comparative analyses among the trichomonas vaginalis , trichomonas tenax , and tritrichomonas foetus...
TRANSCRIPT

Curr Genet (2009) 55:199–210
DOI 10.1007/s00294-009-0237-yRESEARCH ARTICLE
Comparative analyses among the Trichomonas vaginalis, Trichomonas tenax, and Tritrichomonas foetus 5S ribosomal RNA genes
Ana Lilia Torres-Machorro · Roberto Hernández · John F. Alderete · Imelda López-Villaseñor
Received: 4 December 2008 / Revised: 23 February 2009 / Accepted: 28 February 2009 / Published online: 17 March 2009© Springer-Verlag 2009
Abstract The 5S ribosomal RNA (5S rRNA) is an essen-tial component of ribosomes. Throughout evolution, varia-tion is found among 5S rRNA genes regarding theirchromosomal localization, copy number, and intergenicregions. In this report, we describe and compare the genesequences, motifs, genomic copy number, and chromo-somal localization of the Trichomonas vaginalis, Tricho-monas tenax, and Tritrichomonas foetus 5S rRNA genes.T. vaginalis and T. foetus have a single type of 5S rRNA-coding region, whereas two types were found in T. tenax.The sequence identities among the three organisms arebetween 94 and 97%. The intergenic regions are moredivergent in sequence and size with characteristic species-speciWc motifs. The T. foetus 5S rRNA gene has larger andmore complex intergenic regions, which contain either anubiquitin gene or repeated sequences. The 5S rRNA geneswere located in Trichomonads chromosomes by Xuorescentin situ hybridization.
Keywords Ribosome · Intergenic region · RNA polymerase III · Ubiquitin · Repeated sequences · Chromosome
Introduction
Trichomonas vaginalis, T. tenax, and T. foetus are parasiticamitochondriate protozoa that inhabit the urogenital tract ofhumans, the oral cavity of humans, and the urogenital tractof cattle, respectively. Phylogenetic analyses based on the16S ribosomal RNA (rRNA) gene place these organismsnear the base of the eukaryotic tree (Sogin 1991). Thischaracteristic, along with their amitochondriate nature andprokaryotic-like ribosomes, suggest that Trichomonads areancestral eukaryotic organisms that conserve early regula-tory mechanisms (Sogin 1991). Recently, the egalitarianmodel of evolution (Dacks et al. 2008) has challenged theprevious proposal: it divides eukaryotes into six majorsuper-groups, although the relationship among them andthe order of branching remain unknown. This new eukary-otic phylogeny emphasizes that organisms like Trichomo-nads are not primitive, but instead highly evolved parasitesspecialized for their speciWc environments. Today, the con-troversy continues and the phylogenetic location of Tricho-monads remains unclear.
Since ribosomes from Trichomonads have characteris-tics evocative of prokaryotes (Chakrabarti et al. 1992); thestudy of their rRNA genes may lead to the identiWcation ofsome eukaryotic divergent and/or speciWc regulatory pro-cesses. The typical eukaryotic ribosome is composed offour rRNAs and multiple proteins. Three of the rRNA mol-ecules (18S, 5.8S, and 28S) are encoded in the rRNA gene(rDNA) and are co-transcribed by RNA polymerase I (pol I).The fourth rRNA component, the 5S rRNA, is transcribed by
Communicated by S. Hohmann.
Nucleotide sequence data reported are available in the DDBJ/EMBL/GenBank databases under the accession numbers FJ492747, FJ492748, FJ492749, FJ492750, and FJ492751.
A. L. Torres-Machorro · R. Hernández · I. López-Villaseñor (&)Departamento de Biología Molecular y Biotecnología, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Apartado Postal 70-228, CP 04510 Mexico D.F., Mexicoe-mail: [email protected]
J. F. AldereteSchool of Molecular Biosciences, Washington State University, Pullman, WA, USA
123

200 Curr Genet (2009) 55:199–210
RNA polymerase III (pol III) and the genes are not generallylinked to the rDNA unit (Paule and White 2000).
The 5S rRNA gene is generally reiterated in the genomeand organized in tandem in head-to-tail repeats, which canbe encoded in one or in multiple gene clusters (Long andDawid 1980). Great variation is found in the localizationand number of chromosomal sites of the 5S rRNA genesthroughout evolution. Depending on the species, the genecopy number per genome can range from 3 copies in Plas-modium falciparum (Shippen-Lentz and Vezza 1988) toabout 106 copies in Euplotes carassus (Roberson et al.1989). These genes can be localized in a single or in multi-ple chromosomes or chromosome loci. In general, all thecopies of the 5S rRNA gene are considered identical in aparticular organism. However, some eukaryotes such asP. falciparum (Shippen-Lentz and Vezza 1988), Cryptos-poridium parvum (Taghi-Kilani et al. 1994), Neurosporacrassa (Selker et al. 1981), and mouse and human (Hallen-berg et al. 1994) have sequence and length variation in the5S rRNA intergenic region and eventually in the 5S rRNA-coding region. It has been reported that the tandem repeatsof the 5S rRNA genes can be linked to diVerent multigenefamilies. Among these are the rDNA in Dictyosteliumdiscoideum (Cockburn et al. 1978), some tRNA genes inEntamoeba histolytica (Clark et al. 2006), the splicedleader gene in Trypanosoma vivax (Roditi 1992), the his-tone gene cluster in crustaceans (Pelliccia et al. 2001), andthe ubiquitin genes in Tetrahymena pyriformis (Guerreiroet al. 1993).
The 5S rRNA genes are transcribed by pol III. In con-trast to pol I and pol II promoters, the pol III promoter ofthis gene is internal to the rRNA-coding region (internalcontrol region, ICR) (Schramm and Hernandez 2002). Insome organisms, the ICR alone is suYcient to direct polIII transcription, but in others such as yeast, the 5SrRNA gene transcription may also be regulated byupstream regulatory sequences, such as an upstream pro-moter element (upe) and a start site element (sse) (Leeet al. 1997).
In a previous work, we reported the sequence of theT. vaginalis 5S rRNA gene and identiWed putative pol IIIregulatory elements such as an internal promoter, upstreamregulatory sequences, and a palindromic sequence withinthe intergenic region that may indicate a high-energy hair-pin structure (Torres-Machorro et al. 2006). In this study,we compare the 5S rRNA gene sequence, genomic organi-zation, and chromosome localization among three Tricho-monad species. We found at least two types of 5S rRNAgenes in each of the three species, although most of thesequence variation, within and between species, was foundin the 5S rRNA intergenic region. T. vaginalis and T. tenaxare more similar in the 5S rRNA gene sequence, organiza-tion, size, and chromosomal localization, while T. foetus
has larger intergenic regions with particular species-spe-ciWc motifs, such as an ubiquitin open reading frame (ORF)and repeated sequences.
Materials and methods
Trichomonads and culture conditions
Trichomonas vaginalis CNCD147 isolate (Alvarez-Sanchez et al. 2000) and T. foetus (02-97) were grown aspreviously described (Espinosa et al. 2002). The T. tenaxHs-4:NIH was grown in LYI Entamoeba medium supple-mented with 10% fetal bovine serum as recommended byATCC.
Trichomonads 5S rRNA gene cloning and sequencing
To clone the 5S rRNA intergenic regions of the three Tric-homonads, we took advantage of the tandem organizationor these genes and of their sequence conservation (Torres-Machorro et al. 2006). Two independent PCR ampliWca-tions were made with genomic DNA (prepared as describedby Espinosa et al. 2002) and the 5I-F (5�-TACTGGGCTAGGAGACTT-3�) and 5I-R (5�-CGCGACTGCTTAATTTCCA-3�) oligonucleotides (oligos), which align withinthe 5S rRNA gene-coding region and extend toward theintergenic region (Fig. 1a). The ampliWcation reactions con-sisted of 30 cycles of 92°C denaturing for 1 min, 46°Cannealing for 1 min, and 72°C extension for 3 min, withTaq DNA polymerase (recombinant, Invitrogen). The PCRproducts were cloned using the Zero Blunt® TOPO® PCRcloning kit (Invitrogen). Eighteen clones from T. vaginalis,11 from T. tenax, and 9 from T. foetus were sequenced.
To clone the Type B-Tv 5S rRNA intergenic region of theT. vaginalis CNCD147 isolate, a genomic PCR was madeusing the Type B-Tv speciWc oligonucleotide RI335 (5�-CAAAATTAAACAAAATTTTCAGCAGAGAGTGGTGT-3�) and the 5S-R (5�-AAGCAAGCACCGCACGTTCC-3�)oligo. The annealing temperature was 54°C. The PCR productwas cloned using the Zero Blunt TOPO PCR cloning kit(Invitrogen) and four clones were sequenced.
To clone the 5S rRNA gene-coding region, PCR ampliW-cations were made using genomic DNA and the 5S-F (5�-AAGCGGCCACACCCGGCTGG-3�) and 5S-R oligos(Fig. 1a). The PCR ampliWcation conditions used were thesame as described for the intergenic region except forchanging the annealing temperature to 53°C. For each ofthe three organisms, at least three independent clones of thecoding region were sequenced. These 5S gene PCR prod-ucts were also used to estimate the gene copy number. Toobtain the complete 5S rRNA gene sequence, intergenicand coding region sequences were assembled as shown in
123

Curr Genet (2009) 55:199–210 201
Fig. 1a. Two clones were obtained containing severallinked 5S gene copies (see clone 5STtx-tq16, Fig. 1b),which allowed us to discriminate between the two types ofcoding regions in T. tenax.
5S rRNA gene copy number in Trichomonas tenax and Tritrichomonas foetus
The 5S rRNA gene copy number was estimated by a com-parative quantitative hybridization approach essentially asdescribed (Torres-Machorro et al. 2006; Chakrabarti et al.1992), except for the use of a slot blot instead of a South-ern blot approach. BrieXy, genomic DNA from T. tenaxand T. foetus was digested with HindIII, phenol extracted,ethanol precipitated, and resuspended in 0.4 M NaOH/10 mM EDTA solution. Decreasing amounts of this DNAwere loaded in a slot blot apparatus (Hoefer ScientiWc)alongside decreasing amounts of the corresponding 5SrRNA-coding region. This 116 bp 5S rRNA gene PCRproduct represents one gene copy. Additionally, the same5S gene-coding region PCR fragment was radioactivelylabeled (Rediprime II, GE Healthcare) and used as aprobe. The conditions used for hybridization were asdescribed (Espinosa et al. 2002).
Fluorescence in situ hybridization
Trichomonas vaginalis, T. foetus, and T. tenax cells wereprepared for in situ hybridization using the hypotonicswelling technique (Xu et al. 1998) with the followingmodiWcations. Cells grown in TYM medium with 10%horse serum were not treated with colchicine. Cells weregrown to a density of 2 £ 106 cells/ml, diluted to 1 £ 106
cells/ml in the same medium, grown further for 3 h at 37°C,and harvested. With this modiWcation, cells with nuclei inmultiple states of chromosome condensation were obtained.After centrifugation, cells were resuspended in 10 ml of10 mM KCl and incubated at 37°C for 5 min, followed bythe addition of 2 ml of Wxative solution (chilled methanolacetic acid 3:1) as an important pre-Wxing step. The nucleiwere then centrifuged at 200 £ g for 10 min at room tem-perature, the supernatant was removed and the nuclei pelletwas resuspended in 10 ml of ice-cold Wxative solution. Thislast step was repeated two more times. Finally, nuclei wereresuspended in a Wnal volume of 500 �l. As much as 60 �lof the nuclei suspension was dropped from a 1.3 m heightonto clean, grease-free slides. The smears on the slideswere heat-dried. The slides were aged in a nitrogen atmo-sphere for 3 days. Nuclei were Wxed by immersion of the
Fig. 1 Trichomonads 5S rRNA gene cloning and sequencing. a Schematic diagram of the tan-dem organization of the 5S rRNA genes. The regions where oligonucleotides anneal in the 5S rRNA genes are shown with arrows. The PCR ampliWcation products are shown with diVer-ent line styles. The assembly strategy is also shown. b Tricho-monads 5S rRNA gene frag-ments (obtained from TOPO clones) used for the synthesis of probes. c Trichomonads rDNA fragments used as probes for the FISH assays. The T. vaginalis Tv-p921 clone is a genomic clone. RcTx-5 and RcTf-11 are clones obtained by PCR ampliW-cation and cloning of T. tenax and T. foetus rDNA
123

202 Curr Genet (2009) 55:199–210
slides in 1% paraformaldehyde in 2X SSC (pH = 7.4) for30 s, washed twice in 2X SSC (pH = 7.4), and air dried.The slides were then treated with 100 �g/ml ribonuclease A(Sigma) in 2X SSC at 37°C for 1 h. After three washes in2X SSC, the DNA on the slides was denatured in 70%formamide/2X SSC (pH = 7.4) at 75°C for 2 min. Theywere immediately placed in 70% ethanol for 2 min, passedthrough dehydration series of 80 and 100% ethanol for2 min each, and allowed to air dry. As much as 20 �l of theheat-denatured probe (10 ng/�l) in hybridization solution[50% formamide, 10% dextran sulfate (Sigma), 2X SSC]was added to each slide, covered with a plastic coverslipand incubated overnight in a slightly dampened hybridiza-tion box at 37°C. The slides were washed with 50% form-amide/2X SSC for 5 min at 37°C, then in 2X SSC and 4XSSC at room temperature for 5 min. Spreads were stainedwith 0.125 �g/�l DAPI (Sigma) for 1 min, washed withPBS pH = 7.4, and mounted in Vectashield (Vector Labora-tories). Samples were visualized with an epiXuorescencemicroscope (BX5I Olympus) using objective Uplan Fl100X/1.30 Oil 1/0.17. Images were acquired with an Evo-lution VF color digital camera (Media Cybernetics) andwere analyzed with the Adobe Photoshop software.
Preparation of probes for FISH assays
The T. vaginalis 5S gene probe corresponds to the TypeA-Tv 5S rRNA gene from position ¡11 to +194 consider-ing the transcription start point as +1 (Fig. 1b). It includesthe whole 5S rRNA-coding region (118 bp) and 87 bp ofthe intergenic region. The probe was directly labeled withCy™3-dCTP (GE Healthcare) by PCR ampliWcation ofthe T. vaginalis 5S rRNA gene clone 5STv-ag2 (Fig. 1b)with 300 ng of each Pex1 (5�-CGAACAATCCTTTCGAAGCG-3�) and Pex2 (5�-AAGAGTTTTAGCAGGGACAA-3�) oligos (Fig. 1a). The PCR conditions were asdescribed for the ampliWcation of the 5S rRNA intergenicregion, replacing the dCTP with Cy3-dCTP. Salmonsperm DNA (20 �g; Invitrogen) was added to the PCRWnal product, which was precipitated with ethanol. Aftercentrifugation, the pellet was dissolved in distilled waterto a Wnal concentration of » 100 ng/�l. Before use, theprobe was diluted (1:10) in hybridization solution. Allprobes were treated in a similar way before use. TheT. tenax 5S gene probe contains two complete Type A-Ttx5S rRNA gene-coding and intergenic regions and twocomplete Type B-Ttx 5S rRNA gene-coding and inter-genic regions (1,235 bp insert in clone 5STtx-Tq16,Fig. 1b). The T. foetus 5S gene probe contains 94% of the5S rRNA-coding region and the complete Type B-Tf 5SrRNA intergenic region (1,356 bp insert in clone 5STf-pk16, Fig. 1b). The probes were labeled with Cy3-dCTPusing a Nick Translation System (Invitrogen).
The T. vaginalis rRNA gene probe contained the com-plete 6 kb rDNA unit of T. vaginalis (16S-like rRNA, 5.8SrRNA, 28S rRNA, and intergenic region). The Tv-p921genomic clone insert (Fig. 1c) was labeled by nick-transla-tion with Cy3-dCTP. The T. tenax and T. foetus rRNA geneprobes correspond to a 1,894 bp fragment of the rDNA cod-ing region, which includes the complete 16S and 5.8SrRNA-coding sequences, and 29 bp of the 28S rRNA(Fig. 1c). The inserts of clones RCTx-5 and RCTf-11(Fig. 1c) were labeled with Cy3-dCTP using the NickTranslation System (Invitrogen).
Results
Trichomonads 5S rRNA gene cloning, sequencing, and assembly
A T. vaginalis 5S rRNA gene clone was previouslyobtained using a PCR approach (Torres-Machorro et al.2006). The same approach was used for the ampliWcation,cloning, and sequencing of the 5S rRNA genes of T. tenaxand T. foetus, and to extend the T. vaginalis 5S rRNA geneanalysis. For each organism, two diVerent genomic PCRswere made: one to amplify the 5S rRNA-coding region anda second to amplify the 5S rRNA intergenic region. BothPCR products, which overlap in a 90 bp region, werecloned, sequenced, and assembled (Fig. 1a). Clones con-taining one or several tandem 5S rRNA genes wereobtained.
The 5S rRNA gene in Trichomonads
In a previous report, only one type of 5S rRNA gene andintergenic region was experimentally found in the T. vagi-nalis CNCD147 isolate (Torres-Machorro et al. 2006). Incontrast, in an in silico analysis of the T. vaginalis G3strain, whose complete genome has been sequenced (Carl-ton et al. 2007), two types of T. vaginalis 5S rRNA genescould be identiWed. These 5S rRNA gene variants, namedhere by us as Type A-Tv and Type B-Tv 5S rRNA genes,have identical coding sequences and a similar gene copynumber. Their intergenic regions diVer in size by one nucle-otide and have a sequence identity of 84%. The Type B-Tv5S rRNA gene has a TATA-like box located in the putativeupstream regulatory region, which is not present in theType A-Tv 5S gene (Fig. 2a, b; Table 1). The G3 isolateType A-Tv 5S rRNA gene is identical in sequence to theone previously found in the CNCD147 isolate.
Trichomonas tenax has at least two types of 5S rRNAgenes, which measure 307 bp (Type A-Ttx 5S gene) and316 bp (Type B-Ttx 5S gene) (Fig. 2c, d). The codingregions were assigned by alignment with the published
123

Curr Genet (2009) 55:199–210 203
Fig. 2 The 5S rRNA gene organization in Trichomonads. Graphicalrepresentation of the two types of 5S rRNA gene organization in thethree studied Trichomonads. A single copy of each variant is depicted,although the genes are found as tandem repeats in the genome. The 5SrRNA-coding regions (118 bp) are shown as boxes. The pol III pro-moter (ICR, internal control region) is internal to the coding region.The 5S rRNA upstream regulatory region (U) can have an upstreampromoter element (upe, triangle), a start site element (sse, arrow), andan AT-rich region or TATA-like box (diamond), and its compositionvaries depending on the 5S rRNA gene type. The coding region is fol-lowed by runs of Ts (T) of diVerent lengths, which act as the pol III
transcription termination signal. In T. vaginalis, the two types of 5SrRNA genes have a palindromic sequence (H) that can have slight se-quence variations. The T. tenax 5S rRNA gene has two types of codingand intergenic regions. The Type B-Ttx 5S gene has a unique palin-dromic region (HTtx) in the middle of the intergenic region. In T. foe-tus, the Type A-Tf 5S gene is characterized by the presence of anubiquitin ORF (in the intergenic region) with elements for transcrip-tion by pol II: the promoter (Inr, i) and the polyadenylation signal (A).The Type B-Tf 5S gene is characterized by the presence of an inter-rupted ubiquitin ORF and repeated sequences (R) of variable lengthsin the intergenic region
Table 1 Comparison of the 5S rRNA gene sequences and copy number in Trichomonads
Trichomonas vaginalis (%)
Trichomonas tenax Tritrichomonas foetus (%)
Ttx A (%) Ttx B (%)
Coding region identity
Tv 100 96 97 95
Ttx A 100 97 94
Ttx B 100 96
A-Tv (%) B-Tv (%) A-Ttx (%) B-Ttx (%) A-Tf (%) B-Tf (%)
Intergenic region identity
A-Tv (334 bp) 100 84 49 55 42 34
B-Tv (335 bp) 100 43 43 36 43
A-Ttx (307 bp) 100 57 47 46
B-Ttx (316 bp) 100 38 35
A-Tf (862 bp) 100 87
5S gene genome percentage (%) 0.10 0.01 0.04
123
204 Curr Genet (2009) 55:199–210
T. vaginalis sequence. Both variants contain the putativeinternal pol III promoter. The 5S rRNA putative upstreamregulatory region of both types of T. tenax genes is com-posed of a start site element, a GC-rich region, and aTATA-like box (Fig. 2c, d). The two types of 5S rRNAgene-coding regions (118 bp) are 97% identical insequence, diVering only in four nucleotides (Fig. 2;
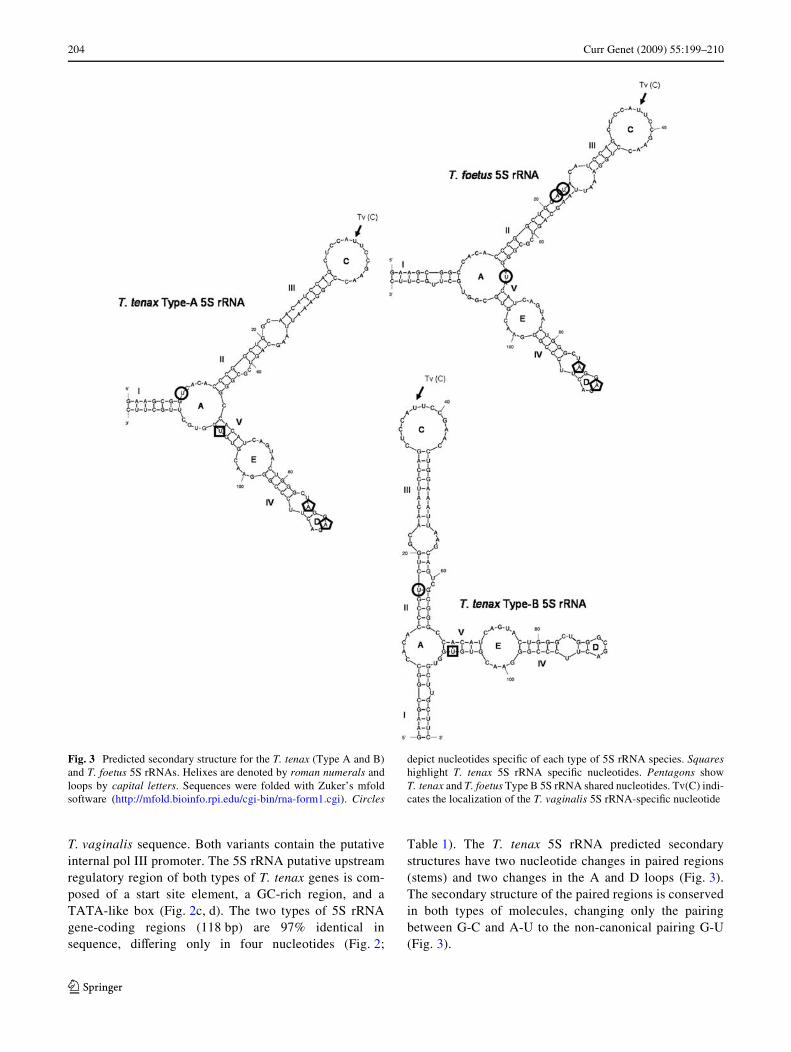
Table 1). The T. tenax 5S rRNA predicted secondarystructures have two nucleotide changes in paired regions(stems) and two changes in the A and D loops (Fig. 3).The secondary structure of the paired regions is conservedin both types of molecules, changing only the pairingbetween G-C and A-U to the non-canonical pairing G-U(Fig. 3).
Fig. 3 Predicted secondary structure for the T. tenax (Type A and B)and T. foetus 5S rRNAs. Helixes are denoted by roman numerals andloops by capital letters. Sequences were folded with Zuker’s mfoldsoftware (http://mfold.bioinfo.rpi.edu/cgi-bin/rna-form1.cgi). Circles
depict nucleotides speciWc of each type of 5S rRNA species. Squareshighlight T. tenax 5S rRNA speciWc nucleotides. Pentagons showT. tenax and T. foetus Type B 5S rRNA shared nucleotides. Tv(C) indi-cates the localization of the T. vaginalis 5S rRNA-speciWc nucleotide
123

Curr Genet (2009) 55:199–210 205
Tritrichomonas foetus has at least two types of 5S rRNAgenes. These genes are identical in the coding region, butdiVer in the intergenic region sequence. We named thesegenes as Type A-Tf 5S rRNA gene (862 bp) and Type B-Tf5S rRNA gene (» 1,300 bp) (Fig. 2e, f). The T. foetus 5SrRNA-coding region (118 bp) conserves the putative pol IIIpromoter sequence and secondary structure described in theT. vaginalis 5S rRNA gene (Fig. 3). Compared to theT. vaginalis 5S rRNA gene-coding region, it has threenucleotide variations (Fig. 4). The putative upstream regu-latory region in both types of 5S rRNA gene are composedof a start site element, an upstream promoter element, andan AT-rich region (Fig. 2e, f). The typical transcription ter-mination signal for pol III, a run of Ts, was found in the 3�-side of the 5S rRNA-coding region of the three Trichomo-nads (Fig. 2).
The 5S rRNA gene-coding region sequence was com-pared among the three Trichomonads, and a very high simi-larity was found among the three with a sequence identityranging from 94 to 97% (Table 1). The Type B-Ttx and theT. vaginalis 5S rRNA gene-coding sequences are the mostrelated pair with 97% sequence identity. We found thateach Trichomonad has speciWc nucleotide variations withinthe 5S rRNA-coding region, and these are “species-spe-ciWc” (Fig. 4) and useful for the delineation among theTrichomonad species. It is noteworthy that the expressionof both types of 5S rRNA genes in T. tenax and T. foetuswas experimentally conWrmed by RT-PCR (data notshown).
The 5S rRNA intergenic region motifs in Trichomonads
Two diVerent 5S rRNA intergenic regions were identiWedin silico in the genome of the T. vaginalis G3 isolate, whichdiVer in size by one nucleotide and have a sequence identityof 84%. Since a single 5S gene sequence was previouslyfound in the T. vaginalis CNCD147 isolate (Torres-Mach-orro et al. 2006), we experimentally searched for the secondtype of 5S rRNA intergenic region in this isolate. Type B-Tv 5S gene could only be identiWed by PCR ampliWcation
using Type B-Tv 5S gene-speciWc oligonucleotides. Withthis approach, eight sequence variants in the palindromicregion were also identiWed (Fig. 5). Nevertheless, sequencevariation in the hairpin region was also found in an in silicoanalysis of the 5S rRNA gene in the T. vaginalis G3 isolate(Fig. 5).
Trichomonas tenax has at least two types of 5S rRNAgene, with 307 and 316 bp in length (Fig. 2c, d). The T.tenax 5S rRNA intergenic regions are only 59% identical insequence (Table 1) and diVer by the presence of a putativehigh-energy hairpin (¡9.6 kcal/mol) located in position¡118 of Type B-Ttx 5S gene intergenic region (Fig. 2d).This hairpin shares no sequence or position similarity withthe one found in the T. vaginalis 5S rRNA gene. Six TypeA and Wve Type B clones were identiWed by sequence, sug-gesting a similar number of 5S rRNA genes from each type.
Trichomonas foetus has at least two types of 5S rRNAgenes with 862 and 1,262–1,367 bp in length. The Type A-Tf 5S rRNA intergenic region (744 bp) contains a completeORF for ubiquitin (Fig. 2e) and a typical pol II promoter,which is identical to the Inr previously described in T. vagi-nalis (Liston and Johnson 1999). In the 3�-end of the ubiq-uitin gene the mRNA polyadenylation signal for T.vaginalis was found (Espinosa et al. 2002). The Type B-Tf5S gene intergenic region can vary in size from 1,144 to1,249 bp, depending on the number of repetitions of a 13 bprepeated element. The intergenic region is characterized bythe presence of two blocks of repeated sequences: the Wrstblock is composed of between 5 and 12 tandem repeatedmotifs (13 bp long), which were found to interrupt the ubiq-uitin-coding region. The second block is composed of twosequences of 47 and 71 bp, with each sequence repeatedonce. The 47 bp repeated sequences are located in positions¡233 and ¡516 considering the tsp as +1 (Fig. 2f). Inter-estingly, the 71 bp block contains the putative 5S geneupstream regulatory region (positions ¡76 and ¡408).Remarkably, the two types of 5S rRNA intergenic regionsin T. foetus are very similar in sequence (87% of identity,Table 1) even though they diVer in size by about 450 bp.This variation in size is mainly due to the repetitive nature
Fig. 4 Sequence comparison of the three Trichomonads 5S rRNA-coding region. Alignment of the 5S rRNA-coding sequences found inTrichomonads. The pol III promoter (ICR) is composed of a box A, anintermediate element (IE) and a box C. T. tenax has two types of 5S
rRNA-coding sequences, named 5STtxA and 5STtxB, while a singlecoding sequence was found in T. vaginalis and in T. foetus. The spe-cies-speciWc sequence variations are denoted in black fonts
123

206 Curr Genet (2009) 55:199–210
of the Type B-Tf intergenic region (Fig. 2f). Three Type A-Tf and six Type B-Tf clones were obtained, suggesting thepresence of more Type B-Tf gene copies in the T. foetusgenome.
5S rRNA gene copy number and chromosomal localization in Trichomonads
To Wnd out the approximate genomic copy number of the5S rRNA gene in T. tenax and T. foetus, a quantitative–qualitative hybridization analysis was used (Fig. 6). Thistype of quantiWcation is based on the known genome size ofa given organism. The genome size of T. vaginalis is con-troversial, ranging in proposed sizes from 2.5 £ 107 to17.6 £ 107 bp (Wang and Wang 1985; Carlton et al. 2007).On the other hand, the size of T. tenax and T. foetusgenomes are also unclear, since their size determinationwas based on the T. vaginalis genome (Zubacova et al.2008). Nevertheless, it is apparent that the genome size issimilar among the three species (Zubacova et al. 2008).Therefore, due to the diYculty in deWning a reliable 5S
rRNA gene copy number, we estimated the percentage ofthe genome sequence that corresponds to the 5S rRNAgene-coding region in each Trichomonad genome(Table 1). The 5S rRNA genes coding region correspond to0.10% of the genome in T. vaginalis and 0.04% of thegenome in T. foetus, whereas in T. tenax this value is aboutone-tenth of the T. vaginalis (0.01% of the genome).
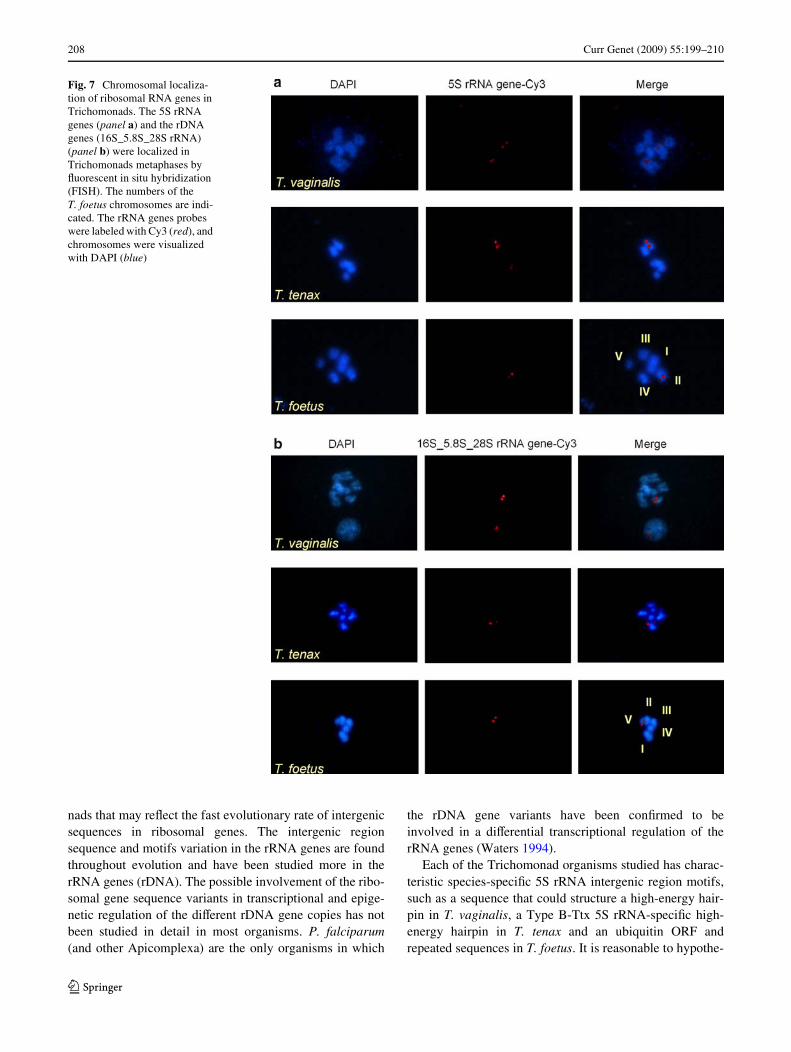
The chromosomal localization of the 5S rRNA genes inTrichomonads was determined by a Xuorescent in situhybridization (FISH) procedure. The chromosomal locali-zation of the rDNA genes was also determined as a control.It has been reported that T. vaginalis and T. tenax have sixchromosomes (Lehker and Alderete 1999; Zubacova et al.2008), while Wve chromosomes have been identiWed inT. foetus (Xu et al. 1998). As shown in Fig. 7, the 5S rRNAgene is located in two chromosomes in T. vaginalis and inT. tenax, whereas this gene is found in a single chromo-some in T. foetus (Fig. 7a).
Due to the similar size of some of the T. vaginalis andT. tenax chromosomes, it was diYcult to classify and/orname them using our microscopy images. The T. vaginalis
Fig. 5 Variation in the palindromic sequence found in the 5S rRNAintergenic region of the T. vaginalis G3 strain and CNCD147 isolate.All the hairpin sequence variations, except G3-2, correspond to se-quenced palindromic region variants found in the 5S rRNA gene inter-
genic region of the CNCD147 isolate. The Wrst two lines correspond tothe hairpin variants found in the G3 isolate. The putative hairpin struc-tures were obtained using the software Mfold (Zuker 2000)
123

Curr Genet (2009) 55:199–210 207
chromosomes have been deWned as three big, two interme-diate sized, and one mini-chromosome (Lehker and Alderete1999). For T. tenax, we identiWed two large and four smallchromosomes. On the other hand, the T. foetus chromo-somes were easily distinguished due to their clear size vari-ation. We were able to identify all T. foetus chromosomes(I–V) according to their size (Fig. 7). In T. vaginalis, the 5SrRNA genes are localized in two of its three large chromo-somes, while the rDNA gene is localized in the mini-chro-mosome (Fig. 7). In T. tenax, the 5S rRNA genes arelocalized in one large and one small chromosome, while therDNA genes are localized in one of the large chromosomes(Fig. 7). In contrast, the T. foetus 5S rRNA genes are foundin a single chromosome (II), whereas the rDNA genes arelocalized in the smallest chromosome (V) (Fig. 7).
Discussion
The high sequence identity in the 5S rRNA gene-codingregions of the three Trichomonad species aYrms a closephylogenetic relationship among them (Felleisen 1997).Likewise, our data show a conservation of the pol III pro-moter. Interestingly, we found two 5S rRNA-coding
sequence variants in T. tenax, which include variation ofthe promoter region. This variation within the promoterregion may result in putative diVerential transcriptional reg-ulation as seen in the Plasmodium rDNA genes (Waters1994). On the other hand, the variation in the codingsequence may result in the synthesis of two types of 5SrRNA mature molecules worthy of future experimentalanalyses to understand the functional implications, if any.
The putative 5S rRNA upstream regulatory region,which includes the start site element, the upstream pro-moter element, and sometimes a TATA-like box or an AT-rich region is relatively well conserved among the threeTrichomonad species. These kinds of elements have beenconWrmed as important transcriptional regulatory motifs inyeast (Lee et al. 1997). Interestingly, the 5S rRNA genevariants in the T. vaginalis G3 isolate have a diVerentialTATA-like motif in the 5S rRNA upstream regulatoryregion whose potential role in transcription is a workinghypothesis in our laboratory.
Since a very high sequence identity was found in the 5SrRNA gene-coding region among the three species, we alsoexpected a relatively high similarity in the sequence andmotifs of their intergenic regions. However, we found inter-esting diVerences in the intergenic regions of the Trichomo-
Fig. 6 5S rRNA gene copy number estimation in T. tenax and T. foetus. A comparative quantitative hybridization was performed to estimate the 5S rRNA gene copy number in T. tenax (a) and T. foetus (b). The amount of DNA loaded in each well is indicated. The probes correspond to the com-plete 5S rRNA-coding region of each organism, labeled with 32P-dCTP by the random primer technique
123
208 Curr Genet (2009) 55:199–210
nads that may reXect the fast evolutionary rate of intergenicsequences in ribosomal genes. The intergenic regionsequence and motifs variation in the rRNA genes are foundthroughout evolution and have been studied more in therRNA genes (rDNA). The possible involvement of the ribo-somal gene sequence variants in transcriptional and epige-netic regulation of the diVerent rDNA gene copies has notbeen studied in detail in most organisms. P. falciparum(and other Apicomplexa) are the only organisms in which
the rDNA gene variants have been conWrmed to beinvolved in a diVerential transcriptional regulation of therRNA genes (Waters 1994).
Each of the Trichomonad organisms studied has charac-teristic species-speciWc 5S rRNA intergenic region motifs,such as a sequence that could structure a high-energy hair-pin in T. vaginalis, a Type B-Ttx 5S rRNA-speciWc high-energy hairpin in T. tenax and an ubiquitin ORF andrepeated sequences in T. foetus. It is reasonable to hypothe-
Fig. 7 Chromosomal localiza-tion of ribosomal RNA genes in Trichomonads. The 5S rRNA genes (panel a) and the rDNA genes (16S_5.8S_28S rRNA) (panel b) were localized in Trichomonads metaphases by Xuorescent in situ hybridization (FISH). The numbers of the T. foetus chromosomes are indi-cated. The rRNA genes probes were labeled with Cy3 (red), and chromosomes were visualized with DAPI (blue)
123

Curr Genet (2009) 55:199–210 209
size that the role of these motifs is for the regulation of 5SrRNA expression. The ubiquitin gene linkage to the 5SrRNA gene has also been observed in an unrelated species,Tetrahymena pyriformis (Guerreiro et al. 1993), makinginteresting the study of the functional relevance of this typeof multigenic families’ genomic linkage.
It is interesting to note that the organization of the T. foe-tus 5S rRNA gene-repeated sequences is similar to theorganization of the rDNA main transcription unit in meta-zoans. In the rDNA, the “spacer promoter” (a sequencesimilar to the ribosomal promoter, which is repeated withinthe intergenic region) has been proposed to have a role inrecruiting the pol I transcription machinery as a mechanismto enhance the rDNA transcriptional eYciency (Paule andWhite 2000). Whether the repeated sequences have a role inregulating the 5S rRNA gene expression requires experi-mental veriWcation in the future.
We have shown that the 5S rRNA gene copies arelocated in at least two unlinked clusters of two diVerentchromosomes in T. tenax and T. vaginalis. In contrast, the5S rRNA genes in T. foetus seem to be located in a singlecluster in one chromosome only. The fact that T. foetus hasone 5S rRNA cluster and one chromosome less than T.tenax and T. vaginalis makes the analysis of karyotypes inthese organisms an interesting topic of research. Whetherthe genes from the sixth chromosome of T. tenax and T.vaginalis were reorganized, compacted or lost in the T. foe-tus genome, are questions with interesting implications inevolution.
The rDNA genes are localized in a single chromosomein the three Trichomonad species, and this may be relatedto the small rDNA copy number in these organisms (Chak-rabarti et al. 1992). In T. vaginalis and T. foetus, the rDNAis found in the smallest chromosome, meaning that therDNA is not linked to the 5S rRNA gene in these organ-isms. In contrast, in T. tenax the 5S rRNA is located in oneof the large chromosomes. Therefore, even though T. tenaxand T. vaginalis are two closely related species, their geno-mic organization does not seem to be completely con-served. Few karyotyping analyses have been done inTrichomonads (Lehker and Alderete 1999; Zubacova et al.2008) and no chromosomal markers have been assigned todate. As we have shown, the 5S rRNA genes and the rDNAgenes have a well-deWned chromosomal localization inTrichomonads. We, therefore, propose these genes as goodchromosomal markers for these organisms.
Acknowledgments This work was supported by grants P45037-Qand CB-2007-079983 from Consejo Nacional de Ciencia y Tecnología(CONACYT) México, and IN214006 from Programa de Apoyo a Pro-yectos de Investigación e Innovación Tecnológica (PAPIIT), Univers-idad Nacional Autónoma de México (UNAM). A scholarship fromCONACYT to A.L. Torres-Machorro is acknowledged. The authorsthank Karla López Pacheco for cloning the T. tenax and T. foetus
rDNA fragments, and Ashwini Kucknoor for help with the T. tenaxcultures. Useful information for the FISH assay was kindly shared byM.E. Gonsebatt’s and F. Recillas-Targa’s laboratories (UNAM).
ConXict of interest statement The authors declare that they have noconXict of interest.
References
Alvarez-Sanchez ME, Avila-Gonzalez L, Becerril-Garcia C, Fattel-Facenda LV, Ortega-Lopez J, Arroyo R (2000) A novel cysteineproteinase (CP65) of Trichomonas vaginalis involved in cytotox-icity. Microb Pathog 28:193–202
Carlton JM, Hirt RP, Silva JC, Delcher AL, Schatz M, Zhao Q,Wortman JR, Bidwell SL, Alsmark UC, Besteiro S, Sicheritz-Ponten T, Noel CJ, Dacks JB, Foster PG, Simillion C, Van de PY,Miranda-Saavedra D, Barton GJ, Westrop GD, Muller S, Dessi D,Fiori PL, Ren Q, Paulsen I, Zhang H, Bastida-Corcuera FD,Simoes-Barbosa A, Brown MT, Hayes RD, Mukherjee M,Okumura CY, Schneider R, Smith AJ, Vanacova S, Villalvazo M,Haas BJ, Pertea M, Feldblyum TV, Utterback TR, Shu CL,Osoegawa K, de Jong PJ, Hrdy I, Horvathova L, Zubacova Z,Dolezal P, Malik SB, Logsdon JM Jr, Henze K, Gupta A, WangCC, Dunne RL, Upcroft JA, Upcroft P, White O, Salzberg SL,Tang P, Chiu CH, Lee YS, Embley TM, Coombs GH, MottramJC, Tachezy J, Fraser-Liggett CM, Johnson PJ (2007) Draftgenome sequence of the sexually transmitted pathogen Tricho-monas vaginalis. Science 315:207–212
Chakrabarti D, Dame JB, Gutell RR, Yowell CA (1992) Characteriza-tion of the rDNA unit and sequence analysis of the small subunitrRNA and 5.8S rRNA genes from Tritrichomonas foetus. MolBiochem Parasitol 52:75–83
Clark CG, Ali IK, Zaki M, Loftus BJ, Hall N (2006) Unique organisa-tion of tRNA genes in Entamoeba histolytica. Mol BiochemParasitol 146:24–29
Cockburn AF, Taylor WC, Firtel RA (1978) Dictyostelium rDNA con-sists of non-chromosomal palindromic dimers containing 5S and36S coding regions. Chromosoma 70:19–29
Dacks JB, Walker G, Field MC (2008) Implications of the neweukaryotic systematics for parasitologists. Parasitol Int 57:97–104
Espinosa N, Hernandez R, Lopez-Griego L, Lopez-Villasenor I (2002)Separable putative polyadenylation and cleavage motifs in Trich-omonas vaginalis mRNAs. Gene 289:81–86
Felleisen RS (1997) Comparative sequence analysis of 5.8S rRNAgenes and internal transcribed spacer (ITS) regions of trichomo-nadid protozoa. Parasitology 115:111–119
Guerreiro P, Neves A, Rodrigues-Pousada C (1993) Clusters of 5S rR-NAs in the intergenic region of ubiquitin genes in Tetrahymenapyriformis. Biochim Biophys Acta 1216:137–139
Hallenberg C, Nederby NJ, Frederiksen S (1994) Characterization of5S rRNA genes from mouse. Gene 142:291–295
Lee Y, Wong WM, Guyer D, Erkine AM, Nazar RN (1997) In vivoanalyses of upstream promoter sequence elements in the 5 SrRNA gene from Saccharomyces cerevisiae. J Mol Biol 269:676–683
Lehker MW, Alderete JF (1999) Resolution of six chromosomes ofTrichomonas vaginalis and conservation of size and numberamong isolates. J Parasitol 85:976–979
Liston DR, Johnson PJ (1999) Analysis of a ubiquitous promoter ele-ment in a primitive eukaryote: early evolution of the initiator ele-ment. Mol Cell Biol 19:2380–2388
Long EO, Dawid IB (1980) Repeated genes in eukaryotes. Annu RevBiochem 49:727–764
123

210 Curr Genet (2009) 55:199–210
Paule MR, White RJ (2000) Survey and summary: transcription byRNA polymerases I and III. Nucleic Acids Res 28:1283–1298
Pelliccia F, Barzotti R, Bucciarelli E, Rocchi A (2001) 5S ribosomal andU1 small nuclear RNA genes: a new linkage type in the genome ofa crustacean that has three diVerent tandemly repeated units con-taining 5S ribosomal DNA sequences. Genome 44:331–335
Roberson AE, WolVe AP, Hauser LJ, Olins DE (1989) The 5S RNAgene minichromosome of Euplotes. Nucleic Acids Res 17:4699–4712
Roditi I (1992) Trypanosoma vivax: linkage of the mini-exon (splicedleader) and 5S ribosomal RNA genes. Nucleic Acids Res 20:1995
Schramm L, Hernandez N (2002) Recruitment of RNA polymerase IIIto its target promoters. Genes Dev 16:2593–2620
Selker EU, Yanofsky C, Driftmier K, Metzenberg RL, Alzner-DeW-eerd B, RajBhandary UL (1981) Dispersed 5S RNA genes in N.crassa: structure, expression and evolution. Cell 24:819–828
Shippen-Lentz DE, Vezza AC (1988) The three 5S rRNA genes fromthe human malaria parasite Plasmodium falciparum are linked.Mol Biochem Parasitol 27:263–273
Sogin ML (1991) Early evolution and the origin of eukaryotes. CurrOpin Genet Dev 1:457–463
Taghi-Kilani R, Remacha-Moreno M, Wenman WM (1994) Three tan-demly repeated 5S ribosomal RNA-encoding genes identiWed,cloned and characterized from Cryptosporidium parvum. Gene142:253–258
Torres-Machorro AL, Hernandez R, Sanchez J, Lopez-Villasenor I(2006) The 5S ribosomal RNA gene from the early diverging pro-tozoa Trichomonas vaginalis. Mol Biochem Parasitol 145:269–273
Wang AL, Wang CC (1985) Isolation and characterization of DNAfrom Tritrichomonas foetus and Trichomonas vaginalis. Mol Bio-chem Parasitol 14:323–335
Waters AP (1994) The ribosomal RNA genes of Plasmodium. AdvParasitol 34:33–79
Xu WD, Lun ZR, Gajadhar A (1998) Chromosome numbers of Tri-trichomonas foetus and Tritrichomonas suis. Vet Parasitol78:247–251
Zubacova Z, Cimburek Z, Tachezy J (2008) Comparative analysis oftrichomonad genome sizes and karyotypes. Mol Biochem Parasi-tol 161:49–54
Zuker M (2000) Calculating nucleic acid secondary structure. CurrOpin Struct Biol 10:303–310
123