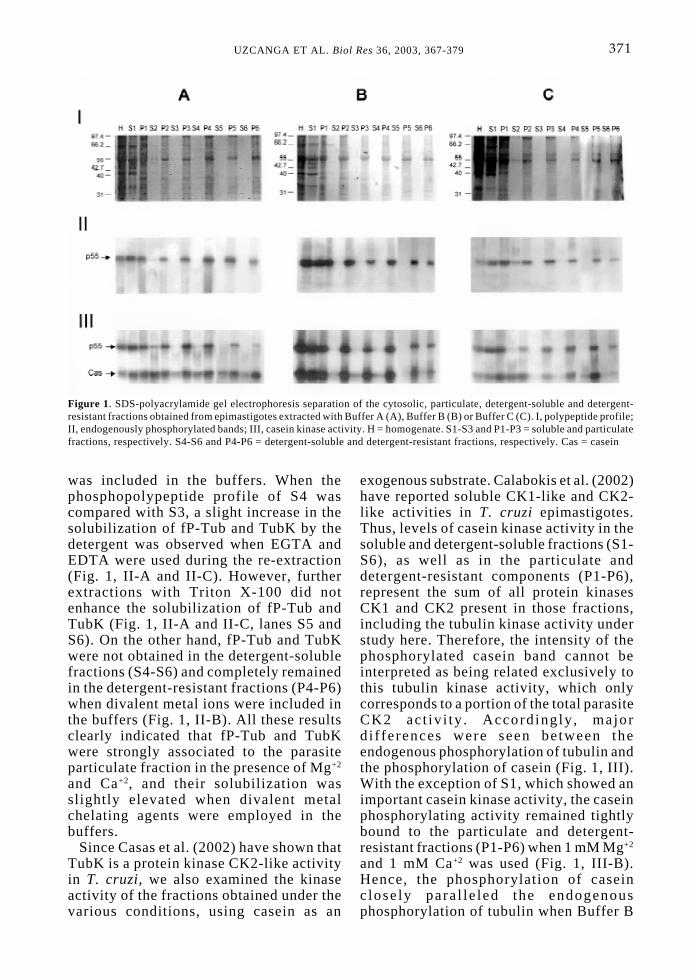
microtubule prote ins. Fol lowingcentrifugation, the supernatants (S) andpellets (P) were separated, yielding thesolubil ized and remaining particulatefractions for each condition. Re-extractionswith the various buffers containing 2%Tri ton X-100 were also carr ied out,generating their corresponding detergent-soluble and detergent-resistant components.Al l the fract ions were consecutivelynumbered and were assayed for their kinaseactivity in the absence or presence of casein.S1-S3 and P1-P3 corresponded to thefractions resulting from the extractions withthe buffers, while S4-S6 and P4–P6 weresubsequently produced by the bufferscontaining 2% Triton X-100. Figure 1summarizes the Coomassie blue stained-polypeptide profile (I), the endogenousprotein kinase (II) and casein kinase (III)activities obtained under the various buffersconditions, following separation of theresult ing fractions by SDS-PAGE. Apredominant 55-kDa-polypeptide band wasdetected by protein staining (Fig. 1, I),which was strongly phosphorylated in theparasite extracts (p55, Fig. 1, II). This bandwas recognized by anti-α and anti-ß tubulinmonoclonal antibodies following Westernblot analys is (data not shown).Additionally, anti-α and anti-ß tubulinmonoclonal antibodies immunoprecipitatedthe phosphorylated 55-kDa polypeptidefrom epimastigote homogenates (data notincluded), verifying that p55 correspondedto the parasite tubulin.
As shown in Figure 1, although thef ract ion of tubul in that becamephosphorylated (fP-Tub) and the tubulinkinase activity (TubK) seemed to remainprimar i ly associated wi th the pel letfractions, both proteins were present inS1 and S2 when divalent metal chelatingagents were employed in the buffers (Fig.1, I I -A and I I -C) . In terest ingly , nodifference was obtained when 1 mM Mg+2
was added together with 2 mM EGTA and0.1 mM EDTA (Fig. 1, II-A). In contrast,TubK and fP-Tub were only obtained inS1 when 1 mM Mg+2 and 1 mM Ca+2 wereused (Fig. 1, II-B), remaining tightlybound to the particulate fractions. Similarresults were obtained when Triton X-100
UZCANGA ET AL. Biol Res 36, 2003, 367-379
371
was included in the buffers. When thephosphopolypeptide profile of S4 wascompared with S3, a slight increase in thesolubilization of fP-Tub and TubK by thedetergent was observed when EGTA andEDTA were used during the re-extraction(Fig. 1, II-A and II-C). However, furtherextractions with Triton X-100 did notenhance the solubilization of fP-Tub andTubK (Fig. 1, II-A and II-C, lanes S5 andS6). On the other hand, fP-Tub and TubKwere not obtained in the detergent-solublefractions (S4-S6) and completely remainedin the detergent-resistant fractions (P4-P6)when divalent metal ions were included inthe buffers (Fig. 1, II-B). All these resultsclearly indicated that fP-Tub and TubKwere strongly associated to the parasiteparticulate fraction in the presence of Mg+2
and Ca+2, and their solubilization wasslightly elevated when divalent metalchelating agents were employed in thebuffers.
Since Casas et al. (2002) have shown thatTubK is a protein kinase CK2-like activityin T. cruzi, we also examined the kinaseactivity of the fractions obtained under thevarious conditions, using casein as an
exogenous substrate. Calabokis et al. (2002)have reported soluble CK1-like and CK2-like activities in T. cruzi epimastigotes.Thus, levels of casein kinase activity in thesoluble and detergent-soluble fractions (S1-S6), as well as in the particulate anddetergent-resistant components (P1-P6),represent the sum of all protein kinasesCK1 and CK2 present in those fractions,including the tubulin kinase activity understudy here. Therefore, the intensity of thephosphorylated casein band cannot beinterpreted as being related exclusively tothis tubulin kinase activity, which onlycorresponds to a portion of the total parasiteCK2 act iv i ty . Accord ing ly , majord i f ferences were seen between theendogenous phosphorylation of tubulin andthe phosphorylation of casein (Fig. 1, III).With the exception of S1, which showed animportant casein kinase activity, the caseinphosphorylating activity remained tightlybound to the particulate and detergent-resistant fractions (P1-P6) when 1 mM Mg+2
and 1 mM Ca+2 was used (Fig. 1, III-B).Hence, the phosphorylation of caseinc losely para l le led the endogenousphosphorylation of tubulin when Buffer B
Figure 1. SDS-polyacrylamide gel electrophoresis separation of the cytosolic, particulate, detergent-soluble and detergent-resistant fractions obtained from epimastigotes extracted with Buffer A (A), Buffer B (B) or Buffer C (C). I, polypeptide profile;II, endogenously phosphorylated bands; III, casein kinase activity. H = homogenate. S1-S3 and P1-P3 = soluble and particulatefractions, respectively. S4-S6 and P4-P6 = detergent-soluble and detergent-resistant fractions, respectively. Cas = casein
UZCANGA ET AL. Biol Res 36, 2003, 367-379
372
was used during the parasite extraction (Fig.1, II-B and III-B). However, some caseinkinase activity was also obtained in the restof the soluble and detergent solublefractions (S2-S6) (Fig. 1, III-B). On theother hand, some differences were observedwhen Figure 1 III-A, III-B and III-C werecompared. The phosphorylation of caseinwas higher in the solubilized fractions (S1-S6) in the presence of 2 mM EGTA, 0.1mM EDTA and 1 mM Mg+2 (Fig. 1, III-A),in contrast with the casein phosphorylationobserved in the same fractions when only 2mM EDTA was included in the buffer (Fig.1, III-C). Under this condition (Fig. 1, III-C), the casein kinase and TubK activitiesremained pr imar i ly conf ined to the
particulate and detergent-resistant fraction,following the two first extractions (S1 andS2).
An aliquot of each reaction mixture wasspotted onto P81 phosphocellulose papersto quantitatively determine the endogenousprotein kinase and casein kinase activitiesof whole-cell homogenates, supernatantsand pellets yielded by the various bufferconditions. The TubK activity was alsomeasured by excising and counting the[ 32P] -phosphory la ted tubul in bands.Initially, the kinetics of [32P] incorporationinto endogenous substrates and exogenouscasein by the various T. cruzi fractions wasexamined at room temperature and showeda linear kinase activity during the first 40
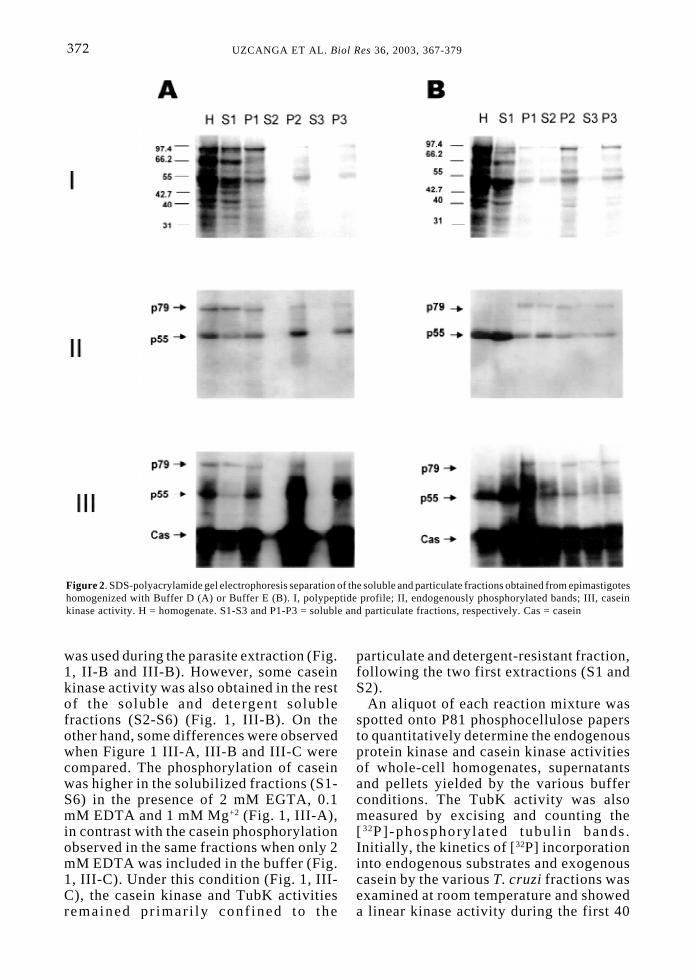
Figure 2. SDS-polyacrylamide gel electrophoresis separation of the soluble and particulate fractions obtained from epimastigoteshomogenized with Buffer D (A) or Buffer E (B). I, polypeptide profile; II, endogenously phosphorylated bands; III, caseinkinase activity. H = homogenate. S1-S3 and P1-P3 = soluble and particulate fractions, respectively. Cas = casein
UZCANGA ET AL. Biol Res 36, 2003, 367-379

373
min (data not shown). Upon longerincubation, the activity reached saturationlevels and started to plateau. Consequently,all subsequent enzymatic assays wereperformed during the linear phase of thecurve, using a total incubation time of 20min. As shown in Table I, the relative valuesdetermined were consistent and agreed withthe results qual i tat ively achieved byautoradiography in Figure 1. Interestingly,the TubK, casein kinase and endogenousprotein kinase activity values obtained inthe original extract were lower than thesum of the corresponding act iv i t iesmeasured in the S1 and P1 fractions, underall conditions. Since these assays wereperformed in a linear mode, our resultsstrongly indicated the presence of anenzyme inh ib i tor in the paras i tehomogenate.
Following our initial observations (Fig.1), two additional parasite extractioncondi t ions were also examined: onecontaining elevated amounts of divalentcations (Buffer D) and another containinghigher concentrations of EDTA and EGTA(Buffer E). The results obtained in Figure1B were corroborated when epimastigoteswere homogenized in the presence of 5 mM
Mg+2 and 5 mM Ca+2 (Fig. 2, A-II). TubKand fP-Tub remained tightly associated withthe pellet, and only a minor amount ofTubK was detected in the first solubilizedfraction (S1). In contrast, a prominentincrease in the solubilization of fP-Tub andTubK was achieved when epimastigoteswere extracted in the presence of 5 mMEDTA and 5 mM EGTA (Fig. 2, B-II, S1).Moreover, fP-Tub and TubK were obtainedin all fractions when divalent ion chelatingagents were included in the buffer (Fig. 2,B-II). When the phosphopolypeptide profileof S4 was investigated, a slight increase inthe solubilization of fP-Tub and TubK incomparison to S3 was evidenced as a resultof re-extracting P3 with Buffer E containing2% Triton X-100 (data not included). Underthese conditions, S5 and S6 also containedfP-Tub and TubK although in lesser amountthan S4 (data not shown). In contrast, fP-Tub and TubK were not obtained in thedetergent-soluble fractions (S4-S6) andremained in the detergent-resistant fractions(P4-P6) when Buffer D in the presence of2% Triton X-100 was employed during there-extractions (data not included). Thephosphorylation of exogenously-addedcasein was also evaluated for comparison
UZCANGA ET AL. Biol Res 36, 2003, 367-379
374
(Fig. 2, A-III and B-III). As previouslydescribed, levels of casein kinase activityin the different fractions may result fromthe various T. cruzi CK1 and CK2 isoformsand cannot be inferred to be solelyassociated to the parasite TubK.
A phosphopolypept ide band ofapproximately 79 kDa (p79) was alsonoticed in Figure 2, which probablycorresponded to an autophosphorylated,unrelated protein kinase or to a parasite-endogenous substrate for another kinase.Interestingly, p79 showed a differentbehavior than fP-Tub and TubK andremained primarily associated with theparasite particulate fraction under bothextraction conditions (Buffers D and E,Fig. 2). These results unequivocal lyindicated that the differential solubilizationof TubK and fP-Tub seen in the absence orpresence of divalent cat ions did notcorrespond to an experimental artifact.
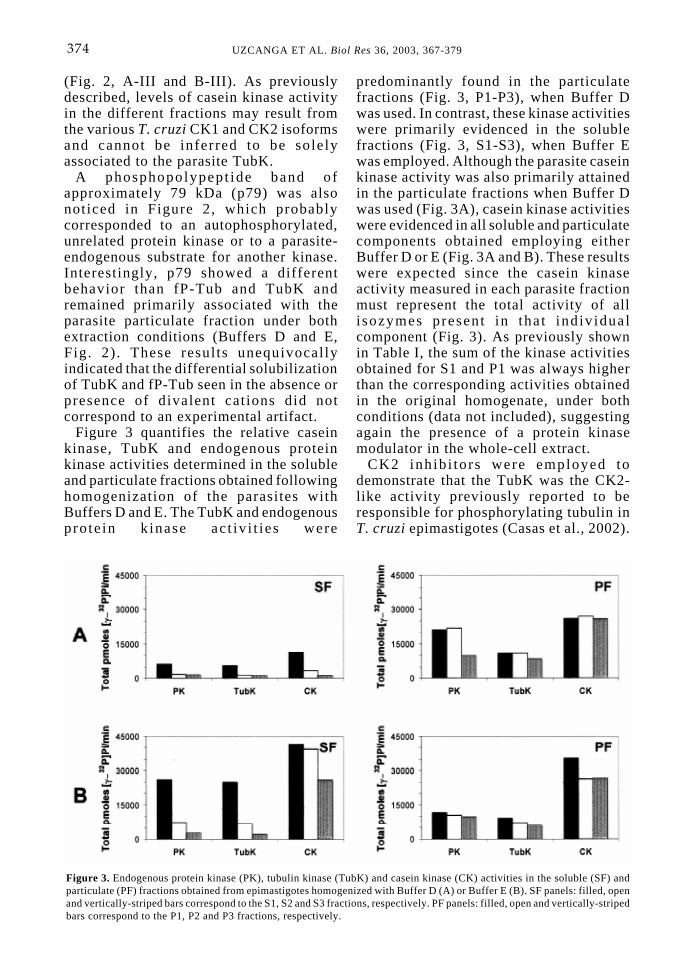
Figure 3 quantifies the relative caseinkinase, TubK and endogenous proteinkinase activities determined in the solubleand particulate fractions obtained followinghomogenization of the parasites withBuffers D and E. The TubK and endogenousprote in k inase act iv i t ies were
predominantly found in the particulatefractions (Fig. 3, P1-P3), when Buffer Dwas used. In contrast, these kinase activitieswere primarily evidenced in the solublefractions (Fig. 3, S1-S3), when Buffer Ewas employed. Although the parasite caseinkinase activity was also primarily attainedin the particulate fractions when Buffer Dwas used (Fig. 3A), casein kinase activitieswere evidenced in all soluble and particulatecomponents obtained employing eitherBuffer D or E (Fig. 3A and B). These resultswere expected since the casein kinaseactivity measured in each parasite fractionmust represent the total activity of allisozymes present in that ind iv idualcomponent (Fig. 3). As previously shownin Table I, the sum of the kinase activitiesobtained for S1 and P1 was always higherthan the corresponding activities obtainedin the original homogenate, under bothconditions (data not included), suggestingagain the presence of a protein kinasemodulator in the whole-cell extract.
CK2 inh ib i tors were employed todemonstrate that the TubK was the CK2-like activity previously reported to beresponsible for phosphorylating tubulin inT. cruzi epimastigotes (Casas et al., 2002).
Figure 3. Endogenous protein kinase (PK), tubulin kinase (TubK) and casein kinase (CK) activities in the soluble (SF) andparticulate (PF) fractions obtained from epimastigotes homogenized with Buffer D (A) or Buffer E (B). SF panels: filled, openand vertically-striped bars correspond to the S1, S2 and S3 fractions, respectively. PF panels: filled, open and vertically-stripedbars correspond to the P1, P2 and P3 fractions, respectively.
UZCANGA ET AL. Biol Res 36, 2003, 367-379

375
Since divalent cations influenced thecompartmentalization of the parasite TubKand fP-Tub, three specific CK2 inhibitors,heparin, 2,3-BPG and GTP (Tuazon &Traugh, 1991) were tested on the TubK-enriched fractions originated when theparasites were extracted with Buffer D orE, i .e. , the corresponding P1 or S1fractions, respectively. Compared to thecontrol assay, a significant decrease in thephosphorylation of tubulin was observedwhen the P1 fraction produced by theextraction with Buffer D was incubatedwith increasing concentrations of heparin(Fig. 4A, top). Similarly, a strong inhibitionin the phosphorylation of tubulin was alsoobserved in the presence of heparin, for theS1 f ract ion that was re leased byhomogenization with Buffer E (Fig. 4B,top). Additionally, 2,3-BPG (Fig. 4A andB, bottom) and GTP (data not shown)inhibited the TubK present in both fractionsin a dose-dependent manner. TubK wasalso assayed in the presence of increasingconcentrations of well-known inhibitors for
other mammalian protein kinases: KN-62for the Ca2+/calmodulin-dependent proteinkinase, CKI-7 for the protein kinase CK1,and PKI (6-22) for the cAMP-dependentprotein kinase. In particular, the effect ofPKI (6-22) was evaluated in the absenceand presence of cAMP. However, noinhibi t ion on the TubK act iv i ty wasobserved with any of these compounds (datanot shown). These results confirmed thatthe TubK under invest igat ion hererepresented a CK2-like enzyme, and arguedagainst the possible contribution of otherprotein kinases in the divalent metaldependent-redistribution of TubK and fP-Tub. When casein was added to the reactionmixtures, a concentrat ion-dependentinhibition of the phosphorylation of bothtubulin and the exogenous substrate caseinwas also evidenced in the presence ofheparin, GTP and 2,3-BPG (data notincluded).
Two synthet ic pept ides, Pep1(RRKDLHDDEEDEAMSITA), which isspecific for CK1 (Marin et al., 1994), and
Figure 4. Inhibition of the endogenous phosphorylation of tubulin by heparin and 2,3-BPG in the P1 fraction originated fromepimastigotes extracted with Buffer D (A) and in the S1 fraction obtained when the parasites were homogenized with BufferE (B). The arrow indicates the migration of phosphorylated tubulin (p55).
UZCANGA ET AL. Biol Res 36, 2003, 367-379

376
Pep2 (RRRADDSDDDDD), which isspecifically recognized by CK2 (Marin etal., 1994) were also examined as substratesfor the TubK-enriched fractions originatedwhen the parasites were extracted withBuffer D or E. The P1 fraction produced bythe extraction with Buffer D phosphorylatedPep2 but not Pep1 (Fig. 5A), indicatingthat P1 contained exclusively proteinkinases of the CK2 type. This result isconsistent with the observation that TubK,which is a CK2, was predominant lyassociated with the parasite particulatefraction under this condition. As shown inFigure 5B, the S1 fraction that was releasedby homogenization with Buffer E wascapable of phosphorylating Pep1 and Pep2.Consequently, both classes of isozymes,protein kinases CK1 and CK2, were presentin S1. However, Pep1 was a better substratefor this fraction than Pep2. Since a solubleCK1-like activity has been previouslyreported (Calabokis et al., 2002), S1probably contains this enzymatic activitybesides the TubK activity that was releasedby extraction with Buffer E.
DISCUSION
Casas et al. (2002) recently reported theco-purification of tubulin and a CK2-like
enzyme responsible for its phosphorylationfrom T. cruzi epimastigotes using DEAE-Sephacel chromatography. In an attempt tofind conditions under which tubulin can beseparated and purified from its kinase, weexamined the ef fect o f var ioushomogenizat ion buf fers on thesolubilization of both proteins. Our resultsclearly indicated a strong associationbetween tubulin and its kinase, since noseparation of tubulin kinase and the fractionof tubulin that became phosphorylated wasobtained under the various conditionstested, even when detergents were includedin the extraction buffers. Moreover, ananalogous solubi l izat ion pattern wasdemonstrated for both proteins fP-Tub andTubK. The presence of Mg2+ and Ca2+ in theextraction buffer produced the associationof a predominant amount of TubK and fP-Tub with the parasite particulate fraction.In contrast, a high proportion of fP-Tuband its kinase was solubilized when bothchelating agents EDTA and EGTA wereincluded in the homogenization step. Co-immunoprecipitation and in vitro bindingstudies have evidenced that CK2 is stronglyassociated to tubulin in mammalian cellextracts (Faust et al., 1999), which arguesin favor of our results with T. cruziepimastigotes.
Other authors have demonstrated theinf luence of divalent cat ions on thecompartmentalization of several proteinkinases and their substrates. Similar effectscaused by Ca2+ have been reported for PKCin a number of tissues. PKC was alsosolubilized in the presence of chelatingagents and remained associated with theparticulate fraction in the presence of Ca2+
(Nikula et al., 1987; Ikeda et al., 1996). InT. cruzi epimastigotes, Ogueta et al. (1994,1996) have reported a Ca+2/calmodulin(CaM)-dependent protein kinase (TcCaMK)the cellular distribution of which reliedprimarily on its phosphorylation state.Based on their results, they established amodel in which the Ca+2/CaM-dependentautophosphorylation of TcCaMK promotesits activation and release into the parasitecytosol, while the dephosphorylated formof TcCaMK is inactive and remains attachedto the parasite cytoskeletal elements
Figure 5. The P1 fraction obtained when epimastigotes werehomogenized with Buffer D (A) and the S1 fraction originatedfrom parasites extracted with Buffer E (B) were assayed forkinase activity using the synthetic peptides Pep1 and Pep2.
UZCANGA ET AL. Biol Res 36, 2003, 367-379

377
(Ogueta et al., 1996). Interestingly, adifferential solubilization of a proteinkinase CK2 and its endogenous substratetubulin, depending on the presence ofdivalent cations, have never been previouslyreported in any other tissue or organism.Here, we clearly showed that Ca2+ and Mg2+
h indered the so lub i l izat ion of th isenzymat ic act iv i ty f rom T. cruz iepimastigotes, suggesting a regulatorymechanism modulating the subcellulartranslocation of both proteins in theparasite.
Calabokis et al. (2002) have recentlyreported the identification of two caseinkinase activities in the clarified cytosolicfraction of T. cruzi epimastigotes. One ofthese, Q-I, belongs to the CK1 family,whereas the other, Q-II, appears to containeither a mixture of CK1 and CK2 or aprote in k inase wi th both act iv i t ies .Evidently, T. cruzi possesses several caseink inases act iv i t ies , and TubK mustcorrespond to one of the parasite CK2isozymes. Previous studies have describedCK2-like enzymes in other trypanosomes.Aboagye-Kwarteng et al. (1991) identifieda protein kinase CK2-like activity in T.brucei brucei and found that thephosphory la t ion of two polypept idesubstrates, p37 and p42, increased as theparasites differentiate from long, slender,rapidly dividing to short, stumpy forms.Additionally, the genes encoding the αsubunits of CK2 from both Leishmaniachagasi and T. brucei have been cloned(Bhatia et al., 1998; Park et al., 2002).Interestingly, two genes encoding for CK2catalytic subunits, CK2α and CK2 α’, wereidentified in T. brucei (Park et al., 2002).Although the alpha isoform is preferentiallynucleolar, it also localizes with the nucleusand in a lower proportion with the cytoplasm(Park et al., 2002).
The mammalian CK2 holoenzyme iscomposed of two catalytic subunits (αα,α’ α’ or αα’), which are essential for cellviability, and a dimer of two non-catalyticß subunits. In humans, Shi et al. (2001)have reported a third catalytic subunitisoform, α’’. Although both the isolatedcatalytic subunits and the holoenzyme areconst i tut ively act ive, the ß subunits
profoundly affect many properties of CK2,whose precise function is stil l poorlyunderstood (Pinna, 2002). Briefly, the CK2ß subuni t has been shown to havetrifunctional properties: it enhances thestability, the activity and the specificity ofthe CK2 catalytic subunits (Guerra &Issinger, 1999). Moreover, a dual role hasalso been reported for the CK2 ß subunit,being an activator or an inhibitor dependingon the substrate employed (Benitez et al.,2001). The phosphorylation of calmodulinby the CK2 catalytic subunits is inhibitedwhen the ß subunit is present. In contrast,the phosphorylation of casein is enhancedwhen both CK2 components are includedin the assay. Addit ionally, given thestructure of the holoenzyme, the ß subunitscould undergo reversible dissociation underphysiological conditions and play a role asanchor ing e lements and/or dock ingplat forms for protein substrates andeffectors (Pinna, 2002). Recently, Park etal (2002) reported the molecular cloning ofthe T. brucei CK2 ß subunit, but its functionhas not yet been elucidated.
By homology with T. brucei, CK2 ßsubunits may also exist in T. cruzi. Here,we evidenced the presence of a negativeregulator of the CK2-like enzyme thatphosphory la tes tubul in in T. cruz i.Interestingly, this TubK regulator alsoseemed to be modulated by divalent cations.Then, if T. cruzi possesses CK2 ß subunits,they could contribute with the modulationof the parasite CK2 catalytic subunitsresponsible for phosphorylating tubulin.However , o ther molecules p lay inginhibitory functions cannot be discarded,since it has been suggested that CK2 ßsubunits might not exist in some organisms(Boldyreff et al., 1993; Dobrowolska et al.,1991; Kikkawa et al., 1992; Ospina et al.,1992).
Various studies have indicated thatcalcium plays an important role in T. cruzihost ce l l invas ion. Pret reatment o ftrypomastigotes with chelating agentsdecreased the association of parasites tomyoblasts by approximately 50% (Morenoet al., 1994). On the contrary, pretreatmentof trypomastigotes with ionomycin, whichelevated the level of Ca2+ in the parasites,
UZCANGA ET AL. Biol Res 36, 2003, 367-379

378
significantly enhanced their infectivitycapability (Yakubu et al., 1994). In addition,T. cruzi macrophage infection was blockedby incubating the parasites with inhibitorsof protein kinases (Vieira et al., 1994). Thephysiological role that divalent cations playin the interaction between tubulin and itskinase calls for further study. However,since the parasite tubulin kinase is a proteinkinase CK2-like activity that is involved inthe modulat ion of processes such asmovement, proliferation and differentiationin other organisms, it is plausible that CK2might also play an important role inregulating these processes in T. cruzi, in acalcium-dependent manner.
ACKNOWLEDGMENTS
This research was supported by grantsfrom the Decanato de Investigación yDesarrollo, Universidad Simón Bolívar (NºDI-CB-S199146-PN) and FONACIT (NºS1-99001075). G. Uzcanga is a recipient ofa graduate research assistantship from theDecanato de Investigación y Desarrollo,Universidad Simón Bolívar.
REFERENCES
ABOAGYE-KWARTENG T, OLE-MOIYOI OK,LONSDALE-ECCLES JD (1991) Phosphorylationd i f fe rences among pro te ins o f b loodst reamdevelopmental stages of Trypanosoma brucei brucei.Biochem J 275: 7-14
ANDREWS NW, HONG K, ROBBINS ES,NUSSENZWEIG V (1987) Stage-specific surfaceantigens expressed during the morphogenesis ofvertebrate forms of Trypanosoma cruzi. Exp Parasitol64: 474-484
BENITEZ MJ, COCHET C, JIMENEZ JS (2001) A surfaceplasmon resonance study of the interactions betweenthe component subunits of protein kinase CK2 andtwo protein substrates, casein and calmodulin. MolCell Biochem 227: 31-36
BHATIA A, SANYAL R, PARAMCHUK W, GEDAMU L(1998) Isolation, characterization and disruption ofthe casein kinase II alpha subunit gene of Leishmaniachagasi. Mol Biochem Parasitol 92: 195-206
BOLDYREFF B, MEGGIO F, DOBROWOLSKA G,PINNA LA, ISSINGER OG (1993) Expression andcharacterization of a recombinant maize CK-2 alphasubunit. Biochim Biophys Acta 1173: 32-38
BURLEIGH BA, ANDREWS NW (1995) The mechanismof Trypanosoma cruzi invasion of mammalian cells.Annual Rev Microbiology 49: 175-200
CALABOKIS M, KURZ L, WILKESMAN J, GALÁN-CARIDAD JM, MÖLLER C, GONZATTI MI, BUBISJ (2002) Biochemical and enzymatic characterizationof a partially-purified casein kinase-1-like activityfrom Trypanosoma cruzi. Parasitol Int 51: 25-39
CARROLL D, SANTORO N, MARSHAK DR (1988)Regulating Cell Growth: Casein-kinase-II-dependentphosphorylation of nuclear oncoproteins. New York:Cold Spring Harbor Laboratory, pp: 91-95
CASAS B, CALABOKIS M, KURZ L, GALÁN-CARIDADJM, BUBIS J, GONZATTI MI (2002) Trypanosomacruzi: in vitro phosphorylation of tubulin by a proteinkinase CK2-like enzyme. Exp Parasitol 101: 129-137
CLEMENT-CHOMIENNE O, WALSH MP (1996)Identification of protein kinase C isoenzymes insmooth musc le : par t ia l pur i f i ca t ion andcharacterization of chicken gizzard PKCζ. BiochemCell Biol 74: 51-65
DESOUZA W (1984) Cell biology of Trypanosoma cruzi.Int Rev Cytol 86: 197-283
DOBROWOLSKA G, BOLDYREFF B, ISSINGER OG(1991) Cloning and sequencing of the casein kinase 2alpha subunit from Zea mays. Biochim Biophys Acta1129: 139-140
DOBROWOLSKA G, LOZEMAN FJ, LI D, KREBS, EG(1999) CK2, a protein kinase of the next millennium.Mol Cell Biochem 191: 3-12
FAUST M, SCHUSTER N, MONTENARH M (1999)Specific binding of protein kinase CK2 catalyticsubunits to tubulin. FEBS Letters 462: 51-56
GROSS SD, ANDERSON RA (1998) Casein kinase I:spat ia l o rgan iza t ion and pos i t ion ing o f amultifunctional protein kinase family. Cell Signal10: 699-711
GUERRA B, BOLDYREFF B, SARNO S, CESARO L,ISSINGER OG, PINNA LA (1999) CK2: a proteinkinase in need of control. Pharmacol Ther 82: 303-313
GUERRA B, ISSINGER OG (1999) Protein kinase CK2and its role in cellular proliferation, development andpathology. Electrophoresis 20: 391–408
GULL K (1999) The cytoskeleton of trypanosomatidparasite. Annual Rev Microbiol 53: 629-655
HILL KL, HUTCHINGS NR, GRANDGENETT PM,DONELSON JE (2000) T Lymphocyte-triggeringfactor of African trypanosomes is associated with theflagellar fraction of the cytoskeleton and represents anew family of proteins that are present in severaldivergent eukaryotes. J Biol Chem 275: 39369-39378
IKEDA M, ONDA T, TOMITA I, TOMITA T (1996) Thediferences in Ca+2 sensitivity of protein kinase C inplatelets from Wistar Kyoto rat and stroke-pronespontaneously hypertensive rat. Thrombosis Res82(5): 417-427
ISSINGER, OG (1993) Casein kinases: pleiotropicmediators of cellular regulation. Pharmacol Ther 59:1-30
KIKKAWA U, MANN SK, FIRTEL RA, HUNTER T(1992) Molecular cloning of casein kinase II alphasubunit from Dictyostelium discoideum and itsexpression in the life cycle. Mol Cell Biol 12: 5711-5723
KOHL L, GULL K (1998) Molecular architecture of thetrypanosome cytoskeleton. Mol Biochem Parasitol93: 1-9
UZCANGA ET AL. Biol Res 36, 2003, 367-379

379
LAEMMLI UK (1970) Cleavage of structural proteinsduring assembly of head bacteriophage T4. Nature227: 680-685
MARIN O, MEGGIO F, PINNA LA (1994) Design andsynthesis of two new peptide substrates for the specificand sensitive monitoring of casein kinases-1 and -2.Biochem Biophys Res Commun 198: 898-905
MORENO SNJ, SILVA J, VERCESI AE, DOCAMPO R(1994) Cytosolic-free calcim elevtion in Trypanosomacruzi is required for cell invasion. J Exp Med 180:1535-1540
NIKULA H, NAOR Z, PARVINEN M, HUHTANIEMI I(1987) Distribution and activation of protein kinase Cin the rat testis tissue. Mol Cell Endocrinol 49: 39-49
OGUETA SB, SOLARI A, TÉLLEZ-IÑON MT (1994)Trypanosoma cruzi epimastigote forms posses a Ca2+-calmodulin dependent protein kinase. FEBS Letters337: 293-297
OGUETA S, MACINTOSH G, TÉLLEZ-IÑON MT (1996)Regulation of Ca2+-calmodulin-dependent proteinkinase from Trypanosoma cruzi. Mol BiochemParasitol 78: 171-183
OSPINA B, NUNEZ A, FERNANDEZ-RENART M (1992)Purification of a soluble casein kinase II fromDictyostelium discoideum lacking the beta subunit:regulation during proliferation and differentiation.Mol Cell Biochem 118: 49-60
PARK JH, BREKKEN D, RANDALL A, PARSONS M(2002) Molecular cloning of Trypanosoma bruceiCK2 catalytic subunits: the α isoform is nucleolarand phosphorylates the nucleolar protein Nopp44/46.Mol Biochem Parasitol 119: 97-106
PINNA LA (1994) A historical view of protein kinaseCK2. Cell Mol Biol Res 40: 383-390
PINNA LA, MEGGIO F (1997) Protein kinase CK2 (“caseinkinase-2”) and its implication in cell division andproliferation. Progress Cell Cycle Res 3: 77-97
PINNA LA (2002) Protein kinase CK2: a challenge tocanons. J Cell Sci 115: 3873-3878
RANGEL-ALDAO R, TOVAR G, LEDESMA DE RUIZ M(1983). The cAMP receptor protein of Trypanosomacruzi. J Biol Chem 258: 6979-6983
ROBINSON D, BEATTIE P, SHERWIN T, KEITH G(1991). Microtubules, tubulin and microtubule-associated proteins of Trypanosomes. MethodsEnzymol 196: 285 –299
SCHERÉVEL J (1994) Protist: an exceptional source ofcell models. Biologie Cellulaire 80: 241-256
SHI X, POTVIN B, HUANG T, HILGARD P, SPRAY DC,SUADICANI SO, WOLKOFF AW, STANLEY P,STOCKERT RJ (2001) A novel casein kinase 2-subunit regulates membrane protein traffic in thehuman hepatoma cell line HuH-7. J Biol Chem 276:2075-2082
TOWBIN H, STAEHELIN T, GORDON J (1979)E lec t rophore t ic t rans fer o f p ro te ins f rompolyacry lamide gels to n i t rocel lu lose sheets:Procedure and some applications. Proc Natl Acad SciUSA 76: 4350-4354
TUAZON PT, TRAUGH JA (1991) Casein kinase I and II- Multipotential serine protein kinases: structure,function, and regulation. Adv Second MessengerPhosphoprotein Res 23: 123-164
URBINA JA, MACHIN I, JURADO L (1993) Thelimitations of paradigms: studies on the intermediarymetabolism of Trypanosoma cruzi. Biol Res 26: 81-88
VIEIRA MCF, DECARVALHO TU, DESOUZA W (1994)Effect of protein kinase inhibitors on the invasionprocess of macrophages by Trypanosoma cruzi.Biochem Biophys Res Commun 203: 967-971
WORLD HEALTH ORGANIZATION (1996) Chagas’Disease. Progress towards elimination of transmission.Weekly Epidemiological Record. WHO 71: 12-15
YAKUBU MA, MAJUMDER S, KIERSZENBAUM F(1994) Changes in Trypanosoma cruzi infectivity bytreatment that affect calcium ion levels. Mol BiochemParasitol 66:119-125
UZCANGA ET AL. Biol Res 36, 2003, 367-379