detection of complement activation by counterimmunoelectrophoresis (cie
TRANSCRIPT

Journal oflmmunological Methods, 13 ( 1 9 7 6 ) 1 0 1 - - 1 1 2 101 © Elsev ie r ]Nor th -Hol land Biomedica l Press
D E T E C T I O N O F C O M P L E M E N T A C T I V A T I O N B Y
C O U N T E R I M M U N O E L E C T R O P H O R E S I S ( C I E ) *
C.M. A R R O Y A V E ** and E.M. 'FAN
Dit, ision o f Allergy and Immunology, Scripps Clinic and Research Foundation, La Jolla, California 92037, U.S.A.
(Received 15 March 1976, accep ted 15 April 1976)
C o u n t e r i m m u n o e l e c t r o p h o r e s i s (CIE) was used as a m e t h o d of de tec t ing ac t iva t ion of the th i rd c o m p o n e n t of the c o m p l e m e n t sys t em (C3). Highly pur i f ied C3, no rma l h u m a n se rum (NHS), EDTA- t r ea t ed plasma and se rum ac t iva ted wi th aggregated h u m a n i m m u n o - g lobul in (agg-IgG) or inul in were used as sources of C3 a n d / o r C3 split p roducts . Activa- t ion o f the a l t e rna t ive p a t h w a y of c o m p l e m e n t was assayed in the presence of EGTA (10 mM) and MgCl2 (0.3 mM), cond i t i ons which b lock ac t iva t ion of the classical pa th- way. When pur i f ied nat ive C3, fresh NHS and fresh EDTA-plasma were tes ted in CIE against e i the r an t i sera to whole C3 or to C3 spli t p roduc t s , on ly one prec ip i t in line was found , which was ident i f ied as nat ive C3. However , w h e n se rum ac t iva ted wi th agg-lgG or inul in were tes ted against the same reagents , two prec ip i t in lines were seen. The first, wi th more ca thoda l mob i l i t y was ident ical to t ha t of na t ive C3. The second line had a more anoda l mobi l i ty , was d i s t inc t ly separa ted f rom the first and c o n t a i n e d C3c and C3d as s h o w n i m m u n o c h e m i c a l l y wi th specific antisera. Native C3 and spli t p roduc t s of C3 were ident i f ied by th is CIE m e t h o d in pa t i en t s showing ev idence of ac t iva ted c o m p l e m e n t by having s u b n o r m a l to ta l c o m p l e m e n t (CH50) levels. When C3 split p roduc t s were ident i f ied , the C3c-C3d prec ip i t in line could always be d is t inguished f rom nat ive C3 by its d i f f e ren t e l e c t r o p h o r e t i c mobi l i ty , even when C3 c o n c e n t r a t i o n s in se rum varied f rom 0.25 mg/ml to 1.5 mg/ml . The sensi t iv i ty of CIE was c o m p a r e d to t h a t of CH50 by as- saying at d i f f e ren t t ime intervals a f te r agg-IgG was added to fresh NHS. C3c-C3d split p r o d u c t s were de t ec t ed by CIE before any fall in CH50 and at all t imes w h e n a s ignif icant decrease in CH50 was present . This s tudy shows t ha t the CIE t e c h n i q u e is a highly sensi- tive, specif ic and rapid m e t h o d for de tec t ing ac t iva t ion of the c o m p l e m e n t sys tem via classical or a l te rna t ive pa thways in h u m a n disease.
I N T R O D U C T I O N
E v i d e n c e h a s a c c u m u l a t e d in r e c e n t y e a r s t h a t r e d u c e d c o m p l e m e n t l e v e l s
in s e r u m is e v i d e n c e o f a n in v i v o c o n t i n u i n g i m m u n e r e a c t i o n p r o c e s s . W i t h
t h e n e w e v i d e n c e f o r a l t e r n a t i v e p a t h w a y ( A P ) a c t i v a t i o n , i t is b e c o m i n g
* This is pub l i ca t i on n u m b e r 994 f rom the Division of Allergy and I m m u n o l o g y , Scr ipps Clinic and Research F o u n d a t i o n , La Jol la , Cal i fornia 92037 . This work was s u p p o r t e d by NIH research grants AI 12594 and AM 12198 . ** Sen io r Inves t igator , The Ar th r i t i s F o u n d a t i o n .

102
clear that low complement levels may also be related to non-immune reac- tions {Osier and Sandberg, 1973).
Involvement of the complement system in human disease is generally manifested by alteration in the total hemolytic activity (Schur and Austen, 1968; Lewis et al., 1971), decrease of complement proteins {Bokisch et al., 1973; Perrin et al., 1974), deposition of proteins of the complement system in tissues (Paronetto and Koffler, 1965; Westberg et al., 1971; Rothfield et al., 1972) and/or increased catabolism of complement components (Charles- worth et al., 1974; Ziegler et al., 1974; Ruddy et al., 1971}.
Activation of complement via the classical or alternative pathways results in the sequential interaction of serum proteins and serial formation of en- zymes. During the activation, reactive sites are generated or uncovered in the component molecules which permit the activated components to interact with and activate the next component in the reaction sequence. The classical and alternative pathways link at the C3 level, and C3 activation results in fragmentation of the native molecule into C3a and C3b. Further degradation of C3b, the protein involved in the activation of the later complement com- ponents, generates C3c and C3d.
Counterimmunoelectrophoresis (CIE) has been used in the past as an effi- cient technique for detecting antibodies to particular antigens (Deborah et al., 1971; Davis and Winfield, 1974; Ferguson et al., 1974; Shackleford et al., 1974; Kurata and Tan, 1976). Since C3 and some of its split products are negatively charged and are antigenic, detection of native C3 and its products was at tempted by using CIE with anti-human C3 as the reagent. As we will describe in this paper, native C3 in serum could be differentiated from its split products, C3c and C3d, since the latter's precipitin lines in CIE had more anodal positions than native C3. This test is rapidly performed and is more sensitive than the traditional assay using both total hemolytic comple- ment activity (CII50 units/ml) for detecting complement activation.
MATERIALS AND METttODS
C3 and C3 fragments
Highly purified C3 was isolated from fresh normal human serum (NHS) by using a modification of a previously described method (Nilsson and Mfiller- Eberhard, 1965). Briefly, the steps used were: 1) euglobulin precipitation by 0.008 M ethylenedinitrilotetraacetic acid (EDTA, pH 5.4) 2) column chro- matography using fresh DE-32 cellulose and stepwise gradient elution with: a) phosphate buffer pH 8.1, 4.2 mmho/cm (starting buffer), b) starting buf- fer adjusted to 6.0 mmho/cm with 1.0 M phosphate monobasic solution, and c) starting buffer adjusted to 7.0 mmho/cm with 1.0 M phosphate mono- basic solution. 3) Pevikon block electrophoresis (Mfiller-Eberhard, 1960), and 4) chromatography of the active material on hydroxylapati te column in phosphate buffer, pll 7.9 and stepwise gradient with 8.0, 10 and 12 mmho/

103
cm buffers as described previously (Nilsson and Miiller-Eberhard, 1965). C3 was fragmented with trypsin according to a published method (Bokisch et al., 1964). C3c and C3d were isolated by a combinat ion of previously de- scribed techniques (Bokisch et al., 1969; Molenaar et al., 1973). The isola- tion of C3d was followed by analysis in immunodiffusion using anti-C3d kindly supplied by Dr. Enrique H. Vallota {Children's llospital, Cincinnati, Ohio). In addition, as sources of native C3 and activated products from C3, the following reagents were used: 1) NHS collected from 40 healthy indivi- duals, 2) EDTA-treated plasma, 3) serum activated by heat-aggregated human immunoglobulins (agg-IgG) at various concentrat ions from 5 mg/ml to 5 pg/ml and 4) serum treated with 10 mM ethylene glycol tetraacetic acid (EGTA) 0.3 mM Mg 2÷ ions (conditions which block activation of the clas- sical pathway) and a final concentrat ion of 10 mg/ml inulin (Sigma Chemical Co., St. Louis, Mo.) by incubation for 30 min at 37°C. Inulin is known to be a po ten t activator of the alternative pathway of complement .
Counterimmunoelectrophoresis (CIE)
Microscope slides of two sizes, 2.5 × 7.5 cm and 5.0 × 7.5 cm, were cleaned with alcohol and coated with 3 and 6 ml of 0.6V~ agarose, respective- ly. Veronal buffer was used at 0.03 M, pit 8.6 containing 0.04 M EDTA. Two to 8 rows of wells were cut with a 4 mm diameter punch, with each row consisting of two wells placed 5 mm apart (circumference to circumference) on each slide. Fifteen pl serum or other samples were placed in the cathodal (left) wells and the same amount of antisera in the anodal (right) wells. CIE was performed at 4°C in 0.03 M barbital buffer, pH 8.6, with a constant cur- rent of 5 ma across the slide. After a precipitin line was clearly seen in the con- trol sample, or the bromophenol blue placed in the cathodal well was in the anodal well, usually at about 60--90 min, CIE was stopped. Slides were examined for precipitin lines immediately after C1E, and the positions of the lines were measured under a magnifying lens containing a calibrated scale. The slides were re-examined 60 min after incubation at 37°C, and results were expressed as electrophoret ic mobility (Rf) of each precipitin line. Rf was calculated as follows:
Rf Distance from nearest circumference of cathodal well to the precipitin line . . . . . . . . . . . . . -D-istance between nearest circumference of wells
Immunochen2ical analysis
Double diffusion experiments were carried out in 1% agar of 1% agarose containing 0.01 M EDTA, 0.03 M veronal buffer, pII 8.6. Immunoelectro- phoresis was performed according to Scheidegger (1955), in the same agar and agarose mentioned above and in veronal buffer at pH 8.6 and ionic strength 0.05. Electrophoresis was carried out for 90--120 min at potential

1 0.1
gradient of five ma per slide and developed with monospecific antisera to C3, C3c and C3d.
ltenzoly tic complement activity
Tim method of Mayer (1964) was employed and the results were ex- pressed as C!I50 units/ml.
Antisera
Highly purified human C3 was injected into a rabbit as described by Goudie et al. {1966). For opt imum results with CIE techniques, antisera to human C3 should contain high titers of antibodies to both C3 and to split products. Since we were interested in an antiserum to C3 with broad specifi- city, trypsinized C$, was used for immunization ill order to obtain antisera with ant ibody activities against C3, C3b, C3c and C3d. One month later, the rabbit was given a second injection of the antigen intramuscularly and sub- cutaneously, and one week later blood was taken. The resulting immune se- rum was analyzed by double diffusion techniques against highly purified complement components , NHS and an euglobulin precipitate from pit 5.4, 0.008 M EDTA-treated serum which is rich in C3. This antiserum was used in all these studies unless otherwise specified. For comparison, four commer- cial 'monospecif ic ' anti-C3 were used (purchased from Mcloy, Springfield, VA; Hyland, Costa Mesa, CA; Cordis, Miami, FL. and Microbiological Ass()- ciates, Albany, CA). When either rabbit anti-trypsinized human C3 or com- mercial anti-C3 was tested in double diffusion against highly purified C3 or fresh NHS, only one precipitin line was detected. Therefore, these antisera were specific for C3 in that they did not react with other serum proteins.
Antisera to C3c and C3d were produced by lymph node injection of 2 rab- bits. Our anti-C3d antiserum reacted with C3d to give a precipitin line that showed complete identi ty with the precipitin line of anti-C3d from Dr. Val- lota. In addition, both anti-C3 and anti-C3d antisera also reacted with native C3, showing that C3d determinants on the native molecule were recognized by the antisera. No reactions were found with C5, C6, properdin, factor B or immunoglobulins (IgG, IgM, lgA). No cross-reactivity was found between anti-C3c and anti-C3d.
Subjects
The group studied included 10 patients with systemic lupus, four with chronic idiopathic urticaria and hypocomplementemia , three with rheuma- toid arthritis and cryoglobulinemia and one with midline granuloma. EDTA- plasma was obtained for complement studies and kept at --70°C. These patients were selected not because of disease categories but because their sera represented clinical examples of complement activation in vivo and con-

105
rained d i f ferent levels of C3. It was necessary to de te rmine if differences in levels of C3 would significantly change the posi t ion of percipit in lines which might cause difficulties in interpret ing the CIE. F rom 40 hea l thy individuals we col lected blood for EDTA-plasma and fresh NHS.
RESULTS
Identification of native C3 in NHS and EDTA-treated plasma
When highly purified C3, fresh NHS and EDTA-plasma f rom normal sub- jects were subjected to CIE against anti-C8 prepared in our l abora tory , a clear line of precipi ta t ion was seen in each instance as illustrated in fig. 1. The precipit in line shown by isolated C3 (in the first row) fused comple te ly with the precipit in lines developed with serum and plasma. These lines did no t disappear af ter washing with saline. Similarly, a single identical precipit in
Fig. 1. Counterimmunoelectrophoresis. Plasma (P), serum (vS), and isolated C3 (C3) were placed in the wells at the cathode and antiserum to C3 at the anode. A single precipitin line is seen with each set of wells and the lines fuse in immunological identity.

1 0 6
l ine was f o u n d w h e n f o u r d i f f e r e n t c o m m e r c i a l an t i s e r a to C3 were used.
To inves t iga te the pos s ib i l i t y t h a t d u r i n g the c o a g u l a t i o n process c o m p l e - m e n t cml be ac t i va t ed in v i t ro , fresh NI IS a n d E D T A - p l a s m a f r o m the same d o n o r s were used in pa i red s tud ies . The a n t i s e r u m used in this e x p e r i m e n t
was m a d e in r a b b i t s aga ins t t r y p s i n i z e d C3. In 5 o u t o f 40 s e r u m samples f rom n o r m a l i nd iv idua l s , a weak p r e c i p i t i n l ine was seen in a m o r e a n o d a l p o s i t i o n t h a n t h a t of na t ive C3. This p r e c i p i t i n l ine was n o t p r e s e n t in p l a s m a s amp le s f rom a n y of the 40 n o r m a l i nd iv idua l s .
lden t ifica t ion o f split products from C3
To d e m o n s t r a t e t h a t an t i -C3 iden t i f i e s C3 as well as C3 spl i t p r o d u c t s , s e r u m a(: t iwlted wi th i nu l in or agg-lgG was used (fig. 2). R o w s 1 a nd 2 con - t a i n e d s e r u m ac t iva t ed wi0~ agg-lgG a n d row 3 wi th i nu l in . The an t i s e r a used in the an()(tal wells were an t i -C3 in rows 1 a nd 3, a nd a n t i - C 3 d in r ow 2.
Rows 1, 2 and 3 had an a n o d a l p r e c i p i t i n l ine which fused c o m p l e t e l y in a siT~gle c o n t i n u o u s l ine of i m m u n o h ) g i e a l i d e n t i t y wi th the p r e c i p i t i n sysr, em
o f a n t i - C 3 d . In rows 1 and 3 the re was a s e c o n d l ine wi th m o r e c a t h o d a l m o b i l i t y . Th i s l ine a p p e a r e d in fresh NHS or in i so la ted a c t i va t e d C3 a n d cor-
- - + I I III
0 Fig. 2. Serum actiw~ted with aggregated Ig(; (IgG) or inulin (I) were placed m the eath- od~d wells and antiserum to (23 (aC3) and to C3d (aC3d) in tile anodal wells. A second preeipitin line, more anodal in position, is seen with each anti-C3 serum. These anodal lines fused with the single line produced by anti-C3d in immunological identity, illustrat- ing that anti-C3 sera were capable of detecting the C3d split products in activated sera. The more eathodal preeipitin lines observed with anti-C3 sera were due to reaction with unaltered natiw, C'3. The preeipitin lines were not reproduced well in the photot~raph and a line drawing was added to help illustrate these features.

107
responded to native C3. As seen in this figure, ant isera to C3 recognized the C3d split p r o d u c t as well as the native molecule . An addi t ional fea ture il- lustrated in the s tudy was tha t C3d had a dis t inct ly faster Rf than native C3. In test ing the commerc ia l anti-C3 antisera, those f rom Meloy, Hyland and Microbiological Associates did no t show the second (anodal) precipi t in line. Only the anti-C3 an t i se rum f rom Cordis was able to show a weak second pre- cipit in line identical to C3d system.
To s tudy the Rf o f C3 and C3 split p roduc t s in CIE, we used as sources of antigens, EDTA-plasma, sera act ivated with agg-IgG or inulin. The ant isera used were: anti-C3, anti-C3c and anti-C3d. The results can be summar ized as fol lows: 1) using fresh EDTA-plasma, on ly one precipi t in line was de tec t ed , which co r responded to the mobi l i ty of native C3. 2) When sera act ivated with agg-IgG or inulin were used, two precipi t in lines were de tec t ed , with the ca thoda l line having the same Rf as native C3. The more anodal precipi t in line was reactive with e i ther an t i se rum to C3c or to C3d. The posi t ions of the anodal precipi t in lines were identical and could no t be dist inguished f rom one a no the r on the basis of Rf. However , immunochemica l ly , they were clearly non-ident ical . With these results it was clear tha t C3c and C3d could no t be d i f fe ren t i a t ed by CIE excep t by using specific antisera to these prote ins .
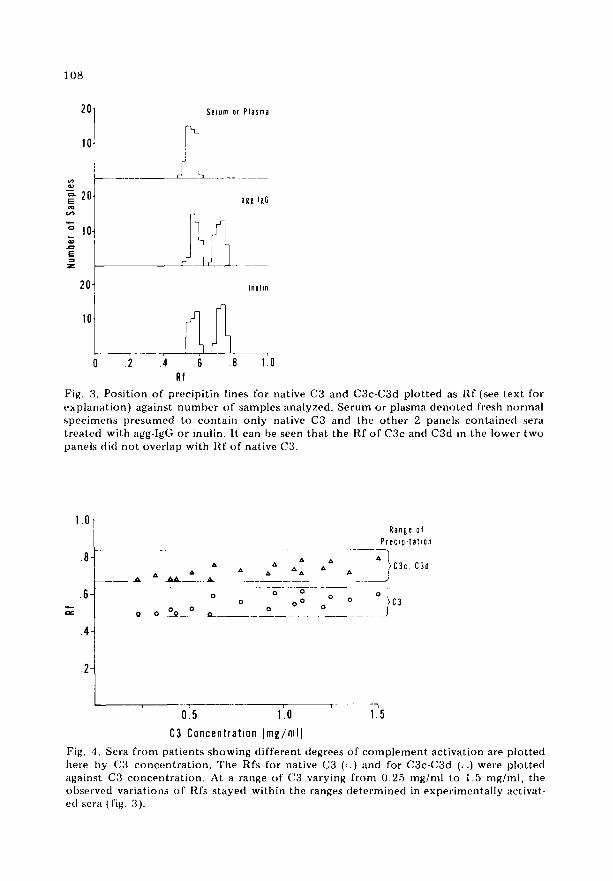
Calculation o f the R f for C3 and split products
In fig. 3 we present the Rfs calculated for each pro te in using d i f f e ren t reagents. In the upper part of the figure, the Rf for native C3 in NllS, EDTA-plasma and hepar in ized plasma are shown. The mean value was 0 .534 ± 0 .117 (2 S.D.). In the middle of the figures are the Rfs calculated for native C3 and C3c-C3d in serum act ivated with agg-IgG. The mean value for C3 was 0 .558 -+ 0 .076 (2 S.D.) and for C3c-C3d, 0 .745 + 0 .096 (2 S.D.). In the lower par t o f the figure the calculated Rfs for C3 and C3c-C3d in serum act ivated with inulin are similar to tha t o f agg-IgG. By statist ical analysis the d i f fe rence be tween the Rfs of the native C3 and C3c-C3d precipi t in systems was significant, the d i f fe rence having a S t u d e n t t test P value < 0.001.
Effect o f C3 concentration on R f
Since the concen t r a t i on o f C3 and C3 split p roduc t s in d i f fe ren t indivi- duals might vary cons iderably dur ing diseases in which c o m p l e m e n t is acti- vated, the fol lowing expe r imen t s were pe r fo rmed . Eighteen EDTA-plasma samples f rom pat ients with d i f f e ren t levels o f to ta l hemoly t i c c o m p l e m e n t act ivi ty (CH50) were analyzed by CIE, using as a reagent anti-C3. The results o f this e x p e r i m e n t are expressed as Rf of precipi t in line in re la t ion to C3 con c e n t r a t i on in radial i m m u n o d i f f u s i o n (fig. 4). There was a t e n d e n c y for Rfs to be lower at the lower concen t r a t i ons of C3 bu t all the values were within the calculated range for C3 or for C3c-C~d split p roduc t s (fig. 3) at concen t ra t ions o f C3 in serum ranging f rom 0.25 mg/ml to 1.5 mg/ml.
108
'=-20, EE
° 10~ i
E
20
Serum or P l a s m a
?
agg IgG
r' I . . . .
Inuhn
0 .2 .4 .6 .8 1~0 Rf
Fig. 3. Posi t ion of precipi t in lines for native C3 and C3c-C3d p lo t ted as I~,f (see text for exp lana t ion) against number of samples analyzed. Serum or plasma deno ted fresh normal spec imcus presumed to conta in only native C3 and the o the r 2 panels conta ined sera t rea ted with agg-lgG or inulin. It can be seen that the Rf of C3c and C3d in the lower two panels did no t overlap with Rf of native C3.
1.0-
.8-
.6-
. 4
.2-
R a n g e of
P r e c t p i t a t t o n
A A C3c. C]d A A A A A ~ A A A '
. . . . ~ A . . . . . A_ _ . . .J
0 0 0 0 0 o o o o iC3
Q 0 O0 0 ~ - _ _ . - - 0 0 J
o15 1'.o ' 1.5
C3 Concentration [mg/mlJ
Fig. 4. Sera f rom pat ients showing d i f ferent degrees of c o m p l e m e n t act ivat ion are p lo t ted here by (33 concen t ra t ion . The Rfs for native C3 (:.) and for C3c-C3d (/~) were p lo t ted against C3 concen t ra t ion . At a range of C3 varying f rom 0.25 mg/ml to 1.5 mg/ml, the observed variations of Rfs s tayed within the ranges de te rmined in exper imenta l ly activat- ed scra (fig. 3).
109
80
• ":'- 60-
o
40-
20-
CIE Plasma -
Serum -
Plasma ÷ agg IgG -
1'0 2'0 3'0 4'0 5'0 6'0 Time at 37°C [min)
+ 4 4 + + 4 4 4 4 4 4 4 +
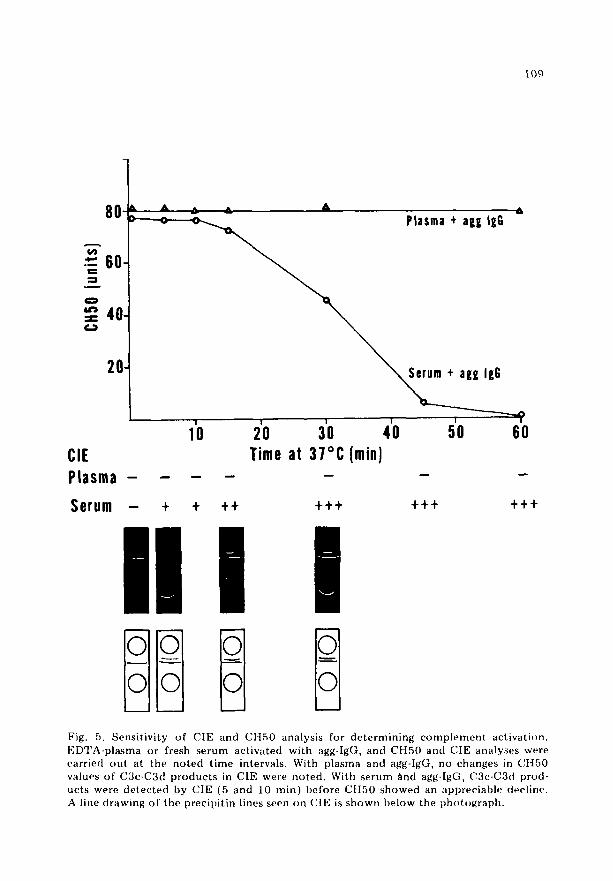
n Fig. 5. Sensit ivity of CIE and CH50 analysis for de te rmin ing c o m p l e m e n t activati()n. EDTA-pla~ma or fresh serum activated with agg-IgG, and CH50 and CIE analyses were carried ou t at the no ted t ime intervals. With plasma and agg-IgG, no changes in Ct t50 values of C3c-C3ct p roduc t s in CIE were no ted . With serum find agg-IgG, C3c-C3d prod- ucts were de tec ted by CIE (5 and 10 znin) before C[I50 showed an appreciable decline. A line drawing of the precipi t in lines seen on CIE is shown below the pht)to~raph,

l l 0
Sensitivity of CIE during in vitro actiualion of complement
In an a t t e m p t to de t e rmine the sensit ivity of the CIE t echn ique as an in- d i ca to r of c o m p l e m e n t act ivat ion, the fol lowing studies were done (fig. 5). EDTA-plasma or NItS was incuba ted with 0.5 mg/ml of agg-IgG at 37°C, and samples were wi thdrawn at d i f f e ren t t ime intervals. When EDTA-plasma was used, no change in the CII50 was observed t h r o u g h o u t the incuba t ion per iod and no anodal precipi t in line co r respond ing to C3c-C3d was seen in CIE. In cont ras t , when serum instead of plasma was act ivated with agg-IgG, a d rop in CTI50 began to be observed at 15 min with comple t e act ivat ion a round 45- -50 min. Yet, by CIE we were able to de t ec t the anodal precipi t in line which co r r e sponded to C3c-C3d within 5 min (fig. 5) at a t ime before CtI50 showed an appreciable fall. Thereaf te r , the precipi t in line became s t ronger with fu r the r falls in CH50.
DISCUSSION
We have adap ted CIE as a simple mid rapid m e t h o d o f de tec t ing act ivat ion o f the c o m p l e m e n t sys tem. The third c o m p o n e n t o f c o m p l e m e n t (C3) was selected as the test p ro te in for this s tudy because it is act ivated by e i ther the classical or al ternat ive pa thway . The fact tha t act ivat ion has occur red is judged by the presence o f split p roduc t s f rom C3, which form on ly af te r ac t ivat ion and are recognizable [)(.'cause they are more negatively charged than the native prote in .
When an t i se rum to C3 of a broad specif ic i ty (reactive with native C3 and C3 split p roduc t s ) was used as the reagent against fresh Nt tS or EDTA- plasma, on ly one precipi t in line was de tec ted . Against serum t rea ted with agg-IgG or inulin, at lea~st two precipi t in lines were de tec ted . By fu r the r analysis, it was shown tha t the more ca thoda l precipi t in line represented native C3 and the more anodal line actual ly consisted o f two lines very close to each o the r which were ident if ied as C3c and C3d. Some of the commer- cially available ant isera against C3 (Meloy, Hyland and Microbiological Asso- ciates) did no t show the anodal precipi t in lines. This may be due to tech- niques used to p roduce ant ibodies on ly against native C3 and, secondar i ly , absorp t ion with reagents conta in ing act ivated or dena tu red C3, to make the antisera immunospec i f i c for the nat iw ~. prote in .
Analyses were made of many sera f rom pat ients with d i f f e ren t diseases, who had d i f fe ren t levels of total c o m p l e m e n t activity. It was clear that , in spite of varying levels of CH50, native C3 could be d i f fe ren t ia ted f rom C3e- C3d split p roduc t s by d i f f e ren t posi t ions of the precipi t in lines. In the course of these studies, the ques t ion arose as to the disposi t ion o f C3b, f rom which C3c and C3d are derived. It was obseiwed tha t a f te r CIE, if the slides were examined af te r pro longed incubat ion , the ca thoda l precipi t in line was some- t imes seen to split in to two lines very close to each o ther , much like the C3c- C3d anodal lines. These lines might have represented two systems, native (;3

11l
and C3b, since studies have shown that they have almost the same electro- phoretic mobili ty (Bokisch et al., 1969). However, we did not carry out further studies, and this can only be considered a possibility. At no time did we observe a precipitin system which might have been considered to repre- sent C3a, either because this split product is rapidly degraded or because it is a basic or positively charged molecule which does not lend itself to detect ion by CIE.
The presence of split products in NItS also was investigated. In a small number of fresh serum samples from healthy individuals (5 of 40 samples), we were able to detect the presence of C3c-C3d precipitin systems. The ob- servation of a fast moving componen t in some NHS was previously reported by another group (Laurell and Lundh, 1967). It is well established that many complement components may be activated by extrinsic or non-complement enzymes such as trypsin, plasmin, lysosomal enzymes and bacterial enzymes (Bokisch et al., 1969; Arroyave and Miiller-Eberhard, 1973). The presence of C3 split products in NHS could be from activation of the complement sys- tem during coagulation, since paired EDTA-plasma run at the same time under the same conditions did not show such anodal precipitin lines. To prevent such complement activation and false positive results, one should ob- tain specimens in the presence of EDTA and complete the electrophoret ic analysis (CIE) as quickly as possible to keep in vitro conversion at a mini- mum.
The detect ion of breakdown products of C3 by using CIE provides an al- ternative to complement consumption or activation tests. Split products of C3 have been found by different research groups in sera of patients with sys- temic lupus erythcmatosus, acute glomerulonephrit is and other glomerulo- nephritides (Morse et al., 1962; Scoothill, 1967; West et al., 1967; t lunder et al., 1974; Perrin et al., 1975). In addition, C3 breakdown products were report- ed in the synovial fluid from patients with rheumatoid arthritis, chronic hepatitis or primary biliary cirrhosis (Zvaifler, 1969; Teisberg and Gijone, 1973). Split products from C3 can be detected by other previously reported techniques. One is the Laurell (1967) technique which requires two different e lectrophoret ic steps with the inclusion of the specific antiserum in the dif- fusion medium in the second step. Another procedure is radial immuno- diffusion (Mancini) using monospecific antisera to C3c or C3d. The CIF technique offers certain practical advantages. Larger numbers of specimens can be tested with standard electrophoresis equipment , the amount of reagent required is small, interpretat ion of the results is quick and unequi- vocal and non-specific reactions are not encountered. The specificity and sensitivity of the assay used in this study were demonstra ted in experiments based on the in vitro generation of C3 split products during activation of normal human serum by agg-IgG and by the study of patients in whom the C3 levels were reduced by at least fiw~ times from the normal concentra t ion of C3 in NHS.

112
A(gKNOWLEDGEMENT
We w o u l d l ike to t h a n k Ms. L. Miles for her t e c h n i c a l ass i s tance .
REFEI(ENCES
Arroyave, C.M. and tl.J. Mfiller-Eberhard, 1973, J. Immunol. 111, 536. Bokisch, V.A., H.J. Mtiller-Eberhard and C.G. Cochrane, 1969, J. Exp. Med. 129, 1109. Bokisch, V.A., F.tl. Top, P.K. Russell, F.J. Dixon and H.J. M[iller-Eberharcl, 1973, New
Engl. J. Med. 289, 996. Charlesworth, J.A., D.G. Williams, E. Sherington, P.J. Lachmann and D.K. Peters, 1974,
J. Clin. Invest. 53, 1578. Davis, J.S. and J.B. Winfield, 1974, Clin. Immunol. Immunopathol. 2, 510. Deborah, A.M., P.F. Kohler and J.W. Singleton, 1971, Allergy Clin. Immunol. 47,315. Ferguson, A.C., H. Kennedy, B.J. Ank, P. Sturgeon and E.R. Stiehns, 1974, A.J.C.P. 62,
861. (k)atne, R.B., C.ll. IIome and P.C. Wilkinson, 1966, Lancet ii, 122,1. Hunder, G.G., B.J. Mullen and F.C. McDuffic, 1974, Ann. Int. Med. 80, ,153. Kurata, N. and E.M. Tan, 1976, Arthritis Rheum. 19, 37. I_,aurell, C.B. and B. Lundh, 1967, Immunol. 12, 313. t, ewis, E.J., C.B. Carpenter anct P.H. Schur, 1971, Ann. hat. Med. 75, 55. Mayer, M.D., 1961, In: Experimental immunochemistry (Charles Thomas, Springfiel(t, Il-
linois USA) 2ncl Ed. p. 133. Molenaar, J.L., A. ten Velde and K.W. Pondman, 1973, J. Immunol. 110, 702. Morse, J.H., H.J. Miiller-Eberhard and H.G. Kunkel, 1962, Bull. N.Y. Acad. Med. 38,641. MiJller-Eberhard, H.J., 1960, Scand. J. Clin. Lab. Invest. 12, 33. Nilsson, U.R. and ft.J. MiJller-Eberhard, 1965, J. Exp. Med. 122, 277. Osier, A. and A.L. Sandberg, 1973, Progr. Allergy. 17, 51. Paronetlo, F. and D. Koffler, 1965, d. Clin. Invest. 44, 1657. Perrin, L.II., P./I. Lambert and P.A. Miescher, 1974, Clin. Exp. Immunol. 16, 575. Perrin, L.II., P.H. Lambert and P.A. Miescher, 1975, J. Clin. Invest. 56, 165. t{othfield, N., A. Ross, J.C. Minta and I.tt. Lepow, 1972, New EngI. J. Med. 287,681. Ruddy, S., C.B. Carpenter, K.W. Chin, J.N. Knostman, N.A. Soter, O. G6tze, ll.J. MiJllcr-
Eherhard and K.F. Austen, 1975, Medicine 54, 165. Schei(legger, J.J., 1955, Int. Arch. Allergy Appl. Immunol. 7, 103. Schur, P.H. and K.F. Austen, 1968, Ann. Rev. Med. 19, 1. Shackelford, P.G., J. Campbell and R.D. Feigin, 1974, J. Pediat. 85,478. Soothill, J.F., 1967, Clin. Exp. Immunol. 2, 83. Teisberg, P. and E. Gijone, 1973, Clin. Exp. Immunol. 14,509. West, C.D., S. Winter, J. Forristal, J.M. McConville and N.C. Davis, 1967, J. Clin. Invest.
.16, 539. Westherg, N.G., G.B. Naff, J.R. Boyer and A.F. Michael, 1971, J. Clin. Invest. 50,642. Ziegler, J.B., F.S. Rosen, W.E. Grupe, I.H. Lepow and C.A. AIper, 1974, J. Clin. Invest.
56,671. Zvaifler, N.J., 1969, J. Clin. Invest..18, 1532.