cytochrome p4501a is induced in endothelial cell lines from the kidney and lung of the bottlenose...
TRANSCRIPT

Aquatic Toxicology 76 (2006) 295–305
Cytochrome P4501A is induced in endothelial cell lines from thekidney and lung of the bottlenose dolphin,Tursiops truncatus
Rita Anne Garricka,b,∗, Bruce R. Woodinb, Joanna Y. Wilsonb,Bobby L. Middlebrooksc, John J. Stegemanb
a Department of Natural Sciences, Fordham University, 113 W. 60th Street, New York, NY 10023, USAb Department of Biology, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USA
c Department of Biological Sciences, University of Southern Mississippi, Hattiesburg, MS 39406-0001, USA
Received 3 August 2005; received in revised form 14 October 2005; accepted 15 October 2005
Abstract
Marine mammals respond to the presence of polycyclic and planar halogenated aromatic hydrocarbons (PAH or PHAH)with the induced expression in endothelium of cytochrome P4501A1, regulated through the aryl hydrocarbon receptor (AHR)transcription factor. Physiological responses in other animals, such as edema and inflammation indicate that the endotheliummay be compromised by exposure to AHR agonists, which are ubiquitous in the marine environment. In other mammalsand fish the cellular and molecular consequences of exposure to AHR agonists have been elucidated in cultured endothelial
ial cells
ey
idneynismsin this
-
cells. We have cultured and characterized cetacean endothelial cells (EC) and used them in induction studies. Endothelwere cultured from the lung and kidney of the bottlenose dolphin,Tursiops truncates, and exposed to the AHR agonists�-naphthoflavone (�NF) and 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). �NF (1–3�M) induced significant increases inCYP1A1 (O-deethylation of 7-ethoxyresorufin to resorufin; EROD) activity to 3.6 and 0.92 pmol/mg/min in lung and kidnEC, respectively. TCDD was more potent than�NF, and more efficacious, with maximum induction of CYP1A1 activity of 10.1and 15.2 pmol/mg/min in lung and kidney EC at 3–10 nM TCDD. The differential response indicates that the lung and kendothelial cells in culture retain the ability to respond in a selective manner to specific stimuli. Both the molecular mechaof induction and the physiological consequences, especially in the vasculature, of toxicant exposure can be studiedsystem.© 2005 Elsevier B.V. All rights reserved.
Keywords: Endothelium; CYP1A; EROD; Dioxin; Cetacean; Microvascular
∗ Corresponding author. Tel.: +1 212 636 6333/+1 508 289 2368;fax: +1 212 636 7213/+1 508 457 2134.
E-mail address: [email protected] (R.A. Garrick).
1. Introduction
Marine organisms are continuously exposed topolycyclic and planar halogenated aromatic hydrocarbons (PAH and PHAH) in their environment. The
0166-445X/$ – see front matter © 2005 Elsevier B.V. All rights reserved.doi:10.1016/j.aquatox.2005.10.005

296 R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305
toxicological consequences of experimental expo-sure to these toxicants in fish and mammals includedevelopmental abnormalities (Guiney et al., 1997),mutagenesis (Shimada and Fujii-Kuriyama, 2004),inflammation (Hennig et al., 2002b), and edema(Teraoka et al., 2003). In marine mammals hightissue levels of PHAH, includingortho-substitutedpolychlorinated biphenyl (PCB) congeners and otherorganochlorines, have been correlated with compro-mised immune systems (Lahvis et al., 1995), repro-ductive abnormalities (Reijnders, 1986), an unusuallyhigh incidence of tumors (Martineau et al., 2002) andepizootic outbreaks (Hall et al., 1992; Aguilar andBorell, 1994). The strongest molecular effect of expo-sure to PAHs or PHAHs in vertebrates is the induc-tion of enzymes of the cytochrome P4501A (CYP1A)family, through the aryl hydrocarbon receptor (AHR)transcription factor (Hahn, 1998; Mimura and Fujii-Kuriyama, 2003). The level of CYP1A1 inductionin laboratory mammals and fish has been shown todepend upon the concentration of the AHR agonistused as an inducer in whole animals and in cells inculture, including in endothelial cells (e.g.Stegemanet al., 1995; Annas et al., 2000b; Garrick et al.,2005).
Toxicant accumulation (Marsili et al., 2001) andthe expression of CYP1A1 (Angell et al., 2004) havebeen documented in cetaceans, primarily in the biopsysamples from field studies; integumentary CYP1A1expression is primarily in endothelial cells (Angelle avee e andC notp eent gi-c ed inc ple,ipe ub-b s( oft l.,2 eenc el-u 1G c-t e
in skin biopsy slices from sperm whale,Physetermacrocephalus, was dose dependent in endothelialtissue.
The endothelium in many vertebrate tissues is rich inAHR expression and responds to xenobiotic exposurewith the induction of CYP1A1 enzymes and physi-ological responses, such as edema and inflammation(Bayou-Denizot et al., 2000; Hennig et al., 2002b),which indicates that the endothelium may be com-promised in exposed animals. Cultures of mammalianand teleost endothelial cells, which show inductionof CYP1A1 by pHAH and PAH, are being used tostudy the mechanisms of the cellular response and thesensitivity of the endothelium in various species tovarious xenobiotics (Farin et al., 1994; Stegeman etal., 1995; Celander et al., 1997; Annas and Brittebo,1998; Bayou-Denizot et al., 2000; Annas et al., 2000b;Hennig et al., 2002a; Garrick et al., 2005). Chat etal. (1998)used immortalized and freshly isolated ratbrain microvessel endothelial cells to demonstrate thatthere is no loss in functional activity in CYP1A1 andrelated enzymes in these endothelial cells even after62 passages.Thum et al. (2000)report that in rataortic endothelial cell cultures loss of induction ofCYP1A1 in response to Aloclor 1254 was only seenin cells which no longer tested positive as endothelialcells.
In cultures of cetacean (dolphin) kidney epithelialcells PAH exposure caused DNA damage and inhibitedcell growth and exposure to fuel oil caused apopto-s afterh eta os eant onsest tiveg entd itsp als( d eta
romt nticbe 1e s inr2
t al., 2004). Controlled exposure studies that hstablished a causal relation between toxicant dosYP1A1 level in laboratory mammals and fish areossible with cetaceans. Thus, the relation betw
oxicant levels, AHR activation and the physioloal consequences of exposure have been inferretaceans from other types of studies. For examn liver microsomes from two cetacean species (Del-hinapterus leucas andGlobicephala melas) CYP1A1xpression level is correlated with the levels in bler of non-ortho and mono-ortho PCB congenerAHR agonists) and with the rate of metabolismhese congeners (White et al., 1994; White et a000). The presence of the AHR receptor has bonfirmed by cloning of the receptor from the bga,Delphinapterus leucas (Jensen and Hahn, 200).odard et al. (2004)demonstrated that in vitro indu
ion of CYP1A1 expression by�-naphthoflavon
is and altered transport and enzyme activitiesigh exposure levels (Carvan et al., 1995; Pfeifferl., 2000; Sweat et al., 2001). However, there are ntudies with endothelial cells cultured from cetacissues which describe the dose dependent respo toxicant exposure. This is an important objeciven the biological roles of endothelium, the frequetection of CYP1A1 in endothelium in biopsies andotential as a marker of exposure in marine mammBen-David et al., 2001; Angell et al., 2004; Godarl., 2004).
We have developed endothelial cell cultures fissue samples from the kidney and lung of an Atlaottlenose dolphin,Tursiops truncatus (Middlebrookst al., 1999). We report here the induction of CYP1Axpression and activity in these endothelial cellesponse to exposure to�-napthoflavone (�NF) and,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD).

R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305 297
2. Materials and methods
2.1. Chemicals
2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD)(>98% pure) was obtained from Ultra Scientific,Hope, RI, USA. 7-Ethoxyresorufin was purchasedfrom Molecular Probes (Eugene, OR). Dimethylsul-foxide (DMSO),�-napthoflavone (�NF) and all otherreagents were the highest grade available from Sigma(St. Louis, MO). Cell culture materials from Sigmawere cell culture tested. Other sources were notedbelow.
2.2. Culture of kidney and lung cells
Dolphin cell lines were developed from kidney andlung biopsies taken from a stillborn Atlantic bottlenosedolphin calf at Marine Life Aquarium, Gulfport, MIand transported in physiological saline to the lab at Uni-versity of Southern Mississippi (Middlebrooks et al.,1999). The parenchymal tissue samples, taken within24 h of the delivery, were trypsinized for 5 h and cul-tured in Liebovitz-15 (L-15) complete medium with10% fetal bovine serum (Hyclone) in tightly cappedflasks (Middlebrooks et al., 1999). Cell lines TuTruK(kidney) and TuTruL (lung) were established fromthese cells.
Cells from passages 19 for kidney (TuTruK) and 18for lung (TuTruL) cell lines were selected as the startingc d forh2 edt oughn urer d onc thatf 99w S,21 i-ct or-t ndi ke
akep sity
lipoprotein (DiI-Ac-LDL, 1,1′-dioctadecyl-1-2,3,3′,3′-tetramethyl-indocarbocyanide perchlorate acetylatedLDL, Biomedical Technologies, Stoughton, MA)(Garrick et al., 2005). The kidney cells were identifiedas homogeneous endothelial cells with Dil-Ac-LDLuptake when viewed on a Zeiss Axiovert S100 flu-orescence microscope. We tested the lung cells withDil-Ac-LDL and found that the smaller cells took upAc-LDL and the larger did not. We subcultured the lungcells with differential plating times and by removing thelarger cells we were able to obtain a homogeneous cellculture in one passage based on Dil-Ac-LDL uptake.Cells grown from these homogeneous cultures were thelung cells used in all subsequent studies. The cells usedin this study were at passages 19–21.
At confluence the cells were subcultured or aliquotscryopreserved in the vapor phase of liquid nitrogen bystandard procedures.
2.3. Treatment of cells
Cells were seeded into 48-well plates (Costar, Cam-bridge, MA) at a density of∼2× 105 cells/well andgrown to confluence in 0.5 ml of M199SE with 10%CCS and incubated at 37◦C in CO2. Subsequent pro-cedures were as described inStegeman et al. (1995)forintact cells. At confluence the medium was removedand the cells were rinsed with serum-free M199SEand 0.5 ml of M199SE with 1% CCS was added toeach well. DMSO, or serial dilutions of TCDD or� in2 atD eptf ew mina5a ure.T anda andi
ara-tt SE( S,sf anda
ultures for the current studies and were evaluateeterogeneity by phase contrast microscopy (Garrick,000; Garrick et al., 2005). The kidney cells appear
o be homogeneous in size and appearance, althot a pure (cloned) culture. The lung cell cultevealed what appeared to be two cell types, baseell size. Culture conditions were changed to thoseavored endothelial cell growth: M199SE [Medium 1ith Earle’s salts and 20 mM NaCl, 20 mM HEPE6 mM NaHCO3, 0.68 mM glutamine, 10�M pyruvate× antibiotic–antimycotic solution (100 U/ml penillin, 100�M/ml streptomycin, 2.5�M/ml ampho-ericin B)] with 10% serum (Cosmic Calf Serum, fified (CCS), Hyclone, Logan, UT) in T-75 flasks ancubated at 37◦C in 5% CO2 (Garrick, 2000; Garrict al., 2005).
Endothelial cells can be identified by the uptattern of fluorescently labeled acetylated low-den
NF dissolved in DMSO, was added to the wells.5�l aliquots from dilutions of stock solutions so thMSO was present at 0.5% (v/v) in all wells exc
or untreated cells (Stegeman et al., 1995). Each dosas added to six wells, the plate was shaken for 1nd the cultures returned to the incubator at 37◦C in% CO2 for 48 h. InStegeman et al. (1995), both�NFnd TCDD produced induction after a 48 h exposhe wells were examined microscopically beforefter treatment to confirm the cells were confluent
ntact.Cells that were used for the microsomal prep
ions were grown in T-75 flasks, exposed to 25�l ofhe appropriate dosing solution in 5 ml of M199to maintain the 0.5% (v/v) of DMSO) with 1% CChaken for 1 min and incubated at 37◦C in 5% CO2or 48 h. The cells were confluent and intact beforefter the treatment.

298 R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305
2.4. Preparation of microsomes from endothelialcells
Microsomal fractions were prepared from endothe-lial cells treated with chemicals in T-75 flasks. After a48 h exposure, the dosing medium was removed andthe flask was rinsed with serum-free M199SE. Onemilliliter of PBS–HEPES pH 7.5 with 2 mM EDTAwas added to the flask. The flask was floated on liq-uid nitrogen to snap freeze the cultures and stored at−80◦C until used. Frozen cells were scraped from theflask into buffer, homogenized, sonicated and subcel-lular fractions obtained by differential centrifugationas described inStegeman et al. (1995). Pellets werere-suspended in buffer (50 mM Tris, pH 7.4, 1 mMdithiothreitol, 1 mM EDTA, 20% glycerol) and frozenin liquid N2 until used. Protein content was determinedusing the bicinchoninic acid method, with serum albu-min as a standard (BCA® Protein Assay Kit, Pierce,Rockford, IL).
2.5. Assays
2.5.1. EROD activityThe O-deethylation of 7-ethoxyresorufin to
resorufin (EROD) a kinetic measure of CYP1A1activity, was determined fluorometrically in intactcells with a Cytofluor 4000 (Millipore) multiwellplate reader, as described for porcine endothelialcells (Stegeman et al., 1995). Incubation proceduresw ta al.,2 mw l ofp pH7K
di-t n( ni-t vesw ine otalp itht fort teda asd an
et al., 1995; Hahn et al., 1996; Ganassin et al.,2000).
Significance of the change in the level of ERODactivity in cells was tested with the Kruskal–Wallis testwith Dunn’s as a post-test for non-parametric ANOVAwith GraphPad InStat.
2.5.2. NADPH-dependent cytochrome c reductaseactivity
NADPH-dependent cytochromec reductase wasmeasured in microsomal preparations from both treatedand untreated cells by the reduction of horse heartcytochromec at 37◦C by the method described pre-viously (Stegeman et al., 1982; Stegeman et al.,1995). Cuvettes contained 15–20�g of protein ina total volume of 500�l. Activity calculated fromthe change in absorbance at 550 nm is expressed asnmol/mg/min.
2.5.3. Immunoblotting for CYP1A1The presence of CYP1A1 was detected by
immunoblot analysis of microsomal preparations asdescribed previously, with slight modification (Hahnet al., 1993; Stegeman et al., 1995). Microsomal pro-tein (30–70�g) and a range of standards of CYP1Afrom scup (Stenotomus chrysops) were resolved ona 4–20% acrylamide gradient gel in TRIS–glycinewith SDS (Jule Inc., Milford, CT). Proteins elec-
hern-1Aedrec-alsa-1A
al.,
alfedism. Then
ere modified as described earlier (Hestermann el., 2000a,b; Hestermann et al., 2002; Garrick et005). At the end of the dosing time the mediuas removed and the cells washed with 0.5 mhosphate-buffered saline (PBS) with HEPES.5 (0.136 M NaCl, 8.1 mM Na2HPO4, 1.47 mM2HPO4, 2.68 mM KCl, 10 mM HEPES).The EROD reaction was initiated by the ad
ion of 200�l/well of a solution of 7-ethoxyresorufi2.67�M in PBS-HEPES at pH 7.8) and moored kinetically for 20 min. Resorufin standard curere prepared in PBS-HEPES pH 7.8 in wellsach plate. At completion of the EROD assay trotein content was measured in each well w
he fluorescamine assay with BSA solutionshe standard curve. EROD activity was calculas pmol of resorufin produced/mg protein/minescribed previously (Kennedy et al., 1993; Stegem
trophoretically transferred to nitrocellulose (Schleic& Schuell, Keene, NH) were incubated with mooclonal antibody Mab 1-12-3 against scup CYP(Park et al., 1986) and a secondary antibody linkto horseradish peroxidase. Mab 1-12-3 stronglyognizes CYP1A1 but not CYP1A2 in mamm(Drahushuk et al., 1999). Detection and quantifiction of the bands relative to those of scup CYPwere as before (Stegeman et al., 1995; Garrick et2005).
2.5.4. EC50 calculationEC50, the inducer concentration producing h
maximal induction of EROD activity, was calculatusing the curve fitting routine of GraphPad Prfor nonlinear regression sigmoidal dose–responsemodified Gaussian equation for EC50 was described i(Stegeman et al., 1995).

R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305 299
3. Results
3.1. Cell preparations
Fig. 1 shows cultures of the cells from the kidney(A and B) and the lung (C and D) of the bottlenosedolphin. The morphology of the two preparations inthe phase contrast micrographs in A and C is similarand cobblestone like, typical of endothelial cells.The appearance of the kidney cells was the sameas in the cultures when first received. In contrast tothe kidney cells, the lung cell cultures were some-what heterogeneous when first received. However,non-endothelial cells lifted off more rapidly thanendothelial cells and by using differential platingtimes we were able to obtain a population of EC fromthe original lung cell culture. In subsequent studieswe used only cells passed from the lung endothelialcell cultures that were homogeneous. The uptake of
Dil-Ac-LDL by the kidney and lung cells, seen inthe fluorescent micrographs inFig. 1B and D, washomogeneous and confirmed that these are endothelialcells; however, the vessel type of origin is not clear.The most likely source of the endothelial cells fromboth the kidney and lung parenchymal tissues was themicrovasculature although there is no specific marker,at present, to confirm that cells are microvascular inorigin.
3.2. Induction of CYP1A1 activity
The kidney and lung cells were grown in 48-wellplates and treated for 48 h with DMSO, TCDD or�NF.The incubation medium contained 1% serum to mini-mize loss of inducer to serum proteins (Hestermann etal., 2000a,b). At the end of the 48 h the EROD activity(CYP1A1 activity) and total protein were measured ineach well as described in Section2.
F ndothep icrogr iss AxiovertS s chara
ig. 1. Micrographs of kidney (A and B) and lung (C and D) ehase contrast micrographs and (B) and (D) are fluorescence m100 at 10×. The uptake pattern for Dil-Ac-LDL in (B) and (D) i
lial cells of the bottlenose dolphinTursiops truncatus. (A) and (C) areaphs showing the uptake of DiI-acetylated-LDL viewed on a Zecteristic of endothelial cells.
300 R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305
Table 1Production of resorufin from 7-ethoxyresorufin in lung and kidney endothelial cells
�NF (�M) DMSO pmol/mg/min
0.01 0.1 1 3 10
Lung cellsAverage 0.90 1.55 1.57 1.20 3.63* 2.47*
±S.E. 0.28 0.25 0.40 0.09 0.37 0.26
Kidney cellsAverage 0.20 0.44 0.29 0.92* 0.33 0.47±S.E. 0.06 0.19 0.19 0.23 0.26 0.52
TCDD (nM) DMSO pmol/mg/min
0.01 0.1 0.3 1 3 10 15 30
Lung cellsAverage 0.30 1.60 6.27* 7.03* 10.12* 6.09* 3.69±S.E. 0.05 0.25 0.75 0.97 1.27 0.52 0.50
Kidney cellsAverage 0.44 0.69 1.61 5.21 6.29* 15.20* 10.22* 7.86* 6.98*
±S.E. 0.05 0.49 0.78 0.51 0.78 1.32 1.73 1.79 0.78
Cells are induced in 48-well plates for 48 h. Values are mean± S.E.;n is a minimum of six wells, a maximum of 18.* Significant difference among column means atp ≤ 0.002 with the Kruskal–Wallis test with Dunn’s as a post-test for non-parametric ANOVA
calculated with GraphPad InStat.
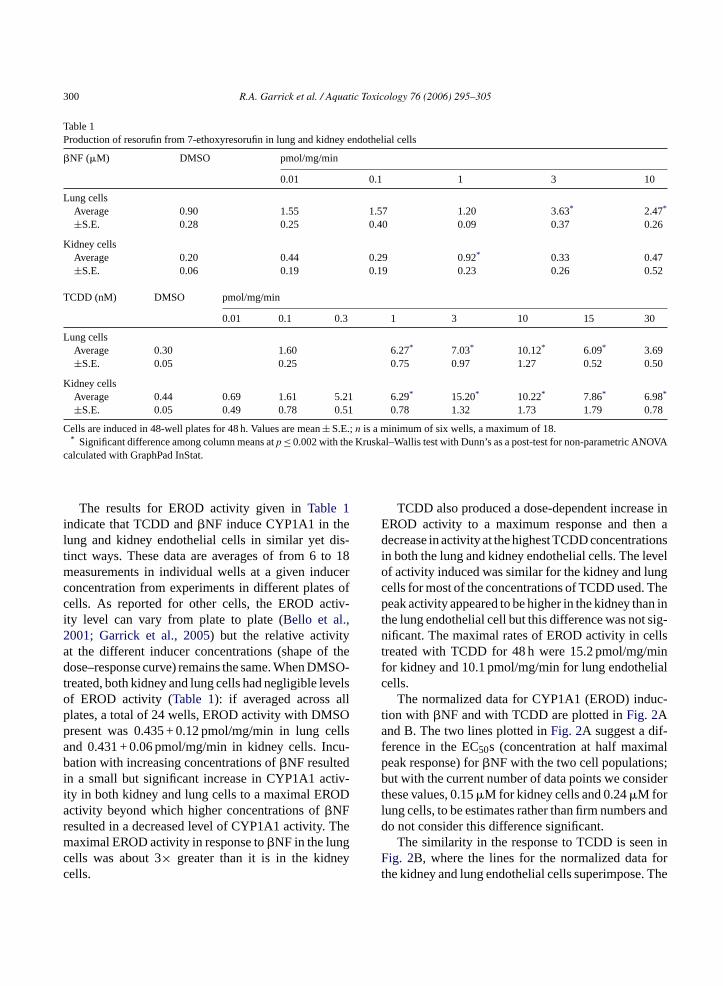
The results for EROD activity given inTable 1indicate that TCDD and�NF induce CYP1A1 in thelung and kidney endothelial cells in similar yet dis-tinct ways. These data are averages of from 6 to 18measurements in individual wells at a given inducerconcentration from experiments in different plates ofcells. As reported for other cells, the EROD activ-ity level can vary from plate to plate (Bello et al.,2001; Garrick et al., 2005) but the relative activityat the different inducer concentrations (shape of thedose–response curve) remains the same. When DMSO-treated, both kidney and lung cells had negligible levelsof EROD activity (Table 1): if averaged across allplates, a total of 24 wells, EROD activity with DMSOpresent was 0.435 + 0.12 pmol/mg/min in lung cellsand 0.431 + 0.06 pmol/mg/min in kidney cells. Incu-bation with increasing concentrations of�NF resultedin a small but significant increase in CYP1A1 activ-ity in both kidney and lung cells to a maximal ERODactivity beyond which higher concentrations of�NFresulted in a decreased level of CYP1A1 activity. Themaximal EROD activity in response to�NF in the lungcells was about 3× greater than it is in the kidneycells.
TCDD also produced a dose-dependent increase inEROD activity to a maximum response and then adecrease in activity at the highest TCDD concentrationsin both the lung and kidney endothelial cells. The levelof activity induced was similar for the kidney and lungcells for most of the concentrations of TCDD used. Thepeak activity appeared to be higher in the kidney than inthe lung endothelial cell but this difference was not sig-nificant. The maximal rates of EROD activity in cellstreated with TCDD for 48 h were 15.2 pmol/mg/minfor kidney and 10.1 pmol/mg/min for lung endothelialcells.
The normalized data for CYP1A1 (EROD) induc-tion with �NF and with TCDD are plotted inFig. 2Aand B. The two lines plotted inFig. 2A suggest a dif-ference in the EC50s (concentration at half maximalpeak response) for�NF with the two cell populations;but with the current number of data points we considerthese values, 0.15�M for kidney cells and 0.24�M forlung cells, to be estimates rather than firm numbers anddo not consider this difference significant.
The similarity in the response to TCDD is seen inFig. 2B, where the lines for the normalized data forthe kidney and lung endothelial cells superimpose. The
R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305 301
Fig. 2. Normalized plots of EROD activity after�NF and TCDDexposure. Normalized plots (percent of maximal response) of theEROD activity (mean± S.E.) fromTable 1(with log transformationof inducer concentration) are given for�NF (A) and TCDD (B) forthe lung and kidney endothelial cells. Normalized plots of the dataare used to calculate EC50. EC50 calculated for�NF is 0.15 and0.24�M for kidney and lung cells, respectively, and EC50 for TCDDis 0.019 and 0.018 nM for kidney and lung cells.
EC50 for CYP1A1 induction by TCDD in the kid-ney cells is 0.019 nM TCDD and in the lung cells is0.018 nM TCDD. Thus, TCDD was more potent than�NF with EC50 in nM versus�M range, respectively(Fig. 2A and B), and more efficacious (Hestermannet al., 2000a,b), with maximum induction of CYP1A1activity 3–10 times higher in lung and kidney endothe-lial cells (Table 1).
3.3. NADPH-dependent cytochrome c reductaseactivity
The levels of NADPH cytochromec (P450) reduc-tase activity measured in microsomal preparations fromtreated and untreated kidney and lung endothelial cellswere similar, 30 nmol/mg/min in each. These rates aresimilar to the 40 nmol/mg/min reported in porcine aor-tic endothelial cells (Stegeman et al., 1995) and shouldbe similarly effective in supporting CYP1A1 activity.
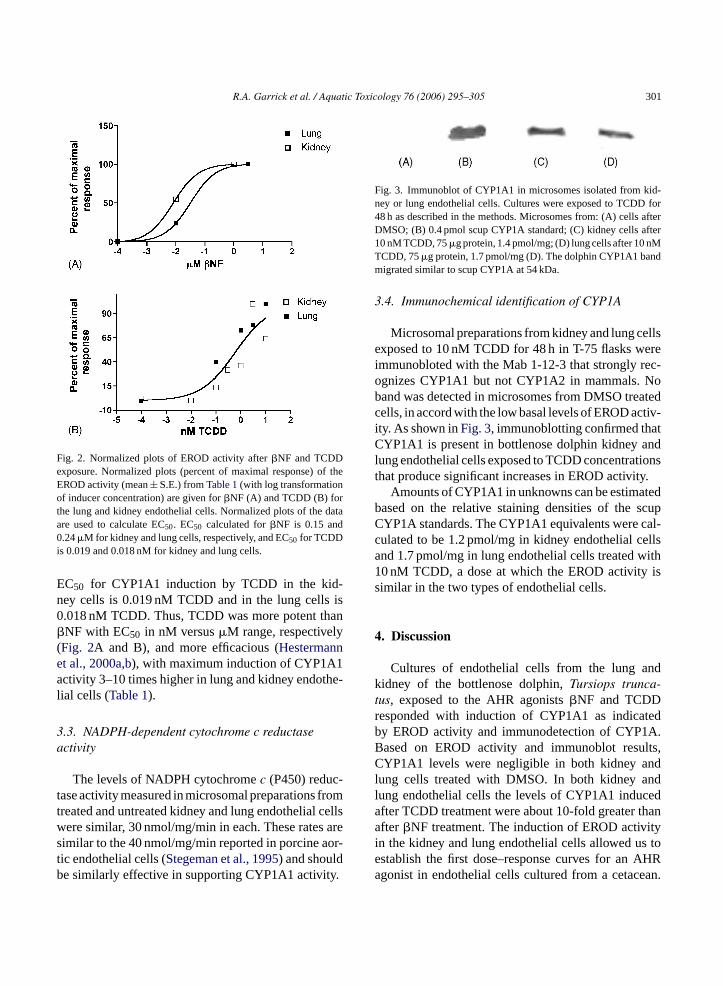
Fig. 3. Immunoblot of CYP1A1 in microsomes isolated from kid-ney or lung endothelial cells. Cultures were exposed to TCDD for48 h as described in the methods. Microsomes from: (A) cells afterDMSO; (B) 0.4 pmol scup CYP1A standard; (C) kidney cells after10 nM TCDD, 75�g protein, 1.4 pmol/mg; (D) lung cells after 10 nMTCDD, 75�g protein, 1.7 pmol/mg (D). The dolphin CYP1A1 bandmigrated similar to scup CYP1A at 54 kDa.
3.4. Immunochemical identification of CYP1A
Microsomal preparations from kidney and lung cellsexposed to 10 nM TCDD for 48 h in T-75 flasks wereimmunobloted with the Mab 1-12-3 that strongly rec-ognizes CYP1A1 but not CYP1A2 in mammals. Noband was detected in microsomes from DMSO treatedcells, in accord with the low basal levels of EROD activ-ity. As shown inFig. 3, immunoblotting confirmed thatCYP1A1 is present in bottlenose dolphin kidney andlung endothelial cells exposed to TCDD concentrationsthat produce significant increases in EROD activity.
Amounts of CYP1A1 in unknowns can be estimatedbased on the relative staining densities of the scupCYP1A standards. The CYP1A1 equivalents were cal-culated to be 1.2 pmol/mg in kidney endothelial cellsand 1.7 pmol/mg in lung endothelial cells treated with10 nM TCDD, a dose at which the EROD activity issimilar in the two types of endothelial cells.
4. Discussion
Cultures of endothelial cells from the lung andkidney of the bottlenose dolphin,Tursiops trunca-tus, exposed to the AHR agonists�NF and TCDDresponded with induction of CYP1A1 as indicatedby EROD activity and immunodetection of CYP1A.Based on EROD activity and immunoblot results,CYP1A1 levels were negligible in both kidney andl ndl eda hana ityi s toe AHRa ean.
ung cells treated with DMSO. In both kidney aung endothelial cells the levels of CYP1A1 inducfter TCDD treatment were about 10-fold greater tfter�NF treatment. The induction of EROD activ
n the kidney and lung endothelial cells allowed ustablish the first dose–response curves for angonist in endothelial cells cultured from a cetac

302 R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305
The dose–response curves demonstrate that endothe-lial cells originating from different tissues (lung andkidney) of the bottlenose dolphin respond similarlyto TCDD induction but have significantly differentmaximal CYP1A1 induction in response to�NF. Thissystem provides the opportunity to both characterizefactors that may determine the similarities and differ-ence in response and to explore how CYP1A1 functions(e.g. production of reactive oxygen or biotransforma-tion) may be associated with conditions in which theendothelium is compromised.
A consistent observation in vertebrates from fishto mammals is that the endothelium in many organsis a major extra-hepatic site of CYP1A1 induction byPHAHs or PAHs (Stegeman et al., 1989; Guiney et al.,1997; Wilson et al., 2005). The induction of CYP1A1in endothelium by�NF was observed also in vitro inskin tissue slices from whales (Godard et al., 2004). Incetacean populations, in vivo, an elevated level of AHRreceptor agonists can be correlated with the expres-sion of CYP1A1 detected immunohistochemically inintegument samples (Angell et al., 2004). Differentlevels of expression of CYP1A1 were reported forendothelium from several organs in populations of bel-uga whale in which the contaminant levels were known(Wilson et al., 2005). The supplementary data (doi:10.1289/eph.7664 athttp://dx.doi.org/) lists the differ-ent levels in endothelium from kidney and lung.
A differential induction of CYP1A1 expression inendothelium in different vessels in the vasculature hasb onice n,2 inv ns,c e ofv 3;G xi-c A1i orc ;G ,2 ndk antr ans.
orp ye im-i ul-
tures (Stegeman et al., 1995; Annas et al., 2000b). Inendothelial cells originally cultured from tissue frommature female eels (Garrick et al., 2005), in humanaortic endothelial cells (Celander et al., 1997) and inintegument tissue slices from sperm whale field sam-ples (Godard et al., 2004), there is a low expression ofCYP1A1 in endothelium even with no inducer presentin the test system. This low level of CYP1A1 expressionin endothelial cells or cetacean tissue slices has beeninterpreted as low levels of environmental induction infield samples, presence of some additional unknowninducers, or constitutive expression (Godard et al.,2004; Garrick et al., 2005). We do question whether thisvery low level of CYP1A1 activation would have sig-nificant physiological or toxicological consequences inthe vasculature.
The peak EROD rates induced with�NF are3× higher in the dolphin lung endothelial cellsthan in the kidney endothelial cells, 3.36 and0.92 pmol/mg/min, respectively, and are similar toEROD activity reported for other mammalian endothe-lial cells, 1–6 pmol/mg/min (Stegeman et al., 1995;Celander et al., 1997; Annas et al., 2000b). Endothelialcells cultured from two different vascular beds of theAmerican eel also have significantly different ERODactivity after exposure to�NF, i.e. 135 pmol/mg/minin endocardial cells and negligible activity in capillarycells from the rete mirabile (Garrick et al., 2005).
The maximum CYP1A1 activity induced by TCDDin both dolphin lung and kidney endothelial cells iss llsf n-s ells(w es-s ta 0bT ndtT arewr 5;G ta 001;T hed t ac herm
een observed in vivo, after either acute or chrxposure to AHR agonists (Schlezinger and Stegema000a; Wilson et al., 2005), and has been reporteditro for endothelial cells cultured from arteries, veiapillaries, endocardium and the microvasculaturertebrates (Annas et al., 2000b; Granberg et al., 200arrick et al., 2005). In cetaceans, exposure to toants also results in differential expression of CYP1n endothelium, i.e. primarily in the microvascularapillary endothelium, in the skin (Angell et al., 2004odard et al., 2004) and in other organs (Wilson et al.005). The availability of bottlenose dolphin lung aidney endothelial cells now allows studies of toxicesponse in cetacean endothelial cells from two org
There is negligible CYP1A1 activity (EROD)rotein present (Fig. 3) in either lung or kidnendothelial cells not exposed to inducer; this is s
lar to most other mammalian endothelial cell c
imilar to that induced in kidney endothelial cerom the American eel (16 pmol/mg/min) and coiderably lower than that in eel endocardial c142 pmol/mg/min) (Garrick et al., 2005) but wellithin the range reported for mammalian large vel endothelial cells, 3–30 pmol/mg/min (Stegeman el., 1995; Celander et al., 1997; Annas et al., 200).CDD is known to be a potent inducer of CYP1A1 a
his is reflected in the EC50s of 0.019 and 0.018 nMCDD for lung and kidney EC, respectively, thatithin the range of EC50s 0.015–0.045 nM TCDD
eported for other endothelial (Stegeman et al., 199arrick et al., 2005) or non-endothelial cells (Hahn el., 1996; Bello et al., 2001; Carlsson and Part, 2om et al., 2001). Thus, the endothelial cells from tolphin respond to in vitro induction by TCDD aoncentration similar to endothelial cells from otammals and teleosts.

R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305 303
In summary, we have shown that endothelial celllines grown from the microvasculature of the lung andthe kidney of the same bottlenose dolphin calf respondto �NF induction with a maximum level of CYP1A1activity similar to other mammalian endothelial cells,although there is a three-fold difference in the maxi-mum activity of the two dolphin cell lines. The responseto TCDD induction is quite similar in the two dolphinendothelial cell lines and is similar to the responses inother mammalian cells. We observed functional diver-sity in different areas of the vasculature in response toinduction with AHR agonists in endothelial cell linescultured from the American eel (Garrick et al., 2005).The data from the dolphin cell endothelial cell linessuggest that there are tissue specific differences in theresponse in marine mammals to some AHR agonistsand we now have the ability to explore this question onthe cellular level.
The similarity in the response to TCDD in the kidneyand lung endothelial cells suggest that the response topHAHs may be similar in other endothelium and thatthis might extend to skin endothelial cells in biopsies.Differences in the�NF response lead to the question ofwhether the response to PAHs might differ and whetherthis difference may be seen in skin endothelial cells aswell. There is need to develop additional cultures ofcells from cetaceans in order to explore the organ andspecies differences in susceptibility to AHR agoniststhat is suggested by field studies.
While the difference in response to�NF in thesec irr thep ce,C oryr nceso int tis-s ins lowl erep tureo
thee itros ust tivityr ncya es of
AHR action and CYP1A1 induction on endothelialcells from different parts of the vasculature, an impor-tant physiological question.
Acknowledgements
Part of this work was completed during a facultyfellowship from Fordham University for RAG. TheFaculty Research Council of Fordham University pro-vided partial support for RAG. This research was sup-ported by NIH grant 5-P42-ES07381 and by U.S.EPAgrant R827102-01-0. This research is an outgrowth andcontinuing impact of Sea Grant Number Grant No.NA90-AA-D-SG480, project NA86RG0075-R/P61.Biological materials fromTursiops truncatus werereceived under permit No. 939-682-00 (NOAA) toWHOI (JJS). Dr. Leslie D. Garrick and Dr. MarkE. Hahn are gratefully acknowledged for readingand commenting on the manuscript. James SaintJohn participated in the study with support from anAmbrose Summer Research Award from FordhamUniversity.
References
Aguilar, A., Borell, A., 1994. Abnormally high polychlorinatedbiphenyl levels in striped dolphins (Stenella coeruleoalba)affected by the 1990–1992 Mediterranean epizootic. Sci. TotalEnviron. 154, 237–247.
A 004.mpli-Mar.
A meyclic129,
A tialothe-dentppl.
B 00.cul-rons.
B uiredkilli-: inab-
ell populations is intriguing, the similarity in theesponses could provide important insights intoathophysiology of the endothelium. For instanYP1A1 expression is linked to an inflammat
esponse both in vivo and in vitro but the consequef inflammation can be quite different if it occurs
he brain or lung compared to in striated muscleue. If there is constitutive expression of CYP1A1ome species or a statistically significant but very
evel of induction of CYP1A1 in some organs, are thhysiological consequences of this in the vasculaf that animal or organ?
In conclusion, we can now extend the study offfects of toxicant exposure in cetaceans to in vtudies with endothelial cell lines. This will allowo explore such questions as inducer structure–acelationships, the basis for the toxic equivalepproach and to explore further the consequenc
ngell, C.M., Wilson, J.Y., Moore, M.J., Stegeman, J.J., 2Cytochrome P450 1A1 expression in cetacean integument: ications for detecting contaminant exposure and effects.Mammal Sci. 20 (3), 554–566.
nnas, A., Brittebo, E.B., 1998. Localization of cytochroP4501A1 and covalent binding of a mutagenic heterocamine in blood vessel endothelia of rodents. Toxicology145–156.
nnas, A., Granberg, A.L., Brittebo, E.B., 2000b. Differenresponse of cultured human umbilical vein and artery endlial cells to AhR receptor agonist treatment: CYP-depenactivation of food and environmental mutagens. Toxicol. APharmacol. 169, 94–101.
ayou-Denizot, C., Daval, J.-L., Netter, P., Minn, A., 20Xenobiotic-mediated production of superoxide by primarytures of rat cerebral endothelial cells, astrocytes and neuBiochim. Biophys. Acta 1497, 115–126.
ello, S., Franks, D.G., Stegeman, J., Hahn, M.E., 2001. Acqresistance to ah receptor agonists in a population of Atlanticfish (Fundulus heteroclitus) inhabiting a marine superfund sitevivo and in vitro studies on the inducibility of xenobiotic matolizing enzymes. Toxicol. Sci. 60, 77–91.

304 R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305
Ben-David, M., Kondratyuk, T., Woodin, B.R., Snyder, P.W., Stege-man, J.J., 2001. Induction of cytochrome P450 1A1 expressionin captive river otters fed prudoe bay crude oil: evaluation byimmunohistochemistry and quantitative RT-PCR. Biomarkers 6(3), 218–235.
Carlsson, C., Part, P., 2001. 7-Ethoxyresorufino-deethylase induc-tion in rainbow trout gill epithelium cultured on permeable sup-ports: asymmetrical distribution of substrate metabolites. Aquat.Toxicol. 54, 29–38.
Carvan, M.J., Flood, L.P., Campbell, B.D., Busbee, D.L., 1995.Effects of benzo(a)pyrene and tetrachlorodibenzo(p)dioxin onfetal dolphin kidney cells: inhibition of proliferation and initia-tion of DNA damage. Chemosphere 30 (1), 187–198.
Celander, M., Weisbrod, R., Stegeman, J.J., 1997. Glucocorti-coid potentiation of cytochrome P4501A1 induction by 2,3,7,8-tetrachlorodibenzo-p-dioxin in porcine and human endothelialcells in culture. Biochem. Biophys. Res. Commun. 232 (3),749–753.
Chat, M., Bayol-Denizoi, C., Suleman, G., Roux, F., Minn, A., 1998.Drug metabolizing enzyme activities and superoxide formationin primary and immortalized rat brain endothelial cells. Life Sci.62 (2), 151–163.
Drahushuk, A.T., McGarrigle, B.P., Slezak, B.P., Stegeman, J.J.,Olson, J.R., 1999. Time- and concentration-dependent induc-tion of CYP1A1 and cyp1a2 in precision-cut rat liver slicesincubated in dynamic organ culture in the presence of 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Appl. Pharmacol. 155,127–138.
Farin, F.M., Pohlman, T.H., Omiencinski, C.J., 1994. Expression ofcytochrome p450s and microsomal epoxide hydrolase in primarycultures of human umbilical vein endothelial cells. Toxicol. Appl.Pharmacol. 124, 1–9.
Ganassin, R.C., Schirmer, K., Bols, N.C., 2000. In: Ostrander, G.K.(Ed.), Cell and Tissue Culture, vol. 38. Academic Press, SanDiego, pp. 631–651.
G the-,
G omeom
G ege-1 bySci.
G 03.A1-n
G 1997.
in(2),
H per-icol.
Hahn, M.E., Lamb, T.M., Schultz, M.E., Smolowitz, R.M., Stege-man, J.J., 1993. Cytochrome P4501A induction and inhibitionby 3,3′,4,4′-tetrachlorobiphenyl in an ah receptor-containing fishhepatoma cell line (PLHC-1). Aquat. Toxicol. 26, 185–208.
Hahn, M.E., Woodward, B.L., Stegeman, J.J., Kennedy, S.W., 1996.Rapid assessment of induced cytochrome P4501A (CYP1A)protein and catalytic activity in fish hepatoma cells grown inmulti-well plates. Environ. Toxicol. Chem. 15 (4), 582–591.
Hall, A.J., Law, R.J., Wells, D.E., Harwood, J., Ross, H.M.,Kennedy, S., Allchin, C.R., Campbell, I.A., Pomeroy, P.P., 1992.Organochloride levels in common seals (phoca vitulina) whichwere victims and survivors of the 1988 phocine distemper epi-zootic. Sci. Total Environ. 115, 145–162.
Hennig, B., Hammonck, B.D., Slim, R., Toberak, M., Saraswathi,V., Robertson, L.W., 2002a. PCB-induced oxidative stress inendothelial cell: modulation by nutrients. Int. J. Hyg. Environ.Health 205, 95–102.
Hennig, B., Meerarani, P., Slim, R., Toberak, M., Daugherty, A.,Silverstone, A., Robertson, L.W., 2002b. Proinflammatory prop-erties of coplanar PCBs: in vitro and in vivo evidence. Toxicol.Appl. Pharmacol. 181, 174–183.
Hestermann, E.V., Stegeman, J.J., Hahn, M.E., 2000a. Relative con-tributions of affinity and intrinsic efficacy to aryl hydrocarbonreceptor ligand potency. Toxicol. Appl. Pharmacol. 168 (2),160–172.
Hestermann, E.V., Stegeman, J.J., Hahn, M.E., 2000b. Serum altersthe uptake and relative potencies of halogenated aromatic hydro-carbons in cell culture bioassays. Toxicol. Sci. 53 (2), 316–325.
Hestermann, E.V., Stegeman, J.J., Hahn, M.E., 2002. Serum with-drawal leads to reduced aryl hydrocarbon receptor expressionand loss of cytochrome P4501A inducibility in PLHC-1 cells.Biochem. Pharmacol. 63 (8), 1405–1414.
Jensen, B., Hahn, M.E., 2001. cDNA cloning and characterizationof a high affinity aryl hydrocarbon receptor in the cetacean, thebeluga,Delphinapterus leucas. Toxicol. Sci. 64, 41–56.
K 993.ringto-racted
L art,free-deral
M E.,ahs)hemo-
M .P.,dy:vion-
M scep-
nnial
arrick, R.A., 2000. Isolation and culture of capillary endolial cells from the eel,Anguilla rostrata. Microvasc. Res. 59377–385.
arrick, R.A., Woodin, B.R., Stegeman, J.J., 2005. CytochrP4501A induced differentially in endothelial cells cultured frAnguilla rostrata. In Vitro Cell Dev. Biol. -Anim. 41, 57–63.
odard, C.A., Smolowitz, R.M., Wilson, J.Y., Payne, R.S., Stman, J.J., 2004. Induction of cetacean cytochrome P4501A�-naphthoflavone exposure of skin biopsy slices. Toxicol.80 (2), 268–275.
ranberg, L., Ostergren, A., Brandt, I., Brittebo, E.B., 20CYP1A1 and cyp1b1 in blood–brain interfaces: CYP1dependent bioactivation of 7,12-dimethylbenz(a)anthracene iendothelial cells. Drug Metab. Dispos. 31 (3), 259–265.
uiney, P.D., Smolowitz, R.M., Peterson, R.E., Stegeman, J.J.,Correlation of 2,3,7,8-tetrachlorodibenzo-p-dioxin induction ofcytochrome P4501A in vascular endothelium with toxicityearly life stages of lake trout. Toxicol. Appl. Pharmacol. 143256–273.
ahn, M.E., 1998. The aryl hydrocarbon receptor: a comparativespective. Comp. Biochem. Physiol. Part C: Pharmacol. ToxEndocrinol. 121, 23–53.
ennedy, S.W., Lorenzen, A., James, C.A., Collins, B.T., 1A rapid and sensitive cell culture bioassay for measuethoxyresorufin-o-deethylase (erod) activity in cultured hepacytes exposed to halogenated aromatic hydrocarbons extfrom wild bird eggs. Chemosphere 27, 367–373.
ahvis, G.P., Wells, R.S., Kuehl, D.W., Stewart, J.L., RhinehH.L., Via, C.S., 1995. Decreased lymphocyte responses inranging bottlenose dolphins (Tursiops truncatus) are associatewith increases concentrations of PCBs and DDT in periphblood. Environ. Health Perspect. 103 (Suppl. 4), 67–72.
arsili, L., Caruso, A., Fossi, M.C., Zanardelli, M., Politi,Focardi, S., 2001. Polycyclic aromatic hydrocarbons (pin subcutaneous biopsies of Mediterranean cetaceans. Csphere 44, 147–154.
artineau, D., Lemberger, K.A.D., Labelle, P., Lipscomb, TMichel, P., Mikaelian, I., 2002. Cancer in wildlife, a case stuBeluga from the St. Lawrence estuary, Quebec, Canada, Enmental Health Perspectives 110, 285–292.
iddlebrooks, R., Colvocoresses, J., Patterson, R.A., 1999. Sutibility of two developmental cell lines fromTursiops truncatusto a range of mammalian viruses. Abstracts of the 13th BieConference on the Biology of Marine Mammals.

R.A. Garrick et al. / Aquatic Toxicology 76 (2006) 295–305 305
Mimura, J., Fujii-Kuriyama, Y., 2003. Functional role of AhR in theexpression of toxic effects by TCDD. Biochim. Biophys. Acta1619, 263–268.
Park, S.S., Miller, H., Klotz, A.V., Kloepper-Sams, P.J., Stegeman,J.J., Gelboin, H.V., 1986. Monoclonal antibodies to liver micro-somal cytochrome p-450e of the marine fishStenotomus chrysops(scup): cross reactivity with 3-methylcholanthrene induced ratcytochrome P-450. Arch. Biochem. Biophys. 249 (2), 339–350.
Pfeiffer, C.J., Sharova, L.V., Gray, L., 2000. Functional and ultra-structural cell pathology induced by fuel oil in cultured dolphinrenal cells. Ecotoxicol. Environ. Safety 47, 210–217.
Reijnders, P.J.H., 1986. Reproductive failure in common seals feed-ing on fish from polluted coastal waters. Nature 324, 456–457.
Schlezinger, J.J., Stegeman, J.J., 2000a. Dose and inducer-dependentinduction of cytochrome P450 1A in endothelia of the eel, includ-ing in the swimbladder rete mirabile, a model microvascularstructure. Drug Metab. Dispos. 28 (6), 701–708.
Shimada, T., Fujii-Kuriyama, Y., 2004. Metabolic activation of poly-cyclic aromatic hydrocarbons to carcinogens by cytochromesP450 1A1 and 1B1. Cancer Sci. 95 (1), 1–6.
Stegeman, J.J., Hahn, M.E., Weisbrod, R., Woodin, B.R., Joy, J.S.,Najibi, S., Cohen, R.A., 1995. Induction of cytochrome P4501A1by aryl hydrocarbon receptor agonists in porcine aorta endothelialcells in culture and cytochrome P4501A1 activity in intact cells.Mol. Pharmacol. 47 (2), 296–306.
Stegeman, J.J., Miller, M.R., Hinton, D.E., 1989. CytochromeP4501A1 induction and localization in endothelium of vertebrate(teleost) heart. Mol. Pharmacol. 36, 723–729.
Stegeman, J.J., Woodin, B.R., Klotz, A.V., Wolke, R.E., Orme-Johnson, N.R., 1982. Cytochrome P-450 and monooxyge-nase activity in cardiac microsomes from the fishStenotomuschrysops. Mol. Pharmacol. 21, 517–526.
Sweat, J.M., Dunigan, D.D., Wright, S.D., 2001. Characterization ofkidney epithelial cells from the florida manatee,trichecus man-atus latirostris. In Vitro Cell Dev. Biol. -Anim. 37, 386–394.
Teraoka, H., Dong, W., Tsujimoto, Y., Iwasa, H., Endoh, D., Ueno,N., Stegeman, J.J., Peterson, R.E., Hiraga, T., 2003. Induction ofcytochrome P450 1A is required for circulation failure and edemaby 2,3,7,8-tetrachlorodibenzo-p-dioxin in zebrafish. Biochem.Biophys. Res. Commun. 304, 223–228.
Thum, T., Haverich, A., Borlak, J., 2000. Cellular dedifferentiationof endothelium is linked to activation and silencing of certainnuclear transcription factors: implication for endothelial dysfunc-tion and vascular biology. FASEB J. 14, 740–751.
Tom, D.J., Lee, L.E., Lew, J., Bols, N.C., 2001. Induction of7-ethoxyresorufin-o-deethylase activity by planar chlorinatedhydrocarbons and polycyclic aromatic hydrocarbons in cell linesfrom the rainbow trout pituitary. Comp. Biochem. Physiol. A:Mol. Integr. Physiol. 128 (2), 185–198.
White, R.D., Hahn, M.E., Lockhart, L.L., Stegeman, J.J., 1994. Cat-alytic and immunochemical characterization of hepatic microso-mal cytochromes P450 in beluga whale,Delphinapterus leucas.Toxicol. Appl. Pharm. 126, 45–57.
White, R.D., Shea, D., Schlezinger, J.J., Hahn, M.E., Stegeman,J.J., 2000. In vitro metabolism of polychlorinated biphenyl con-geners by beluga whale (Delphinapterus leucas) and pilot whales(globicephalea melaena) and relationship to cytochrome P450expression. Comp. Biochem. Physiol. Part C: Pharmacol. Toxi-col. Endocrinol. 126, 267–284.
Wilson, J.Y., Cooke, S.R., Moore, M.J., Martineau, D., Mikaelian,I., Metner, D.A., Lockhart, W.L., Stegeman, J., 2005. Sys-temic effects of arctic pollutants in beluga whales indicated byCYP1A1 expression. Environ. Health Perspect. 113 (11), 1594–1599.