a two-element enhancer-inhibitor transposon system in arabidopsis thaliana
TRANSCRIPT

Mol Gen Genet (1995) 247:555-564 Q Springer-Verlag 1995
Mark G . M . A a r t s • Paul Corzaan • W i l l e m J. Stiekema Andy Pereira
A two-element Enhancer-Inhibitor transposon system in Arabidopsis thaliana
Received: 1 August 1994/Accepted: 11 January 1995
A b s t r a c t The Enhancer-Inhibitor (En-I), also known as Suppressor-mutator (Spm-dSpm), transposable element system of maize was modified and introduced into Ara- bidopsis by Agrobacterium tumefaciens transformation. A stable En/Spm transposase source under control of the CaMV 35S promoter mediated frequent transposition of I/dSpm elements. Transposition occurred continuously throughout plant development over at least seven consec- utive plant generations after transformation. New inser- tions were found at both linked and unlinked positions relative to a transposon donor site. The independent transposition frequency was defined as a transposition parameter, which quantified the rate of unique insertion events and ranged from 7.8% to 29.2% in different popu- lations. An increase as well as a decrease in I/dSpm ele- ment copy number was seen at the individual plant level, but not at the population level after several plant genera- tions. The continuous, frequent transposition observed for this transposon system makes it an attractive tool for use in gene tagging in Arabidopsis.
Key words Transposable element • En/Spm • Arabidopsis thaliana • Continuous transposition • Gene tagging
Introduction
Due to the availability of well characterized endogenous transposons, transposon tagging is the system of choice for isolating genes correlated with specific mutant phe- notypes from Zea mays (maize) (Fedoroff et al. 1984) and Antirrhinum majus (snapdragon) (Martin et al. 1985). Maize (Ac-Ds, En/Spm) and snapdragon (Tam3)
Communicated by H. Saedler
M.G.M. Aarts • P. Corzaan. W. ]. Stiekema • A. Pereira (~) Department of Molecular Biology, DLO-Centre for Plant Breeding and Reproduction Research (CPRO-DLO), Postbus 16, 6700 AA Wageningen, The Netherlands
transposons have also been transferred to other plant species for heterologous transposon tagging (Haring et al. 1991). Successful tagging of the PH6 gene in Petunia hybrida with the maize Ac element was reported by Chuck et al. (1993), while Bancroft et al. (1993) tagged the DRL1 gene using an Ac-Ds system in Arabidopsis.
The autonomous Enhancer (En) or Suppressor-muta- tor (Spm) transposable element, originally identified in maize (Peterson 1953; McClintock 1954), has been molecularly characterized (Pereira et al. 1986; Masson et al. 1987) and also introduced into several heterologous hosts (Frey et al. 1989; Masson and Fedoroff 1989; Pereira and Saedler 1989; Cardon et al. 1993a). We re- cently reported the use of a maize En/Spm-derived sys- tem to tag and clone the MS2 gene from Arabidopsis (Aarts et al. 1993). Non-autonomous derivatives of En/Spm are termed Inhibitor (I) or defective Spin (dSpm) elements. The T-DNA construct we used to introduce an En/Spm-I/dSpm 'in cis two-element' system into plants, contained an unmarked I/dSpm element inserted in the open reading frame of an NPTII marker gene, to monitor excision phenotypically. The same construct contained an En/Spm element as transposase source, from which both the 5' and 3' ends had been removed, creating a stable En/Spm, and replaced by cauliflower mosaic virus (CaMV) 35S promoter (5') and terminator (3') se- quences. These elements control transcription of the two mRNAs that are needed to produce the TnpA and TnpD transposase proteins (Frey et al. 1990; Masson et al. 1991).
This paper gives a detailed description of the En/Spm- I/dSpm in cis two-element tagging system in Arabidop- sis. Four basic aspects of the transposition behaviour of this system have been studied: (1) the frequency of trans- position; (2) the possible limitation of activity of the transposon system after a number of generations; (3) the transpositional loss of I elements; and (4) the transposi- tion to linked sites. These aspects were examined by phe- notypic and molecular genetic means in seven consecu- tive generations. The advantages of this transposon sys- tem for efficient gene tagging in Arabidopsis are dis- cussed.

556
Materials and methods
Binary vector plasmid cwEnN::I
A chimeric neomycin phosphotransferase II (NPTII) gene was inserted into binary vector pGDW3.1 (Wing et al. 1989), contain- ing a chimeric nopaline synthase (nos) promoter-hygromycin phosphotransferase gene (HPT) for selection following transfor- mation. The NPTII gene was modified from Baker et al. (1987), by introduction of a ClaI linker to generate a new translation start (GCGATGG) 5' to a unique BamHI site present in the untranslated leader between the TRI' promoter and the NPTII coding region. Insertion of a 2.2 kb I element with flanking DNA (1-6078; Pereira and Saedler 1989) into the BamHI site and replacement of the original downstream NPTII sequences by sequences downstream of the BamHI site in plasmid pBCK1 (Kaulen et al. 1986) yielding pBHNI, altered the original N-terminus. Upon excision of 1-6078 (e.g. with a 2 bp deletion) the correct reading frame would be MDPACPKTLEVDPAGSG after which follows the normal NPTII reading frame from K 12 onwards. A 7.6 kb BssHII fragment from En-1, treated with Klenow enzyme (Pereira et al, 1986), was cloned between the CaMV 35S promoter-terminator cassette, originating from pD51 (Pietrzak et al. 1986), in pBR322. This wings-clipped En-1 (cwEn) under control of the 35S pro- moter was cloned into pBHNI to produce plasmid cwEnN::I (Fig. 1), which was mobilized into the Agrobacterium strain pGV3101(pMP90RK) Koncz and Schell 1986).
Plant material
For plant transformation, root explants of ecotype Landsberg erecta were infected with Agrobacterium containing cwEnN::I, according to Valvekens et al. (1988). Transformed calli were se- lected on medium containing 20 mg/1 hygromycin. Emerging T t shoots were grown without selection and allowed to self-fertilize and set seed in vitro. Growth conditions were 12-16 h light at 22°C in a growth chamber.
Seeds of T2 and subsequent generations were sterilized if nec- essary by treatment with ethanol 70% (2 rain), 50% commercial bleach (5% sodium hypochlorite, 5 min) and five subsequent washes with sterile water, all in Eppendorf tubes. For selection on antibiotics the seeds were sown on GM medium (Valvekens et al. 1988) or half-strength MS medium (Murashige and Skoog 1962) plus the required antibiotic (100 mg/1 kanamycin, 20 mg/1 hy- gromycin), solidified with 0.8% purified agar. Using no sugar in the medium permitted the sowing of unsterilized seeds on hy- gromycin-containing medium. Segregation for antibiotic resis- tance was scored 5-10 days after germination. Where appropriate, seedlings were transferred to soil (compost:vermiculite:sand 4:1:1) and grown in a climate-controlled greenhouse at 20-22°C with additional light (16 h). To prevent cross-fertilization and dis- persal of seeds, plants were grown in Aracon containers (Beta Developments, Gent, Belgium). For the phenotypic excision assay, seeds were only germinated on GM medium containing 100 mg/1 kanamycin sulphate. Resistance, variegation or sensitivity was scored 5-10 days after germination. Variegated and sensitive seedlings were transferred to GM without antibiotic, before trans- ferring to soil.
Ploidy number was determined by counting chloroplasts in stomatal guard cells of the lower epidermis of young leaves as described by Detrez et al. (1989). At least 25 stomata of different T 2 and T 3 plants were counted.
DNAanalysis
Genomic DNA was isolated according to Dellaporta et al. (1983) or Shure et al. (1983). PCR analysis to amplify empty donor sites of the cwEnN::I T-DNA was performed with 50-200 ng of genom- ic DNA. The primer sequences were 5' CTT-ACG-TCA-CGT-
CTT~GCG-CA 3' (1' promoter) and 5' CCA-GTC-ATA-GCC- GAA-TAG-CC 3' (NPTII gene). For PCR, 30 cycles of denatura- tion (94°C, 30 s), annealing (55-60°C, 1 rain) and extension (72°C, 2 rain) were carried out. PCR products of interest were cloned in Bluescript SK + as blunt-ended fragments after Klenow treatment and gel elution. Inverse PCR (IPCR) was performed on 200 ng HinfI-digested genomic DNA, and amplified fragments were Klenow treated and self-ligated. The self-ligated DNA molecules were linearized by digestion with SaII. Two rounds of 25 PCR cycles (annealing 55°C, extension 3 rain 72°C) were applied, using the nested primers 103-106 described by Masson et al. (1991). A 2-5 I~1 aliquot of the first round products was used for the second round of PCR. Cloning of IPCR fragments was as for PCR frag- ments. To determine the DNA sequence, PCR was performed on most cloned IPCR products using one primer (5' GAC-ACT-CCT- TAG-ATC-TTT-TCT-TGT-AGT-G 3') annealing to both left and right terminal inverted repeats, after which the PCR product was cloned as described previously. Double-stranded supercoiled plas- mid was used for determining the DNA sequence on an automated DNA sequencer (Applied Biosystems). One strand and, if needed, both strands were sequenced.
For DNA blot analysis, 200-400 ng DNA was used per lane, separated in a 0.8% agarose gel in TRIS-acetate running buffer and, after electrophoresis, alkali-denatured and transferred onto Gene Screen Plus or Hybond N + nylon membrane by vacuum blotting. The blots were prehybridized and hybridized following the procedure recommended by the membrane manufacturer. DNA fragments, 32p-labelled by random priming, were used as probes (Fig. 1). A 0.27 kb fragment containing the left border of En (up to a SalI restriction site) was used to detect I elements. The T-DNA right border probe was a 1.1 kb fragment containing the HPT gene. A fragment of 1.8 kb containing the complete NPTII gene plus the octopine synthase (ocs) terminator was used to de- tect excision fragments (see Fig. 1 for probe locations on the T- DNA). After hybridization the membranes were washed with 2×SSC, 1% SDS at 65°C for 2×30 min and autoradiographed (Fuji or Kodak X-ray films) at -80°C using intensifying screens.
Mapping T-DNA and I elements
For mapping of T-DNA and 1 elements, genomic DNA flanking the inserts was used as RFLP probes on a population of recombi- nant inbred lines (RILs) made by Lister and Dean (1993). We routinely used five restriction enzymes (BgIII, DraI, EcoRI, EcoRV and HindIII) to test for RFLPs between the ecotypes Landsberg erecta and Columbia. Probes giving a RFLP were hy- bridized to a blot containing the DNAs of 76 RILs digested with the appropriate enzyme. Recombination frequencies and map dis- tances were calculated with the JoinMap program designed by Stam (1993), using mapping data from RFLP probes provided by Clare Lister and Caroline Dean (Cambridge Laboratory, John In- nes Institute, Norwich, UK).
Results
The in cis two-e l emen t system
For convenience , we wil l use the t e rm En to indica te an e lement express ing t ransposase genes and I to refer to a n o n - a u t o n o m o u s e lement . The I e lement t r ansposon as well as the En t r ansposase genes were c loned on one T - D N A cons t ruc t c w E n N : : I (Fig. 1). In contras t to most pheno typ i c assay cons t ruc ts in which the t r ansposon is inse r t ed in the 5 ' un t rans la t ed leader of a marker gene (Baker et al. 1987), we c loned the I e lement di rect ly downs t r eam of the ATG start codon of a n e o m y c i n phos- pho t rans fe rase (NPTI l ) gene. This seemed necessa ry as I

LB s
cwEn 35s, / / 35s 3' A -m 1, ,,TG NPTIIo= y
:::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: 4 kb ::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: ~ . 1 1 I I
1"-i.:-:-~'~':'2"i.71.2-1-i-??:'2'?1-32.??~.?:'?r+2.32-72+?~.73
, 1 kb , B 1-6078 1
Hvr 4 , r. . . . I
Bg
3
557
Fig. 1 Schematic drawing of the T-DNA construct cwEnN::I, containing the 'in cis two-element En-I' transposon system. A stable wings-clipped En element expressing transposase genes tnpA and tnpD, is controlled by CaMV 35S promoter and termina- tor sequences. A 2.2 kb I element (I-6078) is cloned in the open reading frame of a NPTII gene, in opposite orientation relative to the En transposase. Excision of the element allows expression of the NPTlI kanamycin resistance gene. Hygromycin resistance, conferred by the HPT gene, is used to select for transformation. Promoter sequences are depicted as triangles, pointing in the di- rection of the transcription unit they control. 399 and 8041 are the positions of the BssHII sites in En-1 used for cloning the cwEn. Fragments used as probes for DNA blot hybridizations are indicat- ed with horizontal filled bars numbered 1-3. Probe 1 is a 0.27 kb fragment of the En-I left border (up to the SalI site). Probes 2 and 3 are the full coding plus terminator sequences of the NPTII and HPT genes, respectively. The positions of EcoRI, BglII and HindIII restriction sites are given below the construct as R1, Bg and H. ATG marks the translation start codon of the NPTII marker gene. LB and RB indicate the left and right borders, respectively, of the T-DNA construct
Table 1 Assaying T 2 progeny of primary transformants carrying the cwEnN::I T-DNA for excision. The percentages of kanamycin- resistant (Kr), variegated (K ~) and sensitive (K s) plants among the total number of plants (n) are given
Plant number K~(%) K v (%) K~(%) n
1A 13 17 70 76 1B a 0 32 68 19 1C 2 17 81 48 1D 21 14 65 57
a Tetraploid
Table 2 Assay of T 3 progeny of variegated T 2 plants carrying the cwEnN::I T-DNA for excision. The percentages of kanamycin-re- sistant (Kr), variegated (/~) and sensitive (K') plants among the total number of plants (n) are given
T 2 plant number K r (%) K" (%) K ~ (%) n
1A-H13 element insertion in the untranslated leader of the N P T I I 1A-K20 gene did not completely block expression of kanamycin 1A-9 resistance (Pereira and Saedler 1989; Cardon et al. 1A-54 1993a). The reading frame of the N P T I I gene is restored 1B-6
1B-13 to provide kanamycin resistance when I element excision events add 1 + 3 n bp or delete 2 + 3n bp. Excision of the I 1 c-1 1c-3 element is mediated by transposase proteins expressed lC-8 from a stable En element under control of the CaMV 35S 1D-8 promoter. 1 D- 10
1D-33 1D-51
Phenotypic analysis of I element excision
0 3 97 412 54 7 39 139 38 15 47 143 0 8 92 75 2 3 95 113 0 5 95 104
68 4 28 195 1 7 92 156 0.5 0.5 99 197 0 4 96 165 0.6 3 96.4 179 0.6 6 93.4 174 0.6 6 93.4 180
T-DNA cwEnN::I was introduced into Arabidops is by Agrobac te r ium-med ia ted transformation (Valvekens et al. 1988) and progeny of four transgenic shoots from one transformed callus (1A-1D) that displayed active trans- position, was further analysed. T2 seeds obtained from these T1 plants grown in vitro were germinated and seg- regation of T 2 plants for hygromycin resistance showed that the original callus was hemizygous for two unlinked T-DNA loci (data not shown). DNA blot analysis re- vealed that at these En transposase loci, respectively, five and two right border sequences of T-DNA were integrat- ed; they will be further referred to as the TEn5 and TEn2 loci, respectively. Another sample of the T2 seedlings was screened for I excision on kanamycin-containing medium (Table 1). The ratios of kanamycin resis- tant:variegated:sensitive plants varied substantially be- tween the four lines. Variegation for kanamycin resis- tance, visible as a few green spots on white cotyledons
and caused by excision events in the T2 plant itself, showed that the excision rates in progenies 1A, 1C and 1D were similar. Only progeny from tetraploid T~ plant 1B contained significantly more variegated T 2 plants, probably due to the higher T-DNA (and thus NPTII::I) copy number per cell.
Molecular analysis of I element excision
The frequency of variegated T3 plants (descending from variegated T2 plants) was in general much lower than in the previous generation (Table 2). One reason for this reduction could be that only some of the excision events resulted in a functional N P T l I gene and that, after two generations, there is a substantial reduction in the num- ber of T-DNA inserts still containing an I element in the
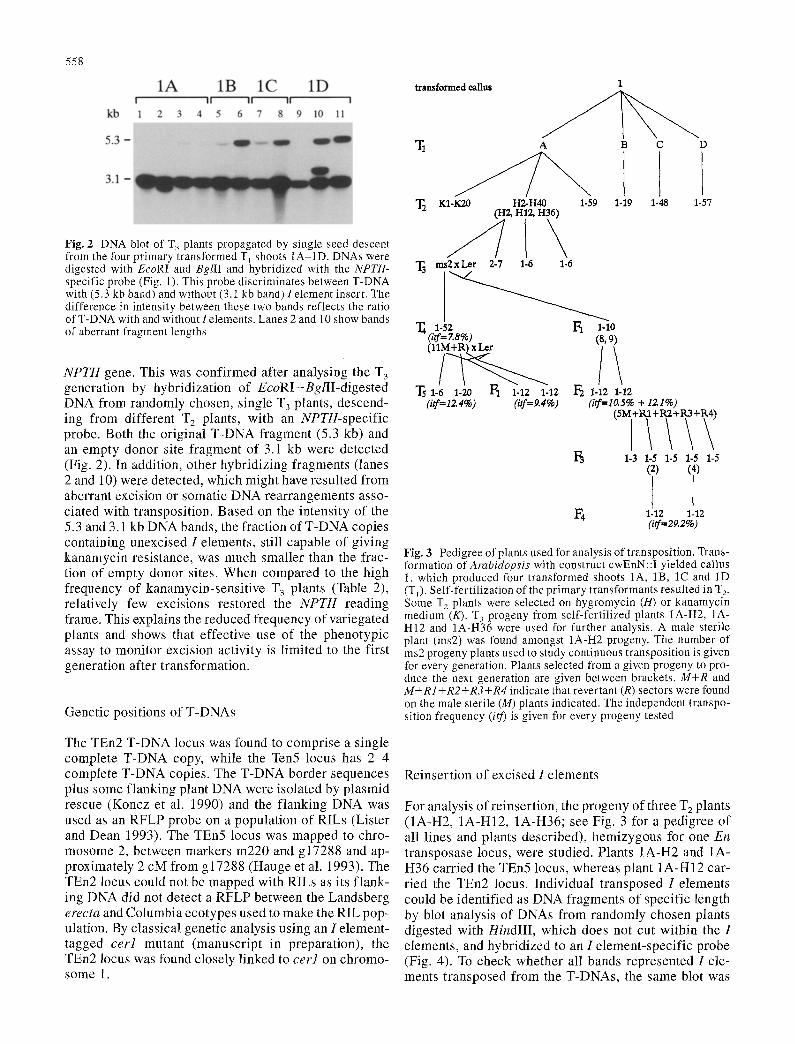
558
kb
5.3
1A 1B 1C 1D l l ! I I I I l
1 2 3 4 5 6 7 8 9 10 l l
3.1
Fig. 2 DNA blot of T 3 plants propagated by single seed descent from the four primary transformed T l shoots 1A-1D. DNAs were digested with EcoRI and BglII and hybridized with the NPTII- specific probe (Fig. 1). This probe discriminates between T-DNA with (5.3 kb band) and without (3.1 kb band) I element insert, The difference in intensity between these two bands reflects the ratio of T-DNA with and without I elements. Lanes 2 and 10 show bands of aberrant fragment lengths
NPTII gene. This was confirmed after analysing the T 3 generation by hybridization of EcoRI+BglII-digested DNA from randomly chosen, single T 3 plants, descend- ing from different T2 plants, with an NPTII-specific probe. Both the original T-DNA fragment (5.3 kb) and an empty donor site fragment of 3.1 kb were detected (Fig. 2). In addition, other hybridizing fragments (lanes 2 and 10) were detected, which might have resulted from aberrant excision or somatic DNA rearrangements asso- ciated with transposition. Based on the intensity of the 5.3 and 3.1 kb DNA bands, the fraction of T-DNA copies containing unexcised I elements, still capable of giving kanamycin resistance, was much smaller than the frac- tion of empty donor sites. When compared to the high frequency of kanamycin-sensitive T 3 plants (Table 2), relatively few excisions restored the NPTII reading frame. This explains the reduced frequency of variegated plants and shows that effective use of the phenotypic assay to monitor excision activity is limited to the first generation after transformation.
Genetic positions of T-DNAs
The TEn2 T-DNA locus was found to comprise a single complete T-DNA copy, while the Ten5 locus has 2 -4 complete T-DNA copies. The T-DNA border sequences plus some flanking plant DNA were isolated by plasmid rescue (Koncz et al. 1990) and the flanking DNA was used as an RFLP probe on a population of RILs (Lister and Dean 1993). The TEn5 locus was mapped to chro- mosome 2, between markers m220 and g17288 and ap- proximately 2 cM from g17288 (Hauge et al. 1993). The TEn2 locus could not be mapped with RILs as its flank- ing DNA did not detect a RFLP between the Landsberg erecta and Columbia ecotypes used to make the RIL pop- ulation. By classical genetic analysis using an 1 element- tagged cerl mutant (manuscript in preparation), the TEn2 locus was found closely linked to cerl on chromo- some 1.
transformed callus
T1
T2
T3
']~ 1-52 (i¢= zs~ )
(i¢=1z4~) 6¢=9.4%)
1
A B C D
K1-K20 H2-H40 1-59 1-19 1-48 1-57 (H2, H12, H36)
ms2 x Ler 2-7 1-6 1-6 --<.____
FI 1-10 (s, 9)
// F2 1-12 1-12
(i¢=1o.~ + lz1%)
F3
F4
1-3 1-5 1-5 1-5 1-5 (2) (4)
I I 1-12 1-12 (i¢=29.2~)
Fig. 3 Pedigree of plants used for analysis of transposition. Trans- formation of Arabidopsis with construct cwEnN::I yielded callus 1, which produced four transformed shoots 1A, 1B, 1C and 1D (T~). Self-fertilization of the primary transformants resulted in T2. Some T a plants were selected on hygromycin (H) or kanamycin medium (K). r 3 progeny from self-fertilized plants 1A-H2, 1A- H12 and 1A-H36 were used for further analysis. A male sterile plant (ms2) was found amongst 1A-H2 progeny. The number of ms2 progeny plants used to study continuous transposition is given for every generation. Plants selected from a given progeny to pro- duce the next generation are given between brackets. M+R and M+RI+R2+R3+R4 indicate that revertant (R) sectors were found on the male sterile (/14) plants indicated. The independent transpo- sition frequency (it¢) is given for every progeny tested
Reinsertion of excised I elements
For analysis of reinsertion, the progeny of three T 2 plants (1A-H2, 1A-H12, 1A-H36; see Fig. 3 for a pedigree of all lines and plants described), hemizygous for one En transposase locus, were studied. Plants 1A-H2 and 1A- H36 carried the TEn5 locus, whereas plant 1A-H12 car- ried the TEn2 locus. Individual transposed I elements could be identified as DNA fragments of specific length by blot analysis of DNAs from randomly chosen plants digested with HindIII, which does not cut within the I elements, and hybridized to an I element-specific probe (Fig. 4). To check whether all bands represented 1 ele- ments transposed from the T-DNAs, the same blot was

H2 H12 H36 I I I I
kb 1 2 3 4 5 6 7 8 9 10 1112n13 14 15 16 17 18 T T T T T T T T T T T T
14.1 -
4 . 8 --
2 .3 -
Fig. 4 DNA blot of T 3 plants from three different 1A T 2 plants demonstrat ing reinsert ions of excised I elements. DNAs were di- gested with both EcoRI and BglII. The blot was hybridized with the I element-specific probe (see Fig. 1). The three progenies shown descended from plants 1A-H2 (lanes 1-6), 1A-HI2 (lanes 7 -12) and 1A-H36 (lanes 13-18). The presence o fEn t ransposase genes is depicted by T above the corresponding lane
hybridized with the HPT probe (see Fig. 1) to discrimi- nate between T-DNAs with and without an I element. Only progeny of 1A-H12 contained an I element inserted in the T-DNA (data not shown). Most plants with trans- posase genes (indicated by T in Fig. 4) displayed a back- ground smear of hybridization, in addition to distinct bands, due to somatic transposition. From the observa- tion of a number of plants which lacked T-DNA but con- tained transposed I elements, it was concluded that most of the I elements had transposed to sites genetically un- linked to the T-DNA. A high frequency of transposition was suggested by the large number of DNA fragments bearing I elements and by the variation in DNA fragment number and pattern between the three T2 lines and the T3 plants from the same T2 line (Fig. 4).
Quantification of transposition
As a measure of transposition in each generation, the independent transposition frequency (i@ was deter- mined. The itf is defined as the number of unique I ele- ment inserts relative to the total number of I element-ho- mologous DNA fragments that appear on a DNA blot made from a population of plants containing En trans- posase, all descending from one plant or cross. A mini- mal population size of eight progeny plants has been chosen, as there is only a small chance that after Mende- lian segregation a plant will be found in a selfed progeny that has a unique I element insert (e.g. the chance of one plant in a progeny of eight having a unique element in- sert, when the parent was hemizygous for this insert, is 3.6?< 10-4). Only bands that had a higher intensity than about half the intensity of the hemizygous bands were counted HindlII digests were used, which best revealed discriminatory bands.
The i t f in progeny of the male sterile plant carrying a mutation caused by an I element insertion in the MS2 gene ofArabidopsis (Aarts et al. 1993) was first deter- mined. This mutant appeared in the T3 generation
559
derived from plant 1A-H2. As some self-fertilization of ms2 mutants takes place at the end of flowering, several generations were obtained after selling in the course of examining the nature of the mutation. In 18 T 4 progeny of the T 3 male sterile mutant, hemizygous for TEn5, DNA blot analysis revealed 10 unique inserts in a total of 127 inserts. The itf for this population was 7.8% (10/ 127×100%). Among the T 4 progeny was a variegated male sterile plant with a large sector of fertile flowers (Aarts et al. 1993). Both male sterile and fertile flowers were allowed to self-pollinate as well as being out- crossed with Landsberg erecta. A blot containing DNA from all four progenies was hybridized with the I ele- ment-specific probe (Fig. 5). When the progenies derived from outcross and self-pollination were consid- ered separately, similar itfvalues were found (9.4%, 10 in 106; and 12.4%, 18 in 145, respectively), which were not significantly different from the T 4 generation itf.
The large number of elements in the T 4 and T 5 progeny increased the probability that elements giving approximately the same restriction fragment lengths could not be distinguished. In order to obtain plants with fewer and preferably only one I element, the original male sterile T 3 mutant was crossed to Landsberg erecta (Fig. 3) and subsequent progenies were analysed for the transposition rate. The F2s derived from two F 1 plants hemizygous for En transposase showed similar itfvalues (10.5%, 2 in 19; and 12.1%, 4 in 3, respectively) to those found for the T 4 and Ts progenies.
One male sterile F2 plant (ms2::I/ms2::I) with only two inserts and hemizygous for En transposase was cho- sen and the F 3 progeny from fertile flowers (resulting after excision of the I element from the ms2::I locus) were screened for plants with only one I element. Two F3 plants (Fig. 6A) were chosen to give the F4 generation (Fig. 6B), which allowed the quantification of the activi- ty of En transposase on one single I element insert. The i t f for this generation was 29.2% (7 in 24), significantly higher than for previous generations.
The activity of the TEn2 T-DNA locus was not exten- sively studied, but in progenies of plant 1A-H12, this transposase source was found to be a factor of 3-5 less capable of activating I elements than the TEn5 T-DNA locus.
Timing of excision
Early transposition in the parental plant at or before bolt- ing stage, generally gives a large sector of the plant con- taining the insert, which will be inherited by a large number of progeny plants if present in the germline. Lat- er transpositions will give correspondingly smaller sec- tors and thus fewer progeny containing the insert, while very late transpositions in single flowers or even gametes will give progeny containing unique inserts. The F 3
progeny shown in Fig. 6A demonstrate the timing of ex- cision when only two elements are involved. Most of the fertile revertant sectors found on the parental Fz plant
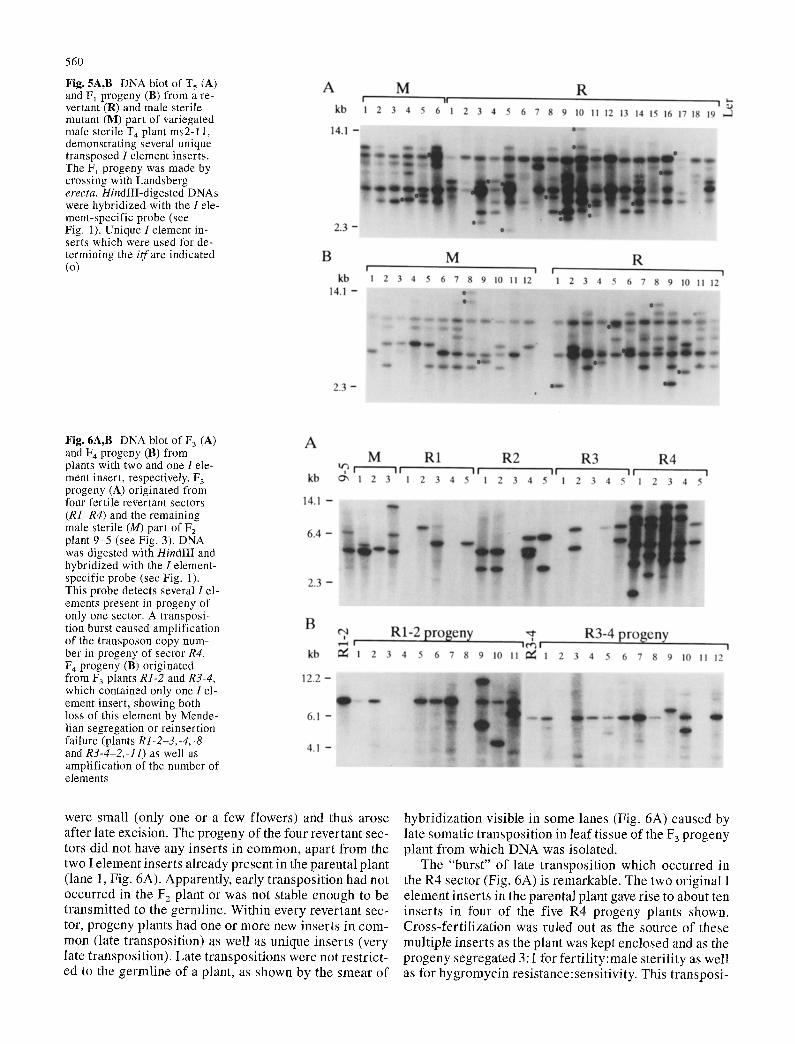
560
Fig. 5A,B DNA blot of T 5 (A) and Fj progeny (B) from a re- vertant (R) and male sterile mutant (M) part of variegated male sterile T 4 plant ms2-11, demonstrating several unique transposed 1 element inserts. The FI progeny was made by crossing with Landsberg erecta. HindlII-digested DNAs were hybridized with the I ele- ment-specific probe (see Fig. 1). Unique 1element in- serts which were used for de- termining the itf are indicated (o)
A M '1 II
kb 2 3 4 5 6 1
14.1 -
2 3 4
R 5 6 7 8 9 10 I1 12 13 14 15 16 17 18 19 ~.d
2 .3 -
B M I I
kb 1 2 3 4 5 6 7 8 9 10 11 12 14.1 -
R ! I
1 2 3 4 5 6 7 8 9 10 11 12
2 . 3 -
Fig. 6A,B DNA blot of F 3 (A) and 174 progeny (B) from plants with two and one I ele- ment insert, respectively. F 3 progeny (A) originated from four fertile revertant sectors (R1-R4) and the remaining male sterile (M) part of F2 plant 9-5 (see Fig. 3). DNA was digested with HindlII and hybridized with the I element- specific probe (see Fig. 1). This probe detects several I el- ements present in progeny of only one sector. A transposi- tion burst caused amplification of the transposon copy num- ber in progeny of sector R4. F 4 progeny (B) originated from F 3 plants R1-2 and R3-4, which contained only one I el- ement insert, showing both loss of this element by Mende- lian segregation or reinsertion failure (plants R1-2-3,-4,-8 and R3-4-2,-11) as well as amplification of the number of elements
A M R1 R2
LI~ I II kb G'~ 1 2 3 1 2 3
14.1 -
6 . 4 -
R 3 R 4 I ! I I I I I
4 5 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
2 .3 - -
B
kb
12.2 -
6 . 1 -
4 . 1 -
~, R1-2 progeny
~1~ 1 2 3 4 5 6 7 8
R 3 - 4 p r o g e n y It'~5 1 I
9 t0 11 ~ 1 2 3 4 5 6 7 8 9 10 II 12
were smal l (on ly one or a few f lowers ) and thus a rose af ter la te excis ion. The p r o g e n y o f the four r ever tan t sec- tors d id not have any in se r t s in c o m m o n , a p a r t f rom the two I e l emen t inse r t s a l r e a d y p re sen t in the pa ren ta l p lan t ( lane 1, Fig. 6A). Apparen t ly , ear ly t r an spos i t i on had not occu r r ed in the F2 p lan t or was not s table enough to be t r a n s m i t t e d to the ge rml ine . Wi th in eve ry r eve r t an t sec- tor, p r o g e n y p lants had one or m o r e new inse r t s in c o m - m o n ( la te t r anspos i t i on ) as wel l as un ique inse r t s (very late t r anspos i t ion) . La te t r anspos i t i ons were not r e s t r i c t - ed to the g e r m l i n e o f a plant , as shown by the s m e a r o f
h y b r i d i z a t i o n v is ib le in some lanes (Fig. 6A) caused by la te soma t i c t r a n s p o s i t i o n in l ea f t i ssue o f the F 3 p r o g e n y p lan t f rom w h i c h D N A was i so la ted .
The "burs t " o f la te t r a n s p o s i t i o n wh ich o c c u r r e d in the R4 sec to r (Fig. 6A) is r emarkab le . The two o r ig ina l I e l emen t i n se r t s in the pa ren ta l p lan t gave r i se to about ten inse r t s in four o f the f ive R4 p r o g e n y p lan ts shown. C r o s s - f e r t i l i z a t i o n was ru led out as the source o f these mu l t i p l e inse r t s as the p lan t was kep t enc losed and as the p r o g e n y s e g r e g a t e d 3:1 for f e r t i l i t y : m a l e s t e r i l i t y as wel l as for h y g r o m y c i n r e s i s t ance : sens i t i v i ty . This t r an spos i -

Fig. 7 Genetic map of Ara- bidopsis showing the positions of transposed I elements (I/dSpm) and flanking mark- ers. I elements marked with an asterisk were not visible as an RFLP fragment after DNA blot analysis and are thought not to be transmitted to progeny. TEn2 and TEn5 are transposase T-DNA loci. Markers between brackets were not mapped by RFLP mapping. TEn2 was mapped with classical markers and 1/dSpm27 was mapped on the physical map after YAC hy- bridization (R. Schmidt, J. West and C. Dean, John Innes Centre, Norwich, UK, unpub- lished results)
85
BB
95 97
104
1
(TEn2)
I/dSpm54* m488
g4552 I/dSpm60*
I/dSpm6= m532
g17311
15 19 21
36 $1 ¢3 48 4,9 52
2
m246
m216 I/dSpm53 rn251
m220 TEn5 917288 94,514
I/dSpm52 336
0 m583 1 g¢523
4 ms2::l/dSpm 8 m228
43 m249 45 I/dSpm18
47 g4564T
57 g2778 61 I/dSpm51* 63 m¢2~ 66
70
78
4
(1,/dSprn27) l/dSpm41G
g5843
pCITd99
I/dSpm6¢
g3713
561
5
0 . 93715
49, m247 52, I/dSpm3
54 ' g4028
91 m555
tion burst most probably occurred in only a part of the revertant sector, as one of the progeny plants contained only the two parental I element inserts. Similar bursts of transposition were observed in other progenies (data not shown), indicating that such events are not uncom- mon.
DNA analysis of excision and insertion sites
To examine whether the activity of this En-I system could be attributed to a change in the transposition mechanism (reflected in altered excision and insertion footprints), some excision and insertion sites were analysed. Two empty T-DNA donor sites from one plant were cloned by PCR and sequenced. In one footprint five nucleotides were deleted, thus restoring the reading frame of the NPTI1 gene. This was in accordance with the kanamycin- resistant phenotype of the plant from which the excision site was cloned. The other empty donor site was the re- sult of an excision that left the 3 bp target site duplication behind but altered 1 bp.
To characterize reinsertion sites, DNA flanking 17 I elements was cloned and sequenced. The insertion of 12 I elements generated a 3 bp target site duplication char- acteristic of the En/Spm transposon family (data not shown, but sequences available on request). In four cases the target site duplication had a 1 bp mismatch, while one insertion revealed no target site duplication. The devia- tions at the target sites are significant, as other regions of the I elements sequenced in the same reactions showed only random deviations of about 0.5%. None of the se- quenced DNAs flanking I elements showed any homolo- gy to sequences present in the databases.
Genetic mapping of I elements
Most I element inserts from which flanking DNA was isolated detected single-copy loci when used as RFLP probes on Landsberg erecta or Columbia DNA. In about half the cases they did not detect an RFLP and could not be positioned on a genetic map. Seven I elements (out of 12 cloned I elements giving major bands in Fig. 6A) were mapped using the RIL population generated by Lister and Dean (1993) (Fig. 7), as well as four additional chimeric somatic insertions (out of seven cloned) not visible by DNA blot analysis. I elements revealing major bands on DNA blots were mostly found to be transmitted to the next generation, whereas somatic insertions were not. Of the two original I elements giving the mapped inserts, only the I element inserted at the ms2 locus could be mapped. Physical or genetic linkage was found for I/dSpm18 and I/dSpmS1, both from the R1 sector, and for I/dSpm41G and I/dSpm27 (mapped after YAC li- brary hybridization; unpublished results f rom R. Schmidt, J. West and C. Dean, John Innes Centre, Nor- wich, UK), both from the same R2 progeny plant. I/dSpm6 and I/dSpm60, though clustered, came from different sectors, M and R4. Two examples of tight clus- tering of I elements were found. In a full sibling of the original ms2 mutant, the I element had excised from the ms2: :I allele and a new insert was found which was insep- arable from the male sterile phenotype in a segregating population of 44 plants (data not shown). Transposition to a very close site was found in one progeny plant of the tagged ms2 mutant (Aarts et al. 1993), where a new in- sertion within the 4.4 kb HindlII fragment containing the MS2 gene was detected by DNA blot analysis (data not shown).

562
Discussion
The isolation of an I element-tagged male sterile mutant in progeny of a particular A. thaliana transformant demonstrated that the En-I in cis two-element system can be used for heterologous gene tagging (Aarts et al. 1993). This system consists of two En transposase lines (TEn2 and TEn5) and many lines with several transposed I elements either with or without En transposase. In this paper various aspects of the transposition behaviour of this system have been described, which will be beneficial for those who are planning to use this system for random or targeted gene isolation.
The system differs in a few characteristics from other transposon systems used in Arabidopsis:
1. The En transposase genes and the/element are located on one T-DNA. Thus only one transformant is needed and new positions of ! elements are generated by self- pollination. The presence of En transposase can be se- lected for by germinating seeds on hygromycin-contain- ing medium. 2. The I element is cloned in the open reading frame instead of in the untranslated leader of an NPTII excision marker gene to abolish background activity. Excision from this position leads to kanamycin resistance in about one-third of cases, which is manifest in the phenotypic assay only in the T2 generation, due to the high frequency of excision. 3. Both En transposase genes tnpA and tnpD are ex- pressed from one stable En element under control of the strong CaMV 35S promoter.
The transposition behaviour of this system in Arabidop- sis showed that I elements can be activated to transpose in the presence of the En transposase source. Correct expression of tnpA and tnpD from wild type maize En-1 in Arabidopsis has already been shown by Cardon et al. (1993a). Apparently Arabidopsis is able to perform the complex differential splicing which is needed to obtain both the tnpA and the tnpD mRNA in the correct relative amounts, even when the endogenous En promoter is re- placed by the CaMV 35S promoter. A non-autonomous I element can be activated in the presence of 35S-tnpA and 35S-tnpD (Cardon et al. 1993b), and the stable En de- scribed here adds to the list of En transposase sources that can be used to activate I elements.
One of the aspects of transposition we studied was the transposition frequency. The itf parameter described here provides another means of examining the potential use of a transposon system for gene tagging as an alter- native to determining a germinal excision frequency with a phenotypic assay (Schmidt and Willmitzer 1989). The itfwas found to be roughly around 10% for populations which carried several I elements, but could increase to 29.2% for progenies of plants containing only one I ele- ment.
The two En transposase loci, TEn2 and TEn5, differ in the frequency of/element transposition (in progeny as
well as somatically), with the TEn5 locus being the more active. Autoregulation of transcription by binding of the TnpA protein to repetitive motifs at the 5' end of the element as described by Fedoroff (1989) and Raina et al. (1993) is abolished, as most of the 5' end of the En trans- posase element has been removed. The difference in transposition rate between TEn2 and TEn5 might there- fore be caused by a positive gene dosage effect, but a position effect of genomic sequences flanking the T- DNAs on the level of expression cannot be ruled out.
The activity of the transposase source was studied over many generations and appeared not to be limited to a few generations. At the plant level, late and indepen- dent transposition events were found. The timing of transposition affects the number of independent inser- tions that are found in the following generation. Our data show that transpositions occur throughout plant develop- ment and as late as in single gametes, which is in accor- dance with the activity profile of the CaMV 35S pro- moter. In tobacco this promoter is active throughout plant development, starting several days after pollination (Benfey and Chua 1990; Scofield et al. 1993). The con- tinuous activity of this En-I system differs from that of 35S transposase based Ac-Ds transposon systems in Ara- bidopsis or tobacco. Scofield et al. (1993) recently pro- posed that Ac transposase expression in tobacco eventu- ally reaches a level that actually inhibits excision; this phenomenon leads to predominantly early transposi- tions, which might explain the few unique Ds element inserts found in progeny of Arabidopsis plants contain- ing a 35S Ac-Ds system (Long et al. 1993). Inhibition of late transposition does not occur in this En-I system, and as the autoregulation system is no longer functional, tim- ing of transposition follows the timing of activity of the 35S promoter.
The strong amplification of the number of transpos- able elements in some progeny plants ("transposition bursts", Fig. 6A), is a phenomenon also known from Ac in maize and in tomato (Osborne et al. 1991), where it results from transposition of an element from a replicat- ed to an unreplicated part of a DNA molecule during mitosis (Greenblatt and Brink 1961). En exhibits this transposition-related phenomenon in maize less fre- quently than Ac. Dash and Peterson (1994) showed that En preferably transposes from replicated to replicated DNA. In Arabidopsis, despite the occasional occurrence of transposition bursts, the number of 1 elements re- mained relatively stable over at least three subsequent generations without selection (T3-Ts), suggesting maintenance of the number of transposons, as seen in maize.
To characterize the transpositional mechanism, we analysed excision and insertion sites of I elements. All the excision footprints obtained after excision of the I element from the NPTII::I T-DNA, as well as from the ms2::l locus (Aarts et al. 1993; unpublished results) showed that excisions occurred according to the transpo- sition model proposed by Saedler and Nevers (1985). Of 17 sequenced DNAs flanking transposed I elements, four

had 1 bp mismatches when compar ing the 3 bp target site duplications directly f lanking the insert, while one in- sert was not f lanked by a 3 bp duplication. Such alter- ations were not observed among 12 En or I element f lanking sequences associated with transposit ion events in maize, tobacco and potato (Gierl et al. 1985; Menssen et al. 1990; Pereira and Saedler 1989; Frey et al. 1989, 1990; Cardon et al. 1991), but analysis of a larger num- ber of insertion sequences f rom both Arabidopsis and maize is needed to conclude that there are differences in the En-I t ransposit ion mechanism between these species.
The availability of a population of RILs (Lister and Dean 1993) greatly facili tated the positioning of I ele- ments on a genetic map using t ransposon-f lanking DNA as RFLP probes. A relative lack of RFLPs, however, al- lowed only a small number of I elements to be mapped and, although both clustered and unlinked transpositions were found, there was no indication of a strong prefer- ence for transposit ion to nearby loci. The observation that ms2: :I and some other I elements seemed to be locat- ed in distal parts of the chromosomes , suggests a tenden- cy of I elements to t ranspose to new chromosomal posi- tions of similar constitution (i.e. nuclear architecture or t ime of replication in the cell cycle). I element transposi- tion in Arabidopsis resembles En t ransposit ion in maize, where it was found to transpose to linked sites in about 25% of the excision events (Peterson 1970). For tobacco, linked transposit ion was found after 44% of dSpm trans- positions (Cardon et al. 1993b). Transposition to linked and unlinked sites has also been shown for Ac and Ds in tomato (Osborne et al. 1991; Belzile and Yoder 1992; Healy et al. 1993; Knapp et al. 1994). The random and linked character of I element transposit ion has a practi- cal consequence for gene tagging. By out-crossing to wild type and subsequent self-ferti l ization, the resulting F2 can contain plants with one or a few stable I element inserts. By segregating the mutation away from the trans- posase source, unstable mutants can be stabilized, and since the number of I elements will be reduced, DNA flanking the mutating I element insert can be more read- ily isolated.
One of the major aims of developing a heterologous t ransposon system for Arabidopsis is to apply t ransposon mutagenesis. "Targeted tagging" can be facili tated by using male sterile I element/En t ransposase plants that are crossed (on a large scale) with the target mutant. By determining the i tf i t is possible to estimate the min imum size of a population that has to be screened to find one plant having a t ransposon insertion in a target gene (as- suming a similar mutation frequency for every gene), Based on the assumption that t ransposon-carrying plants are used with approximately ten randomly distributed I elements and an i t fof 15%, the number of F1 plants to be screened will be around 105 , comparable with that for T -DNA mutagenesis (Feldmann 1991). This figure ac- counts for tagging a gene when no I elements are in close l inkage (within 5 cM). To target a gene of unknown loca- tion, a strategy using En plants that have 10-20 randomly distributed I elements for crosses to the target mutant,
563
will give good chances for success. From experiments with En in maize it is known that the mutation frequency can increase 100-fold when the t ransposon is tightly linked to the target locus (Cone 1994). In Arabidopsis it can be expected that the mutation frequency will similar- ly increase when an I element is genetically linked to the target locus. I f the target gene is mapped, it is worth the effort to first obtain a t ransposon insertion closely linked to the target locus before making F1 populations for screening. The mapped I elements shown (Fig, 7) will enable targeted tagging of closely linked genes. The En-I system described here shows abundant as well as late transpositions and may be an attractive alternative to Ac- Ds based t ransposon systems for targeted gene tagging.
Acknowledgements We thank Maarten Koornneef for helpful suggestions on handling Arabidopsis, for his aid and advice in mapping transposons and for critically reading the manuscript. Renate Schmidt was very helpful in mapping one of the I elements by YAC hybridization. Clare Lister and Caroline Dean were kind enough to distribute seeds (through the Nottingham Arabidopsis Stock Centre) and mapping data of their set of recombinant inbred lines prior to publication. DNA isolation from the RILs was done in cooperation with Ton Peeters, Ben Scheres, Pieter Vos and Casper Vroemen. Prof. Heinz Saedler is acknowledged for making available the En-I plasmid clones originally constructed at the Max-Planck-Institut fiir Ztichtungsforschung (Cologne). Gerrit Stunnenberg cared for most of the plants in the greenhouse, for which we are grateful. This work was supported by a grant (BIOT- CT90-0207) from the European Community under the BRIDGE T-project on 'Development and use of Arabidopsis thaliana as a tool for isolating genes of agronomic importance.'
References
Aarts MGM, Dirkse W, Stiekema WJ, Pereira A (1993) Transpo- son tagging of a male sterility gene in Arabidopsis. Nature 363:715-717
Baker B, Coupland G, Fedoroff N, Starlinger P, Schell J (1987) Phenotypic assay for excision of the maize controlling element Ac in tobacco. EMBO J 6:1547-1554
Bancroft I, Jones JDG, Dean C (1993) Heterologous transposon tagging of the DRL1 locus in Arabidopsis. Plant Cell 5: 631- 638
Belzile F, Yoder FI (1992) Pattern of somatic transposition in a high copy Ac tomato line. Plant J 2:173-179
Benfey PN, Chua N-H (1990) The cauliflower mosaic virus 35S promoter: combinatorial regulation of transcription in plants. Science 250:959-966
Cardon GH, Frey M, Saedler H, Gierl A (1991) Transposition of En/Spm in transgenic tobacco. Maydica 36:305-308
Cardon GI-t, Frey M, Saedler H, Gierl A (1993a) Mobility of the maize transposable element En/Spm in Arabidopsis thaliana. Plant J 3:773-784
Cardon GH, Frey M, Saedler H, Gierl A (1993b) Definition and characterization of an artificial En/Spm-based transposon tag- ging system in transgenic tobacco. Plant Mol Biol 23:157-178
Chuck G, Robbins T, Nijjar C, Ralston E, Courtney-Gutterson N, Dooner HK (1993) Tagging and cloning of a petunia flower colour gene with the maize transposable element Activator. Plant Cell 5:371-378
Cone K (1994) Transposon tagging with Spin. In: Freeling M, Walbot V (eds) The maize handbook, Springer-Verlag, New York, pp 240-242
Dash S, Peterson PA (1994) Frequent loss of the En transposable element after excision and its relation to chromosome replica- tion in maize (Zea mays L.). Genetics 136:653-671

564
Dellaporta SL, Woods J, Hicks JB (1983) A plant DNA miniprepa- ration version II. Plant Mol Biol Rep 4:19-21
Detrez C, Sangwan RS, Sangwan-Norreel BS (1989) Phenotypic and karyotypic status of Beta vulgaris plants regenerated from direct organogenesis in petiole culture. Theor Appl Genet 77:462-468
Fedoroff NV (1989) About maize transposable elements and de- velopment. Cell 56:181-191
Fedoroff N, Furtek D, Nelson O (1984) Cloning of the bronze locus in maize by a simple and generalizable procedure using the transposable controlling element Ac. Proc Natl Acad Sci USA 8I: 3825-3829
Feldmann KA (1991) T-DNA insertion mutagenesis in Arabidop- sis: mutational spectrum. Plant J 1:71-82
Frey M, Tavantzis SM, Saedler H (1989) The maize En-1/Spm element transposes in potato. Mol Gen Genet 217:172-177
Frey M, Reinecke J, Grant S, Saedler H, Gierl A (1990) Excision of the En/Spm transposable element of Zea mays requires two element-encoded proteins. EMBO J 9:4037-4044
Gierl A, Schwarz-Sommer Zs, Saedler H (1985) Molecular inter- actions between the components of the En-I transposable ele- ment system of Zea mays. EMBO J 4:579-583
Greenblatt IM, Brink RA ( 1961) Twin mutations in medium varie- gated pericarp maize. Genetics 47:489-501
Haring MA, Rommens CMT, Nijkamp HJJ, Hille J (1991) The use of transgenic plants to understand transposition mechanisms and to develop transposon tagging strategies. Plant Mol Biol 16: 449-461
Hauge BM, Hanley SM, Cartinhour S, Cherry JM, Gooodman HM, Koornneef M, Stare E Chang C, Kempin S, Medrano L, Meyerowitz EM (1993) An integrated genetic/RFLP map of the Arabidopsis thaliana genome. Plant J 3:745-754
Healy J, Corr C, DeYoung J, Baker B (1993) Linked and unlinked transposition of a genetically marked Dissociation element in transgenic tomato. Genetics 134:571-584
Kaulen H, Schell J, Kreuzaler F (1986) Light-induced expression of the chimeric chalcone synthase-NPTII gene in tobacco cells. EMBO J 5:1-8
Knapp S, Larondell Y, Rossberg M, Furtek D, Theres K (1994) Transgenic tomato lines containing Ds elements at defined ge- nomic positions as tools for targeted transposon tagging. Mol Gen Genet 243:666-673
Koncz C, Schell J (1986) The promoter of TL-DNA gene 5 controls the tissue-specific expression of chimaeric genes carried by a novel type of Agrobacterium binary vector. Mol Gen Genet 204:383-396
Koncz C, Mayerhofer R, Koncz-Kalman Z, Nawrath C, Reiss B, Redei GE Schell J (1990) Isolation of a gene encoding a novel chloroplast protein by T-DNA tagging in Arabidopsis thaliana. EMBO J 9:1337-1346
Lister C, Dean C (1993) Recombinant inbred lines for mapping RFLP and phenotypic markers in Arabidopsis thaliana. Plant J 4:745-750
Long D, Martin M, Sundberg E, Swinburne J, Puangsomlee P, Coupland G (1993) The maize transposable element system AciDs as a mutagen in Arabidopsis: identification of an albino mutation induced by Ds insertion. Proc Natl Acad Sci USA 90:10370-10374
Martin C, Carpenter R, Sommer H, Saedler H, Coen ES (1985) Molecular analysis of instability in flower pigmentation of An- tirrhinum majus, following isolation of the pallida locus by transposon tagging. EMBO J 4:1625-1630
Masson P, Fedoroff N (1989) Mobility of the maize Suppressor- mutator element in transgenic tobacco cells. Proc Natl Acad Sci USA 86:2219-2223
Masson P, Surosky R, Kingsbury J, FedoroffN (1987) Genetic and molecular analysis of the Spm-dependent a-m2 alleles of the maize a locus. Genetics 177:117-137
Masson E Strem M, Fedoroff N (1991) The tnpA and tnpD gene products of the Spm element are required for transposition in tobacco. Plant Cell 3:73-85
McClintock B (1954) Mutations in maize and chromosomal aber- rations in Neurospora. Carnegie Inst Wash Yearbook 53: 254- 260
Menssen A, H6hmann S, Martin W, Schnable PS, Peterson PA, Saedler H, Gierl A (1990) The En/Spm transposable element of Zea mays contains splice sites at the termini generating a novel intron from a dSpm element in the A2 gene. EMBO J 9:3051-3057
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473-497
Osborne BI, Corr CA, Prince JP, Hehl R, Tanksley SD, Mc- Cormick S, Baker B (1991) Ac transposition from T-DNA can generate linked and unlinked clusters of insertions in the toma- to genome. Genetics 129:833-844
Pereira A, Saedler H (1989) Transpositional behaviour of the maize En/Spm element in transgenic tobacco. EMBO J 8:1315-1321
Pereira A, Cuypers H, Gierl A, Schwarz-Sommer Zs, Saedler H (l 986) Molecular analysis of the En/Spm transposable element system of Zea mays. EMBO J 5:835-841
Peterson PA (1953) A mutable pale green locus in maize. Genetics 38:682-683
Peterson PA (1970) The En mutable system in maize. III. Transpo- sition associated with mutational events. Theor Appl Genet 40:367-377
Pietrzak M, Shillito RD, Hohn T, Potrykus 1 (1986) Expression in plants of two bacterial antibiotic resistance genes after proto- plast transformation with a new plant expression vector. Nucle- ic Acids Res 14:5857-5868
Raina R, Cook D, Fedoroff N (1993) Maize Spm transposable element has an enhancer-insensitive promoter. Proc Natl Acad Sci USA 90:6355-6359
Saedler H, Nevers P (1985) Transposition in plants: a molecular model. EMBO J 4:585-590
Schmidt R, Willmitzer L (1989) The maize autonomous element Activator (Ac) shows a minimal germinal excision frequency of 0.2%-0.5% in transgenic Arabidopsis thaliana plants. Mol Gen Genet 220:17-24
Scofield SR, English JJ, Jones JDG (1993) High level expression of the Activator transposase gene inhibits the excision of Dis- sociation in tobacco cotyledons. Cell 75:507-517
Shure M, Wessler S, Fedoroff N (1983) Molecular identification and isolation of the Waxy locus in maize. Cell 35:225-233
Stare P (1993) Construction of integrated genetic linkage maps by means of a new computer package: JoinMap. Plant J 3: 739- 744
Valvekens D, Van Montagu M, Van Lijsebettens M (1988) Agrobacterium tumefaciens mediated transformation of Ara- bidopsis thaliana root explants by using kanamycin selection. Proc Natl Acad Sci USA 85:5536-5540
Wing D, Koncz C, Schell J (1989) Conserved function in Nico- tiana tabacum of a single Drosophila hsp70 promoter heat shock element when fused to a minimal T-DNA promoter. Mol Gen Genet 219:9-16