conserved intramolecular interactions maintain myosin interacting-heads motifs explaining tarantula...
TRANSCRIPT

Article
Lorenzo Alamo1,
0022-2836/© 2016 Elsevi
Conserved Intramolecular InteractionsMaintain Myosin Interacting-Heads MotifsExplaining Tarantula Muscle Super-RelaxedState Structural Basis
1, Dan Qi2, Willy Wrigger
s3, Antonio Pinto1, Jingui Zhu2,Aivett Bilbao †, Richard E. Gillilan4, Songnian Hu2 and Raúl Padrón11 - Centro de Biología Estructural, Instituto Venezolano de Investigaciones Científicas, Apartado 20632, Caracas 1020A, Venezuela2 - Key Laboratory of Genome Sciences and Information, Beijing Institute of Genomics, 1 Beichen West Road, Chaoyang District,Beijing 100101, China3 - Department of Mechanical and Aerospace Engineering, Old Dominion University, 5115 Hampton Boulevard, Norfolk, VA 23529,USA4 - Macromolecular Diffraction Facility, Cornell High Energy Synchrotron Source, 161 Wilson Laboratory, Synchrotron Drive, Ithaca,NY 14853, USA
Correspondence to Raúl Padrón: [email protected]://dx.doi.org/10.1016/j.jmb.2016.01.027Present address: A. Bilbao, SIB Swiss Institute of Bioinformatics, Proteome Informatics Group and Life Sciences MassSpectrometry, School of Pharmaceutical Sciences, University of Geneva, Geneva, Switzerland.Edited by J. Sellers
Abstract
Tarantula striated muscle is an outstanding system for understanding the molecular organization of myosinfilaments. Three-dimensional reconstruction based on cryo-electron microscopy images and single-particleimage processing revealed that, in a relaxed state, myosin molecules undergo intramolecular head–headinteractions, explaining why head activity switches off. The filament model obtained by rigidly docking achicken smooth muscle myosin structure to the reconstruction was improved by flexibly fitting an atomicmodel built by mixing structures from different species to a tilt-corrected 2-nm three-dimensional map offrozen-hydrated tarantula thick filament. We used heavy and light chain sequences from tarantula myosin tobuild a single-species homology model of two heavy meromyosin interacting-heads motifs (IHMs). The flexiblyfitted model includes previously missing loops and shows five intramolecular and five intermolecularinteractions that keep the IHM in a compact off structure, forming four helical tracks of IHMs aroundthe backbone. The residues involved in these interactions are oppositely charged, and their sequenceconservation suggests that IHM is present across animal species. The new model, PDB 3JBH, explains thestructural origin of the ATP turnover rates detected in relaxed tarantula muscle by ascribing the very slow rateto docked unphosphorylated heads, the slow rate to phosphorylated docked heads, and the fast rate tophosphorylated undocked heads. The conservation of intramolecular interactions across animal species andthe presence of IHM in bilaterians suggest that a super-relaxed state should be maintained, as it plays a role insaving ATP in skeletal, cardiac, and smooth muscles.
© 2016 Elsevier Ltd. All rights reserved.
Introduction
Muscle contraction involves the interaction of twosets of filaments: actin-containing thin filaments andmyosin-containing thick filaments. Muscle relaxationis an important step of contraction, and thickfilaments are essential for relaxation [1]. The twomyosin II coiled-coil heavy chains [myosin II heavychains (MHCs)] form a tail with two heads, with
er Ltd. All rights reserved.
attached regulatory light chains (RLCs) and es-sential light chains (ELCs) on each. The tails packtogether, comprising the thick filament backbonewith protruding heads that form helical tracks.Three-dimensional (3D) reconstructions haveshown that the number of helical tracks in striatedmuscle varies between three in cardiac vertebrates(mouse [2], human [3], and zebrafish [4]), four inarthropods (tarantula [5], Limulus [6], and scorpion
J Mol Biol (2016) 428, 1142–1164

1143Myosin Interacting-Heads Motifs
[7]), and seven in mollusks (scallop) [8]. The numberof smooth muscle helical tracks is four in Platyhel-minthes (Schistosome) [9]; in vertebrates, smoothmuscle filaments are non-helical and side polar [10].Electron microscopy studies of smooth muscle
myosin molecules suggest that an off state is achievedby asymmetric, intramolecular interaction between theactin-binding region of one myosin head and theconverter region of the other, which switches off bothheads [11]. This mechanism of relaxation, theso-called myosin interacting-heads motif (IHM) [5],was shown to underlie the relaxed state of thickfilaments from striated muscle [5]. The IHM is presentin species separated by at least 600 million years ofindependent evolution. It has been observed byelectron microscopy in thick filaments from the striatedmuscle of arthropods [5–7] and mollusks [8], thecardiac muscle of vertebrates [2–4], and the smoothmuscle of Platyhelminthes [12]. Electron microscopyhas also shown that the motif is present in isolatedmyosin molecules of intrinsically regulated molecules(like tarantula and Limulus striated muscle andnon-muscle myosin IIA) and in unregulated myosins(like skeletal and cardiac muscles) [13]. Recently, themotif has been detected on isolated myosin moleculesfrom the smooth muscle of the Cnidarian giant seaanemone (Condylactis gigantea) [14], but not inisolated myosins from the amoeba Acanthamoebacastellani [14]. Finally, the presence of the IHM insquid (Loligo peali) has been inferred by comparingthe small-angle X-ray solution scattering (SAXS)profile of its striated muscle with the predictedscattering profile of tarantula Protein Data Bank(PDB) 3DTP structure [15].The uniqueness, wide presence, and high conser-
vation of the IHM establish it as a quintessentialmolecular machine whose simple mechanism explainsthe relaxed (off) state in muscle. The IHM isestablished only under relaxed conditions. In thepresence of Mg·ATP, heads are bent at the “pliantregion” [16], as the switch 2 element is closed,inhibiting the phosphate release. The bending of freeheads and blocked heads produce the requiredconformations for establishing several head–headand head–tail intramolecular interactions, as well asseveral head–head and head–backbone intermolecu-lar interactions, which allow the formation of thickfilaments in regulated muscles. It has been suggestedthat intramolecular interactions are a general mecha-nism for inducing muscle relaxation and switching offmyosin-II-based motile activity in both muscle andnon-muscle cells [17]. The so-called super-relaxedstate (SRX) of myosin II has been reported in striatedmuscle and is characterized by a subpopulation ofmyosin heads with a highly inhibited rate of ATPturnover, lower than the turnover observed for singlemolecules [18]. Myosin IHM has been found to beinvolved in the slow ATP turnover rate observed invertebrate skeletal and cardiac muscle fibers [19–21].
In particular, relaxed tarantula striated muscle fibersexhibit a very slow rate [22].The tarantula striated muscle IHM model (PDB
3DTP) has improved our understanding of theactivation mechanism in arthropods that involvestwo phosphorylatable serines (Ser35 and Ser45)[23]. Based on this structure, a cooperative phos-phorylation activation (CPA) mechanism (Fig. 8c–f)and a model for activation, potentiation, and post-tetanic potentiation involving swaying heads in arelaxed state were proposed for tarantula striatedmuscle [24,25]. A disorder-to-order molecularmechanism occurring in the myosin RLC N-terminalextension (NTE) is proposed to control this arthropodphosphorylation-based activation [26,27]. The IHMmodel has improved the understanding of theactivation mechanism in vertebrate skeletal muscle[28,29] and in cardiac muscle [30,31]. On the otherhand, the smooth muscle IHM (PDB 1I84) structurehas improved the understanding of the smoothmuscle activation mechanism [32], including apossible role of myosin ELC [33].An enhanced model of the IHM is very important for
increasing our understanding of the thick filamentrelaxation and activation mechanisms. The onlystructural information for the IHM comes from cryo-electron microscopy of chicken smooth musclemyosin II two-dimensional crystals [11,34,35] andfrom tarantula striated muscle frozen-hydrated relaxedthick filaments [5,23]. The quasi-atomic model smoothmuscle IHM (PDB 1I84) includes the chicken smoothmuscle subfragment 1 atomic structure (PDB 1BR1),but, as it did not come from a filament, it does notproperly include the subfragment S2 crystal structure[11]. Two reconstructions were calculated from low-dose electron micrographs of frozen-hydrated taran-tula thick filaments. The first reconstruction reached aresolution of 2.5 nm [5] and was used to build an initialquasi-atomic model of the filament by manually fittingthe chicken smooth muscle myosin quasi-atomicstructure PDB 1I84 [11] without the S2. The secondreconstruction, which was deposited in the ElectronMicroscopy Data Bank [36] as EMD-1950 [23],reached a slightly higher resolution (2.0 nm) byusing a higher number of image segments and byincluding the tilt angle made by the filaments with thegrid in the reconstruction. This second reconstructionwas more detailed, clearly showing two new interac-tions in addition to the five shown in the first re-construction [5]. This enhanced reconstruction led to asecond improved tarantula IHM model (PDB 3DTP)[23], which was achieved by flexible fitting of an atomicmodel built from different species: a human cardiacmyosin S2 crystal structure (PDB 2FXM) [37], thepredicted secondary structure homologous quasi-a-tomic model for the tarantula (Avicularia) myosin RLCsequence [23], and the ELC and myosin heads' motordomain from the chicken smooth muscle quasi-atomicmodel (PDB 1I84) [35]. Since three loops (loop 1, loop

Fig. 1. Wide-eye stereo pair of the longitudinal view of the 3D reconstruction of the frozen-hydrated tarantula thickfilament, filtered to 2-nm resolution (EMD-1950) [23] and showing four helical tracks of IHMs (blue), 12 myosinsubfilaments (gray), and the paramyosin core (orange). The 3D map segment shows four 14.5-nm crowns, each of whichhas four IHMs. The quasi-atomic model PDB 3JBH (formed by two IHMs), which is shown as spheres in the right helix, wasflexibly fitted to the 3D map (see Materials and Methods). The myosin heavy chain (MHC) of the blocked head (BH) andfree head (FH) are shown in green and blue. The two myosin ELCs are in magenta (FH) or orange (BH). The two myosinRLCs are in red (FH) or yellow (BH). Bar: 14.5 nm.
1144 Myosin Interacting-Heads Motifs
2, and loop NATP-V, which is defined here as theloop near the nucleotide ATP binding D458 shown inFig. 6b) are missing in chicken MHC (PDB 1BR1),their sequences are not present in two earlier IHMPDB models (PDB 1I84 and PDB 3DTP).Analysis of transcriptional isoforms of tarantula
skeletal muscle protein sequences has determinedits myosin ELC and RLC sequences [38]. The aimsof the present work are (1) to build a homologousIHM quasi-atomic model (PDB 3JBH) based on PDB3DTP using the tarantula myosin II ELC [38], RLC[38], and MHC sequences reported here to flexibly fitthis model to the tarantula thick filament 3Dreconstruction (EMD-1950); (2) to analyze whetherthe residues involved in maintaining the criticalintramolecular interactions of the IHM could es-tablish ionic interactions and whether they areconserved across different species; and (3) toinvestigate whether this new homologous tarantulaIHM model and its interactions provide clues aboutthe structural basis of the SRX state in tarantulastriated muscle.
Results
Tarantula myosin MHC, ELC, and RLCresidue sequences
The tarantula Aphonopelma sp. MHC sequence(GenBank [39] KT619079) was determined as
described in Materials and Methods, which is in-cluded in the supplementary data (see Accessionnumbers) along with the myosin ELC [38] (GenBankKT390185) and RLC [38] (GenBank KT390186).The tarantula Avicularia avicularia myosin RLCsequence (UniProt [40] B4XT43) [23] that wasincluded in PDB 3DTP is very similar to that ofAphonopelma sp., as both have 196 amino acidsand differ only on 9.
Tarantula IHM quasi-atomic model
We built a homologous single-species model usingthe MHC, RLC, and ELC of tarantula Aphonopelmasp. based on PDB 3DTP [23] (see Materials andMethods). The model was flexibly fit into the 2-nm-re-solution 3D map (Fig. 1, EMD-1950) calculated fromelectron micrographs of rapidly frozen tarantulaAphonopelma sp. relaxed thick filaments preservedin vivo in a relaxed state [23]. The model includesthree modeled loops (loop 1, loop 2, and loopNATP-V; Supplementary Table 1) that are missing inthe chicken MHC crystal structure (PDB 1BR1) andthe PDB 1I84 and PDB 3DTP models. Due to theIHM structural asymmetry of free and blocked heads,the loops are located in very different environments(Fig. 2a). Loop 1 faces the solvent in both free andblocked head (Fig. 2a) and is not involved in anyintramolecular or intermolecular interactions. Loop 2was included after being remodeled de novo (Fig. 2a,see Materials and Methods) and is fitted to density

Fig. 2. Wide-eye stereo pairs of the 2-nm resolution 3D reconstruction of frozen-hydrated relaxed thick filament oftarantula (EMD-1950) [23], shown in gray, with the flexibly fitted quasi-atomic model PDB 3JBH, as viewed from the front(a) or back (b) of the filament surface. The model includes the densities where several loops are located in the blockedhead region of interactions “b” and “c” (see Materials and Methods). The MHC in PDB 3JBH shows six surface loops (2, H,CM, 3, C, and I) that are involved in the interactions. Also, the ELC in the PDB 3JBH model shows the extra two aminoacids that are missing in the chicken ELC sequence. In (b), the intramolecular interactions are “a” (FH MD loop 2–S2), “d”(FH MD–BH MD), “e” (FH ELC–BH MD), “f” (S2–BH MD), and “g” (S2–BH ELC). The intermolecular interactions are “b”(BH RLC–FH MD) and “c” (BH ELC–FH MD), which are established with the adjacent IHM in the filament, and “h”, whichoccurs between the blocked head SH3 domain and a neighbor myosin S2 (shown as a 2-nm pink cylinder). Each of theseinteractions is shown in Figs. 6 and 7. MD: motor domain of the myosin head. See legend of Fig. 1. Bar: 50 Å.
1145Myosin Interacting-Heads Motifs
“a” (Fig. 2b). The loop 2 sequence in myosin IIsignificantly affects actin-activated ATPase activity,mechanochemical coupling, and actin binding.Regulation by RLC phosphorylation depends onthe presence of a loop 2 with greater length and aless positive charge density than the skeletal isoform[41–43]. The tarantula loop 2 sequence appears to bedifferent from other species but preserves thepositively charged region, a so-called lysine pocket(Supplementary Figs. 1 and 2) [42]. The CM loop,previously fitted to density “a” [23,37], was found after
remodeling the missing loops to better match thedensities in the interface with the next IHM (Fig. 2aand Supplementary Movies 2–4) and loop 2 to bettermatch interaction “a”. The flexible fitting of the freehead cluster of loops (Fig. 2a, top right) to the 3D maprequired some rearrangement of the myosin subfrag-ment 1 50K domain, including cleft closure betweenthe upper and lower parts of the 50K domain. It waspreviously proposed that the apparent cleft closureand specific CM loop orientation were required toguide the free head, which was detached after the

1146 Myosin Interacting-Heads Motifs
power stroke, to establish the precise electrostaticdocking interaction “a” onto ring 2 of S2, a requisitefor reforming the helical tracks of IHMs [23]. Thisremodeling suggests that it is loop 2, not the CM loop,that is responsible for this electrostatic dockinginteraction.This quasi-atomic model was deposited as entry
3JBH in the PDB [44]. PDB 3JBH and the earlierPDB 3DTP are very similar (Supplementary Movie1). In fact, several PDB 3JBH and PDB 3DTPmolecule regions are very similar to the crystalstructure PDB 1BR1 (Supplementary Movie 1). Asthese missing loops are located in the region ofinteraction of one IHM with a neighboring IHM, theflexible fitting process included the densities of thisIHM–IHM connecting region (Fig. 2a) located alongthe IHM helical track (Fig. 1). This fitting approachallows for the inclusion of densities along helicaltracks and backbone-anchoring intermolecular inter-actions, which was not possible with crystals ofisolated heavy meromyosin (HMM).
Tarantula model improves upon earlierIHM models
The new model, PDB 3JBH, improves upon thePDB 3DTP model since it is built from completesequences of a single species (tarantula Aphono-pelma sp.) rather than a mixture of sequences fromthree species (human, chicken, and tarantula). Inaddition to the three new loops, the new model hassix surface loops (2, 3, H, 4 or C, CM, and I;Supplementary Table 1), as well as the S2, SH3,catalytic, relay, and converter domains (Fig. 2aand Supplementary Table 1), which are involvedin intramolecular and intermolecular interactions(Fig. 2b). The model includes five intramolecularinteractions (involving the S2 and all six surfaceloops) and five intermolecular interactions [two ofwhich are in a cluster of surface loops (C, CM, I,H, and 3), the free and neighboring blocked headregulatory domain, and neighboring myosin tails andthree of which are involved in anchoring with twoadjacent myosin tails]. The main difference betweenPDB 3JBH and PDB 3DTP lies mostly in the freehead motor domain, where the new, remodeledloops are located near the adjacent blocked headRLC of the next IHM along the helical track (Fig. 1).In addition, PDB 3JBH can be used as a startingmodel to be flexibly fitted to a higher-resolution 3Dreconstruction [45].
Free and blocked heads of tarantula model havedifferent conformations
For myosin heads to form helical tracks in tarantulathick filament, their nucleotide pocket switch 2elements should be in the “closed” conformation,preventing phosphate release [46,47]. Also, in the
presence of Mg·ATP, the heads should be bent atthe “pliant region” [16], as the switch 2 element isclosed, inhibiting the phosphate release (Fig. 4). Thetarantula IHM model PDB 3JBH offers furtherinsights into the specific conformation of blockedand free head in relaxed filament with a blockedhead in a closed pre-power stroke conformation(similar to the Mg·ADP-AlF4, PDB 1BR1) [23,35]while the free head, which is also in a pre-powerstroke closed conformation (with a shape similar toblocked head at the limited resolution of the 3Dmap), shows a less angled lever arm than theblocked head (Fig. 4). The angle of the free headlever arm is midway between the lever arms of theblocked head and pre-power stroke transition 1DFL[16] (Fig. 4).
Predicted SAXS profile of tarantula modelagrees with measured profile of squid HMM
X-ray solution scattering of squid HMM under Ca2+--free [ethylene glycol bis(β-aminoethyl ether) N,N´-te-traacetic acid] conditions shows agreement with PDB3DTP, strengthening the evidence of an ancient IHMcompact off state [15]. In this comparison, the 50residues of the RLC NTE (which are unique to thetarantula myosin RLC) were deleted. However, theagreement was maintained when they were included(Drs. Jerry H. Brown and Richard Gillilan, personalcommunication). The computed scattering profile forthe tarantula model (3JBH) closely agrees with thescattering profile computed from PDB 3DTP at anglesbelow q = 0.11 Å−1 (Fig. 5a). While the two computedprofiles separate slightly at wider angles, the degree ofseparation is below the noise level of the currentexperimentally measured squid HMM profile [15]. Thegoodness-of-fit statistic (chi) to the data reported by thesoftware is slightly better for the squid model (chi =1.74) than for the tarantulamodel (chi = 2.01), but giventhe noise levels in the data and the possible systematicdeviations of both models from the data at the widestangle, it is not possible to distinguish between the two.Computations extending to a wide angle (q = 1.0 Å−1
in Fig. 5b) show that the twomodels continue to closelyagree above q = 0.2 Å−1 and are not likely to bedistinguished by future SAXS and wide-angle X-raysolution scattering experiments in that regime.
Tarantula model includes new intramolecularand intermolecular interactions
In the tarantula thick filament, there were intramo-lecular interactions (between the domains insideeach IHM) [5] and intermolecular interactions (head–head; between the domains of one IHM and theadjacent IHM along the helical tracks of myosinheads on the surface of the thick filament backbone;Figs. 1 and 2) [5,23]. When these interactions arebroken, IHMs become less compact as myosin

Fig. 3. (a) Wide-eye stereo pair of three adjacent IHMs forming part of a helix. The intermolecular interactions “b” and “c”are shown with the RLC and ELC of the neighboring blocked head regulatory domain, and interaction “h” is shown with theneighboring S2. The surface of the 3D map corresponding to the IHM in the center is highlighted in yellow. For clarity, theS2 of the model of the two left IHMs has been extended as coiled-coil α-helices (pink). Since the subfilament structure isnot known, the two neighboring subfilaments are depicted as cylinders with diameters of about 2.2 nm. (b) Wide-eyestereo pair of a rotated 90° view of (a), showing that the model of the IHMs is present only in the slice between the two bluedotted lines in (a), causing the neighboring “h” interaction to be far and the “i” and “j” to be closer to the reader. In a relaxedstate, the S2 of the IHM emerges from the top with a slight angle of 6°, causing the helix of IHMs to “float”, separated fromthe backbone surface by about 2 nm. The blocked head is the only part of the IHM that is in contact with the backbone andis covalently connected to it via the S2 and electrostatically connected by three “anchoring” intermolecular interactions: “h”(blocked head SH3 domain) with the extended S2 of an adjacent tail and “i” (blocked head relay/converter) and “j” (blockedhead ELC) with the neighboring S2 (see Fig. 7). For a structure color code, see the legend of Fig. 1.
1147Myosin Interacting-Heads Motifs
heads are released [24,25], firstly with the separationand eventual release of free head from its partnerblocked head and secondly after the release ofblocked head from the thick filament backbone [24].We use the same notation scheme for the fiveintramolecular interactions (“a”, “d”, “e”, “f”, and “g”;Fig. 2b) as suggested previously [23]. In addition tothe three previously described intermolecular inter-actions [5,23] (“b”, “c”, and “h”; Fig. 2), we show twonew intermolecular interactions (head–tail) betweenthe ELC of the blocked head (“i”; Fig. 3b) and the
relay/converter (“j”; Fig. 3b) with the neighboring S2.Interactions “h”, “i”, and “j” cannot be discussed indetail because our myosin tail model does notinclude this interacting part of the S2 (Fig. 3) andwe lack a quasi-atomic model of the subfilament.
Analysis of interactions
A rigorous analysis of the residues involved in theintramolecular interactions that keep the tarantulaIHM structure compact necessarily requires solving

1148 Myosin Interacting-Heads Motifs
its crystal structure, whereas a proper analysis ofintermolecular interactions requires a resolution ofhigher than 2 nm for the 3D map. In the first case,since the tarantula IHM crystal structure had notbeen determined, the interaction analysis had to beperformed using other known myosin head crystalstructures [48]. The crystallization of tarantula IHMhas been hampered by difficulties with purifying asingle homogeneous unphosphorylated tarantulaIHM, as Ser35 monophosphorylation is constitutivelypresent in relaxed tarantula muscle [24,25]. Therecent purification, crystallization, and preliminaryX-ray crystallographic analysis of squid HMM [49]suggest that mollusks' IHM atomic structure will besolved in the near future. In the second case, theclosest available information for the analysis ofintermolecular interactions only comes from2.5-nm-resolution and 2.0-nm-resolution 3D mapswith the densities of intact rapidly frozen relaxedtarantula thick filaments [5,23]. A higher-resolution3D reconstruction should be available in the future[45,50]. In the meantime, we built the new tarantulamodel (Fig. 1), which should allow us to perform afirst analysis of intramolecular and intermolecularinteractions and their conservation. This attempt islimited by the low resolution of the 3D map, whichdoes not allow a proper atomic level analysis, evenafter flexible fitting, but only allows a rough analysisof the possible residues in the general region of eachinteraction, as it is not possible to trace the densitiesof the main or side chains. Therefore, the analysis inthe model below should be considered an informedguess of the interactions that could be formed ratherthan a direct observation of the actual atomiccontacts in a solved structure.
Intramolecular interactions
We analyzed the intramolecular (Fig. 6 andSupplementary Figs. 1–7) and intermolecular (Fig. 7and Supplementary Figs. 8–12) interactions usingPDB 3JBH (Figs. 1–3) and by observing the peptidesand charged residues involved in the contacts:
(1) -Interactions “a” and “f”: interaction “a”involves residues located on S2 ring 2,which interact with free head loop 2 (Fig. 6a;Supplementary Fig. 1 and SupplementaryTables 1 and 2). Interaction “f” involves twosubinteractions: (i) interaction “f.1” (Fig. 6a;Supplementary Fig. 2 and SupplementaryTables 1 and 2) involves S2 ring 1 andblocked head loop 2. In interaction “a”, freehead loop 2 interacts with ring 2, which isnegatively charged. Blocked head loop 2was remodeled de novo and adopted adifferent conformation that increases the
possibility that more residues will interact.The positioning of blocked head loop 2 justin front of S2 ring 1 suggests an electro-static interaction [37]. (ii) Interaction “f.2”(Fig. 6a, Supplementary Fig. 3 and Supple-mentary Tables 1 and 2) involves S2 ring 1,with blocked head H loop and α-helix nearblocked head loop 2 (Y653-H665).
(2) -Interactions “e” and “g”: interaction“e” involves blocked head near loop 2(K608-H622) and two blocked head regionsnear (before and after) the C loop with twofree head ELC loops (Fig. 6b; SupplementaryFig. 4 and Supplementary Tables 1 and 2).We restricted our analysis to ELC tarantula--like sequences with short NTEs (inverte-brates) [51,52]. In our model, we cannotassesswhich residuesare involved inmakingthese contacts. Nevertheless, the interactingloops that are involved agree with thepreviously proposed loops for vertebratesmooth muscle [35] (Supplementary Table2). Interaction “g” involves theS2 at the end ofthe N-terminal “asymmetry zone” marked bythe so-called “kink” [37] at residue N878(M877 in cardiac chicken muscle) with theblocked head catalytic domain and theblocked head ELC (Fig. 6b; SupplementaryFig. 5 and Supplementary Tables 1 and 2)through three subinteractions: (i) freeheadS2with blocked head loopNATP-V, (ii) free headS2 with blocked head ELC, and (iii) blockedhead S2 with blocked head ELC C-terminalextension.
(3) -Interaction “d”: interaction “d” involvestwo dual subinteractions: (i) interaction “d.1”involves the blocked head CM loop withloop NATP-II (defined here as the loop nearthe nucleotide ATP-binding residue G173shown in Fig. 6c; Supplementary Fig. 6 andSupplementary Tables 1 and 2). It must benoted that residues K400 (CM loop) andE165 (catalytic domain) were also pro-posed to interact in a previous loop remodel[17] (Supplementary Table 2). The blockedhead CM loop also interacts with the freehead loop NATP-V and the free head nearloop 2 (Y653-H665). (ii) Interaction “d.2”involves blocked head near interacting loop(I loop) [23] with the free head converterdomain and the blocked head ELC Cloop with the converter and relay domains(Fig. 6c; Supplementary Fig. 7 and Supple-mentary Tables 1 and 2).

Table 1. Conservation of MHC residues involved in critical intramolecular interactions is associated with IHM and SRX presence
Species Muscletype
Filamenttype
MHCtype
Myosinentry
d.1i
d.1ii
d.1iii
d.2i
d.2ii
d.2iii
f.2i
f.2ii
f.2iii
Cs-d
%Cs-f
%Cs
%IHM
detected?SRX
detected?
MHC II Striated-likesequences
Invertebrates CON Sm Bipolar St (a) 100 100 100 n/a 100 n/a 50 100 100 n/a 83 n/a Yes —APH St Bipolar St (b) 100 100 100 100 100 100 100 100 100 100 100 100 Yes YesDROe St Bipolar St P05661 100 0 100 66.7 100 83.3 62.5 100 100 75 88 79 — —DRO St Bipolar St P05661 100 0 100 0 100 83.3 62.5 100 100 64 88 72 — —SCH Sm Bipolar St Q02456 100 100 100 0 100 100 62.5 100 100 83 88 85 Yes —DOR St ? St G4V4Y6 100 100 100 0 100 100 56.3 100 88 83 81 83 Yes (c) —PLA St Bipolar St Q26079 100 100 100 0 100 100 56.3 100 100 83 85 84 Yes —
Vertebrates MUS Ca Bipolar St Q91Z83 100 100 92 0 100 100 87.5 100 88 82 92 85 Yes —HOM Ca Bipolar St P12883 100 100 92 0 100 100 87.5 100 88 82 92 85 Yes Yes (d)DAN Ca Bipolar St Q9PVE1 100 100 92 0 75 100 87.5 100 88 78 92 82 Yes —
Invertebrates AMP St No St I1GH71 100 100 66.7 0 75 75 62.5 100 100 69 87 75 — —MHC II Smooth-& Nm-likesequences
Vertebrates GAL Sm Side polar Sm P10587 100 100 92 92 75 50 87.5 100 96 85 94 88 Yes Yes (e)MUS Nm Bipolar
Mini-filamentsNm Q61879 100 100 92 92 75 50 87.5 100 96 85 94 88 Yes —
Invertebrates AMP Nm No Nm I1GH58 100 100 0 0 66.7 50 68.8 100 79.2 53 83 63 — —Amoebazoa DIC Nm Mini-filaments Nm P08799 75 0 66.7 0 0 0 0 58.3 62.5 24 40 29 — —
ACA Nm Mini-filaments Nm P05659 0 0 100 0 0 66.7 68.8 0 50 28 40 32 No —Myosin Vsequences
Vertebrates Va — — — Q9Y4I1 88 0 92 0 75 50 0 0 92 51 31 44 No —Vb — — — Q9ULV0 63 0 92 0 75 41.7 0 0 79.2 45 26 39 No —Vc — — — Q9NQX4 0 0 66.7 0 75 41.7 0 0 88 31 29 30 No —
The conservation of positive and negative residues required for establishing each one of the critical “d.1”, “d.2”, and “f.2”, intramolecular interactions in a specific species versus tarantula(Aphonopelma), are shown as a percentage (%) and the conservation score Cs (%) integrating these interactions altogether (see Materials and Methods). The score for the contributions of“d.1” and “d.2” (Cs-d) related to IHM stability and “f.2” (Cs-f) related to IHM formation are also shown. The amino acids involved on both sides of the interactions analyzed were as follows: “d.1”i: D391 R164, ii: K394 K400 E165, and iii: E406 K449 R659; “d.2” i: E292 D302 K734, ii: E371 K763 R500, and iii: E376 E379 K721; and “f.2” i: E893 E895 R659 H665, ii: D901 E904 K523,and iii: R905 K908 K913 E532 E533 E534. Acronyms used for the species are as follows: CON: C. gigantea (anemone), APH: Aphonopelma sp. (tarantula), DROe: Drosophila melanogaster(flight muscle embryonic isoform), DRO:D. melanogaster (flight muscle), SCH: Schistosomemansoni, DOR:Doryteuthis pealeii (squid), PLA:Placopecten magellanicus (scallop), MUS:Musmusculus (mouse), HOM: Homo sapiens, DAN: Danio rerio (zebrafish), AMP: Amphimedon (sponge), GAL: Gallus (chicken), ACA: A. castellani, DIC: D. discoideum, Va-c: myosin V H.sapiens isoforms. The MHC entry sequences are detailed in Supplementary Tables 3. Muscle and MHC types: St: Striated, Ca: Cardiac, Sm: Smooth, Nm: Non-muscle. Notes: (a) Foranemone, only partial MHC sequences (Aulactinia verrucosa Q8MV94, Anemonia sulcata Q8MV95, Nematostella vectensis I2G9D5) were available for the interactions as follows: “d.1” i(Nematostella), ii (Nematostella), iii (Nematostella, Aulactinia, Anemonia), “d.2”ii (Nematostella, Aulactinia), and “f.2” i-iii (Aulactinia, Anemonia). n/a represents lack of sequence informationin two anemoneMHC sequences. (b) Tarantula AphonopelmaMHC sequence (KT619079) is shown in Supplementary Data. (c) IHM presence was concluded by comparing the SAXS profileof L. peali (squid) with the predicted scattering profile for tarantula PDB 3DTP [13]. (d) The presence of SRX state has been confirmed in human cardiac muscle (Dr. JamesW. McNamara andRoger Cooke, personal communication) andOryctolagus (rabbit) cardiac muscle whose MHC sequence is very similar to the mouse one; thus, it was not included in Supplementary Table 3.(e) The presence of SRX state has been confirmed in chicken gizzard smooth muscle (preliminary result, Dr. Ed Pate and Dr. Roger Cooke, personal communication).
1149Myosin
Interacting-Heads
Motifs

1150 Myosin Interacting-Heads Motifs
(4) -Interaction at the RLC–RLC interface: theRLCs of the blocked and free heads interactwith themselves across blocked head RLCdomain 1, such that the positively charged Lhelices are packed due to complementarycharges against the blocked head domain 1[23], as analyzed by Brito et al. [24]
Intermolecular interactions
Intermolecular interactions maintain IHMs sus-pended above the backbone surface, contacting itonly in one site:Tarantula thick filament in a relaxed state shows
that the IHMs are located above the backbone surface(Fig. 1), with their S2s slightly angled (~6°) andseparated ~2 nm from the surface in the “swivel”region (Fig. 3b). The model shows that the blockedhead is the only part of the IHM that contacts thebackbone through three backbone-anchoring inter-molecular interactions—“h”, “i”, and “j”—with the twoneighboring extended S2s (Fig. 3b). In other words,the IHMs are above the backbone and covalentlyanchored to it only through their own S2s, while theother eight interactions (“a”, “h”, “i”, “j”, and two pairs of“b” and “c”) are electrostatic (Fig. 3). These sus-pended IHMs are interconnected, forming four coaxialhelical tracks that are separated from the backbonesurface and surrounded only with sarcoplasm solutionmolecules, which shield the IHMs electrostatically.There are five intermolecular interactions (Figs. 2
and 3). Two are between adjacent IHMs (“b” and “c”;Figs. 2b and 7a), establishing the helical tracks ofIHMs, and three are anchoring interactions of eachIHM onto two neighbor S2s (“h”, “i”, and “j”; Figs. 3and 7b and c). Loops involved in the “c” [5] and “b”[23] IHM-interconnecting interactions are shown inSupplementary Figs. 8 and 9, and the loops involvedin the “h”, “i”, and “j” backbone-anchoring interac-tions are shown in Supplementary Figs. 10–12 (seealso Supplementary Tables 1 and 2). In the tarantulamodel, the IHM-interconnecting interactions couldonly be used to analyze the intermolecular interac-tions of species exhibiting four helical tracks, likearthropods [5–7,53] and Platyhelminthes [12], asthese interactions seem to be different in filamentswith three [2,3] or seven [8] helical tracks.
(1) -Interaction “b”: the free head I loop and Cloop are close to the blocked head RLCNTE (Fig. 7a; Supplementary Fig. 8 andSupplementary Table 2).
(2) -Interaction “c”: the free head CM loop isnear blocked head NTE-ELC and free headloop 3 is near blocked head ELC residuesK60-F67 (Fig. 7a; Supplementary Fig. 9 andSupplementary Table 2).
(3) -Interaction “h”: the blocked head SH3domain is involved in an intermolecularinteraction with an adjacent myosin tail [5](Fig. 3, Fig. 7b, and Supplementary Fig. 10).The interaction “h” “holds” the blocked headSH3 domain, anchoring it on top of a neighbormyosin tail every 14.5 nm along each of thefour helical tracks of IHMs [5,23].
(4) -Interaction “i”: this interaction occurs be-tween the blocked head ELC and S2 of aneighbor IHM (Figs. 3b and 7b andc; Supplementary Fig. 11). Together withinteraction “h”, intermolecular interaction“i” can help “park” the blocked head IHM byanchoring it back onto the filament backbone.
(5) -Interaction “j”: this interaction occursbetween the converter and relay regionsof the blocked head with the S2 of aneighboring IHM. Both domains can alsointeract with the neighboring subfilament(Figs. 3b and 7b and c; Supplementary Fig.12). Together with interactions “h” and “i”,intermolecular interaction “j” can also help“park” the blocked head IHM by anchoring itback onto the filament backbone.
Analysis of conservation of interactions
We evaluated the conservation of specific resi-dues involved in intramolecular and intermolecularinteractions by performing alignments of MHCsequences (Supplementary Figs. 1–12) for 68species (Supplementary Tables 3–5) and orderingby their MHC, ELC, and RLC trees (SupplementaryFigs. 13–15). According to the MHC sequencealignment tree (Supplementary Fig. 13), the MHCsof these species can be classified in two generalgroups: (1) striated-like and (2) smooth-like andnon-muscle-like [12]. To analyze the conservation ofintramolecular interactions (Table 1), we includedspecies in which IHMs were detected or are currentlystudied by electron microscopy. The RLC–RLCinteractions [23] and interactions “g”, “f”, “e”, and“d” are very important for the formation and stabilityof IHM. Interaction “f.2” is a key part of the formationof IHM (Figs. 2 and 6), as it docks the blockedhead on top of the S2, forming the precursor S2-BH(Fig. 8b). Interactions “d.1” and “d.2” are essentialfor binding the free head on this precursor S2-BH(Fig. 8b), which is important for IHM stability. Therelative strength of these five interactions shouldcontrol the stability of the assembly of the twomyosin heads once the compact IHM is formed. Forthe analysis, we did not include interactions “a” and“f.1” (Supplementary Figs. 1 and 2) because loop 2exhibits variable length across species, and

1151Myosin Interacting-Heads Motifs
interactions “e” and “g” (Supplementary Figs. 4 and 5)were too complex to be analyzed using our approach.To analyze the conservation of intramolecular interac-tions “d.1”, “d.2”, and “f.2”, we defined a conservationscore (Cs; seeMaterials andMethods) that includes theconservation of the residues involved in each interac-tion, allowing ±3 residues to take into account thelimited resolution of the 3D map. Scores Cs–d and Cs–fwere defined to assess the conservation of interactions“d.1”, “d.2”, and “f.2” in terms of IHM formation andstability. In Table 1, for each of the selected species(Supplementary Tables 3–5), we analyzed the relation-ship between the conservation of residues involved incritical IHM “d.1”, “d.2”, and “f.1” intramolecular interac-tions and their Cs, Cs–d, and Cs–f scores in comparisonwith their muscle type (striated, cardiac, and smooth),MHC type (striated-, smooth- or non-muscle-like), thickfilament type, and IHM. In the alignments, we includedisoforms a–c of myosin V, in which regulation does notinvolve head–head interaction [54]; thus, the conservedresidues involved in potential ionic interactions specificto myosin II could be identified, as was performedpreviously [17] (Supplementary Figs. 1–12).
Conservation of intramolecular interactions
The most conserved interactions seem to be freehead S2 ring 1 loop H (“f.2”) and the CM loop with thecatalytic domain near loop 2 (“d.1”, Y653-H665). Theconverter and C loop, which are also involved in thecontraction process, may be less conserved de-pending on the muscle type, as can be seen forinteraction “d.2”. The sequences of the conservedmyosin ATP-binding site, relay, and converter arefundamentally involved in the myosin ATPasefunction and the production of power stroke force.The sequences of loops 2, 3, C (also called loop 4),CM, and H (Supplementary Tables 1 and 2) areinvolved in the myosin motor function, as well as inthe formation and stability of IHM and the conservedactin-binding interface [55,56]. Table 1 shows thatthe residues involved in interactions “d.1”, “d.2”, and“f.2” (Supplementary Figs. 3, 6, and 7) (cf. Ref. [17])are conserved in all MHC striated-like sequencesin invertebrates (Cs ~ 72–85%, including spongestriated MHC) and vertebrates (Cs ~ 82–85%),smooth-like sequences in vertebrates (Cs ~ 88%),non-muscle-l ike sequences in vertebrates(Cs ~ 88%), and sponge non-muscle MHC(Cs ~ 63%). The Cs values for primitive non-animalAmoebozoans (Cs ~ 29–32%) are close to myosin V(Cs ~ 30–44), suggesting that their interactions areless conserved than in animals. The analysis ofseparate Cs–f and Cs–d contributions suggests thatthe residues involved in interaction “f.2”, which areassociated with the formation of IHM, are moreconserved (Cs–f ~ 81–94%) than in interactions “d.1”and “d.2”, which are associated with the stability ofIHM (Cs–d ~ 53–85%).
Conservation of intermolecular interactions
Conservation of the IHM-interconnecting interac-tions “b” and “c”
We restricted the alignment and conservationanalysis of residues involved in IHM-interconnectinginteractions “b” and “c” to species with long RLC NTE(Supplementary Table 5) and short ELC NTE(Supplementary Table 4) sequences. This wasperformed because in the tarantula CPA mecha-nism, the activation actuator control is located in along RLC NTE where phosphorylatable serines arelocated [26,27]. Light chain NTEs are short or longdepending on the species: RLC NTEs are long in 25invertebrate species (and may exist at the protein ortranscript level in the National Center for Biotech-nology database) but short for the rest, while theconverse is true for ELC NTEs [23,51,52]. Inparticular, the long RLC NTE present in arthropodsand Platyhelminthes has been found to be function-ally involved in the destabilization of helical tracks ofIHMs as a result of the CPA mechanism in tarantulathick filament [23–25]. We did not attempt to analyzethese intermolecular interactions in vertebrate ormollusk IHMs as their 3D map interaction “b” regionis different from the arthropod we are studying.The actual mass density of interaction “b” isclearly seen in the 3D maps of thick filaments fromarthropods [5–7,23] and Platyhelminthes [12], sug-gesting that it plays a key role in establishing the verystable helical tracks of IHMs in thick filaments. Incontrast, in vertebrate and human cardiac muscles,interactions “b” and “c” are only established in one ofevery three crowns, with only interaction “b” presentin the other two crowns. Both interactions “b” and “c”have a neutral net charge and complementarycharges of the free head motor domain and adjacentfree head RLC NTE in interaction “b” and the freehead and adjacent blocked head ELC NTE ininteraction “c” (Supplementary Table 4 and Supple-mentary Fig. 8). Therefore, we conclude thatintermolecular interactions “b” and “c” are criticallyinvolved in maintaining the helical tracks of IHMs inspecies with long RLC NTEs and short ELC NTEs,like arthropods [23–25] and Platyhelminthes [9,23].In both interactions, the MHC side conservesspecific negative and positive residues on I andC loops (interaction “b”) or CM loop and loop 3(interaction “c”), as well as on the long RLC NTE(interaction “b”) or short ELC NTE (interaction “c”)sides (Supplementary Figs. 8 and 9).
Conservation of the backbone-anchoring interac-tions “h”, “i”, and “j”
A quasi-atomic model of the tarantula myosinsubfilaments that comprise the outer myosin layer ofthe thick filament backbone is not available yet; thus,

1152 Myosin Interacting-Heads Motifs
an analysis of the backbone-anchoring intermolec-ular interactions “h”, “i”, and “j” (Fig. 7b and c) cannotbe properly performed at this time. We analyzed onlythe tail side of these three interactions, which seemto involve several conserved negative and positiveresidues (Supplementary Figs. 10–12). Since theSH3 domain of each IHM is located every 14.5 nmalong each of the helical tracks in interaction “h”, itseems plausible that alternating positive and nega-tive charges—which are not present in myosin V,in which regulation does not involve head–headinteractions—on the subfilament surface shouldhelp guide the six SH3 conserved charged residues(K29/K30, E37/K38, K67, and D69) for docking theblocked head back on the backbone of the thickfilament (Supplementary Fig. 10). The presence ofsignificantly longer NTEs in all the SH3 of myosinII—which is not present in myosin V—suggestingthat it is involved in blocked head docking andestablishing the helical tracks of IHMs. Our analysisof interaction “i”, which is restricted only to specieswith short ELC NTEs (Supplementary Fig. 11),shows that six negative (E79, E84, D85, E88, D94,and E140) and four positive (K78, K91, K95, andK144) residues are highly conserved and may makecontact with subfilaments. The analysis of interaction“j” (Supplementary Fig. 12) shows that, in contrast tomyosin Vs, most myosin IIs have two negative (E744and D504) and four positive (K721, R758, H488, andK499) residues that are very conserved in theconverter and relay. We conclude that some specificresidues involved in the intermolecular interactions“h”, “i”, and “j” are crucially involved in the axial andlateral docking of the IHMs on the thick filamentbackbone and are conserved in most myosin IIsequences for interactions “h” and “j” and in shortELC NTE sequences for interaction “i”.In summary, the oppositely charged and conserved
residues involved in the five intramolecular interac-tions maintain the IHM in a compact off structure,explaining the conservation of the motif across animalspecies. The oppositely charged residues involved inthe five intermolecular interactions retain the IHMs,making helical tracks of myosin heads on the surfaceof the backbone, and are conserved in arthropods andPlatyhelminthes.
Discussion
The structure of tarantula thick filaments has beenextensively studied [5,53,57,58]. These filamentsexhibit four helical tracks of myosin heads [53](compacted as IHMs) [5,23] above the backbonesurface. Since the tarantula IHM crystal structurehad not been determined, the quasi-atomic modelPDB 3JBH (see Results) allows for provisionalanalysis of its intramolecular and intermolecularinteractions, as well as building a quasi-atomic
model of the complete tarantula myosin molecule(including its tail), a subfilament model, and thebackbone formed by a ring of 12 subfilamentsaround the paramyosin core (Alamo et al., inprogress). The tarantula myosin RLC NTE, withtwo phosphorylation sites at Ser35 and Ser45, hasbeen shown to be crucial for the relaxation andactivation of tarantula thick filaments [23], for whichwe have proposed a tarantula CPA mechanism [24–27] (Fig. 8c–f). This dual phosphorylation mecha-nism could be extended to other chelicerate striatedmuscles, like that of Limulus [6] and scorpion [7],and possibly other arthropods with thick filamentsthat exhibit four-stranded helical tracks of IHMstogether with myosin RLCs with a long NTE and twophosphorylatable serines. In contrast, a differentactivation mechanism is present in vertebrateskeletal [28,29] and cardiac muscles [30,31], whichhave thick filaments that exhibit a perturbed 3-foldhelical array of IHMs, as well as in vertebrate smoothmuscle [32], which also has IHMs but with aproposed ELC activation role [33], and mollusks,which have 7-fold helical tracks of IHMs with ELCdirect Ca2+-binding activation control. Below wediscuss how the intramolecular and intermolecularinteractions associated with the tarantula model(Figs. 2b, 6, and 7) are sequentially established,forming the IHMs, and how the helical tracks of IHMsin a relaxed state are sequentially disrupted uponactivation and sequentially reformed upon relaxation(Fig. 8, “interaction table”). This discussion is basedon the tarantula CPA mechanism and aims toexplain the structural basis of the ATP turnoverrates detected in tarantula SRX [22].
Interactions form IHMs and their helical tracks ina relaxed state
How are tarantula thick filaments formed in arelaxed state?
The intramolecular interactions are present inisolated switched-off myosin II molecules, whichform IHMs in a solution [17]. Myosin subfilaments areformed by the coiling coiled tails of three myosinmolecules with one pair of heads protruding every43.5 nm. Twelve myosin subfilaments are packedaround a paramyosin core, forming the filamentbackbone (Fig. 1). Initially, the myosin S2s emergehelically over the backbone (Fig. 2b) and the pairs ofheads are disordered and protruding, not yet formingIHMs (precursor IHMs; Fig. 8a). These heads are inthe relaxed closed conformation (Fig. 4) with RLCNTEs that are not phosphorylated. The interactionstable in Fig. 8a–c shows the four stages in which theintramolecular and intermolecular interactions couldbe sequentially involved, establishing the fullyfunctional IHMs (Fig. 8c) and forming the helicalrelaxed structure of the thick filament: (1) only one
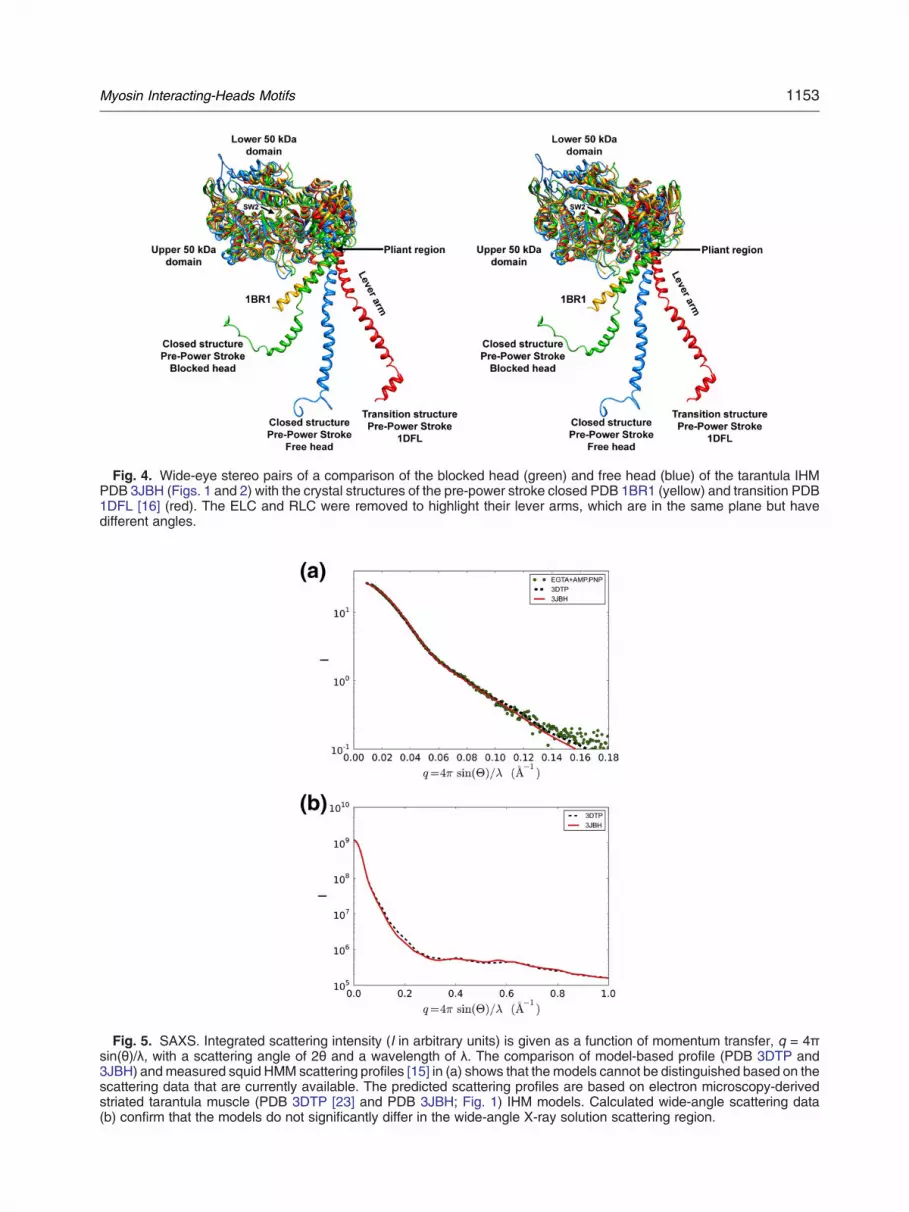
Fig. 4. Wide-eye stereo pairs of a comparison of the blocked head (green) and free head (blue) of the tarantula IHMPDB 3JBH (Figs. 1 and 2) with the crystal structures of the pre-power stroke closed PDB 1BR1 (yellow) and transition PDB1DFL [16] (red). The ELC and RLC were removed to highlight their lever arms, which are in the same plane but havedifferent angles.
Fig. 5. SAXS. Integrated scattering intensity (I in arbitrary units) is given as a function of momentum transfer, q = 4πsin(θ)/λ, with a scattering angle of 2θ and a wavelength of λ. The comparison of model-based profile (PDB 3DTP and3JBH) andmeasured squid HMM scattering profiles [15] in (a) shows that the models cannot be distinguished based on thescattering data that are currently available. The predicted scattering profiles are based on electron microscopy-derivedstriated tarantula muscle (PDB 3DTP [23] and PDB 3JBH; Fig. 1) IHM models. Calculated wide-angle scattering data(b) confirm that the models do not significantly differ in the wide-angle X-ray solution scattering region.
1153Myosin Interacting-Heads Motifs
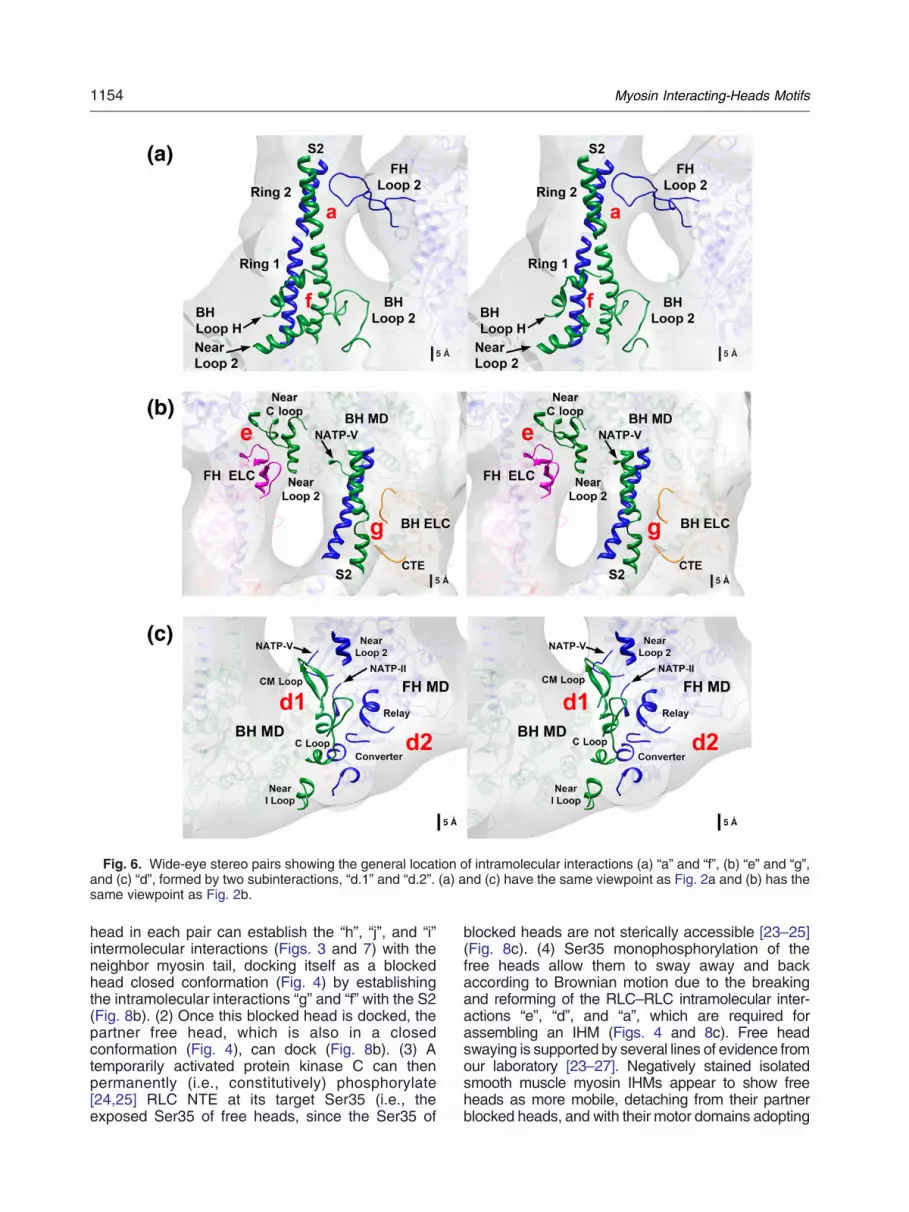
Fig. 6. Wide-eye stereo pairs showing the general location of intramolecular interactions (a) “a” and “f”, (b) “e” and “g”,and (c) “d”, formed by two subinteractions, “d.1” and “d.2”. (a) and (c) have the same viewpoint as Fig. 2a and (b) has thesame viewpoint as Fig. 2b.
1154 Myosin Interacting-Heads Motifs
head in each pair can establish the “h”, “j”, and “i”intermolecular interactions (Figs. 3 and 7) with theneighbor myosin tail, docking itself as a blockedhead closed conformation (Fig. 4) by establishingthe intramolecular interactions “g” and “f” with the S2(Fig. 8b). (2) Once this blocked head is docked, thepartner free head, which is also in a closedconformation (Fig. 4), can dock (Fig. 8b). (3) Atemporarily activated protein kinase C can thenpermanently (i.e., constitutively) phosphorylate[24,25] RLC NTE at its target Ser35 (i.e., theexposed Ser35 of free heads, since the Ser35 of
blocked heads are not sterically accessible [23–25](Fig. 8c). (4) Ser35 monophosphorylation of thefree heads allow them to sway away and backaccording to Brownian motion due to the breakingand reforming of the RLC–RLC intramolecular inter-actions “e”, “d”, and “a”, which are required forassembling an IHM (Figs. 4 and 8c). Free headswaying is supported by several lines of evidence fromour laboratory [23–27]. Negatively stained isolatedsmooth muscle myosin IHMs appear to show freeheads as more mobile, detaching from their partnerblocked heads, and with their motor domains adopting
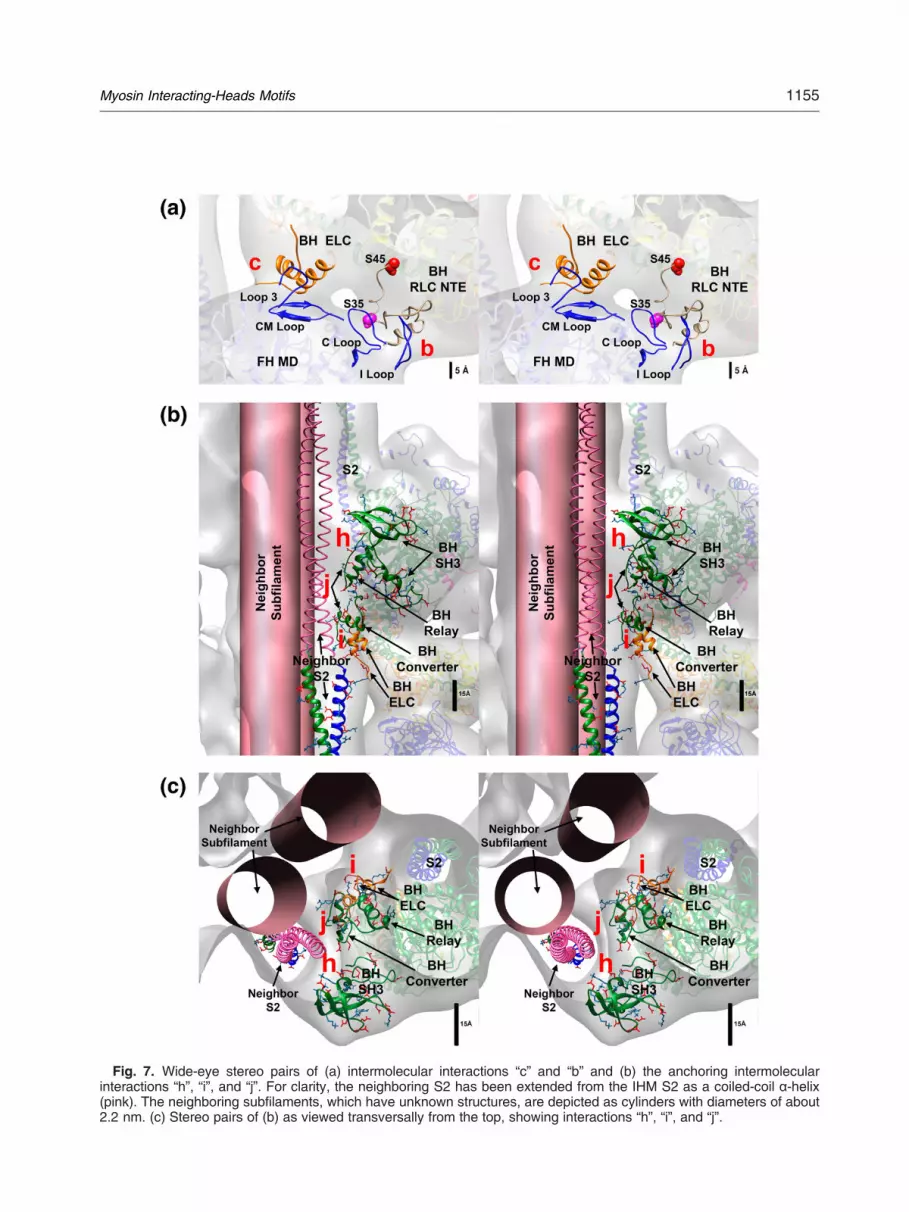
Fig. 7. Wide-eye stereo pairs of (a) intermolecular interactions “c” and “b” and (b) the anchoring intermolecularinteractions “h”, “i”, and “j”. For clarity, the neighboring S2 has been extended from the IHM S2 as a coiled-coil α-helix(pink). The neighboring subfilaments, which have unknown structures, are depicted as cylinders with diameters of about2.2 nm. (c) Stereo pairs of (b) as viewed transversally from the top, showing interactions “h”, “i”, and “j”.
1155Myosin Interacting-Heads Motifs

Fig. 8. Sequential formation, disruption, and reformation of intramolecular and intermolecular interactions (“interactionstable”) in the tarantula IHM PDB 3JBH model upon relaxation (c), activation (c to f, green arrows) and relaxation afteractivation (f to c, red arrows) according to the CPA mechanism (c–f) proposed for tarantula thick filaments [24–27] allowexplanation of the structural basis of the ATP turnover rates detected in tarantula relaxed and Ca2+-activated states [22].(a) Model of a short segment of a precursor tarantula thick filament showing three precursor IHMs with disordered headsand unphosphorylated Ser35 (black circles). The heads do not make any intermolecular or intramolecular interactions.(b) Only one head of each precursor IHM in the pre-power stroke closed state (Fig. 4, green) can establish the threeanchoring intermolecular interactions and dock them as blocked heads (light green) to the backbone. The free head (lightblue), also in the pre-power stroke closed conformation (Fig. 4, blue), can establish intramolecular interactions with thedocked blocked head, which are needed to assemble the IHM. (c) In a relaxed state, these precursor IHMs become fullyfunctional after half the Ser35 are monophosphorylated by a temporarily activated protein kinase C (PKC) (brown arrow).PKC can only phosphorylate the fully exposed Ser35 of the free heads (blue heads), as the Ser35 of the blocked heads(green heads) are not accessible [23,24]. Ser35 monophosphorylation of the free heads allows the free heads to swayaway and back by Brownian motion (“swaying” heads) by breaking and reforming the intramolecular interactions (denotedby “±” in the “interactions table” and by dotted curved arrows in the illustration). (c–f) The tarantula CPA mechanismproposed for tarantula thick filament [24,25]. The interactions table shows how the intramolecular and intermolecularinteractions established in the relaxed state (c) are progressively removed (denoted by crossing them out) upon activation(d–f), ending in a disordered array (f) with fewer interactions. The table also shows how these interactions areprogressively reformed from this final disordered array (f) to a newly achieved ordered relaxed array (c). The IHM modeland CPA mechanism allow explanation of the structural origin of the very slow (N1800 s), slow (250–300 s), and fast(b30 s) ATP turnover rates detected in tarantula striated muscle [22] in a relaxed state: (1) The very slow rate is associatedwith unphosphorylated docked blocked heads in the IHM (green heads with motor domains labeled as “VS”) and the fewunphosphorylated docked free heads in the IHM (data not shown), (2) the slow rate is associated with the Ser35monophosphorylated free heads (blue heads with motor domains labeled as “S”), and (3) the fast rate is associated withthe Ser35 monophosphorylated swaying free heads that are undocked from the IHM (blue heads with motor domainslabeled as “F”). Therefore, the docked (helically ordered) unphosphorylated blocked heads and the few free heads with avery slow rate, as well as the transiently docked Ser35 monophosphorylated free heads with a slow rate, are responsiblefor super-relaxation (SRX) in tarantulas [22], while the remaining (disordered) swaying free heads that move according toBrownian motion and are undocked from the IHM exhibit the fast rate detected in the disordered relaxed state [22]. Incontrast, the very fast (b0.1 s) rate detected in the Ca2+-activated state in tarantula striated muscle [22] is associated withSer35 monophosphorylated free heads (blue heads with motor domains labeled as “VF”), Ser45 monophosphorylatedblocked heads (green heads with motor domains labeled as “VF”) and biphosphorylated free heads (blue heads with motordomains labeled as “VF”) that are bound to actin (yellow spheres) on the activated thin filament. FH: free head, BH: blockedhead.
1156 Myosin Interacting-Heads Motifs

1157Myosin Interacting-Heads Motifs
different orientations [59]. In addition, X-ray diffractionstudies show that a fraction of constitutively on motorsallow the muscle to immediately respond to calciumactivation [60] (cf. Ref. [24]).
Intramolecular interactions are involved informing IHMs
Intramolecular interaction “f.2”, which is involved inthe formation of the IHM, is more conserved in all thestudied animal species than in non-animal species
The IHM is ubiquitous. Table 1 shows that the IHMis present in all animal species with muscles,irrespective of the muscle type (striated skeletal,cardiac, or smooth), filament type (bipolar, sidepolar), or MHC type (striated-, smooth-, or non-mu-scle-like). The wide presence of IHM correlates withthe conservation of residue charge in the “d.1”, “d.2”,and “f.2” interactions in all animal species shownin Table 1 (Cs N ~ 63%). The ranges ofconservation—Cs–f ~ 40–94% and Cs-d ~ 24–85%—for these critical interactions on MHC IIsuggest matched variations in the formation andstability of the IHM as these species evolved. On onehand, the absence of IHM in the Amoebozoan A.castellani correlates with a smaller conservationpercentage (Cs ~ 32%), suggesting the importanceof these critical interactions in the formation andstability of the IHM. On the other hand, the chickenvertebrate smooth muscle head–tail interaction islikely the most stable interaction as it establishesthe motif structure [35] with the smallest rateof product release [61] and results in a higherconservation percentage of the residues involved inthe formation and stability of the IHM (Cs–f and Cs–d,~94%). Also, studies with isolated 10S myosin IIHMM from smooth- and non-muscle-like MHCsequences suggest that the interaction betweenthe blocked head and S2 is required for the formationof a stable IHM [17,61]. This agrees with the similarresidue conservation values in vertebrate smooth-like (Cs–f ~ 94%) and non-muscle-like MHC se-quences (Cs–f ~ 94%). Finally, it has been sug-gested that the strength of the intermolecularinteractions is weaker in unregulated myosin, show-ing that IHMs are less common in solution thanregulated myosin [17]. This is in accordance withthe smaller conservation values (Cs–d ~ 78–82%) invertebrate cardiac muscles from mice, humans, andzebra fish. The consistency of the results amongspecies that was achieved with this interactionscoring approach reassured our confidence in ourinteraction analysis in spite of the clear resolutionlimitations of the 3D map and the lack of definition ofthe precise residue contacts that are involved in theintramolecular interactions. We conclude that “d.1”,“d.2”, and “f.2” intramolecular interactions are criti-cally involved in the formation and stability of the
IHM. The conservation of residues involved inthese interactions supports the conservation of theIHM across all animal species, irrespective of thepresence of muscles (like in sponges that lackmuscle), muscle type, filament type, or MHC II type.The conservation analysis of these interactionssuggests that they are more conserved in animalsthan in primitive non-animals, supporting our con-clusion—which is based on the fact that the IHM wasdetected by electron microscopy [14]—that IHMsevolved after Amoebozoa and animals diverged.
Intermolecular interactions are involved inthe formation of the helical tracks of IHMsalong the filaments
For arthropods such as tarantulas, in whichactivation is dependent upon phosphorylation, thetwo intermolecular interactions “b” and “c” and theremaining anchoring interactions “h”, “i”, and “j”(Figs. 3, 6, and 7) maintain the stability of the helicaltracks of IHMs more than in vertebrates or mollusks.
IHM interconnecting interactions
The “b” and “c” intermolecular interactions arecritically involved in the maintenance of the helicaltracks of IHMs in species with long RLC NTEs andshort ELC NTEs, such as arthropods [23–25]and Platyhelminthes [9,23]. In both interactions, theMHC side conserves specific negative and positiveresidues on I and C loops (interaction “b”) or CM loopand loop 3 (interaction “c”), as well as on the long RLCNTE (interaction “b”) or short ELC NTE (interaction “c”)sides (Supplementary Figs. 8 and 9). This analysiscould be extended to the similar thick filaments of thePlatyhelminthes Schistosome.
Backbone-anchoring interactions
Some specific residues involved in these intermo-lecular interactions are crucially involved in the axialand lateral docking of IHMs on the filamentbackbone, which are conserved in most myosin IIsequences (interactions “h” and “j”) and short ELCNTE sequences (“i”).
MHC type controls filament type formation
According to the average distance tree for MHC IIsequence alignment (Supplementary Fig. 13), MHCscan be classified into three types: striated-, smooth-,and non-muscle-like. Table 1 shows that the MHCtype controls filament type formation: striated-likeMHC forms helical bipolar thick filaments with variousnumbers of helical tracks (3-7), while smooth-likeMHC forms side-polar thick filaments and non-mu-scle-like MHC forms bipolar mini-filaments.

1158 Myosin Interacting-Heads Motifs
Implications of myosin interactions in the evolution-ary origin of the switched off state
This state may have arisen early in species'evolution as a means of switching off non-musclemyosin. Non-muscle myosin II is monomeric in an offstate; thus, only intramolecular interactions betweenheads would have been possible. As musclesevolved, they may have retained these interactions.In filaments, additional (intermolecular) interactionsbecame possible and advantageous in an evolution-ary sense. Thick filaments could be formed first onparamyosin cores, allowing various numbers ofhelical tracks of IHMs: seven in mollusk striatedmuscle, four in arthropod and Platyhelminthesstriated muscle, and three in vertebrate skeletal orcardiac muscle when the paramyosin core was lost.Additionally, IHM regulation could also have evolvedfrom being controlled by direct ELC Ca2+ binding(mollusks) or RLC phosphorylation (in arthropodsand possibly in Platyhelminthes) or being unregulat-ed in vertebrates.
On activation interactions are disrupted,disassembling IHMs and their helical tracks
The “interactions table” in Fig. 8c–f shows whichintramolecular and intermolecular interactions are es-tablished in a relaxed state (Fig. 8c) and which inter-actions are progressively removed after a relaxedtarantula thick filament is activated (Fig. 8d–f), suchthat helical tracks of IHMs are disassembled, with thedisordered free and blocked heads protruding awayfrom the backbone (Fig. 8d–f), as observed bynegative staining [62] and equatorial X-ray diffraction[63]. We have shown that the structural differencesand location within the IHM could preset the orderin which the free and blocked head are released uponphosphorylation, having the free head the rightconformation (Fig. 4) to be released first to interactwith actin, followed by the blocked head (if required)[24,25].
Interactions sequentially reform, comprisingIHMs and their helical tracks upon relaxation
We propose that the disordering of myosin headsproduced by activation (Fig. 8c–f) is restored afterrelaxation in three independent steps, depending onwhich step the motif (Fig. 8f to c) is as follows.
Formation of precursor blocked head-S2 motif
Once the monophosphorylated Ser45 of a blockedhead is dephosphorylated (Fig. 8e to d, top IHM), thedisordered blocked head can eventually re-establishintramolecular interactions “g” and “f” with its own S2(Fig. 8d, top IHM) and reattach to the two neighborS2s (Fig. 7) by re-establishing the anchoring
intermolecular interactions “h”, “j”, and “i”. This allowsthe blocked head to be docked back and anchoredprecisely in the correct axial position (i.e., every14.5 nm). It has been reported that interactionbetween the blocked head and its own S2 is neededto form a stable IHM [17,61]. These intramolecularinteractions can be established between a blockedhead and its S2 in isolated HMM molecules, incontrast to the intramolecular interactions “h”, “i”, and“j”, which can only be established in a filament.
Reassembly of IHM from a blocked head S2precursor motif
Once the partner diphosphorylated free head of adocked blocked head is Ser45 dephosphorylated bymyosin light chain phosphatase, again becominga Ser35 monophosphorylated swaying free head(Fig. 8d to c, middle IHM), they could eventually dockback onto the blocked head S2 precursor motif byre-establishing first the closer RLC–RLC interactionsbetween both blocked and free heads and then theremaining intramolecular interactions “e”, “d”, and“a”. This reassembly should be guided by thecomplementary charges of domain 1 of theblocked and free head RLC NTEs, as we haveproposed [24]. Thus, the RLC domain 1 seems tobe better conserved than the rest of the RLCdomains [64].
Formation of helical tracks of IHMs
When a similar process occurs in axially adjacentreassembled IHMs, pairs of “c ” and “b ”interactions can be re-established between them(Fig. 1). This stabilizes first an initial patch of thehelix, which can extend when additional patches arere-established. This finely tunes the formation ofhelical tracks of IHMs with a subunit distance of14.5 nm, been the 43.5 nm helical repeat enforcedby the 12 myosin subfilament symmetry.
Implications of IHM interactions onsmooth muscle
It was striking to find that vertebrate smoothmuscle IHM [11,34,35] was present in invertebratestriated muscle [5] with similar IHM models (PDB1I84 [35] versus PDB 3DTP [23]). Also, the thickfilaments present in invertebrate striated muscle [5]were similar to invertebrate smooth muscle [12] withsimilar IHMs. In addition, a striking similarity betweenthe myosin II RLCs of Schistosome and tarantulawas observed: both RLCs exhibit a long NTE withtwo putative phosphorylatable serines [23,24],suggesting that our present analysis in this papercould be applied to the smooth muscle IHM ofPlatyhelminthes. Concerning vertebrate smooth

1159Myosin Interacting-Heads Motifs
muscle, which has an unknown specialized side--polar thick filament structure, we cannot infer howthe specific IHM interactions in vertebrate smoothmuscle could be involved in assembling non-helicalside-polar filaments. Trybus et al. [61] have dis-cussed in detail the need for vertebrate smoothmuscle to have two myosin heads, head–headinteractions, and S2 for regulation by RLC phos-phorylation. We can only mention that S2, which issuggested to mediate specific interactions with thehead that are required to achieve an off state invertebrate smooth and non-muscle myosin II [61]and is essential for regulation [61], should beinvolved in IHM intramolecular interactions similarto “a”, “f”, and “g”. Supporting Trybus et al. [61], ourresults with tarantulas suggest that these interac-tions could correspond to interactions “a”, “f”, andperhaps “g”, explaining the necessity of a minimallength of S2 for regulation [61]. Two of theseinteractions, “a” and “f”, are involved in the interac-tion of S2 rings 2 and 1 with free head loop 2 andblocked head, which is part of myosin II regulation insmooth and skeletal muscle [42].
Structural basis of the tarantula muscleSRX state
The two heads in the IHM have inhibited ATPase:the blocked head exhibits a closed conformation [23]of the nucleotide pocket switch 2 (Fig. 4), preventingphosphate release [46,47], and it is “parked”, orlocked in, to the backbone (Fig. 8c) with ATP activitythat is sterically “blocked”, as its actin-bindinginterface is positioned on the converter domain ofits partner free head. This inhibits ATPase activity bystabilizing the converter domain movements neededto release phosphate [11]. Relaxed vertebrateskeletal and cardiac muscle fibers exhibit a slowATP turnover rate [18,19]. Naber et al. [22] proposedthat this is an adaptation to save energy in animalssuch as tarantulas that spend long periods immobilewhile poised to quickly capture prey. This state—which strongly inhibits myosin ATPase activity andhas been observed in a variety of muscle type-s—was called the SRX state in the pioneering workof Cooke et al. [20] Its importance in muscle hasbeen described by many authors [18–20,22,65] (seeRef. [66]). Structurally, SRX has been ascribed to theIHM [18,20]. In relaxed tarantula muscle, three ratesare detected: [22] very slow (N1800 s), slow (250–300 s), and fast (b30 s). Based on the PDB 3JBHmodel, its interactions, and the tarantula CPAmechanism [14,24,26,27] (Fig. 8c–f), we ascribethe very slow rate to the docked unphosphorylatedheads, the slow rate to phosphorylated dockedheads, and the fast rate to phosphorylated undockedheads. Therefore, as shown in Fig. 8c and explainedbelow, the docked (helically ordered) unphosphory-lated blocked and a few free heads, which both have
a very slow rate, and the transiently docked Ser35monophosphorylated free heads, which have aslow rate, are responsible for the two SRX levels intarantulas [22], while the remaining (disordered)swaying free heads with Brownian motion exhibit thefast rate detected in the disordered relaxed state.
Structural basis of the very slow, slow, fast, andvery fast rates in tarantula striated muscle
(1) Very slow rate origin: the very slow rateis ascribed to unphosphorylated blockedheads and the few unphosphorylated Ser35free heads docked in the IHM motif. TheATPase rate is determined by the strengthof the intramolecular interactions, and sincethis muscle is thick filament regulated,some other mechanism momentarily un-docks or activates the myosin. The veryslow rate is expected to exist in otherthick-filament-regulated filaments. In relax-ation, all blocked heads are unphosphory-lated, as endogenous myosin light chainphosphatase is active, dephosphorylatingany Ser45 monophosphorylated blockedheads. The blocked head rate is very slow,as its ATPase activity is sterically “blocked”[11] and locked in to the backbone [26,27].This “parking”, or locking in, of half theavailable myosin heads in a close confor-mation is an effective mechanism for savingenergy by preserving bound ATP.
(2) Slow rate origin: the slow rate is ascribedto docked swaying Ser35 monophosphory-lated free heads (Fig. 8c), for which ATPactivity is transiently inhibited while dockedin the IHM motif.
(3) Fast rate origin: the fast rate is ascribed toundocked swaying Ser35 monophosphory-lated free heads (Fig. 8c) and interpreted asa way to enable at least half of the availableheads to explore as structural sentinels ifthere are any activated thin filaments, tosave energy, and to make a faster transitionto active states if there are activated thinfilaments nearby, favoring quick force pro-duction similar in single twitches or twitchsummation [24]. The Ser35monophosphor-ylation is constitutively (i.e., permanently)present on most free heads, suggestingthat it is a simple way to segregate headsthat are better located to quickly swayaway by Brownian motion and interactwith Ca2+-activated thin filaments [24].

1160 Myosin Interacting-Heads Motifs
(4) Very fast rate origin: in contrast to therelaxed state (Fig. 8c), remnants of thesethree heads' populations (Fig. 8d and e)briefly coexist in an activated state (Fig. 8d–f)together with a quickly increasing fourthpopulation that is formed by released sway-ing Ser35-monophosphorylated free heads,Ser45-monophosphorylated blocked heads,and diphosphorylated free heads, which arein the transition pre-power stroke conforma-tion [23] (Fig. 4) and are activated whenbound to an activated thin filament located~5 nm away [46]. This population is associ-ated with the very fast time constant (b0.1 s)detected in active muscle fibers [18] (Fig. 8f),as a high rate is required after activationduring muscle contraction.
In relaxed tarantula thick filaments, a “closed”conformation that prevents phosphate release isrequired for the formation of helical tracks of IHMs onthe backbone [46]. Blebbistatin, which inhibits myosinII ATPase activity [67], stabilizes helical tracks in arelaxed state by promoting a closed state [68]. Thesetwo results suggest that in tarantula blebbistatin shouldclosely hold the two heads on IHMs, diminishing thefree heads that sway away and implying a very slowrate. In fact, blebbistatin stabilizes the SRX state inskeletal muscle, producing a very long-lived myosin–nucleotide complex with a very slow time constant [65],which favors our interpretation of the very slow rate intarantulas. Additionally, in the presence of blebbistatin,the spin-labeled nucleotides bound to myosin have anoriented spectrum in SRX in both slow and fast skeletalmuscles, similar to those observed in relaxed tarantulafibers without blebbistatin [65]. This strongly supportsthe proposed structural origin of the SRX state (Fig. 8).
Comparison of the SRX states of vertebrateskeletal and cardiac muscles
In contrast to tarantula muscle in which very slow(N1800 s), slow (250–300 s), and fast (b30 s) ratesare detected [22], in vertebrate skeletal [18] andcardiac [19] muscles, only slow (230 and 138–144 s)and fast rates are detected. Relaxed vertebrateand tarantula muscles have several structural andfunctional differences. Thick filaments from tarantu-las exhibit four helical tracks of IHMs with similarmotifs on the three crowns of a 43.5 nm repeat, whilevertebrate and cardiac muscles exhibit three per-turbed helical tracks in which only two crowns exhibitsimilar motifs. Additionally, cardiac muscle featuresthe cMyBP-C zone. Tarantula muscle has thickfilament activation, while in vertebrate skeletal andcardiac muscle, the thick filament acts as a
modulator. This “locks in” the tarantula unpho-sphorylated blocked head to the very slow rate andcauses it to become swaying blocked head (inducingthe slow rate) only after becoming Ser45 monopho-sphorylated blocked head by a myosin light chainkinase [23–27], which can only occur upon Ca2+
activation. In vertebrate skeletal and cardiac mus-cles, in which thick filaments are not regulated, theblocked heads sway without phosphorylation, elim-inating the very slow rate. Studies using bifunctionalfluorescence labels that are bound to intact verte-brate skeletal muscle myosin unphosphorylatedRLC to quantify the orientation of the myosin leverarm by obtaining polarization measurements re-vealed three preferred orientations in the relaxedstate: two with a long axis that is roughly parallel tothe filament axis and one that is roughly perpendic-ular [28]. In vertebrate skeletal muscle RLC, phos-phorylation disrupts the parallel orientation, shiftingthe equilibrium toward a perpendicular orientation,which permanently releases phosphorylated freeand blocked heads. Similar studies on vertebratecardiac muscle, in which almost no endogenous(constitutive) Ser15 monophosphorylation was de-tected, revealed that in a relaxed state (with unpho-sphorylated RLCs) there was a conformationalequilibrium between parallel and perpendicularstates and Ser15 monophosphorylation destabilizedthe parallel conformations, promoting a perpendicu-lar conformation [31]. We believe that the twopreferred parallel orientations are associated withthe slow rate of docked blocked and free head (SRX)in the IHM and that the perpendicular orientation isassociated with the fast rate in undocked swaying freeand blocked heads, as the free and blocked heads areactive and can sway away without being monopho-sphorylated. Therefore, in contrast to tarantula re-laxed muscle, SRX in vertebrate relaxed muscleshould ascribe the only detected rate (slow) to bothswaying blocked and free heads without permanentlydocked blocked heads.
Conservation of the SRX state
The SRX state has been detected in arthropodstriated muscle [22] and vertebrate skeletal (rabbitpsoas and soleus [18]), cardiac (rabbit [19] andhuman [Drs. James W. McNamara and RogerCooke, personal communication]), and smooth mus-cles (preliminary result; Drs. Ed Pate and RogerCooke, personal communication). The conservation ofcritical intramolecular interactions across animals andIHM structure across bilaterians (Table 1) suggeststhat SRX should be a conserved fundamental featureof IHM to save ATP.In conclusion, the tarantula quasi-atomic model
PDB 3JBH, its intramolecular and intermolecularinteractions, and the CPA mechanism show thestructural basis of SRX in tarantula muscle and its

1161Myosin Interacting-Heads Motifs
differences from vertebrate muscle. IHM conserva-tion reveals that intramolecular interactions, whichmaintain the asymmetric structure of IHM, are crucialfor maintaining muscle relaxation in animal species.
Materials and Methods
Characterization of full-length cDNA of tarantula MHC
Aphonopelma MHC sequencing follows the approach ofZhu et al. [38] The total length of the sequenced cDNA is6611 bp, with a 5862-bp open reading frame that begins atposition 126. The deduced residue sequence GenBankKT619079, which is composed of 1953 residues, containscharacteristic features of ATP-binding and actin-binding sitesof myosin. A simple comparison with fully sequenced heavychains shows that 50–64%, 31–35%, and 30% identity withsarcomeric (vertebrate striated and cardiac muscles andinvertebratemuscles), non-sarcomeric (smoothmuscle), andunicellular (Dictyostelium discoideum) MHCs, respectively.The myosin head region is more conserved (57–69% and49–50% identitywith sarcomeric andnon-sarcomericMHCs,respectively) than the coiled-coil rod (43–62% and 26%).
Image processing and 3D reconstruction
The 3D map used for flexible fitting of the tarantulahomology model was EMD-1950 with a 2.0-nm resolution,the same 3D map we used for IHM PDB 3DTP [23].
Homology modeling
Modeling of ELC
The tarantula Aphonopelma sequence was submitted tothe BioInfoBank Meta Server‡. From all the models wereceived, those with the higher scores were chosen. Fromthese, the model with all the residues in the sequence wasselected: PDB 2OVK. Two copies were made, one for thefree head and another one for the blocked head, and theywere superposed on the previous IHM structure (PDB3DTP) by the Chimera MatchMaker tool [69].
RLC modeling
The tarantula Aphonopelma sequence was submitted tothe SwissModel Server to obtain the homology model of theRLC§ using the PDB 3DTP chains E and F as a template.
Modeling of MHC (Motor domain and S2)
The sequences that correspond to the motor domainand S2 of PDB 3DTP were extracted from the full-lengthtarantula Aphonopelma MHC sequence. These 965residues were aligned to the sequence of PDB 3DTPchains A and B (free and blocked head MHC sequences)and submitted to SwissModel Server to obtain thecorresponding free head and blocked head homologymodels. The final model was assembled and saved as aPDB file using Chimera [69].
Flexible fitting
Flexible fitting was performed in seven steps. (1) Initialpreparation of the model (assembly and minimization): aninitial minimization of the structure was performed to avoidproblems during the flexible fitting procedure, as the chainscame from different sources. Using VMD [70], an explicitwater box was generated with the VMD Automatic PSFBuilder. Ions were placed in a separate step using 0.15 Msalt. Then, the solvated and ionized structure wasminimized by 2000 steps using NAMD [71]. To estimatethe effect of the minimization, we calculated the amountsof clashes before and after the minimization procedurewith Chimera. Before minimization, 2757 clashes weredetected, and after minimization, the number of clashesdecreased to 84. (2) Initial rigid-body docking: theminimized model was fitted as a rigid body to the tarantula3D map with the Situs qrange tool. (3) Preparing the PDBfile for flexible fitting: the PDB file was edited and convertedinto a suitable format for X-PLOR [72] refinement. Internalwater molecules were added to empty cavities to stabilizethe structure during flexible fitting using the Dowser program[73]. (4) Flexible fitting using Situs (2.4) and Sculptor (1.1.6):Codebook (CB) vectors were created for the structure usingSitus [74]. The generated connectivity among CB vectorswas edited to remove some bonds and provide the networkwith flexibility. The optimal connectivity network for the mapwas interactively chosen using the Sculptor (1.1.6) graphicinterface. Finally, the flexible fitting step was performedusing X-PLOR. (5) Full water box minimization of the flexiblyfittedmodel: to reduce clashes in the flexibly fittedmodel, weperformed a final minimization in the same way as the initialminimization. (6) Building two adjacent HMMs: the IHMmodel was duplicated and the copy was helically placed atthe next upper crown to generate a model with twointeracting HMMs that allow intermolecular interactions tobe studied. (7) Remodeling of the interacting loops: to seekpossible residues that can create interactions, we usedX-PLOR scripts to allow the lateral chains of interactingloops to move freely in order to find the residues that can belocated at the atomic contact distance range.
Bioinformatics analysis
We used JalView (version 2.8) [75] to analyze thesequences retrieved from the UniProt database¶. Themultiple sequence alignment was performed with Clustal(version 2.0) [76] using default parameters and orderedaccording to the average percent identity in a distance tree(Supplementary Figs. 13–15). For the conservation anal-ysis shown in Supplementary Figs. 1–12 and Table 1, wechose the MHC sequences reported for non-muscle,smooth, skeletal, striated, and cardiac muscles of the 68species shown in Supplementary Table 3. Myosin Vsequences are included for comparison, as they do notestablish IHM [17]. In addition, we chose to align themyosin 28 RLCs and 62 ELCs sequences, as shown inSupplementary Tables 4 and 5. In Table 1, we analyzedthe conservation of the residues involved in “d.1”, “d.2”,and “f.2” intramolecular interactions by ascribing a scorevalue to each subinteraction (d.1 i, ii, and iii; d.2 i, ii, and iii;f.2 i, ii, and iii) as a percentage. In the sequence alignment,each species was compared to the correspondingtarantula sequence (which was considered 100%). Thescore was 100% (1) if both sides of the subinteraction were

1162 Myosin Interacting-Heads Motifs
composed by the same number of conserved residues andhad opposite charges; (2) if the sides of the interactions had adifferent number of residues, all of whichwere conserved; and(3) if the sides of the subinteractions had a different number ofresidues, one residue that was not conserved, and the samenumber of conserved residues on each side. Otherwise, thescore was 75%, 50%, or 25% if the conservation was presentin the previous/following residue in the sequence (±1, 2, or 3amino acids, respectively) or 0% if the difference in positionwas greater than three amino acids, there was no conserva-tion on one side of the interaction, or a residue with theopposite charge was in the same position. We calculated thefinal conservation score, Cs, for each species as the averageof the “d.1”, “d.2”, and “f.2” subinteractions, as well as thescores for the contributionsof “d.1”and “d.2” (Cs–d) or “f.2” (Cs–
f). Only complete sequences with evidence at the transcrip-tional or protein level were selected for the analysis. Foranemone, we only used the few available MHC sequencefragments for different anemone species that are involved inthe “d.1”, “d.2”, and “f.2” intramolecular interactions.
SAXS analysis
The sample preparation and data collection details forsquid HMM were reported in a previous publication [15].Scattering intensity, I(q), is in arbitrary units, with q = 4πsin(θ)/λ in units of Å−1. The scattering angle is 2θ with awavelength of λ = 1.2563 Å. Scattering profiles for PDB3DTP and PDB 3JBH were computed using FoXS [77] andCRYSOL [78] to check consistency, especially at scatter-ing angles beyond q = 0.3 Å−1. CRYSOL required morethan the default number of harmonics (30) for agreementwith FoXS. There were some systematic baseline shiftsbetween the two algorithms at q N 0.3 Å−1 (data notshown), but both algorithms agreed on the location andmagnitude of the difference between the PDB models.Only FoXS results are reported here. FoXS was allowedto fit both the computed profiles to the squid HMMexperimental data using the customary parameters ofhydration layer, excluded volume, and background adjust-ment [15].
Accession numbers
The tarantula (Aphonopelma sp.) myosin ELC [38],RLC [38], and MHC sequences are included in thesupplementary data and have been deposited into theGenBank [39] with accession numbers KT390185,KT390186, and KT619079. Two adjacent HMM tarantulamyosin II IHMs have been deposited in the ResearchCollaboratory for Structural Bioinformatics PDB with theentry code 3JBH.Supplementary data to this article can be found online at
http://dx.doi.org/10.1016/j.jmb.2016.01.027.
Acknowledgments
We thank Dr. Roger Craig for his critical reading,Dr. Guidenn Sulbarán for her comments, Dr.Gustavo Márquez for his help with the manuscript,
and Mr. Jorge Basualdo (ULA) for his initial help inthis project. The images in Figs. 1–4, 6, and 7 andSupplementary Figs. 1–12 were made using UCSFChimera software [69]. Cornell High Energy Syn-chrotron Source is supported by the NationalScience Foundation and National Institutes ofHealth/National Institute of General Medical Sci-ences via National Science Foundation awardDMR-1332208, and the MacCHESS resources aresupported by National Institute of General MedicalSciences award GM-103485 (to R.E.G.). This workwas supported in part by National Institutes of Healthgrant R01GM62968 U.S.A. (to W.W.) and theHoward Hughes Medical Institute, U.S.A. (to R.P.).We dedicate this paper to the memory of Drs. HughE. Huxley and Andrew Szent-Györgyi.
Received 11 September 2015;Received in revised form 15 January 2016;
Accepted 25 January 2016Available online 2 February 2016
Keywords:myosin thick filament;
myosin interacting-heads motif;cryo-electron microscopy;
striated muscle;super-relaxation
‡https://bioinfo.pl/
Abbreviations used:BH, blocked head; C loop, cardiac loop; CM loop,
cardiomyopathy loop; ELC, essential light chain; FH, freehead; HMM, heavy meromyosin; IHM, interacting-heads
motif; MHC, myosin II heavy chain; NTE, N-terminalextension; RLC, regulatory light chain; S2, subfragment2; SAXS, small angle X-ray solution scattering; SRX,
super-relaxed state.
References
[1] R. Craig, R. Padrón, Molecular Structure of the Sarcomere,in: A.G. Engel, C. Franzini-Armstrong (Eds.), Myology, thirded.McGraw-Hill, Inc., New York 2004, pp. 129–166.
[2] M.E. Zoghbi, J.L. Woodhead, R.L. Moss, R. Craig, Three-dimensional structure of vertebrate cardiac muscle myosinfilaments, Proc. Natl. Acad. Sci. U. S. A. 105 (2008) 2386–2390.
[3] H.A. AL-Khayat, R.W. Kensler, J.M. Squire, S.B. Marston,E.P. Morris, Atomic model of the human cardiac musclemyosin filament, Proc. Natl. Acad. Sci. U. S. A. 110 (2013)318–323.
[4] M. González-Solá, H.A. AL-Khayat, M. Behra, R.W. Kensler,Zebrafish cardiacmuscle thick filaments: Isolation technique andthree-dimensional structure, Biophys. J. 106 (2014) 1671–1680.
[5] J.L. Woodhead, F.Q. Zhao, R. Craig, E.H. Egelman, L.Alamo, R. Padrón, Atomic model of a myosin filament in therelaxed state, Nature 436 (2005) 1195–1199.

1163Myosin Interacting-Heads Motifs
[6] F.Q. Zhao, R. Craig, J.L. Woodhead, Head–head interactioncharacterizes the relaxed state of Limulus muscle myosinfilaments, J. Mol. Biol. 385 (2009) 423–431.
[7] A. Pinto, F. Sánchez, L. Alamo, R. Padrón, The myosininteracting-heads motif is present in the relaxed thick filamentof the striated muscle of scorpion, J. Struct. Biol. 180 (2012)469–478.
[8] J.L. Woodhead, F.Q. Zhao, R. Craig, Structural basis of therelaxed state of a Ca2+− regulated myosin filament and itsevolutionary implications, Proc. Natl. Acad. Sci. U. S. A. 110(2013) 8561–8566.
[9] G. Sulbarán, L. Alamo, A. Pinto, G. Márquez, F. Méndez, R.Padrón, R. Craig, Schistosome muscles contain striatedmuscle-like myosin filaments in a smooth muscle-likearchitecture, Biophys. J. 106 (2014) 159a.
[10] R. Craig, J. Megerman, Assembly of smooth muscle myosininto side-polar filaments, J. Cell Biol. 75 (1977) 990–996.
[11] T. Wendt, D. Taylor, K.M. Trybus, K. Taylor, Three-dimensionalimage reconstruction of dephosphorylated smooth muscleheavy meromyosin reveals asymmetry in the interactionbetween myosin heads and placement of subfragment 2,Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 4361–4366.
[12] G. Sulbaran, L. Alamo, A. Pinto, G. Márquez, F. Méndez, R.Padrón, R. Craig, An invertebrate smooth muscle withstriated muscle myosin filaments, Proc. Natl. Acad. Sci. U.S. A. 112 (2015) E5660–E5668.
[13] H.S. Jung, S.A. Burgess, N. Billington, M. Colegrave, H.Patel, J.M. Chalovich, P.D. Chantler, P.J. Knight, Conserva-tion of the regulated structure of folded myosin 2 in speciesseparated by at least 600 million years of independentevolution, Proc. Natl. Acad. Sci. U. S. A. 105 (2008)6022–6026.
[14] Sulbarán, J.Y. Mun, K.H. Lee, L. Alamo, A. Pinto, O. Sato, M.Ikebe, X. Liu, E.D. Korn, R. Padrón, R. Craig, The inhibited,interacting-heads motif characterizes myosin II from theearliest animals with muscle, Biophys. J. 108 (2015) 301a.
[15] R.E. Gillilan, V.S. Kumar, E. O'Neall-Hennessey, C. Cohen,J.H. Brown, X-ray solution scattering of squid heavymeromyosin: Strengthening the evidence for an ancientcompact off state, PLoS ONE 8 (2013), e81994.
[16] A. Houdusse, A.G. Szent-Gyorgyi, C. Cohen, Three confor-mational states of scallop myosin S1, Proc. Natl. Acad. Sci.U. S. A. 97 (2000) 11238–11243.
[17] H.S. Jung, S. Komatsu, M. Ikebe, R. Craig, Head–head andhead–tail interaction: A general mechanism for switchingoffmyosin II activity in cells,Mol. Biol. Cell 19 (2008) 3234–3242.
[18] M.A. Stewart, K. Franks-Skiba, S. Chen, R. Cooke, MyosinATP turnover rate is amechanism involved in thermogenesis inresting skeletal muscle fibers, Proc. Natl. Acad. Sci. U. S. A.107 (2010) 430–435.
[19] P. Hooijman, M.A. Stewart, R. Cooke, A new state of cardiacmyosin with very slow ATP turnover: A potential cardiopro-tective mechanism in the heart, Biophys. J. 100 (2011)1969–1976.
[20] R. Cooke, The role of the myosin ATPase activity in adaptivethermogenesis by skeletal muscle, Biophys. Rev. 3 (2011)33–45.
[21] J.W. McNamara, A. Li, C.G. dos Remedios, R. Cooke, Therole of super-relaxed myosin in skeletal and cardiac muscle,Biophys. Rev. 7 (2015) 5–14.
[22] N. Naber, R. Cooke, E. Pate, Slow myosin ATP turnover in thesuper-relaxed state in tarantula muscle, J. Mol. Biol (2011).
[23] L. Alamo, W. Wriggers, A. Pinto, F. Bártoli, L. Salazar, F.Q.Zhao, R. Craig, R. Padrón, Three-dimensional reconstruction of
tarantula myosin filaments suggests how phosphorylation mayregulate myosin activity, J. Mol. Biol. 384 (2008) 780–797.
[24] R. Brito, L. Alamo, U. Lundberg, J.R. Guerrero, A. Pinto, G.Sulbarán, M.A. Gawinowicz, R. Craig, R. Padrón, Amolecular model of phosphorylation-based activation andpotentiation of tarantula muscle thick filaments, J. Mol. Biol.414 (2011) 44–61.
[25] G. Sulbarán, A. Biasutto, L. Alamo, C. Riggs, A. Pinto, F.Mendéz, R. Craig, R. Padrón, Different head environments intarantula thick filaments support a cooperative activationprocess, Biophys. J. 105 (2013) 2114–2122.
[26] L.M. Espinoza-Fonseca, L. Alamo, A. Pinto, D.D. Thomas, R.Padrón, Sequential myosin phosphorylation activates taran-tula thick filament via a disorder–order transition, Mol.BioSyst. 11 (2015) 2167–2179.
[27] L. Alamo, X.E. Li, L.M. Espinoza-Fonseca, A. Pinto, D.D.Thomas, W. Lehman, R. Padrón, Tarantula myosin free headregulatory light chain phosphorylation stiffens N-terminalextension, releasing it and blocking its docking back, Mol.BioSyst. 11 (2015) 2180–2189.
[28] L. Fusi, Z. Huang, M. Irving, The conformation of myosinheads in relaxed skeletal muscle: Implications for myosin-based regulation, Biophys. J. 109 (2015) 783–792.
[29] J.L. Woodhead, R. Craig, Through thick and thin-interfilamentcommunication in muscle, Biophys. J. 109 (2015) 665–667.
[30] T. Kampourakis, Y.B. Sun, M. Irving, Orientation of the N- and C-terminal lobes of the myosin regulatory light chain in cardiacmuscle, Biophys. J. 108 (2015) 304–314.
[31] T. Kampourakis, M. Irving, Phosphorylation of myosin regulatorylight chain controls myosin head conformation in cardiacmuscle,J. Mol. Cell. Cardiol. 85 (2015) 199–206.
[32] B.A. Baumann, D.W. Taylor, Z. Huang, F. Tama, P.M. Fagnant,K.M. Trybus, K.A. Taylor, Phosphorylated smooth muscle heavymeromyosin shows an open conformation linked to activation, J.Mol. Biol. 415 (2012) 274–287.
[33] K.A. Taylor, M. Feig, C.L. Brooks III, P.M. Fagnant, S. Lowey,K.M. Trybus, Role of the essential light chain in the activation ofsmoothmusclemyosin by regulatory light chain phosphorylation,J. Struct. Biol. 185 (2014) 375–382.
[34] T. Wendt, D. Taylor, T. Messier, K.M. Trybus, K.A. Taylor,Visualization of head–head interactions in the inhibited state ofsmooth muscle myosin, J. Cell Biol. 147 (1999) 1385–1390.
[35] J. Liu, T. Wendt, D. Taylor, K. Taylor, Refined model of the 10Sconformation of smooth muscle myosin by cryo-electronmicroscopy 3D image reconstruction, J. Mol. Biol. 329 (2003)963–972.
[36] C.L. Lawson, M.L. Baker, C. Best, C. Bi, M. Dougherty, P. Feng,G.G. van, B. Devkota, I. Lagerstedt, S.J. Ludtke, R.H. Newman,T.J. Oldfield, I. Rees, G. Sahni, R. Sala, S. Velankar, J. Warren,J.D. Westbrook, K. Henrick, G.J. Kleywegt, H.M. Berman, W.Chiu, EMDataBank.org: Unified data resource for CryoEM,Nucleic Acids Res. 39 (2011) D456–D464.
[37] W. Blankenfeldt, N.H. Thoma, J.S. Wray, M. Gautel, I.Schlichting, Crystal structures of human cardiac beta-myosin IIS2-Delta provide insight into the functional role of theS2 subfragment, Proc. Natl. Acad. Sci. U. S. A. 103 (2006)17713–17717.
[38] J. Zhu, Y. Sun, F.Q. Zhao, J. Yu, R. Craig, S. Hu, Analysis oftarantula skeletal muscle protein sequences and identification oftranscriptional isoforms, BMC Genomics 10 (2009) 117.
[39] D.A. Benson, I. Karsch-Mizrachi, D.J. Lipman, J. Ostell, D.L.Wheeler, GenBank, Nucleic Acids Res. 33 (2005) D34–D38.
[40] UniProt: A hub for protein information, Nucleic Acids Res. 43(2015) D204–D212.

1164 Myosin Interacting-Heads Motifs
[41] T.Q. Uyeda, K.M. Ruppel, J.A. Spudich, Enzymatic activitiescorrelate with chimaeric substitutions at the actin-binding face ofmyosin, Nature 368 (1994) 567–569.
[42] A.S. Rovner, A long, weakly charged actin-binding loop isrequired for phosphorylation-dependent regulation of smoothmuscle myosin, J. Biol. Chem. 273 (1998) 27939–27944.
[43] C.T. Murphy, J.A. Spudich, Variable surface loops and myosinactivity: Accessories to a motor, J. Muscle Res. Cell Motil. 21(2000) 139–151.
[44] H.M. Berman, T.N. Bhat, P.E. Bourne, Z. Feng, G. Gilliland, H.Weissig, J. Westbrook, The Protein Data Bank and thechallenge of structural genomics, Nat. Struct. Biol. 7 (Suppl)(2000) 957–959.
[45] S. Yang, F.Q. Zhao, G. Sulbarán, J.L. Woodhead, L. Alamo, A.Pinto, R. Padrón, R. Craig, Improved imaging, 3D reconstructionand homology modelling of tarantula thick filaments, Biophys. J.(2015) 589a.
[46] M.E. Zoghbi, J.L. Woodhead, R. Craig, R. Padrón, Helical orderin tarantula thick filaments requires the "closed" conformation ofthe myosin head, J. Mol. Biol. 342 (2004) 1223–1236.
[47] S. Xu, G. Offer, J. Gu, H.D. White, L.C. Yu, Temperature andligand dependence of conformation and helical order in myosinfilaments, Biochemistry 42 (2003) 390–401.
[48] I. Rayment, W.R. Rypniewski, K. Schmidt-Base, R. Smith, D.R.Tomchick, M.M. Benning, D.A. Winkelmann, G. Wesenberg,H.M. Holden, Three-dimensional structure of myosin subfrag-ment-1: A molecular motor, Science 261 (1993) 50–58.
[49] E. O'Neall-Hennessey, L. Reshetnikova, V.S. Senthil Kumar, H.Robinson, A.G. Szent-Gyorgyi, C. Cohen, Purification, crystal-lization and preliminary X-ray crystallographic analysis of squidheavy meromyosin, Acta Crystallogr. Sect. F: Struct. Biol. Cryst.Commun. 69 (2013) 248–252.
[50] S. Yang, J.L. Woodhead, F.Q. Zhao, G. Sulbarán, R. Craig, Anapproach to improve the resolution of helical filaments with alarge axial rise and flexible subunits, J. Struct. Biol. (2015).
[51] M.S. Miller, F.N. Soto-Adames, J.M. Braddock, J.M. Wang, J.Robbins, J.O. Vigoreaux, D.W. Maughan, Phylogenic andfunctional analysis of the myosin light chain amino terminalextensions, J. Muscle Res. Cell Motil. 28 (2007) 442.
[52] D.W. Maughan, M.S. Miller, F.N. Soto-Adames, Y. Wang, J.Robbins, J.O. Vigoreaux, Phylogenic and functional analysis ofthe myosin light chain amino terminal extensions, Biophys. J. 94(2008) 306–307.
[53] R.A. Crowther, R. Padrón, R. Craig, Arrangement of the heads ofmyosin in relaxed thick filaments from tarantula muscle, J. Mol.Biol. 184 (1985) 429–439.
[54] J. Liu, D.W. Taylor, E.B. Krementsova, K.M. Trybus, K.A. Taylor,Three-dimensional structure of the myosin V inhibited state bycryoelectron tomography, Nature 442 (2006) 208–211.
[55] M.A. Geeves, R. Fedorov, D.J. Manstein, Molecular mechanismof actomyosin-based motility, Cell. Mol. Life Sci. 62 (2005)1462–1477.
[56] M. Lorenz, K.C. Holmes, The actin-myosin interface, Proc. Natl.Acad. Sci. U. S. A. 107 (2010) 12529–12534.
[57] R. Craig, J.L. Woodhead, Structure and function of myosinfilaments, Curr. Opin. Struct. Biol. 16 (2006) 204–212.
[58] R. Padrón, L. Alamo, Review: The use of negative staining andcryo-electron microscopy to understand the molecular mecha-nism of myosin-linked regulation of striated muscle contraction,Acta Microsc. 13 (2004) 14–29.
[59] S.A. Burgess, S. Yu, M.L. Walker, R.J. Hawkins, J.M. Chalovich,P.J. Knight, Structures of smooth muscle myosin and heavymeromyosin in the folded, shutdown state, J. Mol. Biol. 372(2007) 1165–1178.
[60] M. Linari, E. Brunello, M. Reconditi, L. Fusi, M. Caremani, T.Narayanan, G. Piazzesi, V. Lombardi, M. Irving, Forcegeneration by skeletal muscle is controlled by mechanosensingin myosin filaments, Nature (2015).
[61] K.M. Trybus, Y. Freyzon, L.Z. Faust, H.L. Sweeney, Spare therod, spoil the regulation: Necessity for a myosin rod, Proc. Natl.Acad. Sci. U. S. A. 94 (1997) 48–52.
[62] R. Craig, R. Padrón, J. Kendrick-Jones, Structural changesaccompanying phosphorylation of tarantula muscle myosinfilaments, J. Cell Biol. 105 (1987) 1319–1327.
[63] R. Padrón, N. Panté, H. Sosa, J. Kendrick-Jones, X-raydiffraction study of the structural changes accompanyingphosphorylation of tarantula muscle, J. Muscle Res. Cell Motil.12 (1991) 235–241.
[64] J. Ravaux, A. Hassanin, J. Deutsch, F. Gaill, U. Markmann-Mulisch, Sequence analysis of the myosin regulatory light chaingene of the vestimentiferan Riftia pachyptila, Gene 263 (2001)141–149.
[65] C. Wilson, N. Naber, E. Pate, R. Cooke, The myosin inhibitorblebbistatin stabilizes the super-relaxed state in skeletal muscle,Biophys. J. 107 (2014) 1637–1646.
[66] B.A. Colson, K.J. Petersen, B.C. Collins, D.A. Lowe, D.D.Thomas, The myosin super-relaxed state is disrupted byestradiol deficiency, Biochem. Biophys. Res. Commun. 456(2015) 151–155.
[67] A.F. Straight, A. Cheung, J. Limouze, I. Chen, N.J. Westwood,J.R. Sellers, T.J. Mitchison, Dissecting temporal and spatialcontrol of cytokinesis with a myosin II inhibitor, Science 299(2003) 1743–1747.
[68] F.Q. Zhao, R. Padrón, R. Craig, Blebbistatin stabilizes the helicalorder of myosin filaments by promoting the switch 2 closed state,Biophys. J. 95 (2008) 3322–3329.
[69] E.F. Pettersen, T.D. Goddard, C.C. Huang, G.S. Couch, D.M.Greenblatt, E.C. Meng, T.E. Ferrin, UCSF Chimera—A visual-ization system for exploratory research and analysis, J. Comput.Chem. 25 (2004) 1605–1612.
[70] W. Humphrey, A. Dalke, K. Schulten, VMD: Visual moleculardynamics, J. Mol. Graphs. 14 (1996) 33–38.
[71] L. Kale, R. Skeel, M. Bhandarkar, R. Brunner, A. Gursoy, N.Krawetz, J. Phillips, A. Shinozaki, K.S.K. Varadarajan, NAMD2:Greater scalability for parallel molecular dynamics, J. Comput.Phys. 151 (1999) 283–312.
[72] A.T. Brünger, X-PLOR Version 3.1, A System for X-RayCrystallography and NMR, Yale University Press, 1992.
[73] L. Zhang, J. Hermans, Hydrophilicity of cavities in proteins,Proteins 24 (1996) 433–438.
[74] W. Wriggers, P. Chacon, Modeling tricks and fitting techniquesfor multiresolution structures, Structure 9 (2001) 779–788.
[75] A.M. Waterhouse, J.B. Procter, D.M. Martin, M. Clamp, G.J.Barton, JalView version 2—A multiple sequence alignmenteditor and analysis workbench, Bioinformatics 25 (2009)1189–1191.
[76] M.A. Larkin, G. Blackshields, N.P. Brown, R. Chenna, P.A.McGettigan, H. McWilliam, F. Valentin, I.M. Wallace, A. Wilm, R.Lopez, J.D. Thompson, T.J. Gibson, D.G. Higgins, Clustal Wand clustal X version 2.0. Bioinformatics 23 (2007) 2947–2948.
[77] D. Schneidman-Duhovny, M. Hammel, J.A. Tainer, A. Sali,Accurate SAXS profile computation and its assessmentby contrast variation experiments, Biophys. J. 105 (2013)962–974.
[78] D. Svergun, C. Barberato, M.H.J. Koch, CRYSOL—A programto evaluate X-ray solution scattering of biological macromole-cules from atomic coordinates, J. Appl. Crystallogr. 28 (1995)768–773.