differential responses to distinct psychotropic agents of selectively bred dominant and submissive...
TRANSCRIPT

R
Dd
Ea
b
h
����
a
ARRAA
KDSSAMA
1
finebbplva
C
0h
Behavioural Brain Research 236 (2013) 225– 235
Contents lists available at SciVerse ScienceDirect
Behavioural Brain Research
j ourna l ho mepage: www.elsev ier .com/ locate /bbr
esearch report
ifferential responses to distinct psychotropic agents of selectively bredominant and submissive animals
limelech Neshera,b, Moshe Grossa,b, Serah Lissona,b, Tatiana Tikhonova, Gal Yadidb, Albert Pinhasova,∗
Department of Molecular Biology, Ariel University Center, Ariel, IsraelFaculty of Life Sciences, Bar Ilan University, Ramat Gan, Israel
i g h l i g h t s
Using selective breeding, we developed mice with strong and stable dominant and submissive traits.These two unique animal populations have differential sensitivity to the antidepressant paroxetine.Lithium selectively altered behavior of dominant mice, without altering the same parameters of submissive animals.Diazepam induced paradoxical, stimulatory and anxiogenic effects upon dominant animals.
r t i c l e i n f o
rticle history:eceived 10 June 2012eceived in revised form 24 August 2012ccepted 26 August 2012vailable online xxx
eywords:ominanceubmissivenessocial interaction
a b s t r a c t
Dominance and submissiveness are two opposite poles of behavior representing important functional ele-ments in the development of social interactions. We previously demonstrated the inheritability of thesetraits by selective breeding based upon the dominant–submissive relationships (DSR) food competitionparadigm. Continued multigenerational behavioral selection of Sabra mice yielded animal populationswith strong and stable features of dominance and submissiveness.
We found that these animals react differentially to stressogenic triggers, antidepressants and moodstabilizing agents. The anxiolytic compound diazepam (1.5 mg/kg, i.p.) reduced anxiety-like behavior ofsubmissive animals, but showed anxiogenic effects among dominant animals. In the Forced Swim test, theantidepressant paroxetine (1, 3 and 10 mg/kg, i.p.) markedly reduced immobility of submissive animals,
nxiolyticood stabilizer
ntidepressant
demonstrating antidepressant-like effect. In contrast, when administered to dominant animals, parox-etine caused extreme (frenetic) activity. The mood stabilizer lithium (0.4%, p.o.) selectively influenceddominant mice, without affecting the behavior of submissive animals.
In summary, we describe here two distinct animal populations possessing strong dominant and sub-missive phenotypes. We suggest that these populations hold potential as tools for studying the molecular
ics of
basis and pharmacogenet. Introduction
Social hierarchy is common to many animal phyla such assh, reptiles, birds and mammals, including humans [1–3]. Domi-ance and submissiveness are important functional elements in thestablishment of social hierarchy, situated at opposite poles of theehavioral spectrum [4,5]. In the case of two or more individualsoth broadly classified as dominant or submissive, one of them willrove to be more dominant or more submissive, such that in the
inear hierarchy characterizing a group of social animals, one indi-idual dominates the others, a second dominates all but the first,nd so on, down to the last individual who is dominated by all the
∗ Corresponding author at: Department of Molecular Biology, Ariel Universityenter of Samaria, Ariel 40700, Israel. Tel.: +972 3 9371480; fax: +972 3 9371422.
E-mail address: [email protected] (A. Pinhasov).
166-4328/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.bbr.2012.08.040
dominant and submissive behavior.© 2012 Elsevier B.V. All rights reserved.
others [6]. Blanchard et al. [7] showed that dominant–submissivebehavior in rats develops a few days after grouping and remainslargely unchanged during their lifetime. While dominance and sub-missiveness are generally regarded as adaptive behaviors whichcontribute to maintenance of the social structure, disrupted hierar-chal status formation or its transformation have been implicated insocial dysfunction predicating pathological conditions [8]. Amonghumans, victims of bullying at school or in the workplace havea greater likelihood of developing depression or anxiety, whileanimal studies found that social defeat led to the expression ofdepressive-like behavior [1–3]. Subordinate animals, similarly todepressed humans, show increased defensive behavior, weight lossas well as major alterations in sleep, eating and activity, as well as
elevated glucocorticoid levels [7,9,10].Relationships of dominance and submissiveness can be exac-erbated by scarcity of food, water, territory or sexual partners, andcan be measured by the priority of access to these resources. In such

2 rain R
ecaniifb
sdiiabmat
asaai[frtobaadtb
sodombsucl
2
2
wiwabddiDdog
2
(d
26 E. Nesher et al. / Behavioural B
xperiments, dominance and submissiveness can be quantified byompetitive tests [11,12], in which animals are given restrictedccess to a desired resource [11,13–16]. Measurement of domi-ance as “winner” behavior and submissiveness as “loser” behavior
n triads, or especially in pairs of animals, has better reproducibil-ty than within larger groups. Endpoints such as time spent on aeeder [13,15] or number of sucrose pellets consumed [17,18] cane easily adapted for fast and quantitative behavioral analysis.
Mice are a social species, with a highly developed socialtructure, involving communal nesting and parenting, and wellefined mating, territorial and aggressive behavior [19]. Such social
nteractions lead to the formation of a hierarchy based upon dom-nant and submissive relationships, allowing the use of mice as
model for the study of these relationships. Currently, a num-er of different behavioral paradigms are employed to studyice’s social behavior. For example: the three-chamber social
pproach [19–22], resident–intruder [23–25], partition [26,27] andhe dominant–submissive relationship [13,15,28,29] tests.
The dominant–submissive relationships (DSR) paradigm is social interaction test developed to study dominance andubmissiveness and for the screening of mood stabilizing andntidepressant compounds. Antidepressants from different classesnd generations, such as fluoxetine, imipramine and paroxet-ne, time- and dose-dependently reduced submissive behavior13,15,28,30,31]. The DSR test is based on repeated competition forood between fixed pairs of animals. After repeated interactions, aelationship is formed wherein one animal develops the behavioralrait of dominance and the other, submissiveness. Such formationf hierarchal relationships between mice developed without visi-le demonstration of aggressive behavior. Among wild type micend rats, less than half form clear and consistent DSRs. Most of thenimals form flexible relationships not characterized by consistentominant or submissive behavior. In order to enhance animals’ fea-ures of dominance or submissiveness, we employed a selectivereeding approach based on the DSR paradigm [32].
We previously found that the nature of animals’ social relation-hips in the DSR test is genetically determined [32]. Thus, usingutbred Sabra mice, we found that only 25% of wild type animalseveloped DSR. Four generations of selective breeding for the traitf interest yielded populations in which more than 80% of ani-als developed DSR [32]. Further selection of these mice based on
ehavioral phenotype resulted in the development of animals withtrong and stable dominant and submissive traits. In this paper,sing different behavioral and pharmacological approaches, weharacterize different features of these two unique animal popu-ations.
. Methods
.1. Animals
The populations of dominant (Dom) and submissive (Sub) mice used in this studyere selectively bred on basis of their behavior in the DSR test [32] (see description
n Section 2.2 below). These animals are descendants of the outbred Sabra strain,hich freely develop relationships of dominance and submissiveness in the DSR test
nd are measurably larger than other better known strains [33]. The Sabra strain’sehavioral and biochemical characteristics were recently found to lie within theiapason of those of C57BL/6, Balb/c and ICR mice [33]. Progenies of selectively bredominant and submissive animals from generations F13, F14 and F15 were used
n this study. Animals were given standard laboratory chow and water ad libitum.uring DSR testing, chow was provided according to the DSR protocol [32]; see alsoescription below. The colony room was maintained on a 12 h L:12 h D cycle (lightsn 07:00–19:00 h). The experiments were conducted in compliance with NIH/USDAuidelines, under the approval of the Institutional Animal Care and Use Committee.
.2. Dominant–submissive relationship (DSR) test
The DSR apparatus, made from Plexiglas, consists of two identical chambers12 cm × 8.5 cm × 7 cm) joined by a tunnel (2.5 cm × 2.5 cm × 27 cm) with a 0.5 cmiameter hole in its bottom center. A self-refilling feeder is connected to the tunnel,
esearch 236 (2013) 225– 235
allowing a constant supply of sweetened milk (3% fat, 10% sugar), to which only oneanimal has access at any given moment. The tunnel has narrow slits cut on bothsides of the feeder for easy gate insertion and removal. In this way, the paired micehave an equal starting position at the beginning of each session. The description andschematic presentation of DSR apparatus was presented in detail previously [32].
DSR tests were carried out for five consecutive days per week for a two weekperiod. During each 14 h period preceding testing, the mice were deprived of food;water was provided ad libitum. The animals had free access to food for two daysbetween testing periods, until the night before the next five day testing period.Pairs of mice from different home cages were matched for relatively similar weight(average weight 43.7 ± 2.1 g) and were tested according to the DSR protocol daily.During each 5 min DSR session, milk drinking times were recorded manually. DSRpairs were identified according to criteria described previously [32]. Briefly, DSRpairs required significant difference (p < 0.05) between the average daily drinkingtimes of both animals in a pair, with the difference in drinking score comprising atleast 40%.
2.3. Selective breeding
For breeding, a single dominant or submissive male was housed together withtwo or three females selected for the same trait. The founding generation of dom-inant or submissive animals was designated generation P (parental); offspringgenerations were designated F1, F2, F3, . . . F15, etc. Males of the P generation thatformed DSR during the two week testing period were bred with females with thesame traits: submissive males with submissive females and dominant males withdominant females. In each subsequent generation, the number of animal pairs sub-jected to the DSR test depended on the number of offspring. In consecutive (F1–F15)generations, offspring of the dominant mice were paired and subjected to two weeksof DSR testing with offspring of the submissive animals. Concurrently, we main-tained a colony of background-strain Sabra mice, in which breeders were randomlyassigned.
2.4. Resident–intruder test
This procedure evaluates social behavior in animals [23,24,34]. In the cur-rent experimental setup, cages were divided into two identical compartments(18 cm × 20 cm) by a transparent divider. A resident male (dominant or submissive),was placed in one of the compartments for 30 min, after which a male intruder ofthe opposite behavioral phenotype was placed in the neighboring compartment.In this manner, animals established visual contact without any physical contact.After 10 min, the divider (lattice) was removed and animals (resident and intruder)were physically exposed to each other for 10 min. Time engaged in aggressivebehavior (biting, scratching or chasing after the cagemate) during this time wasrecorded. Aggression expressed in attack initiation toward the partner or fight-ing back in response to attacks was defined as dominant behavior. After 10 minof interaction, animals were placed for 30 min in separate cages before returningto their home cages. Animal pairs (dominant resident/submissive intruder and sub-missive resident/dominant intruder) were consistent during the experiment’s timecourse.
2.5. Open field (OF) test
The OF test is used to estimate spontaneous locomotor (horizontal) andexploratory (vertical) activity [35–40]. The apparatus consists of a clear glass box(30 cm × 40 cm), the floor of which is divided by lines drawn into 20 equally sizedsquares. For better habituation, all animals were placed in the experimental roomone hour prior to the test. Each mouse was placed individually in the center ofthe apparatus, under ambient lighting conditions of 185 lx. Horizontal (number ofsquares crossed) and vertical (number of rearings) activities were recorded manuallyfor 6 min; between subjects, the apparatus was cleaned with 70% ethanol.
2.6. Elevated plus maze (EPM)
The EPM test is used to assess anxiety-like behavior [40–42]. The apparatus iscomprised of two enclosed (10 cm × 45 cm × 40 cm) and two open (10 cm × 45 cm)arms that extend from a common central platform (10 cm × 10 cm). Ambient lightingconditions were set to 720 lx. The black wooden apparatus is elevated to a height of60 cm above floor level. The tests were conducted during the period of illuminationin the mice’s colony room. For better habituation, all animals were placed in theexperimental room 1 h prior to the test. Each animal was placed at the center of themaze, facing one of the enclosed arms. Each mouse spent a 5 min session in the EPM,during which the number of entries into open and closed arms, as well as time spent
in the open and closed arms were manually scored. Animals’ behavior was analyzedas ratios of entries into the open arms to total arm entries (OE/TE), time spent in theopen arms relative to the total time (OT/TT) and as the sum of total entries (TE) intoboth open and closed arms. Between each mouse’s sessions, the maze was cleanedwith 70% ethanol.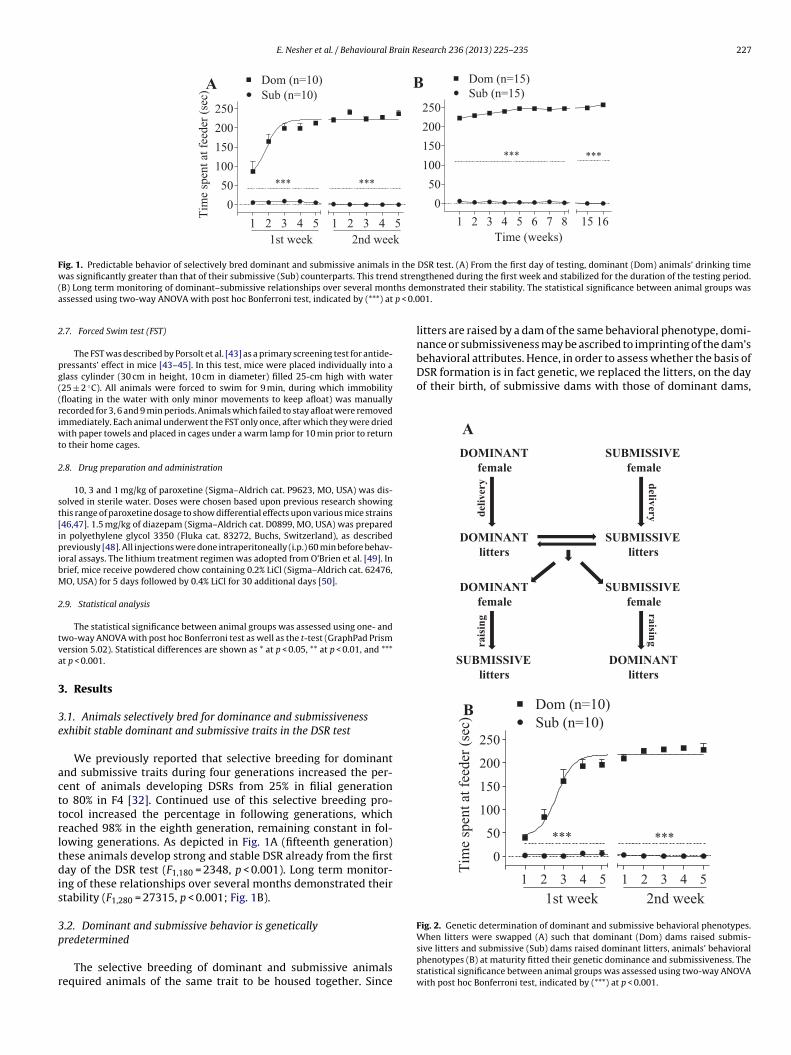
E. Nesher et al. / Behavioural Brain Research 236 (2013) 225– 235 227
050
100150200250
Sub (n=10)Dom (n=10)
1st week1 2 3 4 5
2nd week
***
A
***
1 2 3 4 5
Tim
e sp
ent a
t fee
der (
sec)
1 2 3 4 5 6 7 80
50100150200250
15 16
Dom (n=15)Sub (n=15)
***
B
***
Time (weeks)
Fig. 1. Predictable behavior of selectively bred dominant and submissive animals in the DSR test. (A) From the first day of testing, dominant (Dom) animals’ drinking timew d stren( ths dea p < 0.0
2
pg((riwt
2
st[ipibM
2
tva
3
3e
acttrltdis
3p
r
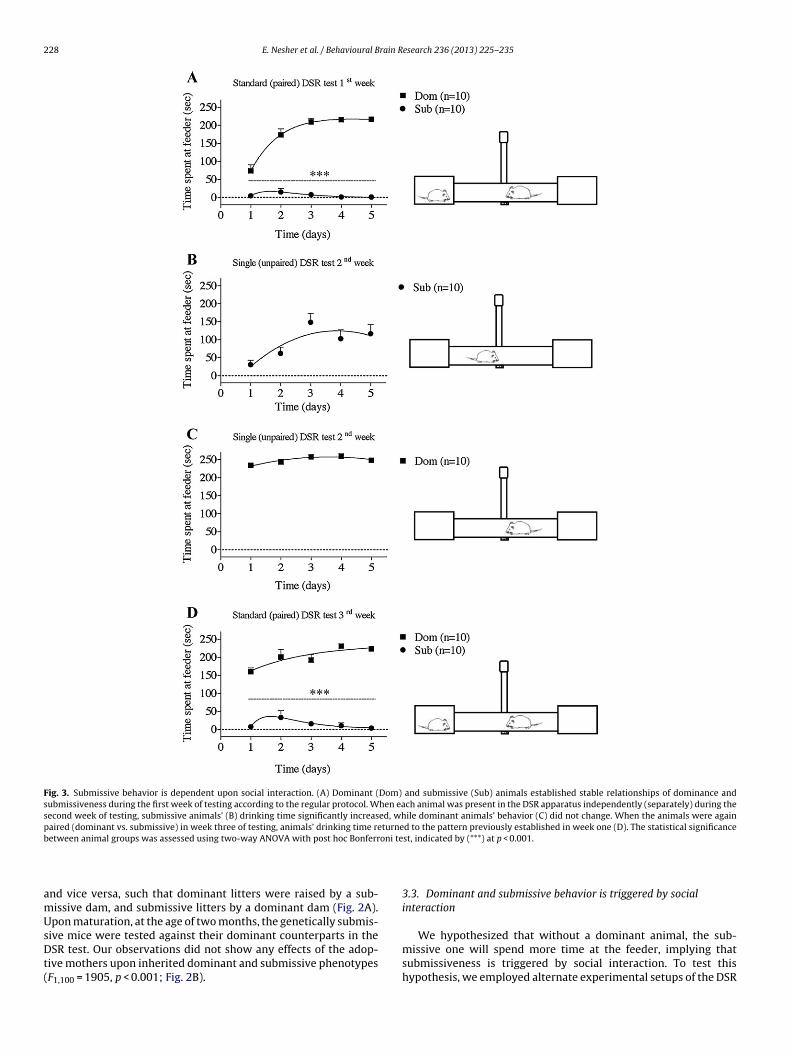
nance or submissiveness may be ascribed to imprinting of the dam’sbehavioral attributes. Hence, in order to assess whether the basis ofDSR formation is in fact genetic, we replaced the litters, on the dayof their birth, of submissive dams with those of dominant dams,
DOMINANTfemale
DOMINANTlitters
DOMINANTfemale
SUBMISSIVElitters
SUBMISSIVEfemale
SUBMISSIVElitters
SUBMISSIVEfemale
DOMINANTlitters
delivery delivery
raising raising
A
0
50
100
150
200
250Sub (n=10)Dom (n=10)
1st week1 2 3 4 5
2nd week
***
B
***
1 2 3 4 5
Tim
e sp
ent a
t fee
der (
sec)
Fig. 2. Genetic determination of dominant and submissive behavioral phenotypes.
as significantly greater than that of their submissive (Sub) counterparts. This trenB) Long term monitoring of dominant–submissive relationships over several monssessed using two-way ANOVA with post hoc Bonferroni test, indicated by (***) at
.7. Forced Swim test (FST)
The FST was described by Porsolt et al. [43] as a primary screening test for antide-ressants’ effect in mice [43–45]. In this test, mice were placed individually into alass cylinder (30 cm in height, 10 cm in diameter) filled 25-cm high with water25 ± 2 ◦C). All animals were forced to swim for 9 min, during which immobilityfloating in the water with only minor movements to keep afloat) was manuallyecorded for 3, 6 and 9 min periods. Animals which failed to stay afloat were removedmmediately. Each animal underwent the FST only once, after which they were dried
ith paper towels and placed in cages under a warm lamp for 10 min prior to returno their home cages.
.8. Drug preparation and administration
10, 3 and 1 mg/kg of paroxetine (Sigma–Aldrich cat. P9623, MO, USA) was dis-olved in sterile water. Doses were chosen based upon previous research showinghis range of paroxetine dosage to show differential effects upon various mice strains46,47]. 1.5 mg/kg of diazepam (Sigma–Aldrich cat. D0899, MO, USA) was preparedn polyethylene glycol 3350 (Fluka cat. 83272, Buchs, Switzerland), as describedreviously [48]. All injections were done intraperitoneally (i.p.) 60 min before behav-
oral assays. The lithium treatment regimen was adopted from O’Brien et al. [49]. Inrief, mice receive powdered chow containing 0.2% LiCl (Sigma–Aldrich cat. 62476,O, USA) for 5 days followed by 0.4% LiCl for 30 additional days [50].
.9. Statistical analysis
The statistical significance between animal groups was assessed using one- andwo-way ANOVA with post hoc Bonferroni test as well as the t-test (GraphPad Prismersion 5.02). Statistical differences are shown as * at p < 0.05, ** at p < 0.01, and ***t p < 0.001.
. Results
.1. Animals selectively bred for dominance and submissivenessxhibit stable dominant and submissive traits in the DSR test
We previously reported that selective breeding for dominantnd submissive traits during four generations increased the per-ent of animals developing DSRs from 25% in filial generationo 80% in F4 [32]. Continued use of this selective breeding pro-ocol increased the percentage in following generations, whicheached 98% in the eighth generation, remaining constant in fol-owing generations. As depicted in Fig. 1A (fifteenth generation)hese animals develop strong and stable DSR already from the firstay of the DSR test (F1,180 = 2348, p < 0.001). Long term monitor-
ng of these relationships over several months demonstrated theirtability (F1,280 = 27315, p < 0.001; Fig. 1B).
.2. Dominant and submissive behavior is genetically
redeterminedThe selective breeding of dominant and submissive animalsequired animals of the same trait to be housed together. Since
gthened during the first week and stabilized for the duration of the testing period.monstrated their stability. The statistical significance between animal groups was01.
litters are raised by a dam of the same behavioral phenotype, domi-
When litters were swapped (A) such that dominant (Dom) dams raised submis-sive litters and submissive (Sub) dams raised dominant litters, animals’ behavioralphenotypes (B) at maturity fitted their genetic dominance and submissiveness. Thestatistical significance between animal groups was assessed using two-way ANOVAwith post hoc Bonferroni test, indicated by (***) at p < 0.001.
228 E. Nesher et al. / Behavioural Brain Research 236 (2013) 225– 235
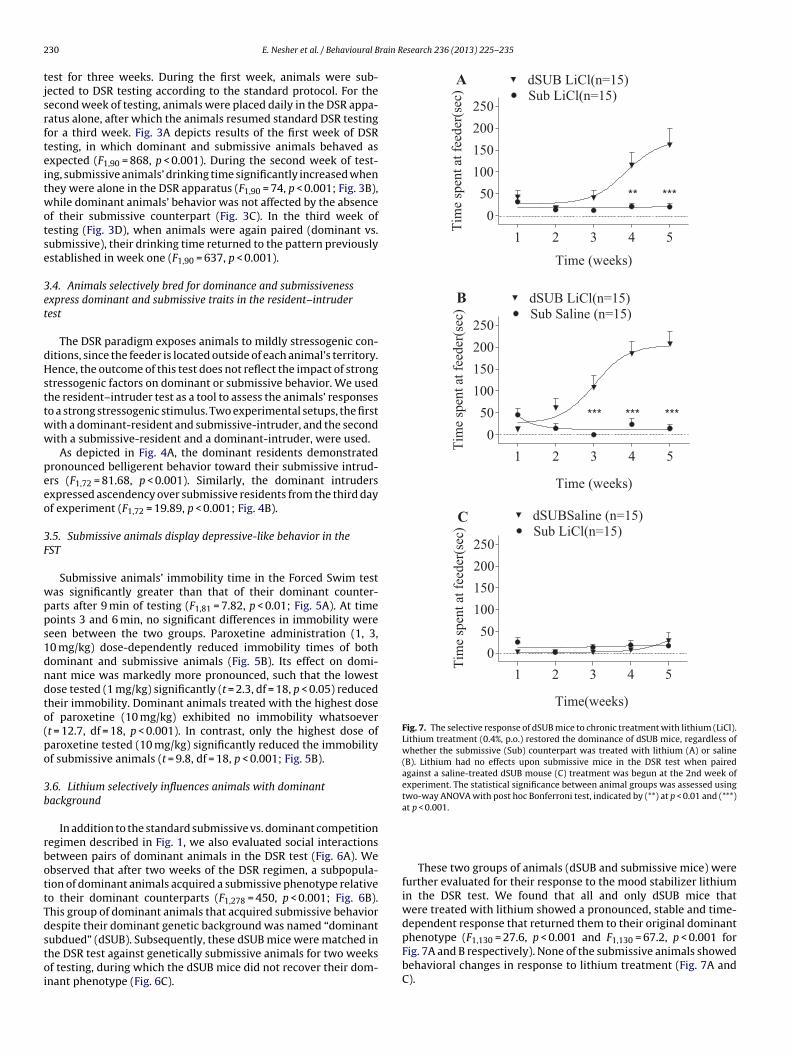
Fig. 3. Submissive behavior is dependent upon social interaction. (A) Dominant (Dom) and submissive (Sub) animals established stable relationships of dominance andsubmissiveness during the first week of testing according to the regular protocol. When each animal was present in the DSR apparatus independently (separately) during thesecond week of testing, submissive animals’ (B) drinking time significantly increased, while dominant animals’ behavior (C) did not change. When the animals were againpaired (dominant vs. submissive) in week three of testing, animals’ drinking time returned to the pattern previously established in week one (D). The statistical significanceb oni te
amUsDt(
etween animal groups was assessed using two-way ANOVA with post hoc Bonferr
nd vice versa, such that dominant litters were raised by a sub-issive dam, and submissive litters by a dominant dam (Fig. 2A).pon maturation, at the age of two months, the genetically submis-
ive mice were tested against their dominant counterparts in theSR test. Our observations did not show any effects of the adop-
ive mothers upon inherited dominant and submissive phenotypesF1,100 = 1905, p < 0.001; Fig. 2B).
st, indicated by (***) at p < 0.001.
3.3. Dominant and submissive behavior is triggered by socialinteraction
We hypothesized that without a dominant animal, the sub-missive one will spend more time at the feeder, implying thatsubmissiveness is triggered by social interaction. To test thishypothesis, we employed alternate experimental setups of the DSR

E. Nesher et al. / Behavioural Brain Research 236 (2013) 225– 235 229
1 2 3 4
0
20
40
60Dom resident (n=10)Sub intruder (n=10)
A
*** ***
Time (days)
Tim
e sp
ent e
ngag
ed in
agre
ssiv
e be
havi
or (s
ec)
1 2 3 4
0
20
40
60Dom intruder (n=10)Sub resident (n=10)
B
* ***
Time (days)
Fig. 4. Expression of dominant and submissive phenotypes in the resident–intruder test. (A) Submissive (Sub) intruders offered no challenge to dominant (Dom) residents.(B) Dominant intruders exerted supremacy over submissive residents. Aggressive behavior is defined as biting, clawing or chasing. The statistical significance between animalgroups was assessed using two-way ANOVA with post hoc Bonferroni test, indicated by (*) at p < 0.05, (**) at p < 0.01 and (***) at p < 0.001.
0
100
200
300
63 9
Sub (n=15)Dom (n=15)
**
A
Time(min)
Imm
obili
ty ti
me
(sec
)
0
100
200
300
400Dom (n=10 for eachdose)Sub (n=10 for eachdose)
Veh 1 3 10 Ve h 1 3 10
*** ***
*
***
B
Treatment dose (mg/kg)
Imm
obili
ty ti
me
(sec
)
Fig. 5. Dominant and submissive mice respond differentially to paroxetine in the Forced Swim test. (A) Naïve submissive (Sub) animals remained immobile significantly(p < 0.01) longer than did naïve dominant (Dom) animals in the 9 min FST. (B) 1 mg/kg paroxetine reduced dominant animals’ immobility (p < 0.05), and higher doses nearlyeliminated immobility (p < 0.001), while submissive animals responded only to 10 mg/kg (p < 0.001). Immobility time was significantly different between experimental groupsas assessed using one- and two-way ANOVA with Bonferroni post hoc corrected analysis, indicated by (*) at p < 0.05, (**) at p < 0.01 and (***) at p < 0.001.
DOMmales
DOMmales
DSRtest
DOMmales
dSUBmales
A
0
50
100
150
200
250
Dom (n=15)dSUB (n=15)
***
B
1st week1 2 3 4 5
2nd week1 2 3 4 5
***
Tim
e sp
ent a
t fee
der(
sec)
0
50
100
150
200
250
Sub (n=15)dSUB (n=15)
C
1st week1 2 3 4 5
2nd week1 2 3 4 5
Fig. 6. Development of dSUB mice using the DSR test. (A) Dominant (Dom) mice were paired against each other in the DSR test. While some of the animals maintained theirdominant features, other became submissive (dSUB). (B) dSUBs acquired stable submissive phenotypes in the DSR test. (C) dSUBs failed to recover their former dominancewhen paired again in the DSR test against submissive (Sub) counterparts. The statistical significance between animal groups was assessed using two-way ANOVA with posthoc Bonferroni test, indicated by (*) at p < 0.05 and (***) at p < 0.001.
2 rain Research 236 (2013) 225– 235
tjsrfteitwotse
3et
dHsttww
peeo
3F
wpps1dndto(po
3b
rbottTdstoi
1 2 3 4 50
50100150200250
*****
dSUB LiCl(n=15)Sub LiCl(n=15)
A
Time (weeks)
Tim
e sp
ent a
t fee
der(
sec)
1 2 3 4 50
50100150200250
***
dSUB LiCl(n=15)Sub Saline (n=15)
B
*** ***
Time (weeks)
Tim
e sp
ent a
t fee
der(
sec)
1 2 3 4 50
50100150200250
Sub LiCl(n=15)dSUBSaline (n=15)C
Time(weeks)
Tim
e sp
ent a
t fee
der(
sec)
Fig. 7. The selective response of dSUB mice to chronic treatment with lithium (LiCl).Lithium treatment (0.4%, p.o.) restored the dominance of dSUB mice, regardless ofwhether the submissive (Sub) counterpart was treated with lithium (A) or saline(B). Lithium had no effects upon submissive mice in the DSR test when pairedagainst a saline-treated dSUB mouse (C) treatment was begun at the 2nd week ofexperiment. The statistical significance between animal groups was assessed usingtwo-way ANOVA with post hoc Bonferroni test, indicated by (**) at p < 0.01 and (***)at p < 0.001.
30 E. Nesher et al. / Behavioural B
est for three weeks. During the first week, animals were sub-ected to DSR testing according to the standard protocol. For theecond week of testing, animals were placed daily in the DSR appa-atus alone, after which the animals resumed standard DSR testingor a third week. Fig. 3A depicts results of the first week of DSResting, in which dominant and submissive animals behaved asxpected (F1,90 = 868, p < 0.001). During the second week of test-ng, submissive animals’ drinking time significantly increased whenhey were alone in the DSR apparatus (F1,90 = 74, p < 0.001; Fig. 3B),hile dominant animals’ behavior was not affected by the absence
f their submissive counterpart (Fig. 3C). In the third week ofesting (Fig. 3D), when animals were again paired (dominant vs.ubmissive), their drinking time returned to the pattern previouslystablished in week one (F1,90 = 637, p < 0.001).
.4. Animals selectively bred for dominance and submissivenessxpress dominant and submissive traits in the resident–intruderest
The DSR paradigm exposes animals to mildly stressogenic con-itions, since the feeder is located outside of each animal’s territory.ence, the outcome of this test does not reflect the impact of strong
tressogenic factors on dominant or submissive behavior. We usedhe resident–intruder test as a tool to assess the animals’ responseso a strong stressogenic stimulus. Two experimental setups, the firstith a dominant-resident and submissive-intruder, and the secondith a submissive-resident and a dominant-intruder, were used.
As depicted in Fig. 4A, the dominant residents demonstratedronounced belligerent behavior toward their submissive intrud-rs (F1,72 = 81.68, p < 0.001). Similarly, the dominant intrudersxpressed ascendency over submissive residents from the third dayf experiment (F1,72 = 19.89, p < 0.001; Fig. 4B).
.5. Submissive animals display depressive-like behavior in theST
Submissive animals’ immobility time in the Forced Swim testas significantly greater than that of their dominant counter-arts after 9 min of testing (F1,81 = 7.82, p < 0.01; Fig. 5A). At timeoints 3 and 6 min, no significant differences in immobility wereeen between the two groups. Paroxetine administration (1, 3,0 mg/kg) dose-dependently reduced immobility times of bothominant and submissive animals (Fig. 5B). Its effect on domi-ant mice was markedly more pronounced, such that the lowestose tested (1 mg/kg) significantly (t = 2.3, df = 18, p < 0.05) reducedheir immobility. Dominant animals treated with the highest dosef paroxetine (10 mg/kg) exhibited no immobility whatsoevert = 12.7, df = 18, p < 0.001). In contrast, only the highest dose ofaroxetine tested (10 mg/kg) significantly reduced the immobilityf submissive animals (t = 9.8, df = 18, p < 0.001; Fig. 5B).
.6. Lithium selectively influences animals with dominantackground
In addition to the standard submissive vs. dominant competitionegimen described in Fig. 1, we also evaluated social interactionsetween pairs of dominant animals in the DSR test (Fig. 6A). Webserved that after two weeks of the DSR regimen, a subpopula-ion of dominant animals acquired a submissive phenotype relativeo their dominant counterparts (F1,278 = 450, p < 0.001; Fig. 6B).his group of dominant animals that acquired submissive behaviorespite their dominant genetic background was named “dominant
ubdued” (dSUB). Subsequently, these dSUB mice were matched inhe DSR test against genetically submissive animals for two weeksf testing, during which the dSUB mice did not recover their dom-nant phenotype (Fig. 6C).These two groups of animals (dSUB and submissive mice) werefurther evaluated for their response to the mood stabilizer lithiumin the DSR test. We found that all and only dSUB mice thatwere treated with lithium showed a pronounced, stable and time-dependent response that returned them to their original dominantphenotype (F1,130 = 27.6, p < 0.001 and F1,130 = 67.2, p < 0.001 forFig. 7A and B respectively). None of the submissive animals showed
behavioral changes in response to lithium treatment (Fig. 7A andC).
rain R
3i
ti(oCrdsidis(
3D
aw(t(
4
saibbsagdpdttfs(isiemhost
sp[siapc
E. Nesher et al. / Behavioural B
.7. Diazepam shows paradoxical effects upon dominant animalsn the EPM and OF tests
We evaluated the response of dominant and submissive animalso the anxiolytic diazepam. In the EPM test, diazepam exerts anx-olytic effects upon submissive animals, reflected in a significantt = 2.8, df = 18, p < 0.05) increase in the ratio of time spent in thepen portions of the maze to the total test time (OT/TT) (Fig. 8F).onversely, dominant animals presented an opposite, anxiogeniceaction to diazepam treatment, as reflected in a significant (t = 2.4,f = 18, p < 0.05) decrease in dominant animals’ OT/TT (Fig. 8E). Noignificant differences were found as a result of diazepam treatmentn other measured parameters of the EPM (Fig. 8A–D). In the OF test,ominant animals showed an unpredicted increase in locomotion
n response to diazepam treatment (t = 3.79, df = 18, p < 0.01), whileubmissive animals demonstrated an anticipated sedative effectt = 2.77, df = 18, p < 0.05) (Fig. 9A and B).
.8. Diazepam increased dominant animals’ drinking time in theSR test
In the DSR test, diazepam-treated (1.5 mg/kg) dominantnimals showed a robust increase in their drinking time,hen matched against vehicle-treated dominant counterparts
F1,45 = 233, p < 0.001; Fig. 10A). In contrast, no effect of diazepamreatment was seen upon submissive animals in the DSR testFig. 10B).
. Discussion
Dominance and submissiveness are two opposite poles of theocial behavior spectrum [3,51–53]. Previous research in humansnd using animal models suggest that extreme expressions of dom-nance and submissiveness may be involved in the etiology ofehavioral disturbances [3,54]. In this work we evaluated animals’ehavior and differential responses to representative antidepres-ant, anxiolytic and mood stabilizing agents of the progeny ofnimals selectively bred for dominance or submissiveness over 15enerations. We previously demonstrated that establishment ofominant–submissive behavior in the DSR paradigm is geneticallyredetermined [32]. The first generations of selectively bred miceeveloped DS relationships only after one week of habituation tohe DSR paradigm [32]. Repeated selective breeding strengthenedhe animals’ behavioral phenotypes to such an extent that alreadyrom the first days of social interaction, animals clearly demon-trated dominant and submissive behavior in the DSR paradigmFig. 1A). These relationships also showed long-term stability dur-ng several months of testing (Fig. 1B). While these experimentstrongly suggest that dominance and submissiveness are genet-cally determined traits, many studies have also shown strongffects of external factors such as the early life environment (i.e.aternal behavior) on animals’ behavioral status [55–59]. We show
ere (Fig. 2) that while the effect of maternal factors cannot be ruledut, dominant and submissive pups raised by mothers of the oppo-ite behavioral phenotype behaved in the DSR test according toheir genetic background.
In contrast to the DSR test, in which animals establish theirocial relationships without visible violence, the resident–intruderaradigm is specifically designed to assess aggressive behavior23,34]. Selectively bred dominant mice easily demonstrated theirupremacy in the resident intruder test by use of aggressive behav-
or toward their submissive counterparts, regardless of their roles resident or intruder (Fig. 4). We suggest that since the DSRaradigm does not force mice into a struggle over territory, but theompetition takes place in a neutral environment not previouslyesearch 236 (2013) 225– 235 231
owned by either animal, it is less crucial to each mouse to estab-lish its supremacy. In this case, the submissive member of the pairhas the option of relinquishing his share of limited resources. Thus,it appears that the dominant behavior of these selectively bredanimals may be expressed differentially, according to the environ-ment and social situation. It is important to stress that submissiveresidents initially employed aggressive behavior toward dominantintruders, who later overcame this resistance. Animals’ aggressioncontributes to the social hierarchy which enables defense, repro-duction and resource access. The invasion of territory presents athreat to this hierarchy, such that even submissive animals willattempt to defend their territory.
In humans, environmental, especially social factors, may serveas triggers and influence the establishment of social status andpersonality traits [60]. In many cases, subordinate or submissivebehavior is a result of social triggers, particularly the existenceof a stronger, dominant partner [61]. In the present case, genet-ically submissive animals drink freely when placed alone in theDSR environment (Fig. 3B), and reassume their submissive statusin the presence of a counterpart. At the same time, the interactionbetween pairs of two submissive animals in DSR test did not evenapproach the proportions seen between dominant and submissivemice (data not shown). Thus, we may suggest that the behaviorof submissive mice is dependent upon environmental and socialtriggers.
Personality formation and development, education, family life,as well as work relationships, all involve social interaction [62,63].For susceptible individuals, an abrupt change in social status maytrigger affective and personality disorders, notably depression andantisocial personality disorder [64,65]. In animals, social inter-actions often lead to aggression and violence, until one of thesubjects’ subduing [6,66,67]. In the present work, when dominantmice were paired against each other in the DSR test, most main-tained their dominance, while others acquired a strong subordinatephenotype (dSUB), despite their dominant genetic background(Fig. 6A). Interestingly, dSUB mice matched in the DSR test againstgenetically submissive animals did not recover their dominantbehavior (Fig. 6C). We treated dSUB animals with the mood sta-bilizer lithium. A number of studies have explored the effect oflithium upon aggression and anxiety in animal models [68,69].Recent studies found that distinct mice strains exhibit antimanicor antidepressant-like effects in response to lithium treatment[70,71]. Lithium’s efficacy as a mood stabilizer has been demon-strated in case studies that produced response rates of 30–65% [72].In the present study, lithium restored the dominance of dSUB ani-mals in a consistent manner, without affecting the behavior of theirsubmissive counterparts (Fig. 7). This lack of effect upon the sub-missive animals was expected, since these animals express featuresfrom the spectrum of depressive behavior. Thus, it appears that themechanisms responsible for the submissive behavior of dSUB ani-mals are different from those governing the behavior of geneticallysubmissive animals.
While the selectively bred animals previously showed stablebehavioral attributes in a battery of behavioral tests [32], wepresently observed paradoxical effects of dominant mice to theanxiolytic compound diazepam. Diazepam belongs to the well-studied benzodiazepine family of anxiolytic drugs acting throughraising the efficiency of GABAergic activity [73,74]. In rodents,diazepam administration induces sedation expressed in reducedlocomotion, as well as anxiolytic-like effects. Among both humansand animals, certain individuals have been reported to demon-strate a paradoxical reaction to GABAA receptor modulators, a
phenomenon whose mechanism is not well understood [75–77]. Inthe present study, administration of diazepam to submissive ani-mals lead to reduced anxiety-like behavior in the EPM (Fig. 8B)and the predicted sedative effect in the OF (Fig. 9B), while the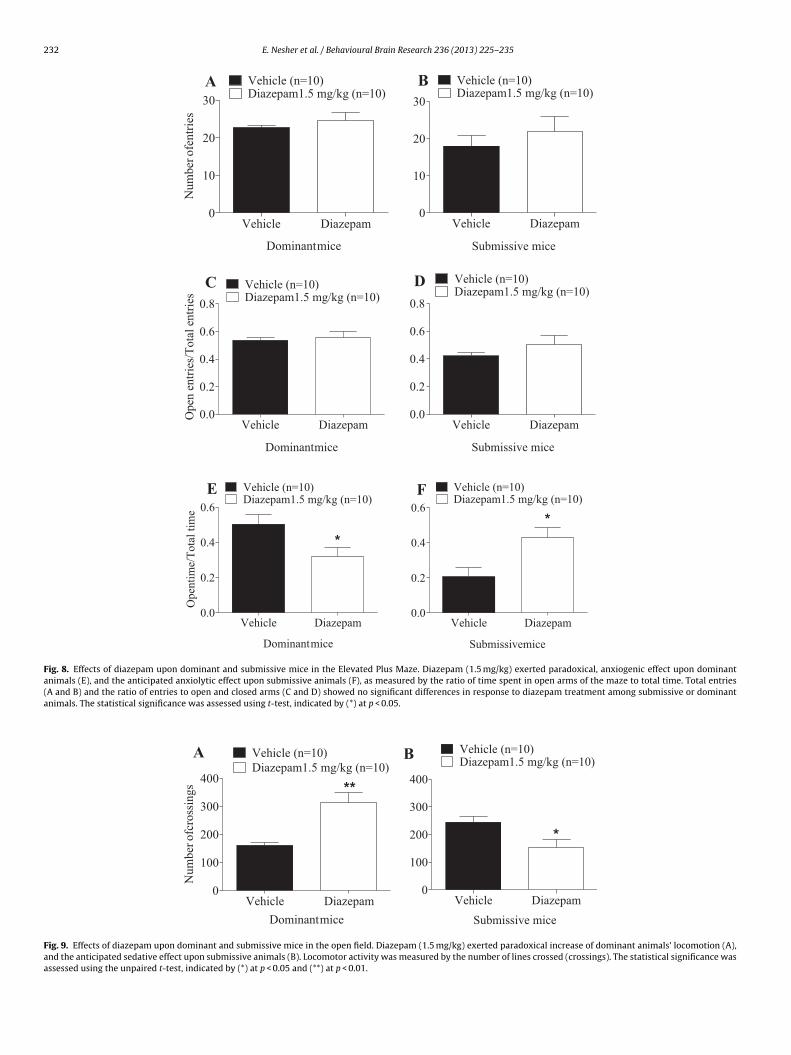
232 E. Nesher et al. / Behavioural Brain Research 236 (2013) 225– 235
Vehicle Diazepam0
10
20
30A Vehicle (n=10)
Diazepam1.5 mg/kg (n=10)
Dominantmice
Num
bero
fent
ries
Vehicle Diazepam0
10
20
30B Vehicle (n=10)
Diazepam1.5 mg/kg (n=10)
Submissive mice
Vehicle Diazepam0.0
0.2
0.4
0.6
0.8
C Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
Dominantmice
Ope
n en
tries
/Tot
al e
ntrie
s
Vehicle Diazepam0.0
0.2
0.4
0.6
0.8
D Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
Submissive mice
Vehicle Diazepam0.0
0.2
0.4
0.6
*
E Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
Dominant mice
Ope
ntim
e/To
tal t
ime
Vehicle Diazepam0.0
0.2
0.4
0.6*
F Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
Submissivemice
Fig. 8. Effects of diazepam upon dominant and submissive mice in the Elevated Plus Maze. Diazepam (1.5 mg/kg) exerted paradoxical, anxiogenic effect upon dominantanimals (E), and the anticipated anxiolytic effect upon submissive animals (F), as measured by the ratio of time spent in open arms of the maze to total time. Total entries(A and B) and the ratio of entries to open and closed arms (C and D) showed no significant differences in response to diazepam treatment among submissive or dominantanimals. The statistical significance was assessed using t-test, indicated by (*) at p < 0.05.
Vehicle Diazepam0
100
200
300
400**
Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
A
Dominantmice
Num
bero
fcro
ssin
gs
Vehicle Diazepam0
100
200
300
400
*
Vehicle (n=10)Diazepam1.5 mg/kg (n=10)B
Submissive mice
Fig. 9. Effects of diazepam upon dominant and submissive mice in the open field. Diazepam (1.5 mg/kg) exerted paradoxical increase of dominant animals’ locomotion (A),and the anticipated sedative effect upon submissive animals (B). Locomotor activity was measured by the number of lines crossed (crossings). The statistical significance wasassessed using the unpaired t-test, indicated by (*) at p < 0.05 and (**) at p < 0.01.

E. Nesher et al. / Behavioural Brain Research 236 (2013) 225– 235 233
1 2 3 4 50
50100150200250300
Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
A
Dominant mice
***
*****
***
**
Time (days)
Tim
e sp
ent a
t fee
der(
sec)
1 2 3 4 50
50100150200250300
Vehicle (n=10)Diazepam1.5 mg/kg (n=10)
B
Submissive miceTime (days)
F t. AniD -treats hoc B
dEdttieiWrwfetpeiarpmp
eia(mswsiitsnsatwcsmci[a
ig. 10. Effects of diazepam upon dominant and submissive mice in the DSR tesiazepam (1.5 mg/kg) increased dominant animals’ dominance (A) relative to vehicle
ignificance between animal groups was assessed using two-way ANOVA with post
rug induced anxiogenic effects among dominant animals in thePM (Fig. 8A), and increased activity in the OF (Fig. 9A). Finally,iazepam significantly strengthened the dominance of animals inhe DSR paradigm relative to their vehicle-treated dominant coun-erparts (Fig. 10). This exaggeration of the dominant phenotypen the DSR test was unexpected, as were the drug’s paradoxicalffects in the EPM and OF. Paradoxical reaction to benzodiazepinesn the clinic include increased hostility, aggression and rage [78].
hile this phenomenon was first reported in 1962, and furtheresearch found this effect to occur in nearly 1% of individuals treatedith benzodiazepines [79], the molecular mechanism responsible
or this reaction is not well understood. Clinical studies have wellstablished that benzodiazepines must be administered with cau-ion to individuals with personality disorders because of the risk ofaradoxical reaction [73,74]. Numerous case studies report adverseffect of diazepam in human patients exhibiting antisocial behav-or [78,80–82]. We suggest that selective breeding of dominantnimals strengthened their genetic predisposition to paradoxicaleaction to benzodiazepines that may be shared with the humanopulation expressing the same phenomena. Thus, these animalsay play a role in the determination of genetic predisposition of
atients to treatment with mood modulation reagents.Additionally, dominant and submissive animals showed differ-
ntial responses to treatment with the selective serotonin reuptakenhibitor (SSRI) paroxetine in the FST (Fig. 5A and B). Submissivenimals responded significantly only to the highest dose tested10 mg/kg). This seeming resistance to antidepressant effect of a
edication commonly used in the clinic appears to be an aspect ofubmissive animals’ depressive-like phenotype. Dominant animalsere considerably more sensitive to paroxetine treatment, and
howed a dose-dependent response, which reached zero immobil-ty in response to 10 mg/kg. Nine consecutive minutes of swimmings highly uncharacteristic behavior for mice, whose behavior inhe FST is normally characterized by a period of swimming and/ortruggle, followed by floating [83,84]. The non-adaptive, freneticature of this behavior was underlined by our need to removeeveral dominant animals who continued to swim beyond theirbility to thereby stay afloat, and began to drown. In such cases,he animals failed to assume the passive floating posture whichould enable their survival without our intervention. A search for
ase studies of adverse reaction to antidepressants found that aubpopulation of depressed patients treated with SSRIs exhibitedanic and hypomanic behavior [85–87]. Moreover, SSRIs are often
ontraindicated in the pediatric setting for the risk of their induc-ng mania, and are contraindicated for patients in a manic state88–90]. These findings appear to offer pharmacological validity,lbeit limited, for dominance to represent manic-like behavior.
[
[
mals were paired dominant vs. dominant (A) and submissive vs. submissive (B).ed dominant mice, without altering submissive animals’ behavior (B). The statisticalonferroni test, indicated by (*) at p < 0.05, (**) at p < 0.01 and (***) at p < 0.001.
This hypothesis, in which dominant and submissive animals eachmodel opposite ends of the manic-depressive spectrum, remainsto be tested thoroughly in future behavioral and pharmacologicalexperiments.
We anticipate that further studies of the molecular mecha-nisms responsible for dominance and submissiveness may leadto the identification of biomarkers of predisposition to behavioraldisorders, enabling early clinical intervention. Additionally, theseanimals may contribute to the characterization of the genetic basisof sensitivity to mood-regulating drugs. Such future study shouldcontribute to the development of more effective clinical strate-gies for the diagnosis and treatment of affective and personalitydisorders.
Acknowledgements
This manuscript is dedicated to the memory of the late Dr. EwaMalatynska, a talented scientist and friend who originally devel-oped the DSR paradigm.
This work was supported by the Israeli Ministry of ImmigrantAbsorption.
References
[1] Bjorkqvist K. Social defeat as a stressor in humans. Physiology and Behavior2001;73:435–42.
[2] Blanchard RJ, McKittrick CR, Blanchard DC. Animal models of social stress:effects on behavior and brain neurochemical systems. Physiology and Behavior2001;73:261–71.
[3] Gardner Jr R. Mechanisms in manic-depressive disorder: an evolutionarymodel. Archives of General Psychiatry 1982;39:1436–41.
[4] Barrett MS, Barber JP. Interpersonal profiles in major depressive disorder. Jour-nal of Clinical Psychology 2007;63:247–66.
[5] Cervenka S, Gustavsson JP, Halldin C, Farde L. Association between striatal andextrastriatal dopamine D2-receptor binding and social desirability. Neuroim-age 2010;50:323–8.
[6] Chase ID, Tovey C, Spangler-Martin D, Manfredonia M. Individual differencesversus social dynamics in the formation of animal dominance hierarchies.Proceedings of the National Academy of Sciences of the United States of America2002;99:5744–9.
[7] Blanchard RJ, Flannelly KJ, Blanchard DC. Life-span studies of dominance andaggression in established colonies of laboratory rats. Physiology and Behavior1988;43:1–7.
[8] Chase ID, Seitz K. Self-structuring properties of dominance hierarchies a newperspective. Advances in Genetics 2011;75:51–81.
[9] Blanchard DC, Spencer RL, Weiss SM, Blanchard RJ, McEwen B, Sakai RR. Visibleburrow system as a model of chronic social stress: behavioral and neuroen-docrine correlates. Psychoneuroendocrinology 1995;20:117–34.
10] Blanchard RJ, Hori K, Tom P, Blanchard DC. Social structure and ethanol con-sumption in the laboratory rat. Pharmacology Biochemistry and Behavior1987;28:437–42.
11] Schutz MT, da Silva VA, Masur J. Genetically selected winner and loser rats:what was selected? Activitas Nervosa Superior 1978;20:178–85.

2 rain R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
34 E. Nesher et al. / Behavioural B
12] Uyeno ET, Graham RA. The effects of food deprivation of rats on swimming toexhaustion. Behaviour 1966;26:351–6.
13] Malatynska E, Goldenberg R, Shuck L, Haque A, Zamecki P, Crites G, et al. Reduc-tion of submissive behavior in rats: a test for antidepressant drug activity.Pharmacology 2002;64:8–17.
14] Malatynska E, Knapp RJ, Stropova D, Varga E, Yamamura HI. The role of theVal353 residue in antagonist binding to rat CCK-B receptors. Neuroreport1995;6:1325–9.
15] Malatynska E, Kostowski W. The effect of antidepressant drugs on dominancebehavior in rats competing for food. Polish Journal of Pharmacology and Phar-macy 1984;36:531–40.
16] Masur J, Martz RM, Carlini EA. Effects of acute and chronic administration ofcannabis sativa and (−) delta9-trans-tetrahydrocannabinol on the behavior ofrats in an open-field arena. Psychopharmacologia 1971;19:388–97.
17] Gentsch C, Lichtsteiner M, Feer H. Competition for sucrose-pellets in triadsof male Wistar rats: the individuals’ performances are differing but stable.Behavioural Brain Research 1988;27:37–44.
18] Strekalova T, Gorenkova N, Schunk E, Dolgov O, Bartsch D. Selective effects ofcitalopram in a mouse model of stress-induced anhedonia with a control forchronic stress. Behavioural Pharmacology 2006;17:271–87.
19] Kaidanovich-Beilin O, Lipina T, Vukobradovic I, Roder J, Woodgett JR. Assess-ment of social interaction behaviors. Journal of Visualized Experiments 2011,pii: 2473, http://dx.doi.org/10.3791/2473
20] Karvat G, Kimchi T. Systematic autistic-like behavioral phenotyping of 4mouse strains using a novel wheel-running assay. Behavioural Brain Research2012;233:405–14.
21] Defensor EB, Pearson BL, Pobbe RL, Bolivar VJ, Blanchard DC, BlanchardRJ. A novel social proximity test suggests patterns of social avoidance andgaze aversion-like behavior in BTBR T+ tf/J mice. Behavioural Brain Research2011;217:302–8.
22] Nadler JJ, Moy SS, Dold G, Trang D, Simmons N, Perez A, et al. Automated appa-ratus for quantitation of social approach behaviors in mice. Genes, Brain andBehavior 2004;3:303–14.
23] Strekalova T, Spanagel R, Bartsch D, Henn FA, Gass P. Stress-induced anhedo-nia in mice is associated with deficits in forced swimming and exploration.Neuropsychopharmacology 2004;29:2007–17.
24] Burokas A, Gutierrez-Cuesta J, Martin-Garcia E, Maldonado R. Operant modelof frustrated expected reward in mice. Addiction Biology 2012;17:770–82.
25] Holmes MM, Niel L, Anyan JJ, Griffith AT, Monks DA, Forger NG. Effects of Baxgene deletion on social behaviors and neural response to olfactory cues in mice.European Journal of Neuroscience 2011;34:1492–9.
26] Kudryavtseva NN. Use of the “partition” test in behavioral and pharmacologicalexperiments. Neuroscience and Behavioral Physiology 2003;33:461–71.
27] Avgustinovich DF, Gorbach OV, Kudryavtseva NN. Comparative analysis ofanxiety-like behavior in partition and plus-maze tests after agonistic inter-actions in mice. Physiology and Behavior 1997;61:37–43.
28] Malatynska E, Knapp RJ. Dominant–submissive behavior as models of maniaand depression. Neuroscience and Biobehavioral Reviews 2005;29:715–37.
29] Pinhasov A, Crooke J, Rosenthal D, Brenneman D, Malatynska E. Reduc-tion of submissive behavior model for antidepressant drug activity testing:study using a video-tracking system. Behavioural Pharmacology 2005;16:657–64.
30] Malatynska E, Pinhasov A, Crooke JJ, Smith-Swintosky VL, Brenneman DE.Reduction of dominant or submissive behaviors as models for antimanic orantidepressant drug testing: technical considerations. Journal of NeuroscienceMethods 2007;165:175–82.
31] Malatynska E, Rapp R, Harrawood D, Tunnicliff G. Submissive behavior in miceas a test for antidepressant drug activity. Pharmacology Biochemistry andBehavior 2005;82:306–13.
32] Feder Y, Nesher E, Ogran A, Kreinin A, Malatynska E, Yadid G, et al. Selec-tive breeding for dominant and submissive behavior in Sabra mice. Journalof Affective Disorders 2010;126:214–22.
33] Nesher E, Peskov V, Rylova A, Raz O, Pinhasov A. Comparative analysis ofthe behavioral and biomolecular parameters of four mouse strains. Journal ofMolecular Neuroscience 2012;46:276–84.
34] Kudryavtseva NN. Agonistic behavior: a model, experimental studies, and per-spectives. Neuroscience and Behavioral Physiology 2000;30:293–305.
35] Archer J. Tests for emotionality in rats and mice: a review. Animal Behaviour1973;21:205–35.
36] Depino AM, Gross C. Simultaneous assessment of autonomic function andanxiety-related behavior in BALB/c and C57BL/6 mice. Behavioural BrainResearch 2007;177:254–60.
37] Gobshtis N, Ben-Shabat S, Fride E. Antidepressant-induced undesirable weightgain: prevention with rimonabant without interference with behavioral effec-tiveness. European Journal of Pharmacology 2007;554:155–63.
38] Carola V, D’Olimpio F, Brunamonti E, Bevilacqua A, Renzi P, Mangia F. Anxiety-related behaviour in C57BL/6 ↔ BALB/c chimeric mice. Behavioural BrainResearch 2004;150:25–32.
39] Carola V, Gross C. BDNF moderates early environmental risk factors for anxietyin mouse. Genes, Brain and Behavior 2010;9:379–89.
40] Carola V, D’Olimpio F, Brunamonti E, Mangia F, Renzi P. Evaluation of the ele-
vated plus-maze and open-field tests for the assessment of anxiety-relatedbehaviour in inbred mice. Behavioural Brain Research 2002;134:49–57.41] Pellow S, Chopin P, File SE, Briley M. Validation of open:closed arm entries in anelevated plus-maze as a measure of anxiety in the rat. Journal of NeuroscienceMethods 1985;14:149–67.
[
esearch 236 (2013) 225– 235
42] Herrera-Ruiz M, Roman-Ramos R, Zamilpa A, Tortoriello J, Jimenez-Ferrer JE.Flavonoids from Tilia americana with anxiolytic activity in plus-maze test. Jour-nal of Ethnopharmacology 2008;118:312–7.
43] Porsolt RD, Bertin A, Jalfre M. Behavioral despair in mice: a primary screeningtest for antidepressants. Archives Internationales de Pharmacodynamie et deTherapie 1977;229:327–36.
44] Petit-Demouliere B, Chenu F, Bourin M. Forced swimming test in mice: a reviewof antidepressant activity. Psychopharmacology (Berl) 2005;177:245–55.
45] Shieh CH, Hong CJ, Huang YH, Tsai SJ. Potential antidepressant propertiesof cysteamine on hippocampal BDNF levels and behavioral despair in mice.Progress in Neuro-Psychopharmacology and Biological Psychiatry 2008;32:1590–4.
46] Sugimoto Y, Yamamoto M, Tagawa N, Kobayashi Y, Mitsui-Saitoh K, Hotta Y,et al. Differences between mice strains in response to paroxetine in the forcedswimming test: involvement of serotonergic or noradrenergic systems. Euro-pean Journal of Pharmacology 2011;672:121–5.
47] Guzzetti S, Calcagno E, Canetta A, Sacchetti G, Fracasso C, Caccia S, et al. Straindifferences in paroxetine-induced reduction of immobility time in the forcedswimming test in mice: role of serotonin. European Journal of Pharmacology2008;594:117–24.
48] Vissiennon C, Nieber K, Kelber O, Butterweck V. Route of administrationdetermines the anxiolytic activity of the flavonols kaempferol, quercetinand myricetin – are they prodrugs? Journal of Nutritional Biochemistry2012;23:733–40.
49] O’Brien WT, Harper AD, Jove F, Woodgett JR, Maretto S, Piccolo S, et al. Glycogensynthase kinase-3beta haploinsufficiency mimics the behavioral and moleculareffects of lithium. Journal of Neuroscience 2004;24:6791–8.
50] Karyo R, Eskira Y, Pinhasov A, Belmaker R, Agam G, Eldar-Finkelman H.Identification of eukaryotic elongation factor-2 as a novel cellular target oflithium and glycogen synthase kinase-3. Molecular and Cellular Neurosciences2010;45:449–55.
51] Sapolsky RM. The influence of social hierarchy on primate health. Science2005;308:648–52.
52] Ginsburg BE. Social behavior and social hierarchy in the formation of person-ality profiles in animals. Proceedings of the Annual Meeting of the AmericanPsychopathological Association 1967;55:95–114.
53] Gottier RF. The dominance–submission hierarchy in the social behavior of thedomestic chicken. Journal of Genetic Psychology 1968;112:205–26.
54] Huhman KL. Social conflict models: can they inform us about human psy-chopathology. Hormones and Behavior 2006;50:640–6.
55] Coutellier L, Friedrich AC, Failing K, Wurbel H. Variations in the postnatalmaternal environment in mice: effects on maternal behaviour and behaviouraland endocrine responses in the adult offspring. Physiology and Behavior2008;93:395–407.
56] Macri S, Wurbel H. Developmental plasticity of HPA and fear responses in rats:a critical review of the maternal mediation hypothesis. Hormones and Behavior2006;50:667–80.
57] Meaney MJ. Maternal care, gene expression, and the transmission of individualdifferences in stress reactivity across generations. Annual Review of Neuro-science 2001;24:1161–92.
58] Levine S. Developmental determinants of sensitivity and resistance to stress.Psychoneuroendocrinology 2005;30:939–46.
59] Schechter M, Pinhasov A, Weller A, Fride E. Blocking the postpartum mousedam’s CB1 receptors impairs maternal behavior as well as offspring develop-ment and their adult social–emotional behavior. Behavioural Brain Research2012;226:481–92.
60] Hari R, Kujala MV. Brain basis of human social interaction: from concepts tobrain imaging. Physiological Reviews 2009;89:453–79.
61] Price JS. Evolutionary aspects of anxiety disorders. Dialogues in Clinical Neuro-science 2003;5:223–36.
62] Klein JT, Shepherd SV, Platt ML. Social attention and the brain. Current Biology2009;19:R958–62.
63] Nucci L. Culture, universals, and the personal. New Directions for Child Devel-opment 1997:5–22.
64] Champion L. Social relationships and social roles. Clinical Psychology & Psy-chotherapy 2012;19:113–23.
65] Murali V, Oyebode F. Poverty, social inequality and mental health. Advances inPsychiatric Treatment 2004;10:216–24.
66] Diaz JL. The psychobiology of aggression and violence: bioethical implications.International Social Science Journal 2010;61:233–45.
67] Sneddon LU, Schmidt R, Fang Y, Cossins AR. Molecular correlates of socialdominance: a novel role for ependymin in aggression. PLoS ONE 2011;6:e18181.
68] O’Donnell KC, Gould TD. The behavioral actions of lithium in rodent models:leads to develop novel therapeutics. Neuroscience and Biobehavioral Reviews2007;31:932–62.
69] Youngs RM, Chu MS, Meloni EG, Naydenov A, Carlezon Jr WA, Konradi C.Lithium administration to preadolescent rats causes long-lasting increasesin anxiety-like behavior and has molecular consequences. Journal of Neuro-science 2006;26:6031–9.
70] Juetten J, Einat H. Behavioral differences in black Swiss mice from separate
colonies: implications for modeling domains of mania. Behavioural Pharma-cology 2012;23:211–4.71] Hannah-Poquette C, Anderson GW, Flaisher-Grinberg S, Wang J, MeinerdingTM, Einat H. Modeling mania: further validation for Black Swiss mice as modelanimals. Behavioural Brain Research 2011;223:222–6.

rain R
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
[
[
E. Nesher et al. / Behavioural B
72] Bauer M, Forsthoff A, Baethge C, Adli M, Berghofer A, Dopfmer S, et al.Lithium augmentation therapy in refractory depression-update 2002. Euro-pean Archives of Psychiatry and Clinical Neuroscience 2003;253:132–9.
73] Ballenger J. Benzodiazepine receptors agonists and antagonists. In: Sadock BJ,Sadock VA, editors. Kaplan & Sadock’s comprehensive textbook of psychiatry.Philadelphia: Lippincott Williams & Wilkins; 2000. p. 2317–23.
74] Aronson J. Meyler’s side effects of drugs: the international encyclopedia ofadverse drug reactions and interactions. 15th ed. Oxford: Elsevier; 2006.
75] Andreen L, Nyberg S, Turkmen S, van Wingen G, Fernandez G, BackstromT. Sex steroid induced negative mood may be explained by the para-doxical effect mediated by GABAA modulators. Psychoneuroendocrinology2009;34:1121–32.
76] Miczek KA, Fish EW, De Bold JF. Neurosteroids, GABAA receptors, and escalatedaggressive behavior. Hormones and Behavior 2003;44:242–57.
77] Gourley SL, Debold JF, Yin W, Cook J, Miczek KA. Benzodiazepines heightenedaggressive behavior in rats: reduction by GABA(A)/alpha(1) receptor antago-nists. Psychopharmacology (Berl) 2005;178:232–40.
78] Hall RC, Zisook S. Paradoxical reactions to benzodiazepines. British Journal ofClinical Pharmacology 1981;11(Suppl. 1):99S–104S.
79] Mancuso CE, Tanzi MG, Gabay M. Paradoxical reactions to benzodi-
azepines: literature review and treatment options. Pharmacotherapy 2004;24:1177–85.80] Jones KA, Nielsen S, Bruno R, Frei M, Lubman DI. Benzodiazepines – their rolein aggression and why GPs should prescribe with caution. Australian FamilyPhysician 2011;40:862–5.
[
esearch 236 (2013) 225– 235 235
81] Arbanas G, Arbanas D, Dujam K. Adverse effects of benzodiazepines in psychi-atric outpatients. Psychiatria Danubina 2009;21:103–7.
82] Workman DG, Cunningham DG. Effect of psychotropic drugs on aggression ina prison setting. Canadian Family Physician 1975;21:63–6.
83] Porsolt RD. Animal model of depression. Biomedicine 1979;30:139–40.84] Cryan JF, Mombereau C. In search of a depressed mouse: utility of models for
studying depression-related behavior in genetically modified mice. MolecularPsychiatry 2004;9:326–57.
85] Aggarwal A, Sharma DD, Sharma RC, Kumar R. Hypomania as a genuine sideeffect of fluoxetine. Journal of Neuropsychiatry and Clinical Neurosciences2011;23:E23–4.
86] Howland RH. Induction of mania with serotonin reuptake inhibitors. Journal ofClinical Psychopharmacology 1996;16:425–7.
87] Lebegue B. Mania precipitated by fluoxetine. African Journal of Psychiatry1987;144:1620.
88] Goldsmith M, Singh M, Chang K. Antidepressants and psychostimulants inpediatric populations: is there an association with mania. Paediatric Drugs2011;13:225–43.
89] Hausmann A, Hortnagl C, Walpoth M, Fuchs M, Conca A. [Are there substantialreasons for contraindicating antidepressants in bipolar disorder? Part II: facts
or artefacts. Neuropsychiatry 2007;21:131–58.90] Baumer FM, Howe M, Gallelli K, Simeonova DI, Hallmayer J, Chang KD. A pilotstudy of antidepressant-induced mania in pediatric bipolar disorder: charac-teristics, risk factors, and the serotonin transporter gene. Biological Psychiatry2006;60:1005–12.