dhea induces 11-hsd2 by acting on ccaat/ enhancer ... - jasn
TRANSCRIPT

DHEA Induces 11�-HSD2 by Acting on CCAAT/Enhancer-Binding ProteinsZoltan Balazs,*† Roberto A.S. Schweizer,† Felix J. Frey,† Francoise Rohner-Jeanrenaud,‡
and Alex Odermatt*†
*Institute of Molecular and Systems Toxicology, University of Basel, Basel, †Department of Nephrology andHypertension, University of Berne, Berne, and ‡Division of Endocrinology, Diabetology and Nutrition, Departmentof Internal Medicine, University Hospital, Geneva, Switzerland
ABSTRACT11�-Hydroxysteroid dehydrogenase (11�-HSD) type 1 and type 2 catalyze the interconversion of inactiveand active glucocorticoids. Impaired regulation of these enzymes has been associated with obesity,diabetes, hypertension, and cardiovascular disease. Previous studies in animals and humans suggestedthat dehydroepiandrosterone (DHEA) has antiglucocorticoid effects, but the underlying mechanisms areunknown. In this study, DHEA treatment markedly increased mRNA expression and activity of 11�-HSD2in a rat cortical collecting duct cell line and in kidneys of C57BL/6J mice and Sprague-Dawley rats.DHEA-treated rats tended to have reduced urinary corticosterone to 11-dehydrocorticosterone ratios.It was found that CCAAT/enhancer-binding protein-� (C/EBP-�) and C/EBP-� regulated HSD11B2transcription and that DHEA likely modulated the transcription of 11�-HSD2 in a phosphatidylinositol-3kinase/Akt-dependent manner by increasing C/EBP-� mRNA and protein expression. Moreover, it isshown that C/EBP-� and C/EBP-� differentially regulate the expression of 11�-HSD1 and 11�-HSD2. Inconclusion, DHEA induces a shift from 11�-HSD1 to 11�-HSD2 expression, increasing conversion fromactive to inactive glucocorticoids. This provides a possible explanation for the antiglucocorticoid effectsof DHEA.
J Am Soc Nephrol 19: 92–101, 2008. doi: 10.1681/ASN.2007030263
Enhanced glucocorticoid effects, mainly as a resultof locally disturbed glucocorticoid metabolism,contribute to diseases such as hypertension and themetabolic syndrome.1–3 The adrenal steroid hor-mone precursor dehydroepiandrosterone (DHEA)is the most abundant circulating steroid in humans,with peak levels between 20 and 30 yr of age, fol-lowed by a steady, age-dependent decline. Manymetabolic effects, including antiobesity, antidia-betic, and antiaging properties, have been attrib-uted to DHEA.4 In rodents, DHEA attenuated isch-emia/reperfusion-induced oxidative stress andrenal dysfunction,5 showed beneficial effects in di-abetic nephropathy,6,7 and inhibited the age-relateddevelopment of proteinuria.8 DHEA seems tocounteract several adverse effects of excessive glu-cocorticoid action, including a negative correlationbetween DHEA concentrations and body mass in-dex, visceral adiposity, and impaired insulin sensi-
tivity in elderly individuals9; however, the molecu-lar mechanisms underlying these antigluco-corticoid effects remain unclear.
In peripheral tissues, local glucocorticoid me-tabolism is mainly controlled by 11�-hydroxys-teroid dehydrogenase (11�-HSD1) and 11�-HSD2. 11�-HSD1 catalyzes the reduction ofinactive 11-ketoglucocorticoids (cortisone, 11-dehydrocorticosterone) into active 11�-hy-
Received March 2, 2007. Accepted August 2, 2007.
Published online ahead of print. Publication date available atwww.jasn.org.
Correspondence: Dr. Alex Odermatt, Institute of Molecular andSystems Toxicology, Department of Pharmaceutical Sciences,University of Basel, Klingelbergstrasse 50, CH-4056 Basel, Swit-zerland. Phone: �41-61-267-1530; Fax: �41-61-267-1515;E-mail: [email protected]
Copyright © 2008 by the American Society of Nephrology
BASIC RESEARCH www.jasn.org
92 ISSN : 1046-6673/1901-92 J Am Soc Nephrol 19: 92–101, 2008
droxyglucocorticoids (cortisol, corticosterone).2 In vitro,11�-HSD1 catalyzes both reductase and dehydrogenase re-action, whereas in vivo and in intact cells expressing hexose-6-phosphate dehydrogenase, providing co-substrateNADPH, it predominantly acts as a reductase and is essen-tial for glucocorticoid reactivation in metabolically relevanttissues.10 –12 11�-HSD2 acts exclusively as a dehydrogenaseusing co-substrate NAD� and has a pivotal role in miner-alocorticoid target tissues by protecting mineralocorticoidreceptors (MR) from activation by cortisol and in placentaby protecting the fetus from high maternal cortisol.13
Several steroid hormones have been associated with theregulation of 11�-HSD gene expression.14 Homma et al.15
showed that administration of DHEA sulfate decreased BPin spontaneously hypertensive rats. Treated rats had re-duced ratios of serum corticosterone to 11-dehydrocortico-sterone, and the oxidation of corticosterone was increasedin kidneys but decreased in the liver, suggesting that altered11�-HSD activities contributed to the observed effects;however, these authors did not clearly distinguish betweenthe activities of 11�-HSD1 and 11�-HSD2 or take into ac-count that 11�-HSD1 also functions as a dehydrogenasewith NAD�.16 We recently reported reduced 11�-HSD1 ex-pression in response to DHEA in 3T3-L1 adipocytes and inliver and adipose tissue of treated mice.17 We showed thatthe CCAAT/enhancer-binding protein (C/EBP) family ofbasic leucin zipper transcription factors C/EBP-�,C/EBP-�, and C/EBP-� is involved in the DHEA-mediatedregulation of 11�-HSD1 expression. C/EBP can act as tran-scriptional activators as well as repressors.18 C/EBP-� andC/EBP-� were shown to bind directly to the HSD11B1 pro-moter and act in concert to regulate gene expression in livercells19 and adipocytes.20 Whether C/EBP might also modu-late HSD11B2 gene expression has not been investigated.
Here, we examined the impact of DHEA on 11�-HSD2 ex-pression and activity in cultured cells and in kidneys ofSprague-Dawley rats and C57BL/6J mice. We investigatedwhether C/EBP are involved in the transcriptional regulationof 11�-HSD2 and compared their effects with those on 11�-HSD1. In particular, we tested whether C/EBP transcriptionfactors might be responsible for the differential regulation of11�-HSD1 and 11�-HSD2.
RESULTS
DHEA Upregulates 11�-HSD2 Expression and Activityin Renal Cortical Collecting Duct CellsRecently, we reported that DHEA decreases 11�-HSD1 mRNAexpression in cultured cells and C57Bl/6J mice.17 Here, wetested the hypothesis that DHEA has antiglucocorticoid effectsby inducing a switch from 11�-HSD1– dependent activationto 11�-HSD2– dependent inactivation of glucocorticoids. Weinvestigated the impact of DHEA on 11�-HSD2 expressionand activity in RCCD2 rat cortical collecting duct cells.21 Incu-
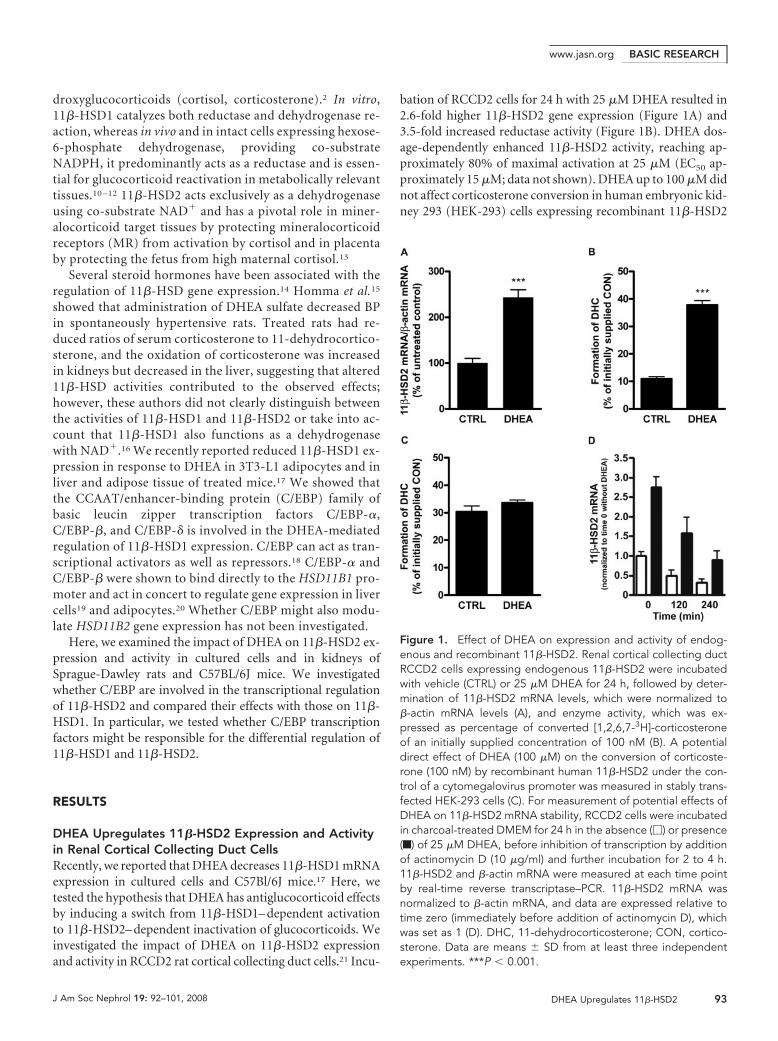
bation of RCCD2 cells for 24 h with 25 �M DHEA resulted in2.6-fold higher 11�-HSD2 gene expression (Figure 1A) and3.5-fold increased reductase activity (Figure 1B). DHEA dos-age-dependently enhanced 11�-HSD2 activity, reaching ap-proximately 80% of maximal activation at 25 �M (EC50 ap-proximately 15 �M; data not shown). DHEA up to 100 �M didnot affect corticosterone conversion in human embryonic kid-ney 293 (HEK-293) cells expressing recombinant 11�-HSD2
Figure 1. Effect of DHEA on expression and activity of endog-enous and recombinant 11�-HSD2. Renal cortical collecting ductRCCD2 cells expressing endogenous 11�-HSD2 were incubatedwith vehicle (CTRL) or 25 �M DHEA for 24 h, followed by deter-mination of 11�-HSD2 mRNA levels, which were normalized to�-actin mRNA levels (A), and enzyme activity, which was ex-pressed as percentage of converted [1,2,6,7-3H]-corticosteroneof an initially supplied concentration of 100 nM (B). A potentialdirect effect of DHEA (100 �M) on the conversion of corticoste-rone (100 nM) by recombinant human 11�-HSD2 under the con-trol of a cytomegalovirus promoter was measured in stably trans-fected HEK-293 cells (C). For measurement of potential effects ofDHEA on 11�-HSD2 mRNA stability, RCCD2 cells were incubatedin charcoal-treated DMEM for 24 h in the absence (�) or presence(f) of 25 �M DHEA, before inhibition of transcription by additionof actinomycin D (10 �g/ml) and further incubation for 2 to 4 h.11�-HSD2 and �-actin mRNA were measured at each time pointby real-time reverse transcriptase–PCR. 11�-HSD2 mRNA wasnormalized to �-actin mRNA, and data are expressed relative totime zero (immediately before addition of actinomycin D), whichwas set as 1 (D). DHC, 11-dehydrocorticosterone; CON, cortico-sterone. Data are means � SD from at least three independentexperiments. ***P � 0.001.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 19: 92–101, 2008 DHEA Upregulates 11�-HSD2 93
under the control of a cytomegalovirus promoter, excluding adirect stimulatory effect of DHEA on enzyme activity (Figure1C). We next examined whether the DHEA-mediated induc-tion of 11�-HSD2 gene expression involves changes in mRNAstability. DHEA treatment increased 11�-HSD2 transcriptionin RCCD2 cells approximately three-fold (Figure 1D). Addi-tion of the transcription inhibitor actinomycin D (10 �g/ml)after 24 h of incubation with vehicle or 25 �M DHEA resultedin a similar decay of 11�-HSD2 mRNA over the course of 4 h,with estimated half-lives of 138 and 148 min, respectively (Fig-ure 1D). Together, these observations suggest that DHEAstimulates 11�-HSD2 activity in RCCD2 cells primarilythrough activation of gene transcription. In contrast to renalcortical collecting duct cells, DHEA did not affect 11�-HSD2expression in Caco-2 and SW620 colon cells (data not shown),suggesting tissue-specific regulation.
DHEA Upregulates 11�-HSD2 Expression and Activityin the KidneyThe effect of DHEA on 11�-HSD2 expression and activity wasthen investigated in vivo by feeding Sprague-Dawley rats stan-
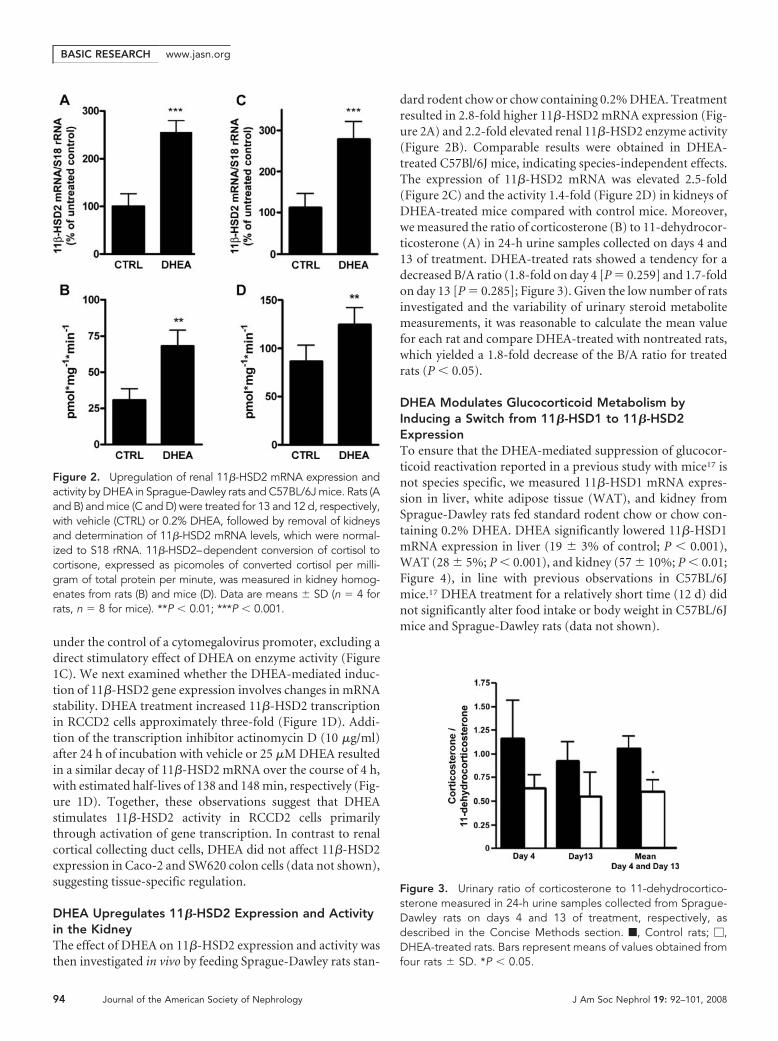
dard rodent chow or chow containing 0.2% DHEA. Treatmentresulted in 2.8-fold higher 11�-HSD2 mRNA expression (Fig-ure 2A) and 2.2-fold elevated renal 11�-HSD2 enzyme activity(Figure 2B). Comparable results were obtained in DHEA-treated C57Bl/6J mice, indicating species-independent effects.The expression of 11�-HSD2 mRNA was elevated 2.5-fold(Figure 2C) and the activity 1.4-fold (Figure 2D) in kidneys ofDHEA-treated mice compared with control mice. Moreover,we measured the ratio of corticosterone (B) to 11-dehydrocor-ticosterone (A) in 24-h urine samples collected on days 4 and13 of treatment. DHEA-treated rats showed a tendency for adecreased B/A ratio (1.8-fold on day 4 [P � 0.259] and 1.7-foldon day 13 [P � 0.285]; Figure 3). Given the low number of ratsinvestigated and the variability of urinary steroid metabolitemeasurements, it was reasonable to calculate the mean valuefor each rat and compare DHEA-treated with nontreated rats,which yielded a 1.8-fold decrease of the B/A ratio for treatedrats (P � 0.05).
DHEA Modulates Glucocorticoid Metabolism byInducing a Switch from 11�-HSD1 to 11�-HSD2ExpressionTo ensure that the DHEA-mediated suppression of glucocor-ticoid reactivation reported in a previous study with mice17 isnot species specific, we measured 11�-HSD1 mRNA expres-sion in liver, white adipose tissue (WAT), and kidney fromSprague-Dawley rats fed standard rodent chow or chow con-taining 0.2% DHEA. DHEA significantly lowered 11�-HSD1mRNA expression in liver (19 � 3% of control; P � 0.001),WAT (28 � 5%; P � 0.001), and kidney (57 � 10%; P � 0.01;Figure 4), in line with previous observations in C57BL/6Jmice.17 DHEA treatment for a relatively short time (12 d) didnot significantly alter food intake or body weight in C57BL/6Jmice and Sprague-Dawley rats (data not shown).
Figure 2. Upregulation of renal 11�-HSD2 mRNA expression andactivity by DHEA in Sprague-Dawley rats and C57BL/6J mice. Rats (Aand B) and mice (C and D) were treated for 13 and 12 d, respectively,with vehicle (CTRL) or 0.2% DHEA, followed by removal of kidneysand determination of 11�-HSD2 mRNA levels, which were normal-ized to S18 rRNA. 11�-HSD2–dependent conversion of cortisol tocortisone, expressed as picomoles of converted cortisol per milli-gram of total protein per minute, was measured in kidney homog-enates from rats (B) and mice (D). Data are means � SD (n � 4 forrats, n � 8 for mice). **P � 0.01; ***P � 0.001.
Figure 3. Urinary ratio of corticosterone to 11-dehydrocortico-sterone measured in 24-h urine samples collected from Sprague-Dawley rats on days 4 and 13 of treatment, respectively, asdescribed in the Concise Methods section. f, Control rats; �,DHEA-treated rats. Bars represent means of values obtained fromfour rats � SD. *P � 0.05.
BASIC RESEARCH www.jasn.org
94 Journal of the American Society of Nephrology J Am Soc Nephrol 19: 92–101, 2008
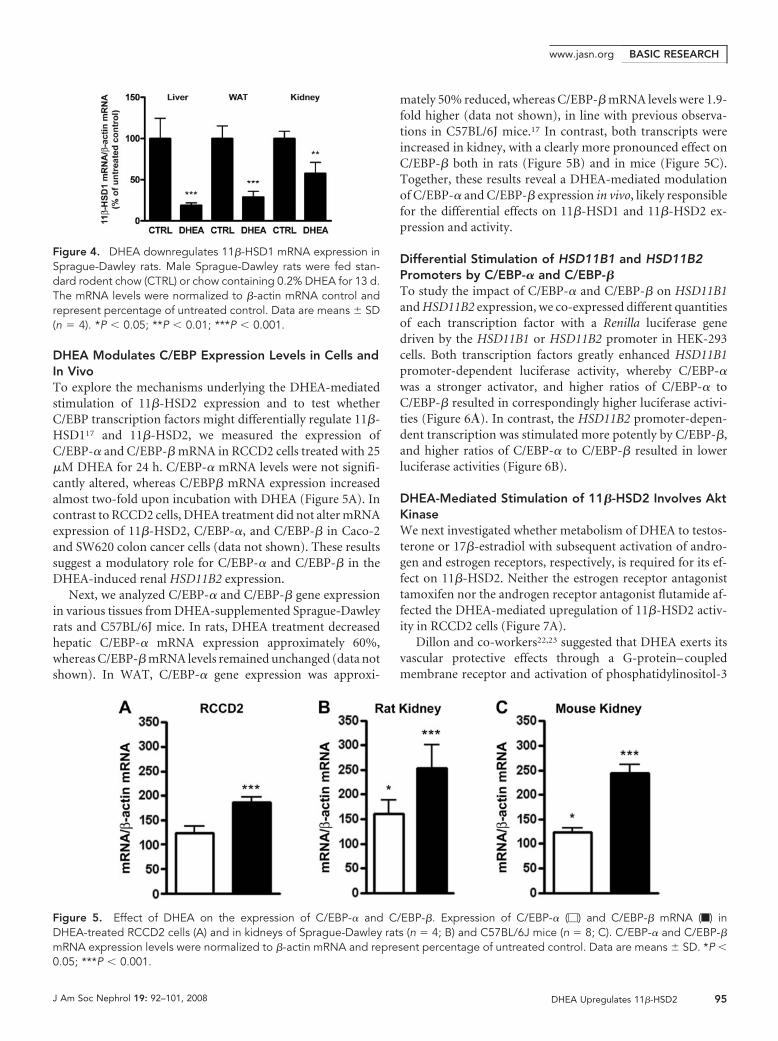
DHEA Modulates C/EBP Expression Levels in Cells andIn VivoTo explore the mechanisms underlying the DHEA-mediatedstimulation of 11�-HSD2 expression and to test whetherC/EBP transcription factors might differentially regulate 11�-HSD117 and 11�-HSD2, we measured the expression ofC/EBP-� and C/EBP-� mRNA in RCCD2 cells treated with 25�M DHEA for 24 h. C/EBP-� mRNA levels were not signifi-cantly altered, whereas C/EBP� mRNA expression increasedalmost two-fold upon incubation with DHEA (Figure 5A). Incontrast to RCCD2 cells, DHEA treatment did not alter mRNAexpression of 11�-HSD2, C/EBP-�, and C/EBP-� in Caco-2and SW620 colon cancer cells (data not shown). These resultssuggest a modulatory role for C/EBP-� and C/EBP-� in theDHEA-induced renal HSD11B2 expression.
Next, we analyzed C/EBP-� and C/EBP-� gene expressionin various tissues from DHEA-supplemented Sprague-Dawleyrats and C57BL/6J mice. In rats, DHEA treatment decreasedhepatic C/EBP-� mRNA expression approximately 60%,whereas C/EBP-� mRNA levels remained unchanged (data notshown). In WAT, C/EBP-� gene expression was approxi-
mately 50% reduced, whereas C/EBP-� mRNA levels were 1.9-fold higher (data not shown), in line with previous observa-tions in C57BL/6J mice.17 In contrast, both transcripts wereincreased in kidney, with a clearly more pronounced effect onC/EBP-� both in rats (Figure 5B) and in mice (Figure 5C).Together, these results reveal a DHEA-mediated modulationof C/EBP-� and C/EBP-� expression in vivo, likely responsiblefor the differential effects on 11�-HSD1 and 11�-HSD2 ex-pression and activity.
Differential Stimulation of HSD11B1 and HSD11B2Promoters by C/EBP-� and C/EBP-�To study the impact of C/EBP-� and C/EBP-� on HSD11B1and HSD11B2 expression, we co-expressed different quantitiesof each transcription factor with a Renilla luciferase genedriven by the HSD11B1 or HSD11B2 promoter in HEK-293cells. Both transcription factors greatly enhanced HSD11B1promoter-dependent luciferase activity, whereby C/EBP-�was a stronger activator, and higher ratios of C/EBP-� toC/EBP-� resulted in correspondingly higher luciferase activi-ties (Figure 6�). In contrast, the HSD11B2 promoter-depen-dent transcription was stimulated more potently by C/EBP-�,and higher ratios of C/EBP-� to C/EBP-� resulted in lowerluciferase activities (Figure 6B).
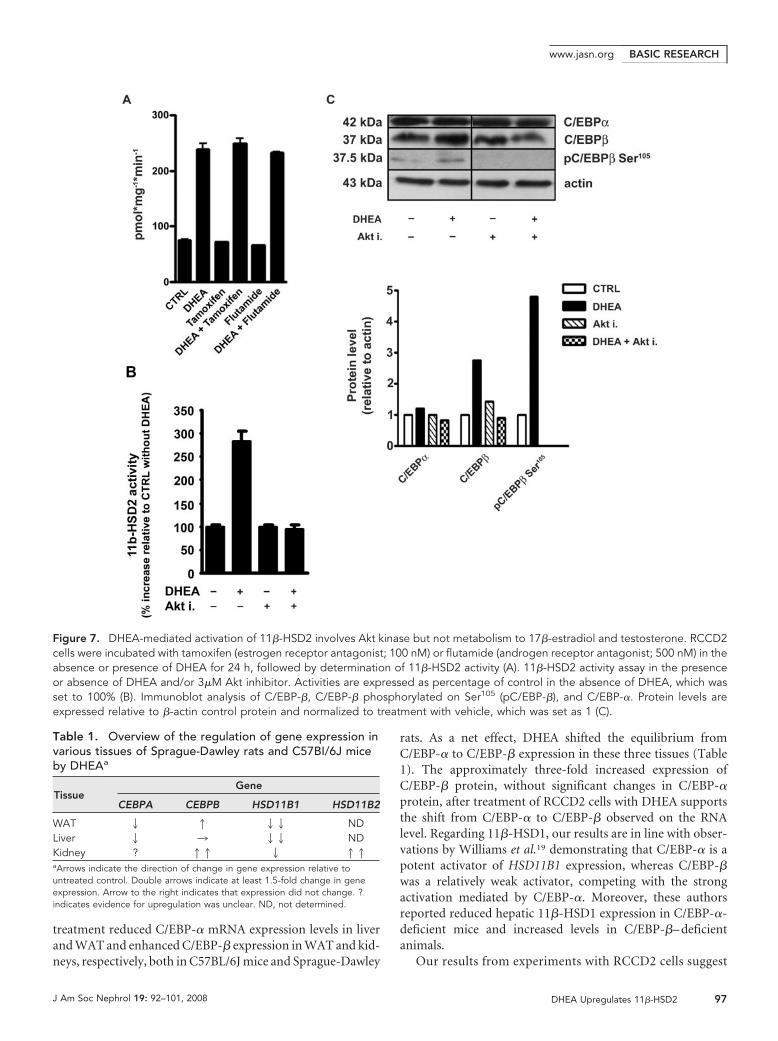
DHEA-Mediated Stimulation of 11�-HSD2 Involves AktKinaseWe next investigated whether metabolism of DHEA to testos-terone or 17�-estradiol with subsequent activation of andro-gen and estrogen receptors, respectively, is required for its ef-fect on 11�-HSD2. Neither the estrogen receptor antagonisttamoxifen nor the androgen receptor antagonist flutamide af-fected the DHEA-mediated upregulation of 11�-HSD2 activ-ity in RCCD2 cells (Figure 7A).
Dillon and co-workers22,23 suggested that DHEA exerts itsvascular protective effects through a G-protein– coupledmembrane receptor and activation of phosphatidylinositol-3
Figure 4. DHEA downregulates 11�-HSD1 mRNA expression inSprague-Dawley rats. Male Sprague-Dawley rats were fed stan-dard rodent chow (CTRL) or chow containing 0.2% DHEA for 13 d.The mRNA levels were normalized to �-actin mRNA control andrepresent percentage of untreated control. Data are means � SD(n � 4). *P � 0.05; **P � 0.01; ***P � 0.001.
Figure 5. Effect of DHEA on the expression of C/EBP-� and C/EBP-�. Expression of C/EBP-� (�) and C/EBP-� mRNA (f) inDHEA-treated RCCD2 cells (A) and in kidneys of Sprague-Dawley rats (n � 4; B) and C57BL/6J mice (n � 8; C). C/EBP-� and C/EBP-�mRNA expression levels were normalized to �-actin mRNA and represent percentage of untreated control. Data are means � SD. *P �0.05; ***P � 0.001.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 19: 92–101, 2008 DHEA Upregulates 11�-HSD2 95
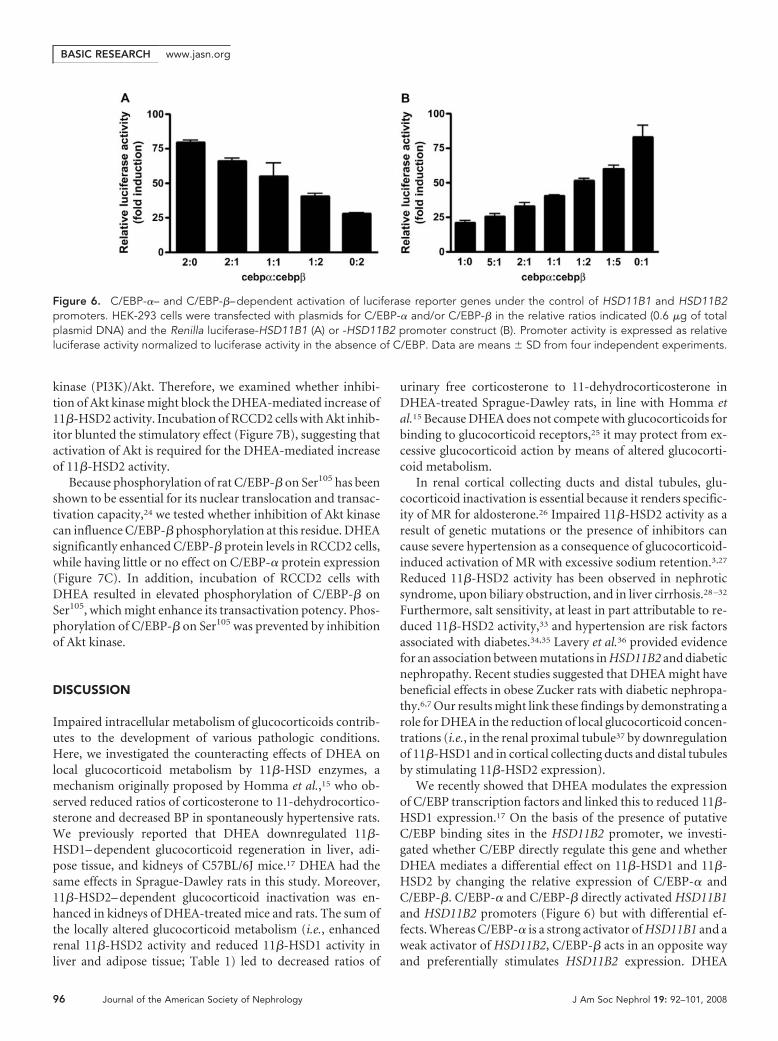
kinase (PI3K)/Akt. Therefore, we examined whether inhibi-tion of Akt kinase might block the DHEA-mediated increase of11�-HSD2 activity. Incubation of RCCD2 cells with Akt inhib-itor blunted the stimulatory effect (Figure 7B), suggesting thatactivation of Akt is required for the DHEA-mediated increaseof 11�-HSD2 activity.
Because phosphorylation of rat C/EBP-� on Ser105 has beenshown to be essential for its nuclear translocation and transac-tivation capacity,24 we tested whether inhibition of Akt kinasecan influence C/EBP-� phosphorylation at this residue. DHEAsignificantly enhanced C/EBP-� protein levels in RCCD2 cells,while having little or no effect on C/EBP-� protein expression(Figure 7C). In addition, incubation of RCCD2 cells withDHEA resulted in elevated phosphorylation of C/EBP-� onSer105, which might enhance its transactivation potency. Phos-phorylation of C/EBP-� on Ser105 was prevented by inhibitionof Akt kinase.
DISCUSSION
Impaired intracellular metabolism of glucocorticoids contrib-utes to the development of various pathologic conditions.Here, we investigated the counteracting effects of DHEA onlocal glucocorticoid metabolism by 11�-HSD enzymes, amechanism originally proposed by Homma et al.,15 who ob-served reduced ratios of corticosterone to 11-dehydrocortico-sterone and decreased BP in spontaneously hypertensive rats.We previously reported that DHEA downregulated 11�-HSD1– dependent glucocorticoid regeneration in liver, adi-pose tissue, and kidneys of C57BL/6J mice.17 DHEA had thesame effects in Sprague-Dawley rats in this study. Moreover,11�-HSD2– dependent glucocorticoid inactivation was en-hanced in kidneys of DHEA-treated mice and rats. The sum ofthe locally altered glucocorticoid metabolism (i.e., enhancedrenal 11�-HSD2 activity and reduced 11�-HSD1 activity inliver and adipose tissue; Table 1) led to decreased ratios of
urinary free corticosterone to 11-dehydrocorticosterone inDHEA-treated Sprague-Dawley rats, in line with Homma etal.15 Because DHEA does not compete with glucocorticoids forbinding to glucocorticoid receptors,25 it may protect from ex-cessive glucocorticoid action by means of altered glucocorti-coid metabolism.
In renal cortical collecting ducts and distal tubules, glu-cocorticoid inactivation is essential because it renders specific-ity of MR for aldosterone.26 Impaired 11�-HSD2 activity as aresult of genetic mutations or the presence of inhibitors cancause severe hypertension as a consequence of glucocorticoid-induced activation of MR with excessive sodium retention.3,27
Reduced 11�-HSD2 activity has been observed in nephroticsyndrome, upon biliary obstruction, and in liver cirrhosis.28 –32
Furthermore, salt sensitivity, at least in part attributable to re-duced 11�-HSD2 activity,33 and hypertension are risk factorsassociated with diabetes.34,35 Lavery et al.36 provided evidencefor an association between mutations in HSD11B2 and diabeticnephropathy. Recent studies suggested that DHEA might havebeneficial effects in obese Zucker rats with diabetic nephropa-thy.6,7 Our results might link these findings by demonstrating arole for DHEA in the reduction of local glucocorticoid concen-trations (i.e., in the renal proximal tubule37 by downregulationof 11�-HSD1 and in cortical collecting ducts and distal tubulesby stimulating 11�-HSD2 expression).
We recently showed that DHEA modulates the expressionof C/EBP transcription factors and linked this to reduced 11�-HSD1 expression.17 On the basis of the presence of putativeC/EBP binding sites in the HSD11B2 promoter, we investi-gated whether C/EBP directly regulate this gene and whetherDHEA mediates a differential effect on 11�-HSD1 and 11�-HSD2 by changing the relative expression of C/EBP-� andC/EBP-�. C/EBP-� and C/EBP-� directly activated HSD11B1and HSD11B2 promoters (Figure 6) but with differential ef-fects. Whereas C/EBP-� is a strong activator of HSD11B1 and aweak activator of HSD11B2, C/EBP-� acts in an opposite wayand preferentially stimulates HSD11B2 expression. DHEA
Figure 6. C/EBP-�– and C/EBP-�–dependent activation of luciferase reporter genes under the control of HSD11B1 and HSD11B2promoters. HEK-293 cells were transfected with plasmids for C/EBP-� and/or C/EBP-� in the relative ratios indicated (0.6 �g of totalplasmid DNA) and the Renilla luciferase-HSD11B1 (A) or -HSD11B2 promoter construct (B). Promoter activity is expressed as relativeluciferase activity normalized to luciferase activity in the absence of C/EBP. Data are means � SD from four independent experiments.
BASIC RESEARCH www.jasn.org
96 Journal of the American Society of Nephrology J Am Soc Nephrol 19: 92–101, 2008
treatment reduced C/EBP-� mRNA expression levels in liverand WAT and enhanced C/EBP-� expression in WAT and kid-neys, respectively, both in C57BL/6J mice and Sprague-Dawley
rats. As a net effect, DHEA shifted the equilibrium fromC/EBP-� to C/EBP-� expression in these three tissues (Table1). The approximately three-fold increased expression ofC/EBP-� protein, without significant changes in C/EBP-�protein, after treatment of RCCD2 cells with DHEA supportsthe shift from C/EBP-� to C/EBP-� observed on the RNAlevel. Regarding 11�-HSD1, our results are in line with obser-vations by Williams et al.19 demonstrating that C/EBP-� is apotent activator of HSD11B1 expression, whereas C/EBP-�was a relatively weak activator, competing with the strongactivation mediated by C/EBP-�. Moreover, these authorsreported reduced hepatic 11�-HSD1 expression in C/EBP-�-deficient mice and increased levels in C/EBP-�– deficientanimals.
Our results from experiments with RCCD2 cells suggest
Figure 7. DHEA-mediated activation of 11�-HSD2 involves Akt kinase but not metabolism to 17�-estradiol and testosterone. RCCD2cells were incubated with tamoxifen (estrogen receptor antagonist; 100 nM) or flutamide (androgen receptor antagonist; 500 nM) in theabsence or presence of DHEA for 24 h, followed by determination of 11�-HSD2 activity (A). 11�-HSD2 activity assay in the presenceor absence of DHEA and/or 3�M Akt inhibitor. Activities are expressed as percentage of control in the absence of DHEA, which wasset to 100% (B). Immunoblot analysis of C/EBP-�, C/EBP-� phosphorylated on Ser105 (pC/EBP-�), and C/EBP-�. Protein levels areexpressed relative to �-actin control protein and normalized to treatment with vehicle, which was set as 1 (C).
Table 1. Overview of the regulation of gene expression invarious tissues of Sprague-Dawley rats and C57BI/6J miceby DHEAa
TissueGene
CEBPA CEBPB HSD11B1 HSD11B2
WAT 2 1 22 NDLiver 2 3 22 NDKidney ? 11 2 11aArrows indicate the direction of change in gene expression relative tountreated control. Double arrows indicate at least 1.5-fold change in geneexpression. Arrow to the right indicates that expression did not change. ?indicates evidence for upregulation was unclear. ND, not determined.
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 19: 92–101, 2008 DHEA Upregulates 11�-HSD2 97

that the effects of DHEA are not due to its metabolism to17�-estradiol or testosterone with subsequent activation ofestrogen receptor and androgen receptor, because neitherantagonists of these receptors (Figure 7A) nor the aro-matase inhibitor formestane (data not shown) had any in-fluence. Instead, inhibition of Akt kinase prevented theDHEA-induced increase of C/EBP-� protein expressionand its phosphorylation on Ser105 as well as stimulation of11�-HSD2 activity. Recent evidence suggested that DHEAmight act through G-protein– coupled membrane receptorsand subsequent activation of PI3K/Akt.23 Phosphorylationof Ser105 on rat C/EBP-� has been shown to enhance itstransactivation potency.24,38 Moreover, activation of PI3K/Akt has been reported to be important for nuclear translo-cation and DNA binding of C/EBP-�.39 Together, these ob-servations support a role for PI3K/Akt in the DHEA-mediated regulation of the ratio of C/EBP-� to C/EBP-�protein expression as well as their phosphorylation state.Elucidation of the mechanism underlying DHEA action re-quires further studies. The phosphorylation state of C/EBPis dynamically regulated by various kinases and phospha-tases, and the pathway downstream from the proposed G-protein– coupled membrane receptor is yet to be character-ized. Exploration of the mechanisms regulating thetransactivation potency of C/EBP-� and C/EBP-� by DHEAis important to understand the fine-tuned control of 11�-HSD1 and 11�-HSD2 activities, which determines glu-cocorticoid action in a highly tissue-specific manner. Byaltering expression and transcriptional activity of C/EBP-�and C/EBP-�, DHEA may exert its antiglucocorticoid ac-tion in C57BL/6J mice and Sprague-Dawley rats by inducinga switch from 11�-HSD1 reductase activity (liver and fat) to11�-HSD2 dehydrogenase activity (kidney), thereby reduc-ing the overall intracellular glucocorticoid availability.
A similar but opposite switch (increased 11�-HSD1 reduc-tase and reduced 11�-HSD2 dehydrogenase activity) was pre-viously shown for the cytokine TNF-�.40 – 42 Importantly, inobese Zucker rats, DHEA treatment reduced serum TNF-�,43
suggesting that the two pathways influence each other. Thedifferential regulation of 11�-HSD1 and 11�-HSD2 is of phys-iologic relevance, because it modulates the intracellular avail-ability of active glucocorticoids.
Our findings indicate that the tissue-specific expressionof 11�-HSD1 and 11�-HSD2 is tightly controlled byC/EBP-� and C/EBP-�. Mechanisms by which DHEA in-duces a switch from 11�-HSD1– dependent glucocorticoidactivation to 11�-HSD2– dependent inactivation, therebycounteracting excessive glucocorticoid action, may involvemodulation of PI3K/Akt and a shift in the expression fromC/EBP-� to C/EBP-�. Although these effects of DHEA arebeneficial in metabolically active tissues to avoid glucocor-ticoid excess, the uncontrolled consumption of DHEAmight cause adverse effects in processes such as cell prolif-eration and differentiation, in which both C/EBP and 11�-HSD also play crucial roles.
CONCISE METHODS
MaterialsCell culture media, Oligo-dT, and Superscript II reverse transcriptase
were from Invitrogen (Carlsbad, CA); steroids were from Steraloids
(Wilton, NH); [1,2,6,7-3H]-cortisol and [1,2,6,7-3H]-corticosterone
were from Amersham Health AG (Wadenswil, Switzerland); [1,2,6,7-3H]-cortisone was from American Radiolabeled Chemicals (St. Louis,
MO), InSolution Akt inhibitor VIII (isozyme selective) was from Cal-
biochem (VWR Int., Dietikon, Switzerland); and reagents for real-
time PCR were from Applied Biosystems (Foster City, CA). All other
chemicals were from Fluka AG (Buchs, Switzerland).
Cell Culture, Transfection, and Transactivation StudiesHEK-293 cells and colon cancer– derived SW620 and Caco-2 cells
were grown in DMEM and supplemented with penicillin, streptomy-
cin, L-glutamine, and 10% FCS. The rat renal cortical collecting duct
cell line RCCD2, provided by Dr. N. Farman (College de France,
Paris, France), was cultured as described previously.21 HEK-293 cells,
grown to 90% confluence, were transfected using the Ca2�-phos-
phate method with different combinations of plasmids containing
full-length rat C/EBP-� or C/EBP-� (in pcDNA3.1; provided by Dr.
P. Johnson,44 National Cancer Institute, Frederick, MD); pGL3-de-
rived plasmids containing the Renilla luciferase gene under the con-
trol of human HSD11B1 or HSD11B2 promoter, consisting of ap-
proximately 1300 bp of sequence upstream of the initiation codon
(provided by Dr. Brigitte Frey,42 Department of Nephrology, Berne,
Switzerland); and �-galactosidase control plasmid. Six hours after
transfection, the medium was replaced by charcoal-treated DMEM,
followed by incubation for another 24 h and determination of lucif-
erase activity using the Dual-Light system (Tropix, Bedford, MA) and
a Fluoroskan Ascent FL luminometer (Thermo Electron Corp, Van-
taa, Finland).
11�-HSD2 mRNA DecayRCCD2 cells were incubated in the absence or presence of 25 �M
DHEA for 24 h, followed by blocking transcription with 10 �g/ml
actinomycin D and further incubation for 2 to 4 h. RNA was isolated,
and 11�-HSD2 and �-actin mRNA were analyzed by real-time re-
verse transcriptase–PCR.
Immunoblot AnalysisCells were washed with PBS and lysed in buffer containing 25 mM
Tris-HCl (pH 7.6), 150 mM NaCl, 1% NP-40, 1% sodium deoxy-
cholate, 0.1% SDS, 1 mM sodium vanadate, 1 mM PMSF, and pro-
teinase inhibitor cocktail (Roche Diagnostics, Rotkreuz, Switzerland).
Equivalent amounts of protein (20 to 30 �g) were loaded to SDS-
PAGE and subsequently transferred to polyvinylidene difluoride
membranes (Hybond-P; Amersham-Pharmacia Biotech, Piscataway,
NJ), followed by incubation with antibodies against C/EBP-�,
C/EBP-� (Santa Cruz Biotechnology, Santa Cruz, CA), C/EBP-�
phosphorylated on Ser105 (Cell Signaling Technology, Beverly, MA),
and �-actin (Santa Cruz Biotechnology) as a loading control. Total
protein was determined using the BCA assay kit (Pierce, Rockford,
IL). Proteins were detected using horseradish peroxidase– conjugated
BASIC RESEARCH www.jasn.org
98 Journal of the American Society of Nephrology J Am Soc Nephrol 19: 92–101, 2008

secondary antibodies (Santa Cruz Biotechnology) and ECL enhanced
chemiluminescence reagent (Amersham-Pharmacia Biotech).
Animal ExperimentsAll animal experiments were approved by the Ethical Committee for
Animal Research of the University of Berne, equivalent to the NIH
guidelines. Male C57BL/6J mice (20 to 22 g body weight; Charles
River, Paris, France) were kept in individual cages. For feeding, they
were transferred to other cages for 2 h and allowed access to 5 g of
standard rodent chow and tap water ad libitum. After 5 d of adapta-
tion, eight animals per condition were fed for 12 d with control chow
or with chow containing DHEA (0.4% DHEA in g/g dry food, corre-
sponding to 0.2% DHEA in fresh food). Body weight and food and
water intake were recorded daily. No significant changes in body
weight were observed during the treatment. On day 12, mice were
killed and tissues were harvested. Serum DHEA sulfate was measured
using a chemiluminescence immunoassay kit and the Immulite One
apparatus following the manufacturer’s instruction (Diagnostics
Products, Los Angeles, CA).
Sprague-Dawley rats (200 to 220 g body weight; Charles River)
were housed in groups of four in a 12:12-h light-dark cycle with stan-
dard laboratory chow and tap water ad libitum. Adaptation to indi-
vidual cages was for 2 d. Four rats received control chow and four
others received chow supplemented with DHEA (0.2% DHEA in
fresh food) for 13 d before they were killed. Plasma samples and major
organs were stored at �70°C.
Analysis of Steroids by Gas Chromatography–MassSpectrometryEvery morning at 9 a.m., 24-h urine samples were collected from rats.
The analysis of urinary steroids was described previously,45 with the
exception that medroxyprogesterone was used as an internal stan-
dard.
RNA Isolation and AnalysisTotal RNA was extracted from adherent cultured cells or from animal
tissues, and mRNA levels were analyzed as described previously.17
Data from the analysis of the relative expression of each gene versus
S18 rRNA or �-actin mRNA was determined using the 2���CT meth-
od.46 Cycle threshold (CT) values were determined from at least four
independent cell experiments or from tissue samples of eight mice or
four Sprague-Dawley rats, each measured in triplicate.
Determination of 11�-HSD2 Enzyme Activity11�-HSD2– dependent conversion of cortisol to cortisone or cortico-
sterone to 11�-dehydrocorticosterone was measured in intact cells as
described previously.47,48 For determination of 11�-HSD2 activity in
tissue homogenates, kidneys were pulverized in liquid nitrogen and
homogenates were incubated for 10 min in buffer TS2 (100 mM NaCl,
1 mM EGTA, 1 mM EDTA, 1 mM MgCl2, 250 mM sucrose, and 20
mM Tris-HCl [pH 7.4]) containing 250 �M NAD� and cortisol or
corticosterone at a final concentration of 100 nM. Steroids were sep-
arated by thin layer chromatography and analyzed by scintillation
counting as described previously.49
Statistical AnalysisStatistical comparisons between groups were performed by ANOVA
or unpaired t test where appropriate. P � 0.05 was considered signif-
icant.
ACKNOWLEDGMENTS
This work was supported by grants from the Cloetta Research Foun-
dation (A.O.), the Swiss National Science Foundation (310000-
112279 to A.O., 3100A0-102153 to F.J.F., and 3100A0-105889 to
F.R.J.), and the Swiss Cancer League (OCS-01402-08-2003 to A.O.).
A.O. is a Novartis Research Foundation Professor. F.R.J. was sup-
ported by the European Community (EC) FP6 funding (LSHM-CT-
2003-503041).
We thank Heidi Jamin, Marcella Klein, and Balazs Legeza for ex-
cellent technical assistance, Dr. Nicolette Farman (Paris, France) for
RCCD2 cells, Dr. Peter Johnson (Frederick, MD) for C/EBP expres-
sion constructs, and Dr. Brigitte Frey (Berne, Switzerland) for
HSD11B1/2 reporter plasmids.
DISCLOSURESNone.
REFERENCES
1. Seckl JR, Walker BR: 11Beta-hydroxysteroid dehydrogenase type 1 asa modulator of glucocorticoid action: From metabolism to memory.Trends Endocrinol Metab 15: 418–424, 2004
2. Atanasov AG, Odermatt A: Readjusting the glucocorticoid balance:An opportunity for modulators of 11b-hydroxysteroid dehydrogenasetype 1 activity? Endocr Metab Immune Disord Drug Targets 7: 125–140, 2007
3. Frey FJ, Odermatt A, Frey BM: Glucocorticoid-mediated mineralocor-ticoid receptor activation and hypertension. Curr Opin Nephrol Hy-pertens 13: 451–458, 2004
4. Tchernof A, Labrie F: Dehydroepiandrosterone, obesity and cardio-vascular disease risk: A review of human studies. Eur J Endocrinol 151:1–14, 2004
5. Aragno M, Cutrin JC, Mastrocola R, Perrelli MG, Restivo F, Poli G,Danni O, Boccuzzi G: Oxidative stress and kidney dysfunction due toischemia/reperfusion in rat: Attenuation by dehydroepiandrosterone.Kidney Int 64: 836–843, 2003
6. Aragno M, Parola S, Brignardello E, Manti R, Betteto S, Tamagno E,Danni O, Boccuzzi G: Oxidative stress and eicosanoids in the kidneysof hyperglycemic rats treated with dehydroepiandrosterone. FreeRadic Biol Med 31: 935–942, 2001
7. Richards RJ, Porter JR, Inserra F, Ferder LF, Stella I, Reisin E, Svec F:Effects of dehydroepiandrosterone and quinapril on nephropathy inobese Zucker rats. Kidney Int 59: 37–43, 2001
8. Pashko LL, Fairman DK, Schwartz AG: Inhibition of proteinuria devel-opment in aging Sprague-Dawley rats and C57BL/6 mice by long-termtreatment with dehydroepiandrosterone. J Gerontol 41: 433–438,1986
9. Villareal DT, Holloszy JO: Effect of DHEA on abdominal fat and insulinaction in elderly women and men: A randomized controlled trial.JAMA 292: 2243–2248, 2004
10. Frick C, Atanasov AG, Arnold P, Ozols J, Odermatt A: Appropriate
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 19: 92–101, 2008 DHEA Upregulates 11�-HSD2 99

function of 11beta-hydroxysteroid dehydrogenase type 1 in the en-doplasmic reticulum lumen is dependent on its N-terminal regionsharing similar topological determinants with 50-kDa esterase. J BiolChem 279: 31131–31138, 2004
11. Atanasov AG, Nashev LG, Schweizer RA, Frick C, Odermatt A: Hex-ose-6-phosphate dehydrogenase determines the reaction direction of11beta-hydroxysteroid dehydrogenase type 1 as an oxoreductase.FEBS Lett 571: 129–133, 2004
12. Lavery GG, Walker EA, Draper N, Jeyasuria P, Marcos J, ShackletonCH, Parker KL, White PC, Stewart PM: Hexose-6-phosphate dehydro-genase knock-out mice lack 11 beta-hydroxysteroid dehydrogenasetype 1-mediated glucocorticoid generation. J Biol Chem 281: 6546–6551, 2006
13. White PC, Mune T, Agarwal AK: 11 Beta-hydroxysteroid dehydroge-nase and the syndrome of apparent mineralocorticoid excess. EndocrRev 18: 135–156, 1997
14. Tomlinson JW, Walker EA, Bujalska IJ, Draper N, Lavery GG, CooperMS, Hewison M, Stewart PM: 11Beta-hydroxysteroid dehydrogenasetype 1: A tissue-specific regulator of glucocorticoid response. EndocrRev 25: 831–866, 2004
15. Homma M, Onodera T, Hirabatake M, Oka K, Kanazawa M, Miwa T,Hayashi T: Activation of 11 beta-hydroxysteroid dehydrogenase bydehydroepiandrosterone sulphate as an anti-hypertensive agent inspontaneously hypertensive rats. J Pharm Pharmacol 50: 1139–1145,1998
16. Arnold P, Tam S, Yan L, Baker ME, Frey FJ, Odermatt A: Glutamate-115 renders specificity of human 11beta-hydroxysteroid dehydroge-nase type 2 for the cofactor NAD(�). Mol Cell Endocrinol 201: 177–187, 2003
17. Apostolova G, Schweizer RA, Balazs Z, Kostadinova RM, Odermatt A:Dehydroepiandrosterone inhibits the amplification of glucocorticoidaction in adipose tissue. Am J Physiol Endocrinol Metab 288: E957–E964, 2005
18. Pei DQ, Shih CH: Transcriptional activation and repression by cellularDNA-binding protein C/EBP. J Virol 64: 1517–1522, 1990
19. Williams LJ, Lyons V, MacLeod I, Rajan V, Darlington GJ, Poli V, SecklJR, Chapman KE: C/EBP regulates hepatic transcription of 11beta-hydroxysteroid dehydrogenase type 1: A novel mechanism for cross-talk between the C/EBP and glucocorticoid signaling pathways. J BiolChem 275: 30232–30239, 2000
20. Gout J, Tirard J, Thevenon C, Riou JP, Begeot M, Naville D: CCAAT/enhancer-binding proteins (C/EBPs) regulate the basal and cAMP-induced transcription of the human 11beta-hydroxysteroid dehydro-genase encoding gene in adipose cells. Biochimie 88: 1115–1124,2006
21. Djelidi S, Beggah A, Courtois-Coutry N, Fay M, Cluzeaud F, Vieng-chareun S, Bonvalet JP, Farman N, Blot-Chabaud M: Basolateral trans-location by vasopressin of the aldosterone-induced pool of latentNa-K-ATPases is accompanied by alpha1 subunit dephosphorylation:Study in a new aldosterone-sensitive rat cortical collecting duct cellline. J Am Soc Nephrol 12: 1805–1818, 2001
22. Liu D, Dillon JS: Dehydroepiandrosterone activates endothelial cellnitric-oxide synthase by a specific plasma membrane receptor cou-pled to Galpha(i2,3). J Biol Chem 277: 21379–21388, 2002
23. Liu D, Si H, Reynolds KA, Zhen W, Jia Z, Dillon JS: Dehydroepiandro-sterone protects vascular endothelial cells against apoptosis through aGalphai protein-dependent activation of phosphatidylinositol 3-ki-nase/Akt and regulation of antiapoptotic Bcl-2 expression. Endocri-nology 148: 3068–3076, 2007
24. Buck M, Poli V, van der Geer P, Chojkier M, Hunter T: Phosphorylationof rat serine 105 or mouse threonine 217 in C/EBP beta is required forhepatocyte proliferation induced by TGF alpha. Mol Cell 4: 1087–1092, 1999
25. Muller C, Cluzeaud F, Pinon GM, Rafestin-Oblin ME, Morfin R: Dehy-droepiandrosterone and its 7-hydroxylated metabolites do not inter-fere with the transactivation and cellular trafficking of the glucocorti-
coid receptor. J Steroid Biochem Mol Biol 92: 469–476, 200426. Funder JW, Pearce PT, Smith R, Smith AI: Mineralocorticoid action:
Target tissue specificity is enzyme, not receptor, mediated. Science242: 583–585, 1988
27. Atanasov AG, Ignatova ID, Nashev LG, Dick B, Ferrari P, Frey FJ andOdermatt A: Impaired protein stability of 11beta-hydroxysteroid de-hydrogenase type 2: A novel mechanism of apparent mineralocorti-coid excess. J Am Soc Nephrol 18: 1262–1270, 2007
28. Ackermann D, Vogt B, Escher G, Dick B, Reichen J, Frey BM, Frey FJ:Inhibition of 11beta-hydroxysteroid dehydrogenase by bile acids inrats with cirrhosis. Hepatology 30: 623–629, 1999
29. Vogt B, Dick B, N�Gankam V, Frey FJ, Frey BM: Reduced 11beta-hydroxysteroid dehydrogenase activity in patients with the nephroticsyndrome. J Clin Endocrinol Metab 84: 811–814, 1999
30. Quattropani C, Vogt B, Odermatt A, Dick B, Frey BM, Frey FJ: Re-duced activity of 11beta-hydroxysteroid dehydrogenase in patientswith cholestasis. J Clin Invest 108: 1299–1305, 2001
31. Stauffer AT, Rochat MK, Dick B, Frey FJ, Odermatt A: Chenodeoxy-cholic acid and deoxycholic acid inhibit 11 beta-hydroxysteroid dehy-drogenase type 2 and cause cortisol-induced transcriptional activationof the mineralocorticoid receptor. J Biol Chem 277: 26286–26292,2002
32. Thiesson HC, Jensen BL, Bistrup C, Ottosen PD, McNeilly AD, AndrewR, Seckl J, Skott O: Renal sodium retention in cirrhotic rats depends onglucocorticoid-mediated activation of mineralocorticoid receptor dueto decreased renal 11beta-HSD-2 activity. Am J Physiol Regul IntegrComp Physiol 292: R625–R636, 2007
33. Lovati E, Ferrari P, Dick B, Jostarndt K, Frey BM, Frey FJ, Schorr U,Sharma AM: Molecular basis of human salt sensitivity: The role of the11beta-hydroxysteroid dehydrogenase type 2. J Clin EndocrinolMetab 84: 3745–3749, 1999
34. Fagerudd JA, Tarnow L, Jacobsen P, Stenman S, Nielsen FS, Petters-son-Fernholm KJ, Gronhagen-Riska C, Parving HH, Groop PH: Predis-position to essential hypertension and development of diabetic ne-phropathy in IDDM patients. Diabetes 47: 439–444, 1998
35. Strojek K, Grzeszczak W, Morawin E, Adamski M, Lacka B, Rudzki H,Schmidt S, Keller C, Ritz E: Nephropathy of type II diabetes: Evidencefor hereditary factors? Kidney Int 51: 1602–1607, 1997
36. Lavery GG, McTernan CL, Bain SC, Chowdhury TA, Hewison M, Stew-art PM: Association studies between the HSD11B2 gene (encodinghuman 11beta-hydroxysteroid dehydrogenase type 2), type 1 diabe-tes mellitus and diabetic nephropathy. Eur J Endocrinol 146: 553–558,2002
37. Brereton PS, van Driel RR, Suhaimi F, Koyama K, Dilley R, Krozowski Z:Light and electron microscopy localization of the 11beta-hydroxys-teroid dehydrogenase type I enzyme in the rat. Endocrinology 142:1644–1651, 2001
38. Trautwein C, Caelles C, van der Geer P, Hunter T, Karin M, Chojkier M:Transactivation by NF-IL6/LAP is enhanced by phosphorylation of itsactivation domain. Nature 364: 544–547, 1993
39. Lee SJ, Kim SG: Role of p90 ribosomal S6-kinase-1 in oltipraz-inducedspecific phosphorylation of CCAAT/enhancer binding protein-beta forGSTA2 gene transactivation. Mol Pharmacol 69: 385–396, 2006
40. Escher G, Galli I, Vishwanath BS, Frey BM, Frey FJ: Tumor necrosisfactor alpha and interleukin 1beta enhance the cortisone/cortisol shut-tle. J Exp Med 186: 189–198, 1997
41. Cooper MS, Bujalska I, Rabbitt E, Walker EA, Bland R, Sheppard MC,Hewison M, Stewart PM: Modulation of 11beta-hydroxysteroid dehy-drogenase isozymes by proinflammatory cytokines in osteoblasts: anautocrine switch from glucocorticoid inactivation to activation. J BoneMiner Res 16: 1037–1044, 2001
42. Kostadinova RM, Nawrocki AR, Frey FJ, Frey BM: Tumor necrosisfactor alpha and phorbol 12-myristate-13-acetate down-regulatehuman 11beta-hydroxysteroid dehydrogenase type 2 through p50/p50 NF-kappaB homodimers and Egr-1. FASEB J 19: 650 – 652,2005
BASIC RESEARCH www.jasn.org
100 Journal of the American Society of Nephrology J Am Soc Nephrol 19: 92–101, 2008

43. Kimura M, Tanaka S, Yamada Y, Kiuchi Y, Yamakawa T, Sekihara H:Dehydroepiandrosterone decreases serum tumor necrosis factor-al-pha and restores insulin sensitivity: Independent effect from second-ary weight reduction in genetically obese Zucker fatty rats. Endocri-nology 139: 3249–3253, 1998
44. Miller M, Shuman JD, Sebastian T, Dauter Z, Johnson PF: Structuralbasis for DNA recognition by the basic region leucine zipper transcrip-tion factor CCAAT/enhancer-binding protein alpha. J Biol Chem 278:15178–15184, 2003
45. Audige A, Dick B, Frey BM, Frey FJ, Corman B, Vogt B: Gluco-corticoids and 11 beta-hydroxysteroid dehydrogenase type 2gene expression in the aging kidney. Eur J Clin Invest 32: 411– 420,2002
46. Livak KJ, Schmittgen TD: Analysis of relative gene expression data
using real-time quantitative PCR and the 2(-Delta Delta C(T)) method.Methods 25: 402–408, 2001
47. Schweizer RA, Zurcher M, Balazs Z, Dick B, Odermatt A: Rapid hepaticmetabolism of 7-ketocholesterol by 11beta-hydroxysteroid dehydro-genase type 1: Species-specific differences between the rat, human,and hamster enzyme. J Biol Chem 279: 18415–18424, 2004
48. Atanasov AG, Nashev LG, Tam S, Baker ME, Odermatt A: Organotinsdisrupt the 11beta-hydroxysteroid dehydrogenase type 2-dependentlocal inactivation of glucocorticoids. Environ Health Perspect 113:1600–1606, 2005
49. Schweizer RA, Atanasov AG, Frey BM, Odermatt A: A rapid screeningassay for inhibitors of 11beta-hydroxysteroid dehydrogenases (11be-ta-HSD): Flavanone selectively inhibits 11beta-HSD1 reductase activ-ity. Mol Cell Endocrinol 212: 41–49, 2003
BASIC RESEARCHwww.jasn.org
J Am Soc Nephrol 19: 92–101, 2008 DHEA Upregulates 11�-HSD2 101