crystal structure of a mutant elongation factor g trapped with a gtp analogue
TRANSCRIPT

FEBS Letters 579 (2005) 4492–4497 FEBS 29808
Crystal structure of a mutant elongation factor G trappedwith a GTP analogue
Sebastian Hanssona, Ranvir Singha, Anatoly T. Gudkovb, Anders Liljasa, Derek T. Logana,*
a Department of Molecular Biophysics, Lund University, Box 124, S-211 00 Lund, Swedenb Institute of Protein Research, Russian Academy of Sciences, 142292 Puschino, Moscow Region, Russia
Received 16 May 2005; revised 21 June 2005; accepted 6 July 2005
Available online 27 July 2005
Edited by Irmgard Sinning
Abstract Elongation factor G (EF-G) is a G protein factor thatcatalyzes the translocation step in protein synthesis on the ribo-some. Its GTP conformation in the absence of the ribosome iscurrently unknown. We present the structure of a mutantEF-G (T84A) in complex with the non-hydrolysable GTP ana-logue GDPNP. The crystal structure provides a first insight intoconformational changes induced in EF-G by GTP. Comparisonof this structure with that of EF-G in complex with GDP sug-gests that the GTP and GDP conformations in solution are verysimilar and that the major contribution to the active GTPaseconformation, which is quite different, therefore comes from itsinteraction with the ribosome.� 2005 Federation of European Biochemical Societies. Publishedby Elsevier B.V. All rights reserved.
Keywords: Translation; Protein synthesis; Elongation factor G;Translocation; Conformational change; GDPNP complex
1. Introduction
Elongation factors Tu and G (EF-Tu and EF-G) are the cat-
alysts in the elongation phase of bacterial protein biosynthesis.
They interact alternately with the ribosome [1,2]. EF-Tu:GTP
delivers an aminoacyl tRNA (aa-tRNA) to the aminoacyl site
(A-site). After ribosome-catalysed peptide bond formation,
EF-G in complex with GTP catalyses the translocation step,
in which the peptidyl-tRNA carrying the nascent polypeptide
in the A-site and the deacylated tRNA in the peptidyl site
(P-site) are moved to the P and exit (E) sites, respectively, with
concomitant advance of the mRNA by one codon. EF-G in
complex with GDP dissociates from the ribosome, which is
now ready for next round of elongation. Both EF-G and
EF-Tu belong to the GTPase superfamily [3]. The GTPase
activity of EF-Tu and EF-G is dramatically enhanced by the
ribosome, which is their effector molecule in its post- and pre-
translocational states (postT and preT), respectively [4,5].
GTPases undergo guanine nucleotide-dependent conforma-
tional changes [6]. While the inactive GDP states of GTPases
are structurally divergent, their active GTP states have very
similar conformations around the GTP binding site. So far,
Abbreviations: EF-TU, elongation factor Tu; EF-G, elongation factorG; aa-tRNA, aminoacyl-tRNA; SAXS, small angle X-ray scattering;cryo-EM, cryo-electron microscopy
*Corresponding author. Fax: +46 46 222 4692.E-mail address: [email protected] (D.T. Logan).
0014-5793/$30.00 � 2005 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2005.07.016
only the GDP and apo conformations of EF-G ‘‘off the ribo-
some’’ have been structurally characterized by X-ray crystal-
lography. Small angle X-ray scattering (SAXS) experiments
suggest that the overall solution conformation of EF-G in its
different states (apo, GDP and GTP) does not change as dra-
matically as does that of EF-Tu [7]. In contrast, according to
cryo-electron microscopy (cryo-EM), EF-G:GTP has a dra-
matically altered conformation on the ribosome compared to
the crystal structures [8–10]. The structure of the GTP confor-
mation of EF-G off the ribosome is therefore of great interest,
as it would reveal how much conformational change is induced
by the nucleotide itself and how much by binding to the ribo-
some. Here we present the crystal structure of the EF-G mu-
tant T84A from Thermus thermophilus in complex with
GDPNP, which shows only modest conformational changes
relative to the GDP structure. The dramatically different con-
formation of EFG:GTP seen on the ribosome must therefore
be induced by its interactions with the latter. In a previous
study we have suggested the possible importance of the ‘‘P-
loop lysine’’ Lys25 and its interactions for the affinity of the
nucleotide [11]. In light of these results, attempts to co-crystal-
lize EF-G with a GTP analogue were undertaken. Trials were
made with wild type and both the G16V and T84A mutants
with several different GTP analogues, although only the
T84A mutant resulted in a structure with GDPNP. The pres-
ent crystal structure once more suggests that one important
conformational change undergone by EF-G upon binding to
the ribosome may involve Lys25 and that this may be one
important component that discriminates the conformational
cycles of EF-G and its eukaryotic counterpart eEF2 from
those of other ribosomal GTPases.
2. Materials and methods
2.1. CrystallizationCloning, expression and purification of T. thermophilus EF-G was
performed as previously described [11]. The T84A mutant was crystal-lized in the presence of 10 mM GDPNP (Sigma�). 5 ll of 100 mMGDPNP was treated with 2 U of shrimp alkaline phosphatase (Amer-sham) at room temperature for 30 min prior to addition to the proteinsolution, in order to remove traces of GDP. Crystals grew from hang-ing drops by vapour diffusion at precipitant conditions identical tothose producing T84A:GDP crystals [11]. 4 ll of a 4 mg/ml proteinsolution was mixed with 4 ll of reservoir solution. The drops werestreak-seeded using T84A:GDP at setup. Crystals resulting from theseexperiments were too small for data collection but were used to streak-seed further drops. Crystals were of suitable size for data collection(0.1 · 0.3 · 0.3 mm) after two days. Preparation of the crystals forcryogenic data collection was as previously described [11].
blished by Elsevier B.V. All rights reserved.

S. Hansson et al. / FEBS Letters 579 (2005) 4492–4497 4493
2.2. Data collection and structure determinationDiffraction data were collected at 100 K to 2.5 A resolution at beam-
line I711 of the MAX-II synchrotron, Lund, Sweden. The data wereintegrated with XDS [12] and further processed with CCP4 programs[13,14]. The structure was solved using the T84A:GDP structure [11]as search model in Molrep [15]. The model was built using XtalView[16] and refined with Refmac5, first using rigid body refinement thenatomic refinement [17] with each domain as a separate TLS group[18]. Density was found at the expected nucleotide binding site andGDP was incorporated into the model. As extra density was visibleat the supposed c-phosphate site (Fig. 1A), GDP was replaced by
Fig. 1. (A) The positive electron density (green) seen for thec-phosphate of the nucleotide in the SIGMAA-weighted m|Fo| � D|Fc|map at a contour level of 3r. The 2m|Fo| � D|Fc| map (blue) comesfrom T84A:GDP, and is contoured at 1r. The figure also shows thepeptide flip between Ile21 and Asp22 of T84A:GDPNP (cyan) andT84A:GDP. (B) Electron density in the SIGMAA-weighted2m|Fo| � D|Fc| map around the nucleotide in the GDPNP structure,also contoured at 1r.
GDPNP. Figures were made using Pymol (http://www.pymol.org).Structural superimpositions were made using SwissPDB Viewer. Radiiof gyration were calculated using MOLEMAN2.
3. Results
Good quality crystals of T84A:GDPNP appeared after ini-
tial streak seeding with fresh T84A:GDP crystals and subse-
quent optimization by streak seeding with T84A:GDPNP
crystals. As for all other EF-G structures the space group is
P212121 [11,19–22]. Interestingly the cell dimensions for
T84A:GDPNP differ from those of T84A:GDP by roughly
16 A in the b-axis (Table 1) and are more similar to those of
native EF-G, despite initial seeding from T84A:GDP crystals.
Electron density maps clearly show extra density at the c-phosphate position, approximately 1.6 A away from the oxy-
gen of the b-phosphate (Fig. 1A). Refining a GDPNP molecule
at occupancy of 0.8 leaves no significant positive or negative
density around the phosphates in the difference map
(Fig. 1B). The position of the c-phosphate group is in agree-
ment with that seen in the GTP analogue complexes of e.g.
EF-Tu and IF2/eIF5B [23–25]. As in several other EF-G struc-
tures, switch II is significantly more flexible than the rest of the
molecule, with B-factors in the range 100–130 A2 for residues
86–90 compared to an average of 53 A2 for the whole struc-
ture. Gly86 and the side chain of His87 in switch II are not vis-
ible in the electron density. Switch I is invisible between
residues 43 and 60, and residues 40–41 and 61–65 can only
be built as poly-alanine. The density for domain III is unclear
in many places, as is the case in several other EF-G structures
[19,21,22].
3.1. Domain movements
An overall comparison of T84A:GDPNP with other EF-G
structures shows that T84A:GDPNP is more similar to the
wild-type EF-G:GDP than to T84A:GDP. The root mean
square deviation (rmsd) from wild type (PDB entry 1DAR)
Table 1Crystallographic data collection and structure refinement statistics
Space group P212121Cell dimensions (A) a = 75.0, b = 103.3, c = 118.9Data resolution (A) 26.9–2.5 (2.6–2.5)No. of observations 233886No. of unique reflections 30656Completeness (overall/outer shell) (%) 96.5/94.6I/rI (overall/outer shell) 11.5/2.7Rmerge (%) 7.0/44.3
RefinementRmodel (%) 21.8Rfree (%) 29.1No. of non-hydrogen atoms 5038No. of water molecules 158
Rms deviations from ideal geometryBond lengths (A) 0.009Bond angles (�) 1.212
Ramachandran plot (%)Most favored 84.1Additionally allowed 15.4Generously allowed 0.2Disallowed 0.4
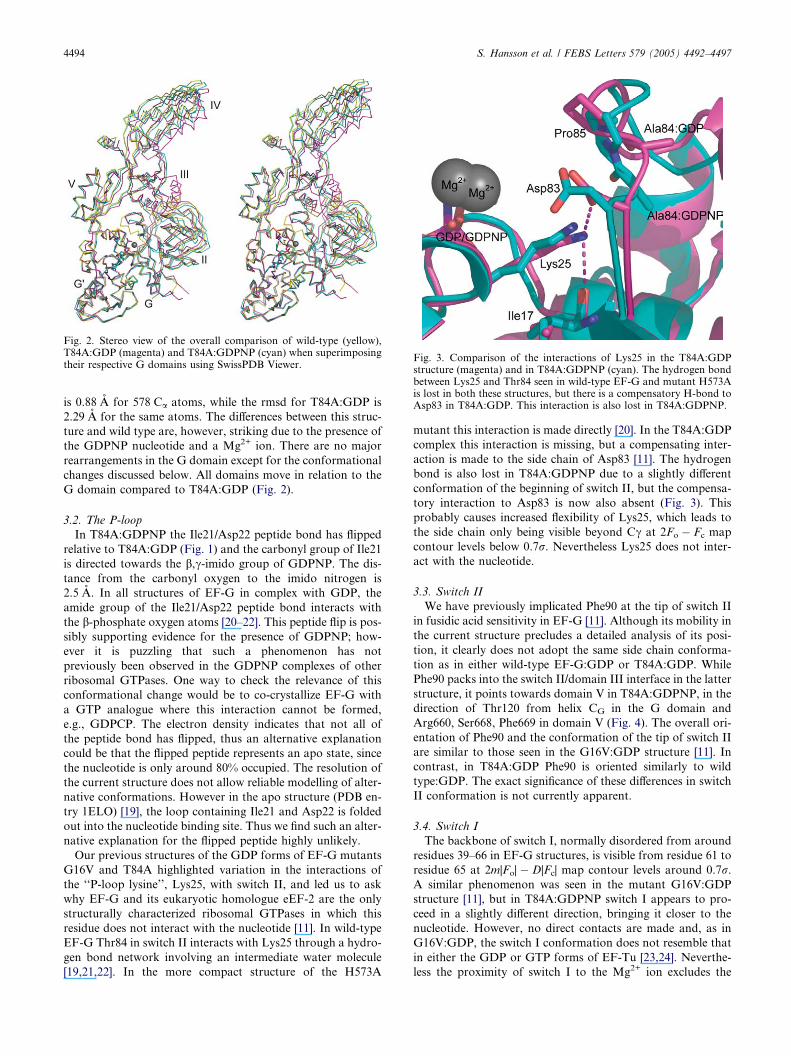
Fig. 2. Stereo view of the overall comparison of wild-type (yellow),T84A:GDP (magenta) and T84A:GDPNP (cyan) when superimposingtheir respective G domains using SwissPDB Viewer.
Fig. 3. Comparison of the interactions of Lys25 in the T84A:GDPstructure (magenta) and in T84A:GDPNP (cyan). The hydrogen bondbetween Lys25 and Thr84 seen in wild-type EF-G and mutant H573Ais lost in both these structures, but there is a compensatory H-bond toAsp83 in T84A:GDP. This interaction is also lost in T84A:GDPNP.
4494 S. Hansson et al. / FEBS Letters 579 (2005) 4492–4497
is 0.88 A for 578 Ca atoms, while the rmsd for T84A:GDP is
2.29 A for the same atoms. The differences between this struc-
ture and wild type are, however, striking due to the presence of
the GDPNP nucleotide and a Mg2+ ion. There are no major
rearrangements in the G domain except for the conformational
changes discussed below. All domains move in relation to the
G domain compared to T84A:GDP (Fig. 2).
3.2. The P-loop
In T84A:GDPNP the Ile21/Asp22 peptide bond has flipped
relative to T84A:GDP (Fig. 1) and the carbonyl group of Ile21
is directed towards the b,c-imido group of GDPNP. The dis-
tance from the carbonyl oxygen to the imido nitrogen is
2.5 A. In all structures of EF-G in complex with GDP, the
amide group of the Ile21/Asp22 peptide bond interacts with
the b-phosphate oxygen atoms [20–22]. This peptide flip is pos-
sibly supporting evidence for the presence of GDPNP; how-
ever it is puzzling that such a phenomenon has not
previously been observed in the GDPNP complexes of other
ribosomal GTPases. One way to check the relevance of this
conformational change would be to co-crystallize EF-G with
a GTP analogue where this interaction cannot be formed,
e.g., GDPCP. The electron density indicates that not all of
the peptide bond has flipped, thus an alternative explanation
could be that the flipped peptide represents an apo state, since
the nucleotide is only around 80% occupied. The resolution of
the current structure does not allow reliable modelling of alter-
native conformations. However in the apo structure (PDB en-
try 1ELO) [19], the loop containing Ile21 and Asp22 is folded
out into the nucleotide binding site. Thus we find such an alter-
native explanation for the flipped peptide highly unlikely.
Our previous structures of the GDP forms of EF-G mutants
G16V and T84A highlighted variation in the interactions of
the ‘‘P-loop lysine’’, Lys25, with switch II, and led us to ask
why EF-G and its eukaryotic homologue eEF-2 are the only
structurally characterized ribosomal GTPases in which this
residue does not interact with the nucleotide [11]. In wild-type
EF-G Thr84 in switch II interacts with Lys25 through a hydro-
gen bond network involving an intermediate water molecule
[19,21,22]. In the more compact structure of the H573A
mutant this interaction is made directly [20]. In the T84A:GDP
complex this interaction is missing, but a compensating inter-
action is made to the side chain of Asp83 [11]. The hydrogen
bond is also lost in T84A:GDPNP due to a slightly different
conformation of the beginning of switch II, but the compensa-
tory interaction to Asp83 is now also absent (Fig. 3). This
probably causes increased flexibility of Lys25, which leads to
the side chain only being visible beyond Cc at 2Fo � Fc map
contour levels below 0.7r. Nevertheless Lys25 does not inter-
act with the nucleotide.
3.3. Switch II
We have previously implicated Phe90 at the tip of switch II
in fusidic acid sensitivity in EF-G [11]. Although its mobility in
the current structure precludes a detailed analysis of its posi-
tion, it clearly does not adopt the same side chain conforma-
tion as in either wild-type EF-G:GDP or T84A:GDP. While
Phe90 packs into the switch II/domain III interface in the latter
structure, it points towards domain V in T84A:GDPNP, in the
direction of Thr120 from helix CG in the G domain and
Arg660, Ser668, Phe669 in domain V (Fig. 4). The overall ori-
entation of Phe90 and the conformation of the tip of switch II
are similar to those seen in the G16V:GDP structure [11]. In
contrast, in T84A:GDP Phe90 is oriented similarly to wild
type:GDP. The exact significance of these differences in switch
II conformation is not currently apparent.
3.4. Switch I
The backbone of switch I, normally disordered from around
residues 39–66 in EF-G structures, is visible from residue 61 to
residue 65 at 2m|Fo| � D|Fc| map contour levels around 0.7r.A similar phenomenon was seen in the mutant G16V:GDP
structure [11], but in T84A:GDPNP switch I appears to pro-
ceed in a slightly different direction, bringing it closer to the
nucleotide. However, no direct contacts are made and, as in
G16V:GDP, the switch I conformation does not resemble that
in either the GDP or GTP forms of EF-Tu [23,24]. Neverthe-
less the proximity of switch I to the Mg2+ ion excludes the

Fig. 4. The conformational changes observed in Phe90 upon compar-ing the T84A:GDP (magenta) and T84A:GDPNP (cyan) structures.From its location in T84A:GDP between domains G and III Phe90moves to be oriented toward Thr120 in helix CG and Arg660 andSer668 in domain V in T84A:GDPNP.
S. Hansson et al. / FEBS Letters 579 (2005) 4492–4497 4495
interaction of Glu295 from a neighboring molecule with the
Mg2+ as seen in other EF-G structures with shorter unit cell
b axis and a visible Mg2+ [11,20].
1 With allowance for the fact that the latter are calculated from Cacoordinates only.
4. Discussion
We have solved the structure of the T84A mutant of T. ther-
mophilus EF-G in complex with the non-hydrolysable GTP
analogue GDPNP. The structure shows an overall conforma-
tion very similar to that of native EF-G in the apo state or
in complex with GDP. No large domain movements are ob-
served. So far the GTP conformation of EF-G has only been
visualized on the ribosome. Cryo-EM reconstructions of
EF-G trapped by GDPNP on translating ribosomes [9,10,26]
reveal large domain movements compared to the crystal struc-
tures [19–22]. It is possible that the T84A:GDPNP structure
does not represent the true GTP conformation of EF-G in
solution due to crystal contacts; however we consider this un-
likely. A strong indicator is that the structure is clearly induced
by the presence of the nucleotide, as its ‘‘wild-type-like’’ con-
formation, with concomitant longer unit cell b axis, was pro-
duced by seeding with T84A:GDP crystals having a different
crystal packing, under conditions identical to those producing
T84A:GDP crystals.
Many attempts have been made to produce a structure of the
GTP conformation of EF-G, both by us and by others. It is
thus puzzling at first glance that the T84A mutation, which re-
sults in 4-fold lower GDPNP affinity compared to wild type
[27], should be the only one to succeed. We currently have
no satisfactory explanation for this, but it could be due to
small technical differences in the way the crystallization exper-
iments were carried out. In any case, the T84A:GDPNP struc-
ture provides evidence that there is little difference between the
GDP and GTP conformations of EF-G in the absence of the
ribosome, as EF-G is clearly capable of binding the GTP ana-
logue without this inducing major conformational changes.
This is in agreement with experiments using small angle X-
ray scattering (SAXS) that demonstrated very little difference
in the radius of gyration of EF-G between the GDP and
GTP states [7]. Interestingly the calculated radii of gyration
of the ‘‘open’’ conformation of EF-G seen in the GDPNP
complex (31.9 A) and that of the more compact conformation
in mutants H573A, T84A:GDP and G16V:GDP (30.5 A) are
much more similar among themselves than they are to that
of EF-G:GDPNP on the ribosome (34.8 A).1 Experiments
with 2-propanol, which stimulates the intrinsic GTPase activ-
ity of E. coli EF-G in solution, show changes in ellipticity indi-
cating tertiary structural changes which may correspond to
domain rearrangement [28]. On the other hand, other data
involving chemical modification of E. coli EF-G have been
interpreted as indicating only local conformational rearrange-
ments at the GTPase site [29].
For EF-G in solution, the GTP affinity has been measured
by most experimenters to be around 1–10 times lower than that
for GDP [27,29,30]. However for both EF-G and the related
ribosomal GTPase RF3, the GTP affinity is significantly ele-
vated when the factor is bound to the ribosome [30,31]. The
classical interpretation has been that, due to the approximately
10-fold higher concentration of GTP in vivo, the EF-G:GDP
and EF-G:GTP forms will be approximately in equilibrium
in solution. The situation is different for EF-Tu, with require-
ment for a guanosine nucleotide exchange factor (GEF), EF-
Ts. However two groups have measured the GDPNP or
GTP affinity of EF-G to be 60 times lower than that of
GDP [27,32]. Based on this ratio and on other experiments,
Ehrenberg and co-workers have proposed, controversially,
that EF-G binds in its GDP form to the ribosome and that
the preT ribosome acts as a GEF for EF-G. In contrast, other
measurements suggest that the EFG:GDP complex has low
affinity for the ribosome [33]. Our structure is compatible with
either the classical hypothesis or that of Ehrenberg et al. Since
the GDP and GTP forms are very similar in solution, the preT
ribosome is able to accept EF-G in either form. What occurs
thereafter is a matter of debate.
Given that the active GTP conformations of switch II are
very much alike in all GTPases [6] it is surprising that there
is no significant ordering of the switch I and II regions in the
T84A:GDPNP structure. In contrast these regions of the
GTP states of EF-Tu and other translational GTPases (tGTP-
ases) are well determined in solution [25,34,35]. Nevertheless
previous crystal structures, for example of IF2/eIF5B, have
shown that switch I is not necessarily involved in the GTP state
of all tGTPases. In IF2/eIF5B, switch I is located at least 13 A
from the c-phosphate moiety with no possibility to reach the
latter [25]. However the EF-G:GDPNP structure is consistent
with a major role for the ribosome in inducing the active
GTPase conformation. How could this take place, thereby
increasing the affinity of EF-G for GTP by a factor of 600
[30]? Various data support the idea that the ‘‘P-loop lysine’’,
Lys25 in EF-G, is involved. We have previously suggested that
a hydrogen bond between Lys25 and Thr84 in switch II may be
an indication that the conformational changes undergone by
EF-G and eEF2 differ from those of other tGTPases [11], in
particular EF-Tu. Thr84 is part of the highly conserved D[T/

Fig. 5. P-loop lysine orientation in ribosomal GTPase structures. IF2(GDP, violet and GDPNP, salmon), aIF2 (GDPNP, light green), SelB(GDP, orange and GDPNP, wheat) and EF-Tu (GDP, light orangeand GDPNP, yellow) belong to group I, where the P-loop lysineinteracts with the nucleotide, while EF-G (GDP, dark green), T84A(GDP, purple and GDPNP, cyan) and eEF2 (slate) belong to group II,where it is locked into an interaction with the beginning of switch II.
4496 S. Hansson et al. / FEBS Letters 579 (2005) 4492–4497
S]PGH motif found in all EF-G/eEF2 sequences (Fig. 6) [3]. In
all other tGTPases for which the GTP and GDP conforma-
tions are known, the P-loop lysine makes contact with both
nucleotides (Fig. 5). In contrast, in all structures of wild-type
EF-G and eEF2 the interaction between the P-loop lysine
and the threonine or serine in switch II is conserved. According
to chemical modification studies [36] one lysine is involved in
nucleotide binding in EF-G. GTP affinity is abolished when
this lysine is modified by pyridoxal phosphate, but this effect
can be eliminated by protecting the lysine through pre-binding
Fig. 6. Sequence alignment of selected P-loop motifs from ribosomalGTPases. Residues in grey are conserved throughout all ribosomalGTPases. * denotes a chemically conserved residue.
of GTP. The K25A mutation in T. thermophilus EF-G de-
creases the GTPase activity (ATG et al., in preparation) and
a similar mutation in EF-Tu (K24E) abolishes the GTP affinity
[37]. This evidence, together with the functional and structural
importance of this universally conserved lysine reported for
several GTPases and ATPases [38,39], seems to imply that
Lys25 is important for the interaction with GTP on the ribo-
some. In order to bind GTP with higher affinity [30] the
Lys25:Thr84 interaction in EF-G may be disturbed such that
Lys25 can interact with the b- and c-phosphates of GTP, most
likely by ribosome-induced conformational changes. Such a
movement of switch II is observed in the eEF1A:eEF1Ba com-
plex during nucleotide exchange [40]. However the role of the
P-loop lysine in nucleotide binding and GTPase activity re-
quires further investigation.
While this manuscript was being evaluated we solved the
structure of the T84A mutant in complex with the closely re-
lated analogue GDPCP. This structure confirms the general
absence of large-scale conformational changes and the lack
of ordering of switch I or II. It does not exhibit the peptide flip
which we have attributed here to the presence of the imido
nitrogen atom. Further details of this structure will be pre-
sented elsewhere.
Acknowledgments:We thank staff at beamline I711 of MAX-lab for ac-cess to beam time. This work was supported by grants from the Swed-ish Research Council (VR), the Crafoord Foundation, the CarlTrygger Foundation and the Royal Swedish Academy of Sciencesto D.L., a grant from VR and EU Grant QLK2-CT-2002-00892 toA.L., and in part by Russian Grant #02-04-48953 from RFBR toA.T.G. Coordinates have been deposited in the Protein Data Bankwith Accession No. 2BV3.
References
[1] Nishizuka, Y. and Lipmann, F. (1966) Comparison of guanosinetriphosphate split and polypeptide synthesis with a purified E. colisystem. Proc. Natl. Acad. Sci. USA 55, 212–219.
[2] Richman, N. and Bodley, J.W. (1972) Ribosomes cannot interactsimultaneously with elongation factors EF Tu and EF G. Proc.Natl. Acad. Sci. USA 69, 686–689.
[3] Bourne, H.R., Sanders, D.A. and McCormick, F. (1991) TheGTPase superfamily: conserved structure and molecular mecha-nism. Nature 349, 117–127.
[4] Spirin, A.S. (1999) Ribosomes, Kluwer Academic/Plenum, NewYork, p. 422.
[5] Miller, D.L. (1972) Elongation factors EF Tu and EF G interactat related sites on ribosomes. Proc. Natl. Acad. Sci. USA 69, 752–755.
[6] Vetter, I.R. and Wittinghofer, A. (2001) The guanine nucleotide-binding switch in three dimensions. Science 294, 1299–1304.
[7] Czworkowski, J. and Moore, P.B. (1997) The conformationalproperties of elongation factor G and the mechanism of trans-location. Biochemistry 36, 10327–10334.
[8] Agrawal, R.K., Penczek, P., Grassucci, R.A. and Frank, J. (1998)Visualization of elongation factor G on the Escherichia coli 70Sribosome: the mechanism of translocation. Proc. Natl. Acad. Sci.USA 95, 6134–6138.
[9] Stark, H., Rodnina, M.V., Wieden, H.J., van Heel, M. andWintermeyer, W. (2000) Large-scale movement of elongationfactor G and extensive conformational change of the ribosomeduring translocation. Cell 100, 301–309.
[10] Valle, M., Zavialov, A., Sengupta, J., Rawat, U., Ehrenberg, M.and Frank, J. (2003) Locking and unlocking of ribosomalmotions. Cell 114, 123–134.
[11] Hansson, S., Singh, R., Gudkov, A.T., Liljas, A. and Logan, D.T.(2005) Structural insights into fusidic acid resistance and sensi-tivity in EF-G. J Mol Biol 348, 939–949.

S. Hansson et al. / FEBS Letters 579 (2005) 4492–4497 4497
[12] Kabsch, W. (1993) Automatic processing of rotation diffractiondata from crystals of initially unknown symmetry and cellconstants. J. Appl. Cryst. 26, 795–800.
[13] Collaborative Computational Project N. (1994) The CCP4 suite:programs for protein crystallography. Acta Crystallogr. D 50,760–763..
[14] Potterton, E., Briggs, P., Turkenburg, M. and Dodson, E. (2003)A graphical user interface to the CCP4 program suite. ActaCrystallogr. D 59, 1131–1137.
[15] Vagin, A. and Teplyakov, A. (2000) An approach to multi-copysearch in molecular replacement. Acta Crystallogr. D 56 (Pt 12),1622–1624.
[16] McRee, D.E. (1999) XtalView/Xfit – a versatile program formanipulating atomic coordinates and electron density. J. Struct.Biol. 125, 156–165.
[17] Murshudov, G.N. (1997) Refinement of macromolecular struc-tures by the maximum-likelihood method. Acta Crystallogr. D 53,240–255.
[18] Winn, M.D., Murshudov, G.N. and Papiz, M.Z. (2003) Macro-molecular TLS refinement in REFMAC at moderate resolutions.Methods Enzymol. 374, 300–321.
[19] Aevarsson, A., Brazhnikov, E., Garber, M., Zheltonosova, J.,Chirgadze, Y., Al-Karadaghi, S., Svensson, L.A. and Liljas, A.(1994) Three-dimensional structure of the ribosomal translocase:elongation factor G from Thermus thermophilus. EMBO J. 13,3669–3677.
[20] Laurberg, M., Kristensen, O., Martemyanov, K.A., Gudkov,A.T., Nagaev, I., Hughes, D. and Liljas, A. (2000) Structure of amutant EF-G reveals domain III and possibly the fusidic acidbinding site. J. Mol. Biol. 303, 593–603.
[21] Al-Karadaghi, S., Aevarsson, A., Garber, M., Zheltonosova, J.and Liljas, A. (1996) The structure of elongation factor G incomplex with GDP: conformational flexibility and nucleotideexchange. Structure 4, 555–565.
[22] Czworkowski, J., Wang, J., Steitz, T.A. and Moore, P.B. (1994)The crystal structure of elongation factor G complexed withGDP, at 2.7 A resolution. EMBO J. 13, 3661–3668.
[23] Kjeldgaard, M., Nissen, P., Thirup, S. and Nyborg, J. (1993) Thecrystal structure of elongation factor EF-Tu from Thermusaquaticus in the GTP conformation. Structure 1, 35–50.
[24] Nissen, P., Kjeldgaard, M., Thirup, S., Polekhina, G., Reshet-nikova, L., Clark, B.F. and Nyborg, J. (1995) Crystal structure ofthe ternary complex of Phe-tRNAPhe, EF-Tu, and a GTP analog.Science 270, 1464–1472.
[25] Roll-Mecak, A., Cao, C., Dever, T.E. and Burley, S.K. (2000) X-ray structures of the universal translation initiation factor IF2/eIF5B: conformational changes on GDP and GTP binding. Cell103, 781–792.
[26] Agrawal, R.K., Heagle, A.B., Penczek, P., Grassucci, R.A. andFrank, J. (1999) EF-G-dependent GTP hydrolysis induces trans-location accompanied by large conformational changes in the 70Sribosome. Nat. Struct. Biol. 6, 643–647.
[27] Martemyanov, K.A., Liljas, A., Yarunin, A.S. and Gudkov, A.T.(2001) Mutations in the G-domain of elongation factor G fromThermus thermophilus affect both its interaction with GTP andfusidic acid. J. Biol. Chem. 276, 28774–28778.
[28] Masullo, M., Parlato, G., De Vendittis, E. and Bocchini, V. (1989)Effect of propan-2-ol on enzymic and structural properties ofelongation factor G. Biochem. J. 261, 725–731.
[29] Kaziro, Y. (1978) The role of guanosine 5 0-triphosphate inpolypeptide chain elongation. Biochim. Biophys. Acta 505, 95–127.
[30] Baca, O.G., Rohrbach, M.S. and Bodley, J.W. (1976) Equilibriummeasurements of the interactions of guanine nucleotides withEscherichia coli elongation factor G and the ribosome. Biochem-istry 15, 4570–4574.
[31] Zavialov, A.V., Buckingham, R.H. and Ehrenberg, M. (2001) Aposttermination ribosomal complex is the guanine nucleotideexchange factor for peptide release factor RF3. Cell 107, 115–124.
[32] Zavialov, A., Hauryliuk, V. and Ehrenberg, M. (2005) Guano-sine-nucleotide exchange on ribosome-bound elongation factor Ginitiates the translocation of tRNAs. J. Biol. 4, 9.
[33] Martemyanov, K.A. and Gudkov, A.T. (2000) Domain III ofelongation factor G from Thermus thermophilus is essential forinduction of GTP hydrolysis on the ribosome. J. Biol. Chem. 275,35820–35824.
[34] Berchtold, H., Reshetnikova, L., Reiser, C.O., Schirmer, N.K.,Sprinzl, M. and Hilgenfeld, R. (1993) Crystal structure of activeelongation factor Tu reveals major domain rearrangements.Nature 365, 126–132.
[35] Polekhina, G., Thirup, S., Kjeldgaard, M., Nissen, P., Lippmann,C. and Nyborg, J. (1996) Helix unwinding in the effector region ofelongation factor EF-Tu-GDP. Structure 4, 1141–1151.
[36] Giovane, A., Balestrieri, C. and Gualerzi, C. (1982) Structure–function relationships in Escherichia coli translationalelongation factor G: modification of lysine residues by the site-specific reagent pyridoxal phosphate. Biochemistry 21, 5224–5230.
[37] Hwang, Y.W., McCabe, P.G., Innis, M.A. and Miller, D.L.(1989) Site-directed mutagenesis of the GDP binding domain ofbacterial elongation factor Tu. Arch. Biochem. Biophys. 274,394–403.
[38] Leipe, D.D., Wolf, Y.I., Koonin, E.V. and Aravind, L. (2002)Classification and evolution of P-loop GTPases and relatedATPases. J. Mol. Biol. 317, 41–72.
[39] Saraste, M., Sibbald, P.R. and Wittinghofer, A. (1990) The P-loop – a common motif in ATP- and GTP-binding proteins.Trends Biochem. Sci. 15, 430–434.
[40] Andersen, G.R., Pedersen, L., Valente, L., Chatterjee, I., Kinzy,T.G., Kjeldgaard, M. and Nyborg, J. (2000) Structural basis fornucleotide exchange and competition with tRNA in the yeastelongation factor complex eEF1A:eEF1Balpha. Mol. Cell 6,1261–1266.