developing a synthetic signal transduction system in plants
TRANSCRIPT

C H A P T E R T W E N T Y - F I V E
M
IS
D
ethods
SN 0
epartm
Developing a Synthetic Signal
Transduction System in Plants
Kevin J. Morey, Mauricio S. Antunes, Kirk D. Albrecht, Tessa
A. Bowen, Jared F. Troupe, Keira L. Havens, and June I. Medford
Contents
1. In
in
076
en
troduction
Enzymology, Volume 497 # 2011
-6879, DOI: 10.1016/B978-0-12-385075-1.00025-1 All rig
t of Biology, Colorado State University, Fort Collins, Colorado, USA
Else
hts
582
1
.1. S ignal transduction and synthetic biology 5821
.2. E ndogenous kinase-based signaling systems in plants 5832. F
oundation for Developing a Molecular Testing Platformfor HK Systems
5862
.1. E arly work showing plant HKs function in bacteria 5862
.2. D evelopment of a synthetic signaling systembased on bacterial TCS
5883. T
echnical Considerations in Developing a Eukaryotic SyntheticSignal Transduction System Based on Bacterial TCS Components
5893
.1. P roper membrane and compartment targeting 5903
.2. B acterial RRs: Signal dependent nuclear translocation andadaptation for transcriptional activation
5903
.3. C onsideration of codon bias when designing syntheticsignaling systems
5914. A
Partial Synthetic Signal Transduction System UsingCytokinin Input
5925. A
Eukaryotic Synthetic Signal Transduction Pathway 5936. C
onclusions 5957. P
rotocols 5977
.1. F luorometric GUS assay (fluorometer) 5977
.2. F luorometric GUS assay (microplate reader) 599Ackn
owledgments 599Refe
rences 599Abstract
One area of focus in the emerging field of plant synthetic biology is the
manipulation of systems involved in sensing and response to environmental
signals. Sensing and responding to signals, including ligands, typically involves
vier Inc.
reserved.
581

582 Kevin J. Morey et al.
biological signal transduction. Plants use a wide variety of signaling systems to
sense and respond to their environment. One of these systems, a histidine
kinase (HK) based signaling system, lends itself to manipulation using the tools
of synthetic biology. Both plants and bacteria use HKs to relay signals, which in
bacteria can involve as few as two proteins (two-component systems or TCS).
HK proteins are evolutionarily conserved between plants and bacteria and plant
HK components have been shown to be functional in bacteria. We found that
this conservation also applies to bacterial HK components which can function in
plants. This conservation of function led us to hypothesize that synthetic HK
signaling components can be designed and rapidly tested in bacteria. These
novel HK signaling components form the foundation for a synthetic signaling
system in plants, but typically require modifications such as codon optimization
and proper targeting to allow optimal function. We describe the process and
methodology of producing a synthetic signal transduction system in plants. We
discovered that the bacterial response regulator (RR) PhoB shows HK-depen-
dent nuclear translocation in planta. Using this discovery, we engineered a
partial synthetic pathway in which a synthetic promoter (PlantPho) is activated
using a plant-adapted PhoB (PhoB–VP64) and the endogenous HK-based cyto-
kinin signaling pathway. Building on this work, we adapted an input or sensing
system based on bacterial chemotactic binding proteins and HKs, resulting in a
complete eukaryotic signal transduction system. Input to our eukaryotic signal
transduction system is provided by a periplasmic binding protein (PBP), ribose-
binding protein (RBP). RBP interacts with the membrane-localized chemotactic
receptor Trg. PBPs like RBP have been computationally redesigned to bind
small ligands, such as the explosive 2,4,6-trinitrotoluene (TNT). A fusion
between the chemotactic receptor Trg and the HK, PhoR, enables signal trans-
duction via PhoB, which undergoes nuclear translocation in response to phos-
phorylation, resulting in transcriptional activation of an output gene under
control of a synthetic plant promoter. Collectively, these components produce
a novel ligand-responsive signal transduction system in plants and provide a
means to engineer a eukaryotic synthetic signaling system.
1. Introduction
1.1. Signal transduction and synthetic biology
A broad goal of synthetic biology is to engineer novel genetic circuits toproduce a specific and regulatable behavior (Khalil and Collins, 2010).Endogenous biological circuits that involve signal transduction allow phys-ical and molecular cues from an organism’s environment to activate acellular response (Kiel et al., 2010). In plants, signaling pathways are acti-vated in response to specific molecules, such as hormones and secondarymessengers ( Jaillais and Chory, 2010; Zhang and McCormick, 2009), andto a variety of environmental stimuli, such as light, temperature, abiotic

Developing a Synthetic Signal Transduction System in Plants 583
stress, certain types of bacteria, viruses, and herbivores (De Moraes et al.,2004). One approach to engineer novel synthetic genetic circuits takesadvantage of the mechanisms naturally used by endogenous signal transduc-tion pathways. These natural signaling pathways can then be modified bythe addition of synthetic components that interact in a similar way.
1.2. Endogenous kinase-based signaling systems in plants
Kinase-based signal transduction pathways represent a major means ofinformation transfer in plants and include: serine–threonine kinases(Rodriguez et al., 2010), histidine kinases (HKs; To and Kieber, 2008),and kinases with dual specificity for threonine and tyrosine (Oh et al., 2009;Sessa et al., 1996). These systems have been studied extensively, and there-fore could serve as starting points for building synthetic signal transductionpathways.
1.2.1. Light sensingLight is essential for every aspect of plant life. Plants regulate growth anddevelopment in response to light by modulating seed germination, seedlingde-etiolation, phototropism, circadian rhythms, and flowering time. Light issensed by at least three photoreceptors; phototropins, cryptochromes, andphytochromes, all of which have been shown to have kinase activity in thepresence of the appropriate wavelength (Christie, 2007; Osgur and Sancar,2006; Yeh and Lagarias, 1998). Phototropins respond to UV-A light, whilecryptochromes respond to blue light, and phytochromes respond to red andfar-red light. By integrating complex signals from different photoreceptors,plants are able to respond as needed to widely differing light conditions.
Phytochromes are the best studied class of light sensors. Phytochromescontain an N-terminal photo-sensory domain and a C-terminal regulatoryHK-related domain. There are five known phytochromes, each with bothunique and overlapping functions (Castillon et al., 2007). All switchbetween two reversible states depending on exposure to red or far-redlight. Red light switches phytochromes into a biologically active state,whereas “dark-decay” or far-red light reverses this process and inactivatesphytochromes. The active phytochromes translocate to the nucleus anddirectly interact with transcription factors to regulate expression ofthousands of light responsive genes (Sharrock, 2008). While light sensingproteins have been previously used in synthetic biology (Levskaya et al.,2009), a plant synthetic signaling pathway that is based on phytochromesignaling would most likely be influenced by light, making it difficult tocontrol such a pathway independent of endogenous processes.

584 Kevin J. Morey et al.
1.2.2. Plant signaling based on MAP kinasesAnother endogenous kinase system, mitogen-activated protein kinase(MAPK) signal transduction, mediates responses to environmental and devel-opmental cues in plants. MAPK signaling typically involves a tyrosine orserine–threonine phosphorylation cascade between its membrane receptorand MAP3Ks, MAP2Ks, and MAPKs. MAPKs in turn regulate activity ofdownstream kinases, other enzymes, and transcription factors, effectingresponses to abiotic stress and hormones. In Arabidopsis, 60 MAP3Ks, 10MAP2Ks, and 20 MAPKs have been identified, although the function ofmany of these proteins has yet to be determined (Rodriguez et al., 2010).This multitude of MAP kinases requires a multitude of spatial–temporal con-trols, specific phosphatases, scaffolding proteins, and feedback controls tomaintain signal specificity.
One type of MAP kinase signaling involves the plant hormone ethylene.Ethylene is a regulator of a variety of processes including germination, growth,development, fruit ripening, and stress responses. Plant transmembrane recep-tors for ethylene are localized to the endoplasmic reticulum (ER). Whenethylene is bound the receptor complex is inactivated and a MAPK signalingcascade is initiated, activating transcription of primary ethylene response genes.However, the sensitivity of this response is thought to differ among tissues anddevelopmental stages (Stepanova andAlonso, 2009;Yoo et al., 2009), revealingyet another measure of MAP kinase signaling complexity.
The complexity ofMAPkinase-based signaling is amajor drawback in usingthese components to build a synthetic signal transduction pathway; a largenumber of MAPK proteins need to be expressed to obtain a desired response.Furthermore, a synthetic signal pathway built out of parts from this systemwould likely suffer from significant unwanted crosstalk with endogenous sig-naling components. This complication would make testing a synthetic systemand obtaining the desired specificity with MAP kinase components difficult.
1.2.3. Cytokinin signaling based on histidine kinasesIn addition to tyrosine and serine–threonine kinase-based signaling, plantsuse a HK signal transduction system to respond to the hormone cytokinin.Like other plant hormones, cytokinins affect a number of processes such asbranching, chloroplast development, leaf senescence, some stress responses,and pathogen resistance (Heyl and Schmulling, 2003). Early work in this arearevealed that plant HKs used in cytokinin perception have homology to thefirst HK systems discovered, bacterial two-component systems (TCS).
Bacterial TCS range from simple to complex and control a variety ofresponses that include: chemotaxis (nutrient sensing), osmotic sensing,quorum sensing, regulation of pathogenesis, redox sensing, and sporulation(Parkinson and Kofoid, 1992). The simplest bacterial TCS use only twoproteins: a membrane-localized HK, and a soluble, cytoplasm-localized

Developing a Synthetic Signal Transduction System in Plants 585
response regulator (RR). Structurally, a typical membrane-localized HKprotein consists of an N-terminal extracellular receptor, a transmembranealpha helix, and a C-terminal cytoplasmic region, which contains a dimer-ization and histidine phosphorylation (DHP) domain and a hATPasedomain. In a simple TCS, a receptor HK binds a ligand or recognizes astimulus outside the cell. The receptor then undergoes a conformationalchange that is propagated by a transmembrane alpha helix (transmembranesignaling) leading to activation of the cytoplasmic region. This activationcauses autophosphorylation of a histidine residue on the DHP domain. Thehigh energy phosphate is then transferred from theHK to an aspartate residueon its cognate RR (Fig. 25.1A). Upon phosphorylation, many RRs (e.g.,OmpR/PhoB family) undergo a conformational change that activates theireffector domain. In many cases the effector domain binds DNA and activatestranscription, controlling expression of downstream genes involved in the
HK
Ligand Ligand
DHP
hATPase
DHP
hATPase
HHP
PD
RRHK
Receiverdomain
HP
P
D
D
RR
Hpt
RR
HK
Membrane
PBP-Ligandcomplex
H
W
A
CheW
CheAP
D
cytoplasm
Two-component systems discussed in chapter
A B C
Figure 25.1 HK-based signaling systems in bacteria and plants: (A) “Simple” bacterialTCS—extracellular input is provided by a transmembrane HK. In the HK molecule, ahistidine residue in the dimerization and histidine phosphorylation (DHP) domainautophosphorylates in response to ligand binding. The high energy phosphate is thentransferred to a cytoplasmically localized response regulator (RR). (B) Hybrid HK(e.g., AHKs, RcsC) signaling found in plants and bacteria—these systems have morecomplex HKs and signaling components. In response to a ligand binding, a histidineresidue in the DHP domain autophosphorylates. The high energy phosphate is thentransferred internally to an aspartate in the receiver domain, and subsequently to ahistidine in a cytoplasmic Hpt protein (called AHPs in Arabidopsis), which thenphosphorylates an RR on its aspartate residue. (C) Bacterial chemotactic TCS—achemotactic receptor (e.g., Trg) binds a PBP–ligand complex or directly binds a ligand.This binding event initiates a signal that is transmitted to a cytoplasm-localized histidinekinase CheA. CheW acts as an adaptor protein for the chemotactic receptor/CheAcomplex. CheA then phosphorylates RRs that do not activate transcription.

586 Kevin J. Morey et al.
response to the ligand or stimulus. More elaborate TCS include morecomplex HK proteins, multiple RRs, and a third component, histidinephosphotransfer (Hpt) proteins (Fig. 25.1B). Regardless of the added com-plexity, one hallmark of all known TCS is that the high energy phosphatemoiety is always relayed between histidine and aspartate residues.
Plant HK systems are similar to the hybrid TCS found in bacteria andinvolve complex HK receptors with multiple RRs. Three cytokinin-responsive receptor HKs have been identified in Arabidopsis, AHK2,AHK3, and AHK4/CRE1. Upon binding cytokinin, the AHKs autopho-sphorylate at a histidine residue, then undergo an internal phosphotransferto an aspartate residue located on the C-terminal receiver domain. Thisphosphate is transferred to a histidine residue on a Hpt protein (known asAHPs in Arabidopsis) (Fig. 25.1B). AHPs have been shown to relay thephosphate moiety to nuclear-localized RRs (known as ARRs in Arabidop-sis), as well as cytoplasm-localized cytokinin response factors (CRFs). Uponactivation by aspartate phosphorylation, ARRs control expression of genesinvolved in cytokinin responses (Argueso et al., 2009; To and Kieber, 2008).
This similarity in the signaling mechanism is corroborated by evolution-ary conservation of the components involved in bacterial TCS and plantHK systems. Protein sequence alignment of the respective phosphotransferand/or phospho-accepting domains of Escherichia coli and Arabidopsis HKs,Hpts, and RRs show significant homology, showing these domains areevolutionarily conserved (Fig. 25.2). Moreover, phylogenetic analyses sug-gest plants obtained these genes from bacteria via lateral gene transfer(Koretke et al., 2000). Collectively, these data and results from the Mizunolaboratory (Section 2.1), lead us to hypothesize that bacteria and plant HKcomponents may have interchangeable functionality.
The evolutionary conservation of plant and bacterial HK signaling sys-tems provides an attractive platform for developing a eukaryotic syntheticsignal transduction pathway. First, because bacterial TCS use only twoproteins from stimulus to a response, they provide a framework for a syntheticsystem with a minimal number of components. Second, if the componentsconserve functionality, it will provide a rapid testing platform (bacteria)where components can be rationally designed and techniques such as directedevolution applied prior to introduction into a eukaryotic host.
2. Foundation for Developing a Molecular
Testing Platform for HK Systems
2.1. Early work showing plant HKs function in bacteria
Evidence that the Arabidopsis proteins AHK2, AHK3, and AHK4/CRE1function as cytokinin receptorswas obtained frombacterial studies.These plantHKs in bacterial cells complement a deletion mutant of the E. coli HK RcsC

Sequence conservation Black=good Grey=badPhosphorylation site is marked by a box
A HK DHP domain
B Hpt phosphotransfer domain
C RR receiver domain
Figure 25.2 T-Coffee (Notredame et al., 2000) alignment of conserved phosphotrans-fer and phosphoaccepting domains of Arabidopsis and E. coli HKs, Hpts, and RRs.These domains are conserved in both prokaryotes and eukaryotes.
Developing a Synthetic Signal Transduction System in Plants 587
and properly initiate a signal in the presence of the plant hormone cytokinin(Suzuki et al., 2001;Yamada et al., 2001).RcsC is a hybridHK (Fig. 25.1B) thatsignals, via phosphorelay, to an Hpt factor (YojN) and a RR (RcsB), andactivates the capsular polysaccharide synthesis operon (RcsC ! YojN !RcsB) (Takeda et al., 2001). The Arabidopsis HKs are able to functionallyreplace RcsC and interact with the bacterial Hpt YojN to produce thesignaling pathway, AHK ! YojN ! RcsB. In addition, AHK4 has also

588 Kevin J. Morey et al.
been coexpressed with the Arabidopsis Hpts (AHP1, 2, 3, and 5) in a signaltitration assaywhich established thatAHK4 is capable of interactingwithAHPsin bacteria (Suzuki et al., 2001). These results demonstrate that not only is itpossible to functionally express heterologous plant HK signaling proteins inbacteria, but also that these heterologous components can interact or “crosstalk” with endogenous bacterial components.
These experiments provided a basis for our work, whereby various com-ponents fromplant or syntheticHK systems can be rapidly tested and improvedin a bacterial system prior to functional adaptation for plants. The ability to usebacteria to test synthetic eukaryotic signaling components facilitates the use ofsynthetic design and other experimental approaches, such as directed evolu-tion. In addition, basic questions about aspects of transmembrane signaling andreceptor function can be addressed using this system.
2.2. Development of a synthetic signaling systembased on bacterial TCS
The functional expression of plant HK signaling components in bacteria(Suzuki et al., 2001) lead us to hypothesize that the reciprocal might also bepossible, that is, bacterial components could function in plants. If plant HKcomponents were used to engineer a plant synthetic signal transduction systemthere is likely to be unwanted interference from endogenous HK signaling,which in Arabidopsis involves 28 AHPs and ARRs (Dortay et al., 2006).
When designing a synthetic signaling pathway one needs to address howthe signal is initiated (input), propagated (transmission), and finally, what isactivated by the signal (output). The signal transmission needs to be regulatedby an input dedicated to the synthetic signaling pathway to prevent nonspe-cific activation of the pathway. Periplasmic binding proteins (PBPs), specifi-cally those from the bacterial chemotactic TCS, constitute a framework forsynthetic signaling input. PBPs are a family of proteins involved in thescavenging of a wide variety of substances in bacteria. They undergo a distinctconformational change from an open form (without ligand) to a ligand boundclosed form able to initiate signal transduction. This conformational changepresented us with a relatively “clean” input, that is, an unbound PBP will notinitiate signal transduction. In addition, the PBP–ligand binding pocket ishighly specific to its particular ligand and the protein itself can be used as ascaffold for computational redesign, providing a broad potential for input(Dwyer and Hellinga, 2004). Computationally redesigned PBPs have beenshown to bind novel molecules, including a surrogate for a nerve toxin, themetal zinc, L-lactate, and 2,4,6-trinitrotoluene (TNT) (Allert et al., 2004;Dwyer and Hellinga, 2004; Dwyer et al., 2003, 2004; Looger et al., 2003).Initial attempts to change binding specificity of PBPs focused on computa-tionally redesigning the binding site, but simply modifying the residues in thebinding site may introduce unintentional problems into the structural and

Developing a Synthetic Signal Transduction System in Plants 589
binding stability of the PBP (Schreier et al., 2009; Vercillo et al., 2007).Despite stability issues, computationally redesigned PBPs have been shownto activate TCS in bacteria (Looger et al., 2003; Rodrigo et al., 2007).
Once the signal is initiated by the PBP outside of the cell, it needs to betransmitted to the interior. Transmembrane signaling for a synthetic pathwaycan also be based on bacterial TCS. In E. coli, a subset of PBPs are capable ofinteracting with the chemotactic receptors Trg, Tar, Tsr, and Tap, and initiat-ingaTCSsignalingcascade that allows thebacteria to respond to thepresenceofcertain sugars (glucose, ribose,maltose, galactose), aminoacids (asp, glu, ser, ala,gly), and dipeptides, by directing the cells to environments containing thesenutrients (Mowbray and Sandgren, 1998; Fig. 25.1C). However, while thesignal is transmitted to the interior of the cell through the chemotactic receptor,theRRs receiving the chemotactic signal do not activate transcription.Hence,to produce a transcriptional output with PBP input, we engineered a fusionprotein between the extracellular portion of a chemotactic receptor and thecytoplasmic portion of an HK that is capable of interacting with a RRthat can activate transcription. These receptor/HK fusions were developedand tested in the rapidbacterial testing systemand further discussed inSection5.
The third component of a synthetic signaling system is the “output,” whichtypically consists of transcriptional activation of one or more genes, leading to adetectable phenotype. Therefore, for a eukaryotic organism, the synthetic sig-naling system requires a RR capable of both accepting the signal from atransmembrane HK and subsequently localizing to the nucleus to activatetranscription in response to the signal. Work in our laboratory (described inSection4) established that somebacterialRRs (PhoB andOmpR) are capable ofsignal-dependent nuclear translocation inArabidopsis (Antunes et al., 2009).Wechose PhoB as theRR for our synthetic signaling systembecause it shows strongsignal-dependent nuclear translocation and it has a simple activationmechanism,allowing us to develop a plant synthetic signal transduction pathway.
In the following sections, we describe considerations for properlyexpressing synthetic components in bacteria or plants, how the partialsignaling system was engineered, how a complete pathway was assembled,and finally how system output was quantified using the reporter geneb-glucuronidase (GUS).
3. Technical Considerations in Developing a
Eukaryotic Synthetic Signal Transduction
System Based on Bacterial TCS Components
During the process of developing a functional synthetic signal trans-duction pathway in plants we found it necessary to address a number oftechnical issues to ensure the bacterial components were properly expressed.

590 Kevin J. Morey et al.
Differences in cell structure, transcription activity, and codon usagebetween prokaryotes and eukaryotes can all produce problems if notaccounted for. The following sections address these concerns.
3.1. Proper membrane and compartment targeting
When developing a eukaryotic synthetic signaling system that uses bacterialcomponents, it is important to consider some fundamental differencesbetween prokaryotic and eukaryotic cells, one of which is their distinctsubcellular structure. In membrane-localized bacterial HKs, the transmem-brane segments are sufficient for proper targeting to the plasma membrane.Eukaryotic cells have multiple membrane-enclosed organelles; thus, anyprotein localized in a particular membrane needs to contain a signal peptidethat specifically targets it to that membrane. To ensure proper membranelocalization in plants, we added the signal peptide from the plasma mem-brane-localized Arabidopsis flagellin receptor FLS2 (Gomez-Gomez andBoller, 2000) to the N-terminal end of HKs.
We also had to target the aforementioned PBPs as plants do not have aperiplasm. Instead, plants have a space outside the cell plasma membraneknown as the apoplast, where small proteins like PBPs can freely diffuse andcontact exogenous substances (Somerville et al., 2004).We targeted the PBPsto the apoplast using a plant signal peptide that is normally used to secreteproteins to the apoplastic pollen-stigma matrix (Baumberger et al., 2003).
3.2. Bacterial RRs: Signal dependent nuclear translocationand adaptation for transcriptional activation
The evolutionary conservation of TCS genes between bacteria and plantsallows some functional interchange of individual proteins from one organ-ism to another. However, a fundamental difference between bacteria andplants is the compartmentalization of transcriptional responses in plants tothe nucleus. And, although bacterial cells do not have a nucleus, manybacterial proteins related to pathogenicity show nuclear localization ineukaryotic cells (Kay and Bonas, 2009). In addition, protein sequencesthat function as nuclear localization signals are commonly short sequencesof positively charged amino acids. DNA-binding domains, such as thosefound in bacterial RRs, are also known to contain positively charged aminoacids. This foundational knowledge provided us a framework to investigatebacterial RRs in plants.
In order to build a functional synthetic system in plants, a bacterial RRmust receive the transmembrane signal in the cytoplasm and translocate tothe nucleus to activate transcription. Previous work in our laboratory(below and Antunes et al. 2009) established that some bacterial RRs arecapable of signal-dependent nuclear translocation. We then modified the

Developing a Synthetic Signal Transduction System in Plants 591
bacterial RR PhoB to interact with eukaryotic RNA polymerase II andactivate transcription in plants. A commonly used transcriptional activator isthe C-terminal domain of VP16, a herpes simplex virus encoded protein.VP16 has been shown to activate transcription in eukaryotic cells(Triezenberg et al., 1988), and tetrameric repeats (VP64) of amino acids437–447 of VP16 fused to DNA-binding domains have been shown toproduce efficient transcriptional activators in eukaryotes (Seipel et al., 1992).Hence, we fused VP64 to the C-terminus of PhoB to produce PhoB–VP64,a modified or synthetic PhoB that can activate endogenous eukaryotictranscription machinery.
3.3. Consideration of codon bias when designing syntheticsignaling systems
One last consideration when engineering a synthetic signaling system iscodon usage bias. The presence of codon usage bias among differentorganisms has been well documented (Batard et al., 2000; Lessard et al.,2002; Suo et al., 2006). The preference of one codon over another by anorganism can be a barrier to expressing bacterial genetic circuits in plants ortesting plant proteins in bacteria. Codon optimization of heterologouslyexpressed genes can improve expression levels and, in some cases, simplyallow a gene to be heterologously expressed (Perlak et al., 1991).
Our system presents a stark example of conflicting codon usage bias.Four of the seven rare codons utilized by E. coli (used at a frequency< 0.5%) code for the amino acid arginine (Chen and Texada, 2006).These codons account for over 74% of the arginine codon usage in Arabi-dopsis (Nakamura et al., 2000), with the extremely rare AGG and AGAcodons making up 57% of Arabidopsis usage. Hence, if not addressed whenexpressing heterologous genes, rare codon clustering can lead to drasticallyreduced protein levels and mRNA degradation (Li et al., 2006; Sunoharaet al., 2004). For instance, the AHK4/CRE1 gene contains 40 of the rarearginine codons, including two arginine codons rare to E. coli appearingcontiguously at positions 16 and 17, as well as at positions 133 and 134. Ourlaboratory has encountered gene instability/toxicity issues when expressingthis protein in E. coli, a result that has also been reported elsewhere (Mizunoand Yamashino, 2010).
The effect of codon usage bias can be complicated during cloning inbacterial cells by the unwanted activity of promoters used to drive plantgenes. The CaMV35S promoter, commonly used for constitutive expres-sion of genes in plants, has been shown to have activity in E. coli (Assaad andSigner, 1990). Therefore, cloning a gene that has been codon-optimizedfor plants downstream of the CaMV35S promoter could potentially leadto instability in bacterial cells if the gene has a large number of rareE. coli codons. Hence, codon usage may need to be considered

592 Kevin J. Morey et al.
when simply cloning plant genes in E. coli as well as when developing abacterial testing system.
4. A Partial Synthetic Signal Transduction
System Using Cytokinin Input
One challenging aspect of developing a eukaryotic signal transductionpathway that is activated by an external input is moving the signal from themembrane to bring about a transcriptional response in the nucleus. Plasmamembrane to transcriptional responses in eukaryotes typically involve mul-tiple proteins which themselves interact with numerous other proteins. Incontrast bacterial signaling systems can use as few as one protein frommembrane to transcriptional response.
Given these considerations, we experimentally tested a simple membraneto nucleus signaling system in eukaryotes. Components from simpler systems(bacteria and yeast) were expressed in plants as N-terminal fusions to smGFP(Davis and Vierstra, 1996). As an initial means to activate HK signaling inplants we used application of exogenous cytokinin. We tested the bacterialRRsOmpR (Mizuno et al., 1982;Wurtzel et al., 1982), PhoB (Makino et al.,1986, 1989), andRcsB (Chen et al., 2001), a bacterialHpt, YojN (Chen et al.,2001), and the yeast Hpt, Ypd1 (Posas et al., 1996) in transient assays. PhoBand OmpR appeared to show signal-dependent nuclear accumulation inplant cells, whereas the responses of RcsB, YojN, and Ypd1 were equivocal.Therefore, we focused our subsequent work on PhoB and OmpR.
To corroborate these transient assay results, we then generated stabletransgenic Arabidopsis plants that constitutively expressed either PhoB-GFPor OmpR-GFP (Antunes et al., 2009). Plants were grown for 6-8 days, andimages were collected showing the subcellular distribution of GFP in roots,leaves, and hypocotyls. The same tissues were treated with 1 mM t-zeatin (acytokinin) and re-imaged to determine the effect of cytokinin mediated HKactivation on the bacterial RRs. To verify whether the compartment whereRR-GFP localized corresponded to nuclei, the roots were stained with1 ng/mL DAPI for 10 min. Images were collected using either a NikonDiaphot fluorescence microscope, or a Carl Zeiss LSM 510META confocalmicroscope. These results, and others detailed in Antunes et al. (2009)indicated that bacterial RRs accumulate in the nucleus in response to acytokinin-initiated signal and could, therefore, be used as a component of asynthetic signal transduction pathway in plants.
To produce a PhoB responsive promoter in a plant, we designed andconstructed a synthetic promoter entitled PlantPho, which consists of fourcopies of PhoB’s DNA-binding element (Pho box) placed upstream of aminimal promoter (�46 region from the CaMV35S promoter) (Antunes

Developing a Synthetic Signal Transduction System in Plants 593
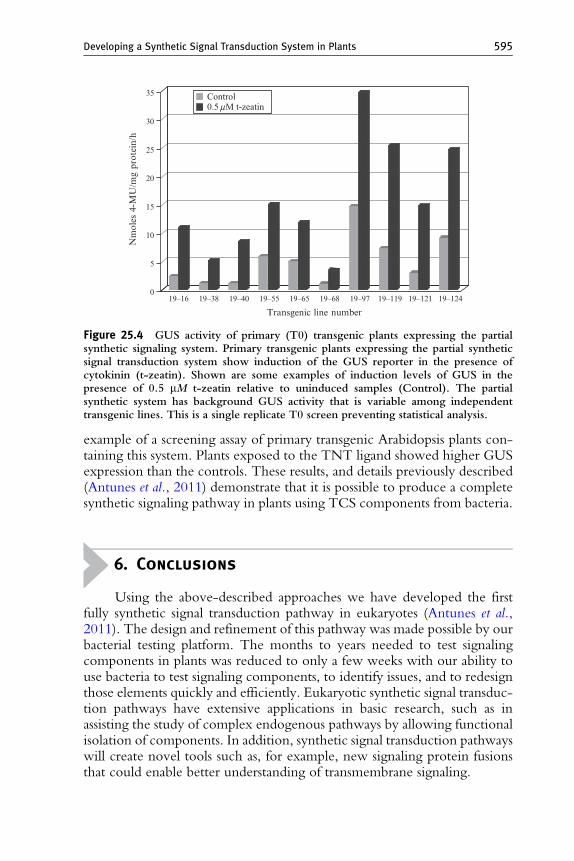
et al., 2009). To test if the PlantPho promoter was responsive to cytokinin-mediated HK activation, we placed the GUS reporter gene ( Jefferson et al.,1987) under its control and generated transgenic Arabidopsis plants thatconstitutively expressed PhoB–VP64. The partial synthetic signaling systemis diagrammed in Fig. 25.3. GUS expression is assayed by incubatingdevelopmentally equivalent leaves (leaf 1 and leaf 2) from transgenic plantsin the presence or absence of 10 mM t-zeatin for 16 h, followed by totalprotein extraction and GUS activity measurement in the presence of theenzyme’s substrate, 4-methylumbelliferyl b-D-glucuronide (MUG).Enzyme activity results in the production of a fluorescent product,4-methylumbeliferone (4-MU). GUS activity is then normalized to a4-MU standard curve and the total protein content of the sample, as deter-mined by the Bradford assay. Figure 25.4 is an example of a screening assay ofprimary transgenic Arabidopsis plants containing the partial synthetic system.Results indicate that cytokinin can activate a partial synthetic signalingsystem in plants. These experiments provide information on two criticalsteps in developing a eukaryotic signaling pathway with bacterial compo-nents: bacterial RRs are capable of signal-dependent nuclear translocationand, once nuclear localized, they are able to activate a synthetic promoter(provided they contain a eukaryotic transcriptional activation domain).
5. A Eukaryotic Synthetic Signal
Transduction Pathway
While the partial synthetic system provides two key aspects of apathway, transmission and output, it still lacks specific input from anexogenous signal and transmembrane signaling. To produce an entirelysynthetic signal transduction pathway, we replaced the input from thenative cytokinin sensing components of the partial system described abovewith synthetic components that are capable of sensing a specific substance ofinterest and transmitting this information across the membrane. As men-tioned previously, when a PBP binds its ligand a conformational change isinduced, leading to increased affinity of the PBP–ligand complex for theextracellular domain of a bacterial chemotactic receptor, Trg, with a specificPBP, ribose-binding protein (RBP). We used the computationally rede-signed RBP, TNT.R3, that binds the explosive TNT (Looger et al., 2003)to provide a specific input to our synthetic signal transduction pathway. TheRR in our partial synthetic signaling system, PhoB–VP64 is capable ofreceiving a phosphate from PhoB’s cognate HK, PhoR (Lamarche et al.,2008).We therefore linked input from the computationally redesigned PBPsto our partial signal transduction system by producing a functional receptorHK fusion between Trg (to bind the ligand-redesigned PBP complex) and

CK
P P
GUS
AHK
PhoB-VP64
AHP
Nucleus
– 46 CaMV35S
PhoB DNAbinding site
Apoplast
Cell wall
Membrane
Figure 25.3 Diagram of the partial synthetic signaling system in plants. Endogenousmembrane-localized cytokinin receptor HKs (AHK2, AHK3, and AHK4) autopho-sphorylate upon binding cytokinin resulting in a phosphorelay to endogenous AHPs.We postulate the AHPs then phosphorylate the synthetic RR, PhoB–VP64. PhoB–VP64 translocates to the nucleus and activates a synthetic PlantPho promoter consistingof four PhoB DNA-binding domains upstream of a minimal 35S promoter. CK,cytokinin; P, phosphate moiety.
594 Kevin J. Morey et al.
PhoR to activate PhoB–VP64. A Trg:PhoR fusion that links Trg at itsHAMP domain to position M19 of PhoR was produced by experimentalanalysis in bacteria (Antunes et al., 2011). We re-engineered the Trg–PhoRfusion for plant expression by adding an N-terminal plasma membranetargeting signal peptide from FLS2 (Gomez-Gomez and Boller, 2000) toproduce Fls:Trg:PhoR. The genes encoding ssTNT.R3 and Fls:Trg:PhoRwere placed downstream of the constitutive CaMV35S and nopalinesynthase (PNOS) promoters, respectively, then combined with the partialsynthetic system to generate transgenic Arabidopsis plants expressing thesynthetic signal transduction pathway: ssTNT.R3 ! Fls:–Trg:–PhoR !PhoB–VP64 ! PlantPho promoter::GUS (Fig. 25.5). To test the functionof our system in plants, we measured GUS activity in primary transgenicplants by incubating developmentally equivalent leaves (leaf 1 and leaf 2) inthe presence or absence of 10 mMTNT for 16 h. Protein extraction andGUSactivity were performed as described in Section 7. Figure 25.6 shows an
25
30
35 Control0.5mM t-zeatin
20
15
10
Nm
oles
4-M
U/m
g pr
otei
n/h
5
019–16 19–38 19–40 19–55 19–65 19–68 19–97 19–119 19–121 19–124
Transgenic line number
Figure 25.4 GUS activity of primary (T0) transgenic plants expressing the partialsynthetic signaling system. Primary transgenic plants expressing the partial syntheticsignal transduction system show induction of the GUS reporter in the presence ofcytokinin (t-zeatin). Shown are some examples of induction levels of GUS in thepresence of 0.5 mM t-zeatin relative to uninduced samples (Control). The partialsynthetic system has background GUS activity that is variable among independenttransgenic lines. This is a single replicate T0 screen preventing statistical analysis.
Developing a Synthetic Signal Transduction System in Plants 595
example of a screening assay of primary transgenic Arabidopsis plants con-taining this system. Plants exposed to the TNT ligand showed higher GUSexpression than the controls. These results, and details previously described(Antunes et al., 2011) demonstrate that it is possible to produce a completesynthetic signaling pathway in plants using TCS components from bacteria.
6. Conclusions
Using the above-described approaches we have developed the firstfully synthetic signal transduction pathway in eukaryotes (Antunes et al.,2011). The design and refinement of this pathway was made possible by ourbacterial testing platform. The months to years needed to test signalingcomponents in plants was reduced to only a few weeks with our ability touse bacteria to test signaling components, to identify issues, and to redesignthose elements quickly and efficiently. Eukaryotic synthetic signal transduc-tion pathways have extensive applications in basic research, such as inassisting the study of complex endogenous pathways by allowing functionalisolation of components. In addition, synthetic signal transduction pathwayswill create novel tools such as, for example, new signaling protein fusionsthat could enable better understanding of transmembrane signaling.
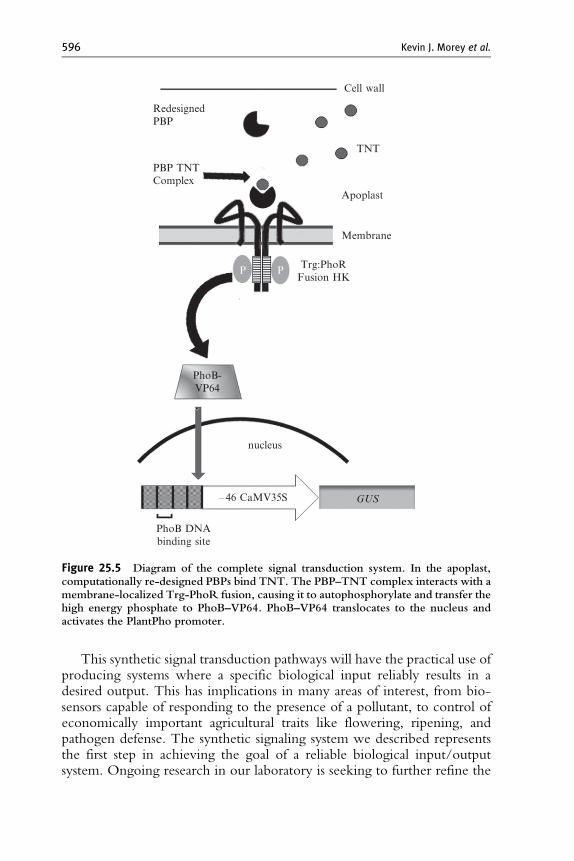
Cell wall
TNT
Apoplast
RedesignedPBP
PBP TNTComplex
Membrane
Trg:PhoRFusion HK
P P
GUS
PhoB-VP64
nucleus
– 46 CaMV35S
PhoB DNAbinding site
Figure 25.5 Diagram of the complete signal transduction system. In the apoplast,computationally re-designed PBPs bind TNT. The PBP–TNT complex interacts with amembrane-localized Trg-PhoR fusion, causing it to autophosphorylate and transfer thehigh energy phosphate to PhoB–VP64. PhoB–VP64 translocates to the nucleus andactivates the PlantPho promoter.
596 Kevin J. Morey et al.
This synthetic signal transduction pathways will have the practical use ofproducing systems where a specific biological input reliably results in adesired output. This has implications in many areas of interest, from bio-sensors capable of responding to the presence of a pollutant, to control ofeconomically important agricultural traits like flowering, ripening, andpathogen defense. The synthetic signaling system we described representsthe first step in achieving the goal of a reliable biological input/outputsystem. Ongoing research in our laboratory is seeking to further refine the
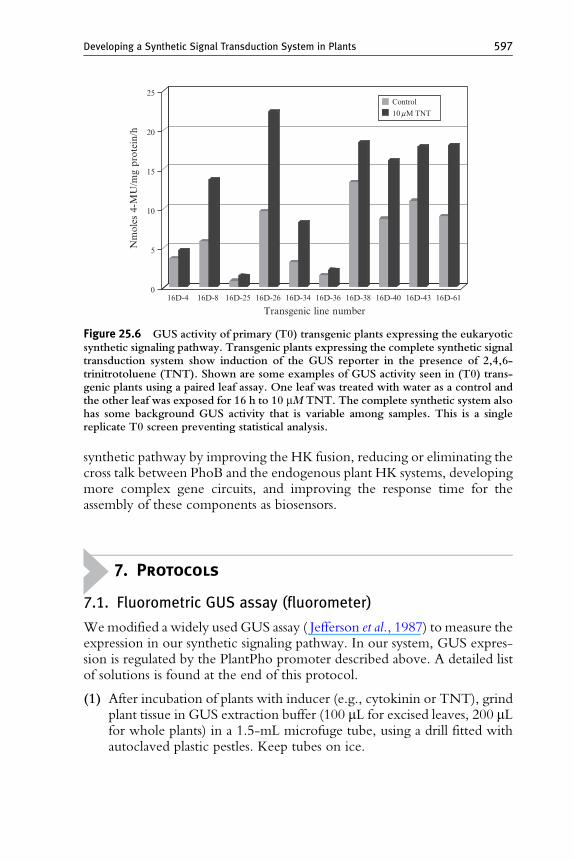
25Control10mM TNT
20
15
10
Nm
oles
4-M
U/m
g pr
otei
n/h
5
016D-4 16D-8 16D-25 16D-26 16D-34
Transgenic line number16D-36 16D-38 16D-40 16D-43 16D-61
Figure 25.6 GUS activity of primary (T0) transgenic plants expressing the eukaryoticsynthetic signaling pathway. Transgenic plants expressing the complete synthetic signaltransduction system show induction of the GUS reporter in the presence of 2,4,6-trinitrotoluene (TNT). Shown are some examples of GUS activity seen in (T0) trans-genic plants using a paired leaf assay. One leaf was treated with water as a control andthe other leaf was exposed for 16 h to 10 mM TNT. The complete synthetic system alsohas some background GUS activity that is variable among samples. This is a singlereplicate T0 screen preventing statistical analysis.
Developing a Synthetic Signal Transduction System in Plants 597
synthetic pathway by improving the HK fusion, reducing or eliminating thecross talk between PhoB and the endogenous plant HK systems, developingmore complex gene circuits, and improving the response time for theassembly of these components as biosensors.
7. Protocols
7.1. Fluorometric GUS assay (fluorometer)
Wemodified a widely used GUS assay ( Jefferson et al., 1987) to measure theexpression in our synthetic signaling pathway. In our system, GUS expres-sion is regulated by the PlantPho promoter described above. A detailed listof solutions is found at the end of this protocol.
(1) After incubation of plants with inducer (e.g., cytokinin or TNT), grindplant tissue in GUS extraction buffer (100 mL for excised leaves, 200 mLfor whole plants) in a 1.5-mL microfuge tube, using a drill fitted withautoclaved plastic pestles. Keep tubes on ice.

598 Kevin J. Morey et al.
(2) Centrifuge samples at 10,000 rpm (9,000 g) for 10–15 min at 4 �C.(3) Transfer the cleared supernatant to a new tube and place it on ice.(4) In a 0.5-mL microfuge tube combine 50 mL of the protein extract
obtained above with 50 mL of GUS assay buffer (2 mM MUG in GUSextraction buffer).
(5) Mix well and centrifuge for 15 s.(6) Incubate reaction at 37 �C for 1 h.(7) Stop the reaction by mixing 50 mL of the reaction mixture into 1.95 mL
carbonate stop buffer (0.2MNa2CO3, see Solutions) contained in a 4-mLtube (avoid excessive light exposure of tubes because 4-MU is lightsensitive).
(8) Read 4-MU fluorescence units of samples in a DyNA Quant 200Fluorometer (Hoefer, Inc., San Francisco, CA), excitation 365 nm,emission 460 nm, using a 4-MU standard solution to calibrate theinstrument. Calibrate by first adding 1.9 mL of carbonate stop bufferto the cuvette and zero the instrument, then add 100 mL of the 4-MUstandard solution to the stop buffer (see Solutions), and set the instru-ment to 500 units to standardize.
(9) Transfer 20 mL of the remaining protein extract to a new 1.5 mLcentrifuge tube and read the protein concentration using the Bradfordprotein assay at 595 nm. Use bovine serum albumin solution diluted inextraction buffer (described above) as a standard, ranging in concentra-tion from 0 to 0.6 mg/mL.
Solutions:
(1) GUS extraction buffer: final concentrations: 50 mM NaHPO4, pH 7.0,10 mM b-mercaptoethanol, 10 mMNa2EDTA, pH 8.0, 0.1% sarcosyl,0.1% Triton X-100.
(2) GUS assay buffer (2 mMMUG in GUS extraction buffer): Mix 22 mg ofMUG with 25 mL of GUS extraction buffer.
(3) Carbonate stop buffer (0.2 M Na2CO3): Weigh out 6.2 g Na2CO3�H2Oand dissolve it in 250 mL of diH2O.
(4) 4-MU standard solution. First, prepare a concentrated MU calibrationstock solution containing 1 mM 7-hydroxy-4-methylcoumarin, MU, indiH2O. Mix 19.8 mg (0.0198 g) of MU into 100 mL of diH2O toproduce a 1 mM solution. Dilute 10 mL of the 1 mMMU solution into10 mL of diH2O to make the 1 mM MU stock (store this solution at4 �C, protected from light).
(5) For calibration of the fluorometer, make a 50 nM MU solution, bydiluting 100 mL of the 1 mM concentrated MU calibration stocksolution prepared above into 1.9 mL carbonate stop buffer. This solu-tion must be made fresh just before use.

Developing a Synthetic Signal Transduction System in Plants 599
7.2. Fluorometric GUS assay (microplate reader)
(1) Remove leaves 1 and 2 from 14-day-old Arabidopsis and incubatethem overnight in the presence or absence of the inducer of the genecircuit.
(2) Gently blot the leaves dry with a soft paper wipe and transfer the leavesto 1.5 mL centrifuge tubes containing 150 mL extraction buffer.
(3) Grind leaves using a drill fitted with a plastic pestle, keeping samples onice at all times.
(4) Centrifuge tubes at 10,000 rpm (9,000 g) for 10 min at 4 �C.(5) Transfer 25 mL of the protein extract to a 96-well microplate containing
25 mL of GUS assay buffer. Cover and incubate at 37 �C for at least anhour.
(6) After the incubation period, add 250 mL of carbonate stop buffer (0.2MNa2CO3) to each well to stop the reaction.
(7) Read the fluorescence on a microplate reader; we used a BiotekSynergy HT: excitation 365 nm, emission 460 nm. Use a 1 mM 4-MU standard solution (described above) to prepare a standard curveranging from 0 to 400 nM 4-MU.
(8) Accomplish protein normalization as described in step 9 of 7.1
ACKNOWLEDGMENTS
This work was supported by grants from Defense Advanced Research Project Agency andOffice of Naval Research.
REFERENCES
Allert, M., Rizk, S. S., Looger, L. L., and Hellinga, H. W. (2004). Computational design ofreceptors for an organophosphate surrogate of the nerve agent soman. Proc. Natl. Acad.Sci. USA 101, 7907–7912.
Antunes, M. S., Morey, K. J., Tewari-Singh, N., Bowen, T. A., Smith, J. J., Webb, C. T.,Hellinga, H. W., and Medford, J. I. (2009). Engineering key components in a syntheticeukaryotic signal transduction pathway. Mol. Syst. Biol. 5, 270.
Antunes, M. S., Morey, K. J., Smith, J. J., Albrecht, K. D., Bowen, T. A., Zdunek, J. K.,Troupe, J. F., Cuneo, M. J., Webb, C. T., Hellinga, H. W., and Medford, J. I. (2011).Programmable ligand detection in plants through a synthetic signal transduction pathway.PloS ONE 6(1):e16292.
Argueso, C. T., Ferreira, F. J., and Kieber, J. J. (2009). Environmental perception avenues:The interaction of cytokinin and environmental response pathways. Plant Cell Environ.32, 1147–1160.
Assaad, F. F., and Signer, E. R. (1990). Cauliflower mosaic-virus P35S promoter activity inEscherichia coli. Mol. Gen. Genet. 223, 517–520.

600 Kevin J. Morey et al.
Batard, Y., Hehn, A., Nedelkina, S., Schalk, M., Pallett, K., Schaller, H., and Werck-Reichhart, D. (2000). Increasing expression of P450 and P450-reductase proteins frommonocots in heterologous systems. Arch. Biochem. Biophys. 379, 161–169.
Baumberger, N., Doesseger, B., Guyot, R., Diet, A., Parsons, R. L., Clark, M. A.,Simmons,M. P., Bedinger, P., Goff, S. A.,Ringli, C., andKeller, B. (2003).Whole-genomecomparison of leucine-rich repeat extensins in Arabidopsis and rice. A conserved family ofcell wall proteins form a vegetative and a reproductive clade. Plant Physiol. 131, 1313–1326.
Castillon, A., Shen, H., and Huq, E. (2007). Phytochrome interacting factors: Centralplayers in phytochrome-mediated light signaling networks. Trends Plant Sci. 12,514–521.
Chen, D. Q., and Texada, D. E. (2006). Low-usage codons and rare codons of Escherichiacoli. Gene Ther. Mol. Biol. 10A, 1–12.
Chen, M. H., Takeda, S., Yamada, H., Ishii, Y., Yamashino, T., and Mizuno, T. (2001).Characterization of the RcsC ! YojN ! RcsB phosphorelay signaling pathwayinvolved in capsular synthesis in Escherichia coli. Biosci. Biotechnol. Biochem. 65, 2364–2367.
Christie, J. M. (2007). Phototropin blue-light receptors. Annu. Rev. Plant Biol. 58, 21–45.Davis, S. J., and Vierstra, R. D. (1996). Soluble derivatives of green fluorescent protein
(GFP) for use in Arabidopsis thaliana. Weeds World 3, 43–48.De Moraes, C. M., Schultz, J. C., Mescher, M. C., and Tumlinsoni, J. H. (2004). Induced
plant signaling and its implications for environmental sensing. J. Toxicol. Environ. Health A67, 819–834.
Dortay, H., Mehnert, N., Burkle, L., Schmulling, T., and Heyl, A. (2006). Analysis ofprotein interactions within the cytokinin-signaling pathway of Arabidopsis thaliana. FEBSJ. 273, 4631–4644.
Dwyer, M. A., and Hellinga, H. W. (2004). Periplasmic binding proteins: A versatilesuperfamily for protein engineering. Curr. Opin. Struct. Biol. 14, 495–504.
Dwyer, M. A., Looger, L. L., and Hellinga, H. W. (2003). Computational design of a Zn2þ
receptor that controls bacterial gene expression. Proc. Natl. Acad. Sci. USA 100,11255–11260.
Dwyer, M. A., Looger, L. L., and Hellinga, H. W. (2004). Computational design of abiologically active enzyme. Science 304, 1967–1971.
Gomez-Gomez, L., and Boller, T. (2000). FLS2: An LRR receptor-like kinase involved inthe perception of the bacterial elicitor flagellin in Arabidopsis. Mol. Cell 5, 1003–1011.
Heyl, A., and Schmulling, T. (2003). Cytokinin signal perception and transduction. Curr.Opin. Plant Biol. 6, 480–488.
Jaillais, Y., and Chory, J. (2010). Unraveling the paradoxes of plant hormone signalingintegration. Nat. Struct. Mol. Biol. 17, 642–645.
Jefferson, R. A., Kavanagh, T. A., and Bevan, M. W. (1987). GUS fusions: Beta-glucuroni-dase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6,3901–3907.
Kay, S., and Bonas, U. (2009). How Xanthomonas type III effectors manipulate the hostplant. Curr. Opin. Microbiol. 12, 37–43.
Khalil, A. S., and Collins, J. J. (2010). Synthetic biology: Applications come of age.Nat. Rev.Genet. 11, 367–379.
Kiel, C., Yus, E., and Serrano, L. (2010). Engineering signal transduction pathways. Cell140, 33–47.
Koretke, K. K., Lupas, A. N., Warren, P. V., Rosenberg, M., and Brown, J. R. (2000).Evolution of two-component signal transduction. Mol. Biol. Evol. 17, 1956–1970.
Lamarche, M. G., Wanner, B. L., Crepin, S., and Harel, J. (2008). The phosphate regulonand bacterial virulence: A regulatory network connecting phosphate homeostasis andpathogenesis. FEMS Microbiol. Rev. 32, 461–473.

Developing a Synthetic Signal Transduction System in Plants 601
Lessard, P. A., Kulaveerasingam, H., York, G. M., Strong, A., and Sinskey, A. J. (2002).Manipulating gene expression for the metabolic engineering of plants. Metab. Eng. 4,67–79.
Levskaya, A., Weiner, O. D., Lim, W. A., and Voigt, C. A. (2009). Spatiotemporal controlof cell signalling using a light-switchable protein interaction. Nature 461, 997–1001.
Li, X., Hirano, R., Tagami, H., and Aiba, H. (2006). Protein tagging at rare codons is causedby tmRNA action at the 3’ end of nonstop mRNA generated in response to ribosornestalling. RNA 12, 248–255.
Looger, L. L., Dwyer, M. A., Smith, J. J., and Hellinga, H.W. (2003). Computational designof receptor and sensor proteins with novel functions. Nature 423, 185–190.
Makino, K., Shinagawa, H., Amemura, M., and Nakata, A. (1986). Nucleotide sequence ofthe phoR gene, a regulatory gene for the phosphate regulon of Escherichia coli. J. Mol. Biol.192, 549–556.
Makino, K., Shinagawa, H., Amemura, M., Kawamoto, T., Yamada, M., and Nakata, A.(1989). Signal transduction in the phosphate regulon of Escherichia coli involves phospho-transfer between PhoR and PhoB proteins. J. Mol. Biol. 210, 551–559.
Mizuno, T., and Yamashino, T. (2010). Biochemical characterization of plant hormonecytokinin-receptor histidine kinases using microorganisms. Methods Enzymol. 471,335–356.
Mizuno, T., Wurtzel, E. T., and Inouye, M. (1982). Cloning of the regulatory genes(OmpR and EnvZ) for the matrix proteins of the Escherichia coli outer-membrane.J. Bacteriol. 150, 1462–1466.
Mowbray, S. L., and Sandgren, M. O. J. (1998). Chemotaxis receptors: A progress report onstructure and function. J. Struct. Biol. 124, 257–275.
Nakamura, Y., Gojobori, T., and Ikemura, T. (2000). Codon usage tabulated from interna-tional DNA sequence databases: Status for the year 2000. Nucleic Acids Res. 28, 292.
Notredame, C., Higgins, D. G., and Heringa, J. (2000). T-Coffee: A novel method for fastand accurate multiple sequence alignment. J. Mol. Biol. 302, 205–217.
Oh, M. H., Wang, X., Kota, U., Goshe, M. B., Clouse, S. D., and Huber, S. C. (2009).Tyrosine phosphorylation of the BRI1 receptor kinase emerges as a component ofbrassinosteroid signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 106, 658–663.
Osgur, S., and Sancar, A. (2006). Analysis of autophosphorylating kinase activities ofArabidopsis and human cryptochromes. Biochemistry 45, 13369–13374.
Parkinson, J. S., and Kofoid, E. C. (1992). Communication modules in bacterial signalingproteins. Annu. Rev. Genet. 26, 71–112.
Perlak, F. J., Fuchs, R. L., Dean, D. A., Mcpherson, S. L., and Fischhoff, D. A. (1991).Modification of the coding sequence enhances plant expression of insect control proteingenes. Proc. Natl. Acad. Sci. USA 88, 3324–3328.
Posas, F., WurglerMurphy, S. M., Maeda, T., Witten, E. A., Thai, T. C., and Saito, H.(1996). Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelaymechanism in the SLN1-YPD1-SSK1 “two-component” osmosensor. Cell 86,865–875.
Rodrigo, G., Montagud, A., Aparici, A., Aroca, M. C., Baguena, M., Carrera, J., Edo, C.,Fernandez-de-Cordoba, P., Ferrando, A., Fuertes, G., Gimenez, D., Mata, C., et al.(2007). Vanillin cell sensor. IET Synth. Biol. 1, 74–78.
Rodriguez, M. C., Petersen, M., and Mundy, J. (2010). Mitogen-activated protein kinasesignaling in plants. Annu. Rev. Plant Biol. 61, 621–649.
Schreier, B., Stumpp, C., Wiesner, S., and Hocker, B. (2009). Computational design ofligand binding is not a solved problem. Proc. Natl. Acad. Sci. USA 106, 18491–18496.
Seipel, K., Georgiev, O., and Schaffner, W. (1992). Different activation domains stimulatetranscription from remote (enhancer) and proximal (promoter) positions. EMBO J. 11,4961–4968.

602 Kevin J. Morey et al.
Sessa, G., Raz, V., Savaldi, S., and Fluhr, R. (1996). PK12, a plant dual-specificity proteinkinase of the LAMMER family, is regulated by the hormone ethylene. Plant Cell 8,2223–2234.
Sharrock, R. A. (2008). The phytochrome red/far-red photoreceptor superfamily. GenomeBiol. 9, 230.
Somerville, C., Bauer, S., Brininstool, G., Facette, M., Hamann, T., Milne, J., Osborne, E.,Paredez, A., Persson, S., Raab, T., Vorwerk, S., and Youngs, H. (2004). Toward asystems approach to understanding plant cell walls. Science 306, 2206–2211.
Stepanova, A. N., and Alonso, J. M. (2009). Ethylene signaling and response: Wheredifferent regulatory modules meet. Curr. Opin. Plant Biol. 12, 548–555.
Sunohara, T., Jojima, K., Tagami, H., Inada, T., and Aiba, H. (2004). Ribosome stallingduring translation elongation induces cleavage of mRNA being translated in Escherichiacoli. J. Biol. Chem. 279, 15368–15375.
Suo, G. L., Chen, B., Zhang, J. Y., Duan, Z. Y., He, Z. Q., Yao, W., Yue, C. Y., andDai, J. W. (2006). Effects of codon modification on human BMP2 gene expression intobacco plants. Plant Cell Rep. 25, 689–697.
Suzuki, T., Miwa, K., Ishikawa, K., Yamada, H., Aiba, H., and Mizuno, T. (2001). TheArabidopsis sensor His-kinase, AHK4, can respond to cytokinins. Plant Cell Physiol. 42,107–113.
Takeda, S., Fujisawa, Y., Matsubara, M., Aiba, H., and Mizuno, T. (2001). A novel featureof the multistep phosphorelay in Escherichia coli: A revised model of the RcsC ! YojN! RcsB signalling pathway implicated in capsular synthesis and swarming behaviour.Mol. Microbiol. 40, 440–450.
To, J. P. C., and Kieber, J. J. (2008). Cytokinin signaling: Two-components and more.Trends Plant Sci. 13, 85–92.
Triezenberg, S. J., Kingsbury, R. C., and Mcknight, S. L. (1988). Functional dissection ofVp16, the trans-activator of herpes-simplex virus immediate early gene-expression.GenesDev. 2, 718–729.
Vercillo, N. C., Herald, K. J., Fox, J. M., Der, B. S., and Dattelbaum, J. D. (2007). Analysisof ligand binding to a ribose biosensor using site-directed mutagenesis and fluorescencespectroscopy. Protein Sci. 16, 362–368.
Wurtzel, E. T., Chou, M. Y., and Inouye, M. (1982). Osmoregulation of gene-expression.1. DNA-sequence of the Ompr gene of the Ompb operon of Escherichia coli andcharacterization of its gene-product. J. Biol. Chem. 257, 3685–3691.
Yamada, H., Suzuki, T., Terada, K., Takei, K., Ishikawa, K., Miwa, K., Yamashino, T., andMizuno, T. (2001). The Arabidopsis AHK4 histidine kinase is a cytokinin-bindingreceptor that transduces cytokinin signals across the membrane. Plant Cell Physiol. 42,1017–1023.
Yeh, K.-C., and Lagarias, J. C. (1998). Eukaryotic phytochromes: Light-regulated serine/threonine protein kinases with histidine kinase ancestry. Proc. Natl. Acad. Sci. USA 95,13976–13981.
Yoo, S. D., Cho, Y., and Sheen, J. (2009). Emerging connections in the ethylene signalingnetwork. Trends Plant Sci. 14, 270–279.
Zhang, Y., and McCormick, S. (2009). AGCVIII kinases: At the crossroads of cellularsignaling. Trends Plant Sci. 14, 689–695.