cumulative influence of elastin peptides and plasminogen on matrix metalloproteinase activation and...
TRANSCRIPT

Clinical & Experimental Metastasis 19: 107–117, 2002.© 2002 Kluwer Academic Publishers. Printed in the Netherlands.
107
Cumulative influence of elastin peptides and plasminogen on matrixmetalloproteinase activation and type I collagen invasion by HT-1080fibrosarcoma cells
Eric Huet1,∗, Bertrand Brassart1,∗, Jean-Hubert Cauchard1, Laurent Debelle1, Philippe Birembaut2,Jean Wallach3, Herve Emonard1, Myriam Polette2 & William Hornebeck1
1FRE 2260 CNRS; 2INSERM U514, Institut Federatif de Recherche Biomolecules (IFR 53), Universite de Reims (URCA),France; 3Laboratoire de Biochimie Analytique, Universite Claude Bernard, Lyon, France
Received 8 May 2001; accepted in revised form 13 September 2001
Key words: cell invasion, elastin, elastin receptor, matrix metalloproteinase, plasmin, type I collagen
Abstract
HT-1080 fibrosarcoma cells express at their plasma membrane the elastin-binding protein (EBP). Occupancy of EBP byelastin fragments, tropoelastin or XGVAPG peptides was found to trigger procollagenase-1 (proMMP-1) overproductionby HT-1080 cells at the protein and enzyme levels. RT-PCR analysis indicated that elastin peptides did not modify theMMP-1 mRNA steady state levels, suggesting the involvement of a post-transcriptional mechanism. We previously reportedthat binding of elastin peptides to EBP induced other matrix metalloproteinases (MMP-2 and MT1-MMP) expression [20].Since those peptides were here found to also accelerate the secretion of urokinase from HT-1080 cells, culture medium wassupplemented with plasminogen together with elastin peptides at aims to induce or potentiate MMPs activation cascades. Insuch conditions, plasmin activity was generated and exacerbate proMMP-1 and proMMP-2 activation. As a consequence,elastin peptides and plasminogen-treated HT-1080 cells displayed a significant type I collagen matrix invasive capacity.
Abbreviations: ECM – extracellular matrix; EBP – elastin-binding protein; EDPs – elastin-derived peptides; HLE – humanleukocyte elastase; kE – kappa-elastin peptides; MMP – matrix metalloproteinase; PAI – plasminogen activator inhibitor;Plgn – plasminogen; TIMP – tissue inhibitor of metalloproteinase; uPA – urokinase-type plasminogen activator
Introduction
Extracellular matrix (ECM) degradation is required for ma-lignant progression. Acting mainly in the pericellular en-vironment, neutral proteinases belonging to the plasminsystem and to the matrix metalloproteinase (MMP) fam-ily actively participate in tumor invasion and subsequentmetastatic dissemination [1–5]. Such focalized proteolysiscreates several cryptic sites within the ECM and releasesmatrix fragments [6]. Matrix and matrix-derived peptideswere shown to modulate proteinase expression, further en-hancing or repressing tissue invasion by cancer cells [7–10].The contribution of several types of collagen and adhesiveglycoproteins in this process is now largely documented, butthe participation of elastin has been mostly ignored. Bindingof peptide motifs present, not only in elastin, but also inlaminin, to a plasma membrane-associated protein, also des-ignated as elastin-binding protein or EBP, has been reportedto trigger cell activation [11]. EBP was characterized as a
∗These two authors contributed equally to this work.
Correspondence to: Dr William Hornebeek, CNRS FRE 2260, Facultede Medecine, 51, rue Cognacq-Jay, 51100 Reims, France. Tel: +33-3-26913535; Fax: +33-3-26918055; E-mail: [email protected]
67 kDa splice variant of beta-galactosidase, and thereforewas distinct from integrins and another laminin-receptor ofsimilar molecular mass [11]. In elastin-producing cells, thisprotein was identified in endosomal compartments whereit binds tropoelastin, facilitating its secretion and assemblyinto elastic fibers. It is recycled to the cell membrane andacts as a true receptor for elastin peptide motifs mediat-ing signal transduction and cell activation [12]. Interactionbetween elastin peptides and EBP can direct cell migra-tion and was reported to increase the rate of proliferationof human skin fibroblasts [13, 14]; it also modifies cal-cium fluxes in human skin fibroblasts and smooth musclecells [15], and induces the release of H2O2 and elastase inmonocytes [16]. Moreover, the presence of EBP in severalhighly metastasizing cell lines was reported to be correlatedwith their potentiality to metastasize to elastin-rich tissuesuch as lung [17, 18]. We initially demonstrated that thebinding of elastin-derived peptides to metastatic lung carci-noma cell lines increased intracellular Ca2+ concentration,induced cell chemotaxis towards elastin peptide gradient aswell as the secretion of metalloelastase-type activity [19].More recently, we evidenced that occupancy of EBP in HT-1080 cells by elastin peptides led to increased expression ofMMP-2 and MT1-MMP [20]; elastin-mediated overexpres-

108 E. Huet et al.
sion of those enzymes was accompanied by an enhancedcapacity of those cells to invade matrigel (our unpublisheddata).
Along with degradation of type IV collagen and lamininin basement membrane, destruction of the stromal colla-gens is a key step in the invasive/metastatic behavior oftumor cells. Several proteinases were shown to displaycollagen-degrading capacity, including collagenases (MMP-1 and MMP-13) but also MT1-MMP and MMP-2 [21, 22].Their de novo synthesis, activation and interaction with theirtissue specific inhibitors, TIMPs, regulate their enzyme ac-tivity and therefore collagenolysis. Overproduction of thoseproenzymes is not sufficient for the acquisition of an invasivephenotype. Processing of their N-terminal part at the peri orextracellular environment represents an important level ofregulation controlling the matrix invasive capacity of cancercells [23]. Activation of MMP-1 involves a proteolytic cas-cade, which can be initiated by removal of its N-terminaldomain by the plasminogen/plasmin system, generating a42 kDa active enzyme [24]. ProMMP-2 processing to activeenzyme is more complex and is mediated by the formationof a membrane-bound ternary complex consisting of MT1-MMP, TIMP-2 and latent proMMP-2. Another uncomplexedactivated MT1-MMP molecule, located close to this ternarycomplex was proposed to cleave, in a second step, proMMP-2 bound to MT1-MMP/TIMP-2 at the cell plasma membrane[25, 26].
Our aims was therefore to investigate whether elastin-derived peptides could initiate these activation cascadesand to study their respective contributions in the capacityof HT-1080 cells to invade a type I collagen matrix. Wedemonstrated that binding of elastin peptides to EBP in HT-1080 cells, could stimulate and accelerate the production ofMMP-1 and urokinase (uPA). Consequently, simultaneoustreatment of those cells by elastin peptides and plasminogengenerated plasmin activity, and stimulated the conversion ofproMMP-1 and proMMP-2 to active enzyme species. Suchinduced activation of those proteolytic cascades, by elastinpeptides and plasminogen, further led to enhanced ability ofcancer cells to invade a type I collagen matrix.
Materials and methods
Reagents
Culture reagents were from Gibco BRL (Life Technologies,Cergy Pontoise, France). Gelatin, bovine alpha-casein, Ma-trigel (ECM gel), lactose and actinomycin D were purchasedfrom Sigma (Saint Quentin Fallavier, France). Human pro-collagenase (proMMP-1), human recombinant progelati-nase A (proMMP-2), human recombinant TIMP-2, sheeppolyclonal anti-human MMP-1 and MMP-3, human plas-minogen, plasmin and urokinase were purchased from Cal-biochem (Merck Eurolab SA, Fontenay-sous-Bois, France).Type I collagen was prepared from rat-tail as described[27]. H-D-Val-Leu-Lys-pNA (S-2251) was purchased fromChromogenix (Biogenic SA, Maurin, France) and plasminactivity was determined as described [27].
Preparation of elastin-derived peptides (EDPs)
Insoluble elastin was prepared from bovine ligamentumnuchae by hot alkali treatment and its purity assessed byamino acid analysis [28]. The lack of hexoses and hex-osamines was determined in the preparation. Soluble kappa-elastin peptides (kE) were obtained from purified insolubleelastin by organo-alkaline hydrolysis [29]. Peptides with anaverage molecular weight of 75 kDa were isolated by gelpermeation on Sephadex G-100. For production of elas-tase hydrolysates, 75 mg of insoluble elastin were groundto a < 100 mesh, dispersed in 7.5 ml of 100 mM Tris,5 mM CaCl2, pH 8.0 and maintained under constant me-chanical stirring for 1 h at 37 ◦C. 37.5 µg (1.25 µmol)of human leukocyte elastase (HLE) (Elastin Products Co.,Owensville, USA) were added to elastin dispersion. Flaskswere incubated at 37 ◦C under constant stirring for 30 to1,980 min, and reaction was stopped by addition of phenyl-methanesulfonylfluoride (Pmsf; 1 mM). Elastin dispersionswere centrifuged at 12,000 g for 15 min, supernatants wereremoved and pellets were washed 3 times with 10 ml of dis-tilled water, lyophilized and weighed. The protein content ofsupernatants was determined using kE as standard [28].
Peptides synthesis
Peptides were synthesized using 9-fluoromethoxycarbonyl(Fmoc) chemistry, using a Fmoc-Val-resin (0.22 meq/g).Couplings were performed with Fmoc-amino acid-pentafluo-rophenyl esters (Pfb) (four molar excess). Each Fmocdeprotection step involved treatment with 20% piperi-dine/dimethylformamide for 10 min. Cleavage of the peptidefrom the resin was achieved by a 6 h treatment with tri-fluoroacetic acid/water (95:5; v/v), followed by successivewashings of the resin with ether. Purity of peptides wasconfirmed by HPLC and by fast atom bombardment massspectrometry. Both VXVAPG and XGVAPG libraries wereconstructed using standard solid-phase peptide chemistrywith Fmoc protected amino acids on Wang resins usingsplit-synthesis methodology [30, 31]. Fmoc-VAPGV wasfirst synthesized, starting with a Fmoc-Val-Wang resin (sub-stitution: 0.64 mmol/g), and separated in two respectivefractions, for VXVAPGV and XGVAPGV syntheses. Theformer was further divided into 19 subfractions (20 mgeach), in a Multipeptide Biotech BT 400 device. In each wellwas coupled a Fmoc-amino acid-OPfp ester (except Cys).After washings and drying, the resins were pooled beforethe final addition of valine. The latter library was constructedin a similar way, except that Fmoc-Gly-OPfp was added toVAPGV resin, before the final ‘split-combine’ process.
Cell culture
Human fibrosarcoma cells (HT-1080 cells) were obtainedfrom the American Type Culture Collection (Rockville,USA). Cells were cultured in DMEM supplemented with10% (v/v) fetal calf serum (FCS), 2 mM glutamine,100 units/ml penicillin and 100 µg/ml streptomycin. Whenthe cells reached subconfluence, they were washed twice

Stimulation of HT-1080 cells invasion by elastin peptides 109
Figure 1. Elastin-derived peptides stimulate the production of collagenase-1 (MMP-1) by HT-1080 fibrosarcoma cells in a dose-dependent manner. Cells atsubconfluence have been treated with kappa-elastin (kE) in serum-free DMEM for 24 h. Abscissa: kE concentration in µg/ml. Ordinates: MMP-1 amountin ng/105 cells as determined by ELISA. The data in triplicate show the mean ± SD. ∗∗: significantly different from the untreated cells (0.001 < P < 0.01).Insert: Western blot analysis of concentrated conditioned media (24 h of culture). 1 – Control in absence of kE. 2 – HT-1080 cells have been treated with50 µg/ml of kE.
with PBS to remove residual FCS and incubated for 6 to24 h in serum-free medium, with or without EDPs. Condi-tioned media were harvested and centrifuged at 500 g for10 min at 4 ◦C to remove cellular debris. The influence ofelastin-derived peptides on cell growth was evaluated usingthe Crystal violet assay and cell toxicity was assessed bythe trypan blue exclusion technique. The protein content ofconditioned medium was determined by the Lowry methodusing bovine serum albumin as standard.
Zymographic analyses
Gelatin zymographyConditioned media from HT-1080 cells cultures were an-alyzed for gelatin degradation by electrophoresis undernon-reducing conditions on a SDS-polyacrylamide gel con-taining 1 mg/ml denatured type I collagen. The volumeof conditioned medium loaded per lane was standardizedon the basis of the cell count at harvest. Gels were incu-bated overnight at room temperature in 50 mM Tris-HCl,150 mM NaCl, 10 mM CaCl2, pH 7.4. White lysis zones,indicative of degrading activity, were revealed by stainingwith Coomassie brilliant blue. As positive controls, ei-ther proMMP-2 or a 1 mM aminophenylmercuric acetate(APMA)-activated MMP-2 was used. As a negative control,10 mM EDTA was added to the incubation buffer.
Casein/plasminogen zymographyFor the determination of plasminogen activators, condi-tioned media were analyzed on SDS-polyacrylamide gelscontaining 1 mg/ml alpha-casein and 10 µg/ml plasminogen.Following electrophoresis, gels were incubated overnight atroom temperature in 100 mM glycine buffer, EDTA 5 mM,pH 8.0. Caseinolytic activity resulting from plasminogenactivation was indicated here by white lysis zones whichwere revealed by staining with Coomassie brilliant blue. Asa positive control, urokinase was used; as a negative control,2 mM Pmsf was added into the incubation buffer.
Gelatin reverse zymographyFor tissue inhibitors of metalloproteinase (TIMPs) identifi-cation, SDS gels containing 0.1% gelatin, 15% polyacry-lamide and 20 ng/ml activated MMP-2 were used. Reversezymography revealed inhibitory activity, which appeared asblue zones against a clear background, demonstrating inhi-bition of gelatin lysis in the gels. Either TIMP-2 (0.1 ng) orTIMP-1 (0.1 ng) was used as marker in the gels.
Western blot analysis
Conditioned media were concentrated in NanosepTM (Poly-labo, Strasbourg, France) and samples, adjusted to the sameprotein concentration, were electrophoresed in a 0.1% SDS,10% polyacrylamide gel, under reducing conditions. Theywere then transferred onto Immobilon-P membranes (Milli-pore, Saint-Quentin en Yvelines, France). The membranes
110 E. Huet et al.
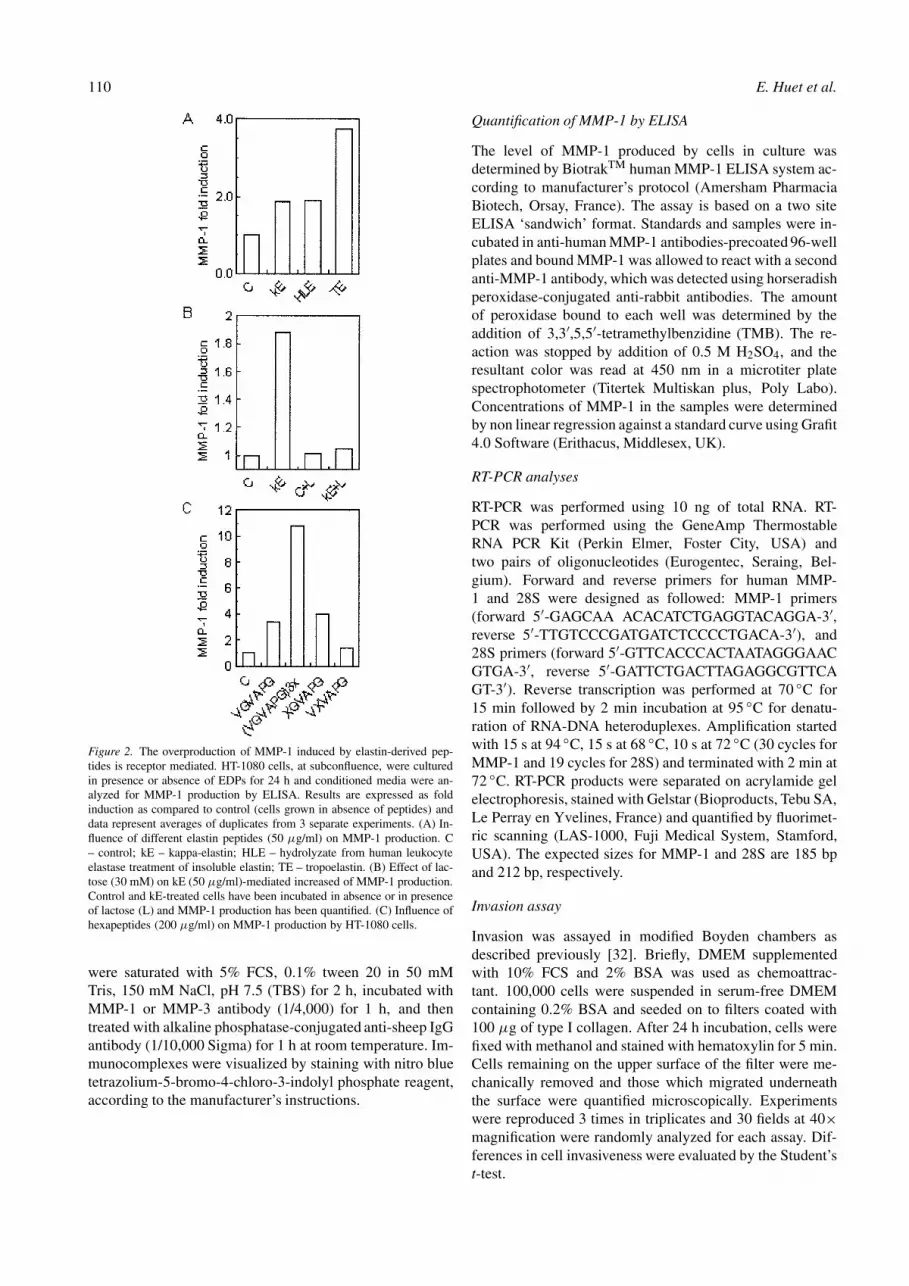
Figure 2. The overproduction of MMP-1 induced by elastin-derived pep-tides is receptor mediated. HT-1080 cells, at subconfluence, were culturedin presence or absence of EDPs for 24 h and conditioned media were an-alyzed for MMP-1 production by ELISA. Results are expressed as foldinduction as compared to control (cells grown in absence of peptides) anddata represent averages of duplicates from 3 separate experiments. (A) In-fluence of different elastin peptides (50 µg/ml) on MMP-1 production. C– control; kE – kappa-elastin; HLE – hydrolyzate from human leukocyteelastase treatment of insoluble elastin; TE – tropoelastin. (B) Effect of lac-tose (30 mM) on kE (50 µg/ml)-mediated increased of MMP-1 production.Control and kE-treated cells have been incubated in absence or in presenceof lactose (L) and MMP-1 production has been quantified. (C) Influence ofhexapeptides (200 µg/ml) on MMP-1 production by HT-1080 cells.
were saturated with 5% FCS, 0.1% tween 20 in 50 mMTris, 150 mM NaCl, pH 7.5 (TBS) for 2 h, incubated withMMP-1 or MMP-3 antibody (1/4,000) for 1 h, and thentreated with alkaline phosphatase-conjugated anti-sheep IgGantibody (1/10,000 Sigma) for 1 h at room temperature. Im-munocomplexes were visualized by staining with nitro bluetetrazolium-5-bromo-4-chloro-3-indolyl phosphate reagent,according to the manufacturer’s instructions.
Quantification of MMP-1 by ELISA
The level of MMP-1 produced by cells in culture wasdetermined by BiotrakTM human MMP-1 ELISA system ac-cording to manufacturer’s protocol (Amersham PharmaciaBiotech, Orsay, France). The assay is based on a two siteELISA ‘sandwich’ format. Standards and samples were in-cubated in anti-human MMP-1 antibodies-precoated 96-wellplates and bound MMP-1 was allowed to react with a secondanti-MMP-1 antibody, which was detected using horseradishperoxidase-conjugated anti-rabbit antibodies. The amountof peroxidase bound to each well was determined by theaddition of 3,3′,5,5′-tetramethylbenzidine (TMB). The re-action was stopped by addition of 0.5 M H2SO4, and theresultant color was read at 450 nm in a microtiter platespectrophotometer (Titertek Multiskan plus, Poly Labo).Concentrations of MMP-1 in the samples were determinedby non linear regression against a standard curve using Grafit4.0 Software (Erithacus, Middlesex, UK).
RT-PCR analyses
RT-PCR was performed using 10 ng of total RNA. RT-PCR was performed using the GeneAmp ThermostableRNA PCR Kit (Perkin Elmer, Foster City, USA) andtwo pairs of oligonucleotides (Eurogentec, Seraing, Bel-gium). Forward and reverse primers for human MMP-1 and 28S were designed as followed: MMP-1 primers(forward 5′-GAGCAA ACACATCTGAGGTACAGGA-3′,reverse 5′-TTGTCCCGATGATCTCCCCTGACA-3′), and28S primers (forward 5′-GTTCACCCACTAATAGGGAACGTGA-3′, reverse 5′-GATTCTGACTTAGAGGCGTTCAGT-3′). Reverse transcription was performed at 70 ◦C for15 min followed by 2 min incubation at 95 ◦C for denatu-ration of RNA-DNA heteroduplexes. Amplification startedwith 15 s at 94 ◦C, 15 s at 68 ◦C, 10 s at 72 ◦C (30 cycles forMMP-1 and 19 cycles for 28S) and terminated with 2 min at72 ◦C. RT-PCR products were separated on acrylamide gelelectrophoresis, stained with Gelstar (Bioproducts, Tebu SA,Le Perray en Yvelines, France) and quantified by fluorimet-ric scanning (LAS-1000, Fuji Medical System, Stamford,USA). The expected sizes for MMP-1 and 28S are 185 bpand 212 bp, respectively.
Invasion assay
Invasion was assayed in modified Boyden chambers asdescribed previously [32]. Briefly, DMEM supplementedwith 10% FCS and 2% BSA was used as chemoattrac-tant. 100,000 cells were suspended in serum-free DMEMcontaining 0.2% BSA and seeded on to filters coated with100 µg of type I collagen. After 24 h incubation, cells werefixed with methanol and stained with hematoxylin for 5 min.Cells remaining on the upper surface of the filter were me-chanically removed and those which migrated underneaththe surface were quantified microscopically. Experimentswere reproduced 3 times in triplicates and 30 fields at 40×magnification were randomly analyzed for each assay. Dif-ferences in cell invasiveness were evaluated by the Student’st-test.

Stimulation of HT-1080 cells invasion by elastin peptides 111
Figure 3. Elastin-derived peptides (kE) increase MMP-1 and MMP-2 pro-duction from HT-1080 cells by distinct mechanisms. (A) RT-PCR analysisof MMP-1 mRNA steady state levels (see text for details). HT-1080 cellshave been treated or not (control: C) with 50 µg/ml of kE for 30 min to 24 h.(B) Variation of MMP-1 mRNA levels following treatment of HT-1080 cellswith kE (50 µg/ml) for 30 min to 24 h. 28S mRNA levels were used asreporters for MMP-1. � control; � elastin peptides-treated cells. (C) Effectof actinomycin D (1 µg/ml) on the production of MMP-1 and MMP-2 inkE (50 µg/ml)-treated HT-1080 cells for 6 h. Production of MMP-1 andMMP-2 has been analyzed by Western blot and gelatin zymography, re-spectively. 1 – Control. 2 – kE-treated cells. 3 – kE-treated cells in presenceof actinomycin D.
Statistical analysis
Data were analyzed by the Student’s t-test; P < 0.05 wasconsidered to be statistically significant.
Results
Elastin peptides stimulate the production of proMMP-1
Supplementation of serum-free culture media with elastin-derived peptides (1–100 ng/ml) did not induce HT-1080cells growth (from 6–72 h of culture). Those peptides, also,had no cell toxicity and did not modify the overall proteincontent of conditioned media over 24 h of culture.
To investigate the effect of elastin-derived peptides onMMP-1 production by HT-1080 cells, conditioned mediafrom cells treated with kE (10 to 100 µg/ml) for 24 h wereanalyzed by western blot or ELISA. Media from control cul-tures exhibited two immunoreactive species with apparentmolecular weights of 53 kDa and 57 kDa corresponding tothe non-glycosylated and glycosylated proMMP-1 isoforms.The level of proMMP-1 secretion was significantly enhancedwhen cells were grown in the presence of kE (50 µg/ml) for24 h (Figure 1 insert).
Quantitative analysis of proMMP-1 production byELISA indicated that kE-mediated effect was concentration-dependent; a significant 1.4-fold increase in enzyme se-cretion was observed following supplementation of culturemedium with 10 µg/ml of kE and the maximum level ofproMMP-1 production by HT-1080 cells grown in presenceof kE was found equal to 2.5 ng/105 cells (Figure 1).
Next, we examined whether other elastin peptides couldalso modulate MMP-1 production by HT-1080 cells. Aftera 24 h hydrolysis of insoluble elastin by elastase, the su-pernatant was withdrawn and added to fibrosarcoma cellsat 50 µg/ml for 24 h. Following this, the level of secretedMMP-1 increased 2-fold; similarly, tropoelastin (50 µg/ml),the soluble precursor of insoluble elastin synthesized bymesenchymal cells, induced a nearly 4-fold enhancement ofMMP-1 secretion (Figure 2A).
Increased production of proMMP-1 by elastin peptides isreceptor mediated
The S-gal elastin receptor (EBP) was shown to exhibit sev-eral characteristics. Particularly, it possesses galactolectin-like property and interacts with a VGVAPG hexapeptidesequence repeated 6-fold in human tropoelastin [11]. Toevaluate whether the effect of elastin fragments on MMP-1 production was receptor-mediated, the influence of lactoseand VGVAPG peptides on the amount of enzyme productionby HT-1080 cells was studied. Data from Figure 2B showsthat lactose, used at a concentration as low as 30 mM, cannearly totally suppressed the kE-mediated MMP-1 inductionby HT-1080 cells; glucose, at the same concentration, hadno effect (not shown). In addition, collagenase inductioncould be reproduced by VGVAPG elastin-derived peptides.Interestingly, increasing the number of VGVAPG repeats ledto increased level of MMP-1 production (Figure 2C). Ourprevious data indicated that only peptides with a GXXPGconsensus sequence were able to induce MMP-1 in humanskin fibroblasts [33]. To delineate the relative importance ofthe two first terminal amino acids on MMP-1 secretion byHT-1080 cells, XGVAPG and VXVAPG peptide librarieswere synthesized. Figure 2C shows that XGVAPG but notVXVAPG libraries exhibited collagenase inducible effect.This result further emphasized the crucial importance of theG residue, and thus the conformation of the hexapeptide onEBP binding and MMP expression by fibrosarcoma cells.
Total RNAs obtained from HT-1080 cells cultured for30 min to 24 h in absence or presence of kE (50 µg/ml)were analyzed by RT-PCR. kE treatment induced only minoralteration of MMP-1 mRNA levels (Figures 3A–B). On the

112 E. Huet et al.
Figure 4. Influence of elastin-derived peptides on urokinase expression and generation of plasmin activity by HT-1080 cells. (A) Urokinase production.Analyzed by plasminogen/casein zymography; the position of urokinase run in the same gel is indicated. Lanes 1 and 3 correspond to conditioned mediafrom controls at 6 and 24 h of culture. Lanes 2 and 4 correspond to conditioned media from kE (50 µg/ml)-treated cells at 6 and 24 h of culture. (B) Plasminactivity. Culture media from either control (1 and 3) or kE (50 µg/ml)-treated cells (2 and 4) has been supplemented with plasminogen and plasmin activity(expressed as mU/105 cells) has been determined following 6 h (1 and 2) and 24 h (3 and 4) of culture using S-2251 as substrate. The data in triplicateshow the mean ± SD. ∗∗∗ Significantly different from the control cells (P < 0.001).
contrary, as we previously reported [20], occupancy of EBPby elastin peptides led to a significant 3-fold enhancement ofMMP-2 mRNA levels. Consistently, actinomycin D, an an-tibiotic which blocks RNA transcription, inhibited kE effecton MMP-2 production while having no influence on MMP-1secretion (Figure 3C).
Elastin fragments stimulate the rate of secretion ofurokinase from HT-1080 cells
Although elastin peptides had major influence on MMP-1production, under no conditions could enzyme activationbe evidenced. Generation of active MMP-1 in differentcell culture conditions, generally necessitates the combinedproduction of active plasmin through overexpression ofplasminogen activators and/or decreased expression of plas-minogen activator inhibitors (PAIs) [34]. Such reaction is
accelerated by stromelysin-1 (MMP-3) which completes theultimate step of collagenase activation [35].
Similarly, as reported by other investigators, we couldnot detect MMP-3 as well as MMP-13 expression by HT-1080 cells [9, 10] (data not shown). Conditioned mediafrom HT-1080 cells were analyzed by casein-plasminogenzymography to reveal plasminogen activators. Those cellswere found to secrete uPA, but not the tissue-type plasmino-gen activator (tPA), in a time-dependent manner (Figure 4A).Following 6 h in culture, in absence of kE, the level of uPAwas low; it reached a maximum level at 24 h. At this time,higher molecular weight caseinolytic bands, probably cor-responding to uPA-PAIs complexes, could be also detected[34]. When cells were grown in presence of kE (50 µg/ml),secretion of uPA was considerably accelerated and the max-imum secreted enzyme level was reached as early as 6 h. At

Stimulation of HT-1080 cells invasion by elastin peptides 113
Figure 5. Combined effect of elastin-derived peptides (50 µg/ml) and plasminogen (20 µg/ml) on MMP-1 expression by HT-1080 cells following 6 h(1–4) and 24 h (5–8) of culture. MMP-1 levels were determined in conditionned media by Western blot. 1 and 5: Controls. 2 and 6: kE-treated cells. 3 and7: plasminogen-treated cells. 4 and 8: kE and plasminogen-treated cells.
24 h, the zymographic pattern was similar in absence or inpresence of kE.
We further investigated whether supplementation of cul-ture media with Plgn (20 µg/ml) could generate plasminactivity in kE-treated or untreated HT-1080 cells. Plasminactivity, as determined by S-2251 substrate hydrolysis, wasnot detected in the conditioned media from cells cultured inabsence of plasminogen following 6 and 24 h in culture. Ad-dition of Plgn generated 5.8 mU/105 cells and 21.1 mU/105
cells plasmin activity at 6 and 24 h in culture (Figure 4B).kE alone did not induce plasmin activity. A combinationof both kE and plasminogen led to a transient 13-fold in-crease in plasmin activity. At 24 h, the level of plasminactivity in medium from cells treated with Plgn and kE wassignificantly reduced.
Influence of plasminogen on MMP activation in control andkE-treated HT-1080 cells
It is now well documented that generation of active plasminis a prerequisite for proMMP-1 activation in different celltypes and cell culture systems [2, 34]. As shown in Figure 5,supplementation of culture media from HT-1080 cells with20 µg/ml Plgn led to conversion of proMMP-1 to active42 kDa enzyme species, following 24 h of culture. Furtheraddition of kE (50 µg/ml) to cells accelerated this process(Figure 5), consistent with the effect of elastin fragments onuPA production and induction of plasmin activity.
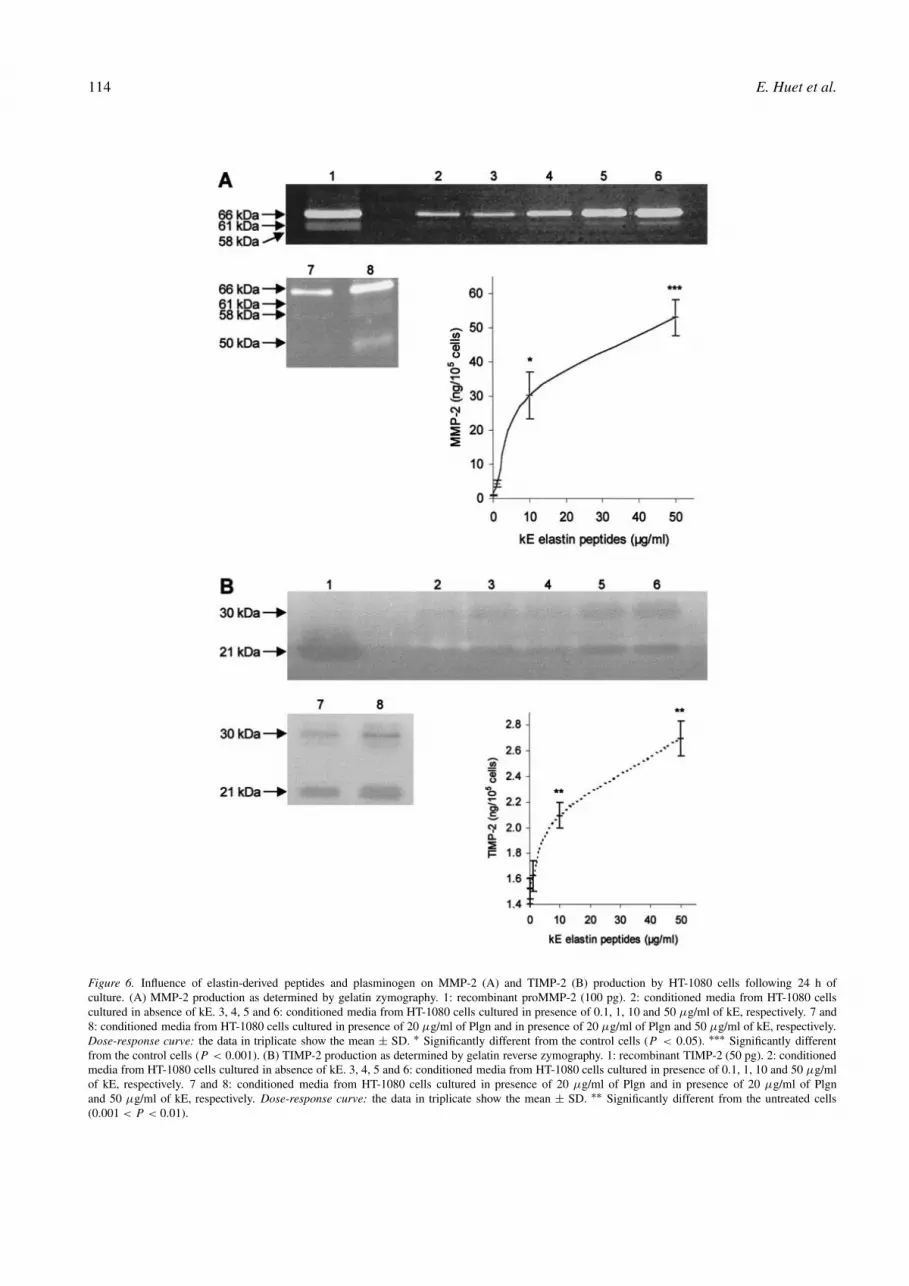
Besides its action on proMMP-1 activation, thePlgn/plasmin system has been involved not only in the sec-ond step of proMMP-2 activation [36], but also in the degra-dation of that enzyme [37]. We therefore analyzed by gelatinzymography the quantitative and qualitative production ofMMP-2 from HT-1080 cells cultured in different conditions,i.e., absence or presence of kE (0.1 to 50 µg/ml) and theabsence or presence of Plgn (20 µg/ml). kE significantlyenhanced the production of MMP-2 in a dose-dependentmanner (Figure 6A). Following 24 h of culture in serum-freeconditions, control and kE (50 µg/ml)-treated cells secreted4.5 and 51 ng of MMP-2, respectively. Plgn, alone, hadonly low influence on MMP-2 level and slightly increasedenzyme activation (Figure 6A, lane 7). Such enzyme acti-vation was particularly evident when HT-1080 cells wereincubated simultaneously with kE and Plgn, where both58 kDa and 61 kDa active MMP-2 species could be iden-tified (Figure 6A, lane 8). In such conditions, the level of
proMMP-2 was reduced by half with appearance of a 50 kDagelatinolytic band, suggesting that generated plasmin hada dual effect on MMP-2, inducing both its activation anddegradation.
Since levels of TIMP-2 were reported to participatenot only in proMMP-2 activation but also in its protectionfrom plasmin degradation [37], we evaluated the varia-tion of secreted TIMP-2 from HT-1080 cells. Here again,elastin-derived peptides were found to stimulate TIMP-2secretion from HT-1080 cells in a dose-dependent manner(Figure 6B).
At 24 h of culture,105 control cells and kE-treated cellssecreted 1.5 ng and 2.7 ng of TIMP-2, respectively. Thus, ona molar basis, the level of MMP-2 exceeded that of inhibitorin kE-treated but not in control cells. Plgn had no signifi-cant influence on TIMP-2 production from either control orelastin-treated HT-1080 cells (Figure 6B, lanes 7 and 8).
Influence of elastin peptides and plasminogen on thecapacity of HT-1080 cells to invade a type I collagen matrix
Under our experimental conditions, untreated HT-1080 cellshad a very low ability to penetrate a type I collagen ma-trix, thus contrasting with the high capacity of those cellsto invade a basement membrane equivalent ([10]; our un-published observation). When 105 HT-1080 cells were incu-bated with either kE (50 µg/ml) or Plgn (20 µg/ml), beforeseeding onto collagen gels, only 309 ± 99 or 141 ± 60 cellswere recovered in the lower compartment of the chambersafter 24 h (Figure 7). When both agents were combinedin conditions that induced plasmin activity and proMMPsactivation, the collagen invasive potential of HT-1080 cellsincreased 5-fold as compared to cells treated with kE alone(Figure 7).
Discussion
The modulation of neutral proteinase expression and acti-vation induced in HT-1080 cells by elastin-derived peptideswas here investigated. We first provided evidence that thosepeptides could stimulate the production of proMMP-1; a2-fold increase in proMMP-1 secretion was reached at aconcentration of elastin peptides, from alkaline or elastasehydrolysates of insoluble elastin, close to those determinedin physiological fluids [38]. This effect could be inhibited by
114 E. Huet et al.
Figure 6. Influence of elastin-derived peptides and plasminogen on MMP-2 (A) and TIMP-2 (B) production by HT-1080 cells following 24 h ofculture. (A) MMP-2 production as determined by gelatin zymography. 1: recombinant proMMP-2 (100 pg). 2: conditioned media from HT-1080 cellscultured in absence of kE. 3, 4, 5 and 6: conditioned media from HT-1080 cells cultured in presence of 0.1, 1, 10 and 50 µg/ml of kE, respectively. 7 and8: conditioned media from HT-1080 cells cultured in presence of 20 µg/ml of Plgn and in presence of 20 µg/ml of Plgn and 50 µg/ml of kE, respectively.Dose-response curve: the data in triplicate show the mean ± SD. ∗ Significantly different from the control cells (P < 0.05). ∗∗∗ Significantly differentfrom the control cells (P < 0.001). (B) TIMP-2 production as determined by gelatin reverse zymography. 1: recombinant TIMP-2 (50 pg). 2: conditionedmedia from HT-1080 cells cultured in absence of kE. 3, 4, 5 and 6: conditioned media from HT-1080 cells cultured in presence of 0.1, 1, 10 and 50 µg/mlof kE, respectively. 7 and 8: conditioned media from HT-1080 cells cultured in presence of 20 µg/ml of Plgn and in presence of 20 µg/ml of Plgnand 50 µg/ml of kE, respectively. Dose-response curve: the data in triplicate show the mean ± SD. ∗∗ Significantly different from the untreated cells(0.001 < P < 0.01).

Stimulation of HT-1080 cells invasion by elastin peptides 115
Figure 7. Influence of plasminogen (Plgn, 20 µg/ml) and elastin-derivedpeptides (kE, 50 µg/ml) on type I collagen invasive capacity of HT-1080cells. �: Control; �: kE-treated cells. The data in triplicate show the mean± SD. ∗ Significantly different from the control cells (0.01 < P < 0.05).∗∗ Significantly different from the control cells (0.001 < P < 0.01).
lactose and reproduced at even higher extent by tropoelastinand VGVAPG peptides, thus indicating the involvement ofEBP in triggering proMMP-1 production. We have recentlyshown that the type VIII beta-turn conformation adopted byelastin peptides with GXXPG sequence was essential for in-ducing MMP-1 expression in fibroblasts [33]. The criticalimportance of the first glycine residue in that pentapeptidesequence was here confirmed as well as the non-absolute re-quirement of a N-terminal valine in the VGVAPG hexapep-tide sequence repeated several fold in human tropoelastin.It must be underlined that GXXPG consensus sequence isalso present in several other matrix proteins including fi-bronectin (GLTPG), tenascin C (GLAPG), thrombospondin-2 (GNTPG), fibrillin-2 (GGIPG) as well as in several typesof collagen suggesting that proteolysis of those macro-molecules and liberation of peptides bearing that sequencecould also participate in overproduction of MMP-1 by EBP-expressing cancer cells. Thus, a specific sequence found inseveral ECM molecules might induce MMP-1 production bytumor cells, further facilitating stromal invasion.
In HT-1080 cells, binding of elastin peptides to EBP didnot modify MMP-1 mRNA steady state levels. By contrast,EBP occupancy by elastin-derived peptides was shown toincrease both MMP-1 and MMP-3 mRNA steady state lev-els in cultured fibroblasts [33]. MMP-3 was undetected inHT-1080 cells treated or not with elastin peptides. The sig-nal pathway propagation leading to induction of MMPs byelastin in fibroblasts was found analogous to that previouslydescribed in human leukocytes where the elastin receptorwas coupled to phospholipase C through a pertussis toxinsensitive GTP binding G protein and further protein kinase Cactivation [16]. Such discrepancies between normal and can-cer cells in the mechanism leading to MMP-1 overproduc-tion is unclear but might involve differences in the generalorganization of the elastin receptor system. Indeed, in fibrob-lasts, EBP interacts with two membrane-associated proteins,a 61 kDa protein with neuraminidase activity and a 55 kDagalactosidase-binding protein which has been reported to beinvolved in transducing signal events [12]. Although EBP
was evidenced in the plasma membrane preparations of HT-1080 cells, those cells overexpressed the 61 kDa proteinmoiety but the 55 kDa galactosidase binding protein wasundetected [20]. Moreover, in agreement with our previ-ous data, interaction between elastin-derived peptides andEBP stimulated MMP-2 expression both at the protein andmRNA levels in HT-1080 cells, and adding actinomycin D,an antibiotic which blocks RNA transcription, suppressedthis effect. In presence of actinomycin D, the amount ofMMP-1 production was not modified after interaction be-tween elastin peptides and EBP. All together, these datasuggested that a post-transcriptional mechanism is involvedin the elastin-derived peptides-mediated overproduction ofMMP-1 by HT-1080 cells.
While exhibiting major effect on MMP-1 production,elastin peptides had no influence on enzyme activation.However, in presence of plasminogen, conditioned mediafrom kE-treated HT-1080 cells developed plasmin activity asearly as following 6 h of culture in serum-free conditions andsuch generation of plasmin activity paralleled processing ofproMMP-1 to a 42 kDa activated enzyme. Elastin peptideswere also shown to increase urokinase secretion by HT-1080cells. Similarly, Lim et al. [39] have found that treatmentof cancer cell lines with phorbol ester led to a temporaryincrease of uPA expression which further declined later inculture with a concomitant enhancement of plasminogen ac-tivator inhibitors, i.e., PAI-1 and PAI-2. The Plgn/plasminsystem has been reported to intervene in proMMP-2 process-ing as well as in MMP-2 and TIMP-2 degradations [36, 37,40]. Implication of plasmin in the conversion of proMMP-2to mature MMP-2 was here similarly observed; such en-zyme processing could be amplified in the presence of kE. Inthe extracellular milieu, plasmin might also exhibit differentrole acting as a degrading enzyme for both proMMP-2 andTIMP-2. Cleavage of proMMP-2 by plasmin was reported tobe inhibited by equimolar concentrations of proMMP-2 andTIMP-2 [37]. Although kE was found to increase TIMP-2 aswell as MMP-2 expression, the level of the enzyme exceededthe amount of its inhibitor, on a molar basis. Thus, plas-min activity was found to decrease pro-MMP-2 level withgeneration of a faint 50-kDa gelatinolytic species and otherMMP-2 degradation fragments, which can be identified onlyby Western blot analysis (not shown).
Since elastin-derived peptides induced MMP expressionand activation, their effect on the capacity of HT-1080 cellsto invade a type I collagen matrix was investigated. In ab-sence of elastin peptides, HT-1080 exhibited a very lowcapacity to penetrate a type I collagen matrix (0.4�of thetotal loaded cell). In presence of elastin peptides, the inva-sive capacity of cells was significantly enhanced (3.1� ofloaded cell invade collagen matrix). Stimulation of MMP-2and MT1-MMP expression by elastin peptides in HT-1080cells was accompanied by MMP-2 maturation to fully activeenzyme at the cell plasma membrane [20] which paralleledan increased capacity of cells to invade Matrigel, a recon-stituted basement membrane equivalent (our unpublisheddata). Since treatment of cells with elastin peptides did notlead to MMP-1 activation, kE-mediated invasion of type I

116 E. Huet et al.
collagen by HT-1080 cells was probably directed by theMT1-MMP/MMP-2 collagenolytic system. Moreover, plas-minogen was found to act in a cumulative manner withelastin peptides in the production and activation of MMP-2and MMP-1 activation cascades. Consequently, the invasivecapacity of elastin and plasminogen-treated HT-1080 cellstoward a type I collagen matrix was significantly increased(36-fold) as compared to cells treated with elastin (8-fold) orplasminogen alone (4-fold). Each component of the MMP-2 cascade (either MT1-MMP or MMP-2) or MMP-1 couldcontribute to such an enhanced type I collagen invasive ca-pacity of fibrosarcoma cells. Also, since elastin peptideswere found to induce MMP-3 expression in fibroblasts [33],host-cells could further participate in proMMP-1 overacti-vation and matrix invasion by cancer cells expressing theelastin receptor.
To summarize, initial elastolysis by serine elastases frominflammatory cells or several members of the MMP familyin elastin-rich tissues such as lung, skin, and breast, must beconsidered as a main catalytic event in tumor progression.
Acknowledgements
This work was supported by funds from the Prefecturede Champagne-Ardenne, a grant from ARERS (Reims-Champagne Ardenne), CNRS (FRE 2260) and INSERM(U514). The authors thank Mrs M. Decarme for her skillfultechnical assistance.
References
1. Stetler-Stevenson WG, Hewitt R, Corcoran M. Matrix metallopro-teinases and tumor invasion: From correlation and causality to theclinic. Semin Cancer Biol 1996; 7 (3): 147–54.
2. DeClerck YA, Laug WE. Cooperation between matrix metallopro-teinases and the plasminogen activator-plasmin system in tumorprogression. Enzyme Protein 1996; 49 (1–3): 72–84.
3. Basset P, Okada A, Chenard MP et al. Matrix metalloproteinasesas stromal effectors of human carcinoma progression: Therapeuticimplications. Matrix Biol 1997; 15 (8–9): 535–41.
4. Chambers AF, Matrisian LM. Changing views of the role of matrixmetalloproteinases in metastasis. J Natl Cancer Inst 1997; 89 (17):1260–70.
5. Stamenkovic II. Matrix metalloproteinases in tumor invasion andmetastasis. Semin Cancer Biol 2000; 10 (6): 415–33.
6. DeClerck YA. Interactions between tumour cells and stromal cellsand proteolytic modification of the extracellular matrix by metallo-proteinases in cancer. Eur J Cancer 2000; 36 (10): 1258–68.
7. Reich R, Blumenthal M, Liscovitch M. Role of phospholipase D inlaminin-induced production of gelatinase A (MMP-2) in metastaticcells. Clin Exp Metastasis 1995; 13 (2): 134–40.
8. Deryugina EI, Bourdon MA, Reisfeld RA et al. Remodeling of colla-gen matrix by human tumor cells requires activation and cell surfaceassociation of matrix metalloproteinase-2. Cancer Res 1998; 58 (16):3743–50.
9. Stanton H, Gavrilovic J, Atkinson SJ et al. The activation of ProMMP-2 (gelatinase A) by HT1080 fibrosarcoma cells is promoted by cultureon a fibronectin substrate and is concomitant with an increase inprocessing of MT1-MMP (MMP-14) to a 45 kDa form. J Cell Sci1998; 111 (18): 2789–98.
10. Maquoi E, Frankenne F, Noel A et al. Type IV collagen induces matrixmetalloproteinase 2 activation in HT1080 fibrosarcoma cells. Exp CellRes 2000; 261 (2): 348–59.
11. Privitera S, Prody CA, Callahan JW et al. The 67-kDa enzymaticallyinactive alternatively spliced variant of beta-galactosidase is identicalto the elastin/laminin-binding protein. J Biol Chem 1998; 273 (11):6319–26.
12. Hinek A. Biological roles of the non-integrin elastin/laminin receptor.Biol Chem 1996; 377 (7–8): 471–80.
13. Blood CH, Zetter BR. Membrane-bound protein kinase C modulatesreceptor affinity and chemotactic responsiveness of Lewis lung carci-noma sublines to an elastin-derived peptide. J Biol Chem 1989; 264(18): 10614–20.
14. Kamoun A, Landeau JM, Godeau G et al. Growth stimulation of hu-man skin fibroblasts by elastin-derived peptides. Cell Adhes Commun1995; 3 (4): 273–81.
15. Jacob MP, Fulop T, Jr., Foris G et al. Effect of elastin peptides onion fluxes in mononuclear cells, fibroblasts, and smooth muscle cells.Proc Natl Acad Sci USA 1987; 84 (4): 995–9.
16. Varga Z, Jacob MP, Robert L et al. Identification and signal trans-duction mechanism of elastin peptide receptor in human leukocytes.FEBS Lett 1989; 258 (1): 5–8.
17. Yusa T, Blood CH, Zetter BR. Tumor cell interactions with elastin:implications for pulmonary metastasis. Am Rev Respir Dis 1989; 140(5): 1458–62.
18. Svitkina TM, Parsons DF. Binding of some metastatic tumor cell linesto fibrous elastin and elastin peptides. Int J Cancer 1993; 53 (5): 824–8.
19. Timar J, Lapis K, Fulop T et al. Interaction between elastin and tu-mor cell lines with different metastatic potential; in vitro and in vivostudies. J Cancer Res Clin Oncol 1991; 117 (3): 232–8.
20. Brassart B, Randoux A, Hornebeck W et al. Regulation of ma-trix metalloproteinase-2 (gelatinase A, MMP-2), membrane-typematrix metalloproteinase-1 (MT1-MMP) and tissue inhibitor ofmetalloproteinases-2 (TIMP-2) expression by elastin-derived peptidesin human HT-1080 fibrosarcoma cell line. Clin Exp Metastasis 1998;16 (6): 489–500.
21. Aimes RT, Quigley JP. Matrix metalloproteinase-2 is an interstitialcollagenase. Inhibitor-free enzyme catalyzes the cleavage of collagenfibrils and soluble native type I collagen generating the specific 3/4-and 1/4-length fragments. J Biol Chem 1995; 270 (11): 5872–6.
22. Hotary K, Allen E, Punturieri A et al. Regulation of cell invasionand morphogenesis in a three-dimensional type I collagen matrix bymembrane-type matrix metalloproteinases 1, 2, and 3. J Cell Biol2000; 149 (6): 1309–23.
23. Murphy G, Stanton H, Cowell S et al. Mechanisms for pro matrixmetalloproteinase activation. Apmis 1999; 107 (1): 38–44.
24. Suzuki K, Enghild JJ, Morodomi T et al. Mechanisms of activationof tissue procollagenase by matrix metalloproteinase 3 (stromelysin).Biochemistry 1990; 29 (44): 10261–70.
25. Deryugina EI, Ratnikov B, Monosov E et al. MT1-MMP initiates ac-tivation of pro-MMP-2 and integrin alphavbeta3 promotes maturationof MMP-2 in breast carcinoma cells. Exp Cell Res 2001; 263 (2):209–23.
26. Zigrino P, Drescher C, Mauch C. Collagen-induced proMMP-2 acti-vation by MT1-MMP in human dermal fibroblasts and the possiblerole of alpha2beta1 integrins. Eur J Cell Biol 2000; 80 (1): 68–77.
27. Berton A, Lorimier S, Emonard H et al. Contribution of the plas-min/matrix metalloproteinase cascade to the retraction of humanfibroblast populated collagen lattices. Mol Cell Biol Res Commun2000; 3 (3): 173–80.
28. Jacob MP, Hornebeck W. Isolation and characterization of insolubleand kappa-elastin. Frontiers Matrix Biology 1985; 10: 92–129.
29. Samouillan V, Lamure A, Maurel E et al. Alterations in the chaindynamics of insoluble elastin upon proteolysis by serine elastases.Biopolymers 2001; 58 (2): 175–85.
30. Lam KS, Salmon SE, Hersh EM et al. A new type of synthetic pep-tide library for identifying ligand-binding activity. Nature 1991; 354(6348): 82–4.
31. Houghten RA, Pinilla C, Blondelle SE et al. Generation and use ofsynthetic peptide combinatorial libraries for basic research and drugdiscovery. Nature 1991; 354 (6348): 84–6.
32. Polette M, Gilles C, de Bentzmann S et al. Association of fibroblastoidfeatures with the invasive phenotype in human bronchial cancer celllines. Clin Exp Metastasis 1998; 16 (2): 105–12.

Stimulation of HT-1080 cells invasion by elastin peptides 117
33. Brassart B, Fuchs P, Huet E et al. Conformational dependence of col-lagenase (matrix metalloproteinase-1) up-regulation by elastin pep-tides in cultured fibroblasts. J Biol Chem 2001; 276 (7): 5222–5227.
34. Lijnen HR. Molecular interactions between the plasminogen/plasminand matrix metalloproteinase system. Fibrinolysis & Proteolysis2000; 14 (2–3): 175–181.
35. Unemori EN, Bair MJ, Bauer EA et al. Stromelysin expression regu-lates collagenase activation in human fibroblasts. Dissociable controlof two metalloproteinases by interferon-gamma. J Biol Chem 1991;266 (34): 23477–82.
36. Baramova EN, Bajou K, Remacle A et al. Involvement of PA/plasminsystem in the processing of pro-MMP-9 and in the second step ofpro-MMP-2 activation. FEBS Lett 1997; 405 (2): 157–62.
37. Farina AR, Tacconelli A, Teti A et al. Tissue inhibitor ofmetalloproteinase-2 protection of matrix metalloproteinase-2 from
degradation by plasmin is reversed by divalent cation chelator EDTAand the bisphosphonate alendronate. Cancer Res 1998; 58 (14): 2957–60.
38. Fulop T, Jr, Wei SM, Robert L et al. Determination of elastin peptidesin normal and arteriosclerotic human sera by ELISA. Clin PhysiolBiochem 1990; 8 (6): 273–82.
39. Lim YT, Sugiura Y, Laug WE et al. Independent regulation of matrixmetalloproteinases and plasminogen activators in human fibrosar-coma cells. J Cell Physiol 1996; 167 (2): 333–40.
40. Maquoi E, Frankenne F, Baramova E et al. Membrane type 1 matrixmetalloproteinase-associated degradation of tissue inhibitor of metal-loproteinase 2 in human tumor cell lines. J Biol Chem 2000; 275 (15):11368–78.