contribution of the nucleus to the mechanical properties of endothelial cells
TRANSCRIPT

Journal of Biomechanics 35 (2002) 177–187
Contribution of the nucleus to the mechanical properties ofendothelial cells
Nathalie Caille, Olivier Thoumine, Yanik Tardy, Jean-Jacques Meister*
Biomedical Engineering Laboratory, Swiss Federal Institute of Technology, PSE-A, 1015 Lausanne, Switzerland
Accepted 22 September 2001
Abstract
The cell nucleus plays a central role in the response of the endothelium to mechanical forces, possibly by deforming during cellularadaptation. The goal of this work was to precisely quantify the mechanical properties of the nucleus. Individual endothelial cellswere subjected to compression between glass microplates. This technique allows measurement of the uniaxial force applied to the cell
and the resulting deformation. Measurements were made on round and spread cells to rule out the influence of cell morphology onthe nucleus mechanical properties. Tests were also carried out with nuclei isolated from cell cultures by a chemical treatment. Thenon-linear force–deformation curves indicate that round cells deform at lower forces than spread cells and nuclei. Finite-element
models were also built with geometries adapted to actual morphometric measurements of round cells, spread cells and isolatednuclei. The nucleus and the cytoplasm were modeled as separate homogeneous hyperelastic materials. The models simulate thecompression and yield the force–deformation curve for a given set of elastic moduli. These parameters are varied to obtain a best fit
between the theoretical and experimental data. The elastic modulus of the cytoplasm is found to be on the order of 500 N/m2 forspread and round cells. The elastic modulus of the endothelial nucleus is on the order of 5000 N/m2 for nuclei in the cell and on theorder of 8000 N/m2 for isolated nuclei. These results represent an unambiguous measurement of the nucleus mechanical propertiesand will be important in understanding how cells perceive mechanical forces and respond to them. r 2002 Elsevier Science Ltd. All
rights reserved.
Keywords: Cellular mechanics; Microplate manipulation; Cell spreading; Nucleus
1. Introduction
Hemodynamic factors are known to influence bloodvessel pathology (Nerem, 1992). In particular, athero-sclerosis lesions develop preferentially in regions whereparticular fluctuations in wall shear stress occur (Kuet al., 1985). The endothelium, in direct contact withblood flow, is likely to transduce such mechanical forcesinto biochemical signals to the inner wall of bloodvessels (Gimbrone et al., 1995). Endothelial cells areknown to adapt their shape and function in response toapplied shear stress, as demonstrated by many in vivoand in vitro studies (Davies, 1995). Stress and strainbeing primary triggering signals for cellular remodeling,it is important to know the mechanical properties of
endothelial cells in order to better understand theirresponse to flow.
Studies using micropipet suction, magnetocytometryor atomic force microscopy have shown that the cellularstiffness increases when endothelial cells are allowed tospread (Wang and Ingber, 1994) or after adaptation toflow (Sato et al., 1987), both conditions being associatedwith a remodeling of the cytoskeleton. These measure-ments also show regional variations (Miyazaki andHayashi, 1999; Sato et al., 2000). However, these reportsconsider an overall cellular stiffness and do notdistinguish the relative contribution of the variousstructural elements of the cell, in particular that of thenucleus.
Deformation of the nucleus of endothelial cells couldvery well play a role in shear stress transduction. Forexample, a theoretical study has shown that endothelialcells minimize the force on their nuclei during adapta-tion to flow (Hazel and Pedley, 2000). The nucleus iscapable of large deformations concomitant to changes in
*Corresponding author. Tel.: +41-21-693-8345; fax: +41-21-693-
8305.
E-mail address: [email protected] (J.-J. Meister).
0021-9290/02/$ - see front matter r 2002 Elsevier Science Ltd. All rights reserved.
PII: S 0 0 2 1 - 9 2 9 0 ( 0 1 ) 0 0 2 0 1 - 9

cellular shape (Maniotis et al., 1997a). For example, thenucleus shrinks when cells are compressed (Guilak,1995), extends when cells are stretched (Caille et al.,1998), spreads when cells flatten on a substrate (Ingber,1990) and conversely rounds up when cells are inducedto contract (Sims et al., 1992; Pienta and Coffey, 1992).In addition, a slight elongation of the nucleus occurswhen cells are exposed to flow (Galbraith et al., 1998).The nucleus is filled with DNA and is connected to thecytoskeleton. Therefore, deformation of the nucleusaccompanying cytoskeletal remodeling could result inaltered DNA packing (Maniotis et al., 1997b) and thusaffect gene regulation (Gimbrone et al., 1997).
Only limited information exists on the mechanicalproperties of the nucleus of endothelial cells. By pullingon microspheres attached to integrins on endothelial cellmembranes, Maniotis et al. (1997a) found that thenucleus inside the cell is about 9 times stiffer than thecytoplasm. Studies on mechanically and chemicallyisolated nuclei of neutrophils using micropipet aspira-tion have shown that the elasticity and viscosity of thenucleus is about 10 times larger than that of thecytoplasm (Dong et al., 1991). Similar experiments onnuclei of articular chondrocytes have shown that theyare 3–4 times stiffer and nearly twice as viscous as thecytoplasm (Guilak et al., 2000). All of these estimationsare based on local strains applied to the cell or to thenucleus. However, no absolute value of mechanicalproperties of endothelial cells or isolated nuclei ofendothelial cells subjected to a global strain is currentlyavailable.
The objective of this study was to precisely quantifythe mechanical resistance of the nucleus of endothelialcells. Mechanical tests were carried out on individualcells and isolated nuclei using a recently developedmicroplate manipulation technique. Experimental datawere interpreted with simulations from a finite elementprogram, leading to the intrinsic material properties ofthe cytoplasm and nucleus.
2. Materials and methods
2.1. Cells and nuclei
Freshly excised bovine aortas were obtained from alocal slaughterhouse. Endothelial cells were isolatedusing the method of Booyse et al. (1975). Cells werecultured in Dulbecco’s Modified Eagle’s Mediumsupplemented with 10% fetal calf serum and 50 mg/mlgentamycin (culture medium). Cells were passaged everyweek and finally used at passages 4–12. Before theexperiments, cells were detached with 0.05% trypsin–EDTA and resuspended in culture medium without fetalcalf serum containing 20 mM Hepes (observationmedium). Nuclei were extracted from confluent en-
dothelial cells by treatment with a non-ionic detergent(Igepal) and citric acid as described by Thoumine et al.(1999a), and then suspended in observation medium.The suspension of cells or nuclei was then placed in anobservation chamber for manipulation.
2.2. Micromanipulation system
The set-up has been described in detail previously(Thoumine and Ott, 1997; Thoumine et al., 1999a, b)and a diagram of the micromanipulation system is givenin Fig. 1. Briefly, it consists of an inverted microscopeplaced on an anti-vibration table and equipped on bothsides with micromanipulators. Microplates pulled fromglass bars are held by steel arms and maneuvered withthe micromanipulators. No coating or other treatment isapplied to the microplates. In practice, a rigid micro-plate is used to capture a cell, and is displaced finely inthe transverse axis using a piezoelectric translator. Onthe other side, a more flexible microplate is used; itsdeflection in response to load is used to estimate theuniaxial force. The deformation sequences are filmedunder bright-field illumination using a 40� /0.75 objec-tive, a 1.6� lens and a CCD camera. The video imagesare then digitized on a computer. Using image analysissoftware, the deflection of the flexible microplate ismeasured by tracking the image intensity profile. Thisgives pixel accuracy corresponding to a resolution of70.12 mm. The force applied on the cell is calculated bymultiplying the deflection of the flexible microplate byits rigidity, obtained after calibration with a referencemicroneedle (Thoumine et al., 1999a, b). The error inforce measurements is approximately 10%.
2.3. Mechanical test
Experiments were carried out at room temperature onthree bodies: round cells, spread cells and isolatednuclei. The untreated rigid microplate is placed in theobservation chamber and positioned beneath a sus-pended endothelial cell or a nucleus settling down due togravity. The microplate is maintained in this positionuntil the cell or the nucleus adheres to it (o2 min). Theobservation medium is then changed for medium with10% fetal calf serum containing 20 mM Hepes. Forround cells and nuclei, the mechanical test is done
rigid microplate
flexible microplate
piezo-driventranslation
fixed
cell nucleus
support
Fig. 1. Sketch of the experimental set-up.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187178

immediately. For the so-called spread cells, the chamberand the rigid microplate are connected using a custom-made system and transferred to an incubator (371C, 5%CO2) (Thoumine et al., 1999a). After a 45 min incuba-tion period, sufficient to promote cell spreading, thechamber is brought back to the microscope and themicroplate re-attached to the steel arm. The microplateis rotated by 901 around its axis to provide a side-view.After initial contact with the flexible microplate,compression is imposed by displacing the rigid micro-plate carrying the cell or nucleus by steps of 2.5 mm every10 s towards the flexible microplate (Fig. 2). The rigidityof the flexible microplate is chosen in order to obtain acompression of the cell or nucleus on the order of 1.5 mmand a deflection of the flexible microplate about 1 mm forthe first 2.5 mm displacement of the rigid microplate. Inthis manner, 4–7 points, depending on the initial heightof the cell or the nucleus, can be acquired for eachstress–strain curve measured. This corresponds to a totalcompression of 50–60%. Relaxation is done by with-drawing the rigid microplate using the same timesequence. The total cycle (loading and unloading) lastsfrom 1.5 to 3 min, depending on the cell. The mechanicaltest is carried out in the shortest possible time in order tomeasure only the passive mechanical properties and
avoid cellular adaptation. It was noted that the appliedforces and resulting cell or nuclei deformation remainconstant throughout each 10 s step.
2.4. Morphometry
The cell height and the length of the contact linebetween the cell or nucleus and the microplate weredetermined by analysis of side-view images at eachcompression step. The cell or nucleus deformation isdefined as the relative reduction in height, i.e. 100 �ðH=H0 � 1Þ; where H is the height in the deformed state,H0 the height in the initial undeformed state and theresult is expressed in %. Moreover, the principaldimensions of the cells and nuclei on microplates weremeasured from top views before starting the compres-sion (zero stress state). To obtain the relative position ofthe nucleus within the cell, additional cells from aculture well were treated with the fluorescent DNAmarker Syto 13, as described by Caille et al. (1998).These cells were not compressed between the micro-plates but the same protocol as for compressionmeasurements was then followed except that the cellswere allowed to settle down into the deformationchamber, not on a microplate, for visualization with aconfocal laser scanning microscope (Zeiss LSM 410).
3. Finite-element model
3.1. Assumptions and geometry
An endothelial cell is a very complex organizedsystem. Its principal structural elements are the cyto-plasm, which is composed of the hyaloplasm and thecytoskeleton, the membrane and different organelles ofwhich the most prominent is the nucleus. In thistheoretical model, the system is simplified to consideronly two materials, one for the nucleus and one for therest of the cell, here after called cytoplasm. Thisselection is based on the importance of these elementsin the mechanical properties of the endothelial cell. Inorder to obtain a constitutive law that well describes thebehavior of the cell, it is assumed that both materials arecontinuous, homogeneous, incompressible, isotropicand hyperelastic. In addition, the cell is considered aspassive; this implies that the mechanical properties ofthe cell are considered before any remodeling occurs.Because the deformation sequence is relatively short,this assumption is rather valid. Three finite elementmeshes were built to represent the three different bodies,one for round cells, one for spread cells, and one forisolated nuclei (Fig. 3a, d and g). The shapes anddimensions of each model are based on averagemorphometric dimensions measured for all round cells,spread cells and isolated nuclei. The geometries of all
Fig. 2. Digitized video images of a round cell (a–c), a spread cell (d–f),
and an isolated nucleus (g–i) exposed to compression. After adhering
to a rigid microplate (thick bottom contour), the body is pushed
against a flexible microplate (thin upper contour). Cells and nuclei are
shown at rest, i.e. at the start of the experiment (left panels: a, d, g),
and at two different stages of compression. Note the deformation of
the cells and nucleus, and the concomitant upward deflection of the
flexible microplate, which is used to estimate the applied force.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187 179

models are chosen axisymmetric. The geometry of thespread cell is defined as if the spreading is circular with aradius chosen to completely encircle the cell. Measure-ments of isolated nuclei and those in the cells show thatthey have either a pure ellipsoidal shape or the form ofan ellipse revolved about its minor axis. The latter formis chosen for implementation in the finite element model.For a majority of round and spread cells, the nucleusappears to be centered in the cell when seen from a topview. When viewed from the side, the nucleus appears tobe located in the lower portion of round cells and tooccupy almost the entire height of spread cells. Themodel geometries are based on these observations. Theboundary nodes that are initially in contact with therigid microplate are fixed to the microplate while allother boundary nodes are able to contact the flexible orrigid microplates but not adhere to them. For eachgeometry, the assumptions and the constitutive laws forthe materials are the same.
3.2. Material constitutive law
The assumption that the materials used in the modelare hyperelastic implies that these materials are elasticand that the stress tensor, S; can be derived from a strainenergy density function, W ; which is a scalar function ofthe strain. This can be expressed mathematically by
S ¼qWqe
¼ 2qWqC
; ð1Þ
where e is the material strain tensor and C is the rightCauchy–Green tensor.
The strain energy density function of the Mooney–Rivlin law is used to represent the stress–strain behaviorof a nearly incompressible material (Truesdell and Noll,1992). It has the additional advantage of relying on onlytwo material constants. Using Eq. (1), the stress tensorderived from the Mooney–Rivlin strain energy densityfunction may be expressed as
SðCÞ ¼ �pC�1 þ m1
2þ b
� �Cþ m
1
2� b
� �I; ð2Þ
where p is the hydrostatic pressure, m is the shearmodulus, b a constant and I is the identity tensor.
This constitutive law is integrated in the finite elementsoftware (ANSYS version 5.4, Swanson AnalysisSystems, Houston, PA, USA) run on a personalcomputer.
Two parameters, a1 and a2; are required to character-ize the material. These are related to m and b as follows:
a1 ¼1
2m
1
2þ b
� �; ð3Þ
a2 ¼1
2m
1
2� b
� �: ð4Þ
These parameters do not have a simple physicalinterpretation and obtaining them experimentally re-quires a large number of experiments that are not allpossible at the scale of the cell. In order to express these
Fig. 3. Finite element simulations of compression experiments. Model geometries for: (a) round cells, (d) spread cells, and (g) isolated nuclei.
Corresponding deformed meshes calculated at 30% compression (b, e, h) and 50% compression (c, f, i). The nucleus within the cell is outlined with a
bold line.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187180

parameters as a function of known parameters, a term-by-term identification of the Mooney–Rivlin law(Eq. (2)) to the Kirchhoff–Saint Venant constitutivelaw is carried out. From this operation, a newexpression of the strain tensor is obtained from whichthe strain energy density function can be expressed as
W ¼ a1ðI1 � 3Þ; ð5Þ
where a1 ¼ m=2 ¼ E=4ð1 þ nÞ; I1 is the first invariant, nis Poisson’s ratio (=0.5 due to the assumption ofincompressibility) and E is the modulus of elasticity(unknown).
This expression of the Mooney–Rivlin strain energydensity function corresponds to that of a neo-Hookeanmaterial (Zahalak et al., 1990). In addition, it is afunction of a single parameter depending only on themodulus of elasticity of the material, E:
The cytoplasm and the nucleus materials aremodeled with this behavioral law in order to findthe two unknown parameters in the model, the elasticmodulus of the cytoplasm Ecyto, and that of the nucleusEnucleus:
Ecyto ¼ 4ð1 þ ncytoÞa1cyto; ð6Þ
Enucleus ¼ 4ð1 þ nnucleusÞa1nucleus: ð7Þ
3.3. Fitting the model with experimental data
The values of Ecyto and Enucleus are manually suppliedfor the three model geometries described above.Those values are varied from 10 to 106 N/m2 by regularincrements allowing 4 values per order of magnitude (i.e.1000, 2500, 5000 and 7500 for the order of magnitude103) in order to cover the range of experimental andtheoretical values found in the literature. The compres-sion of the three models is done by increments of 0.5 mm,resulting in flattening of the model cells and nuclei,similar to that observed in the experiments (Fig. 2). Theprogram gives the average force and the reduction inheight is calculated. The corresponding force–deforma-tion curve is plotted and compared to the experimentalresults. To identify the parameter values that mostclosely match the simulation to the experiment, adeviation parameter, d; is used (Schmidt et al., 1994).This parameter is a numerical representation of thedifference between the simulation and the experimentand is defined as
d ¼Pn
i¼1 ððFexpðiÞ � FsimðiÞÞ=FexpðiÞÞ2
n; ð8Þ
where n is the number of measurements for anexperiment, FexpðiÞ is the ith force measurementand FsimðiÞ is the corresponding simulated forceobtained by linear interpolation. Graphs of d versusEnucleus are plotted for each value of Ecyto and used todetermine the values of these parameters that minimize
d; the difference between the simulation and theexperiment.
4. Results
4.1. Experimental measurements
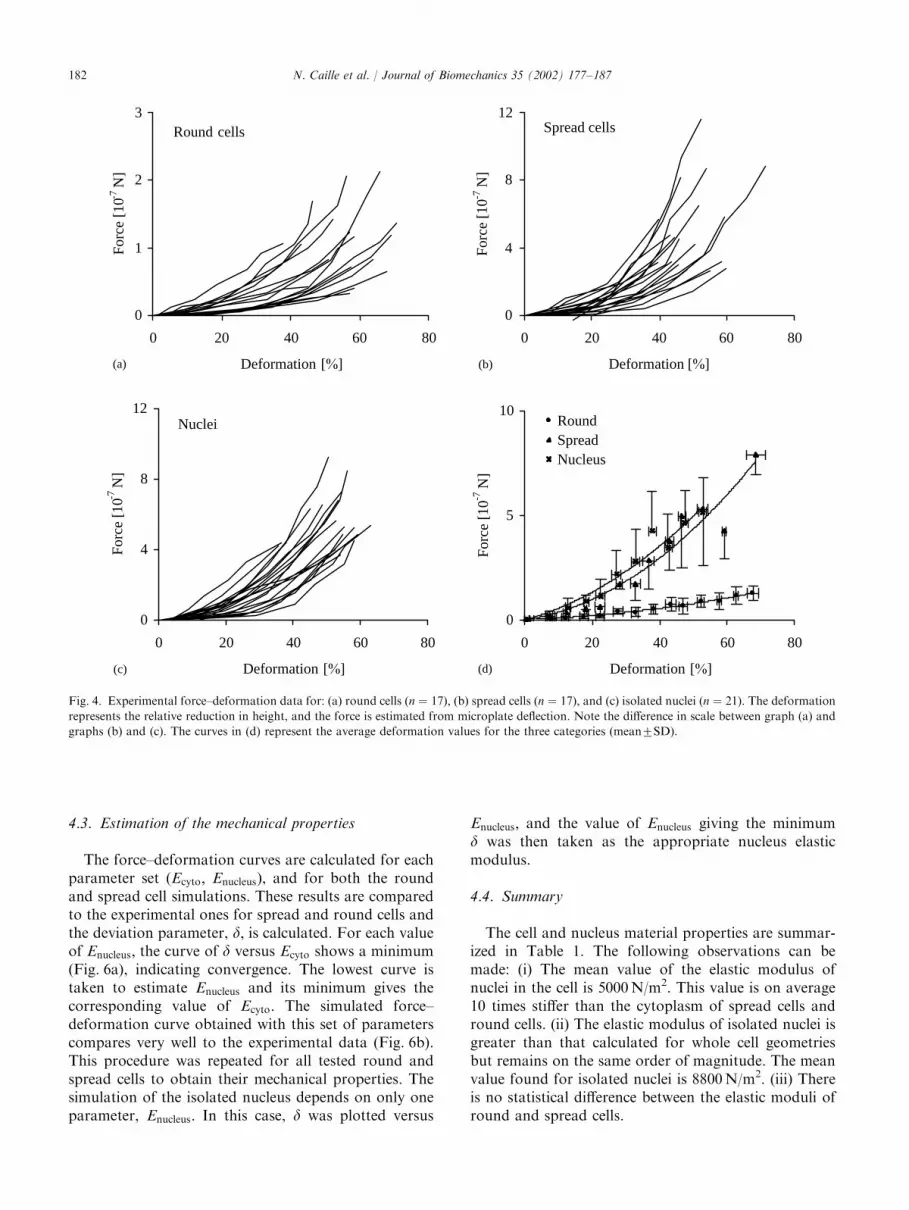
When compressed between the microplates, both thecells and the nuclei are immediately deformed at thetime scale of the experiment. This deformation ischaracterized by a reduction of their height and anincrease in their base (Fig. 2). For spread cells andisolated nuclei, there are only small residual deforma-tions at the end of the mechanical tests and a negligiblehysteresis in the loading/unloading cycle (not shown),indicating essentially no viscous or plastic behavior.However, measurable residual deformations and ahysteresis in the force–deformation relationship for aloading–unloading cycle are observed for round cells(not shown). For both the cells and the nuclei, theimposed compressive force increases non-linearly as afunction of the height reduction, i.e. the force necessaryto deform the cells and nuclei increases non-linearly astheir deformation increases (Fig. 4a–c). This behavior isobserved for each individual trace, as well as for theaverages (Fig. 4d). Considering cell-to-cell variation andthe precision in the force measurements, one can assumethat the spread cells and the isolated nuclei have thesame behavior. Round cells, on the other hand, requireforces approximately one-fourth of those required forspread cells and isolated nuclei for the same amount ofdeformation. For example, the force required to create adeformation of 50% is approximately 10�7 N for roundcells, 4� 10�7 N for spread cells and 4.5� 10�7 N forisolated nuclei (Fig. 4d).
4.2. Simulations
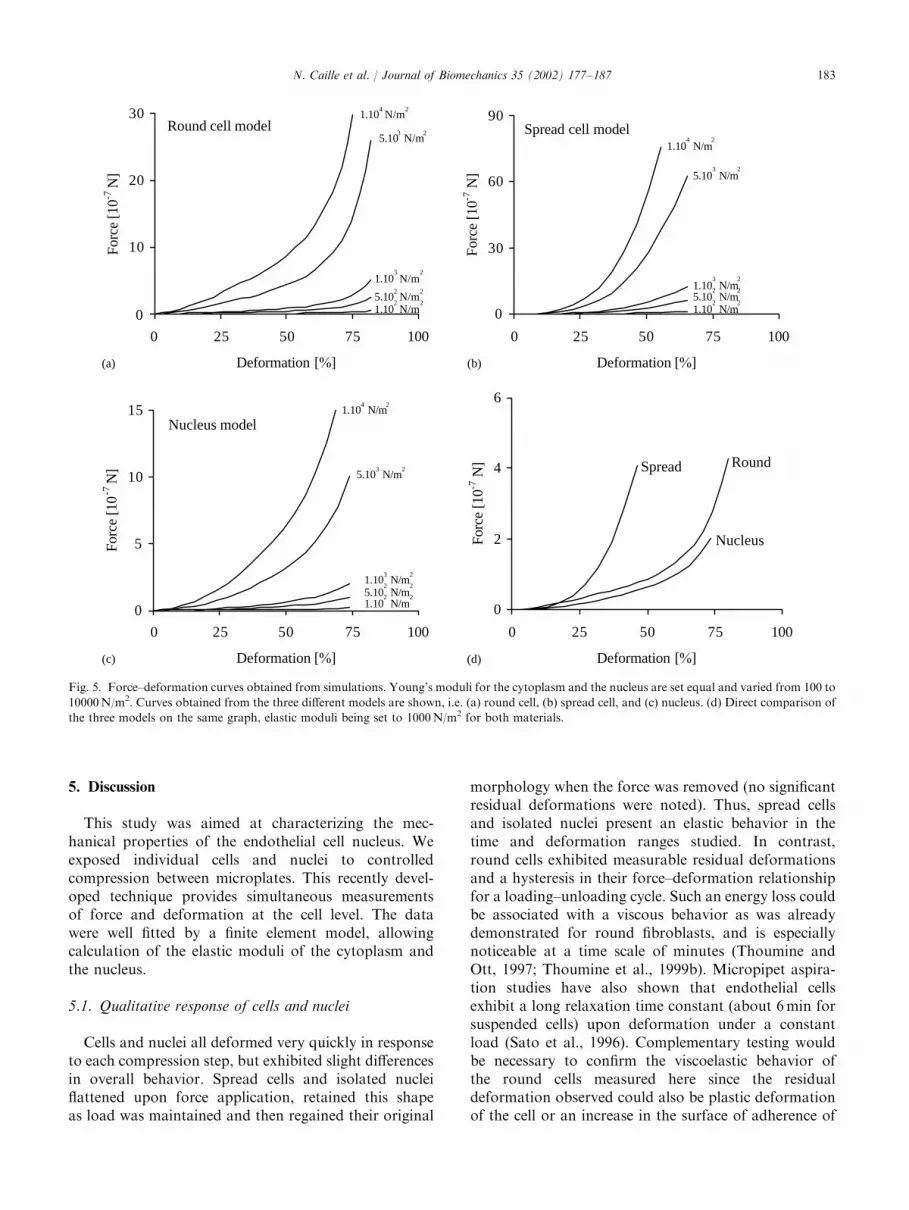
Several test cases were explored in order to observethe behavior of the model and its sensitivity tovariations in the material properties. These simulationsclearly show the non-linear force–deformation behaviorfor each geometry similar to that observed experi-mentally (Fig. 5). Choosing the same value for theYoung’s moduli of the cytoplasm and the nucleusallows the demonstration that the non-linearity of thecurves is due to the material behavior and not theinhomogeneity caused by the presence of the nucleus(Fig. 5a–c). Furthermore, by comparing the curvesobtained for 1000 N/m2 for the three geometries(Fig. 5d), one can see that in order to obtain thesame percentage of deformation, less force is requiredfor the round cells and the isolated nuclei than thespread cells.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187 181
4.3. Estimation of the mechanical properties
The force–deformation curves are calculated for eachparameter set (Ecyto; Enucleus), and for both the roundand spread cell simulations. These results are comparedto the experimental ones for spread and round cells andthe deviation parameter, d; is calculated. For each valueof Enucleus; the curve of d versus Ecyto shows a minimum(Fig. 6a), indicating convergence. The lowest curve istaken to estimate Enucleus and its minimum gives thecorresponding value of Ecyto: The simulated force–deformation curve obtained with this set of parameterscompares very well to the experimental data (Fig. 6b).This procedure was repeated for all tested round andspread cells to obtain their mechanical properties. Thesimulation of the isolated nucleus depends on only oneparameter, Enucleus: In this case, d was plotted versus
Enucleus; and the value of Enucleus giving the minimumd was then taken as the appropriate nucleus elasticmodulus.
4.4. Summary
The cell and nucleus material properties are summar-ized in Table 1. The following observations can bemade: (i) The mean value of the elastic modulus ofnuclei in the cell is 5000 N/m2. This value is on average10 times stiffer than the cytoplasm of spread cells andround cells. (ii) The elastic modulus of isolated nuclei isgreater than that calculated for whole cell geometriesbut remains on the same order of magnitude. The meanvalue found for isolated nuclei is 8800 N/m2. (iii) Thereis no statistical difference between the elastic moduli ofround and spread cells.
0
1
2
3
0 20 40 60 80
Deformation [%]
Forc
e [1
0-7 N
]Round cells
0
4
8
12
0 20 40 60 80
Deformation [%]
Forc
e [1
0-7 N
]
Spread cells
0
4
8
12
0 20 40 60 80
Deformation [%]
Forc
e [1
0-7 N
]
Nuclei
0
5
10
0 20 40 60 80
Deformation [%]
Forc
e [1
0-7 N
]
RoundSpreadNucleus
(a) (b)
(d)(c)
Fig. 4. Experimental force–deformation data for: (a) round cells (n ¼ 17), (b) spread cells (n ¼ 17), and (c) isolated nuclei (n ¼ 21). The deformation
represents the relative reduction in height, and the force is estimated from microplate deflection. Note the difference in scale between graph (a) and
graphs (b) and (c). The curves in (d) represent the average deformation values for the three categories (mean7SD).
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187182
5. Discussion
This study was aimed at characterizing the mec-hanical properties of the endothelial cell nucleus. Weexposed individual cells and nuclei to controlledcompression between microplates. This recently devel-oped technique provides simultaneous measurementsof force and deformation at the cell level. The datawere well fitted by a finite element model, allowingcalculation of the elastic moduli of the cytoplasm andthe nucleus.
5.1. Qualitative response of cells and nuclei
Cells and nuclei all deformed very quickly in responseto each compression step, but exhibited slight differencesin overall behavior. Spread cells and isolated nucleiflattened upon force application, retained this shapeas load was maintained and then regained their original
morphology when the force was removed (no significantresidual deformations were noted). Thus, spread cellsand isolated nuclei present an elastic behavior in thetime and deformation ranges studied. In contrast,round cells exhibited measurable residual deformationsand a hysteresis in their force–deformation relationshipfor a loading–unloading cycle. Such an energy loss couldbe associated with a viscous behavior as was alreadydemonstrated for round fibroblasts, and is especiallynoticeable at a time scale of minutes (Thoumine andOtt, 1997; Thoumine et al., 1999b). Micropipet aspira-tion studies have also shown that endothelial cellsexhibit a long relaxation time constant (about 6 min forsuspended cells) upon deformation under a constantload (Sato et al., 1996). Complementary testing wouldbe necessary to confirm the viscoelastic behavior ofthe round cells measured here since the residualdeformation observed could also be plastic deformationof the cell or an increase in the surface of adherence of
0
10
20
30
0 25 50 75 100
Deformation [%]
Forc
e [1
0-7 N
]Round cell model
1.104 N/m
2
1.102 N/m
25.10
2 N/m
2
1.103 N/m
2
5.103 N/m
2
0
30
60
90
0 25 50 75 100
Deformation [%]
Forc
e [1
0-7 N
]
Spread cell model1.10
4 N/m
2
5.102 N/m
2
1.102 N/m
2
1.103 N/m
2
5.103 N/m
2
0
5
10
15
0 25 50 75 100
Deformation [%]
Forc
e [1
0-7
N]
Nucleus model1.10
4 N/m
2
1.102 N/m
25.102 N/m
21.10
3 N/m
2
5.103 N/m
2
0
2
4
6
0 25 50 75 100
Deformation [%]
Forc
e [1
0-7 N
] Spread
Nucleus
Round
(a) (b)
(d)(c)
Fig. 5. Force–deformation curves obtained from simulations. Young’s moduli for the cytoplasm and the nucleus are set equal and varied from 100 to
10000 N/m2. Curves obtained from the three different models are shown, i.e. (a) round cell, (b) spread cell, and (c) nucleus. (d) Direct comparison of
the three models on the same graph, elastic moduli being set to 1000 N/m2 for both materials.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187 183

the cell on its basal surface to the rigid microplate.Because the experiments are kept short (the average timefor the overall cycle is 2 min), this time-varying
component, if it exists, was neglected in the calculations.While adhesion of the cells to the flexible microplate isblocked by the presence of serum in the medium(Thoumine et al., 1999a), the isolated nuclei adherestrongly to the glass surface. The adherence is so strongthat nuclei cannot be detached from the microplateafter compression. This introduces an additional error inthe force–deformation measurements for nuclei, espe-cially for the unloading sequence. Since the measure-ments made during the unloading are not sufficient toconclude on the existence of a viscoelastic behavior ofround cells or isolated nuclei, only the loading sequencewas taken into account for modeling. However,viscoelasticity can be considered as two separateperfectly elastic behaviors: one in loading and anotherin unloading. The form of the material constitutive lawis the same for both, but the elastic parameters aredifferent. Therefore, the fact that only the loading wasmodeled here is not in contradiction with a possibleviscoelastic behavior.
5.2. Quantitative mechanical behavior
The forces necessary for the compression of endothe-lial cells or isolated nuclei are in the range of 1–10� 10�7 N. In particular, the forces obtained on roundendothelial cells (up to 10�8 N) agree with thosereported for similar compression of round fibroblastsbetween two microplates (Thoumine and Ott, 1997),giving confidence in the reproducibility of the technique.For comparison, the forces involved in pokingexperiments (Miyazaki and Hayashi, 1999) or micro-pipet suction (Sato et al., 1987; Thoumine et al., 1999b)are on the order of 10�9 N. The difference is due tothe fact that only local regions are probed with thelatter techniques, whereas microplates allow whole celldeformations.
Another important result is that the resistance todeformation increases as the force does for all objectstested. Such non-linear behavior has already beendocumented in various cell types using the cell poker(Petersen et al., 1982), magnetocytometry (Wang et al.,1993; Wang and Ingber, 1994) and atomic forcemicroscopy (Miyazaki and Hayashi, 1999; Sato et al.,
Ecyto [N/m2]
Dev
iati
on p
aram
eter
δ105
103
101
10-1
10-3
101 102 103 104 105
0
0.4
0.8
0 10 20 30 40
Deformation [%]
Forc
e [1
0-7 N
]
simulation
measure
(a)
(b)
Fig. 6. Example of a parameter fit for a round cell. (a) Plot of the
parameter d; expressing the deviation between the experimental and
simulated data, versus the cytoplasm elastic modulus Ecyto; for
different values of the nucleus elastic modulus, Enucleus: In this
particular case, the minimum d is obtained for Ecyto ¼ 1000 N/m2
and Enucleus ¼ 2500 N/m2. (b) Force–deformation plot showing both
the experimental data (triangles) and the simulated results (line) after
using the parameters obtained from (a). This is a representative
example of all parameter adjustments carried out on cells and nuclei.
Table 1
Summary of the estimated mechanical properties for cells and nuclei
Elastic modulus (N/m2) Min Max Mean Standard deviation
Round Ecyto 100 1000 323a 252
n ¼ 13 Enucleus 500 10 000 4462a 3608
Spread Ecyto 100 2500 775b 757
n ¼ 10 Enucleus 1000 10 000 5100b 2951
Nucleus Enucleus 5000 25 000 8800 4769
n ¼ 20
aEcyto and Enucleus of round cell are significantly different (po0:05) using a paired Student’s t-Test.bEcyto and Enucleus of spread cell are significantly different (po0:05) using a paired Student’s t-Test.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187184

2000). Similar behavior is obtained from the modelswhen the same mechanical properties are attributed tothe cytoplasm and the nucleus (Fig. 5d). This impliesthat the hyperelastic properties of the models, not thefact that they contain two different materials,account for this non-linear behavior. These modelconfigurations also demonstrate that for the samepercentage of deformation, less force must be appliedto the round cells and to the isolated nuclei thanto the spread cells (Fig. 5d). This difference is certainlyrelated to the difference in the geometries and especiallythe contact surfaces with the microplates. Duringcompression, the contact surface with the flexiblemicroplate is larger for the spread cell than forround cells and isolated nuclei and therefore greaterforces are needed to obtain the same amount ofdeformation. The difference between the contactsurfaces of the round cells and the isolated nuclei issmaller hence the force–deformation curves for the samemechanical properties are closer. From this observationand the experimental knowledge that similar forces arerequired to obtain similar percentages of deformation inthe spread cells and the isolated nuclei, one expectsthat the isolated nuclei will have a greater elasticmodulus than the global elastic modulus of the spreadand round cells.
5.3. Cytoplasmic properties
The elastic modulus found for the cytoplasm ofboth round and spread cells is on the order of 500 N/m2.Despite dispersion of the results that could be reducedby parameterizing the model dimensions in order toallow the entry of the measured dimensions of eachcell and nucleus instead of working with averagedimensions, this value is in agreement with the esti-mations done by Theret et al. (1988) based on acontinuum analysis of micropipet experiments. Theo-retical (Satcher and Dewey, 1996; MacIntosh et al.,1995) and experimental (Janmey, 1991) estimationsbased solely on the mechanical properties of actinfilaments are 2–3 orders of magnitude greater than thevalue found here. Although the actin cytoskeletoncertainly plays a key role in the cell mechanicalproperties, the cell is a complex assembly and there-fore its global behavior may be different than that ofone of its isolated components, even the most abundant.The fact that similar mechanical properties arefound for both round and spread cells indicates thatthere is no rigidification of the cell during spreading.However, the forces necessary to compress the spreadcells are greater than those for the round ones,illustrating the role that the cell geometry plays inthe determination of the global compression rigidityof the cell. This is shown by the behavioral studyof the model where for the same material properties
the spread cell is found to require a greater compressionforce than the round cell to obtain the same percentageof deformation (Fig. 5d). An increase in the rigidityassociated with spreading has been shown on endo-thelial cells (Sato et al., 1987; Theret et al., 1988; Wangand Ingber, 1994) and fibroblasts (Thoumine et al.,1999b). Reorganization of the cytoskeleton duringthis process has been proposed to explain the results.Here, the incubation time for cell spreading is rela-tively short (45 min) compared to those used in theliterature. This time limit is due to technical limitationscaused by the migration of the cells towards theedge of the microplate when longer incubationperiods are used. One can thus conclude that thereorganization of the cytoskeleton at this point isnot sufficient to yield changes in the mechanicalproperties that are measurable with the system used inthis study.
5.4. Nucleus properties
The nucleus was long assumed to be a stiff structure,i.e. it was suspected of getting plugged in micropipetsin the course of cell aspiration (Schmid-Sch .onbeinet al., 1981). The elastic modulus of nuclei in roundand spread cells was found to be around 5000 N/m2,on average 10 times more rigid than the cytoplasm.The value found for the elastic modulus of isolatednuclei, around 8000 N/m2, is greater than that calculatedfor nuclei in the cell but remains on the same orderof magnitude. The difference between these twovalues may be due to the method used to expulse thenuclei and the fact that they are not in their normalphysiological environment. The results obtained fornuclei in the cell are in good agreement withother studies found in the literature. Guilak et al.(2000) have found that the modulus of elasticity ofnuclei of articular chondrocytes is on the orderof 1000 N/m2, 3–4 times stiffer and nearly as viscous asthe cytoplasm when using the micropipet techniqueon mechanically and chemically isolated nuclei. Inmore qualitative studies, Dong et al. (1991) have shown,using a similar technique, that the elasticity of thenucleus of isolated neutrophils is about 10 times largerthan that of the cytoplasm. Maniotis et al. (1997a)have also shown that the nucleus presents a rigiditygreater than that of the rest of the cell and haveestimated it to be 9 times stiffer than the cytoplasm;their technique consists of the application of localstrain to the cell with a micropipet and the measurementof movement of intracellular organelles. Miyazakiand Hayashi (1999) measured the local mechanicalproperties of intact endothelial cells in fresh arteriesby poking the surface of the cells and they found agreater stiffness at locations above the nucleus. Incontrast, experiments with a cell poker (Petersen et al.,
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187 185

1982) or an atomic force microscope (Sato et al., 2000)on cells adherent to a rigid substrate demonstrate agreater stiffness at the cell periphery. This is probablybecause these distal regions are very thin, i.e. o0.5 mm,and as soon as the probe touches them, it feels theresistance of the underlying substrate. The fact that thenucleus is more rigid than the cytoplasm can explainwhy the nucleus deforms less than the cells when theirsubstrate is stretched (Caille et al., 1998). Our measure-ments of the nucleus rigidity may help to develop abetter understanding of how endothelial cells transduceexternal forces like shear stress into specific geneticallyregulated responses.
Acknowledgements
The authors thank Ms. A. Kottelat and Ms. D.-E.Bakman for their contributions to the experimentalmeasurements. This work was supported by the FondsNational Suisse de la Recherche Scientifique (Grant No.2100-042340.94/1).
References
Booyse, F.M., Sedlak, B.J., Rafelson Jr., M.E., 1975. Culture of
arterial endothelial cells. characterization and growth of bovine
aortic cells. Thrombosis et Diathesis Haemorrhagica. 34 (3),
825–839.
Caille, N., Tardy, Y., Meister, J.-J., 1998. Assessment of strain field in
endothelial cells subjected to uniaxial deformation of their
substrate. Annals of Biomedical Engineering 26, 409–416.
Davies, P.F., 1995. Flow-mediated endothelial mechanotransduction.
Physiological Reviews 75, 519–560.
Dong, C., Skalak, R., Sung, K.-L.P., 1991. Cytoplasmic rheology of
passive neutrophils. Biorheology 28, 557–567.
Galbraith, C.G., Skalak, R., Chien, S., 1998. Shear stress induces
spatial reorganization of the endothelial cell cytoskeleton. Cell
Motility and the Cytoskeleton 40, 317–330.
Gimbrone, M.A., Cybulsky, M.I., Kume, N., Collins, T., Resnick, N.,
1995. Vascular endothelium, an integrator of pathophysiological
stimuli in atherogenesis. Annals of the New York Academy of
Sciences 748, 122–131.
Gimbrone, M.A., Resnick, N., Nagel, T., Khachigian, L.M., Collins,
T., Topper, J., 1997. Hemodynamics, endothelial gene expression
and atherogenesis. Annals of the New York Academy of Sciences
811, 1–10.
Guilak, F., 1995. Compression-induced changes in the shape and
volume of the chondrocyte nucleus. Journal of Biomechanics 28,
1529–1541.
Guilak, F., Tedrow, J.R., Burgkart, R., 2000. Viscoelastic properties of
the cell nucleus. Biochemical and Biophysical Research Commu-
nications 269, 781–786.
Hazel, A.L., Pedley, T.J., 2000. Vascular endothelial cells minimize the
total force on their nuclei. Biophysical Journal 78, 47–54.
Ingber, D.E., 1990. Fibronectin controls capillary endothelial cell
growth by modulating cell shape. Proceeding of the National
Academy of Sciences of the United States of America 87,
3579–3583.
Janmey, P.A., 1991. Mechanical properties of cytoskeletal polymers.
Current Opinion in Cell Biology 2, 4–11.
Ku, D.N., Giddens, D.P., Zarins, C.K., Birchmeier, W., 1985.
Pulsative flow and atherosclerosis in the human carotid bifurca-
tion: positive correlation between plaque localization and low and
oscillating shear stress. Arteriosclerosis 5, 293–302.
MacIntosh, F.C., K.as, J., Janmey, P.A., 1995. Elasticity of semi-
flexible biopolymer networks. Physical Review Letters 75,
4425–4428.
Maniotis, A.J., Chen, C.S., Ingber, D.E., 1997a. Demonstration of
mechanical connections between integrins, cytoskeletal filaments,
and nucleoplasm that stabilize nuclear structure. Proceeding of the
National Academy of Sciences of the United States of America 94,
849–854.
Maniotis, A.J., Bojanowski, K., Ingber, D.E., 1997b. Mechanical
continuity and reversible chromosome disassembly within intact
genomes removed from living cells. Journal of Cellular Biochem-
istry 65, 114–130.
Miyazaki, H., Hayashi, K., 1999. Atomic force microscopic measure-
ment of the mechanical properties of intact endothelial cells in fresh
arteries. Medical and Biological Engineering and Computing 37,
530–536.
Nerem, R.M., 1992. Vacular fluid mechanics, the arterial wall, and
atherosclerosis. Journal of Biomechanical Engineering 114,
274–282.
Petersen, N.O., McConnaughey, W.B., Elson, E.L., 1982. Dependence
of locally measured cellular deformability on position on the cell,
temperature, and cytochalasin B. Proceeding of the National
Academy of Sciences of the United States of America 79,
5327–5331.
Pienta, K.J., Coffey, D.S., 1992. Nuclear–cytoskeletal interactions:
evidence for physical connections between the nucleus and cell
periphery and their alteration by transformation. Journal of
Cellular Biochemistry 49, 357–365.
Satcher, R.L., Dewey, C.F., 1996. Theoretical estimates of mechanical
properties of the endothelial cell cytoskeleton. Biophysical Journal
71, 109–118.
Sato, M., Murina, J., Nerem, R.M., 1987. Micropipet aspiration of
cultured bovine aortic endothelial cells exposed to shear stress.
Arteriosclerosis 7, 276–286.
Sato, M., Ohshima, N., Nerem, R.M., 1996. Viscoelastic properties of
cultured porcine aortic endothelial cells exposed to shear stress.
Journal of Biomechanics 29, 461–467.
Sato, M., Nagayama, K., Kataoka, N., Sasaki, M., Hane, K., 2000.
Local mechanical properties measured by atomic force microscopy
for cultured bovine endothelial cells exposed to shear stress.
Journal of Biomechanics 33, 127–135.
Schmid-Sch .onbein, G.W., Sung, K.-L.P., T .ozeren, H., Skalk, R.,
Chien, S., 1981. Passive mechanical properties of human leuko-
cytes. Biophysical Journal 36, 243–256.
Schmidt, C.E., Chen, T., Lauffenburger, D.A., 1994. Simulation of
integrin–cytoskeletal interactions in migrating fibroblasts. Biophy-
sical Journal 67, 461–474.
Sims, J.R., Karp, S., Ingber, D.E., 1992. Altering the cellular
mechanical force balance results in integrated changes in cell,
cytoskeletal and nuclear shape. Journal of Cell Science 103,
1215–1222.
Theret, D.P., Levesque, M.J., Sato, M., Nerem, R.M., Wheeler, L.T.,
1988. The application of a homogeneous half-space model in the
analysis of endothelial cell micropipet measurements. Journal of
Biomechanical Engineering 110, 190–199.
Thoumine, O., Ott, A., 1997. Time scale dependent viscoelastic and
contractile regimes in fibroblasts probed by microplate manipula-
tion. Journal of Cell Science 110, 2109–2116.
Thoumine, O., Ott, A., Cardoso, O., Meister, J.-J., 1999a. Microplates:
a new tool for manipulation and mechanical perturbation of
individual cells. Journal of Biochemical and Biophysical Methods
39, 47–62.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187186

Thoumine, O., Cardoso, O., Meister, J.-J., 1999b. Changes in the
mechanical properties of fibroblasts during spreading: a micro-
manipulation study. European Biophysics Journal 28, 222–234.
Truesdell, C., Noll, W., 1992. The non-linear field theories of
mechanics. Springer, Berlin, Heidelberg.
Wang, N., Ingber, D.E., 1994. Control of cytoskeletal mechanics by
extracellular matrix, cell shape, and mechanical tension. Biophy-
sical Journal 66, 2181–2189.
Wang, N., Butler, J.P., Ingber, D.E., 1993. Mechanotransduction
across the cell surface and through the cytoskeleton. Science 260,
1124–1127.
Zahalak, G.I., McConnaughey, W.B., Elson, E.L., 1990. Determina-
tion of cellular mechanical properties by cell poking, with an
application to leukocytes. Journal of Biomechanical Engineering
112, 28–294.
N. Caille et al. / Journal of Biomechanics 35 (2002) 177–187 187