controlling methane emissions from ruminants employing bacteriocin
TRANSCRIPT

140 Climate Resilient Livestock & Production System
Chapter-13
Controlling Methane Emissions from Ruminants Employing Bacteriocin Renuka, A.K. Punia*, Syma Ashraf, P. Srinivasulu, T. Sridhar Goud, A.K. Singh, Lovely Sharma, Sonia Saini, Rajni Devi, Anil Kumar, S.V. Singh and R.C. Upadhyay
Dairy Cattle Physiology Division, *Dairy Microbiology Division, National Dairy Research Institute, Karnal-132001 (Haryana) INDIA
Bacteriocins are proteinaceous toxins produced by bacteria to inhibit the growth of similar or closely related bacterial strain(s). These are typically considered to be of narrow spectrum antibiotics. An attempt has been made in this article to understand the contribution of ruminants to methane emission and to mitigate the methane emission from ruminants by using bacteriocins. The greenhouse gas (GHG) emissions from agriculture sector accounts for about 25.5% of the total global and over 60% of anthropogenic emissions. Animal husbandry accounts for 18% of GHG emissions that cause global warming. CH4 is considered to be the largest potential contributor to the global warming phenomenon. Ruminant livestock such as cattle, buffalo, sheep and goats contributes the major proportion of total agricultural emission of methane. Indian livestock system is a large contributor to GHGs and therefore to the global warming phenomenon. Methane emission from enteric fermentation from Indian livestock ranged from 7.26 to 10.4 MT/year. Nisin obtained from Lactobaccilus lactis ssp. lactis, has also been shown to decrease methane production in vitro. Bovicin HC5, another bacteriocin produced by Streptococcus bovis from the rumen, has been reported to suppress methane production by 50%. The potential for rumen-produced bacteriocins to suppress methanogenesis is unknown, but their potential as a new generation of rumen modifiers is already being exploited. Altogether, the use of bacteriocins may be prospective for inhibiting methanogen populations in the rumen.
GHG emissions have become an increasingly important topic worldwide due to their effects on global warming. The agricultural emissions of methane account for about 60% of the total methane from anthropogenic sources, of which one-fourth arises from enteric fermentation in livestock (Olivier et al., 2005). Globally, livestock produces about 80 million tonnes of enteric methane annually. Most of methane from ruminants originates from microbial fermentation of carbohydrates in the rumen and lower digestive tract, referred to as enteric methane emissions. Methane emissions in ruminants also account for a 2 to 12% of gross energy loss of feeds depending upon the type of diets (Johnson and Johnson, 1995). Therefore, inhibition of methane production

141Climate Resilient Livestock & Production System
in the rumen has been attempted for the past couple of decades to increase the utilization of feed energy for production purposes. The continuous growth of livestock production will further stimulate specialization and industrialization of farming and exacerbate GHG problems in the absence of adequate mitigation stragies (Steinfeld et al., 2006). Hence, there are needs for developing and application of GHG mitigation technologies in livestock production. A number of different strategies to reduce methane production in ruminants have been applied and investigated such as dietary manipulations, vaccines (Boadi et al., 2004; Anderson et al., 2008,), ionophores, plant extracts (Beauchemin et al., 2008; Martin et al., 2010), halogenated methane analogues etc (Patra et al., 2011). However, to date, very few of these strategies have been adopted on farm due to their effect on animal performance.
Livestock and methane productionThe carbohydrates breakdown in the digestive tract of herbivores results in
the production of methane (Adam, 2000). The initial steps are performed either by facultative anaerobic bacteria or by obligate anaerobes. Methanogenic archaea bacteria are obligate anaerobes that are very sensitive to oxygen and prefer environments without any other electron acceptors. They perform the final steps in the fermentation and they convert H2 and CO2 produced by the other organisms to methane or they can convert acetic acid to methane. Nearly, 90-95% of the CO2 produced is converted to methane, and the energy derived is used to fix the remaining CO2 into cellular materials. Strictly, this methane formation action by the methanogens is not a fermentation, but is rather a strange form of respiration.
A number of factors affect methane emission; including diet, barn conditions, and whether or not the cow is lactating, but an average cow produces approximately 550 to 600 liters per day (Adam, 2000).
Role of Indian livestock and methane emissions Indian livestock are large contributors to GHGs and therefore, to the global
warming. These play a significant role in Indian economy by contributing a large amount of food (milk, meat etc.); fibre (wool); skin and manure. Estimated values of methane emissions from domesticated animals varied widely in different reports from 70-220 Million tones/year (Ehhalt, 1974; Baker-Blocker et al., 1977; Sheppard et al., 1982; Blake, 1984).
142 Climate Resilient Livestock & Production System
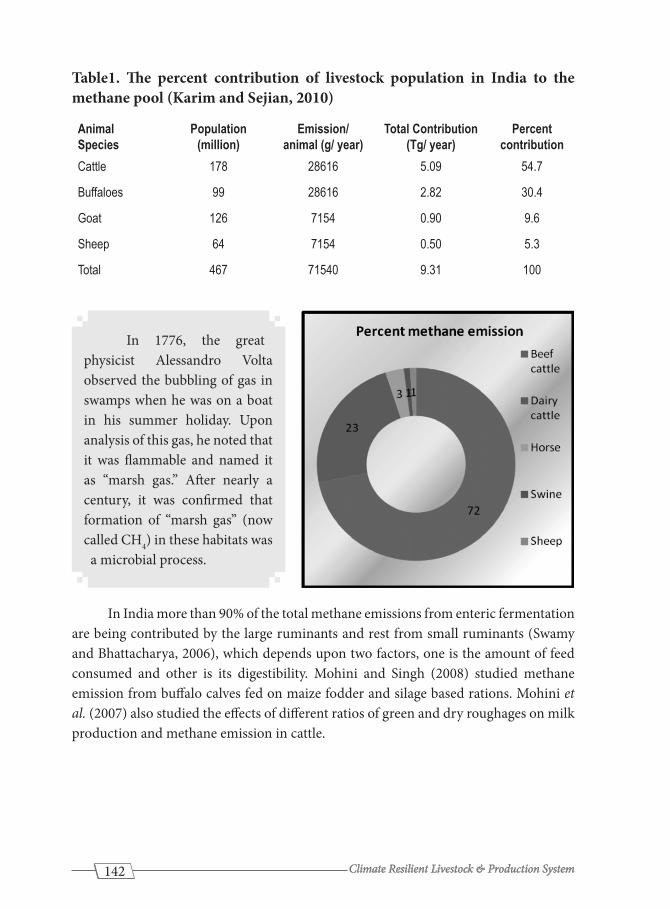
Table1. The percent contribution of livestock population in India to the methane pool (Karim and Sejian, 2010)
Animal Species
Population (million)
Emission/ animal (g/ year)
Total Contribution (Tg/ year)
Percent contribution
Cattle 178 28616 5.09 54.7
Buffaloes 99 28616 2.82 30.4
Goat 126 7154 0.90 9.6
Sheep 64 7154 0.50 5.3
Total 467 71540 9.31 100
In 1776, the great physicist Alessandro Volta observed the bubbling of gas in swamps when he was on a boat in his summer holiday. Upon analysis of this gas, he noted that it was flammable and named it as “marsh gas.” After nearly a century, it was confirmed that formation of “marsh gas” (now called CH4) in these habitats was
a microbial process.
In India more than 90% of the total methane emissions from enteric fermentation are being contributed by the large ruminants and rest from small ruminants (Swamy and Bhattacharya, 2006), which depends upon two factors, one is the amount of feed consumed and other is its digestibility. Mohini and Singh (2008) studied methane emission from buffalo calves fed on maize fodder and silage based rations. Mohini et al. (2007) also studied the effects of different ratios of green and dry roughages on milk production and methane emission in cattle.

143Climate Resilient Livestock & Production System
Table 2. Methane emission per animal per year (Source: NASA’s Goddard Institute for Space Science)
Livestock category Methane emission(kg/ animal/ year)
Livestock category
Methane emission(kg/ animal/ year)
Western cattle 120 Sheep 8.0Non-western cattle 60 Pig 1.5
In 1994, Indian livestock emitted about 10.08 Tg methane due to enteric fermentation, out of which crossbred cattle, indigenous cattle, buffaloes, goats and sheep and other livestock emitted about 4.6, 48.5, 39, 4.7, 1.8 and 1.4 percent, respectively (Singhal et al., 2005). Methane emission from enteric fermentation from Indian livestock ranged from 7.26 to 10.4 MT/year (Garg and Shukla, 2002; US-EPA, 1994). The lowest annual methane production for dairy (180 kg/herd) and non-dairy cattle was reported in Indian subcontinent (Sharma et al., 2006) while comparing with other regions of the world (IPCC, 1996 Guideline for National Green House Gas Inventories reference manual).
Methanogenesis
In ruminants and pseudo-ruminants, the major portion of the methanogenesis occurs in the the rumen, where feeds including fibrous plant structures are fermented primarily to short-chain volatile fatty acids, CO2, hydrogen , and methane by bacteria,
Among 28 genera and 113 species of methanogens known to be present in nature, only seven species have commonly been cultured from the rumen (Janssen and Kirs, 2008). These are Methanobacterium formicicum, Methanobacterium bryantii, Methanobrevibacter ruminantium, Methanobrevibacter millerae, Methanobrevibacter olleyae, Methanomicrobium mobile, and Methanoculleus olentangyi. Methanosarcina spp. has also been cultured from the rumen but are not normally a major part of the archaeal community. Analysis of molecular-based studies reveals that the members of family Methanobacteriaceae (which includes Methanobrevibacter spp., Methanobacterium spp., and Methanosphaera spp.) are the dominant members (30% to 99% of archaea) of the rumen archaea. Members of the order Methanomicrobiales (which includes Methanomicrobium spp.) are less abundant (0% to 54%), and members of the order Methanosarcinales (which includes Methanimicrococcus) are rare (2% to 3%). Usually, CH4 is produced by two types of methanogens, the slow-growing methanogens (generation time about 130 h) that produces CH4 from acetate (e.g., Methanosarcina) and fast growing methanogens (generation time 4–12 h) that reduce CO2 with H2.

144 Climate Resilient Livestock & Production System
protozoa, fungi and methanogens. The methanogens belong to a separate domain archaea in the kingdom of Euryarchaeota and are found in a wide range of other anaerobic environments (Liu and Whitman, 2008). Most rumen methanogens derive energy for their growth through a series of biochemical reduction of CO2 with H2, and some methanogens use acetate and methyl group-containing compounds to produce methane (methanogenesis): 4H2 + CO2 = CH4 + 2H2O
Methanogenesis promotes more complete oxidation of fermented substrates and greater energy recovery by fermenting organisms. It also helps in maintaining the low partial pressure of H2 in the rumen, thus providing a favorable environment for degradation of cell wall carbohydrates (Liu and Whitman, 2008). In the rumen, methanogenesis occurs mostly by the fast-growing methanogens as rumen retention times are too short to permit establishment of the slow growing species (Weimer, 1998). Unlike methanogens, acetogens produce acetate by utilizing H2. They act as important H2 sinks in the hindgut fermentation of mammals. Reductive acetogenesis occurs in the intestine of non-ruminants, sometimes along with methanogenesis and sometimes replacing methanogenesis (Liu and Whitman, 2008).
4H2+2CO2 = CH3COOH+2H2O
Acetogens such as Acetomaculum ruminis have been isolated from the rumen of most of the domestic species (Atwood and McSweeney, 2008) but population densities of acetogens are highly variable. Acetogens are the normal flora in the rumen, but methanogens out compete acetogens as methanogens have lower utilization thresholds for H2 than acetogens and also due to thermodynamically more favorable nature of methanogenesis over acetogenesis (Atwood and McSweeney, 2008).
Bacteriocins Bacteriocins are a heterogeneous group of ribosomally synthesized antibacterial
peptides and proteins. They are usually capable of inhibiting bacteria closely related to the producing strain, which are presumably competing for the same ecological niche, but they may also inhibit a wider range of target organisms. In Gram-positive bacteria these substances consist primarily of cationic, amphipathic peptides which are divided into two main groups: lantibiotic (class I), and non-lantibiotic (class II) bacteriocins (Klaenhammer, 1993; Eijsink et al., 2002; Nes et al., 2007).
The bacteriocins produced by Gram-positive organisms also include large, enzymatically active peptides (class III) and complex peptide-containing molecules (class IV). Bacteriocins produced by Gram-negative bacteria and archaea are similarly diverse (Riley and Wertz, 2002; Gillor et al., 2008). Environmental surveys have

145Climate Resilient Livestock & Production System
identified prevalence levels of bacteriocin-producing isolates of 3 to 90% (Gordon and O’Brien, 2006; Gillor et al., 2008), and it has been suggested that most bacteria produce at least one bacteriocin (Klaenhammer, 1988).
Types and mode of actionThe bacteriocins produced by Gram-negative bacteria fall into two classes, colicins
and microcins. The colicins, first described in E. coli are large, heat labile peptides that generally kill their target organism by membrane permeabilization or by nucleic acid degradation (Riley and Wertz, 2002). Microcins appear to be similar in many respects to the class I and II bacteriocins of Gram-positive bacteria, but kill through a variety of mechanisms (Duquesne et al., 2007). They are not as well characterized as the colicins. The bacteriocins produced by Gram-positive bacteria, historically most extensively studied among the lactic acid bacteria, comprise four distinct classes. The lantibiotics (class I) are small heat-stable peptides. The original translated peptide, or prebacteriocin, is generally significantly larger and undergoes extensive post-translational modification that includes cleavage of a leader peptide and modification of amino acid residues to form dehydroalanine, lanthionine, and/or 3-methyllanthionine residues (Twomey et al., 2002). They generally act by forming pores in the cytoplasmic membrane of the target cell or by interfering with cell wall synthesis, and in some cases have been shown to require interaction with specific target or docking molecules for optimal activity (Gillor et al., 2008).
The class II bacteriocins are also small heat-stable peptides that, like the lantibiotics, are synthesized with a leader peptide. However, post-translational processing is largely limited to cleavage of the leader peptide, and in most cases they do not contain modified amino acid residues (Drider et al., 2006). They act by forming pores in the cytoplasmic membrane of the target cell (Eijsink et al., 2002; Gillor et al., 2008).
Commercial Bacteriocins
Several of the bacteriocin-producing bacteria have been patented, but to the end of 2005 none of them were at the commercialization stage. A mixture of Lactobacillus strains has shown promising results to reduce the fecal shedding of E. coli O157:H7 in cattle and is currently being marketed as a probiotic with the name of Bovamine®. These lactobacilli were originally selected on the basis of its in vitro inhibitory activity against pathogenic E. coli, but there was little evidence that they produced a bacteriocin (Brashears et al., 2003). The most recent patent of bacteriocins for livestock utilization was a really broad invention of a mixture of sorbic acid with bacteriocins or bacteriocin producing bacteria to be included in feed rations (Raczek, 2004).

146 Climate Resilient Livestock & Production System
The class III bacteriocins are large proteins that possess bactericidal enzyme activity (Nilsen et al., 2003), while class IV bacteriocins have lipid or carbohydrate moieties that are required for activity (Vermeiren et al., 2006). Less is known about the distribution or significance of these latter groups.
Methane mitigation through BacteriocinsNisin obtained from Lactobaccilus lactis ssp. lactis, has also been shown to
decrease methane production in vitro. Although the mechanism is still unclear, nisin has been shown to reduce rumen methanogenesis by 36% (Callaway et al., 1997). A combination of nisin & nitrate, an alternative electron receptor, has been reported to reduce methane emissions in sheep (Sar et al., 2005).
Bovicin HC5, another bacteriocin produced by Streptococcus bovis from the rumen, has been reported to suppress methane production by 50% (Lee et al., 2002). In vitro studies of the class I lantibiotic bovicin HC5, produced by S. bovis HC5 revealed that it may be equally as useful as monensin in limiting methane production and amino acid degradation in the rumen (Lee et al., 2002; Lima et al., 2009).
Most studies of bacteriocin effects on rumen function have been less defined. Cell free supernatant from the bacteriocin producer L. plantarum 80 inhibits methanogenesis (Nollet et al. 1998), but the mechanism of this effect has not been determined. The studies on the effects of bacteriocins in vivo are limited and only few specific bacteriocins have been examined, and found effective in rumen environment. First is the enterocin CCM 4231, produced by the rumen isolate Enterococcus faecium CCM 4231 (Laukova and Czikkova, 1998). McAllister and Newbold (2008) reported that bacteriocins could prove effective in directly inhibiting methanogens and redirecting H2 to other reductive bacteria, such as propionate producers or acetogens. Since many lactic acid bacteria produce bacteriocins, reduced methane production observed at very low pH may be due to bacteriocins effects on methanogens may not be through direct pH effect. The potential for rumen produced bacteriocins to suppress methanogenesis is unknown, but their potential as a new generation of rumen modifiers is already being exploited (Teather and Forster, 1998). Altogether, the use of bacteriocins may be prospective for inhibiting methanogen populations in the rumen.
Live microbial supplements for methane mitigationThere is an increasing interest in exploiting probiotics as natural feed additives to
solve problems in livestock production as alternatives to the antibiotics . Probiotics and/or direct-fed microbials are used in the diet of ruminants to improve the health status, rumen fermentation and ultimately the animal performance that could also reduce

147Climate Resilient Livestock & Production System
methane emissions. Only limited information is available on the effect of probiotics on methane production. Although, it is assumed that yeasts reduces methane production by increasing butyrate or propionate (Lila et al., 2004) reducing protozoa (Newbold et al., 1998), promoting acetogenesis (Chaucheyras et al., 1995) and improving animal productivity. Probiotics improved productivity by 7 to 8 percent resulting in reduced methane per unit of product in cattle. Joblin (1999) and Klieve (2007) reported the use of probiotics in mitigation of methane from ruminants. Acetogenic bacteria in in vitro mixed rumen cultures resulted in a modest decrease in methane production (lopez et al., 1999). Lynch and Martin (2002) reported that 20 % decrease in methane after 48 h of incubation of mixed rumen microbes in presence of alfalfa and live yeasts. Mwenya et al. (2004) reported that a yeast Trichosporon sericeum (4 g/ day) decreased methane by 10 % in sheep fed on a roughage-based diet and also reported that sheep produced significantly less methane when 0.4 % yeasts were included in a basal hay and 30 % concentrate diet. However, short term incubations are inappropriate to study the effects of yeasts on rumen fermentation. The effects of S. cerevisiae are mediated through an effect on the numbers and activity of microbes in rumen and periods longer than 24 to 48 h may be required to fully realize the effects. Yeast culture probably stimulates acetogens to compete with methanogens or to co-metabolize hydrogen thus, decreasing methane formation (Chaucheyras et al., 1995). A. oryzae and S. cerevisiae increased methane (Martin and Nisbet, 1990) while Mathieu et al. (1996) reported that S. cerevisiae could not affect methane release in vivo. These contradictory results on methane might be due to the strain differences between yeasts and the type of diets (Newbold and Rode, 2006). Methane reduction effects of probiotics are not consistent; hence, there is a need to identify the dietary situation in which probiotics can give reliable results. Overall, more research is required to use probiotics in livestock for methane reduction and improving the micro flora.
ConclusionThe utilization of bacteriocins or bacteriocin-producing bacteria and probiotics in
livestock is a field with enormous possibilities for both research and commercialization. As more countries develop antibiotic-limiting policies, the need for alternative antimicrobial and the use of probiotics will probably be the main driving force to continue identifying novel bacteriocins/probiotics and testing existing ones. Because of the relative specificity of bacteriocins as compared with antibiotics, it can be anticipated that the identification of broader spectrum bacteriocins will be an active research endeavor. Similarly, it is fore seen that researchers will likely utilize combinations of bacteriocins to obtain a broader spectrum for target organisms.

148 Climate Resilient Livestock & Production System
AcknowledgementAuthors are thankful to National Initiative on Climate Resilient Agriculture
(NICRA) project of Indian Council of Agricultural Research (ICAR), New Delhi, for providing the support in developing the manuscript.
ReferencesAdam, D. 2000. Tracking methane emissions from cows just got a little easier, URL:
http://www.nature.com/nsu/000907/000907-6.html, date: Sep 5, 2000.
Anderson, R.C., Krueger, N.A., Stanton, T.B., Callaway, T.R., Edrington, T.S., Harvey, R.B., Jung, Y.S. and Nisbet, D.J. 2008. Effects of select nitrocompounds on in vitro ruminal fermentation during conditions of limiting or excess added reductant. Bioresour. Technol., 99: 8655 – 8661.
Atwood, G. and McSweeney, C.S. 2008. Methanogen genomics to discover targets for methane mitigation technologies and options for alternative H2 utilization in the rumen. Aus. J. Exp. Agri., 48: 28 – 37.
Baker-Blocker, A., Donohue, T.M. and Maney K.H. 1977. Methane flux from wetland areas. Tellus, 29: 245 – 250.
Boadi, D., Benchaar, C., Chiquette, J. and Masse, D. 2004. Mitigation strategies to reduce enteric methane emissions from dairy cows: Update review. Canad. J. Ani. Sc., 84: 319 - 335.
Beauchemin, K.A., Kreuzer, M., O’Mara, F. and McAllister, T.A. 2008. Nutritional management for enteric methane abatement: A review. Aus. J. Exp. Agri., 48: 21 – 27.
Blake, D.R. 1984. Increasing concentrations of atmospheric methane, 1979-1983. Ph.D. Thesis, University of California, Irvine, CA, pp. 213.
Brashears, M.M., Galyean, M.L., Loneragan, G.H., Mann,J.E., and Killinger-Mann, K. 2003. Prevalence ofEscherichia coli O157:H7 and performance by beeffeedlot cattle given Lactobacillus direct-fed microbials. J. Food Prot., 66: 748 – 754.
Drider, D., Fimland, G., Hechard, Y., McMullen, L.M. and Prevost, H. 2006. The continuing story of class IIa bacteriocins. Microbiol. Mol. Biol. Rev., 70: 564 - 582.
Callaway, T.R., Alexandra, M.S., Carneiro, De.Melo. and Russell, J.B. 1997. The effect of nisin and monensin on ruminal fermentations in-vitro. Curr. Microbiol., 35: 90 – 96.

149Climate Resilient Livestock & Production System
Chaucheyras, F.G., Fonty, G., Bertin, G. and Gouet, P. 1995. In-vitro H2 utilization by a ruminal acetogenic bacterium cultivated alone or in association with an archaea methanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. App. Environ. Microbiol., 61: 3466 – 3467.
Duquesne, S., Petit, V., Peduzzi, J. and Rebuffat, S. 2007. Structural and functional diversity of microcins, gene-encoded antibacterial peptides from enterobacteria. J. Mol. Microbiol. Biotechnol., 13: 200 - 209.
Eijsink, V.G., Axelsson, L., Diep, D.B., Havarstein, L.S., Holo, H. and Nes, I.F. 2002. Production of class II bacteriocins by lactic acid bacteria; an example of biological warfare and communication. A. Van. Leeuw., 81: 639 - 654.
Ehhalt, D.H. 1974. The atmospheric cycle of methane. Tellus, 26: 58-70.
Garg, A. and. Shukla, P.R. 2002. Emission Inventory of India. Tata McGraw Hill, New Delhi, pp: 84-89.
Gillor, O., Etzion, A., Riley, M.A. 2008. The dual role of bacteriocins as anti- and probiotics. Appl. Microbiol. Biotechnol., 81: 591 - 606.
Gordon, D.M. and O’Brien, C.L. 2006. Bacteriocin diversity and the frequency of multiple bacteriocin production in Escherichia coli. Microbiol., 152: 3239-3244.
IPCC. 1996. Guideline for National Green House Gas Inventories reference manual, pp. 4-11.
Janssen, P.J. and Kirs, M. 2008. Structure of the archaeal community of the rumen. App. and Environ. Microbiol., 74: 3619 – 3625.
Joblin, K.N. 1999. Ruminal acetogens and their potential to lower ruminant methane emissions. Austral. J. Agri. Research., 50: 1307 – 1313.
Johnson, K.A. and Johnson, D.E. 1995. Methane emissions from cattle. J. Ani. Sci., 73: 2483 - 2492.
Karim, S.A. and Sejian, V. 2010. Sheep productivity adapting to climate change. Proceedings of ‘National Symposium on Climate Change and Livestock Productivity in India’ October 7-8, at NDRI, Karnal, 107-118.
Klaenhammer, T.R. 1993. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev., 12: 39 - 86.
Klaenhammer, T. R. 1988. Bacteriocins of lactic acid bacteria. Biochimie., 770: 337- 349.

150 Climate Resilient Livestock & Production System
Klieve, A.V. and Joblin, K. 2007. Comparison in hydrogen utilisation of ruminal and marsupial reductive acetogens. In R. Kennedy, (eds) 5 Year Research Progress Report 2002 - 2007 , The Pastoral Greenhouse Gas Research Consortium, Well- ington, New Zealand, pp. 34 - 35.
Laukova, A. and Czikkova, S. 1998. Inhibition effect of enterocin CCM 4231 in the rumen fluid environment. Lett. Appl. Microbiol., 26: 215 - 218.
Lee, S.S., Hsu, J.T., Mantovani, H.C. and Russell, J.B. 2002. The effect of bovicin HC5, a bacteriocin from Streptococcus bovis HC5, on ruminal methane production in-vitro. FEMS Microbiol. Lett., 217: 51 - 55.
Lila, Z.A., Mohammed, N., Tatsuoka, Ajisaka, N., Kanda, S., Kurokawa, Y. and Itabashi, H. 2004. Effect of cyclodextrin diallyl maleate on methane production, ruminal fermentation and microbes in vitro and in vivo. Ani. Sci. Journ., 75: 15 – 22.
Lima, J.R., Ribon, Ade O., Russell, JB. and Mantovani, H.C. 2009. Bovicin HC5 inhibits wasteful amino acid degradation by mixed ruminal bacteria in-vitro. FEMS Microbiol. Lett., 292: 78 - 84.
Liu, Y., and Whitman, W.B. 2008. Metabolic, phylogenetic, and ecological diversity of the methanogenic archaea. Anna. NewYork Acad. Sci., 1125: 171 – 189.
Lopez, S., McIntosh, F.M., Wallace, R.J. and Newbold, C.J. 1999. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Ani. Feed Sci. and Technol., 78: 19.
Lynch, H.A. and Martin, S.A. 2002. Effects of Saccharomyces cerevisiae culture and Saccharomyces cerevisiae live cells on in vitro mixed ruminal microorganism fermentation. J. Dairy Sci., 85: 2603 – 2608.
Martin, C., Morgavi, D. and Doreau, M. 2010. Methane mitigation in ruminants: From microbe to the farm scale. Animal, 4: 351 - 365.
Martin, S.A. and Nisbet, D.J. 1990. Effects of Aspergillus oryzae fermentation extract on fermentation of amino acids, bermudagrass and starch by mixed ruminal micro-organisms in vitro. J. of Ani. Sci., 68: 2142 - 2149.
Mathieu, F., Jouany, J.P., Senaud, J., Bohatier, J., Berthin, G. and Mercier, M. 1996. The effect of Saccharomyces cerevisiae and Aspergillus oryzae on fermentations in the rumen of faunated and defaunated sheep; protozoal and probiotic interactions. Reprod. N. Develop., 36: 271 – 287.

151Climate Resilient Livestock & Production System
McAllister, T.A., and Newbold, C.J. 2008. Redirecting rumen fermentation to reduce methanogenesis. Aust. J. Exp. Agricul., 48: 7- 13.
Mohini, M. and Singh, G.P. 2008. Methane emission from buffalo calves fed on maize fodder and silage based rations. Indian Journal Animal Nutrition, 25(4): 313-317.
Mohini, M., Mani, V. and Singh, G.P. 2007. Effect of different ratios of green and dry roughages on milk production and methane emission in cattle. Indian Journal of Animal Nutrition, 77 (1): 79-82.
Mwenya, B., Santoso, B., Sar, C., Gamo, Y., Kobayashi, T., Arai, I. and Takashi, J. 2004. Effects of including 1,4-galactooligosaccharides, lactic acid bacteria or yeast culture on methanogenesis as well as energy and nitrogen metabolism in sheep. Ani. Feed Sci. Technol., 115: 313 – 326.
NASA’s Goddard Institute for Space Studies. www.giss.nasa.gov/
Newbold, C.J. and Rode, L.M. 2006. Dietary additives to control methanogenesis in the rumen. In Soliva CR, Takahashi J and Kreuzer M Eds. Greenhouse Gases and Animal Agriculture: An Update pp. 138– 147. Amsterdam: Elsevier. International Congress Series No. 1293.
Newbold, C.J., Melntosh, F.M. and Wallace, R.J. 1998. Changes in the microbial population of a rumen-simulating fermenter in response to live yeast. Canad. J. Ani. Sci., 78: 241 - 244.
Nilsen, T., Nes, I.F. and Holo, H. 2003. Enterolysin A, a cell wall-degrading bacteriocin from Enterococcus faecalis LMG 2333. Appl. Envron. Microbiol., 69: 2975 - 2984.
Nes, I.F., Diep, D.B. and Holo, H. 2007. Bacteriocin diversity in Streptococcus and Enterococcus. J. Bacteriol., 189: 1189 - 1198.
Nollet, L., Mbanzamihigo, L., Demeyer, D. and Verstraete, W. 1998. Effect of the addition of Peptosteptococcus productus ATCC 35244 on reductive acetogenesis in the ruminal ecosystem after inhibition of methanogenesis by cell-free supernatantm of Lactobacilus plantarum 80. Anim. Feed Sci. Technol., 71: 49 - 66.
Olivier, J.G.J., van Aardenne, J.A., Dentener, F., Ganzeveld, L. and Peters, J.A.H.W. 2005. Recent trends in global greenhouse gas emissions: Regional trends and spatial distribution of key sources. In A. van Amstel (eds) Non-CO2 greenhouse gases (NCGG-4). Rotterdam: Millipress. pp. 325–330.

152 Climate Resilient Livestock & Production System
Patra, A.K., Kamra, D.N., Bhar, R., Kumar, R. and Agarwal, N. 2011. Effect of Terminalia chebula and Allium sativum on in vivo methane emission by sheep. J. Anim. Physiol. Anim. Nutr., 95: 187–191.
Raczek, N. 2004. Bacteriocin-containing sorbic acid product as addition to feedstuffs in agricultural livestock rearing (USA patent 6780447).
Riley, M.A. and Wertz, J.E. 2002. Bacteriocin diversity: ecological and evolutionary perspectives. Biochimie., 84: 357 - 64.
Sar, C., Mwenya, B., Santoso, B., Takaura, K., Morikawa, R., Isogai, N., Asakura, Y., Toride, Y. and Takahashi, J. 2005. Effect of Escherichia coli wild type or its derivative with high nitrite reductase activity on in vitro ruminalmethanogenesis and nitrate/nitrite reduction. J. Ani. Sci., 83: 644 – 652.
Sharma, S., Bhattacharya, A. and Garg, A. 2006. Green house gas emission from India: A perspective. Curr. Sci., 90(3): 326-333.
Sheppard, J.C., Westberg, J.F., Hopper, Ganesan, K. and Zimmerman, P. 1982. Inventory of global methane sources and H. their production rates. J. Geophys. Res., 87: 1305-1312.
Singh, S.N. Perturbing the heat Radiation Balance of the Earth. Report, NBRI, India.
Singhal, K.K., Mohini, M., Jha, A.K. and Gupta, P.K. 2005. Methane emission estimates from enteric fermentation in Indian livestock: Dry matter intake approach. Current Science, 88: 119-127.
Steinfeld, H., Costales, A., Rushton, J., Scherf, B., Bennett, T., and Hall, D. 2006. Livestock report 2006. Rome: FAO.
Swamy, M. and Bhattacharya, S. 2006. Budgeting anthropogenic greenhouse gas emission from Indian livestock using country-specific emission coefficients. Curr. Sci., 91(10): 1340-1353.
Teather, R.M. and Forster, R.J. 1998. Manipulating the rumen microflora with bacteriocins to improve ruminant production. Canad. J. Ani. Sci., 78: 57 - 69.
Twomey, D., Ross, R.P., Ryan, M., Meaney, B. and Hill, C. 2002. Lantibiotics produced by lactic acid bacteria: structure, function and application. A. Van. Leeuw., 82: 165 - 185.
US-EPA. 1994. International Anthropogenic Methane Emission. Washington DC, EPA-23-R-93 - 010.
