cryptic open reading frames in plasmid vector backbone sequences can provide highly immunogenic...
TRANSCRIPT

[CANCER RESEARCH 58, 3087-3093. July 15, 1998]
Cryptic Open Reading Frames in Plasmid Vector Backbone Sequences Can ProvideHighly Immunogenic Cytotoxic T-Lymphocyte Epitopes1
Thorbald van Hall, Nastasja E. van de Rhee, Stephen P. Schoenberger, Michel P. M. Vierboom,Frank A. W. Verreck, Cornelis J. M. Melief, and Rienk Offringa2
Department of Inimunohematology and Blood Bank. Leiden Universi^ Medical Center, 2300 RC Leiden, the Netherlands
ABSTRACT
Murine tumor cells obtained through transfection of expression plas-
mids carrying activated cellular and/or viral oncogenes constitute formidable tools for immunological tumor research. As reported previously,mouse embryo cells of C57BL/6 origin, transformed by mutated p53 orhuman papilloma virus type 16 (HPV16), present, at their surface, MHC-bound peptides that are derived from the p53 and the HPV16 E7 onco-
proteins, respectively, which can serve as a target for a highly effectiveantitumor T-cell response. Here, we describe the identification, through
molecular cloning, of an additional, highly immunodominant peptide thatis presented by the aforementioned HPV16- and p53-transformed cells.
This peptide is encoded by a cryptic open reading frame in the backbonesequences of the plasmids that had been used to generate these cells.Considerable amounts of transcripts encompassing this open readingframe were detected in the cells concerned. These transcripts were theresult of the bidirectional nature of the retroviral long terminal repeat(LTR) present in the expression plasmids used for transfection, whichresulted in transcription of the gene of interest, as well as in transcriptionof the vector sequences positioned at the other side of the LTR. Due to thismechanism, all tumor cells harboring LTR-driven expression plasmids
expressed the highly immunogenic peptide, whereas cells containing plasmids driven by more unidirectional promoters exhibited lower levels ofthis peptide. LTR-driven expression plasmids were also shown to encode
this peptide epitope when used for DNA vaccination, as mice vaccinatedwith such a plasmid developed a ('II. response against this peptide. Our
data show that awareness of plasmid backbone-derived epitopes is of
crucial importance for the correct interpretation of preclinical experiments and for the design of DNA vaccines.
INTRODUCTION
Murine tumor models have proven to be of great value for thedetailed analysis of tumor-specific T cell responses and for the development of T cell-based immunotherapeutic strategies against cancer (reviewed in Refs. 1-4). Clear advantages of such murine modelsare the well-defined genetic background of mice, the possibility of
generating and modifying syngeneic tumor cell lines, and the opportunity to test the efficacy of immune interventions in vivo. Mostroutinely studied murine tumors have been obtained through inductionwith potent doses of chemical or physical carcinogens, through infection with oncogenic viruses or through transfection of oncogenes intoprimary cells.
Identification at the molecular level of MHC-bound immunogenicpeptides representing target antigens for tumor-specific T cells was
first performed with cells that express viral oncogenes. Cumulativeresearch showed that almost all T-cell epitopes defined thus far onvirus-induced tumor cells were of viral origin (5). For instance.
Received 2/18/98; accepted 5/18/98.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1This study was financed by the Netherlands Cancer Foundation Grant RUL 94-854
(to T. v. H. and N. E. v. d. R.).2 To whom requests for reprints should be addressed, at Leiden University Medical
Center, Department of Immunohematology and Blood Bank. P.O. Box 9600. 2300 RCLeiden, the Netherlands. Phone: 31-71-5263845; Fax: 31-71-5216751.
MECs3 of B6 origin that were transformed by HPV16 were shown toexpress a H-2Db-bound peptide derived from the HPV16 E7 protein
(RAHYNIVTF; Refs. 6 and 7). CTLs that were capable of eradicatingthe HPV 16-transformed cells in vitro as well as in vivo could be
isolated from mice that had been vaccinated with this peptide. Moreover, vaccination of mice with the HPV16E7 peptide induced strong,protective CTL immunity against a subsequent challenge with atumorigenic dose of the HPV 16-transformed cells (6). The CTLepitopes expressed by the carcinogen-induced murine P815 mastocy-
toma and its immunogenic mutagenized variants were the first examples of epitopes that were encoded by cellular genes. The identification through molecular cloning of these epitopes and the cognategenes marked both a technical and conceptual breakthrough, whichboosted the identification of multiple tumor-associated T-cell epitopes
that were presented by tumors of murine and human origin (reviewedin Ref. 2). Such T-cell epitopes of nonviral origin can be encoded bycell lineage-specific (e.g., melanocyte-specific) genes, by genes that
are not expressed by normal somatic tissues but are induced as a resultof the transformation process (e.g., MAGE family of antigens), or bythe mutated or ectopically expressed oncogenes themselves. Recently,we described an example of the latter category, a H-2Kh-bound
peptide derived from the wild-type sequence of murine p53, which is
presented by B6 MECs transformed by mutated murine p53 andactivated Ras. CTLs against this peptide, raised in p53 knockout mice,were shown to eradicate p53-overexpressing tumor cells in p53+/+
mice in the absence of any detectable immunopathological damage tothe normal p53+/+ tissues (8).
Here, we describe the identification of a novel highly immunogenicCTL epitope, which is presented by the above-mentioned HPV 16- andp53- transformed cells in addition to the HPV16E7 and p53-derivedpeptides that have already been defined. With respect to the HPV 16-
transformed cells, an as yet undefined epitope was previously reportedto be immunodominant over the viral, HPV16-derived epitope (7),whereas the p53-transformed cells express an unknown epitope that isat least codominant to the p53-derived peptide in p53 knockout mice.4
Because these CTL epitopes were found to play a major role in theT-cell immune response against both tumors, we set out to identify the
nature of these peptides. We now present data indicating that thehighly immunogenic epitopes presented by the HPV 16- and p53-transformed cells are, in fact, the same H-2Db-bound peptide. Iden
tification of the gene encoding this peptide through molecular cloningrevealed that the epitope was encoded by a cryptic ORF that waspresent in the backbone sequences of the expression plasmids that hadbeen used to generate the HPV 16- and p53-transformed cells through
DNA transfection. Importantly, these expression plasmids all contained a retroviral LTR, the bidirectional nature of which was foundto be instrumental in the expression of significant levels of plasmidbackbone-derived transcripts by the cells. The implications of these
3 The abbreviations used are: MEC, mouse embryo cell; B6. C57BL/6; HPV 16, human
papilloma virus type 16; LTR, long terminal repeat; MuLV, murine leukemia virus; TCR.T-cell receptor; TNF, tumor necrosis factor; HPLC. high-pressure liquid chromatography;
Ad, adenovirus; RSV. Rous sarcoma virus; ORF, open reading frame; aa, amino acid(s);CMV. cytomegalovirus.
4 Unpublished observations.
3087
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

PLASM») BACKBONE-DERIVED CTL EPITOI'l S
findings for the use of plasmid-transfected tumor cells in immunization studies, as well as for the use of LTR-driven plasmids for DNA
vaccination, are discussed.
MATERIALS AND METHODS
Cell Lines. Primary MECs of B6. B10.A (4R). and B10.A (5R) origin wereprepared as described elsewhere (9). After two to four passages in vitro.primary MECs were transtected with expression vectors containing activatedcellular oncogenes using the calcium phosphate precipitation method ( 10). Theoncogene expression constructs that were used for transfection arepLTRp53cG (Val-135 mutation; Réf.Il), pRSV-H-ras (Lys-12 mutation: Ref.12), and pRSV-v-fos (Phe-15, Glu-64, Val-138, and Asn-291 mutations andD228-240. D259-267, and D316-38I deletions: Ref. 13). Several cell lines
were obtained through transfection of primary MECs with the expressionconstructs pLTRp53cG + pRSV-H-ras. as well as with thepLTRp53cG + pRSV-v-fos constructs. The pTkneo construct (9) was cotrans-fected in all cultures to generate neomycin-resistant clones. The p53 + H-ras-transformed cell line (clones 4J and 4LI) and the p53 + fos-transformed cellline clone (6J3) arc described in a previous report (8). The HPV16 + EJras-transformed cells (clone HPV-C3, containing pRSVneo). as well as theAd5E I-transformed cells (clone 11.2. containing pTkneo) were describedelsewhere (6. 14). EL-4 cells are chemically induced thymoma cells, and RMAis a Rauscher MuLV-induced lymphoma cell line, both of B6 origin. 771 and786 cells are MCFI233 MuLV-induced lymphoma cells of C57BL/10 origin(15). HeLa.Dh and COS.Dh cells were obtained through transfection of the
human cervix carcinoma cell line HeLa and monkey kidney cell line COS,respectively, with a genomic H-2Db expression construct in the context ofpcDNA-1. All cell lines were cultured in medium consisting of Iscove'smodified Dulbecco's medium (Life Technologies. Inc.). supplemented with
8# heat-inactivated PCS (Bodinco. Alkmaar. the Netherlands). 2 mM L-
glutamine (ICN Biomcdicals Inc.. Costa Mesa, CA). 100 units/ml penicillin(Yamanouchi Pharma. Leiderdorp. the Netherlands), and 30 /xM 2-mercapto-ethanol (Merck, Darmstadt. Germany) at 37°Cin humidified air with 5% CO2.
Generation and Analysis of CTL and T-Cell Hybridomas. B6 micewere immunized twice by s.c. injection with IO7 IFN-y-treated. irradiated
p53/H-ras MECs (clone 4J). Spleen cells were restimulated in \-ilro in the
presence of 5% irradiated (25-Gy) tumor cells. Viable responder cells were
maintained in complete culture medium (see above) supplemented with 2%supernatant of ConA- and PMA-stimulated rat spleen cells (rat factor). Bulkcultures were restimulated weekly in U-bottomed 96-well plates (Costar) using
tumor cells and irradiated syngeneic splenic feeder cells in the presence of ratfactor. All CTL cultures were TCR-a/3*. CD4 . and CD8 +, as tested by flow
cytometry analysis. CTL clones raised against HPV16- and Ad5E I-trans
formed cell lines have been described previously (7, 9).Several of the CTL clones were fused with the LacZ BWZ36 CD8a
hybridoma cell line, as described previously ( 16. 17), resulting in continuouslygrowing T-cell hybridomas that exhibit the target specificity of the parental
CTL clones and that produce LacZ upon TCR triggering.Cytolytic activity of T-cell cultures was measured by means of europium
(Eu' +) release assays, as described elsewhere (6). The mean percentage
specific lysis of triplicate wells was calculated as follows: % specific lysis = [(cpm experimental release —¿�cpm spontaneous release)/(cpm maximumrelease - cpm spontaneous release)] x 100.
Measurement of secreted TNF-a by stimulated CTLs was performed with a
bioassay. using WEHI 164 clone 13 cells, as described previously(18). Percentage TNF-a release of triplicate wells was calculated as follows: % TNF-a release = [W550_650nm experimental wells —¿�/4550_65anm
wells containing medium only)/C4550_fi50nm wells containing 500 pg/mlTNF-a —¿�A550_65()nm wells containing medium only)].
Construction and Screening of cDNA Expression Library. Poly(A)-purified mRNA of HPV16/EJ-ras MEC (HPV-C3) was used as template foroIigo(dT)-primed cDNA synthesis according to the manufacturer's instructions
(Time Saver cDNA synthesis kit; Pharmacia Biotech) with the Not-dT18
primer d[AACTGGAAGAATTCGCGGCCGCAGGAAT(18l]p and EcoRladaptor AATTCGGCACGAG. cDNA was unidirectionally cloned into thevector pcDNA-1 (InVitrogen). The library was transformed into Escherichia
coli strain Sure (Life Technologies, Inc.), after which plasmid DNA was
isolated from pools of 50-70 bacterial colonies using the Qiaprep system
(Qiagen).Screening of the library involved transient transfection of cDNA-pools into
HeLa.Dh cells, seeded onto flat-bottomed 96-well plates, with DEAE dextran.
as described elsewhere (14). Two days after transfection. LacZ T-cell hybri
domas were added. The following day. cells were fixated, and the induced LacZwas visualized by incubation with a 5-bromo-4-chloro-3-indolyl-ß-D-galactopyr-anoside substrate, as described (17). The number of blue-stained hybridoma cells
in each of the wells was determined visually using a microscope.Peptides. Peptides were generated by solid-phase strategies on a multiple
peptide synthesizer (Abimed model AMS 422), as described previously (6).Purity of the peptides was determined by reverse-phase HPLC. Peptides (5 mg)were dissolved in 100 ju.1of DMSO. diluted in PBS. and stored at -20°C. Final
concentration of DMSO in assays was <0.2%.
RESULTS
An Immunogenic CTL Epitope Shared by p53- and HPV16-
transformed Cells. MECs of B6 origin that were transformed by amutated murine p53 gene and an activated murine H-rcis gene (clone4J; see "Materials and Methods") were previously used to immunize
p53 knockout mice for the isolation of p53-specific CTLs. The re
sulting CTLs were shown to recognize a peptide derived from thewild-type murine p53 sequence in the context of H-2Kh (8). During
these experiments, we observed a second CTL specificity that appeared to be codominant to the CTL response against p53 in p53knockout mice and that dominated the CTL response against thep53/H-ras MEC in p53+/+ mice. Because this additional CTL spec
ificity played a major role in the T-cell response against this tumor, we
set out to identify the immunogenic peptide concerned. CTLs with thespecificity of interest were isolated from the spleens of syngeneicp53+/+ mice that had been immunized with irradiated p53/H-ras
MECs (clone 4J). Cultures of such splenocytes that had been restimulated in vitro with the same tumor cells exhibited strong cytolyticactivity against the relevant tumor cells but not against untransformedcells (data not shown). From six independent CD4"/CD8+a/3+ T-cell
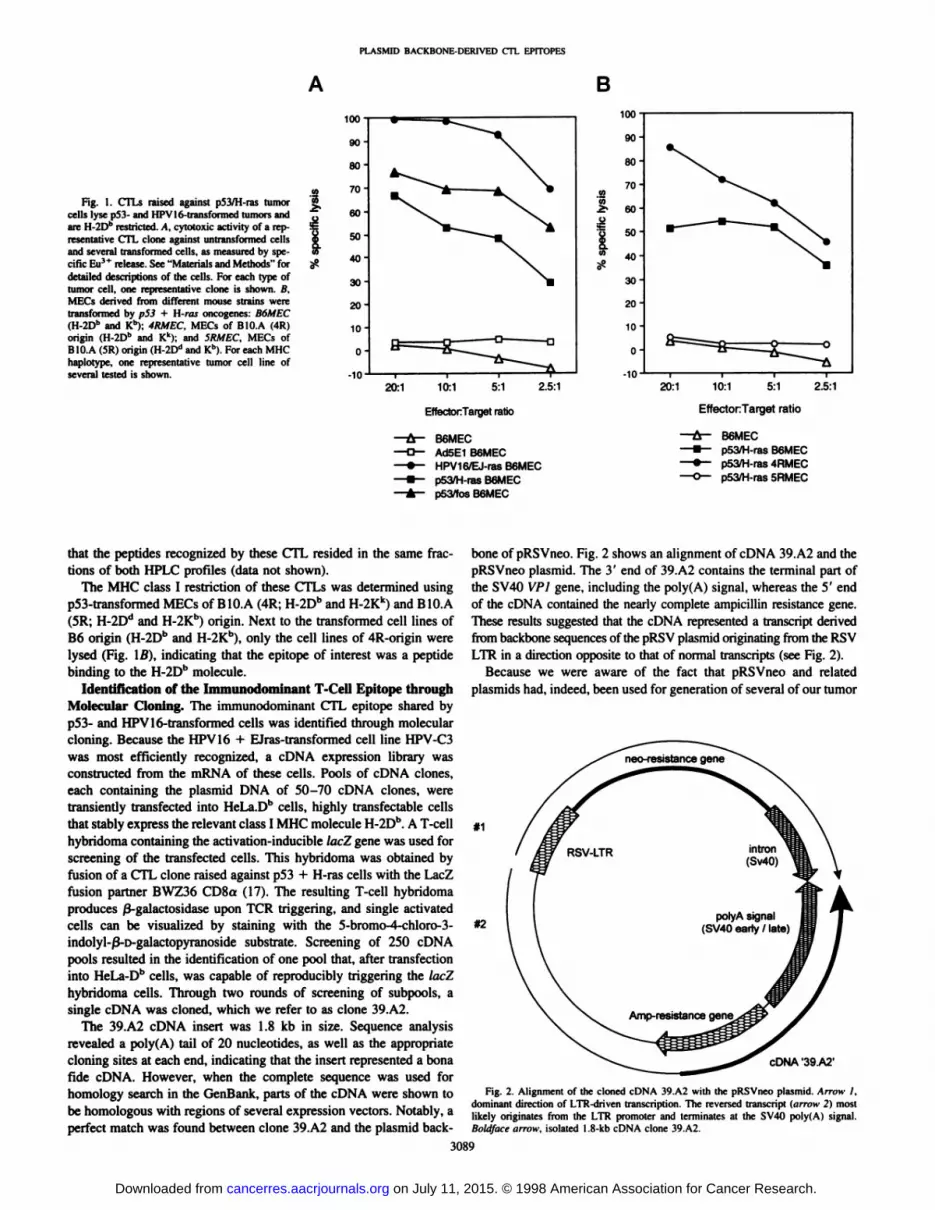
cultures, several clones were isolated. All these CTL clones exhibitedan identical target cell specificity, as is shown for a representativeCTL clone (Fig. 1/4). MECs transformed by p53 + H-ras as well as by
p53 + fos were efficiently lysed by the CTLs. On the other hand,untransformed MEC were not lysed, nor were various tumor cells ofB6 origin, including cells transformed by the El region of Ad5. B16melanoma cells, T-cell lymphomas EL4 and RMA, and B-cell lym-phomas 771 and 786 (Fig. \A and data not shown; see "Materials andMethods" for detailed description).
The results suggested that the sensitivity of the tumor cells to theCTLs was related to the fact that these cells were p53 transformed.Interestingly, cells transformed by the combination HPV16 + Eiras(e.g., clone HPV-C3) were also efficiently lysed by these CTLs (Fig.IA). Analysis of several independently generated p53- and HPV 16-
transformed cell lines showed that all these cell lines were lysed bythe CTLs (data not shown), indicating that p53- and HPV-transformed
cells apparently shared the CTL epitope of interest. Our group hasreported previously that the HPV16-transformed cell line HPV-C3
presents at least two different CTL epitopes, the subdominant HPV16E7-derived peptide RAHYNIVTF and an unknown ¡mmunodominant
epitope (7). Strikingly, we now found the CTLs raised againstHPV-C3 cells and reactive to this latter immunodominant epitope to
display a target cell specificity identical to that of the CTL raisedagainst p53-transformed cells (data not shown), supporting the notionthat our p53- and HPV 16-transformed cell lines share this dominantT-cell epitope. Moreover, analysis of HPLC-fractionated peptidepools eluted from both p53- and HPV 16-transformed cells revealed
3088
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
PLASMID BACKBONE-DERIVED CTL EPITOl'l S
B
Fig. 1. CTLs raised against p53/H-ras tumorcells lyse p53- and HPV16-transformed tumors andare H-2Dh restricted. A. cytotoxic activity of a rep
resentative CTL clone against untransformed cellsand several transformed cells, as measured by specific Eu3* release. See "Materials and Methods" for
detailed descriptions of the cells. For each type oftumor cell, one representative clone is shown. B.MECs derived from different mouse strains weretransformed by p53 + H-ras oncogenes: B6MEC(H-2Dh and Kh); 4RMEC, MECs of BIO.A (4R)origin (H-2Db and Kk); and SRMEC, MECs ofBIO.A (5R) origin (H-2Dd and Kh). For each MHC
haplotype, one representative tumor cell line ofseveral tested is shown.
100
20:1 10:1 5:1
EffectorTarget ratio
2.5:1
B6MECAd5E1 B6MECHPV16/EJ-rasB6MECp53/H-ras B6MEC
p53/fos B6MEC
90-
80-
70-
60 -
50-
40 -
30-
20-
10-
o-
-1020:1 10:1 5:1 2.5:1
EffectorTarget ratio
—¿�A- B6MEC—¿�•—p53/H-ras B6MEC—¿�•—p53/H-ras 4RMEC—¿�O—p53/H-ras 5RMEC
that the peptides recognized by these CTL resided in the same fractions of both HPLC profiles (data not shown).
The MHC class I restriction of these CTLs was determined usingp53-transformed MECs of B10.A (4R; H-2Dh and H-2Kk) and B10.A(5R; H-2D" and H-2Kb) origin. Next to the transformed cell lines ofB6 origin (H-2Db and H-2Kb), only the cell lines of 4R-origin were
lysed (Fig. Iß),indicating that the epitope of interest was a peptidebinding to the H-2Dh molecule.
Identification of the Immunodominant T-Cell Epitope throughMolecular Cloning. The immunodominant CTL epitope shared byp53- and HPV16-transformed cells was identified through molecularcloning. Because the HPV16 + EJras-transformed cell line HPV-C3
was most efficiently recognized, a cDNA expression library wasconstructed from the mRNA of these cells. Pools of cDNA clones,each containing the plasmid DNA of 50-70 cDNA clones, weretransiently transfected into HeLa.Db cells, highly transfectable cellsthat stably express the relevant class I MHC molecule H-2Db. A T-cell
hybridoma containing the activation-inducible lacZ gene was used for
screening of the transfected cells. This hybridoma was obtained byfusion of a CTL clone raised against p53 + H-ras cells with the LacZfusion partner BWZ36 CD8a (17). The resulting T-cell hybridomaproduces ß-galactosidase upon TCR triggering, and single activatedcells can be visualized by staining with the 5-bromo-4-chloro-3-indolyl-ß-D-galactopyranoside substrate. Screening of 250 cDNA
pools resulted in the identification of one pool that, after transfectioninto HeLa-Db cells, was capable of reproducibly triggering the lacZ
hybridoma cells. Through two rounds of screening of subpools, asingle cDNA was cloned, which we refer to as clone 39.A2.
The 39.A2 cDNA insert was 1.8 kb in size. Sequence analysisrevealed a poly(A) tail of 20 nucleotides, as well as the appropriatecloning sites at each end, indicating that the insert represented a bonafide cDNA. However, when the complete sequence was used forhomology search in the GenBank, parts of the cDNA were shown tobe homologous with regions of several expression vectors. Notably, aperfect match was found between clone 39.A2 and the plasmid back
bone of pRSVneo. Fig. 2 shows an alignment of cDNA 39. A2 and thepRSVneo plasmid. The 3' end of 39.A2 contains the terminal part ofthe SV40 VPÃŒgene, including the poly(A) signal, whereas the 5' end
of the cDNA contained the nearly complete ampicillin resistance gene.These results suggested that the cDNA represented a transcript derivedfrom backbone sequences of the pRSV plasmid originating from the RSVLTR in a direction opposite to that of normal transcripts (see Fig. 2).
Because we were aware of the fact that pRSVneo and relatedplasmids had, indeed, been used for generation of several of our tumor
#1
cDNA '39.A21
Fig. 2. Alignment of the cloned cDNA 39.A2 with the pRSVneo plasmid. Arrow I,dominant direction of LTR-driven transcription. The reversed transcript (iirrow 2) most
likely originates from the LTR promoter and terminates at the SV40 poly(A) signal.Boldface arrow, isolated 1.8-kh cDNA clone 39.A2.
3089
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
PLASMID BACKBONE-DERIVED CTL EPITOPliS
Oui
g
a
OUI
u111s
>a.uHI
UUI
-a
cDNA 39.A2
GAPOH
I I I I
12345
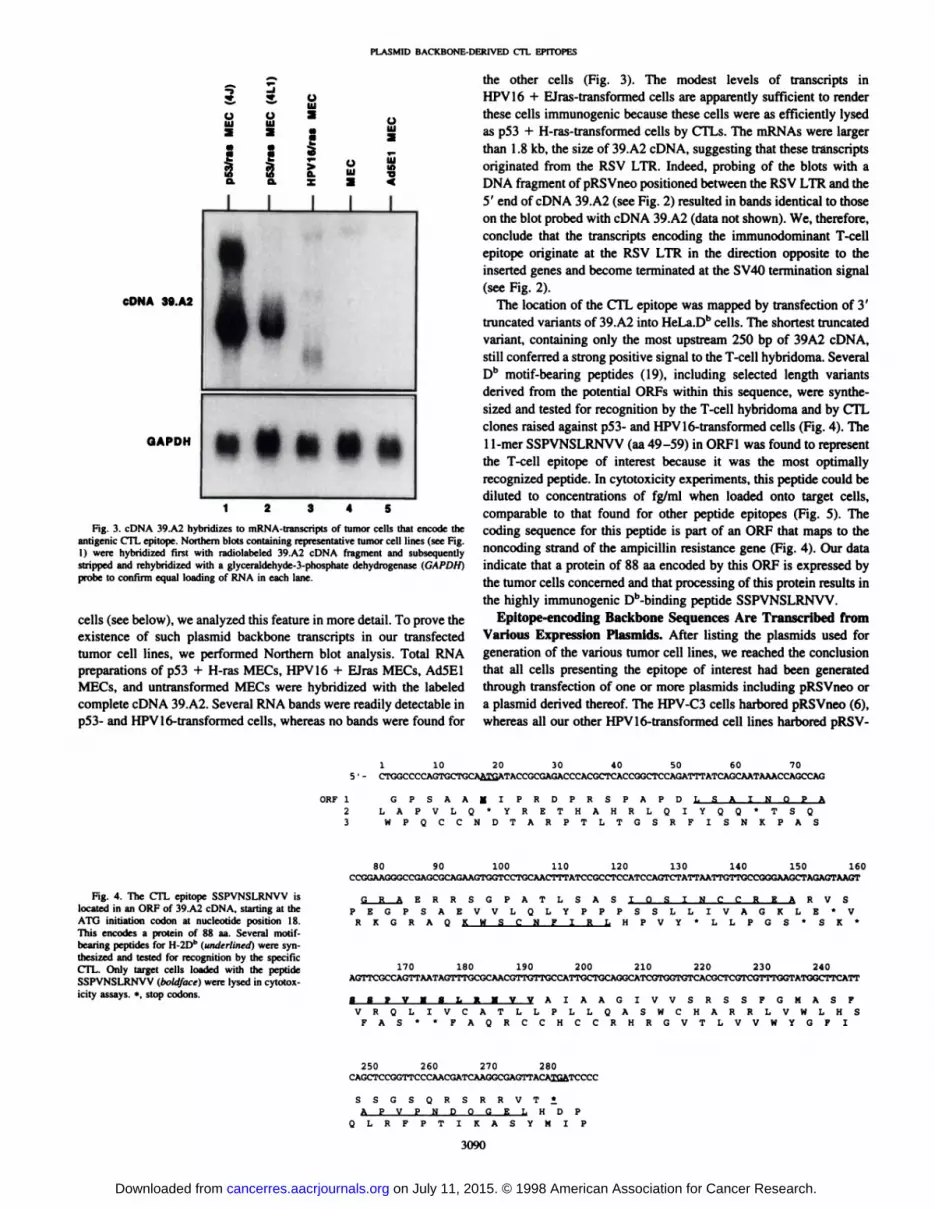
Fig. 3. cDNA 39.A2 hybridizes to mRNA-transcripts of tumor cells that encode the
antigenic CTL epitope. Northern blots containing representative tumor cell lines (see Fig.1) were hybridized first with radiolabeled 39.A2 cDNA fragment and subsequentlystripped and rehybridized with a giyceraldehyde-3-phosphate dehydrogenase (GAPDHÌ
probe to confirm equal loading of RNA in each lane.
cells (see below), we analyzed this feature in more detail. To prove theexistence of such plasmid backbone transcripts in our transfectedtumor cell lines, we performed Northern blot analysis. Total RNApreparations of p53 + H-ras MECs, HPV16 + EJras MECs, AdSEl
MECs, and untransformed MECs were hybridized with the labeledcomplete cDNA 39.A2. Several RNA bands were readily detectable inp53- and HPV16-transformed cells, whereas no bands were found for
the other cells (Fig. 3). The modest levels of transcripts inHPV 16 + EJras-transformed cells are apparently sufficient to render
these cells immunogenic because these cells were as efficiently lysedas p53 + H-ras-transformed cells by CTLs. The mRNAs were larger
than 1.8 kb, the size of 39.A2 cDNA, suggesting that these transcriptsoriginated from the RSV LTR. Indeed, probing of the blots with aDNA fragment of pRSVneo positioned between the RSV LTR and the5' end of cDNA 39.A2 (see Fig. 2) resulted in bands identical to those
on the blot probed with cDNA 39.A2 (data not shown). We, therefore,conclude that the transcripts encoding the immunodominant T-cell
epitope originate at the RSV LTR in the direction opposite to theinserted genes and become terminated at the SV40 termination signal(see Fig. 2).
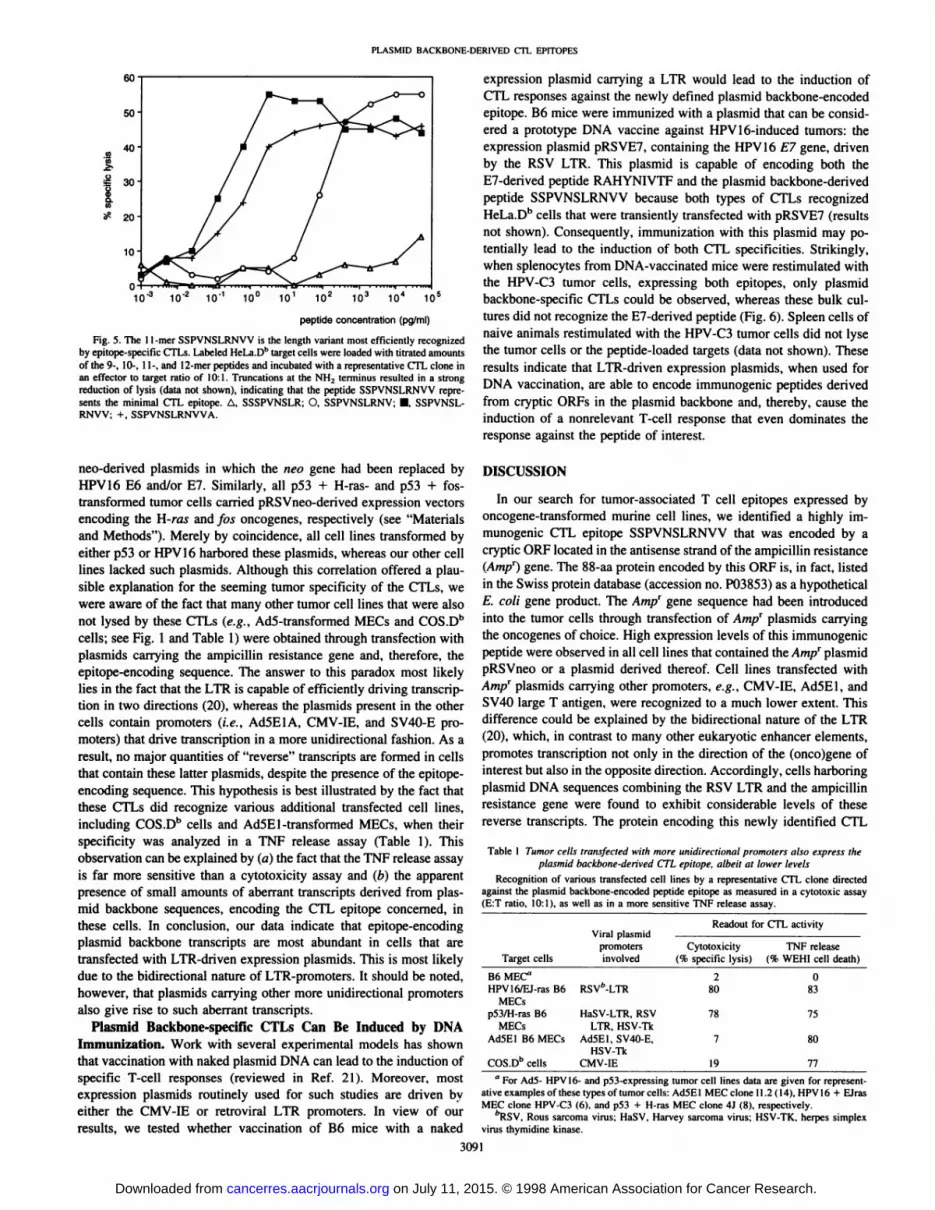
The location of the CTL epitope was mapped by transfection of 3'truncated variants of 39.A2 into HeLa.Db cells. The shortest truncated
variant, containing only the most upstream 250 bp of 39A2 cDNA,still conferred a strong positive signal to the T-cell hybridoma. SeveralDb motif-bearing peptides (19), including selected length variants
derived from the potential ORFs within this sequence, were synthesized and tested for recognition by the T-cell hybridoma and by CTLclones raised against p53- and HPV 16-transformed cells (Fig. 4). The11-mer SSPVNSLRNVV (aa 49-59) in ORF1 was found to representthe T-cell epitope of interest because it was the most optimally
recognized peptide. In cytotoxicity experiments, this peptide could bediluted to concentrations of fg/ml when loaded onto target cells,comparable to that found for other peptide epitopes (Fig. 5). Thecoding sequence for this peptide is part of an ORF that maps to thenoncoding strand of the ampicillin resistance gene (Fig. 4). Our dataindicate that a protein of 88 aa encoded by this ORF is expressed bythe tumor cells concerned and that processing of this protein results inthe highly immunogenic Db-binding peptide SSPVNSLRNVV.
Epitope-encoding Backbone Sequences Are Transcribed from
Various Expression Plasmids. After listing the plasmids used forgeneration of the various tumor cell lines, we reached the conclusionthat all cells presenting the epitope of interest had been generatedthrough transfection of one or more plasmids including pRSVneo ora plasmid derived thereof. The HPV-C3 cells harbored pRSVneo (6),whereas all our other HPV 16-transformed cell lines harbored pRSV-
1 10 20 30 40 50 60 705 ' - CTGGCCCCAGTGCTGCAA^ATACCGCGAGACCCACGCTCACCGGCTCCAGATTTATCAGCAATAAACCAGCCAG
ORF 1 GPSAAHIPRDPRSPAPD L S A I2 LAPVLQ*YRETHAHRLQIYQQ
3 WPQCCNDTARPTLTGSRFISN
' T S Q
K P A S
80 90 100 110 120 130 140 150 160
Fig. 4. The CTL epitope SSPVNSLRNVV islocated in an ORF of 39.A2 cDNA, starting at theATO initiation codon at nucleotide position 18.This encodes a protein of 88 aa. Several motif-bearing peptides for H-2Dh (underlined) were syn
thesized and tested for recognition by the specificCTL. Only target cells loaded with the peptideSSPVNSLRNVV (boldface) were lysed in cytotoxicity assays. *, stop codons.
G R A ERRSGPATLSAS IOSINCCREA R V SPEGPSAEVVLQLYPPPSSLLIVAGKLE'V
R K G R A Q KWSCNFIRL HPVY*LLPGS*SK*
170 180 190 200 210 220 230 240AGTTCGCCAGTTAATAGTTTGCGCAACGTTGTTGCCATTGCTGCAGGCATCGTGGTGTCACGCTCGTCGTTTGGTATGGCTTCATT
SSPVHSLRNVV AIAAGIVVSRSSFGMASFVRQLIVCATLLPLLQASWCHARRLVWLHS
FAS * * FAQRCCHCCRHRGVTLVVWYGF I
250 260 270 280CAGCTCCGGTTCCCAACGATCAAGGCGAGTTACAI2ATCCCC
SSGSQRSRRVT!APVPNDOGEL HDP
QLRFPTIKASYMIP
3090
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
PLASMID BACKBONE-DERIVED CTL EPITOPHS
5s 20-
10- 10'' 10'10
peptide concentration (pg/ml)
Fig. 5. The 11-mer SSPVNSLRNVV is the length variant most efficiently recognizedby epitope-specific CTLs. Labeled HeLa.D11target cells were loaded with titrated amounts
of the 9-, 10-, 11-, and 12-merpeptides and incubated with a representative CTL clone inan effector to target ratio of 10:1. Truncations at the NH2 terminus resulted in a strongreduction of lysis (data not shown), indicating that the peptide SSPVNSLRNVV represents the minimal CTL epitope. A, SSSPVNSLR; O, SSPVNSLRNV: •¿�SSPVNSLRNVV; + . SSPVNSLRNVVA.
neo-derived plasmids in which the neo gene had been replaced byHPV16 E6 and/or E7. Similarly, all p53 + H-ras- and p53 + fos-transformed tumor cells carried pRSVneo-derived expression vectorsencoding the H-ras and fas oncogenes, respectively (see "Materialsand Methods"). Merely by coincidence, all cell lines transformed by
either p53 or HPV16 harbored these plasmids, whereas our other celllines lacked such plasmids. Although this correlation offered a plausible explanation for the seeming tumor specificity of the CTLs, wewere aware of the fact that many other tumor cell lines that were alsonot lysed by these CTLs (e.g., Ad5-transformed MECs and COS.Dh
cells; see Fig. 1 and Table 1) were obtained through transfection withplasmids carrying the ampicillin resistance gene and, therefore, theepitope-encoding sequence. The answer to this paradox most likely
lies in the fact that the LTR is capable of efficiently driving transcription in two directions (20), whereas the plasmids present in the othercells contain promoters (i.e., AdSEl A, CMV-IE, and SV40-E pro
moters) that drive transcription in a more unidirectional fashion. As aresult, no major quantities of "reverse" transcripts are formed in cells
that contain these latter plasmids, despite the presence of the epitope-
encoding sequence. This hypothesis is best illustrated by the fact thatthese CTLs did recognize various additional transfected cell lines,including COS.Db cells and AdSEl-transformed MECs. when their
specificity was analyzed in a TNF release assay (Table 1). Thisobservation can be explained by (a) the fact that the TNF release assayis far more sensitive than a cytotoxicity assay and (b) the apparentpresence of small amounts of aberrant transcripts derived from plas-
mid backbone sequences, encoding the CTL epitope concerned, inthese cells. In conclusion, our data indicate that epitope-encoding
plasmid backbone transcripts are most abundant in cells that aretransfected with LTR-driven expression plasmids. This is most likelydue to the bidirectional nature of LTR-promoters. It should be noted,
however, that plasmids carrying other more unidirectional promotersalso give rise to such aberrant transcripts.
Plasmid Backbone-specific CTLs Can Be Induced by DNA
Immunization. Work with several experimental models has shownthat vaccination with naked plasmid DNA can lead to the induction ofspecific T-cell responses (reviewed in Ref. 21). Moreover, most
expression plasmids routinely used for such studies are driven byeither the CMV-IE or retroviral LTR promoters. In view of our
results, we tested whether vaccination of B6 mice with a naked
expression plasmid carrying a LTR would lead to the induction ofCTL responses against the newly defined plasmid backbone-encoded
epitope. B6 mice were immunized with a plasmid that can be considered a prototype DNA vaccine against HPV16-induced tumors: the
expression plasmid pRSVEV, containing the HPV16 E7 gene, drivenby the RSV LTR. This plasmid is capable of encoding both theE7-derived peptide RAHYNIVTF and the plasmid backbone-derived
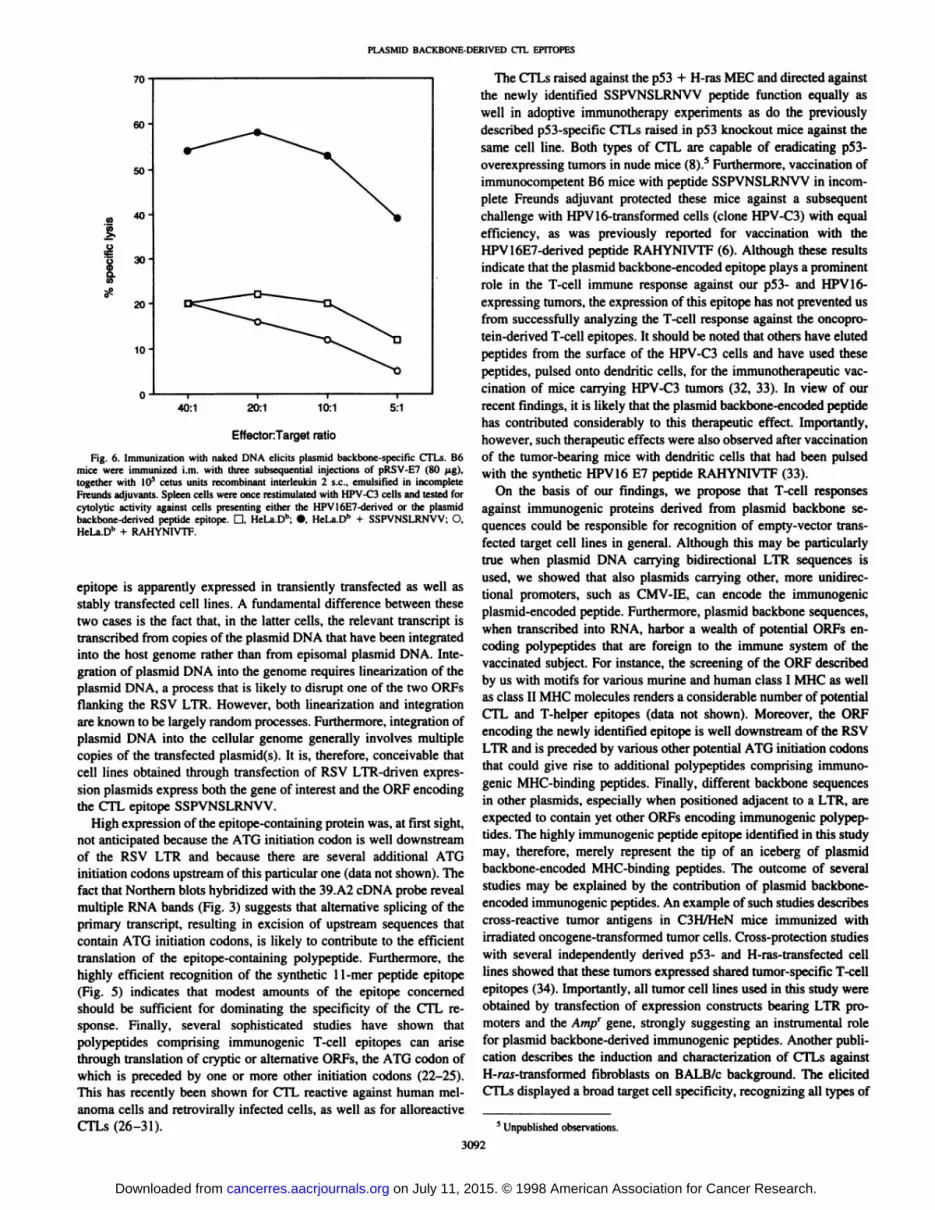
peptide SSPVNSLRNVV because both types of CTLs recognizedHeLa.Dh cells that were transiently transfected with pRSVEV (results
not shown). Consequently, immunization with this plasmid may potentially lead to the induction of both CTL specificities. Strikingly,when splenocytes from DNA-vaccinated mice were restimulated withthe HPV-C3 tumor cells, expressing both epitopes, only plasmidbackbone-specific CTLs could be observed, whereas these bulk cultures did not recognize the E7-derived peptide (Fig. 6). Spleen cells ofnaive animals restimulated with the HPV-C3 tumor cells did not lysethe tumor cells or the peptide-loaded targets (data not shown). Theseresults indicate that LTR-driven expression plasmids, when used for
DNA vaccination, are able to encode immunogenic peptides derivedfrom cryptic ORFs in the plasmid backbone and, thereby, cause theinduction of a nonrelevant T-cell response that even dominates the
response against the peptide of interest.
DISCUSSION
In our search for tumor-associated T cell epitopes expressed byoncogene-transformed murine cell lines, we identified a highly im
munogenic CTL epitope SSPVNSLRNVV that was encoded by acryptic ORF located in the antisense strand of the ampicillin resistance(Am//) gene. The 88-aa protein encoded by this ORF is, in fact, listed
in the Swiss protein database (accession no. P03853) as a hypotheticalE. coli gene product. The Amp' gene sequence had been introduced
into the tumor cells through transfection of Am// plasmids carryingthe oncogenes of choice. High expression levels of this immunogenicpeptide were observed in all cell lines that contained the Am// plasmidpRSVneo or a plasmid derived thereof. Cell lines transfected withAm// plasmids carrying other promoters, e.g., CMV-IE, AdSEl, and
SV40 large T antigen, were recognized to a much lower extent. Thisdifference could be explained by the bidirectional nature of the LTR(20), which, in contrast to many other eukaryotic enhancer elements,promotes transcription not only in the direction of the (onco)gene ofinterest but also in the opposite direction. Accordingly, cells harboringplasmid DNA sequences combining the RSV LTR and the ampicillinresistance gene were found to exhibit considerable levels of thesereverse transcripts. The protein encoding this newly identified CTL
Table 1 Tumor celia transfected with more iinitlirectiontil promoters alno express theplasmiti backbone-derived CTL epitope. aÃbeiÃti¡lower levels
Recognition of various transfected cell lines by a representative CTL clone directedagainst the plasmid backbone-encoded peptide epitope as measured in a cytotoxic assay(E:T ratio. 10:1), as well as in a more sensitive TNF release assay.
TargetcellsB6MEC"
HPV16/EJ-rasB6MECsp53/H-ras
B6
MECsAdSElB6MECsCOS.D"
cellsViral
plasmidpromoters
involvedRSV''-LTRHaSV-LTR.
RSVLTR,HSV-TkAd5El.SV40-E,HSV-TkCMV-IEReadout
for CTLactivityCytotoxicity
(% specificlysis)2
8078719TNF
release(% WEHI celldeath)0
83758077
" For Ad5- HPVI6- and p53-ex pressing tumor cell lines data are given for represent
ative examples of these types of tumor cells: Ad5El MEC clone 11.2 (14), HPV16 + EJrasMEC clone HPV-C3 (6), and p53 + H-ras MEC clone 4J (8), respectively.
'RSV, Rous sarcoma virus; HaSV. Harvey sarcoma virus; HSV-TK, herpes simplex
virus ihymidine kinase.
3091
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
PLASMID BACKBONE-DERIVED CTL EPITOPES
70
I
i
60-
50-
40-
30 H
20 H
10
40:1 20:1 10:1 5:1
EffectonTarget ratio
Fig. 6. Immuni/ation with naked DNA elicits plasmid backbone-specific CTLs. B6mice were immunized i.m. wilh three subsequential injections of pRSV-E7 (80 (ig),together with I0~s cetus units recombinant interleukin 2 s.c.. emulsified in incomplete
Freunds adjuvants. Spleen cells were once restimulated with HPV-C3 cells and tested forcytolytic activity against cells presenting either the HPVI6E7-derived or the plasmidbackbone-derived peptide epitope. D, HeLa.D"; •¿�.HeLa.Dh + SSPVNSLRNVV; O,HeLa.D" + RAHYNIVTF.
epitope is apparently expressed in transiently transfected as well asstably transfected cell lines. A fundamental difference between thesetwo cases is the fact that, in the latter cells, the relevant transcript istranscribed from copies of the plasmid DNA that have been integratedinto the host genome rather than from episomal plasmid DNA. Integration of plasmid DNA into the genome requires linearization of theplasmid DNA, a process that is likely to disrupt one of the two ORFsflanking the RSV LTR. However, both linearization and integrationare known to be largely random processes. Furthermore, integration ofplasmid DNA into the cellular genome generally involves multiplecopies of the transfected plasmid(s). It is, therefore, conceivable thatcell lines obtained through transfection of RSV LTR-driven expres
sion plasmids express both the gene of interest and the ORF encodingthe CTL epitope SSPVNSLRNVV.
High expression of the epitope-containing protein was, at first sight,
not anticipated because the ATG initiation codon is well downstreamof the RSV LTR and because there are several additional ATGinitiation codons upstream of this particular one (data not shown). Thefact that Northern blots hybridized with the 39.A2 cDNA probe revealmultiple RNA bands (Fig. 3) suggests that alternative splicing of theprimary transcript, resulting in excision of upstream sequences thatcontain ATG initiation codons, is likely to contribute to the efficienttranslation of the epitope-containing polypeptide. Furthermore, thehighly efficient recognition of the synthetic 11-mer peptide epitope
(Fig. 5) indicates that modest amounts of the epitope concernedshould be sufficient for dominating the specificity of the CTL response. Finally, several sophisticated studies have shown thatpolypeptides comprising immunogenic T-cell epitopes can arise
through translation of cryptic or alternative ORFs, the ATG codon ofwhich is preceded by one or more other initiation codons (22-25).
This has recently been shown for CTL reactive against human melanoma cells and retrovirally infected cells, as well as for alloreactiveCTLs (26-31).
The CTLs raised against the p53 + H-ras MEC and directed against
the newly identified SSPVNSLRNVV peptide function equally aswell in adoptive immunotherapy experiments as do the previouslydescribed p53-specific CTLs raised in p53 knockout mice against thesame cell line. Both types of CTL are capable of eradicating p53-overexpressing tumors in nude mice (8).5 Furthermore, vaccination of
immunocompetent B6 mice with peptide SSPVNSLRNVV in incomplete Freunds adjuvant protected these mice against a subsequentchallenge with HPV16-transformed cells (clone HPV-C3) with equal
efficiency, as was previously reported for vaccination with theHPV16E7-derived peptide RAHYNIVTF (6). Although these resultsindicate that the plasmid backbone-encoded epitope plays a prominentrole in the T-cell immune response against our p53- and HPV 16-
expressing tumors, the expression of this epitope has not prevented usfrom successfully analyzing the T-cell response against the oncopro-tein-derived T-cell epitopes. It should be noted that others have elutedpeptides from the surface of the HPV-C3 cells and have used these
peptides, pulsed onto dendritic cells, for the immunotherapeutic vaccination of mice carrying HPV-C3 tumors (32, 33). In view of ourrecent findings, it is likely that the plasmid backbone-encoded peptide
has contributed considerably to this therapeutic effect. Importantly,however, such therapeutic effects were also observed after vaccinationof the tumor-bearing mice with dendritic cells that had been pulsed
with the synthetic HPV 16 E7 peptide RAHYNIVTF (33).On the basis of our findings, we propose that T-cell responses
against immunogenic proteins derived from plasmid backbone sequences could be responsible for recognition of empty-vector trans
fected target cell lines in general. Although this may be particularlytrue when plasmid DNA carrying bidirectional LTR sequences isused, we showed that also plasmids carrying other, more unidirectional promoters, such as CMV-IE, can encode the immunogenicplasmid-encoded peptide. Furthermore, plasmid backbone sequences,
when transcribed into RNA, harbor a wealth of potential ORFs encoding polypeptides that are foreign to the immune system of thevaccinated subject. For instance, the screening of the ORF describedby us with motifs for various murine and human class I MHC as wellas class II MHC molecules renders a considerable number of potentialCTL and T-helper epitopes (data not shown). Moreover, the ORF
encoding the newly identified epitope is well downstream of the RSVLTR and is preceded by various other potential ATG initiation codonsthat could give rise to additional polypeptides comprising immunogenic MHC-binding peptides. Finally, different backbone sequences
in other plasmids, especially when positioned adjacent to a LTR. areexpected to contain yet other ORFs encoding immunogenic polypeptides. The highly immunogenic peptide epitope identified in this studymay, therefore, merely represent the tip of an iceberg of plasmidbackbone-encoded MHC-binding peptides. The outcome of severalstudies may be explained by the contribution of plasmid backbone-
encoded immunogenic peptides. An example of such studies describescross-reactive tumor antigens in C3H/HeN mice immunized withirradiated oncogene-transformed tumor cells. Cross-protection studieswith several independently derived p53- and H-ras-transfected celllines showed that these tumors expressed shared tumor-specific T-cell
epitopes (34). Importantly, all tumor cell lines used in this study wereobtained by transfection of expression constructs bearing LTR promoters and the Amp' gene, strongly suggesting an instrumental role
for plasmid backbone-derived immunogenic peptides. Another publi
cation describes the induction and characterization of CTLs againstH-ra.c-transformed fibroblasts on BALB/c background. The elicited
CTLs displayed a broad target cell specificity, recognizing all types of
5 Unpublished observations.
3092
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

PI-ASMID BACKBONE-DERIVED CTL EPITOPES
generated H-ras transfectants (35). Again, the expression of the H-ras
gene in the transfected cell lines was driven by an LTR, leading toplasmid backbone transcripts and expression of highly immunogenicCTL epitopes. In view of our results, it is likely that the tumor-specific
antigens described in these studies can be defined as plasmid backbone derived.
Apart from influencing the outcome of immunological experimentsin tumor models, immunogenic peptide epitopes encoded by plasmidbackbone sequences may also play a role in DNA vaccination. Micevaccinated with plasmid DNA encoding both a viral epitope and thebackbone-derived epitope raised strong CTL responses against the
latter peptide. Vaccination with plasmid DNA containing LTR promoters is now being applied in a wide variety of experimental models(reviewed in Ref. 36) and even in clinical trials (37, 38). It should benoted that at least the majority of problems concerning the inductionof T-cell responses against vector backbone sequences can be avoided
by using markedly different expression vectors, either plasmids orviruses, for in vivo priming and boosting, as well as for transfection ofcells to be used for in vitro restimulation of T-cell responses.
In conclusion, plasmid backbone sequences can encode highlyimmunogenic T-cell epitopes, the T-cell response against which can
even dominate the response against a viral peptide. Expression of suchpeptides by transfected cells influences the outcome of immunologicaltumor studies, whereas the use of plasmids for DNA vaccination canlead to the induction of T-cell immunity against such nonrelevant
epitopes.
REFERENCES
1. Melief, C. J. M. Tumor eradication by adoptive transfer of cytotoxic T lymphocytes.Adv. Cancer Res., 58: 143-175, 1992.
2. Boon, T.. Cerottini, J-C., van den Eynde, B., van der Bruggen. P., and Van Pel. A.Tumor antigens recognized by T lymphocytes. Annu. Rev. Immunol., 12: 337-365.
1994.3. Jaffe, M. J.. and Pardoll. D. M. Murine tumor antigens: is it worth the search? Curr.
Opin. Immunol., 8: 622-627. 1996.4. Melief. C. J. M.. Offringa. R.. Toes. R. E. M., and Kast. W. M. Peptide-based cancer
vaccines. Curr. Opin. Immunol., 8: 651-657, 1996.
5. Toes, R. E. M., Offringa. R., Feltkamp, M. C. W., Visseren, M. J. W.. Schoenberger.S. P., Melief, C. J. M.. and Kast, W. M. Tumor rejection antigens and tumor specificcytotoxic T lymphocytes. Behring Inst. Mitt.. 94: 72-86, 1994.
6. Feltkamp, M. C. W., Smits. H. L.. Vierboom, M. P. M., Minnaar, R. P., De Jongh,B. M.. Drijfhout. J. W.. Ter Schegge!. J., Melief. C. J. M., and Kasl, W. M.Vaccination with a cytotoxic T lymphocyte epitope-containing peptide protectsagainst a tumor induced by human papillomavirus type 16-transformed cells. Eur.J. Immunol., 23: 2242-2249. 1993.
7. Feltkamp, M. C. W., Vreugdenhil, G. R., Vierboom, M. P. M., Ras, E.. Van der Burg,S. H.. Ter Schegge!, J., Melief, C. J. M., and Kast, W. M. CTL raised against asubdominant epitope offered as a synthetic peptide eradicate human papillomavirustype 16-induced tumors. Eur. J. Immunol.. 25: 2638-2641. 1995.
8. Vierboom, M. P. M., Nijman. H. W., Offringa. R., van der Voort, E. I. H., van Hall.T.. van den Broek, L., Fleuren, G., Kenemans. P., Kast, W. M., and Melief, C. J. M.Tumor eradication by wild-type p53-specific cytotoxic T lymphocytes. J. Exp. Med.,186: 695-704, 1997.
9. Kast, W. M.. Offringa. R.. Peters, P. J., Voordouw. A. C.. Meloen, R. H., Van der Eb.A. J., and Melief. C. J. M. Eradication of adenovirus El-induced tumors by Ela-specific cytotoxic T lymphocytes. Cell, 59: 603-615, 1989.
10. Van der Eb, A. J.. and Graham, F. L. Assay of transforming activity of tumor virusDNA. Methods Enzymol., 654: 826-839, 1980.
11. Kac/.marek. L.. Oren. M., and Baserga, R. Co-operation between the p53 proteintumor antigen and platelet-poor plasma in the induction of cellular DNA synthesis.Exp. Cell Res.. 762: 268-272. 1986.
12. Reddy, E. P.. Lipman, D.. Anderson, P. R.. Tronick. S. R.. and Aaronson, S. A.Nucleotide sequence analysis of the BALB/c murine sarcoma virus transforminggene. J. Virol., 53: 984-987, 1985.
13. Van Beveren. C., van Straaten. F., Curran, T.. Muller. R., and Verma. I. M. Analysisof FBJ-MuSV provirus and c-fos (mouse) gene reveals that viral and cellularfos geneproducts have different carboxy termini. Cell, 32: 1241-1255, 1983.
14. Toes, R. E. M., Offringa, R., Blom, H. J. J.. Brandt. R. M. P., Van der Eb, A. J..Melief, C. J. M.. and Kast. W. M. An adenovirus type 5 early region 1B-encoded CTL
epitope-mediating tumor eradication by CTL clones is down-modulated by an activated ras oncogene. J. Immuno]., 154: 3396-3405, 1995.
15. Sijts, E. J. A. M., Ossendorp. F.. Mengedé,E. A. M., Van den Eisen, P. J., and Melief.C. J. M. An immunodominant MCF murine leukemia virus encoded CTL epitope,identified by its MHC class I-binding motif, explains MuLV type specificity ofMCF-directed CTL. J. Immunol., /52: 106-116, 1994.
16. Karttunen, J.. and Shastri, N. Measurement of ligand-induced activation in singleviable T cells using the lacZ reporter gene. Proc. Nati. Acad. Sci. USA, 88: 3972-3976, 1991.
17. Sanderson, S.. and Shastri. N. LacZ inducible, antigen/MHC-specific T cell hybrids.
Int. Immunol.. 6: 1994.18. Traversai! C., van der Bruggen, P.. van den Eynde, B.. Hainaut. P., Lemoine, C.,
Ohta, N., Old, L.. and Boon, T. Transfection and expression of a gene coding for ahuman melanoma antigen recognized by autologuous CTL. Immunogenetics, 35:145-152, 1992.
19. Rammensee, H. G.. Friede, T.. and Stevanovic, S. MHC ligands and peptide motifs:first listing. Immunogenetics, 41: 178-228, 1995.
20. Christy, R. J., and Huang. R. C. C. Functional analysis of the long terminal repeatsof intracisterna! A-particle genes: sequences within th U3 region determine both theefficiency and direction of promoter activity. Mol. Cell. Biol., 8: 1093-1102, 1988.
21. Donnelly. J. J.. Ulmer, J. B.. Shiver. J. W., and Liu. M. A. DNA vaccines. Annu. Rev.Immunol.. /5: 617-648, 1997.
22. Bullock, T. N. J.. and Eisenlohr. L. C. Ribosomal scanning past the primary initiationcodon as a mechanism for expression of CTL epitopes encoded in alternative readingframes. J. Exp. Med., 184: 1319-1329, 1996.
23. Bullock, T. N. J., Patterson. A. E., Franlin. L. L.. Notidis. E., and Eisenlohr, L. C.Initiation codon scanthrough versus termination codon readthrough demonstratesstrong potential for major histocompatibility complex class I-restricted cryptic epitopeexpression. J. Exp. Med.. 186: 1051-1058. 1997.
24. Elliott. T.. Bodmer, H.. and Townsend. A. Recognition of out-of-frame major hislo-compatibility complex class I-restricted epitopes in vivo. Eur. J. Immunol.. 26:1175-1179, 1996.
25. Fetten. J. V.. Roy. N.. and Gilboa, E. A frameshift mutation at the NH, terminus ofthe nucleoprotein gene does not affect generation of cytotoxic T lymphocyte epitopes.J. Immunol.. ¡47:2697-2705, 1991.
26. Wang. R., Parkhurst, M. R., Kawakami, Y., Robbins, P. F., and Rosenberg, S. A.Utilization of an alternative open reading frame of a normal gene in generating anovel human cancer antigen. J. Exp. Med., 183: 1131-1140, 1996.
27. Guilloux, Y., Lucas, S., Brichard. V. G., Van Pel, A., Viret. C.. De Plaen. E.,Brasseur, F., Lethe, B.. Jotereau, F., and Boon. T. A peptide recognized by humancytolytic T lymphocytes on HLA-A2 melanomas is encoded by an intron sequence ofthe A'-acetylglucosaminyltransferese V gene. J. Exp. Med., 183: 1173-1183. 1996.
28. Coulie, P. G., Lehmann, F.. Lethe, B.. Herman. J.. Lurquin, C., Andrawiss. M.. andBoon. T. A mutated intron sequence codes for an antigenic peptide recognized bycytolytic T lymphocytes on a human melanoma. Proc. Nati. Acá.Sci. USA. 92:7976-7980. 1995.
29. Malarkannan, S.. Afkarian. M., and Shastri. N. A rare cryptic translation product ispresented by Kb major histocompatibility complex class I molecule to alloreactive Tcells. J. Exp. Med., 182: 1739-1750, 1995.
30. Malarkannan. S.. Gonzalez, F.. Nguyen. V.. Adair, G.. and Shastri, N. AlloreactiveCD8+ T cells can recognize unusual, rare, and unique processed peptide/MHCcomplexes. J. Immunol.. 157: 4464-4473, 1996.
31. Mayrand. S., Schwarz, D. A., and Green. W. R. An alternative translational readingframe encodes an immunodominant retroviral CTL determinant expressed by animmunodeficiency-causing retrovirus. J. Immunol.. 760: 39-50. 1998.
32. Zitvogel, L.. Mayordomo, J. L, Tjandrawan, T.. Deleo. A. B.. Clarke. M. R.. Lotze,M. T.. and Storkus. W. J. Therapy of murine tumors with tumor peplide-pulseddendritic cells: dependence of T cells. B7 costimulation. and T helper cell 1-associated cytokines. J. Exp. Med.. 183: 87-97. 1996.
33. Mayordomo, J. I., Zorina, T.. Storkus, W. J., Zitvogel. L.. Celluzzi, C., Falo. L. D..Melief. C. J. M., Ildstad, S. T.. Kast, W. M.. Deleo, A. B.. and Lotze. M. T. Bonemarrow-derived dendritic cells pulsed with synthetic tumour peptides elicit protectiveand therapeutic antitumour immunity. Nat. Med., /: 1297-1302. 1995.
34. Appleman. L.. Uyeki. J., and Frey. A. B. Mouse embryo fibroblasts transformed byactivated ras or dominant-negative p53 express cross-reactive tumor rejection antigens. Int. J. Cancer, 61: 887-894, 1995.
35. Ehrlich. T., Wishniak. O., Isakov, N.. Cohen. O.. Segal. S.. Rager-Zisman, B., andGopas, J. The effect of H-ras expression on tumorigenicity and immunogenicity ofBalb/c 3T3 fibroblasts. Immunol. Lett., 39: 3-8, 1994.
36. Spooner. R. A.. Dconarain. M. P.. and Epenetos, A. A. DNA vaccination for cancertreatment. Gene Ther.. 2: 173-180. 1995.
37. Nabel. G. J., Nabel, E. G.. Yang, Z.. Fox. B. A.. Plautz. G. E.. Gao, X.. Huang, L.,Shu, S., Gordon, D.. and Chang. A. E. Direct gene transfer with DNA-liposome
complexes in melanoma: expression, biologic activity, and lack of toxicity in humans.Proc. Nati. Acad. Sci. USA, 90: 11307-11311. 1993.
38. Nabel. G. J.. Gordon. D., Bishop, D. K.. Nickoloff. B. J.. Yang. Z.. Aruga, A.,Cameron. M. J.. Nabel, E. G., and Chang. A. E. Immune response in humanmelanoma after transfer of an allogeneic class I major histocompatibility complexgene with DNA-liposome complexes. Proc. Nati. Acad. Sci. USA. 93: 15388-15393.
1996.
3093
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1998;58:3087-3093. Cancer Res Thorbald van Hall, Nastasja E. van de Rhee, Stephen P. Schoenberger, et al. T-Lymphocyte EpitopesSequences Can Provide Highly Immunogenic Cytotoxic Cryptic Open Reading Frames in Plasmid Vector Backbone
Updated version
http://cancerres.aacrjournals.org/content/58/14/3087
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on July 11, 2015. © 1998 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from