comparison of voltage-activated cl − channels in rat parotid acinar cells with clc2 in a mammalian...
TRANSCRIPT

Comparison of Voltage-activated Cl− Channels in Rat Parotid Acinar Cells with ClC-2 ina Mammalian Expression System
K. Park 1, J. Arreola1,2, T. Begenisich2, J.E. Melvin1
1Rochester Institute for Biomedical Sciences, Center for Oral Biology, University of Rochester Medical Center, Rochester, NY 14642, USA2Department of Pharmacology and Physiology, University of Rochester Medical Center, Rochester, NY 14642, USA
Received: 10 November 1997/Revised: 19 February 1998
Abstract. Rat parotid acinar cells express Cl− currentsthat are activated in a time-dependent manner by hyper-polarized potentials. ClC-2, a member of the ClC genefamily, codes for a voltage-gated, inward rectifying an-ion channel when expressed inXenopusoocytes. In thepresent study, we found that cDNA derived from indi-vidual parotid acinar cells contained sequence identicalto that reported for ClC-2 in rat brain and heart. A poly-clonal antibody generated against the N-terminal cyto-plasmic domain of ClC-2 recognized an approximately100 kD protein on western blots of both brain and parotidgland. ClC-2 expressed in oocytes has different kineticsfrom the currents found in parotid acinar cells. Since theClC-2 channel was cloned from and its transcripts areexpressed in mammalian tissue, we compared the chan-nel properties of acinar cells to a mammalian expressionsystem. We expressed ClC-2 channels in human embry-onic kidney cells, HEK 293, using recombinant ClC-2DNA and ClC-2 DNA fused with DNA coding for jel-lyfish green fluorescent protein (GFP). Confocal mi-croscopy revealed that the expressed ClC-2-GFP chi-mera protein localized to the plasma membrane. Wholecell Cl− currents from HEK 293 cells expressing ClC-2-GFP were similar, if not identical, to the Cl− currentsrecorded from cells transfected with ClC-2 cDNA (noGFP). The voltage-dependence and kinetics of ClC-2channels expressed in HEK 293 cells were quite similarto those in acinar cells. Channels in parotid acinar andHEK 293 cells activated at more positive membrane po-tentials and with a faster time course than the channelsexpressed inXenopusoocytes. In summary, we foundthat ClC-2 message and protein are expressed in salivarycells and that the properties of voltage-activated, inward
rectifying Cl− channels in acinar cells are similar to thosegenerated by the ClC-2-GFP construct expressed in HEK293 cells. The properties of the ClC-2 anion channelseem to be dependent on the type of cell background inwhich it is expressed.
Key words: Salivary acinar cells — HEK 293 — Greenfluorescent protein
Introduction
The ClC gene family codes for several types of voltage-gated chloride channel proteins (Jentsch et al., 1995;Jentsch, 1996; Jentsch & Gu¨nther, 1997). One memberof this family, ClC-2, was first cloned from rat brain andheart and its transcripts are widely expressed in mam-malian tissues and cell lines (Thiemann et al., 1992).The functional properties of this channel have been char-acterized by transient expression inXenopusoocytes(Thiemann et al., 1992; Gru¨nder et al., 1992; Jordt &Jentsch, 1997). When expressed in oocytes, ClC-2 is ac-tivated by hyperpolarization and cell swelling. Hyper-polarization-activated chloride currents have been iden-tified in several cell types such as neurons (Smith et al.,1995), pancreatic acinar cells (Carew & Thorn, 1996),mandibular gland duct cells (Dinudom, Young & Cook,1993; Komwatana et al., 1994), Leydig cells from maturetestis (Noulin & Joffre, 1993), intestinal T84 cells(Fritsch & Edelman, 1996), osteoblasts (Chesnoy-Marchais & Fritsch, 1994), and parotid acinar cells(Arreola et al., 1996).
The properties of these Cl− currents are generallysimilar to the ClC-2 Cl− channels expressed in oocytes.However, several distinct differences are evident, e.g., inanion selectivity and inhibitor sensitivity, making it dif-ficult to positively identify endogenous voltage-activatedCorrespondence to:J.E. Melvin
J. Membrane Biol. 163, 87–95 (1998) The Journal of
MembraneBiology© Springer-Verlag New York Inc. 1998

Cl− currents as ClC-2. In parotid acinar cells the hyper-polarization-activated channels are equally permeable toCl− and I− (Arreola et al., 1996). When expressed inoocytes, rat ClC-2 displays strong selectivity for Cl− overI− (Thiemann et al., 1992) whereas rabbit ClC-2 favors I−
over Cl− (Malinowska et al., 1995). ClC-2 is apparentlyactivated by hypotonic shock and is sensitive to the Cl−
channel blockers DPC and 9-AC (Thiemann et al., 1992;Grunder et al., 1992; Jordt & Jentsch, 1997), while theClC-2-like currents in mouse mandibular gland duct cells(Komwatana et al., 1995) and rat osteoblasts (Chesnoy-Marchais & Fritsch, 1994) are inhibited by cell swellingand are insensitive to these same antagonists. The volt-age-activated Cl− current in rat osteoblastic cells is muchmore sensitive (Chesnoy-Marchais & Fritsch, 1994) toDIDS inhibition than ClC-2 channels expressed in oo-cytes (Thiemann et al., 1992). It is unclear whether thedifferent properties of the Cl− currents in oocytes ex-pressing ClC-2 are the result of altered characteristics ofthese Cl− currents when expressed in a non-mammalianbackground or that the hyperpolarization-activated Cl−
currents in mammalian cells are not due to the ClC-2protein.
The transient expression of cloned ion channels inXenopusoocytes is a common method to study channelproperties. However, Cl− channel expression in theseoocytes is complicated by the presence of endogenouschloride channels (Kowdley et al., 1994). On the otherhand, the identification of transfected cells is difficult inmammalian transient expression systems. One strategyfor easily identifying transfected cells is labeling chan-nels with the green fluorescent protein (GFP) as has beensuccessfully used to study the functional expression ofNMDAR1 receptor-activated anion channels (Marshallet al., 1995). GFP, cloned from the jellyfishAequoreavictoria, is a cytosolic polypeptide composed of 238amino acid residues which emits green fluorescent lightin the absence of exogenous substrates and cofactorswhen expressed inE. coli or mammalian cells (Prasher etal., 1992). It has been shown to be an excellent tool forfollowing protein expression, localization, and traffick-ing without any noticeable functional disruption of theprotein being studied (Chalfie et al., 1994; Marshall etal., 1995).
The purpose of the present study was to compare thekinetic properties of native voltage-activated Cl− currentswith ClC-2 expressed in a well-defined mammalian ex-pression system. In this communication we demonstratethat ClC-2 transcript and protein are expressed in ratparotid glands. We also describe the properties of a chi-mera ClC-2 chloride channel fused to the GFP reportergene. The chimera ClC-2-GFP channels produced afluorescent protein localized to the plasma membranewhen expressed in the human embryonic kidney cell lineHEK 293. The electrophysiological properties of the
ClC-2-GFP channel activated by hyperpolarizing mem-brane potentials were similar to those of parotid acinarcells. In contrast, the voltage sensitivity and activationkinetics of the ClC-2 channels from these mammaliancells were quite different than those of the ClC-2 channelexpressed inXenopusoocytes.
Materials and Methods
PREPARATION OF RAT PAROTID ACINAR CELLS
Parotid glands were removed from male 150–250 gm Wistar strain ratsand dispersed by trypsin-collagenase digestion as described previously(Arreola et al., 1996). Acinar cells were plated onto poly-l-lysinecoated glass coverslips for patch clamp experiments. Individual acinarcells were collected using glass micropipettes for RT-PCR experi-ments.
RT-PCR AMPLIFICATION OF RAT PAROTID
ACINAR CELLS
Total RNA was isolated from 5–10 acinar cells using either the Hirtsupernatant method (1967) or TRIzol reagent (Gibco BRL, Grand Is-land, NY). RNA was reverse-transcribed to cDNA according to themanufacturer’s instructions (1st-STRAND cDNA Synthesis kit, Clon-tech, Palo Alto, CA). The cDNA was amplified using PCR primer setsthat covered the entire protein coding region of the rat ClC-2 gene.The 50 ml PCR reaction mixture contained 5ml of the cDNA-containing solution, 0.2mM of the upper and lower primers, 3ml of 25mM MgCl2, 4 ml of 2.5 mM dNTPs, 5ml of 10× PCR buffer (500 mMKCl, 100 mM Tris-HCl, pH 8.3) and 0.4ml (2 U) of AmpliTaq DNApolymerase (Perkin-Elmer, Norwalk, CT). PCR amplification was car-ried out for 30 cycles in a DNA thermal cycler (MJ Research, Water-town, MA) and each cycle was set to: 94°C for 30 sec, 68°C for 20 sec,72°C for 2 min. A 2-ml aliquot of the first-round PCR reaction mixturewas re-amplified using the same PCR conditions. PCR amplificationproducts were separated on 1% TBE-agarose gels and further analyzedby restriction endonucleases and by Sanger DNA sequencing (Sanger,Nicklen & Coulson, 1977).
Based on published sequence (Thiemann et al., 1992), rat ClC-2specific primers were synthesized as follows:
1. ClC2U1, 58-(-33)AAGCAAGAGGAGGCAAGAGGAC(-12)-382. ClC2U2, 58-(309)GGGGCTCCTCATGGCATTGGTCAGT-
(333)-383. ClC2U3, 58-(885)TCGGGTCTTGGCAGTGTGGAACCGTGA-
T(912)-384. ClC2U4, 58-(2036)TCCGCTTCCAGGTGAACACAGAGGAC-
T(2062)-385. ClC2L1, 58-(1025)GCTCCCCCGAAGCCACTAGCAATAC-
(1001)-386. ClC2L2, 58-(2127)GGGCCCTCTCTTTAGAGCAGGCTTCA-
G(2101)-387. ClC2L3, 58-(2902)TTCTCAAAAGACGAGGCAGGGG(2881)-38
the numbers in parentheses refer to the nucleotide positions in theClC-2 gene assigning the value +1 to the translation initiation site.
CLONING OF THE CODING REGION FOR THEClC-2 GENE
FROM RAT BRAIN
Poly(A+) RNA was prepared from rat brain by oligo(dT)-cellulosechromatography of total RNA isolated by TRIzol reagent. First strand
88 K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells

cDNA was reverse-transcribed from Poly(A+) RNA as above. Oligo-nucleotide primers ClC2U1 and ClC2L3 were used to amplify a 2935bp product containing the entire protein coding sequence for ClC-2.PCR amplifications were carried out for 30 cycles under the above PCRconditions. The amplified ClC-2 cDNA was gel purified and sub-cloned into the pCRII vector (Invitrogen, San Diego, CA) for furtheranalyses. The ClC-2 DNA sequence was confirmed by sequencing.
CONSTRUCTION OFClC-2 cDNA IN pcDNA1
EcoR1 endonuclease (New England Biolabs, Beverly, MA) was used todigest ClC-2 cDNA from pCRII-ClC-2. ClC-2 expression was drivenby the cytomegalovirus mammalian promoter after insertion into thepcDNA1 vector (Invitrogen, San Diego, CA), pcDNA1-ClC-2. Thecorrect orientation for translation of the DNA was confirmed by se-quencing.
SITE-DIRECTED MUTAGENESIS AND CONSTRUCTION OF
ClC-2-GFP cDNAIN THE pMT3 VECTOR
ClC-2 cDNA was mutated using a PCR-based technique (Saiki et al.,1988) to introduce an AgeI restriction endonuclease site and to removethe 38 end stop codon. The synthesized lower primer to produce thesemutations had the following sequence:
58-(2726)GGTCACcGGtaACTTGTCA(2709)-38
This lower primer and the ClC2U1 upper primer (see above) wereemployed to amplify and mutate pCRII-ClC-2 cDNA. The lower caseletters in the mutating primer sequence indicate the base changes. ThePCR products were digested by AgeI and gel purified. The purifiedfragment was subcloned into the pMT3-GFP vector which had beenlinearized by AgeI and dephosphorylated with calf intestine alkalinephosphatase (Boehringer Mannheim Biochemica, Germany). ThepMT3-GFP plasmid contains GFP cDNA inserted in the KpnI and SpeIsites of the pMT3 vector. The pMT3 vector was derived from pMT2by replacing the multi-cloning sites for the PstI and EcoRI cloning sites(Franke et al., 1988). After ligation of the AgeI sticky-ends of thepMT3-GFP vector and the mutated ClC-2 cDNA, the free ends wereblunt-ended using T4 DNA polymerase (New England Biolabs, Bev-erly, MA) and self-ligated. The pMT3-ClC-2-GFP fusion constructwas sequenced to confirm the mutation and orientation.
TRANSIENT EXPRESSION OFGFP, CLC-2-GFP,AND
ClC-2 PROTEINS IN HEK 293 CELLS
The human embryonic cell line HEK 293 (American Type CultureCollection, Rockville, MD) was maintained in Dulbecco’s modifiedEagle’s medium containing 10% newborn calf serum, 2 mM L-glutamine, penicillin (100 units ml−1) and streptomycin (100mg ml−1;Gibco/BRL, Grand Island, NY). One day prior to transfection, cellswere replated (5 × 105 cells per 60 mm plate) and then transfected with10mg of plasmid DNA (pcDNA1-ClC-2, pMT3-ClC-2-GFP, or pMT3-GFP plasmid DNA) using the calcium precipitation method (Chen &Okayama, 1987). Fiftym l of 2.5 M CaCl2 was added to450 ml of plasmid DNA (10mg) in 0.1× TE (1 mM Tris-HCl, 0.1 mM
EDTA, pH 4 8.0). This mixture was added to 500ml of 2× HEPES-buffered saline (280 mM NaCl, 50 mM HEPES, 1.5 mM Na2HPO4, pH4 7.1) and precipitated for 30 min at room temperature. The cellswere incubated with the Ca2+ phosphate-DNA precipitates for 6 hr,followed by a 10% glycerol shock for 3 min and then incubated withfresh medium.
RECOVERY OF pcDNA-1-ClC-2FROM HEK 293 CELLS
Plasmid DNA from HEK 293 cells was rescued using the Hirt extrac-tion method (1967) to verify that the recorded inward rectifying Cl−
currents were due to expression of ClC-2 cDNA lacking the GFP label.The cells were collected with a glass micropipette and lysed in 0.8 mlof 0.6% SDS containing 0.01M EDTA (pH 4 7.5) for 20 min at roomtemperature. Two hundredml of 5 M NaCl was added to the lysate,mixed gently by inverting the tube 10 times, and stored at 4°C for atleast 8 hr. After centrifugation at 17,000 ×g for 30 min at 4°C, thesupernatant was removed and precipitated with 100% ethanol. Therecovered plasmid DNA was transformed into theE. coli strain JM109using electroporation (1 × 109 cfu permg plasmid DNA) and analyzedby restriction endonuclease digestion with KpnI.
IDENTIFICATION OF EXPRESSED ANDNATIVE ClC-2USING WESTERN BLOT ANALYSIS
A rabbit polyclonal antibody was prepared to the N-terminal region ofrat ClC-2 corresponding to amino acids Met 16 to Glu 35 (oligopeptideMEPRALQYEQTLMYGRYTQE). The anti-ClC-2 antibody was pu-rified using an antigen (the synthesized oligopeptide)-linked affinitycolumn.
Rat brain homogenates and rat parotid crude membranes wereused to identify native ClC-2 protein. Tissues were prepared as previ-ously described by Turner, George & Baum (1986). Rat brains andparotid glands were homogenized twice by 10-sec strokes at powerlevel 5 with a Polytron homogenizer (Brinkmann Instruments, West-bury, NY) in 10 ml per gram tissue of a solution containing: 10 mM
HEPES adjusted to pH 7.4 with Tris, 10% sucrose, 1 mM EDTA and 1mM PMSF. Homogenates were centrifuged at 2,500 ×g for 15 min at4°C and the supernatants were saved. Rat parotid glands were furtherpurified to prepare crude membrane proteins by centrifugation of thesupernatant at 22,000 ×g for 20 min at 4°C. The supernatant wasdiscarded and the pellet was resuspended in buffer (10 mM HEPESadjusted to pH 7.4 with Tris, 100 mM mannitol, 1 mM EDTA and 1 mM
PMSF), passed once through a 25-gauge needle and once through a30-gauge needle. Aliquots were quickly frozen in liquid N2 and storedat −85°C until use.
Cell extracts from transfected HEK 293 cells were used to iden-tify expressed ClC-2 protein. Extracts were prepared 48 hr after trans-fection. HEK 293 cells were washed twice with PBS and centrifugedat 700 × g for 10 min. Proteins were extracted in 10% SDS withboiling for 5 min followed by addition of an equal volume of lysisbuffer (250 mM sucrose, 5 mM MOPS, 1 mM EDTA, pH 4 7.5, and 1mM PMSF) and stored at −85°C until use.
Protein concentration was determined using the BCA (bicin-chonic acid) protein assay reagent (Pierce, Rockford, IL). Proteins (70mg/lane) were resolved by 10% SDS-PAGE and transferred to a nitro-cellulose membrane (hybond C, Amersham, Arlington Heights, IL).Membranes were blocked in PBS containing 0.1% tween 20, 10%normal goat serum and 4% BSA and probed with the affinity columnpurified primary antibody (anti-ClC-2) or pre-immune serum. Proteinswere then detected with a horseradish peroxidase-conjugated goat anti-rabbit IgG antibody (Jackson ImmunoResearch Lab., West Grove, PA)by ECL chemiluminescence (Amersham, Buckinghamshire, England).
IDENTIFICATION OF EXPRESSEDGFPOR ClC-2-GFPIN
HEK 293 CELLS BY CONFOCAL MICROSCOPY
The fluorescence from cells expressing either the ClC-2-GFP fusion orGFP gene product was viewed on an Ultima confocal laser cytometer(Meridian Instr., Okemos, MI) using an Olympus 40× UV objective
89K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells
(NA 0.85). GFP was excited with the 488 nm wavelength of an En-terprise argon laser (Coherent, CA) and the emitted fluorescence wasmonitored at >500 nm. The estimated thickness of the image was 5mm. Forty-eight hours after transfection, the Cl− currents of fluores-cent cells were recorded with the whole-cell patch clamp technique.
WHOLE-CELL PATCH CLAMP
The whole cell configuration of the patch clamp technique was used torecord macroscopic Cl− currents from rat parotid acinar cells and fromHEK 293 cells 48 hr after transfection. Cl− currents were recordedwith a PC-501A amplifier (Warner Instrument). Pipettes had 1–2MV
resistance when filled with the internal solution. Cation currents wereminimized using the non-permeant cation TEA inside and outside thecell. To facilitate comparison with previous observations of voltage-activated Cl− channels in native parotid acinar cells (Arreola et al.,1996) we used the same intracellular solution which was composed of(in mM): 135 TEACl, 5 TEAF, 15 EGTA-TEA, and 20 TES pH 7.3.Cells were bathed in a pH 7.3 hypertonic solution containing (in mM):135 TEACl, 15 EGTA-TEA, 20 TES, and 60 D-mannitol. This hyper-tonic bath solution inactivates the endogenous volume-sensitive Cl−
current present in parotid acinar cells and HEK 293 cells.Whole cell current recordings began 5–8 min after rupturing the
plasma membrane beneath the pipette tip except for experiments de-signed to monitor the time dependent increase in the magnitude of thevoltage-gated, inward rectifying Cl− currents. The holding potentialwas 0 mV. To activate ClC-2 channels the membrane potential wasstepped every 10 sec to values from −140 to +80 mV in 20 mV incre-ments of 500–700 msec duration each. In most experiments the voltagewas repolarized to −50 mV for an additional 300–500 msec beforereturning to the holding potential. Capacitative transients were partlycompensated by the amplifier circuitry. Macroscopic currents werefiltered at 500 Hz and sampled at 250 or 667 Hz.
Results
VOLTAGE-GATED, INWARD RECTIFYING Cl− CURRENTS IN
RAT PAROTID ACINAR CELLS
Rat parotid acinar cells express at least three types ofanion channels (Arreola et al., 1996). One of these ex-
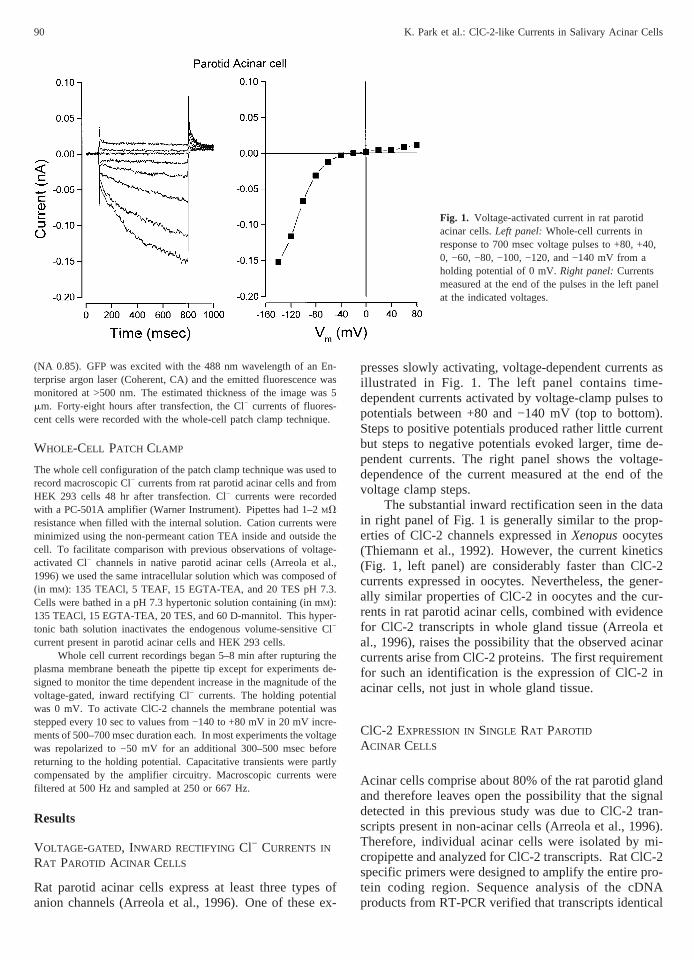
presses slowly activating, voltage-dependent currents asillustrated in Fig. 1. The left panel contains time-dependent currents activated by voltage-clamp pulses topotentials between +80 and −140 mV (top to bottom).Steps to positive potentials produced rather little currentbut steps to negative potentials evoked larger, time de-pendent currents. The right panel shows the voltage-dependence of the current measured at the end of thevoltage clamp steps.
The substantial inward rectification seen in the datain right panel of Fig. 1 is generally similar to the prop-erties of ClC-2 channels expressed inXenopusoocytes(Thiemann et al., 1992). However, the current kinetics(Fig. 1, left panel) are considerably faster than ClC-2currents expressed in oocytes. Nevertheless, the gener-ally similar properties of ClC-2 in oocytes and the cur-rents in rat parotid acinar cells, combined with evidencefor ClC-2 transcripts in whole gland tissue (Arreola etal., 1996), raises the possibility that the observed acinarcurrents arise from ClC-2 proteins. The first requirementfor such an identification is the expression of ClC-2 inacinar cells, not just in whole gland tissue.
ClC-2 EXPRESSION INSINGLE RAT PAROTID
ACINAR CELLS
Acinar cells comprise about 80% of the rat parotid glandand therefore leaves open the possibility that the signaldetected in this previous study was due to ClC-2 tran-scripts present in non-acinar cells (Arreola et al., 1996).Therefore, individual acinar cells were isolated by mi-cropipette and analyzed for ClC-2 transcripts. Rat ClC-2specific primers were designed to amplify the entire pro-tein coding region. Sequence analysis of the cDNAproducts from RT-PCR verified that transcripts identical
Fig. 1. Voltage-activated current in rat parotidacinar cells.Left panel:Whole-cell currents inresponse to 700 msec voltage pulses to +80, +40,0, −60, −80, −100, −120, and −140 mV from aholding potential of 0 mV.Right panel:Currentsmeasured at the end of the pulses in the left panelat the indicated voltages.
90 K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells

to rat brain ClC-2 were present in rat parotid acinar cells(Fig. 2).
An affinity-purified polyclonal antibody was used toconfirm that ClC-2 protein was expressed in rat parotidgland. PanelB of Fig. 3 shows the results testing thespecificity of the antibody generated against the N-terminal domain of ClC-2. Proteins isolated from HEK293 cells (Lane 1) showed no reactivity with the anti-body, whereas the antibody recognized a distinct band inprotein extracts from HEK 293 cells transfected withboth a ClC-2 containing plasmid (Lane 2) and a ClC-2-GFP containing plasmid (Lane 3). ClC-2-GFP migratedwith a molecular weight of about 125 kD, the expectedsize of the fusion protein, whereas ClC-2 expressed inHEK 293 cells migrated somewhat faster than antici-pated with a molecular weight of 85–90 kD. PanelA ofFig. 3 shows that pre-immune serum (PI) did not reactwith proteins isolated from rat parotid gland (P) or brain(B), the tissue from which ClC-2 was cloned. In con-trast, the affinity-purified antibody (Ab) recognized aprotein band in both parotid gland and brain of about 100kD, the size protein predicted from the ClC-2 cDNAsequence analysis. Additional smaller molecular weightbands are also present in brain tissue homogenates.Modifying the protease inhibitor cocktail did not changethe relative abundance of the various protein bands inbrain homogenates suggesting that proteolysis during theprotein isolation procedure was not responsible for the
multiple bands on the Western blot. These smaller mo-lecular weight bands may represent some post-translation event or cross-reactivity with a related proteinin brain tissue.
The data of Figs. 2 and 3 demonstrate that ClC-2was expressed in rat parotid acinar cells—a necessaryrequirement to identifying the inwardly rectifying cur-rent with this channel protein. However, the differencesin kinetics between ClC-2 expressed in oocytes and thekinetics of the inwardly rectifying current in acinar cellsmake this identification less certain. To examine if theClC-2 kinetics depended on the expression system used,we studied ClC-2 properties as expressed in mammalianHEK 293 cells.
IDENTIFICATION OF TRANSFECTEDCELLS BY
GFP FLUORESCENCE
The identification of transfected cells was facilitated byfusing GFP cDNA to the carboxyl terminus of ClC-2cDNA. The carboxyl terminus was chosen because mu-tation analysis of ClC-2 expressed inXenopusoocytesshowed that this domain is not involved in channel gat-ing, whereas the amino terminus is necessary for channelactivation by volume and voltage (Gru¨nder et al., 1992;Jordt & Jentsch, 1997). GFP fluorescence was viewedby confocal microscopy 48 hr after transfection. Cellstransfected with GFP cDNA produced a homogenousfluorescence throughout the cytosol (Fig. 4, upper panel).In contrast, the expressed chimera ClC-2-GFP produced
Fig. 2. RT-PCR of ClC-2 transcripts in rat parotid acinar cells. Indi-vidual acinar cells and acini were isolated by micropipette. The positionof the upper and lower rat ClC-2 specific primers used to amplify theentire protein coding region are shown in the upper panel. In the lowerpanel, the primer pairs used and the expected size products (given inparentheses) are shown above the amplified products from brain (B)and parotid acinar cells (P). Molecular weight markers (M) are shown(1 Kb ladder, GibcoBRL). The image was digitized with an IS-1000imaging system (Alpha Innotech, San Leandro, CA). Sequence analysisof the products from RT-PCR of RNA isolated from these cells verifiedthat transcripts identical to rat brain ClC-2 were present in rat parotidacinar cells.
Fig. 3. Western analysis of ClC-2 protein in rat parotid gland andbrain. An affinity-purified polyclonal antibody was used to confirm thatClC-2 protein is present in rat parotid gland.Panel A: pre-immuneserum (PI) did not react with proteins isolated from rat parotid gland(P) or brain (B). The affinity-purified antibody (Ab) recognized a pro-tein band in both parotid gland and brain of about 100 kD, the sizeprotein predicted from ClC-2 cDNA sequence analysis.Panel B:Pro-teins isolated from HEK 293 cells (Lane 1) showed no reactivity withthe antibody, whereas the antibody recognized distinct bands in proteinextracts from HEK 293 cells transfected with both a ClC-2 containingplasmid (Lane 2) and a ClC-2-GFP containing plasmid (Lane 3). Theimage was digitized with an IS-1000 imaging system (Alpha Innotech,San Leandro, CA).
91K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells
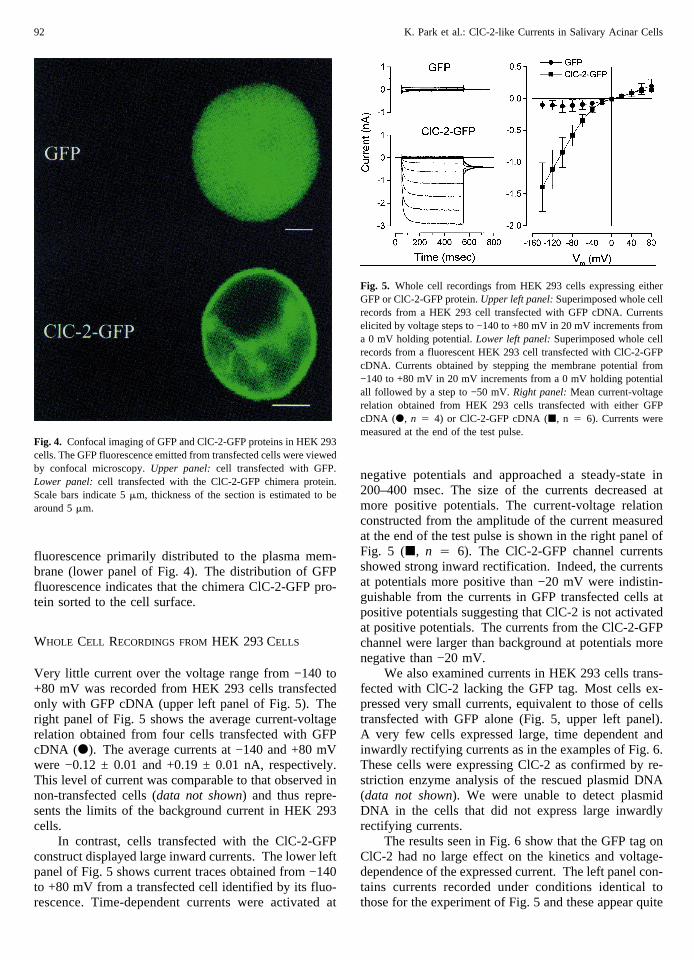
fluorescence primarily distributed to the plasma mem-brane (lower panel of Fig. 4). The distribution of GFPfluorescence indicates that the chimera ClC-2-GFP pro-tein sorted to the cell surface.
WHOLE CELL RECORDINGS FROMHEK 293 CELLS
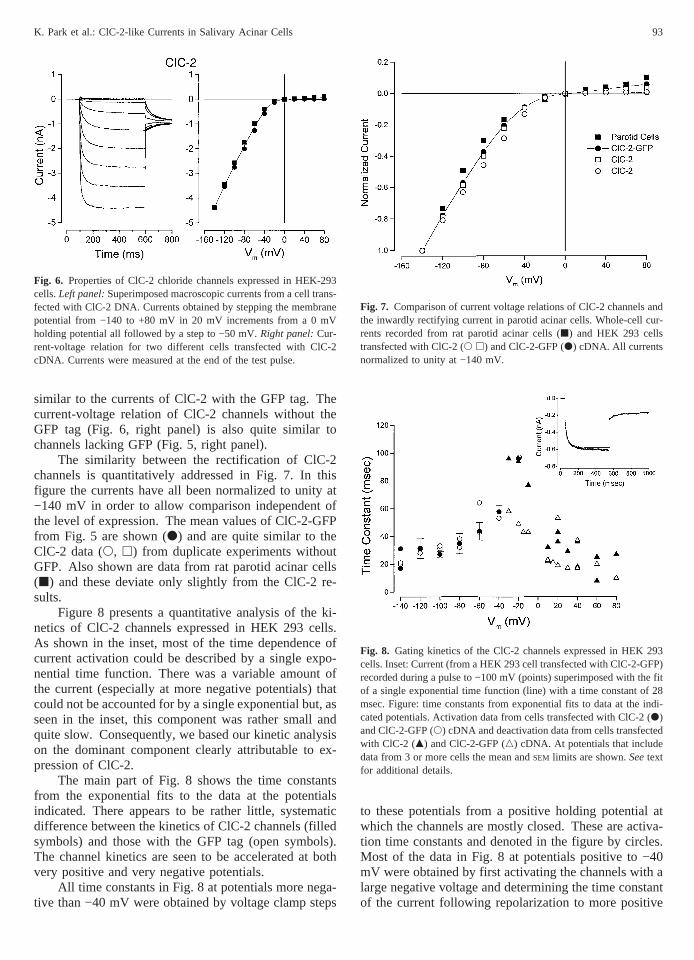
Very little current over the voltage range from −140 to+80 mV was recorded from HEK 293 cells transfectedonly with GFP cDNA (upper left panel of Fig. 5). Theright panel of Fig. 5 shows the average current-voltagerelation obtained from four cells transfected with GFPcDNA (d). The average currents at −140 and +80 mVwere −0.12 ± 0.01 and +0.19 ± 0.01 nA, respectively.This level of current was comparable to that observed innon-transfected cells (data not shown) and thus repre-sents the limits of the background current in HEK 293cells.
In contrast, cells transfected with the ClC-2-GFPconstruct displayed large inward currents. The lower leftpanel of Fig. 5 shows current traces obtained from −140to +80 mV from a transfected cell identified by its fluo-rescence. Time-dependent currents were activated at
negative potentials and approached a steady-state in200–400 msec. The size of the currents decreased atmore positive potentials. The current-voltage relationconstructed from the amplitude of the current measuredat the end of the test pulse is shown in the right panel ofFig. 5 (j, n 4 6). The ClC-2-GFP channel currentsshowed strong inward rectification. Indeed, the currentsat potentials more positive than −20 mV were indistin-guishable from the currents in GFP transfected cells atpositive potentials suggesting that ClC-2 is not activatedat positive potentials. The currents from the ClC-2-GFPchannel were larger than background at potentials morenegative than −20 mV.
We also examined currents in HEK 293 cells trans-fected with ClC-2 lacking the GFP tag. Most cells ex-pressed very small currents, equivalent to those of cellstransfected with GFP alone (Fig. 5, upper left panel).A very few cells expressed large, time dependent andinwardly rectifying currents as in the examples of Fig. 6.These cells were expressing ClC-2 as confirmed by re-striction enzyme analysis of the rescued plasmid DNA(data not shown). We were unable to detect plasmidDNA in the cells that did not express large inwardlyrectifying currents.
The results seen in Fig. 6 show that the GFP tag onClC-2 had no large effect on the kinetics and voltage-dependence of the expressed current. The left panel con-tains currents recorded under conditions identical tothose for the experiment of Fig. 5 and these appear quite
Fig. 4. Confocal imaging of GFP and ClC-2-GFP proteins in HEK 293cells. The GFP fluorescence emitted from transfected cells were viewedby confocal microscopy.Upper panel: cell transfected with GFP.Lower panel:cell transfected with the ClC-2-GFP chimera protein.Scale bars indicate 5mm, thickness of the section is estimated to bearound 5mm.
Fig. 5. Whole cell recordings from HEK 293 cells expressing eitherGFP or ClC-2-GFP protein.Upper left panel:Superimposed whole cellrecords from a HEK 293 cell transfected with GFP cDNA. Currentselicited by voltage steps to −140 to +80 mV in 20 mV increments froma 0 mV holding potential.Lower left panel:Superimposed whole cellrecords from a fluorescent HEK 293 cell transfected with ClC-2-GFPcDNA. Currents obtained by stepping the membrane potential from−140 to +80 mV in 20 mV increments from a 0 mVholding potentialall followed by a step to −50 mV.Right panel:Mean current-voltagerelation obtained from HEK 293 cells transfected with either GFPcDNA (d, n 4 4) or ClC-2-GFP cDNA (j, n 4 6). Currents weremeasured at the end of the test pulse.
92 K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells
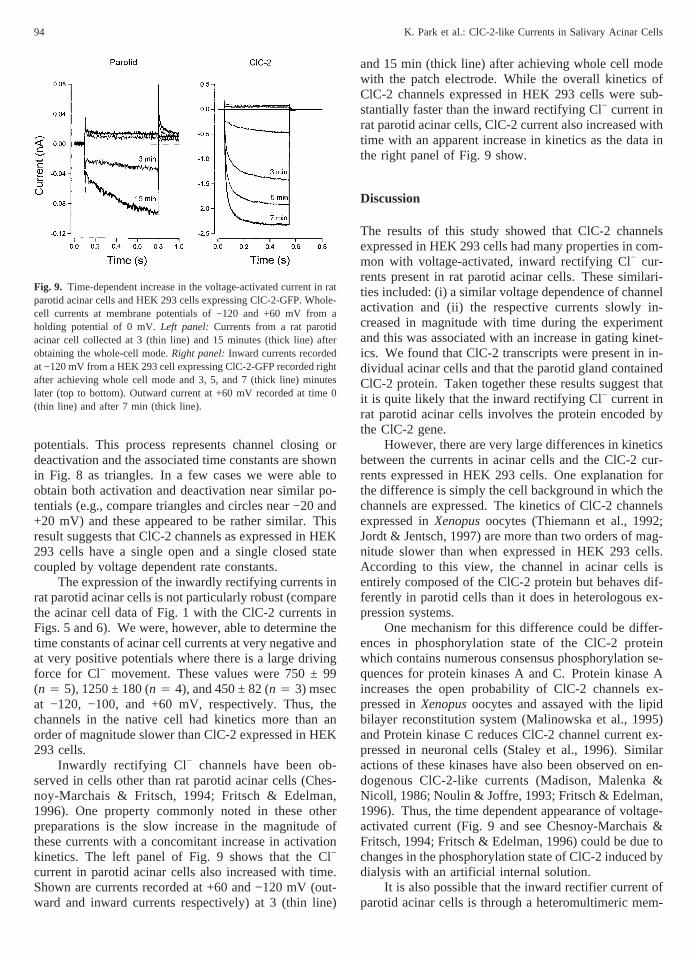
similar to the currents of ClC-2 with the GFP tag. Thecurrent-voltage relation of ClC-2 channels without theGFP tag (Fig. 6, right panel) is also quite similar tochannels lacking GFP (Fig. 5, right panel).
The similarity between the rectification of ClC-2channels is quantitatively addressed in Fig. 7. In thisfigure the currents have all been normalized to unity at−140 mV in order to allow comparison independent ofthe level of expression. The mean values of ClC-2-GFPfrom Fig. 5 are shown (d) and are quite similar to theClC-2 data (s, h) from duplicate experiments withoutGFP. Also shown are data from rat parotid acinar cells(j) and these deviate only slightly from the ClC-2 re-sults.
Figure 8 presents a quantitative analysis of the ki-netics of ClC-2 channels expressed in HEK 293 cells.As shown in the inset, most of the time dependence ofcurrent activation could be described by a single expo-nential time function. There was a variable amount ofthe current (especially at more negative potentials) thatcould not be accounted for by a single exponential but, asseen in the inset, this component was rather small andquite slow. Consequently, we based our kinetic analysison the dominant component clearly attributable to ex-pression of ClC-2.
The main part of Fig. 8 shows the time constantsfrom the exponential fits to the data at the potentialsindicated. There appears to be rather little, systematicdifference between the kinetics of ClC-2 channels (filledsymbols) and those with the GFP tag (open symbols).The channel kinetics are seen to be accelerated at bothvery positive and very negative potentials.
All time constants in Fig. 8 at potentials more nega-tive than −40 mV were obtained by voltage clamp steps
to these potentials from a positive holding potential atwhich the channels are mostly closed. These are activa-tion time constants and denoted in the figure by circles.Most of the data in Fig. 8 at potentials positive to −40mV were obtained by first activating the channels with alarge negative voltage and determining the time constantof the current following repolarization to more positive
Fig. 6. Properties of ClC-2 chloride channels expressed in HEK-293cells.Left panel:Superimposed macroscopic currents from a cell trans-fected with ClC-2 DNA. Currents obtained by stepping the membranepotential from −140 to +80 mV in 20 mV increments from a 0 mVholding potential all followed by a step to −50 mV.Right panel:Cur-rent-voltage relation for two different cells transfected with ClC-2cDNA. Currents were measured at the end of the test pulse.
Fig. 7. Comparison of current voltage relations of ClC-2 channels andthe inwardly rectifying current in parotid acinar cells. Whole-cell cur-rents recorded from rat parotid acinar cells (j) and HEK 293 cellstransfected with ClC-2 (s h) and ClC-2-GFP (d) cDNA. All currentsnormalized to unity at −140 mV.
Fig. 8. Gating kinetics of the ClC-2 channels expressed in HEK 293cells. Inset: Current (from a HEK 293 cell transfected with ClC-2-GFP)recorded during a pulse to −100 mV (points) superimposed with the fitof a single exponential time function (line) with a time constant of 28msec. Figure: time constants from exponential fits to data at the indi-cated potentials. Activation data from cells transfected with ClC-2 (d)and ClC-2-GFP (s) cDNA and deactivation data from cells transfectedwith ClC-2 (m) and ClC-2-GFP (n) cDNA. At potentials that includedata from 3 or more cells the mean andSEM limits are shown.Seetextfor additional details.
93K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells
potentials. This process represents channel closing ordeactivation and the associated time constants are shownin Fig. 8 as triangles. In a few cases we were able toobtain both activation and deactivation near similar po-tentials (e.g., compare triangles and circles near −20 and+20 mV) and these appeared to be rather similar. Thisresult suggests that ClC-2 channels as expressed in HEK293 cells have a single open and a single closed statecoupled by voltage dependent rate constants.
The expression of the inwardly rectifying currents inrat parotid acinar cells is not particularly robust (comparethe acinar cell data of Fig. 1 with the ClC-2 currents inFigs. 5 and 6). We were, however, able to determine thetime constants of acinar cell currents at very negative andat very positive potentials where there is a large drivingforce for Cl− movement. These values were 750 ± 99(n 4 5), 1250 ± 180 (n 4 4), and 450 ± 82 (n 4 3) msecat −120, −100, and +60 mV, respectively. Thus, thechannels in the native cell had kinetics more than anorder of magnitude slower than ClC-2 expressed in HEK293 cells.
Inwardly rectifying Cl− channels have been ob-served in cells other than rat parotid acinar cells (Ches-noy-Marchais & Fritsch, 1994; Fritsch & Edelman,1996). One property commonly noted in these otherpreparations is the slow increase in the magnitude ofthese currents with a concomitant increase in activationkinetics. The left panel of Fig. 9 shows that the Cl−
current in parotid acinar cells also increased with time.Shown are currents recorded at +60 and −120 mV (out-ward and inward currents respectively) at 3 (thin line)
and 15 min (thick line) after achieving whole cell modewith the patch electrode. While the overall kinetics ofClC-2 channels expressed in HEK 293 cells were sub-stantially faster than the inward rectifying Cl− current inrat parotid acinar cells, ClC-2 current also increased withtime with an apparent increase in kinetics as the data inthe right panel of Fig. 9 show.
Discussion
The results of this study showed that ClC-2 channelsexpressed in HEK 293 cells had many properties in com-mon with voltage-activated, inward rectifying Cl− cur-rents present in rat parotid acinar cells. These similari-ties included: (i) a similar voltage dependence of channelactivation and (ii) the respective currents slowly in-creased in magnitude with time during the experimentand this was associated with an increase in gating kinet-ics. We found that ClC-2 transcripts were present in in-dividual acinar cells and that the parotid gland containedClC-2 protein. Taken together these results suggest thatit is quite likely that the inward rectifying Cl− current inrat parotid acinar cells involves the protein encoded bythe ClC-2 gene.
However, there are very large differences in kineticsbetween the currents in acinar cells and the ClC-2 cur-rents expressed in HEK 293 cells. One explanation forthe difference is simply the cell background in which thechannels are expressed. The kinetics of ClC-2 channelsexpressed inXenopusoocytes (Thiemann et al., 1992;Jordt & Jentsch, 1997) are more than two orders of mag-nitude slower than when expressed in HEK 293 cells.According to this view, the channel in acinar cells isentirely composed of the ClC-2 protein but behaves dif-ferently in parotid cells than it does in heterologous ex-pression systems.
One mechanism for this difference could be differ-ences in phosphorylation state of the ClC-2 proteinwhich contains numerous consensus phosphorylation se-quences for protein kinases A and C. Protein kinase Aincreases the open probability of ClC-2 channels ex-pressed inXenopusoocytes and assayed with the lipidbilayer reconstitution system (Malinowska et al., 1995)and Protein kinase C reduces ClC-2 channel current ex-pressed in neuronal cells (Staley et al., 1996). Similaractions of these kinases have also been observed on en-dogenous ClC-2-like currents (Madison, Malenka &Nicoll, 1986; Noulin & Joffre, 1993; Fritsch & Edelman,1996). Thus, the time dependent appearance of voltage-activated current (Fig. 9 and see Chesnoy-Marchais &Fritsch, 1994; Fritsch & Edelman, 1996) could be due tochanges in the phosphorylation state of ClC-2 induced bydialysis with an artificial internal solution.
It is also possible that the inward rectifier current ofparotid acinar cells is through a heteromultimeric mem-
Fig. 9. Time-dependent increase in the voltage-activated current in ratparotid acinar cells and HEK 293 cells expressing ClC-2-GFP. Whole-cell currents at membrane potentials of −120 and +60 mV from aholding potential of 0 mV.Left panel: Currents from a rat parotidacinar cell collected at 3 (thin line) and 15 minutes (thick line) afterobtaining the whole-cell mode.Right panel:Inward currents recordedat −120 mV from a HEK 293 cell expressing ClC-2-GFP recorded rightafter achieving whole cell mode and 3, 5, and 7 (thick line) minuteslater (top to bottom). Outward current at +60 mV recorded at time 0(thin line) and after 7 min (thick line).
94 K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells

brane protein composed of ClC-2 and other subunits thatalter channel gating kinetics. Homomultimeric structureappears to be a general property of the ClC gene family(Steinmeyer et al., 1994; Ludewig, Pusch & Jentsch,1996; Middleton, Phessant & Miller, 1996), however it isnot known if these channels associate with as yet un-identified regulatory subunits.
The pMT3-GFP vector was a gift from Dr. P. Stampe, Department ofPharmacology and Physiology, University of Rochester. We thank Dr.A.J. Persechini for a critical appraisal of this manuscript. This workwas supported in part by National Institutes of Health grant R01DE09692 (J.E.M.). Current address for J.A. is Instituto de Fisica, Uni-versidad Autonoma de San Luis Potosi, San Luis Potosi, Mexico.
References
Arreola, J., Park, K., Melvin, J.E., Begenisich, T. 1996. Three distinctchloride channels control anion movements in rat parotid acinarcells.J. Physiol.490:351–362
Carew, M.A., Thorn, P. 1996. Identification of ClC-2-like chloridecurrents in pig pancreatic acinar cells.Pfluegers Arch433:84–90
Chalfie, M., Tu, Y., Euskirchen, G., Ward, W.W., Prasher, D.C. 1994.Green fluorescent protein as a marker for gene expression.Science263:801–805
Chen, C., Okayama, H. 1987. High-efficiency transformation of mam-malian cells by plasmid DNA.Mol. Cell. Biol. 7:2745–2752
Chesnoy-Marchais, D., Fritsch, J. 1994. Activation of hyperpolariza-tion and atypical osmosensitivity of a Cl− current in rat osteoblasticcells.J. Membrane Biol.140:173–188
Dinudom, A., Young, J.A., Cook, D.I. 1993. Na+ and Cl− conductancesare controlled by cytosolic Cl− concentration in the intralobularduct cells of mouse mandibular glands.J. Membrane Biol.135:289–295
Franke, R.R., Sakmar, T.P., Oprian, D.D., Gobind Khorana, H. 1988. Asingle amino acid substitution in rhodopsin (lysine 248 leucine)prevents activation of transducin.J. Biol. Chem.263:2119–2122
Fritsch, J., Edelman, A. 1996. Modulation of the hyperpolarization-activated Cl− current in human intestinal T84 epithelial cells byphosphorylation.J. Physiol.490:115–128
Grunder, S., Thiemann, A., Pusch, M., Jentsch, T.J. 1992. Regionsinvolved in the opening of ClC-2 chloride channel by voltage andcell volume.Nature360:759–762
Hirt, B. 1967. Selective extraction of polyoma DNA from infectedmouse cell cultures.J. Mol. Biol. 26:365–369
Jentsch, T.J. 1996. Chloride channels: a molecular perspective.Curr.Opinion Neurobiol.6:303–310
Jentsch, T.J., Gu¨nther, W. 1997. Chloride channels: an emerging mo-lecular picture.BioEssays19:117–126
Jentsch, T.J., Gu¨nther, W., Pusch, M., Schwappach, B. 1995. Propertiesof voltage-gated chloride channels of the ClC gene family.J. Phys-iol. 482:19S–25S
Jordt, S.-E., Jentsch, T.J. 1997. Molecular dissection of gating in theClC-2 chloride channel.EMBO J.16:1582–1592
Komwatana, P., Dinudom, A., Young, J.A., Cook, D.I. 1994. Charac-terization of the Cl− conductance in the granular duct cells of mousemandibular glands.Pfluegers Arch.428:641–647
Komwatana, P., Dinudom, A., Young, J.A., Cook, D.I. 1995. Osmoticsensitivity of the hyperpolarization-activated Cl− current in mousemandibular duct cells.Cell Physiol. Biochem.5:243–251
Kowdley, G.C., Ackerman, S.J., John, J.E. III, Jones, L.R., Moorman,J.R. 1994. Hyperpolarization-activated chloride currents inXeno-pusoocytes.J. Gen. Physiol.103:217–230
Ludewig, U., Pusch, M., Jentsch, T.J. 1966. Two physically distinctpores in the dimeric ClC-0 chloride channel.Nature383:340–343
Madison, D.V., Malenka, R.C., Nicoll, R.A. 1986. Phorbol esters blocka voltage-sensitive chloride current in hippocampal pyramidal cells.Nature321:695–697
Malinowska, D.H., Kupert, E.Y., Bahinski, A., Sherry, A.M., Cupo-letti, J. 1995. Cloning, Functional expression, and characterizationof a PKA-activated gastric Cl− channel.Am. J. Physiol.268:C191–C200
Marshall, J., Molloy, R., Moss, G.W., Howe, J.R., Hughes, T.E. 1995.The jellyfish green fluorescent protein: A new tool for studying ionchannel expression and function.Neuron14:211–215
Middleton, R.E., Phessant, D.J., Miller, C. 1996. Homodimeric archi-tecture of a ClC-type Cl− channel.Nature383:337–340
Noulin, J.F., Joffre, M. 1993. Characterization and cyclic AMP-dependence of a hyperpolarization-activated chloride conductancein Leydig cells from mature rat testis.J. Membrane Biol.133:1–15
Prasher, D.C., Eckenrode, V.K., Ward, W.W., Prendergast, F.G., andCormier, M.J. 1992. Primary structure of the Aequorea victoriagreen-fluorescent protein.Gene111:229–233
Saiki, R.K., Gelfand, D.H., Stoffel, S., Scharf, S.J., Higuchi, R., Horn,G.T., Mullis, K.B., Erlich, H.A. 1988. Primer-directed enzymaticamplification of DNA with a thermostable DNA polymerase.Sci-ence239:487–491
Sanger, F., Nicklen, S., Coulson, A.R. 1977. DNA sequencing withchain-terminating inhibitors.Proc. Natl. Acad. Sci. USA74:5463–5467
Smith, R.L., Clayton, G.H., Wilcox, C.L., Escudero, K.W., Staley, K.J.1995. Differential expression of an inwardly rectifying chlorideconductance in rat brain neurons: a potential mechanism for cell-specific modulation of postsynaptic inhibition.J. Neurosci.15:4057–4067
Staley, K., Smith, R., Schaack, J., Wilcox, C., Jentsch, T.J. 1996.Alteration of GABAA receptor function following gene transfer ofthe ClC-2 chloride channel.Neuron17:543–551
Steinmeyer, K., Lorenz, C., Pusch, M., Koch, M.C., Jentsch, T.J. 1994.Multimeric structure of ClC-1 chloride channels as revealed bymutations in dominant myotonia congenita (Thomsen).EMBO J.13:7373–743
Thiemann, A., Gru¨nder, S., Pusch, M., Jentsch, T.J. 1992. A chloridechannel widely expressed in epithelial and non-epithelial cells.Na-ture 356:57–60
Turner, R.J., George, J., Baum, B.J. 1986. Evidence for a Na+/K+/Cl−
cotransport system in basolateral membrane vesicles from rabbitparotid.J. Membrane Biol.94:143–152
95K. Park et al.: ClC-2-like Currents in Salivary Acinar Cells