distribution of calanus finmarchicus in the northern north atlantic and arctic ocean—expatriation...
TRANSCRIPT

ARTICLE IN PRESS
0967-0645/$ - see
doi:10.1016/j.ds
�CorrespondiE-mail addre
Deep-Sea Research II 54 (2007) 2729–2747
www.elsevier.com/locate/dsr2
Distribution of Calanus finmarchicus in the northernNorth Atlantic and Arctic Ocean—Expatriation and
potential colonization
Hans-Jurgen Hirchea,�, Ksenia Kosobokovab
aAlfred Wegener Institute for Polar and Marine Research, Columbusstrasse 1, D-27568 Bremerhaven, GermanybP.P. Shirshov Institute of Oceanology, Russian Academy of Sciences, 36 Nakhimov ave., Moscow 117997, Russia
Received in revised form 1 July 2007; accepted 10 August 2007
Available online 23 October 2007
Abstract
The distribution of Calanus finmarchicus was studied on a transect across the central Greenland Sea, and on five
transects from the Eurasian shelves across the Atlantic Inflow in the Arctic Ocean. Stage composition was used as an
indicator for successful growth; gonad maturity and egg production were taken as indicators for reproductive activity. On
the Arctic Ocean transects, these parameters were measured simultaneously from the sibling species Calanus glacialis.
Response of egg production rate to different temperatures at optimal food conditions was very similar between both
species in the laboratory. C. finmarchicus was present at all stations studied, but young developmental stages were only
present close to the regions of submergence of Atlantic water under the Polar water. This together with a decreasing
abundance and biomass from west to east along the Atlantic Inflow in the Arctic Ocean and reproductive failure indicates
that C. finmarchicus is expatriated in the Arctic Ocean. We hypothesize that the late availability of food in the Arctic
Ocean, rather than low temperature per se, limits reproductive success. Better reproductive success in the very low
temperature regions of the Return Atlantic Current and the marginal ice zone in the Greenland Sea supports this
hypothesis. The possibility for a replacement of C. glacialis by C. finmarchicus and consequences for the ecosystem after
increasing warming of the Arctic are discussed.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Calanus finmarchicus; Calanus glacialis; Expatriation; Climate change; Arctic Ocean
1. Introduction
Calanus finmarchicus is a key species in thezooplankton of the North Atlantic, where it is preyfor many species of fish. Due to its biomass, its rolein carbon flux is also important. C. finmarchicus isfound all over the North Atlantic (Jaschnov, 1970;
front matter r 2007 Elsevier Ltd. All rights reserved
r2.2007.08.006
ng author. Fax: +49471 4831 1918.
ss: [email protected] (H.-J. Hirche).
Conover, 1988). Its centre of activity is confined toice-free water, while its congeners, Calanus hyper-
boreus and Calanus glacialis, inhabit the seasonalice-covered seas and the Arctic Ocean (Conover,1988). Due to the circulation system in the NorthAtlantic, high numbers of C. finmarchicus aretransported by the North Atlantic Current (NAC)into subarctic and arctic seas. The main northwardtransport of Atlantic water is through the NAC andits northern continuation, the West Spitsbergen
.

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472730
Current (WSC), which, after its passage through theGreenland Sea, finally drains into the Arctic Oceanin Fram Strait. In the Greenland Sea, part of theAtlantic water recirculates and flows southwards asthe Return Atlantic Current (RAC) along the EastGreenland Shelf (EGS) underneath the Polar watersof the East Greenland Current (EGC) (Gascardet al., 1988). Via troughs, Atlantic water alsoreaches far onto the EGS (Budeus and Schneider,1995). The large frontal systems of the Arctic Front(AF) in the east and the East Greenland Polar Front(EGPF) in the west are interfaces between Atlantic,Arctic and Polar water masses and allow exchangeof faunistic elements. Atlantic water is advectedonto the Barents Shelf from the Lofoten Basinthrough the Barents Sea Opening into the BarentsSea (Loeng, 1991). Portions of Atlantic watersubmerge under the lighter Arctic water at thePolar Front and flow north following troughs onthe shelf (Schauer et al., 2002). From the north,branches of Atlantic water enter the Barents Seabetween Nordaustlandet and Franz-Josef-Land as anear-bottom water mass (Pfirman et al., 1994).
In the Arctic Ocean, water of Atlantic originforms a layer several 100m thick between 200 and1000m depth. Atlantic water enters mainly via theFram Strait (Rudels et al., 1994) and forms aboundary current running counter-clockwise alongthe perimeter of the Arctic Ocean. Recirculatingbranches of Atlantic water are deflected seawardwhere mid-ocean ridges, like the Nansen-GakkelRidge, the Lomonosov Ridge (Anderson et al.,1989), and the Alpha-Mendeleev Ridge, meet theEurasian Shelf (Rudels et al., 1994).
While in many of these regions C. finmarchicus
constitutes a large part of the biomass (Hirche andMumm, 1992; Kosobokova and Hirche, 2000), wesuggest that it is not actively growing and reprodu-cing there, but is expatriated. The low temperaturesin Polar water have been suggested as the cause forlow growth and reproduction of C. finmarchicus inthe Arctic Ocean (Jaschnov, 1970). Corkett et al.(1986) observed embryonic development at 0 1C,development to CI at 2 1C, and older stages at 5 1C,but do not comment why they did not conduct allmeasurements at 0 1C. Campbell et al. (2001) for C.
finmarchicus and Thompson (1982) for Calanus spp.studied the development at 4, 8, and 12 1C andfound successful growth at all temperatures. Tandeet al. (1985) and Hansen et al. (1996) assumed fromfield observations in the Barents Sea that tempera-ture hindered gonad development of CV and ovary
maturation in adult females. On the other hand,females collected in the EGC in June at �1.6 1Ccontinued to spawn at 0 1C for 22 days (end ofexperiment), and females collected in the WSC inApril spawned continuously for 77 days at 0 1C(Hirche, 1990).
Recent climate models forecast large environ-mental changes, especially in the subarctic andarctic regions (Polyakov et al., 2002). These changesinclude increase of water temperatures, thinning ofsea ice, and reduction of the duration of icecoverage (Johannessen et al., 2002). Increasedinflow of Atlantic water into the Arctic Ocean hasbeen observed, which caused a shift in the balancebetween Pacific and Atlantic waters there (Carmacket al., 1995; McLaughlin et al., 1996, 2002; Swiftet al., 1997). Although we do not know yet whichphysiological or other barriers prevent C. finmarch-
icus from inhabiting these regions, it is possible thatthe environmental changes associated with theclimate changes will allow future colonizationand replacement of the autochthonous congenersC. glacialis and C. hyperboreus. This would drama-tically affect the ecosystems, as C. finmarchicus issmaller and has a different life cycle strategy thanthe other species.
Here we describe the distribution of C. finmarch-
icus on transects across the central Greenland Seaand the Atlantic Inflow in the Arctic Oceancombining observations from several expeditions.By comparing total abundance with stage composi-tion (as an indicator for active growth), and withgonad stage and egg production rate (as indicatorsof reproductive activity) on transects across differ-ent water masses, we try to distinguish expatriatedpopulations from actively growing and reproducingones. As reduced growth and reproduction ofC. finmarchicus could instead be caused by foodlimitation, we used the sibling species C. glacialis asan indicator for feeding conditions, assuming thatfood requirements and preferences are similar.Finally, we compared egg production rates of thetwo species at different temperatures in the labora-tory and in the field.
2. Material and methods
2.1. Zooplankton collection
Calanus spp. were collected during severalexpeditions of the RV ‘Polarstern’ to the GreenlandSea and the Arctic Ocean. Cruise dates and station

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2731
locations are listed in Table 1, transects and stationpositions are shown in Fig. 1. On transect GS(Fig. 1) zooplankton was collected with verticaltows by a bongo net (310 mm mesh) in the upper80m. On transect BS it was collected with a bongonet (310 mm mesh) in the upper 100m and with aMulti-net (Hydrobios, Kiel, 0.25m2 mouth opening,150 mm mesh) in five depth strata down to thebottom (Table 1). At transects W, E, F five stratadown to 1500m or to the bottom were sampled, attransects B and H from five to nine depth layerswere sampled down to the bottom with the samemodel of Multi-net. Samples were preserved in 4%borax–buffered formaldehyde. All copepoditestages of the two species were counted in thesamples.
Table 1
Sampling locations, bottom depth and maximum sampling depth (m)
Cruise Transect,
station no.
Latitude, N Longitude Date
ARK VI/3 Transect GS
66 74130 14130W 11 Jun
67 74135 14100 11 Jun
68 74142 13130 11 Jun
69 74145 131 11 Jun
70 74145 12120 11 Jun
71 741145 11140 12 Jun
72 74145 11100 12 Jun
73 74145 10125 12 Jun
74 74145 9145 12 Jun
76 74145 8130 13 Jun
78 74145 7117 13 Jun
80 74145 6100 13 Jun
82 74145 4144 13 Jun
84 74145 3138 14 Jun
86 74145 2113W 14 Jun
88 74145 0157W 14 Jun
91 74145 1102E 15 Jun
93 74145 2113 15 Jun
95 74145 3128 15 Jun
97 74145 4146 16 Jun
99 74145 6103 16 Jun
100 74145 6140 16 Jun
101 74145 7115 16 Jun
102 74145 7155 16 Jun
103 74145 8135 17 Jun
104 74145 9112 17 Jun
105 74145 9150 17 Jun
106 74145 10129 17 Jun
107 74145 11105 17 Jun
108 74145 11145 17 Jun
110 74145 13100 18 Jun
112 74145 14115 18 Jun
114 74145 15132E 18 Jun
2.2. Taxonomy and prosome length
For differentiation between C. finmarchicus andC. glacialis, prosome length established by Hircheet al. (1994) in the Greenland Sea was used (Table 2).
The prosome length of at least 60 individuals ofeach stage was measured with a micrometer at 25�magnification to the nearest 0.04mm between thetip of the cephalosome and the end of the lastthoracic segment. As prosome length, among otherfactors, is controlled by the temperature duringdevelopment, different body sizes are found for thesame species in different regions. As prosomelengths often overlap between species, this criterionis not absolutely reliable. Especially when onespecies is represented by a very small fraction, the
Bottom
depth, (m)
Maximum depth
of haul (m)
Egg production
experiments
e 89 250 80 Pooled
e 89 400 80 Pooled
e 89 518 80 Pooled
e 89 770 80 Pooled
e 89 2150 80 Pooled
e 89 2700 80 Pooled
e 89 3050 80 Pooled
e 89 3190 80 Pooled
e 89 3270 80 Pooled
e 89 3345 80 Pooled
e 89 3450 80 Pooled
e 89 3524 80 Pooled
e 89 3600 80 Pooled
e 89 3655 80 Pooled
e 89 3680 80 Pooled
e 89 3400 80 Pooled
e 89 3770 80 Pooled
e 89 3771 80 Pooled
e 89 3314 80 Pooled
e 89 3450 80 Pooled
e 89 2980 80 Pooled
e 89 2250 80 Pooled
e 89 2130 80 Pooled
e 89 2430 80 Pooled
e 89 3250 80 Pooled
e 89 2580 80 Pooled
e 89 2585 80 Pooled
e 89 2515 80 Pooled
e 89 2490 80 Pooled
e 89 2448 80 Pooled
e 89 2250 80 Pooled
e 89 1950 80 Pooled
e 89 820 80 Pooled

ARTICLE IN PRESS
Table 1 (continued )
Cruise Transect,
station no.
Latitude, N Longitude Date Bottom
depth, (m)
Maximum depth
of haul (m)
Egg production
experiments
ARK IX/4 Transect W
6 811120 301360E 12 Aug 93 188 160 Single and pooled
7 811280 301530 13 Aug 93 525 490 Single and pooled
14 811400 301160 13 Aug 93 2701 1500 Single and pooled
19 821120 341300 15 Aug 93 2456 1500 Single and pooled
ARK IX/4 Transect F
32 781430 1321210 2 Sep 93 2987 1500 Single and pooled
35 781230 1331040 4 Sep 93 2062 1500 Single and pooled
38 781100 1331230 5 Sep 93 982 950 Single and pooled
39 781060 1331340 5 Sep 93 446 450 Single and pooled
ARK IX/4 Transect E
54 791110 1191540 13 Sep 93 3067 1500 Single and pooled
56 781400 1181430 14 Sep 93 2615 1500 Single and pooled
58 781000 1181330 15 Sep 93 1930 1500 Single and pooled
60 771330 1181260 16 Sep 93 1178 1000 Single and pooled
62 771240 1181110 17 Sep 93 556 500 Single and pooled
ARK XI/1 Transect B
25 811060 1051230E 7 Aug 95 2642 2500 Single and pooled
27 811140 1061450 8 Aug 95 3133 3000 Single and pooled
31 801460 1031230 11 Aug 95 1435 1400 Single and pooled
32 801390 1031030 11 Aug 95 621 500 Single and pooled
33 801250 1011590 12 Aug 95 266 245 Single and pooled
ARK XI/1 Transect H
75 801550 1221400 4 Sep 95 3566 3500 Single and pooled
47 801550 1321000 20 Aug 95 3907 3500 Single and pooled
49 811030 1361320 22 Aug 95 2708 2600 Single and pooled
51a 811070 1381470 23 Aug 95 1830 1700 Single and pooled
52 811100 1401060 24 Aug 95 1292 1200 Single and pooled
55 811110 1431240 25 Aug 95 1693 1600 Single and pooled
57 811120 1501150 27 Aug 95 2643 2500 Single and pooled
MN ¼ multi-net, Bo ¼ bongo net. Egg production experiments: single ¼ single females in 250mL, pooled ¼ 20–30 females in 3L beakers.
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472732
overlapping tails of the length frequency distribu-tion lead to a substantial over-estimation of thesmaller fraction (Unstad and Tande, 1991).
2.3. Dry mass
Dry mass was calculated using dry mass sizecoefficients of copepodids and adults of C. finmar-
chicus and C. glacialis from Hirche and Kosobokova(2003, see their Table 2).
2.4. Gonad maturation stages
Gonad maturation of preserved females used inegg production experiments was examined under astereo microscope. Females were stained with boraxcarmine (2%) in ethanol (70%). The criteria given inSmith (1990), Niehoff and Hirche (1996) and
Kosobokova (1999) were adapted to distinguishthe following five states of gonad maturation:
I.
Immature: ovary compact, no oocytes in diver-ticulae and oviducts.II.
Semi-mature: one or several rows of smallimmature oocytes in diverticulae and oviducts.III.
Mature: several rows of immature oocytesdensely packed into diverticulae; pouches ofoocytes in oviducts; ventral row of oocytes indiverticulae and oviducts formed by largemature oocytes.IV.
Semi-spent: few small oocytes in the ovary,single oocytes spread irregularly in diverticulaeand oviducts.V.
Spent: diverticulae and oviducts as thin bands;no oocytes in the ovary; posterior of ovaryextends to the third thoracic segment.
ARTICLE IN PRESS
Fig. 1. Circulation of Atlantic water in the northern North Atlantic with location of stations (first and last station number indicated) and
transects used in this study. White arrow ¼ AW on the surface; black arrow ¼ submerged flow of AW. Hatched area in inserted
map ¼ marginal ice zone (MIZ). RAC ¼ Return Atlantic Current, WSC ¼West Spitsbergen Current.
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2733
2.5. Egg production experiments
Females for in-situ egg production experimentswere collected with bongo hauls (0.6m diameter,310 mm mesh) in the upper 80 or 100m. Femaleswere sorted immediately after collection. Singlefemales were incubated in plexiglass cylinders
(150mL) having mesh (330 mm) false bottoms toseparate eggs from females. Cylinders were thensuspended in 250-mL beakers with surface pre-screened (100 mm) seawater at ambient temperature(mean of water column sampled). On other cruises,between 20 and 30 females were pooled andincubated in 3-L beakers. Incubation methods used

ARTICLE IN PRESS
Table 2
Prosome lengths (mm) from the literature and values used to distinguish copepodids and adult female Calanus finmarchicus and C. glacialis
in this study
C. finmarchicus C. glacialis
Stage Tande et al.
(1985)
Unstad and
Tande (1991)
Hirche et al.
(1994)
Kosteyn and
Kwasniewski (1992)
Hirche et al.
(1994)
CI o0.85 o0.82 0.84–1.04 40.82
CII o1.2 o1.24 1.20–1.45 41.24
CIII o1.65 o1.6 1.70–2.05 41.6
CIV o2.3 o2.2 2.35–2.80 42.2
CV 2.5–2.9 o3.0 o3.0 3.05–3.95 43.0
AF 2.7–3.1 o3.2 o3.2 3.50–4.40 43.2
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472734
during the different cruises are included in Table 1.For long-term incubations to study the effect oftemperature on egg production, 8–16 females werefed cultures of Thalassiosira antarctica grown on f/4medium at concentrations 4400 mmCL�1 for10–16 days at 0 1C. Thereafter, single females weretransferred to containers as described above andexposed to temperatures between �1.8 and 8 1C fora period of 15 days. Egg production at eachtemperature was calculated as the mean of all daysat a given temperature except the first 2 days afterstart of the experiment. In C. finmarchicus this delayis required to adjust egg production to the newtemperature (Hirche et al., 1997). Light wasprovided continuously by a daylight fluorescentbulb at 4 mEm�2. After 24 h, cylinders weretransferred to new containers with fresh food, andeggs were counted.
2.6. Chlorophyll
Samples for the determination of chlorophyll-awere taken from a Rosette sampling system in stepsof 10m from the surface to 50m. Concentrations ofchlorophyll-a and phaeopigments were determinedon a Turner Designs fluorometer after grinding andextracting (90% acetone) filters (GF/C) from watercasts. For calibration, pure chlorophyll-a in 90%acetone (Sigma) was used.
2.7. Statistics
C. finmarchicus egg production rates, watertemperature and chlorophyll-a concentration weretested for correlation using non-parametric Spear-man’s correlation analysis. Then, multiple regressionanalysis was conducted using C. finmarchicus eggproduction rates as dependent variable. Variables
were log or log(1+x) transformed where necessaryto obtain normality and to homogenize variances(Zar, 1996). More than one independent variablewere only included in the model if they led to a morethan 20% increase in the adjusted R2. The adjustedR2 is a modification of R2 that adjusts for thenumber of explanatory terms in the model. UnlikeR2, the adjusted R2 increases only if the newindependent variable improves the model morethan would be expected by chance (Zar, 1996).Statistical analyses were performed at a ¼ 0.05 withSTATISTICA 6.1 (StatSoft Inc.).
3. Results
3.1. Central Greenland Sea—Transect GS
Our transect crossed the central Greenland Seafrom the EGS to the foot of the Barents Shelf nearBear Island, and thus included the three hydro-graphic domains of the Greenland Sea: the Polardomain on the EGS, the Arctic domain of theGreenland Sea Gyre (GSG), and the Atlanticdomain of the WSC. Topography and distributionof the potential temperature across our transect isshown in Budeus et al. (1993), the general hydro-graphy of the Greenland Sea has been described indetail by Meincke et al. (1992) and Budeus et al.(1993). In the ice covered Polar Domain (stations66–69) temperatures were close to the freezing pointdown to about 40m depth. The Polar waters wereconfined to relatively shallow depths as they wereunderlain by Atlantic influenced waters from 150mdownwards. Therefore the EGPF at stations 69/70did not reach to the bottom. The RAC flowingsouthward under Polar water at stations 69–73 hadmaximum temperatures of up to 2 1C (station 69) inits core at 150m. In the central basin, temperatures

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2735
in the upper 80m ranged from �0.75 to 0.5 1C. Atthe AF (stations 101/102), which separates theArctic from the Atlantic domain, the temperatureas well as the salinity front extended almostvertically to the surface in the upper 200m betweenstations 99 and 102. Between 200 and 300m depth,a warm and saline westward intrusion indicatedsome cross frontal exchange. Temperatures in theupper 80m of the WSC were between 4 and 6.5 1C.
Transect GS shows high concentrations of C. finmar-
chicus in the WSC east of the AF (Fig. 2). The elevatedconcentrations in the west at stations 69–72 (EGPF)are clearly associated with the RAC. Fig. 2 showsalso, that C. finmarchicus is present in the centralGSG and on the EGS, however, at much lowerconcentrations. Only the stages CIV to CVI weredistinguished, but their ratios varied between thedifferent water masses: 11.576.7% (CIV), 53.979.0% (CV) and 34.679.3% (females) in the WSC;35.578.4%, 48.679.3%, and 15.977.0% in theGSG; and 29.471.6%, 53.775.5%, and 16.873.9% on the EGS, respectively.
Egg production was measured at 17 stations withC. finmarchicus and two stations with both C.
finmarchicus and C. glacialis (Fig. 3A). In the WSC,mean egg production was 4.3 eggs female�1 d�1,
66
67
68
69
70
71
72
73
74
76
78
80
82
84
86
0
100
200
300
400
500
600
AF
CV
CIV
RAC
GSGEGC
Ab
un
dan
ce (
n m
-3)
S
Fig. 2. Distribution of copepodids CIV, CV and adult females of Ca
EGC ¼ East Greenland Current, RAC ¼ Return Atlantic Current, GS
ranging from 0.1 to 13.3. In the frontal area of theAF (stations 99–101), at temperatures from 0.5 to1.5 1C, mean egg production rate was 4.8 (range4.6–5.0). In the GSG there were not sufficientfemales to set up experiments. At stations 69–71,located on top of the RAC, C. finmarchicus
produced between 0.2 and 2.6 eggs female�1 d�1,respectively. On the EGS no eggs were produced byC. finmarchicus, but C. glacialis laid 2 and 1.5 eggsfemale�1 d�1 at stations 67 and 68, respectively.
3.2. Marginal ice zone (MIZ) of the Greenland
Sea—MIZ
The hydrography of the MIZ in the Fram Straitduring our study has been described in detail byJohannessen et al. (1987), Manley (1987) andGascard et al. (1988); the zooplankton dynamicsby Smith et al. (1985), Smith (1987), and Hirche(2004). The position of the MIZ in the GreenlandSea is mainly controlled by the position of theEGPF (Bourke et al., 1988) and therefore relativelyinvariant in space in contrast to the MIZ in otherregions, where a receding ice edge in summer isfollowed by ice-edge blooms. The position of the iceedge in 1984 changed rapidly during the cruise due
88
91
93
95
97
99
100
101
102
103
104
105
106
107
108
110
112
114
WSC
tation
lanus finmarchicus across the central Greenland Sea (741450N).
G ¼ Greenland Sea Gyre, WSC ¼West Spitsbergen Current.

ARTICLE IN PRESS
9876543210-1-20
20
40
60
80
Chl a
Ch
loro
ph
yll a
(m
g m
-2)
Eg
g p
rod
ucti
on
rate
(eg
gs f
em
ale
-1 d
-1)
0
20
40
60
80
100
C.finmarchicus
C.glacialis
C.finmarchicus
C.glacialis
Greenland SeaBarents Sea
C.finmarchicus
Eg
g p
rod
ucti
on
rate
(eg
gs f
em
ale
-1 d
-1)
Temperature (°C)
9876543210-1-2
(A)
(B)
Fig. 3. Egg production (eggs female�1 d�1) of Calanus finmarchicus in relation to temperature in the upper 80m (A) on Transect GS across
the central Greenland Sea and on Transect BS in the Barents Sea; (B) in the Marginal Ice Zone of the Greenland Sea (MIZ in Fig. 1). Solid
line ¼ egg production rate of C. finmarchicus at optimum food conditions (see Fig. 8).
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472736
to changes in winds and currents. In addition,various eddies shaped its appearance by their owndrift and by advecting ice into the open water.
A study by Smith et al. (1985) in the MIZ of theGreenland Sea found C. finmarchicus present in thethree neighbouring water masses. It dominatedwaters of Atlantic origin and was represented byall copepodite stages, while in Arctic and Polarwaters mostly CV and females were found, similarto the western part of Transect GSG. During thesame study, egg production was measured at 71
stations in the MIZ (Fig. 1) and plotted against themean seawater temperature in the upper 80m,where females were collected (Fig. 3B). Femaleswere laying eggs at temperatures down to o�1 1C.A comparison with the experimental egg productionrates at maximum food concentrations from Hircheet al. (1997) shows that C. finmarchicus females werealways food limited (Fig. 2C). C. finmarchicus eggproduction rates, seawater temperature and chlor-ophyll-a concentration were all positively correlatedwith each other. multiple linear regression analysis

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2737
showed that the best prediction of egg productionrates was obtained with a model using log (x+1)egg production rates as dependent variable and logchlorophyll-a concentration and water temperatureas independent variables. Adjusted R2 using bothvariables was 0.48, clearly better than the adjustedR2 of 0.38 and 0.33 obtained for simple linearregression models using only water temperature andlog chlorophyll-a as independent variables, respec-tively. Regression slopes were significantly differentfrom zero in both simple linear regression modelsand for the multiple linear regression model(po0.0001, unadjusted overall R2
¼ 0.50, n ¼ 46).
3.3. Barents Sea—Transect BS
Two meridional transects along 331E across thePolar Front of the northern Barents Sea (Fig. 1) inMay and June 1997 have been published before andare here summarized for completeness (Hircheand Kosobokova, 2003). All copepodite stages ofC. finmarchicus were found south of the front inAtlantic water, but stages CI and CII were rare inthe Polar domain north of the front. Egg produc-tion rates, the portion of spawning females, andyoung copepodids were much higher in C. glacialis
than in C. finmarchicus in the Polar domain close tothe front, while farther to the north, insufficientfemale C. finmarchicus were found to set upexperiments. It is worthwhile mentioning, however,that C. finmarchicus had relatively high egg produc-tion rates at temperatures around �1 1C, whencompared with individuals obtained on the Green-land Sea Transect (Fig. 3A). Results of incubationexperiments were supported by a tremendousabundance of C. glacialis eggs in the water column.In contrast, no eggs of C. finmarchicus were foundin the Polar domain 430 nm north of the front.
3.4. Arctic Ocean—Transect W
Hydrography for Transect W covering fourstations from the Barents Shelf to the WesternNansen Basin in 1993 was described by Schaueret al. (1997, their Transect I). Under a layer of cold,low-salinity Arctic Surface water of ca. 100mthickness, a relatively cold, low salinity layer ofNorthern Barents Sea Water was situated on theslope between stations 6 and 7, and was replaced bywarm (43 1C at 200m) Atlantic water 30–40 kmseaward of the shelf break (stations 7 and 14).
Station 19 was to the north of the Atlantic Inflowand dominated by Arctic water.
A large percentages of young stages of bothC. finmarchicus and C. glacialis were found at thesouthern end of the transect (especially at station 7on the shelf break), while older stages dominatedthe deep stations (Fig. 4). Pronounced maxima ofboth abundance and biomass of C. finmarchicus
were centred on the shelf break (sta. 7, Fig. 5). Byfar the majority were in the upper 100m, but at thedeep stations 14 and 16 a few specimens were alsofound in the deepest samples (Fig. 6). On a transectin 1987, which started at a similar location butextended at the Nansen-Gakkel Ridge, Hirche andMumm (1992) found C. finmarchicus dominated themesozooplankton biomass in the Atlantic Inflow,but noted a drastic drop in abundance and almostcomplete disappearance of younger stages asso-ciated with an abrupt change in water masses at ca.831N (Anderson et al., 1989).
C. finmarchicus laid few eggs (range 0.41–3.25) atthree of four stations, while C. glacialis spawnedbetween 16 and 22 eggs female�1 d�1 at the twostations where experiments were set up (Fig. 7).Gonad maturation data were consistent with theseresults. Between 14% and 26% of the C. finmarch-
icus females were mature, whereas in C. glacialis,
except for station 7, more than half of the femaleswere mature (Fig. 7). In the study by Hirche andMumm (1992, their Tables 2 and 3) C. finmar-
chicus produced eggs at only one station in theAtlantic Inflow water (0.3 eggs female�1 d�1), whileC. glacialis spawned at four stations. After feedingfemales of the two species with superabundant foodfor 11 days, C. glacialis laid slightly more eggs. Atthe one station where such experiments were alsoconducted with C. finmarchicus, egg production alsoincreased from 0.3 to 1 eggs female�1 d�1.
3.5. Arctic Ocean—Transect B
Hydrography for Transect B covering five sta-tions from the Laptev Sea Shelf to the easternNansen Basin in 1995 was described by Rudels et al.(2000, their Transect B). A cold, low-salinity watercolumn over the shelf and slope reaching down tomore than 1000m, the Barents Sea branch of theAtlantic Inflow, was separated by a 40-km widefrontal zone from a warm (almost 3 1C), high-salinity core located at 300m depth in the basin atthe northern side of the transect, the Fram Straitbranch.
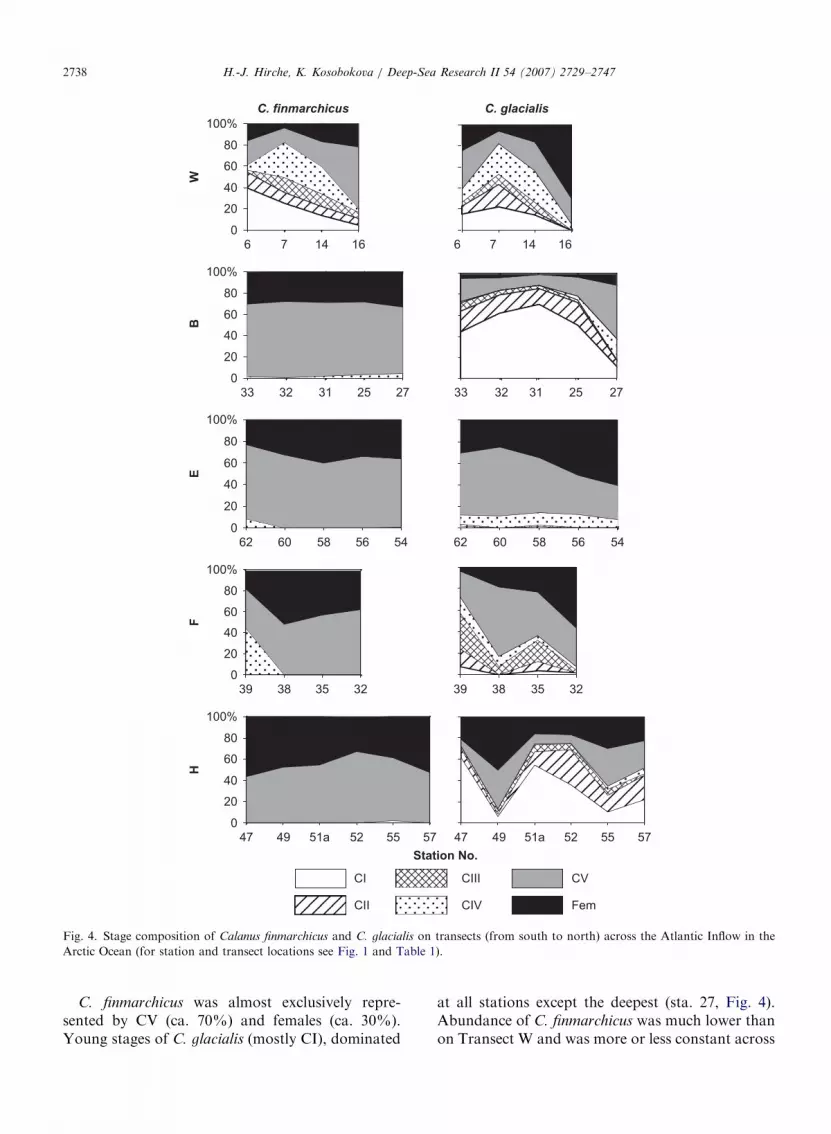
ARTICLE IN PRESS
BF
HE
W
Station No.
C. finmarchicus C. glacialis
0
20
40
60
80
100%
33 32 31 25 27 33 32 31 25 27
6 7 14 16 6 7 14 160
20
40
60
80
100%
47 49 51a 52 55 57 47 49 51a 52 55 570
20
40
60
80
100%
62 60 58 56 54 62 60 58 56 540
20
40
60
80
100%
39 38 35 32 39 38 35 320
20
40
60
80
100%
Fem
CV
CIV
CIII
CII
CI
Fig. 4. Stage composition of Calanus finmarchicus and C. glacialis on transects (from south to north) across the Atlantic Inflow in the
Arctic Ocean (for station and transect locations see Fig. 1 and Table 1).
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472738
C. finmarchicus was almost exclusively repre-sented by CV (ca. 70%) and females (ca. 30%).Young stages of C. glacialis (mostly CI), dominated
at all stations except the deepest (sta. 27, Fig. 4).Abundance of C. finmarchicus was much lower thanon Transect W and was more or less constant across
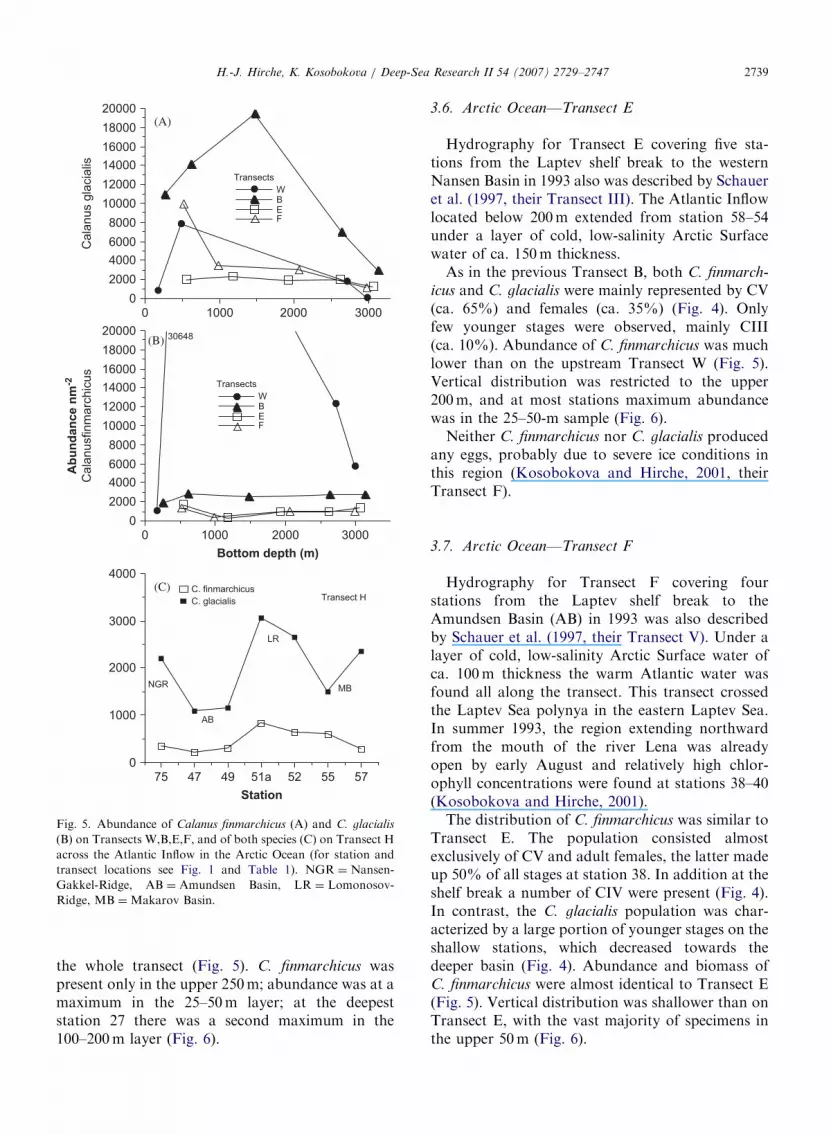
ARTICLE IN PRESS
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
WBEF
Cala
nus g
lacia
lis
30002000100000
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
Bottom depth (m)
75 47 49 51a 52 55 570
1000
2000
3000
4000
C. finmarchicus
C. glacialis
Station
Transects
30648
Ab
un
dan
ce n
m-2
Cala
nusfinm
arc
hic
us
WBEF
Transects
Transect H
3000200010000
NGR
AB
LR
MB
(A)
(B)
(C)
Fig. 5. Abundance of Calanus finmarchicus (A) and C. glacialis
(B) on Transects W,B,E,F, and of both species (C) on Transect H
across the Atlantic Inflow in the Arctic Ocean (for station and
transect locations see Fig. 1 and Table 1). NGR ¼ Nansen-
Gakkel-Ridge, AB ¼ Amundsen Basin, LR ¼ Lomonosov-
Ridge, MB ¼Makarov Basin.
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2739
the whole transect (Fig. 5). C. finmarchicus waspresent only in the upper 250m; abundance was at amaximum in the 25–50m layer; at the deepeststation 27 there was a second maximum in the100–200m layer (Fig. 6).
3.6. Arctic Ocean—Transect E
Hydrography for Transect E covering five sta-tions from the Laptev shelf break to the westernNansen Basin in 1993 also was described by Schaueret al. (1997, their Transect III). The Atlantic Inflowlocated below 200m extended from station 58–54under a layer of cold, low-salinity Arctic Surfacewater of ca. 150m thickness.
As in the previous Transect B, both C. finmarch-
icus and C. glacialis were mainly represented by CV(ca. 65%) and females (ca. 35%) (Fig. 4). Onlyfew younger stages were observed, mainly CIII(ca. 10%). Abundance of C. finmarchicus was muchlower than on the upstream Transect W (Fig. 5).Vertical distribution was restricted to the upper200m, and at most stations maximum abundancewas in the 25–50-m sample (Fig. 6).
Neither C. finmarchicus nor C. glacialis producedany eggs, probably due to severe ice conditions inthis region (Kosobokova and Hirche, 2001, theirTransect F).
3.7. Arctic Ocean—Transect F
Hydrography for Transect F covering fourstations from the Laptev shelf break to theAmundsen Basin (AB) in 1993 was also describedby Schauer et al. (1997, their Transect V). Under alayer of cold, low-salinity Arctic Surface water ofca. 100m thickness the warm Atlantic water wasfound all along the transect. This transect crossedthe Laptev Sea polynya in the eastern Laptev Sea.In summer 1993, the region extending northwardfrom the mouth of the river Lena was alreadyopen by early August and relatively high chlor-ophyll concentrations were found at stations 38–40(Kosobokova and Hirche, 2001).
The distribution of C. finmarchicus was similar toTransect E. The population consisted almostexclusively of CV and adult females, the latter madeup 50% of all stages at station 38. In addition at theshelf break a number of CIV were present (Fig. 4).In contrast, the C. glacialis population was char-acterized by a large portion of younger stages on theshallow stations, which decreased towards thedeeper basin (Fig. 4). Abundance and biomass ofC. finmarchicus were almost identical to Transect E(Fig. 5). Vertical distribution was shallower than onTransect E, with the vast majority of specimens inthe upper 50m (Fig. 6).

ARTICLE IN PRESS
Transect B
0
200
400
600
800
1000
1200
1400
0 10 20 30 40
Sta 33
Sta 32
Sta 31
Sta 25
Sta 27
Transect E
0
200
400
600
800
1000
1200
1400
0 10 15
Abundance, ind m-3
Sta 62
Sta 60
Sta 58
Sta 54
Transect F
0
200
400
600
800
1000
1200
1400
0 10 15 20 25
Dep
th, m
Sta 39
Sta 38
Sta 35
Sta 32
Transect H
0
200
400
600
800
1000
1200
1400
0 10 15
Sta 75
Sta 47
Sta 49
Sta 51
Transect H
0
200
400
600
800
1000
1200
1400
0 10 15
Sta 55
Sta 57
Sta 52
Transect W
0
200
400
600
800
1000
1200
1400
0 50 100 150
Dep
th, m
Sta 6
Sta 7
Sta 16
Sta 14
5
5 5 5
Fig. 6. Vertical distribution of Calanus finmarchicus on transects across the Atlantic Inflow in the Arctic Ocean (for station and transect
locations see Fig. 1 and Table 1).
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472740
In Fig. 7 gonad stage and egg production rateson Transect F are compared for C. finmarchicus andC. glacialis (C. glacialis from Kosobokova andHirche, 2001, their Transect H). Between 62%and 92% of the C. finmarchicus females were spentor showed signs of resorption, the rest of thefemales were immature. No eggs were produced
during experiments at five stations. In contrast,young copepodids of C. glacialis were present atfour stations on the slope near the ice edge(Kosobokova et al., 1998, their Table 6). Femalesproduced up to 8 eggs female�1 d�1 at four stations,in most females gonads were spent, with up to 30%immature.

ARTICLE IN PRESS
Go
nad
matu
rity
(%
)
0
20
40
60
80
100
Cala
nu
s f
inm
arc
hic
us
EPR 3.3 0.4 0 1.0
immaturesemi-riperipesemi-spentspent
31
38
43
53
41
72
40
243
39
513
38
982
35
2062
32
2975
Transect FTransect W
Station
Depth (m)
0
20
40
60
80
100
Cala
nu
s g
lacia
lis
6
180
14
2720
19
2994
67
490
22.3 15.7
Go
nad
matu
rity
(%
)
EPR
-- 0 -- 0 0 0
-- -- --3.1 0 2.6 2.5 8.3 00.1
0 0
Fig. 7. Gonad maturation stages and egg production rates (EPR) (eggs female�1 d�1) of Calanus finmarchicus and C. glacialis on Transects
W and F across the Atlantic Inflow in the Arctic Ocean (see Fig. 1). Data for C. glacialis from Kosobokova and Hirche, 1991).
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2741
3.8. Arctic Ocean—Transect H
Hydrography and ice cover during Transect Hfrom the AB across the Lomonosov Ridge to theMakarov Basin (MB) were described in detail byRudels et al. (2000, part of their Transect C) andsummarized by Kosobokova and Hirche (2000).The core of the Atlantic water flows to the north onthe Eurasian side of the Lomonosov Ridge (stations51a, 52). Two branches of Atlantic water from theNorwegian Sea were identified by salinity maxima,representing the Barents Sea branch and the FramStrait branch. The Atlantic water probably crossesthe ridge at various latitudes south of 86.51N andflows southward along the MB side of the ridge(Woodgate et al., 2001).
While all copepodite stages of C. glacialis werepresent at almost all stations, C. finmarchicus wasalmost exclusively represented as CV and adults inalmost equal proportion (Fig. 4). Highest concen-trations of both species were observed on the crestof the Lomonosov Ridge (stations 51a, 52), wherethe core of Atlantic water was located. Abundanceof C. finmarchicus on this transect ranged from 204to 830 individuals m�2 and was the lowest of all
transects (Fig. 5). This is in good agreement withhydrographic observations that near the Laptevshelf about half of the Atlantic Inflow flow isdiverted north along the Eurasian Basin side of theLomonosov Ridge (Woodgate et al., 2001). As onall other transects, C. finmarchicus were concen-trated in the upper 100m with the maximumabundance in the 20–50-m layer. At some stationsa secondary maximum was observed between 100and 200m (Fig. 6).
Although egg production experiments were set upat all stations, neither C. finmarchicus nor C. glacialis
spawned any eggs.
4. Temperature and reproduction—laboratory
studies
In egg production experiments, C. finmarchicus
collected in the northern Norwegian Sea at 9 1Cwere incubated at five temperatures after acclima-tion at 0 1C for 4–7 days (Hirche et al., 1997). Theylaid eggs in regular intervals during ca. 2 weeks.Egg production rate increased exponentially withtemperature over the range between �1.5 and 8 1C.

ARTICLE IN PRESS
9876543210-1-20
20
40
60
80C.glacialis
C.finmarchicusR
2 = 0.979
R2 = 0.976
Eg
g p
rod
ucti
on
rate
(eg
gs f
em
ale
-1 d
-1)
Temperature (°C)
Fig. 8. Egg production rates of Calanus finmarchicus and
C. glacialis at different temperatures and optimum food concen-
trations (data for C. finmarchicus from Hirche et al., 1997),
(vertical bars7st.dev.).
H.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472742
The temperature response of C. glacialis treatedequally in the experiments was very similar (Fig. 8).
5. Discussion
5.1. Distribution and hydrography
During this study C. finmarchicus was observed atall stations studied in the Greenland Sea, theBarents Sea, in the Eurasian Basins, and the MBin the Arctic Ocean. Including published informa-tion on Davis Strait (Huntley et al., 1983) andBaffin Bay (Ringuette et al., 2000), we can general-ize that the species is found all over the NorthAtlantic and the eastern Arctic Ocean (Jaschnov,1970; Conover, 1988) in close association with theflow of Atlantic water. This makes the species agood tracer, as suggested earlier by Jaschnov (1970).Depending on the depth distribution and flowpattern of the Atlantic water, three differentexpatriation patterns are possible. (1) On transectGS across the AF in the Greenland Sea, Atlanticand Arctic water masses are aligned in the upperlayer. The biomass of C. finmarchicus decreased by afactor 10 within 10 nm of the frontal zone indicatinglittle cross-frontal exchange, despite the numerouseddies in this there (Trees et al., 1992; Walczowskiand Maslowski, 2003) that could accelerate cross-frontal exchange processes. (2) On Transect BSacross the Polar Front of the Barents Sea (Hircheand Kosobokova, 2003) Atlantic and Polar watersare also aligned in the upper layer, but Atlantic
water, including its fauna, submerges under thePolar water below ca. 100m. (3) While in thenorthern Barents Sea Atlantic water submerges overa wide area, the RAC and Atlantic Inflow formrelatively narrow submerged bands of flow travel-ling long distances from their origin in the FramStrait (Bourke et al., 1988; Woodgate et al., 2001).C. finmarchicus preferred the surface or near-surfacelayer, despite the fact that the submerged Atlanticwater is covered by a layer of cold Polar water oftenmore than 200m in thickness. At most stations, thevertical distribution maximum was between 25 and50m. In addition, at some stations a secondmaximum was found between 100 and 200m. Incontrast, in the Atlantic layer between 200 and900m, hardly any C. finmarchicus were observed(Fig. 6). Similarly, in the northern Barents SeaC. finmarchicus was found in the upper layersdirectly north of the front, indicating an immediateascent after submergence of the Atlantic water(Hirche and Kosobokova, 2003). This verticaldistribution should have strong consequences forthe advection of C. finmarchicus, as the submergedAtlantic water and the surface layer often flow indifferent directions (Woodgate et al., 2001).Although for the Arctic Ocean no data on thevertical distribution in winter are available, weassume, based on observations from the GreenlandSea (Hirche, 1991) and Norwegian Sea (Østvedt,1955), that C. finmarchicus performs seasonalmigrations and spends most of its life in deeperwaters, where it is then advected along with theAtlantic Inflow farther into the Arctic Ocean.Furthermore, we assume that overwintering stagesmaintain their depth when Atlantic water sub-merges. As a consequence, the speed and directionof advection of C. finmarchicus in all areas ofsubmergence of Atlantic water should underlieseasonal variations.
5.2. Expatriation and factors controlling recruitment
Our observations on stage composition, abun-dance and reproductive activity clearly suggest thatC. finmarchicus is not recruiting successfully in thenorthern Barents Sea nor in the Arctic Ocean.Young developmental stages were only present nearthe source regions, as on Transect W shortly pastthe entrance to the Arctic Ocean, and on TransectBS just north of the Polar Front (Hirche andKosobokova, 2003), where they were probablyadvected only recently. Furthermore, a downstream

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2743
decrease of abundance and biomass along theAtlantic Inflow and across the Lomonossov Ridgewas observed for consecutive transects during thesame years (1995: B, H; 1997: W, E, F). Gonadstages and egg production measurements furthersupport this view. In contrast, in the westernGreenland Sea, where Atlantic water of the RACis squeezed in between the GSG and the EGC, theC. finmarchicus population seemed to thrive well onour transect, judging from the presence of spawningfemales despite water temperatures between +11and �1 1C. Apparently its ability to survive longperiods with little energetic effort makes C. finmar-
chicus able to enter far into regions of expatriation.Indeed, it is known to over winter for as alongas 9 months. CIV, CV, and females can all beover wintering stages (Hirche, 1996), whereas theyounger stages have little to no lipid reserves(Kattner and Krause, 1987) and therefore cannotsurvive long starvation periods. But which factorslimit successful development in Polar Regions?
Temperature is the factor blamed most often inthe literature (Jaschnov, 1970; Sameoto, 1984;Tande et al., 1985). It may affect hatching, juveniledevelopment, gonad maturation, and overwinteringphysiology. According to Sameoto (1984), thestages CII, CIII and CIV have a narrowertemperature tolerance than CV and adults. So farthere is little knowledge about the effect of sustainedlow temperature on the development of C. finmarch-
icus, and our study cannot contribute to this issue.Reproductive biology has been studied moreintensively. Previous egg production experiments(Hirche, 1990; Hirche et al., 1997) and fieldobservations from this study in the MIZ of theGreenland Sea indicate that temperature per se maynot be limiting reproduction.
Other factors that may be important for advectedspecimens are the match of food availability andintrinsic timing of development. In the NorthAtlantic, moulting of adults and subsequent fertili-zation of the females by the short-lived males takesplace in late winter (Marshall and Orr, 1955). Whenfood is available, females produce eggs and spawn.Males were not observed in our samples. Theproduction of males is a sensitive phase in the lifecycle of C. finmarchicus (Irigoien et al., 2000). Eithermale development was inhibited by low tempera-tures or they died before our sampling period.The dominance of females in stage composition inthe eastern Arctic Ocean clearly indicates en route
development, assuming that the specimens collected
in late summer entered the Arctic Ocean during theprevious fall and winter as CV, as this stage is by farthe dominant over wintering stage in the GreenlandSea (Hirche, 1991). This assumption is based onmooring data near the Lomonossov Ridge obtainedduring our sampling (Woodgate et al., 2001), whichshow that Barents Sea winter water was present onour sampling locations and that the Atlantic Inflowwas advected at approximately 5 cm s�1 (Woodgateet al., 2001). While in the North Atlantic thephytoplankton usually blooms in late spring (Braar-ud et al., 1958), in the Arctic Ocean blooms areobserved in August/September (Grahl et al., 1999),which is several months later. This delay may havecaused the gonad absorption observed on TransectF. Indeed, such absorption processes occur whenfemales are exposed to long starvation periods(Kosobokova, 1999; Niehoff, 2000) and may thusexplain their reproductive failure. Feeding super-abundant food to such females for 11 days inducedegg production only in very few specimens (Hircheand Mumm, 1992). Regional differences in thebreeding cycle of C. finmarchicus are common andwere often attributed to differences in the timing ofphytoplankton development (Huntley et al., 1983;Plourde and Runge, 1993; Ringuette et al., 2000).However, in the Arctic, phytoplankton may developjust too late to be utilized by C. finmarchicus forgonad maturation.
Observations of reproductive activity in the RACand MIZ of the Greenland Sea or near-frontal regionof the Barents Sea support the hypothesis ofreproductive failure due to mismatch between thetiming of the female reproductive cycle and foodavailability. In the RAC, specimens ascending intothe polar layer often find favourable food conditionsin the MIZ. Due to its association with the EGPF,this region is ice-free or covered only with thin ice allyear round. Stability induced by melt water allowsearly phytoplankton growth (Smith, 1987). Conse-quently, egg production was found at low tempera-tures when food was sufficient (Fig. 3). However, inBelgica Trough on the EGS, not far away from theMIZ, but with low food concentrations due to heavyice cover for most of the year (Lara et al., 1994), nosigns of growth or reproductive activity weredetected (Hirche et al., 1994; Hirche and Kwasniews-ki, 1997). In the near-frontal region of the BarentsSea, hydrographic conditions are similar to the MIZin the Greenland Sea. The specimens found theremay have been advected only recently together withphytoplankton produced in the MIZ.

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472744
5.3. Comparison of C. finmarchicus and C. glacialis
Comparison of the congeners C. finmarchicus andC. glacialis shows that the latter is at present betteradapted to life in the Arctic, because it can betterutilize the patchy and mostly late phytoplanktonoccurrence that is often associated with polynyassuch as the Northwater polynya in Baffin Bay(Ringuette et al., 2000), Northeast Water polynyaon the EGS (Hirche et al., 1994; Ashjian et al., 1995,1997; Hirche and Kwasniewski, 1997), and LaptevSea polynya (Kosobokova and Hirche, 2001).Females of C. glacialis are long-lived (Kosobokova,1999) and are able to spawn if fed after longstarvation periods (Hirche, 1989). The use of icealgae (Runge and Ingram, 1988) further expands itsgrowth period.
5.4. Future perspectives
Even if mostly inactive, the export of a largebiomass of C. finmarchicus represents a big loss forthe North Atlantic, whereas for the Arctic, thisexport represents both prey for local predators anda large source of carbon that will sink in the areas ofexpatriation. Under continuing warming andstrengthening of the Atlantic Inflow, different scenar-ios are expected in the future. An increase of advectionof Atlantic populations may significantly increase thesedimentation of advected biogenic material. Furtherwarming could favour the survival of the highlyproductive Atlantic communities, which finally couldreplace the Arctic fauna, which is characterized by lowbiomass and low production (Hirche and Mumm,1992; Kosobokova and Hirche, 2000). Especially inthe subarctic shelf seas, a shift from C. glacialis toC. finmarchicus is likely under continued climatechange, as not only the survival of C. finmarchicus
would be favoured by higher temperatures and earlierblooms due to earlier ice melt, but C. glacialis wouldbe inhibited. Kosobokova (1999) concluded fromcomparison of gonad stages and the vertical distribu-tion of C. glacialis collected in the White Sea indifferent years that the key factor causing thetermination of the spawning period of C. glacialis inearly summer was the increase of temperature in thesurface layer. When daily average temperatures nearthe surface reached 5 1C, females left the surface layersand stopped spawning. This also was observed in theLurefjord by Niehoff and Hirche (2005). In both theWhite Sea and the Lurefjord, 5 1C seems to be astrong threshold above which dormancy is induced in
female C. glacialis. Thus, a replacement of C. glacialis
by C. finmarchicus may happen first only in the laterpart of the year, when surface temperatures aresurpassing this limit.
Further colonization should depend on the estab-lishment of over wintering populations and changesin the regional ecosystem. We assume that it will becrucial whether climate change leads to an earlierstart of phytoplankton development. The transitionperiod could be very hazardous to upper trophiclevels due to the differences in size between the twospecies. C. finmarchicus has a maximum femaleprosome length of 3.2mm as compared to 4.6mmin C. glacialis. Correspondingly, maximum carboncontent is 100–225mg versus 600mg (Hirche andKwasniewski, 1997). Little auks (Alle alle) from BearIsland only sporadically take the abundant copepodC. finmarchicus, but actively select the much lessabundant and larger C. glacialis (Weslawski et al.,1999; Karnovsky et al., 2003).
Acknowledgements
We thank the captains and crew of the variousRV ‘‘Polarstern’’ cruises. We appreciate the help ofmany colleagues who assisted the sampling, experi-ments, sample analysis and preparation of themanuscript, among others R.N. Bohrer, U. Holtz,S. Kwasniewski, D. Mengedoht, U. Meyer and T.Scherzinger. V. Kosobokov and A. Basilico helpedwith the maps, R. Schwamborn with the statistics.The work of KNK was supported by RussianFoundation for Basic research Grants nos. 03-05-64871 and 06-05-65187 and visiting scientist grantsof the AWI. This paper was first presented in theGLOBEC-ESSAS Symposium on effects of climatevariability on sub-arctic marine ecosystems, hostedby PICES in Victoria, BC, May 2005.
References
Anderson, L.G., Jones, E.P., Koltermann, K.P., Schlosser, P.,
Swift, J.H., Wallace, D.W.R., 1989. The first oceanographic
section across the Nansen Basin in the Arctic Ocean. Deep-
Sea Research 36, 475–482.
Ashjian, C.J., Smith, S.L., Lane, P.V.Z., 1995. The Northeast
Water Polynya during summer 1992: distribution and aspects
of secondary production of copepods. Journal of Geophysical
Research 100 (C3), 4371–4388.
Ashjian, C.J., Smith, S.L., Bignami, F., Hopkins, T., Lane,
P.V.Z., 1997. Distribution of zooplankton in the Northeast
Water Polynya during summer 1992. Journal of Marine
Systems 10, 279–298.

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2745
Bourke, R.H., Weigel, A.M., Paquette, R.G., 1988. The
westward turning branch of the West Spitsbergen Current.
Journal of Geophysical Research 93, 14065–14077.
Braarud, T.K., Ringdal, G., Nordli, J., 1958. Seasonal changes in
the phytoplankton at various points off the Norwegian West
Coast (observations at the permanent oceanographic station,
1945–1946). Fiskeridirektoratets Skrifter Serie Havundersø-
kelser 12/3, 1–77.
Budeus, G., Schneider, W., 1995. On the hydrography of the
Northeast Water Polynya. Journal of Geophysical Research
100, 4287–4299.
Budeus, G., Maul, A.-A., Krause, G., 1993. Variability in the
Greenland Sea as revealed by a repeated high spatial
resolution conductivity-temperature-depth survey. Journal
of Geophysical Research 98 (C6), 9985–10000.
Campbell, R.G., Wagner, M.M., Teegarden, G.J., Boudreau, C.A.,
Durbin, E.G., 2001. Growth and development rates of Calanus
finmarchicus reared in the laboratory. Marine Ecology Progress
Series 221, 161–183.
Carmack, E.C., Macdonald, R.W., Perkin, R.G., McLaughlin, F.A.,
Pearson, R.J., 1995. Evidence for warming of Atlantic water in
the southern Canadian Basin of the Arctic Ocean: results from
the Larsen-93 expedition. Geophysical Research Letters 22,
1061–1064.
Conover, R.J., 1988. Comparative life histories in the genera
Calanus and Neocalanus in high latitudes of the northern
hemisphere. Hydrobiology 167/168, 127–142.
Corkett, C.J., McLaren, I.A., Sevigny, J.M., 1986. The rearing of
the marine calanoid copepods Calanus finmarchicus (Gun-
nerus), C. glacialis (Jaschnov) and C. hyperboreus (Krøyer)
with comment on the equiproportional rule. In: Schriever, G.,
Schminke, H.K., Shih, C.T. (Eds.), Proceedings of 2nd
International Conference on Copepoda, 13–17 August 1984,
Ottawa: Syllogeus, vol. 58. National Museum Canada,
pp. 539–546.
Gascard, J.C., Kergomard, C., Jeannin, P.F., Fily, M., 1988.
Diagnostic study of the Fram Strait Marginal Ice Zone during
summer from 1983 and 1984 Marginal Ice Zone Experiment.
Lagrangian observations. Journal of Geophysical Research
93, 3613–3641.
Grahl, C., Boetius, A., Noethig, E.-M., 1999. Pelagic-benthic
coupling in the Laptev Sea affected by ice cover. In:
Kassens, H., Bauch, H.A., Dmitrenko, I.A., Eicken, H.,
Hubberten, H.-W., Melles, M., Thiede, J., Timokhov, L.A.
(Eds.), Land-Ocean Systems in the Siberian Arctic. Springer,
Berlin, Heidelberg, New York, pp. 143–150.
Hansen, B., Christiansen, S., Pedersen, G., 1996. Plankton
dynamics in the marginal ice zone of the central Barents Sea
during spring: carbon flow and structure of the grazer food
chain. Polar Biology 16, 115–128.
Hirche, H.J., 1989. Egg production of the Arctic copepod
Calanus glacialis—laboratory experiments. Marine Biology
103, 311–318.
Hirche, H.J., 1990. Egg production of Calanus finmarchicus at
low temperatures. Marine Biology 10, 53–58.
Hirche, H.J., 1991. Distribution of dominant calanoid copepod
species in the Greenland Sea during late fall. Polar Biology 11,
351–362.
Hirche, H.J., 1996. Diapause in the marine copepod, Calanus
finmarchicus—A review. Ophelia 44, 129–143.
Hirche, H.J., 2004. Zooplankton habitats of the Greenland Sea—
an experimental laboratory for studies of pelagic ecology.
In: Skreslet, S. (Ed.), Jan Mayen Island in Scientific Focus.
NATO Sciences Series. IV. Earth and Environmental
Sciences, vol. 45, pp. 123–133.
Hirche, H.J., Kosobokova, K.N., 2003. Early reproduction and
development of dominant calanoid copepods in the sea ice
zone of the Barents Sea—need for a change of paradigms?
Marine Biology 143, 769–781.
Hirche, H.J., Kwasniewski, S., 1997. Distribution, reproduction
and development of Calanus species in the Northeast Water in
relation to environmental conditions. Journal of Marine
Systems 10, 299–317.
Hirche, H.J., Mumm, N., 1992. Distribution of dominant
copepods in the Nansen Basin, Arctic Ocean, in summer.
Deep-Sea Research 39 (Suppl. 2), S485–S505.
Hirche, H.J., Hagen, W., Mumm, N., Richter, C., 1994. The
Northeast Water Polynya, Greenland Sea. III. Meso- and
macrozooplankton distribution and production of dominant
herbivorous copepods during summer. Polar Biology 14,
491–503.
Hirche, H.J., Meyer, U., Niehoff, B., 1997. Egg production of
Calanus finmarchicus: effect of temperature, food and season.
Marine Biology 127, 609–620.
Huntley, M., Strong, K.W., Dengler, A.T., 1983. Dynamics and
community structure of zooplankton in the Davis Strait and
northern Labrador Sea. Arctic 36, 143–161.
Irigoien, X., Obermueller, B., Head, R.N., Harris, R.P.,
Rey, C., Hansen, B.W., Hygum, B.H., Heath, M.R.,
Durbin, E.G., 2000. The effect of food on the determination
of sex ratio in Calanus spp.: evidence from experimental
studies and field data. ICES Journal of Marine Science 57,
1752–1763.
Jaschnov, W.A., 1970. Distribution of Calanus species in the seas
of the northern hemisphere. Internationale Revue der
Gesamten Hydrobiologie 55, 197–212.
Johannessen, J.A., Johannessen, O.M., Svendsen, E., Shuchman, R.,
Manley, T.O., Campbell, W., Josberger, E., Sandven, S.,
Gascard, J.C., Olaussen, T., Davidson, K., Van Leer, J., 1987.
Mesoscale Eddies in the Fram Strait Marginal Ice Zone During
MIZEX 1983 and 1984. Journal of Geophysical Research 92
(C7), 6754–6772.
Johannessen, O.M., Bengtsson, L., Miles, M.W., Kuzmina, S.I.,
Semenov, V.A., Alekseev, G.V., Nagurnyi, A.P., Zakharov,
V.F., Bobylev, L., Pettersson, L.H., Hasselmann, K., Cattle,
H.P., 2002. Arctic climate change-observed and modeled
temperature and sea ice variability. Technical Report No. 218,
Nansen Environmental and Remote Sensing Center, Bergen,
pp. 1–22.
Karnovsky, N.J., Kwasniewski, S., Weslawski, J.M., Walkusz, W.,
Beszczynska-Moller, A., 2003. Foraging behavior of little auks
in a heterogeneous environment. Marine Ecology Progress
Series 253, 289–303.
Kattner, G., Krause, M., 1987. Changes in lipids during the
development of Calanus finmarchicus s.l. from copepodid I to
adult. Marine Biology 96, 511–518.
Kosobokova, K.N., 1999. The reproductive cycle and life history
of the Arctic copepod Calanus glacialis in the White Sea.
Polar Biology 22, 254–263.
Kosobokova, K.N., Hirche, H.J., 2000. Zooplankton distribution
across the Lomonosov Ridge, Arctic Ocean: species inven-
tory, biomass and vertical structure. Deep-Sea Research I 47,
2029–2060.

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–27472746
Kosobokova, K.N., Hirche, H.J., 2001. Reproduction of Calanus
glacialis in the Laptev Sea, Arctic Ocean. Polar Biology 24,
33–43.
Kosobokova, K.N., Hanssen, N., Hirche, H.J., Knickmeier, K.,
1998. Composition and distribution of zooplankton in the
Laptev Sea and adjacent Nansen Basin in the summer 1993.
Polar Biology 19, 63–76.
Kosteyn, Y., Kwasniewski, S., 1992. The near shore zooplankton
of the Tikhaia Bay (Franz Josef Land) in August 1991. In:
Gjertz, I., Mørkved, B. (Eds.), Environmental Studies from
Franz Josef Land with Emphasis on Tikhaia Bay, Hooker
Island, vol. 120. Norsk Polarinstitutt, Meddelser, pp. 23–34.
Lara, R.J., Kattner, G., Tillmann, U., Hirche, H.J., 1994. The
North East Water polynya (Greenland Sea). II. Mechanisms
of nutrient supply and influence on phytoplankton distribu-
tion. Polar Biology 14, 483–490.
Loeng, H., 1991. Features of the physical oceanographic
conditions of the Barents Sea. Proceedings of the Pro Mare
Symposium on Polar Marine Ecology, Trondheim, 12–16
May 1990. Polar Research 10, 5–18.
Manley, T.O., 1987. Effects of sub-ice mesoscale features within
the marginal ice zone of Fram Strait. Journal of Geophysical
Research 92 (C4), 3944–3960.
Marshall, S.M., Orr, A.P., 1955. The Biology of a Marine
Copepod Calanus finmarchicus Gunnerus. Oliver and Boyd,
Edinburgh.
McLaughlin, F.A., Carmack, E.C., Macdonald, R.W., Bishop,
J.K.B., 1996. Physical and geochemical properties across the
Atlantic/Pacific water mass front in the southern Canadian
Basin. Journal of Geophysical Research 101, 1183–1197.
McLaughlin, F., Carmack, E., MacDonald, R.W., Weaver, A.J.,
Smith, J., 2002. The Canada Basin 1989–1995: Upstream
events and far-field effects of the Barents Sea. Journal of
Geophysical Research 107 (C7), 3082.
Meincke, J., Jonsson, S., Swift, J.H., 1992. Variability of
convective condition in the Greenland Sea. ICES Marine
Science Symposium 1095, 32–39.
Niehoff, B., 2000. The effect of starvation on the reproductive
potential of Calanus finmarchicus. ICES Journal of Marine
Systems 57, 1764–1772.
Niehoff, B., Hirche, H.J., 1996. Oogenesis and gonad maturation
in the copepod Calanus finmarchicus and the prediction of egg
production from preserved samples. Polar Biology 16,
601–612.
Niehoff, B., Hirche, H.J., 2005. Reproduction of Calanus glacialis
in the Lurefjord (western Norway): indication for tempera-
ture-induced female dormancy. Marine Ecology Progress
Series 285, 107–115.
Østvedt, O.J., 1955. Zooplankton investigations from weather-
ship ‘‘M’’ in the Norwegian Sea, 1948–49. Hvalradets Skrifter
40, 1–93.
Pfirman, S.L., Bauch, D., Gammelsrød, T., 1994. The northern
Barents Sea: water mass distribution and modification. The
Polar Oceans and their role in shaping the global environ-
ment. Geophysical Monographs Series 85, 77–93.
Plourde, S., Runge, J.A., 1993. Reproduction of the planktonic
copepod Calanus finmarchicus in the Lower St. Lawrence
Estuary: relation to the cycle of phytoplankton production
and evidence for a Calanus pump. Marine Ecology Progress
Series 102, 217–227.
Polyakov, I., Alekseev, G., Bekryaev, R., Bhatt, U., Colony, R.,
Johnson, M., Karklin, V., Makshtas, A., Walsh, D., Yulin, A.,
2002. Observationally based assessment of polar amplification
of global warming. Geophysical Research Letters 29 (18),
1878–1881.
Ringuette, M., Fortier, L., Fortier, M., Runge, J., Belanger, S.,
Larouche, P., Weslawski, J.M., Kwasniewski, S., 2000. Spring
and summer population development of Arctic copepods in
and outside polynyas: the North Water versus Barrow Strait.
Deep-Sea Research II 22–23, 5081–5099.
Rudels, B., Jones, E.P., Anderson, L.G., Kattner, G., 1994. On
the intermediate depth waters of the Arctic Ocean. The Polar
Oceans and their role in shaping the global environment.
Geophysical Monographs Series 85, 33–46.
Rudels, B., Muench, R.D., Gunn, J., Schauer, U., 2000.
The Arctic Ocean north of the Siberian shelves: advec-
tion, lateral mixing, slope convection and intermediate and
deep water characteristics. Journal of Marine Systems 25,
77–99.
Runge, J.A., Ingram, R.G., 1988. Underice grazing by plank-
tonic, calanoid copepods in relation to a bloom of ice
microalgae in southeastern Hudson Bay. Limnology and
Oceanography 33, 280–286.
Sameoto, D.D., 1984. Vertical distribution of zooplankton
biomass and species in northeastern Baffin Bay related to
temperature and salinity. Polar Biology 2, 213–224.
Schauer, U., Muench, R.D., Rudels, B., Timokhov, L., 1997.
Impact of eastern shelf waters on the Nansen Basin
intermediate layers. Journal of Geophysical Research 102
(C2), 3371–3382.
Schauer, U., Loeng, H., Rudels, B., Ozhigin, V.K., Dieck, W.,
2002. Atlantic Water flow through the Barents and Kara Sea.
Deep-Sea Research I 49, 2281–2298.
Smith, S.L., Smith, W.O., Codispoti, L.A., Wilson, D.L.,
1985. Biological observations in the marginal ice zone
of the East Greenland Sea. Journal of Marine Research 43,
693–717.
Smith, W.O., 1987. Phytoplankton dynamics in marginal ice
zones. Oceanography and Marine Biology Annual Review 25,
11–38.
Smith, S.L., 1990. Egg production and feeding by copepods
prior to the spring bloom of phytoplankton in the Fram
Strait area of the Greenland Sea. Marine Biology 106,
59–69.
Swift, J.H., Jones, E.P., Aagaard, K., Carmack, E.C., Hingston, M.,
MacDonald, R.W., McLaughlin, F.A., Perkin, R.G., 1997.
Waters of the Makarov and Canada basins. Deep-Sea Research
II 44, 1503–1529.
Tande, K.S., Hassel, A., Slagstad, D., 1985. Gonad maturation
and possible life cycle strategies in Calanus finmarchicus and
Calanus glacialis in the northwestern part of the Barents Sea.
In: Gray, J.S., Christiansen, M.E. (Eds.), Marine Biology of
Polar Regions and Effects of Stress on Marine Organisms.
Wiley, Chichester, New York, pp. 141–155.
Thompson, B.M., 1982. Growth and development of Pseudoca-
lanus elongatus and Calanus sp. in the laboratory. Journal of
the Marine Biological Association of the United Kingdom 62,
359–372.
Trees, C.C., Aiken, J., Hirche, H.J., Groom, S.B., 1992. Bio-
optical variability across the Arctic Front. Polar Biology 12,
455–461.
Unstad, K.H., Tande, K.S., 1991. Depth distribution of Calanus
finmarchicus and Calanus glacialis in relation to environ-

ARTICLE IN PRESSH.-J. Hirche, K. Kosobokova / Deep-Sea Research II 54 (2007) 2729–2747 2747
mental conditions in the Barents Sea. Polar Research 10,
389–407.
Walczowski, W., Maslowski, W., 2003. Mesoscale structures in
the subarctic seas-observations and modelling. Geophysical
Research Abstracts 5, 05662.
Weslawski, J.M., Stempniewicz, L., Mehlum, F., Kwasniewski, S.,
1999. Summer feeding strategy of the little auk (Alle alle) from
Bjørnøya, Barents Sea. Polar Biology 21, 129–134.
Woodgate, R.A., Aagaard, K., Muench, R.D., Gunn, J., Bjork, G.,
Rudels, B., Roach, A.T., Schauer, U., 2001. The Arctic Ocean
Boundary Current along the Eurasian slope and the adjacent
Lomonosov Ridge: water mass properties, transports and
transformations from moored instruments. Deep-Sea Research
I 48, 1757–1792.
Zar, J., 1996. Biostatistical Analysis, third ed. Prentice-Hall,
Englewood Cliffs, NJ.