disruption of the b-cell specific transcriptional program in hhv-8 associated primary effusion...
TRANSCRIPT

Disruption of the B-cell specific transcriptional program in HHV-8
associated primary effusion lymphoma cell lines
Meztli Arguello1,2, Marco Sgarbanti1, Eduardo Hernandez1, Yael Mamane1,2, Sonia Sharma1,2,Marc Servant1,3, Rongtuan Lin1,3 and John Hiscottn,1,2,3
1Terry Fox Molecular Oncology Group, Lady Davis Institute for Medical Research, 3755 Cote-Ste., Catherine Road, Montreal,Quebec, Canada H3T-1E2; 2Department of Microbiology and Immunology, McGill University, Montreal, Canada H3T- 1E2;3Department of Medicine, McGill University, Montreal, Canada H3T-1E2
Primary effusion lymphoma (PEL) is a lymphoprolifera-tive disease of B-cell origin that is associated with HHV-8infection. PEL cells harbor a non-B, non-T phenotype andlack significant surface immunoglobulin (Ig) expression, acharacteristic that has not been fully explained. In thepresent study, we demonstrate that PEL cells constitu-tively express interferon regulatory factor (IRF)-4, atranscription factor that regulates the activity of theimmunoglobulin light-chain enhancer elements kB andjE30 through binding to a composite Ets-IRF site. IRF-4activity requires its physical interaction with PU.1, an Etsfamily member involved in the activation of genes essentialfor B-cell development. However, in PEL-derived B-celllines, PU.1 expression was completely abrogated; expres-sion of the B cell specific transcription factor Oct-2, whichis known to regulate PU.1 expression, was also abolished.Moreover, the B-cell-specific coactivator of octamerfactors, BOB-1/OcaB, was expressed at very decreasedlevels in PEL cells. Ectopic expression of Oct-2 was ableto fully restore PU.1 promoter activity in the PEL cell lineBCBL-1, while PU.1 expression also reconstituted theactivity of the kB Ets-IRF site. In addition, protein levelsof BSAP/Pax-5 and IRF-8/ICSBP were undetectable inPEL cells. The pattern of transcription factor ablationobserved in PEL was found to be comparable to thatobserved in classical Hodgkin’s disease-derived cell lines,which also lack B-cell-specific surface markers. Theseobservations indicate that disruption of the B-cell-specifictranscriptional program is likely to contribute to theincomplete B-cell phenotype characteristic of PEL cells.Oncogene (2003) 22, 964–973. doi:10.1038/sj.onc.1206270
Keywords: primary effusion lymphoma (PEL); IRF-4;PU.1; Oct-2; HHV-8
Introduction
Primary effusion lymphoma (PEL) is an unusual andaggressive lymphoproliferative disease associated withinfection by Kaposi’s sarcoma-associated herpesvirus(KSHV/HHV-8) (Chang et al., 1994; Cesarman et al.,1995). PEL occurs predominantly in immunocompro-mised individuals such as AIDS patients (Drexler et al.,1998). The disease has been characterized as being of B-cell origin and although the exact maturation stage ofPEL cells is still controversial (Matolcsy et al., 1998;Carbone et al., 2000b), it is thought to arise from cells ofa mature, postgerminal center, preterminal differentia-tion stage (Nador et al., 1996; Gaidano et al., 1997).PEL cells lack expression of most typical B-cell surfacemarkers such as CD20, CD22, and CD19, as well asexpression of surface immunoglobulin (Ig) (Drexleret al., 1998). In fact, identification of PEL cells as beingof B-cell origin was ultimately based on genotypicanalysis demonstrating Ig gene rearrangement in allPEL cases.
B-cell-specific expression of immunoglobulin genes isregulated by the complex interaction between regulatoryDNA elements and B-cell-specific transcription factors(reviewed in Singh, 1994; Reya and Grosschedl, 1998).Among the regulatory DNA elements, the l2-4 and kE30
light-chain enhancers contain a composite Ets/IRFelement that is essential for their activity (Pongubalaet al., 1992; Eisenbeis et al., 1993, 1995). This site iscoordinately regulated by the transcription factors PU.1and IRF-4, which form a ternary complex andsynergistically activate transcription (Brass et al., 1996,1999). Such a composite Ets-IRF element is present aswell in the CD20 promoter, among others (Himmel-mann et al., 1997). Interferon regulatory factor (IRF)-4is a member of the IRF family of transcription factorsthat regulate cytokine expression, cell growth, andimmunomodulation (reviewed in Marecki and Fenton,2002; Mamane et al., 2002b; Pernis, 2002). IRF-4expression is specifically restricted to the lymphoid andmyeloid compartments (Eisenbeis et al., 1995), and in Bcells expression is associated with the final steps ofintragerminal center B-cell differentiation and matura-tion towards plasma cells (Falini et al., 2000). At the
Received 24 June 2002; revised 22 November 2002; accepted 22November 2002
*Correspondence: J Hiscott, Lady Davis Institute for MedicalResearch, 3755 Cote-Ste.-Catherine Road, Montreal, Quebec, CanadaH3T 1E2; E-mail: [email protected]
Oncogene (2003) 22, 964–973& 2003 Nature Publishing Group All rights reserved 0950-9232/03 $25.00
www.nature.com/onc

molecular level, IRF-4 has a weak intrinsic DNAbinding activity and its transactivation potentialwith respect to the Ets-IRF element depends onthe formation of a complex with PU.1 (Pongubalaet al., 1992; Eisenbeis et al., 1993, 1995; Brass et al.,1996).
PU.1 is a member of the Ets family of transcriptionfactors that plays an essential role in the development ofB cell and myeloid lineages (Scott et al., 1994; DeKoterand Singh, 2000). In B cells, PU.1 is expressed at allstages of maturation (excluding plasma cells) and itregulates B-cell-specific genes including the m heavychain (Nelsen et al., 1993; Rao et al., 1997), the Ig lightchains (l and k) (Pongubala et al., 1992; Eisenbeis et al.,1993), Iga (Feldhaus et al., 1992), Igb (Omori and Wall,1993), Bruton’s Tyrosine kinase (Btk) (Muller et al.,1996), and the Ig J chain (Shin and Koshland, 1993).Expression of PU.1 is regulated at the transcriptionallevel by the dual action of an Ets binding site and anoctamer motif (50-ATGCAAAT-30) (Chen et al., 1996).The Ets site binds PU.1, thus leading to an autoregu-latory stimulatory loop. The octamer site can be boundby either Oct-1 or Oct-2 transcription factors, with Oct-2 exhibiting a higher transactivation potential than Oct-1 (Chen et al., 1996).
Oct-1 and Oct-2 possess different patterns of expres-sion in vivo: whereas Oct-1 is ubiquitously expressed,Oct-2 is mainly restricted to the B lymphoid compart-ment, and it is involved in B-cell-specific gene expres-sion. The transactivation potential of Oct-1 andOct-2 is greatly enhanced by interactions with theB-cell-specific coactivator BOB-1/OcaB (Gstaiger et al.,1995; Strubin et al., 1995). Although knockout studieshave failed to show an essential role for either Oct-2(Corcoran et al., 1993) or BOB-1/OcaB (Schubartet al., 2001) in immunoglobulin gene expression, theOct-2�/�, BOB-1/OcaB�/�, or double knockout miceshowed defects in events that occur after B-cellactivation such as germinal center formation andmaintenance of the peripheral B-cell pool. Moreover,studies of somatic cell hybrids demonstrated thatB-cell-specific genes were coordinately silenced inmature B cells when fused to a non-B-cell line (Lieber-man et al., 1993). The extinction phenomenon wascorrelated with loss of Oct-2 expression and couldbe reversed by ectopic expression of Oct-2(Radomska et al., 1994; Sharif et al., 2001). Bothknockout mice and somatic cell hybrid studies thusascribe an essential role to Oct-2 in late-stage B-celldevelopment.
In the present study, we sought to examine theexpression profile of B-cell-specific transcription factorsin HHV-8-induced primary effusion lymphoma cells.Our results indicate that in B-cell lines derived fromPEL, transformation is associated with an overexpres-sion of IRF-4, as well as with the loss of expression ofPU.1,Oct-2, IRF-8/ICSBP, and BSAP/Pax5. Thesedramatic alterations in B-cell-specific transcriptionfactors may thus contribute to lymphomagenesis andto the development of the non-B, non-T phenotype inPEL cells.
Results
Constitutive expression of IRF-4 in PEL is concomitantwith a downregulation of PU.1
The transcription factor IRF-4 requires physical asso-ciation with PU.1 in order to efficiently bind andtransactivate target Ets-IRF sequences found in thelight-chain gene enhancers (Eisenbeis et al., 1995).Analysis of IRF-4 protein levels by immunoblotdemonstrated constitutive IRF-4 levels in all PEL celllines examined (BCBL-1, BC-3, and BCP-1), comparedto the control Burkitt’s lymphoma BJAB cells, whichshowed no significant level of expression (Figure 1a). Incontrast, no PU.1 protein could be detected in any of thePEL cell lines (Figure 1b). Surprisingly, the expressionpattern of IRF-8, the IRF family member most closelyrelated to IRF-4, was inversely related to IRF-4; that is,IRF-8 was expressed in BJAB, but not in PEL cells(Figure 1c). The levels of Spi-B, another Ets familymember that binds similar DNA elements and activatesmany of the same target genes as PU.1, remainedconstant in two of the three PEL cell lines compared toBJAB (Figure 1d).
To determine if the overexpression of IRF-4, togetherwith the abrogation of PU.1 expression in PEL cellsdisrupted the regulation of B-cell-specific genes, electro-phoretic mobility shift assay (EMSA) analysis wasperformed using the composite Ets-IRF element foundin the lB and kE30 light-chain promoters (Figure 2). Incontrol BJAB cells, a protein–DNA complex was easilyobserved using the lB or the kE30 probe (Figure 2,lane 1) and the protein–DNA complex was supershifted
BJAB
BCBL-1
BCP-1
BC-3COS-7
PU.1
actin
ba
IRF-4
actin
BJAB
BCBL-1
BC-3BCP-1
1 2 3 4 1 2 3 4 5
IRF-8/ICSBP
BJAB
BCBL-1
c
1 2
d BJAB
BCBL-1
BCP-1
BC-3
Spi-B
actin
1 2 3 4
Figure 1 Expression of IRF-4 and PU.1 in PEL cells. Immunoblotanalysis was performed using 100mg of whole-cell extracts (WCE)from PEL cell lines BCBL-1, BC-3, and BCP-1 as well as controlCos7 and BJAB cells, as described in Materials and methods.Immunoblots were probed with anti-IRF-4 (a), anti-PU.1 (b), anti-ICSBP/IRF-8 (c), and anti-Spi-B (d) antisera. In a, b, and d themembranes were reblotted with anti-b-actin antibody to verifyprotein loading homogeneity. These immunoblots are representa-tive of at least three independent experiments
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
965
Oncogene
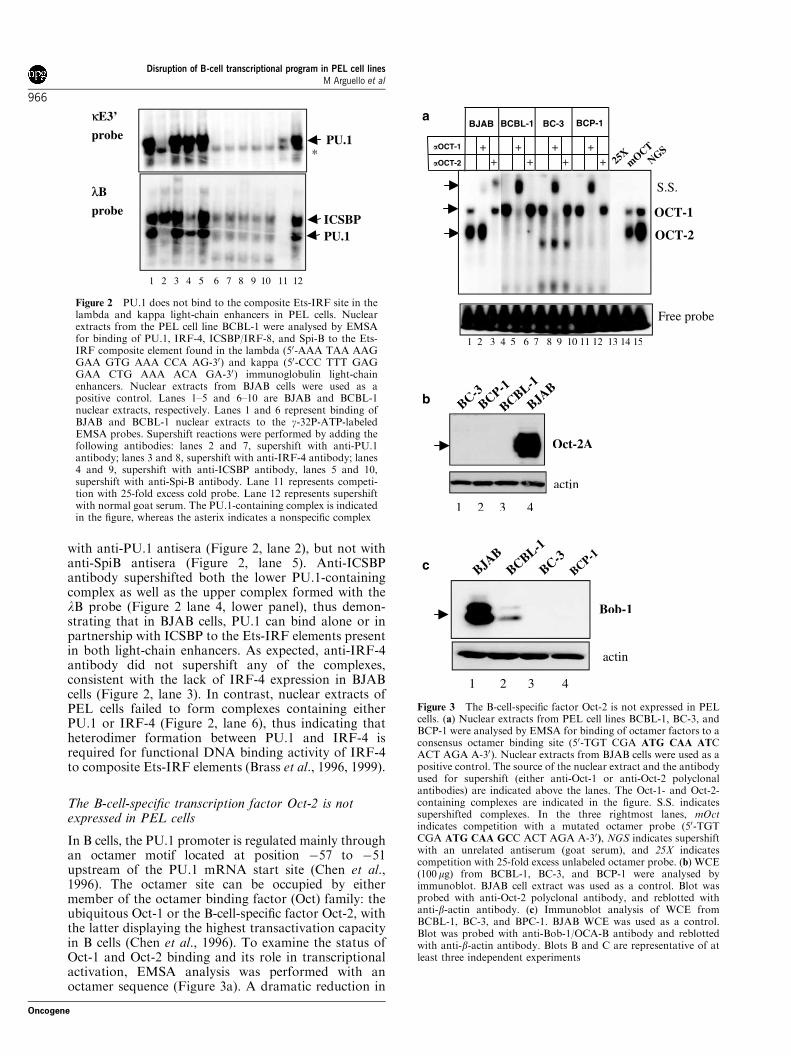
with anti-PU.1 antisera (Figure 2, lane 2), but not withanti-SpiB antisera (Figure 2, lane 5). Anti-ICSBPantibody supershifted both the lower PU.1-containingcomplex as well as the upper complex formed with thelB probe (Figure 2 lane 4, lower panel), thus demon-strating that in BJAB cells, PU.1 can bind alone or inpartnership with ICSBP to the Ets-IRF elements presentin both light-chain enhancers. As expected, anti-IRF-4antibody did not supershift any of the complexes,consistent with the lack of IRF-4 expression in BJABcells (Figure 2, lane 3). In contrast, nuclear extracts ofPEL cells failed to form complexes containing eitherPU.1 or IRF-4 (Figure 2, lane 6), thus indicating thatheterodimer formation between PU.1 and IRF-4 isrequired for functional DNA binding activity of IRF-4to composite Ets-IRF elements (Brass et al., 1996, 1999).
The B-cell-specific transcription factor Oct-2 is notexpressed in PEL cells
In B cells, the PU.1 promoter is regulated mainly throughan octamer motif located at position �57 to �51upstream of the PU.1 mRNA start site (Chen et al.,1996). The octamer site can be occupied by eithermember of the octamer binding factor (Oct) family: theubiquitous Oct-1 or the B-cell-specific factor Oct-2, withthe latter displaying the highest transactivation capacityin B cells (Chen et al., 1996). To examine the status ofOct-1 and Oct-2 binding and its role in transcriptionalactivation, EMSA analysis was performed with anoctamer sequence (Figure 3a). A dramatic reduction in
κκE3’
probe
probe
λB
PU.1
PU.1ICSBP
*
1 2 3 4 5 6 7 8 9 10 11 12
Figure 2 PU.1 does not bind to the composite Ets-IRF site in thelambda and kappa light-chain enhancers in PEL cells. Nuclearextracts from the PEL cell line BCBL-1 were analysed by EMSAfor binding of PU.1, IRF-4, ICSBP/IRF-8, and Spi-B to the Ets-IRF composite element found in the lambda (50-AAA TAA AAGGAA GTG AAA CCA AG-30) and kappa (50-CCC TTT GAGGAA CTG AAA ACA GA-30) immunoglobulin light-chainenhancers. Nuclear extracts from BJAB cells were used as apositive control. Lanes 1–5 and 6–10 are BJAB and BCBL-1nuclear extracts, respectively. Lanes 1 and 6 represent binding ofBJAB and BCBL-1 nuclear extracts to the g-32P-ATP-labeledEMSA probes. Supershift reactions were performed by adding thefollowing antibodies: lanes 2 and 7, supershift with anti-PU.1antibody; lanes 3 and 8, supershift with anti-IRF-4 antibody; lanes4 and 9, supershift with anti-ICSBP antibody, lanes 5 and 10,supershift with anti-Spi-B antibody. Lane 11 represents competi-tion with 25-fold excess cold probe. Lane 12 represents supershiftwith normal goat serum. The PU.1-containing complex is indicatedin the figure, whereas the asterix indicates a nonspecific complex
a
S.S.
25XmOCT
NGS
OCT-1
OCT-2
Free probe
++ + + ++ + + +
BJAB BCBL-1 BC-3 BCP-1
�OCT-1
�OCT-2
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
c BC-3BCP-1
BCBL-1
BJAB
Bob-1
actin
1 2 3 4
b BC-3BCP-1
BCBL-1
BJAB
Oct-2A
actin
1 2 3 4
Figure 3 The B-cell-specific factor Oct-2 is not expressed in PELcells. (a) Nuclear extracts from PEL cell lines BCBL-1, BC-3, andBCP-1 were analysed by EMSA for binding of octamer factors to aconsensus octamer binding site (50-TGT CGA ATG CAA ATCACT AGA A-30). Nuclear extracts from BJAB cells were used as apositive control. The source of the nuclear extract and the antibodyused for supershift (either anti-Oct-1 or anti-Oct-2 polyclonalantibodies) are indicated above the lanes. The Oct-1- and Oct-2-containing complexes are indicated in the figure. S.S. indicatessupershifted complexes. In the three rightmost lanes, mOctindicates competition with a mutated octamer probe (50-TGTCGA ATG CAA GCC ACT AGA A-30), NGS indicates supershiftwith an unrelated antiserum (goat serum), and 25X indicatescompetition with 25-fold excess unlabeled octamer probe. (b) WCE(100 mg) from BCBL-1, BC-3, and BCP-1 were analysed byimmunoblot. BJAB cell extract was used as a control. Blot wasprobed with anti-Oct-2 polyclonal antibody, and reblotted withanti-b-actin antibody. (c) Immunoblot analysis of WCE fromBCBL-1, BC-3, and BPC-1. BJAB WCE was used as a control.Blot was probed with anti-Bob-1/OCA-B antibody and reblottedwith anti-b-actin antibody. Blots B and C are representative of atleast three independent experiments
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
966
Oncogene
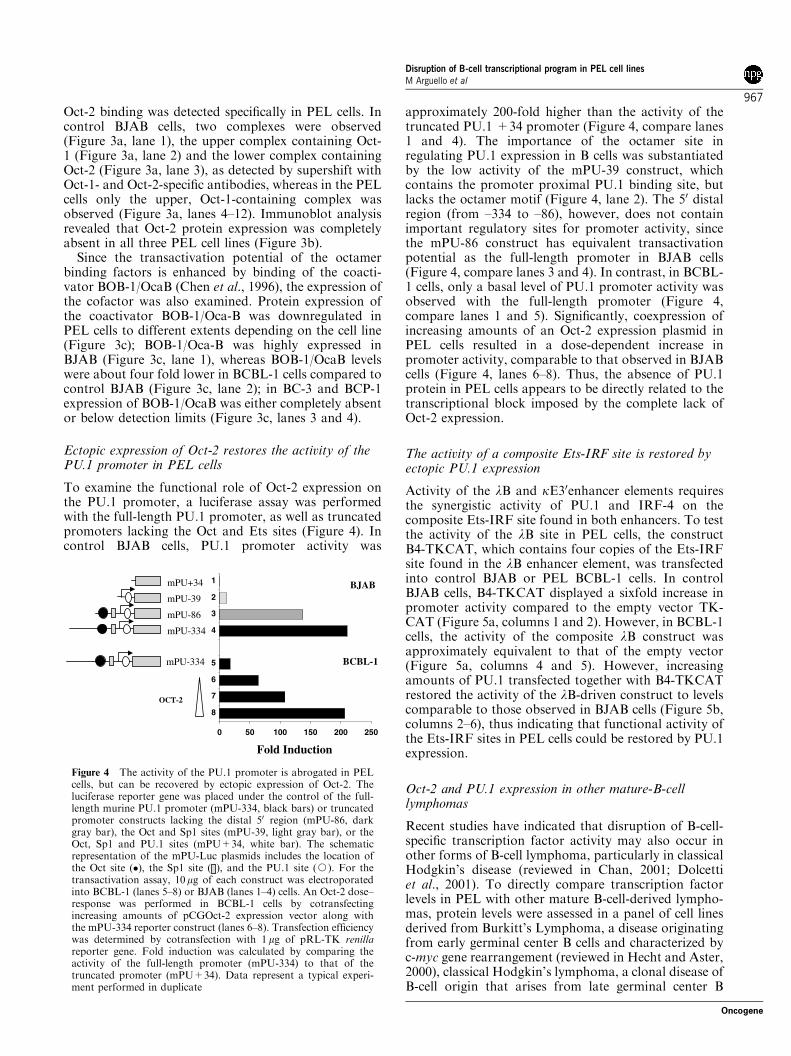
Oct-2 binding was detected specifically in PEL cells. Incontrol BJAB cells, two complexes were observed(Figure 3a, lane 1), the upper complex containing Oct-1 (Figure 3a, lane 2) and the lower complex containingOct-2 (Figure 3a, lane 3), as detected by supershift withOct-1- and Oct-2-specific antibodies, whereas in the PELcells only the upper, Oct-1-containing complex wasobserved (Figure 3a, lanes 4–12). Immunoblot analysisrevealed that Oct-2 protein expression was completelyabsent in all three PEL cell lines (Figure 3b).
Since the transactivation potential of the octamerbinding factors is enhanced by binding of the coacti-vator BOB-1/OcaB (Chen et al., 1996), the expression ofthe cofactor was also examined. Protein expression ofthe coactivator BOB-1/Oca-B was downregulated inPEL cells to different extents depending on the cell line(Figure 3c); BOB-1/Oca-B was highly expressed inBJAB (Figure 3c, lane 1), whereas BOB-1/OcaB levelswere about four fold lower in BCBL-1 cells compared tocontrol BJAB (Figure 3c, lane 2); in BC-3 and BCP-1expression of BOB-1/OcaB was either completely absentor below detection limits (Figure 3c, lanes 3 and 4).
Ectopic expression of Oct-2 restores the activity of thePU.1 promoter in PEL cells
To examine the functional role of Oct-2 expression onthe PU.1 promoter, a luciferase assay was performedwith the full-length PU.1 promoter, as well as truncatedpromoters lacking the Oct and Ets sites (Figure 4). Incontrol BJAB cells, PU.1 promoter activity was
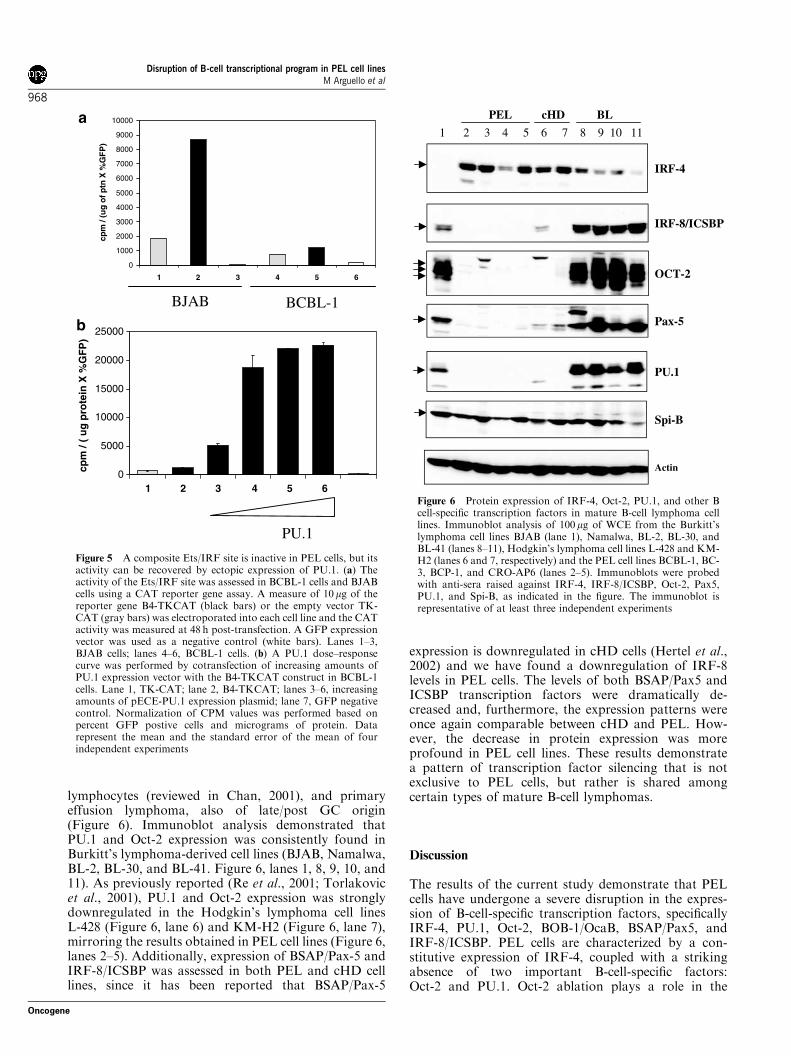
approximately 200-fold higher than the activity of thetruncated PU.1 +34 promoter (Figure 4, compare lanes1 and 4). The importance of the octamer site inregulating PU.1 expression in B cells was substantiatedby the low activity of the mPU-39 construct, whichcontains the promoter proximal PU.1 binding site, butlacks the octamer motif (Figure 4, lane 2). The 50 distalregion (from –334 to –86), however, does not containimportant regulatory sites for promoter activity, sincethe mPU-86 construct has equivalent transactivationpotential as the full-length promoter in BJAB cells(Figure 4, compare lanes 3 and 4). In contrast, in BCBL-1 cells, only a basal level of PU.1 promoter activity wasobserved with the full-length promoter (Figure 4,compare lanes 1 and 5). Significantly, coexpression ofincreasing amounts of an Oct-2 expression plasmid inPEL cells resulted in a dose-dependent increase inpromoter activity, comparable to that observed in BJABcells (Figure 4, lanes 6–8). Thus, the absence of PU.1protein in PEL cells appears to be directly related to thetranscriptional block imposed by the complete lack ofOct-2 expression.
The activity of a composite Ets-IRF site is restored byectopic PU.1 expression
Activity of the lB and kE30enhancer elements requiresthe synergistic activity of PU.1 and IRF-4 on thecomposite Ets-IRF site found in both enhancers. To testthe activity of the lB site in PEL cells, the constructB4-TKCAT, which contains four copies of the Ets-IRFsite found in the lB enhancer element, was transfectedinto control BJAB or PEL BCBL-1 cells. In controlBJAB cells, B4-TKCAT displayed a sixfold increase inpromoter activity compared to the empty vector TK-CAT (Figure 5a, columns 1 and 2). However, in BCBL-1cells, the activity of the composite lB construct wasapproximately equivalent to that of the empty vector(Figure 5a, columns 4 and 5). However, increasingamounts of PU.1 transfected together with B4-TKCATrestored the activity of the lB-driven construct to levelscomparable to those observed in BJAB cells (Figure 5b,columns 2–6), thus indicating that functional activity ofthe Ets-IRF sites in PEL cells could be restored by PU.1expression.
Oct-2 and PU.1 expression in other mature-B-celllymphomas
Recent studies have indicated that disruption of B-cell-specific transcription factor activity may also occur inother forms of B-cell lymphoma, particularly in classicalHodgkin’s disease (reviewed in Chan, 2001; Dolcettiet al., 2001). To directly compare transcription factorlevels in PEL with other mature B-cell-derived lympho-mas, protein levels were assessed in a panel of cell linesderived from Burkitt’s Lymphoma, a disease originatingfrom early germinal center B cells and characterized byc-myc gene rearrangement (reviewed in Hecht and Aster,2000), classical Hodgkin’s lymphoma, a clonal disease ofB-cell origin that arises from late germinal center B
OCT-2
0 50 100 150 200 250
8
7
6
5
4
3
2
1mPU+34
mPU-39
mPU-86
mPU-334
BJAB
BCBL-1mPU-334
Fold Induction
Figure 4 The activity of the PU.1 promoter is abrogated in PELcells, but can be recovered by ectopic expression of Oct-2. Theluciferase reporter gene was placed under the control of the full-length murine PU.1 promoter (mPU-334, black bars) or truncatedpromoter constructs lacking the distal 50 region (mPU-86, darkgray bar), the Oct and Sp1 sites (mPU-39, light gray bar), or theOct, Sp1 and PU.1 sites (mPU+34, white bar). The schematicrepresentation of the mPU-Luc plasmids includes the location ofthe Oct site (�), the Sp1 site ( ), and the PU.1 site (J). For thetransactivation assay, 10mg of each construct was electroporatedinto BCBL-1 (lanes 5–8) or BJAB (lanes 1–4) cells. An Oct-2 dose–response was performed in BCBL-1 cells by cotransfectingincreasing amounts of pCGOct-2 expression vector along withthe mPU-334 reporter construct (lanes 6–8). Transfection efficiencywas determined by cotransfection with 1 mg of pRL-TK renillareporter gene. Fold induction was calculated by comparing theactivity of the full-length promoter (mPU-334) to that of thetruncated promoter (mPU+34). Data represent a typical experi-ment performed in duplicate
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
967
Oncogene
lymphocytes (reviewed in Chan, 2001), and primaryeffusion lymphoma, also of late/post GC origin(Figure 6). Immunoblot analysis demonstrated thatPU.1 and Oct-2 expression was consistently found inBurkitt’s lymphoma-derived cell lines (BJAB, Namalwa,BL-2, BL-30, and BL-41. Figure 6, lanes 1, 8, 9, 10, and11). As previously reported (Re et al., 2001; Torlakovicet al., 2001), PU.1 and Oct-2 expression was stronglydownregulated in the Hodgkin’s lymphoma cell linesL-428 (Figure 6, lane 6) and KM-H2 (Figure 6, lane 7),mirroring the results obtained in PEL cell lines (Figure 6,lanes 2–5). Additionally, expression of BSAP/Pax-5 andIRF-8/ICSBP was assessed in both PEL and cHD celllines, since it has been reported that BSAP/Pax-5
expression is downregulated in cHD cells (Hertel et al.,2002) and we have found a downregulation of IRF-8levels in PEL cells. The levels of both BSAP/Pax5 andICSBP transcription factors were dramatically de-creased and, furthermore, the expression patterns wereonce again comparable between cHD and PEL. How-ever, the decrease in protein expression was moreprofound in PEL cell lines. These results demonstratea pattern of transcription factor silencing that is notexclusive to PEL cells, but rather is shared amongcertain types of mature B-cell lymphomas.
Discussion
The results of the current study demonstrate that PELcells have undergone a severe disruption in the expres-sion of B-cell-specific transcription factors, specificallyIRF-4, PU.1, Oct-2, BOB-1/OcaB, BSAP/Pax5, andIRF-8/ICSBP. PEL cells are characterized by a con-stitutive expression of IRF-4, coupled with a strikingabsence of two important B-cell-specific factors:Oct-2 and PU.1. Oct-2 ablation plays a role in the
a
b
0
5000
10000
15000
20000
25000
1 2 3 4 5 6
cpm
/ (
ug
pro
tein
X %
GF
P)
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
1 32 4 5 6
cpm
/ (u
g o
f ptn
X %
GF
P)
BJAB BCBL-1
PU.1
Figure 5 A composite Ets/IRF site is inactive in PEL cells, but itsactivity can be recovered by ectopic expression of PU.1. (a) Theactivity of the Ets/IRF site was assessed in BCBL-1 cells and BJABcells using a CAT reporter gene assay. A measure of 10mg of thereporter gene B4-TKCAT (black bars) or the empty vector TK-CAT (gray bars) was electroporated into each cell line and the CATactivity was measured at 48 h post-transfection. A GFP expressionvector was used as a negative control (white bars). Lanes 1–3,BJAB cells; lanes 4–6, BCBL-1 cells. (b) A PU.1 dose–responsecurve was performed by cotransfection of increasing amounts ofPU.1 expression vector with the B4-TKCAT construct in BCBL-1cells. Lane 1, TK-CAT; lane 2, B4-TKCAT; lanes 3–6, increasingamounts of pECE-PU.1 expression plasmid; lane 7, GFP negativecontrol. Normalization of CPM values was performed based onpercent GFP postive cells and micrograms of protein. Datarepresent the mean and the standard error of the mean of fourindependent experiments
1 2 3 4 5 6 7 8 9 10 11
IRF-4
IRF-8/ICSBP
OCT-2
Pax-5
PU.1
Spi-B
PEL cHD BL
Actin
Figure 6 Protein expression of IRF-4, Oct-2, PU.1, and other Bcell-specific transcription factors in mature B-cell lymphoma celllines. Immunoblot analysis of 100mg of WCE from the Burkitt’slymphoma cell lines BJAB (lane 1), Namalwa, BL-2, BL-30, andBL-41 (lanes 8–11), Hodgkin’s lymphoma cell lines L-428 and KM-H2 (lanes 6 and 7, respectively) and the PEL cell lines BCBL-1, BC-3, BCP-1, and CRO-AP6 (lanes 2–5). Immunoblots were probedwith anti-sera raised against IRF-4, IRF-8/ICSBP, Oct-2, Pax5,PU.1, and Spi-B, as indicated in the figure. The immunoblot isrepresentative of at least three independent experiments
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
968
Oncogene

downregulation of PU.1 expression in PEL cells, whichin turn leads to an impaired activity of PU.1 responsiveelements such as the Ets-IRF sites found in the lB andkE30 immunoglobulin (Ig) light-chain enhancers and theCD20 promoter. Ectopic expression of PU.1 in BCBL-1cells fully restored the activity of the composite Ets-IRFsite, indicating that it is the absence of PU.1 thataccounts for the loss of Ig light-chain enhancer activityin PEL cells.
PU.1 expression is controlled in part by an octamersite located in the �57 to �51 region of the PU.1promoter and composed of the sequence 50-ATG-CAAAC-30. This site is activated preferentially by thelymphoid-specific factor Oct-2, with the activity beingfurther enhanced by interaction with the coactivatorBOB-1/Oca-B. Mature, activated B cells express highlevels of Oct-2 and BOB-1 (Re et al., 2001). In PEL cells,however, Oct-2 expression was completely abrogated,although Oct-1 levels were normal. As a consequence ofthe loss of Oct-2, the activity of a reporter gene underthe control of the PU.1 promoter was negligible inBCBL-1 cells. Interestingly, transient expression of Oct-2 reconstituted PU.1 promoter activity to control levels,whereas endogenous Oct-1 was unable to restore PU.1promoter activity and thus could not functionallyreplace Oct-2. BCBL-1 is the only PEL cell line thatretained BOB-1/OcaB expression, albeit at significantlyreduced levels. It is possible that partial expression ofthe coactivator could explain why recovery of the PU.1promoter activity was observed when Oct-2 activity wasreconstituted in BCBL-1 cells. Our studies thus suggestthat it is the loss of the B-cell-specific Oct-2 protein thatcontributes to the transcriptional shutoff of PU.1. Incontrast to the dramatic alterations in IRF-4, Oct-2, andPU.1, expression of the Ets family member Spi-B wasgenerally not altered in PEL cell lines. Although it hasbeen reported that Spi-B is able to bind to the sameDNA motifs as PU.1, in our hands endogenous Spi-Bwas unable to replace PU.1 in the transactivation of thelB element. In recent experiments, IRF-4 was alsoshown to be constitutively expressed in HTLV-1-infected leukemia T cells (Sharma et al., 2002).Furthermore, in the T-cell context, IRF-4 was shownto act as a global transcriptional repressor of genesinvolved in cell cycle regulation, apoptosis, DNA repairand cell adhesion (Mamane et al., 2002a). In the B-cellcontext, IRF-4 acts as a strong transcriptional activatorof downstream target genes by virtue of its associationwith PU.1. In the absence of its transcriptional partner,it will be interesting to determine if IRF-4 also acts as arepressor of B-cell-specific transcription in PEL andcHD.
Among the genes downregulated when Oct-2 is absentin PEL cells are PU.1 and BOB-1/OcaB, two factors thatare key for B-cell development, maturation, andsurvival. Once deprived of Oct-2, BOB-1/OcaB, andPU.1 expression, PEL cells may lose the ability toexpress surface molecules in a silencing-like event, suchas previously observed in hybrid B cells (Liebermanet al., 1993; Radomska et al., 1994). Oct-2 haspreviously been shown to regulate the expression of an
array of B-cell-specific genes, as demonstrated in hybridB cells, fused either to myeloma or T cells (Junker et al.,1990; Radomska et al., 1994). Fusion of an Ig-secretingcell (plasmacytoma) to a T-cell lymphoma led tocessation Oct-2 production as well as immunoglobulin,J chain, and other B-cell-specific genes. However, whenOct-2 extinction was prevented through stable transfec-tion of an Oct-2 expression plasmid, all B-cell-specificgenes escaped silencing, including the endogenous Oct-2. Interestingly, endogenous or exogenous Oct-1 couldnot rescue the cells from B-cell-specific gene silencing(Radomska et al., 1994; Sharif et al., 2001), a result thatis consistent with the present study. We are currentlyinvestigating the role of PU.1 and Oct-2 abrogation inthe expression of other B-cell molecules such as the mand d heavy chains, the J chain, and CD20, in the hopeof elucidating the role of these two factors in the‘incomplete’ phenotype observed in PEL cells.
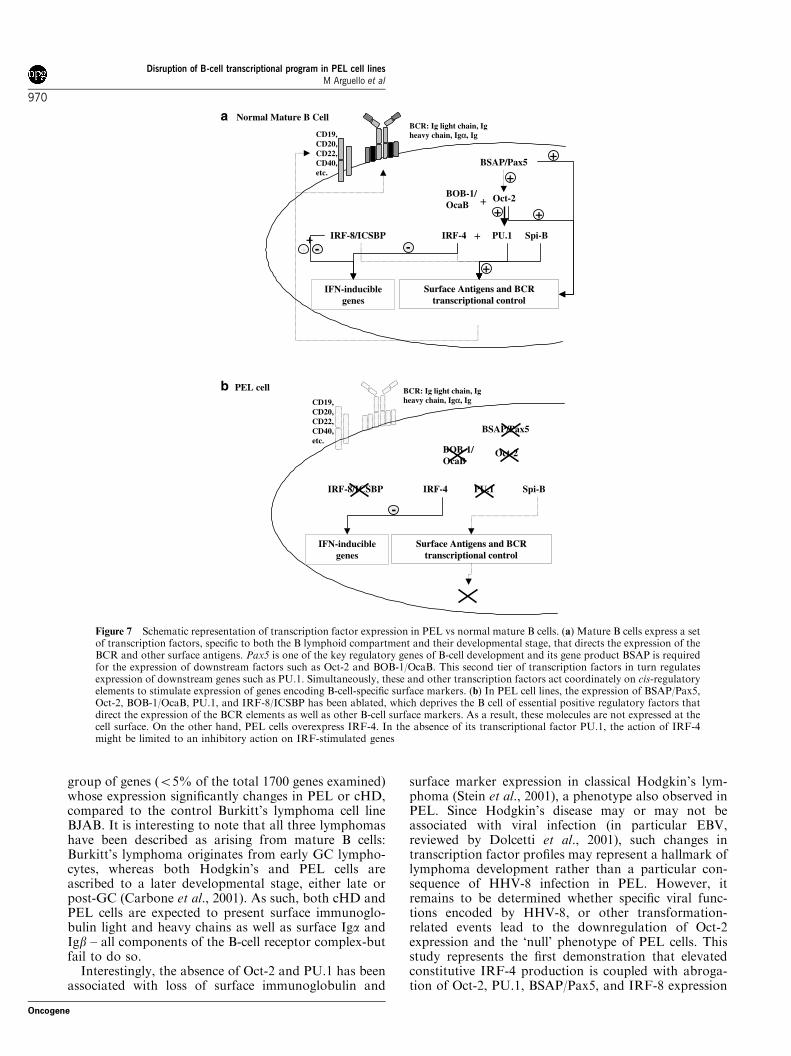
The results of this study together with other recentexperimentation (Re et al., 2001; Stein et al., 2001;Torlakovic et al., 2001; Hertel et al., 2002) provide apreliminary model describing B-cell transcription factorablation in mature B-cell lymphomas, and is representedschematically in Figure 7; in normal mature B cells,expression of surface antigens is regulated both byextracellular signals and by developmentally regulatedtranscription factors. These transcription factors actcoordinately to direct the expression of B-cell surfaceantigens, and in particular, B-cell receptor (BCR)components (Figure 7a). In PEL cells, however,abrogation of the expression of essential B-cell tran-scription factors such as BSAP/Pax5, Oct-2, and BOB-1/OcaB by as yet uncharacterized mechanisms, leads tosilencing of the expression of downstream factors (forexample PU.1, a gene under the transcriptional controlof Oct-2 and BOB-1/OcaB) in a cascade-like event. Theablation of Oct-2, BSAP/Pax5, BOB-1/OcaB, PU.1, andIRF-8 expression is a potential cause of the lack of BCRas well as other B-cell surface markers observed in PELcells (Figure 7b).
A limited survey of the expression patterns of Oct-2,PU.1, IRF-4, IRF-8, and BSAP/Pax5 in PEL and othermature B-cell-derived lymphomas demonstrated thatclassical Hodgkin’s disease (cHD) cells have a transcrip-tion factor profile very similar to that of PEL, withincreased levels of IRF-4 and reduced or ablatedexpression of the remaining transcription factors: Oct-2, PU.1, IRF-8, and BSAP/Pax5. In contrast, the profileof all Burkitt’s lymphoma cell lines was exactly oppositeto that of PEL cells. The direct comparison of two ormore cell lines derived from different lymphomas raisesthe question as to whether the observed changes arespecific to the phenotypic characteristics of the diseaseor if they represent global differences caused by cell-lineskewing. To address this question, microarray studieswere conducted using a human array containing 1700genes to compare gene expression between the PEL cellline BCBL-1, the cHD cell line L-428, and Burkitt’slymphoma cell line BJAB (data not shown). Preliminaryresults indicated that transcription factors Oct-2, PU.1,IRF-4, IRF-8, and BSAP/Pax5 belong to a limited
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
969
Oncogene
group of genes (o5% of the total 1700 genes examined)whose expression significantly changes in PEL or cHD,compared to the control Burkitt’s lymphoma cell lineBJAB. It is interesting to note that all three lymphomashave been described as arising from mature B cells:Burkitt’s lymphoma originates from early GC lympho-cytes, whereas both Hodgkin’s and PEL cells areascribed to a later developmental stage, either late orpost-GC (Carbone et al., 2001). As such, both cHD andPEL cells are expected to present surface immunoglo-bulin light and heavy chains as well as surface Iga andIgb – all components of the B-cell receptor complex-butfail to do so.
Interestingly, the absence of Oct-2 and PU.1 has beenassociated with loss of surface immunoglobulin and
surface marker expression in classical Hodgkin’s lym-phoma (Stein et al., 2001), a phenotype also observed inPEL. Since Hodgkin’s disease may or may not beassociated with viral infection (in particular EBV,reviewed by Dolcetti et al., 2001), such changes intranscription factor profiles may represent a hallmark oflymphoma development rather than a particular con-sequence of HHV-8 infection in PEL. However, itremains to be determined whether specific viral func-tions encoded by HHV-8, or other transformation-related events lead to the downregulation of Oct-2expression and the ‘null’ phenotype of PEL cells. Thisstudy represents the first demonstration that elevatedconstitutive IRF-4 production is coupled with abroga-tion of Oct-2, PU.1, BSAP/Pax5, and IRF-8 expression
BSAP/Pax5
Oct-2
PU.1IRF-4IRF-8/ICSBP
BOB-1/ OcaB
Surface Antigens and BCR Surface Antigens and BCR transcriptional controltranscriptional control
CD19, CD20, CD22, CD40, etc.
BCR: Ig light chain, Ig heavy chain, Igα, Ig
+
Spi-B+
IFN-inducible genes
++
--
a
b
++
++++
++
++
--
BSAP/Pax5
Oct-2
PU.1IRF-4IRF-8/ICSBP
BOB-1/ OcaB
Surface Antigens and BCR Surface Antigens and BCR transcriptional controltranscriptional control
CD19, CD20, CD22, CD40, etc.
BCR: Ig light chain, Ig heavy chain, Igα, Ig
Spi-B
IFN-inducible genes
--
Normal Mature B Cell
PEL cell
Figure 7 Schematic representation of transcription factor expression in PEL vs normal mature B cells. (a) Mature B cells express a setof transcription factors, specific to both the B lymphoid compartment and their developmental stage, that directs the expression of theBCR and other surface antigens. Pax5 is one of the key regulatory genes of B-cell development and its gene product BSAP is requiredfor the expression of downstream factors such as Oct-2 and BOB-1/OcaB. This second tier of transcription factors in turn regulatesexpression of downstream genes such as PU.1. Simultaneously, these and other transcription factors act coordinately on cis-regulatoryelements to stimulate expression of genes encoding B-cell-specific surface markers. (b) In PEL cell lines, the expression of BSAP/Pax5,Oct-2, BOB-1/OcaB, PU.1, and IRF-8/ICSBP has been ablated, which deprives the B cell of essential positive regulatory factors thatdirect the expression of the BCR elements as well as other B-cell surface markers. As a result, these molecules are not expressed at thecell surface. On the other hand, PEL cells overexpress IRF-4. In the absence of its transcriptional factor PU.1, the action of IRF-4might be limited to an inhibitory action on IRF-stimulated genes
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
970
Oncogene

in PEL cells, and provides insight into the moleculartranscriptional changes that occur in Primary EffusionLymphoma. Altogether, this work and other recentstudies suggest that transcription factor ablation may bea common theme in the development of certain subtypesof late-stage B-cell lymphoma.
Materials and methods
Cell lines
BCBL-1, a primary effusion lymphoma cell line infected withHHV-8 but negative for EBV or HIV-1, and BJAB, an EBV-negative Burkitt’s lymphoma cell line, were a kind gift from DrJae U Jung (Harvard Medical School, MA, USA). Both celllines were cultured in RPMI 1640 (Wisent) supplemented with10% FBS, 10�5
m b-mercaptoethanol, and 8mg/ml gentamicin.BC-3 (ATCC no. CRL-2277) is a primary effusion lymphomacell line infected with HHV-8, but negative for both HIV andEBV. BCP-1 (ATCC product number CRL-2294) is an HHV-8positive, HIV and EBV negative, clonal lymphoma cell lineisolated from peripheral blood. Both PEL cell lines weremaintained in RPMI 1640 supplemented with 20% FBS and8mg/ml gentamicin. CRO-AP6 is an HIV-positive, EBV-negative PEL cell line established from the pleural effusionof a 26-year-old, HIV+ patient (described in Carbone et al.,2000a, a kind gift from Dr A Carbone, Centro di ReferimentoOncologico, IRCCS, Italy). KM-H2 and L-428 are two well-characterized Hodgkin’s lymphoma cell lines of B-cell origin(described in Drexler, 1993; Hertel et al., 2002), BL-2, BL-30,and BL-41 are Burkitt’s lymphoma cell lines previouslydescribed (Ehlin-Henriksson et al., 1987), all a kind gift fromDr Sigrun Smola. Namalwa cells are an EBV-positive Burkitt’slymphoma cell line obtained from the ATCC (product numberCRL-1432). They were cultured in RPMI 1640 with 10% FBSand 1% sodium pyruvate.
Plasmid constructions
The B4-TKCAT reporter construct was a kind gift fromDr H Singh (University of Chicago, Eisenbeis et al, 1993).B4-TKCAT contains four copies of the immunoglobulin lBsite subcloned upstream of the HSV thymidine kinasepromoter in the TK-CAT vector. The PU.1 expression vectorPU.1/pECE was a kind gift from Dr M Fenton. The PU.1cDNA is placed under the transcriptional control of the SV40early promoter. The luciferase reporters under the control ofthe murine full-length PU.1 promoter (mPU-334) and dele-tions (mPU-86, mPU-39, and mPU+34) as well as the Oct-2and Oct-1 expression vectors (pCGOct-1 and pCGOct-2) werea kind gift from Dr D Tenen (Harvard University) and aredescribed in Chen et al. (1996).
Whole-cell extract preparation and immunoblot analysis
Whole-cell extracts (WCE) and immunoblot analysis wereperformed using standard protocols as described (Servantet al., 2001). Briefly, WCE (50–100 mg) prepared using NonidetP-40 lysis buffer were subjected to electrophoresis on 10–12%SDS–PAGE gels. Proteins transferred to Trans-Blots nitro-cellulose membrane (Bio-Rad Laboratories) were blocked inPBS containing 5% nonfat dry milk and 0.05% Tween 20 thenincubated with 1mg/ml primary antibody dilutions. Afterwashes, the blots were incubated with a secondary antibodycoupled to HRP. Blots were visualized using the ECLdetection system from NEN Life Science Products. The
following antibodies were used in this study: Anti-IRF-4 goatpolyclonal antibody, anti-ICSBP goat polyclonal antibody,anti-PU.1 rabbit polyclonal antibody, anti-Oct-2 rabbit poly-clonal antibody, anti-Bob-1 rabbit polyclonal antibody, anti-Spi-B goat polyclonal antibody, anti-Pax5 goat polyclonalantibody, all from Santa Cruz Biotechnology (Santa Cruz,CA, USA). Anti-b-actin mouse monoclonal antibody waspurchased from Sigma and used at a 1 : 10 000 dilution.
Transient transfections and reporter assays
Electroporation was performed at 950 mF and 250 V for bothBJAB and BCBL-1 cells. Luciferase assays were performedfollowing a transient transfection of 10 mg of the reporter gene(mPU-334, mPU-86, mPU-39, or mPU+34) and 1.0 mg of therenilla internal control (pRL-TK) into 10� 106 cells. The dose–response for Oct-2 was performed in a similar manner, butcotransfecting 0, 1, 2, or 4mg of the pCGOct-2 expressionvector (in which the octamer factor cDNA is under the controlof a CMV promoter). The luciferase assay was performed 48 hafter transfection using the Promega Dual Luciferase ReporterAssay System. Transfection efficiency was normalized usingthe renilla activity counts.
CAT assays were performed after transient transfection of10 mg of the reporter constructs B4-TKCAT, TK-CAT, orpEGFP-C1 into 10� 106 cells. PU.1 dose–response wasperformed by cotransfecting 0, 10, 50, 150, or 300 ng of thePU.1/pECE expression vector. The CAT assay was performed48 h after transfection as follows: cell pellet was resuspended in0.25m Tris pH 7.8 and lysed by three rounds of freeze–thaw.Equal volumes of protein extract (50 ml) were placed in a 7mlglass scintillation vial and heated at 601C for 10min todeactivate deacetylases, then cooled on ice. To the extractswere added 1mm chloroamphenicol (Sigma) and 0.4 nCi of 3H-acetyl-coenzyme A (200.00mCi/mmol, Perkin Elmer LifeSciences) in a total volume of 250ml buffered with 100 mm
Tris pH 7.8. The mixture was overlaid with Beta Max waterimmiscible scintillation fluid (ICN Radiochemicals) andincubated at 371C for 6 h. The samples were then analysed ina scintillation counter for 3H measurement. Transfectionefficiency was normalized by %GFP and microgram ofprotein.
Electrophoretic mobility shift assay (EMSA)
BJAB, BCBL-1, BC-3, and BCP-1 nuclear extracts wereprepared as described elsewhere (Osborn et al., 1989). EMSAanalysis using the octamer consensus sequence was performedas described in Chen et al. (1996). Briefly, 2� 105 cpm of the[g-32P]ATP consensus Octamer probe (50-TGT CGA ATG
CAA ATC ACT AGA A-30, Sta. Cruz Biotechnology) wasincubated with 5–10 mg of nuclear extracts in binding buffer(10 mm HEPES, pH 7.5, 50mm KCl, 5mm MgCl2, 1 mm
EDTA, 1 mm dithiothreitol, 0.1mg/ml poly(dI/dC), 0.1mg/ml BSA, and 5% glycerol) on ice for 30min. For supershiftexperiments, 1ml of anti-Oct-2 antibody (Santa Cruz Biotech-nology) or anti-Oct-1 antibody (Sta. Cruz Biotechnology) wasadded to the reaction. Competition with mutant probe wasperformed using 1 ml of a mutated octamer probe (50-TGTCGA ATG CAA GCC ACT AGA A-30, Santa CruzBiotechnology). Supershift with an unrelated antibody wasperformed by incubation with 1 ml of normal goat serum.Binding reactions were subjected to electrophoresis on a 5%polyacrylamide gel in 0.25�TBE at 160V for 3 h.
Electromobility shift assays using the lB (50-AAA TAAAAG GAA GTG AAA CCA AG-30) and kE30 (50-CCC TTTGAG GAA CTG AAA ACA GA-30) probes were performed
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
971
Oncogene

as described above. For supershift experiments, 5 ml of anti-IRF-4 antibody, anti-PU.1 antibody, anti-ICSBP antibody, oranti-Spi-B antibody (Santa Cruz Biotechnology) was added tothe reaction.
AcknowledgmentsWe gratefully acknowledge gifts of Oct-1 and Oct-2 expressionvectors as well as murine PU.1 promoter luciferase reporterconstructs from Dr Daniel G Tenen (Harvard University); theB4-TKCAT reporter construct from Dr Harinder Singh
(University of Chicago) as well as the PU.1 expression vectorfrom Dr Matthew Fenton (Boston University). We extendour gratitude to Dr Antonino Carbone for his kind gift ofthe PEL cell line CRO-AP6 and to Dr Sigrun Smola for herkind gift of Burkitt’s lymphoma and Hodgking’s lymphomacell lines used in this study. Many thanks to Maria Ricciand to the members of the Molecular Oncology Group,Lady Davis Institute, McGill University, for their helpfuldiscussion and technical assistance. This work was supportedby Grants MOP42562 from the Canadian Institutes of HealthResearch.
References
Brass AL, Kehrli E, Eisenbeis CF, Storb U and Singh H.(1996). Genes Dev., 10, 2335–2347.
Brass AL, Zhu AQ and Singh H. (1999). EMBO J., 18,977–991.
Carbone A, Cilia AM, Gloghini A, Capello D, Fassone L,Perin T, Rossi D, Canzonieri V, De Paoli P, Vaccher E,Tirelli U, Volpe R and Gaidano G. (2000a). Leukemia, 14,1301–1309.
Carbone A, Gloghini A, Cozzi MR, Capello D, Steffan A,Monini P, De Marco L and Gaidano G. (2000b). Br. J.Haematol., 111, 247–257.
Carbone A, Gloghini A, Larocca LM, Capello D, Pierconti F,Canzonieri V, Tirelli U, Dalla-Favera R and Gaidano G.(2001). Blood, 97, 744–751.
Cesarman E, Chang Y, Moore PS, Said JW and Knowles DM.(1995). N. Engl. J. Med., 332, 1186–1191.
Chan WC. (2001). Hematol. Oncol., 19, 1–17.Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J,
Knowles DM and Moore PS. (1994). Science, 266, 1865–1869.
Chen H, Zhang P, Radomska HS, Hetherington CJ, ZhangDE and Tenen DG. (1996). J. Biol. Chem., 271, 15743–15752.
Corcoran LM, Karvelas M, Nossal GJ, Ye ZS, Jacks T andBaltimore D. (1993). Genes Dev., 7, 570–582.
DeKoter RP and Singh H. (2000). Science, 288, 1439–1441.Dolcetti R, Boiocchi M, Gloghini A and Carbone A. (2001).
Eur. J. Cancer, 37, 1276–1287.Drexler HG. (1993). Leuk. Lymphoma, 9, 1–25.Drexler HG, Uphoff CC, Gaidano G and Carbone A. (1998).
Leukemia, 12, 1507–1517.Ehlin-Henriksson B, Manneborg-Sandlund A and Klein G.
(1987). Int. J. Cancer, 39, 211–218.Eisenbeis CF, Singh H and Storb U. (1993). Mol. Cell Biol.,13, 6452–6461.
Eisenbeis CF, Singh H and Storb U. (1995). Genes Dev., 9,1377–1387.
Falini B, Fizzotti M, Pucciarini A, Bigerna B, Marafioti T,Gambacorta M, Pacini R, Alunni C, Natali-Tanci L,Ugolini B, Sebastiani C, Cattoretti G, Pileri S, Dalla-FaveraR and Stein H. (2000). Blood, 95, 2084–2092.
Feldhaus AL, Mbangkollo D, Arvin KL, Klug CA and SinghH. (1992). Mol. Cell Biol., 12, 1126–1133.
Gaidano G, Carbone A, Pastore C, Capello D, Migliazza A,Gloghini A, Roncella S, Ferrarini M, Saglio G and Dalla-Favera R. (1997). Blood, 89, 3755–3762.
Gstaiger M, Knoepfel L, Georgiev O, Schaffner W andHovens CM. (1995). Nature, 373, 360–362.
Hecht JL and Aster JC. (2000). J. Clin. Oncol., 18, 37073721.
Hertel CB, Zhou XG, Hamilton-Dutoit SJ and Junker S.(2002). Oncogene, 21, 4908–4920.
Himmelmann A, Riva A, Wilson GL, Lucas BP, Thevenin Cand Kehrl JH. (1997). Blood, 90, 3984–3995.
Junker S, Pedersen S, Schreiber E and Matthias P. (1990). Cell,61, 467–474.
Lieberman SA, Hines MD, Bergsagel PL, Kuehl WM andEckhardt LA. (1993). J. Immunol., 151, 2588–2600.
Mamane Y, Grandvaux N, Hernandez E, Sharma S, InnocenteSA, Lee JM, Azimi N, Lin R and Hiscott J. (2002a).Oncogene, 21, 6751–6765.
Mamane Y, Sharma S, Grandvaux N, Hernandez E andHiscott J. (2002b). J. Interferon Cytokine Res., 22,
135–143.Marecki S and Fenton MJ. (2002). J. Interferon Cytokine Res.,22, 121–133.
Matolcsy A, Nador RG, Cesarman E and Knowles DM.(1998). Am. J. Pathol., 153, 1609–1614.
Muller S, Sideras P, Smith CI and Xanthopoulos KG. (1996).Oncogene, 13, 1955–1964.
Nador RG, Cesarman E, Chadburn A, Dawson DB,Ansari MQ, Sald J and Knowles DM. (1996). Blood, 88,645–656.
Nelsen B, Tian G, Erman B, Gregoire J, Maki R, Graves Band Sen R. (1993). Science, 261, 82–86.
Omori SA and Wall R. (1993). Proc. Natl. Acad. Sci. USA, 90,11723–11727.
Osborn L, Kunkel S and Nabel GJ. (1989). Proc. Natl. Acad.Sci. USA, 86, 2336–2340.
Pernis AB. (2002). J. Interferon Cytokine Res., 22, 111–120.
Pongubala JM, Nagulapalli S, Klemsz MJ, McKercher SR,Maki RA and Atchison ML. (1992). Mol. Cell Biol., 12, 368–378.
Radomska HS, Shen CP, Kadesch T and Eckhardt LA. (1994).Immunity, 1, 623–634.
Rao E, Dang W, Tian G and Sen R. (1997). J. Biol. Chem.,272, 6722–6732.
Re D, Muschen M, Ahmadi T, Wickenhauser C, Staratschek-Jox A, Holtick U, Diehl V and Wolf J. (2001). Cancer Res.,61, 2080–2084.
Reya T and Grosschedl R. (1998). Curr. Opin. Immunol., 10,158–165.
Schubart K, Massa S, Schubart D, Corcoran LM, Rolink AGand Matthias P. (2001). Nat. Immunol., 2, 69–74.
Scott EW, Simon MC, Anastasi J and Singh H. (1994).Science, 265, 1573–1577.
Servant MJ, ten Oever B, LePage C, Conti L, Gessani S,Julkunen I, Lin R and Hiscott J. (2001). J. Biol. Chem., 276,355–363.
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
972
Oncogene

Sharif MN, Radomska HS, Miller DM and Eckhardt LA.(2001). J. Immunol., 167, 4421–4429.
Sharma S, Grandvaux N, Mamane Y, Genin P, Azimi N,Waldmann T and Hiscott J. (2002). J. Immunol., 169, 3120–3130.
Shin MK and Koshland ME. (1993). Genes Dev., 7, 2006–2015.
Singh H. (1994). Immunol. Res., 13, 280–290.
Stein H, Marafioti T, Foss HD, Laumen H, Hummel M,Anagnostopoulos I, Wirth T, Demel G and Falini B. (2001).Blood, 97, 496–501.
Strubin M, Newell JW and Matthias P. (1995). Cell, 80, 497–506.
Torlakovic E, Tierens A, Dang HD and Delabie J. (2001). Am.J. Pathol., 159, 1807–1814.
Disruption of B-cell transcriptional program in PEL cell linesM Arguello et al
973
Oncogene