co-occurrence patterns of wood-decaying fungi on picea abies logs: does fomitopsis pinicola...
TRANSCRIPT

Václav POUSKA1, 2*, Miroslav SVOBODA1, Jan LEPŠ2, 3
1 Faculty of Forestry and Wood Sciences, Czech University of Life Sciences in Prague, Kamýcká 129, 165 21 Praha 6 – Suchdol, Czech Republic, *e-mail: [email protected] (corresponding author)
2 Faculty of Science, University of South Bohemia, Branišovská 31, 370 05 České Budějovice, Czech Republic
3 Institute of Entomology, Academy of Sciences of the Czech Republic, Branišovská 31, 370 05 České Budějovice, Czech Republic
CO-OCCURRENCE PATTERNS OF WOOD-DECAYING FUNGI ON PICEA ABIES LOGS: DOES FOMITOPSIS PINICOLA
INFLUENCE THE OTHER SPECIES?
POLISH JOURNAL OF ECOLOGY(Pol. J. Ecol.)
61 1 119–134 2013
Regular research paper
ABSTRACT: Dominant fungi, especially primary decayers, probably influence other fungi growing together with them. Fomitopsis pinicola is one of the important primary decayers, and it has been shown that several other species regularly co-occur together with it. We asked whether the presence of common species (es-pecially F. pinicola) affects the species richness and composition of other fungi. This study was conducted in an old-growth mountain spruce forest in the Bohemian Forest, Czech Republic. We surveyed logs on the ground for sporocarps of fungi in three successive years. Characteris-tics of logs such as dimensions, stage of decay and the cause of tree death (wind, competition, butt rot, bark beetles and unascertained) were recorded. F. pinicola was abundant mostly on logs that originated from trees infested by bark beetles. Analysis of covariance with the volume of logs and decay stage as covariables showed significant effect of these covariables and of F. pinicola presence on species numbers – logs in middle decay stages with the sporocarps of F. pinicola had more species than other logs. Based on Canonical Correspondence Analysis with vol-ume, decay stage and the cause of tree death as covariables, the species composition on logs was also influenced by F. pinicola. We found such statistical effects in several other species. Red-listed species Antrodiella citrinella and Camarops tubulina co-occurred with F. pinicola.
KEY WORDS: coarse woody debris, macro-mycetes, Norway spruce, species interaction
1. INTRODUCTION
Although interactions between wood de-cay fungi are mostly antagonistic and succes-sion often proceeds as the resource capture by combat (B oddy 2000), some fungi, when se-nescent, may facilitate establishing of certain specialised species (Niemelä et al. 1995). However, regardless of positive or negative relationships, the co-occurrence of some spe-cies is merely due to their similar habitat re-quirements (O vaskainen et al. 2010) and most fungi in late stages of wood decomposi-tion seem to be independent, appearing and disappearing without evident correlations to other decayers (Niemelä et al. 1995). My-corrhizal fungi also grow in dead wood (e.g. Tedersoo et al. 2003, R aja la et al. 2011) and interact with wood-decayers; some of them, for example, are able to capture nutri-ents from wood-decaying fungi (Dighton et al. 1987, L indahl et al. 1999).
Wood-decaying fungi spread predomi-nantly by spores but some of them can in-vade wood through the ground. Spore size and shape differ in many species and so their
journal 33.indb 119 2013-04-30 10:05:39

Václav Pouska et al.
abilities to disperse and establish in the sub-strate (Kauserud et al. 2008). Several fungi such as Amylostereum areolatum (Chail-let: Fr.) Boidin (Vasi l iauskas et al. 1998) or Graphium spp. (Mül ler at al. 2002) are also regularly spread by insects. While spore dispersal by air is non-targeted, insects pre-fer different types of spruce wood (Jonsel l and Wesl ien 2003) and probably make the spread of associated fungi far less random. Some insects are also sensitive to fungi al-ready present in wood (Jonsel l et al. 2005, Abrahamsson et al. 2008, Wesl ien et al. 2011). Wind, insects and fungi are common mortality agents of trees (Worra l l et al. 2005, L ännenpää et al. 2008) and it is often difficult to distinguish primary cause of tree death. The occurrence of fungi is intercon-nected with various causes of tree mortality (Renval l 1995, Edman et al. 2007, Pouska et al. 2011).
The major changes in fungal species com-position and richness occur with the progres-sion in wood decay (Renval l 1995, Høi land and B endiksen 1997, Hei lmann-Clau-sen 2001, Fukasawa et al. 2009, R aja la et al. 2011). There is also a strong influence of the size of spruce logs on species composition (Høi land and B endiksen 1997, Pouska et al. 2011, R aja la et al. 2011) and richness (e.g. Høi land and B endiksen 1997). Dif-ferences in the inoculation of wood by fungi influence subsequent species turnover (e.g. Fukami et al. 2010). We may ask whether and how some dominant fungi, especially pri-mary decayers, influence other fungi growing in the same logs. Fomitopsis pinicola (Swartz: Fr.) P. Karst. is one of the important primary decayers of spruce wood and Renval l (1995) showed that several other species regularly co-occur together with it, similarly Spirin and Shirokov (2002). However, L indblad (1998) did not find any influence of F. pini-cola on the diversity of other species. O vas-kainen et al. (2010) found either positive or negative correlations with several species.
Our sporocarp-based study has a limited potential to reveal causal relationships in the occurrence of fungal species. The only un-equivocal proof of the effect of any species on the others would be through a manipulative experiment, in which logs would be artificial-ly inoculated by a fungus. Only experiments
in vitro were done (e.g. Holmer and Sten-l id 1997, Iakovlev and Stenl id 2000, Fu-kami et al. 2010) until a recent experiment in situ (L indner et al. 2011). However, it is extremely difficult do to similar experiments in the field, especially to control or reveal spe-cies assemblages. Therefore, relying on spe-cies co-occurrence patterns with accounting for possible effects of other factors still seems to be a good way (O vaskainen et al. 2010).
Since 1990s, large areas of spruce stands in the Bohemian Forest have been affected by bark beetle outbreaks (e.g. Mül ler et al. 2008). This has led to the multiple increase of the amount of dead wood in formerly man-aged and in some protected forests. F. pinicola has become very abundant in unmanaged stands (Bässler and Mül ler 2010), which has probably influenced the community of wood-decaying fungi. We ask whether the presence of F. pinicola, or the presence of oth-er common species, affects the species rich-ness and composition of other fungi in logs.
2. MATERIAL AND METHODS
2.1. Study site
We conducted this study in an old-growth mountain spruce forest in the southern part of the Bohemian Forest in the Czech Repub-lic (48°46’30’’N, 13°49’30’’E). The locality is on the northern slope of the mountain Tro-jmezná (Bayerischer Plöckenstein), in a part of the reserve officially protected since 1933 (current area of the reserve is 598 ha). This reserve is a part of the Šumava National Park. Vegetation at the locality was classified as Calamagrostio villosae-Piceetum and Athyrio alpestris-Piceetum associations (Neuhäus-lová 2001). During the study period, Nor-way spruce (Picea abies (L.) Karsten) was the only dominant tree species. Another species, rowan (Sorbus aucuparia L.), was restricted to understorey. Based on historical manage-ment plans, study area has not been affected by major logging. Groups of trees killed by bark beetles occurred in the area after 1995. As a measure against bark beetles, selective salvage logging was carried out between 1999 and 2006; some trees were cut, their bark peeled off, and the logs were left at the site. After our sampling period, a windstorm in
120
journal 33.indb 120 2013-04-30 10:05:39

Co-occurrence of wood-decaying fungi on Picea abies
2007 exacerbated the bark beetle outbreak and by 2008 most of the canopy trees were dead.
2.2. Fomitopsis pinicola
Fomitopsis pinicola is a polypore with perennial sporocarps, causing brown cubi-cal rot, widely distributed in coniferous for-est regions of the northern hemisphere; it grows both on conifers and broadleaved trees and is able to live both as parasite and sapro-troph (Ryvarden and Gi lber tson 1993). In former Czechoslovakia, it was commonly collected on P. abies, Abies alba Mill., Betula pendula Roth, Prunus avium L. and Pinus syl-vestris L. (Kot laba 1984). It does not spread by mycelium through the ground, probably only by spores (Nordén 1997). Many inver-tebrates like mites, collembolans and insects graze on fungi. Sporocarps of F. pinicola pro-vide habitat for a variety of insects, such as Coleoptera, Diptera, Lepidoptera and Hyme-noptera (Jonsel l and Nordlander 2002).
2.3. Wood and fungi sampling
We recorded characteristics of individual spruce logs and the presence of fungal spe-cies on these logs in 12 circular plots (2000 m2 each) that lay at the elevation from 1220 to 1340 m. The minimal distance of plot cen-tres is 100 m. All parts of lying spruce stems with a diameter ≥ 10 cm at the larger end that originated from the plots were included in this study. The setting of these plots and the measurement of logs was described in detail in Svoboda and Pouska (2008) and Pous-ka et al. (2010). In 2006, the volume of natu-ral logs varied from 39 to 206 m3 ha–1, and the average log volume was 1 m3. The basic sampling unit of this study is “a log”. In some cases, tree stems broke upon falling, and such separate parts were pooled together to form one sampling unit. In other cases, however, the upper tree part broke and fell first, fol-lowed several years later by the breaking of lower part of the stem at the base. Such parts, although originally belonging to the same tree, undergo different progressions in de-velopment, and thus were not pooled (each part is a separate sampling unit). Although one can argue that if species A grows on one
part of the original stem, and species B on another, detached, part of the stem, little can be said about the co-occurrence of species A and B, there are reasons to pool broken parts in studies focused on communities in logs. The most important is a prevalence of several species on tree tops or bases (see Renval l 1995). Separating such accidentally detached parts would disturb the co-occurrence pat-tern, whereas some other stems (broken at the base or windthrows) remain almost com-plete. In addition to the fact that most logs are not complete but broken higher at the stem, this would introduce even more noise into the data.
We assessed the stage of decay for logs on a five-degree scale according to Sippola and Renvall (1999) using an iron spike (length 20 cm, max. diameter 7 mm; a log was hit with a certain force) as follows: 1 – recently dead trunk/piece of wood; wood is hard (spike pen-etrates only a few mm into the wood), com-pletely covered with bark, phloem fresh in at least some places; 2 – wood mostly hard (spike penetrates 1–2 cm into the wood), most of the bark left (though not necessarily for bark bee-tle-infested trunks), but no fresh phloem pres-ent; 3 – wood partly decayed on the surface or in the centre (spike penetrates 3–5 cm into the wood at most places), large pieces of bark usually loosened or detached; 4 – most of the wood soft throughout, the whole spike (15–20 cm) penetrates into the wood (depending on wood diameter). However, the central parts can remain hard, while the surface layers of the wood can be missing; 5 – wood very soft, disintegrates when lifted; trunk usually cov-ered by field-layer vegetation.
We also assessed causes of tree death as in Pouska et al. (2011): wind – windthrows with a root plate or crown breaks of living trees, competition, butt rot, bark beetles, and unascertained. More than one cause was as-sessed for some logs and probabilities of the causes were combined using fuzzy coding. This means that the probabilities of all death causes together sum up to one for each log. Cut logs (recently peeled and cut long ago) were not included in this study.
We carried out inventories of wood-de-caying fungi in three successive years (2004–2006), in the period from June to November. We visited each plot four times. Logs were
121
journal 33.indb 121 2013-04-30 10:05:40
Václav Pouska et al.
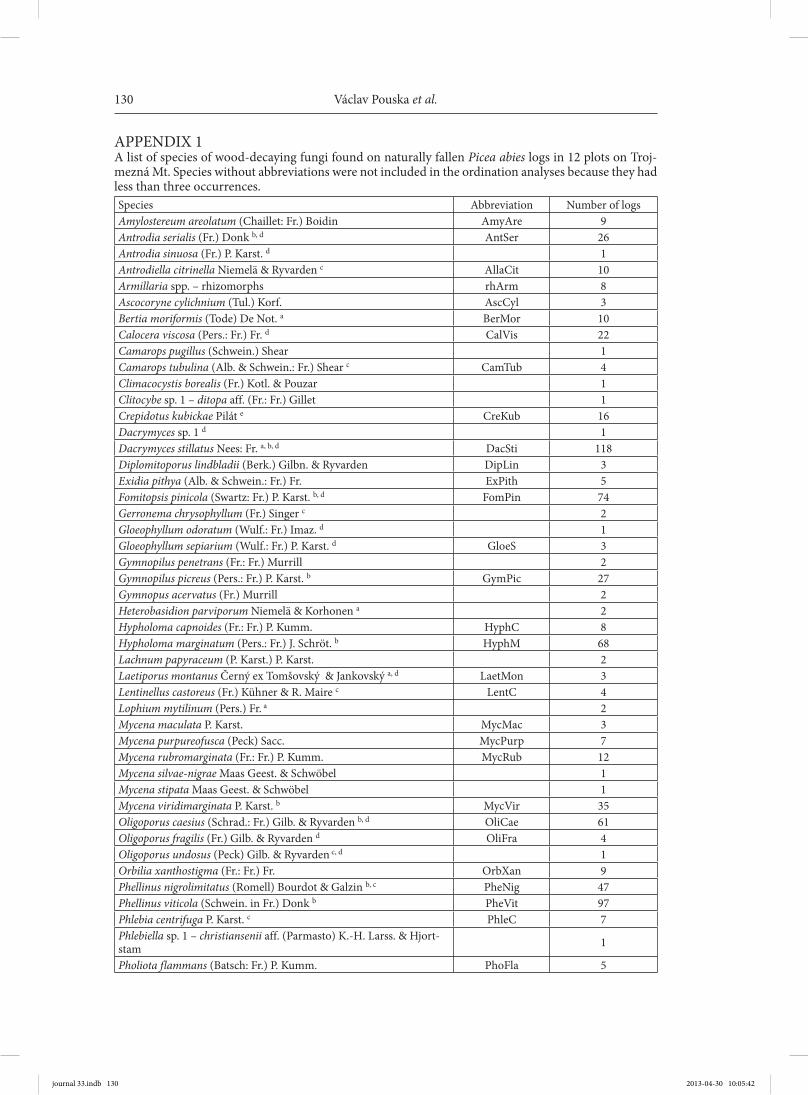
examined for the presence of sporocarps, though branches were not. Dead sporocarps were omitted, including dead annual sporo-carps that could not be identified. Records from all visits were pooled. For the most part we did not identify thin corticioid sporocarps (Corticiaceae s.l.) or Galerina spp., and these groups were not included in the study. The list of species is in Appendix 1 and other details in Pouska et al. (2010, 2011). Species names follow: Horak (2005) for agarics, Ryvarden and Gi lber tson (1993, 1994) for polypores, Hansen and Knudsen (1997) for other aphyllophoroid fungi, Hansen and Knud-sen (2000) for ascomycetes, and Index Fun-gorum. We assigned red-listed species ac-cording to Holec and B eran (2006).
2.4. Data analysis
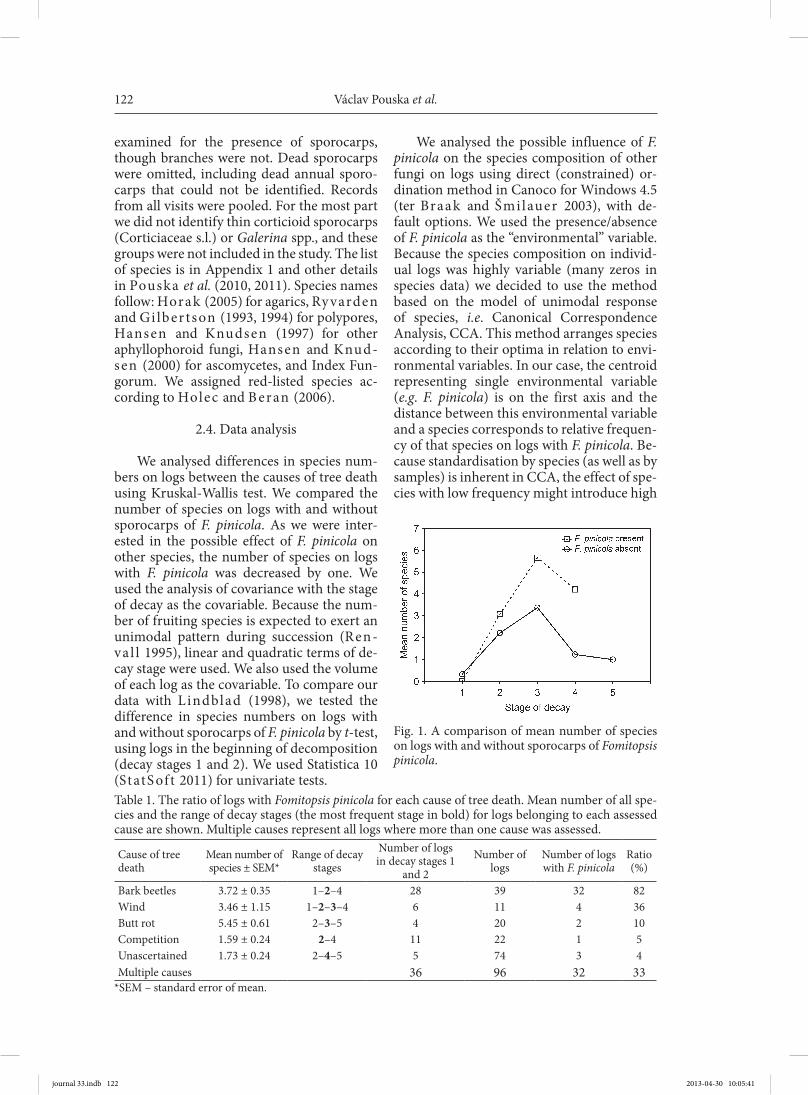
We analysed differences in species num-bers on logs between the causes of tree death using Kruskal-Wallis test. We compared the number of species on logs with and without sporocarps of F. pinicola. As we were inter-ested in the possible effect of F. pinicola on other species, the number of species on logs with F. pinicola was decreased by one. We used the analysis of covariance with the stage of decay as the covariable. Because the num-ber of fruiting species is expected to exert an unimodal pattern during succession (Ren-va l l 1995), linear and quadratic terms of de-cay stage were used. We also used the volume of each log as the covariable. To compare our data with L indblad (1998), we tested the difference in species numbers on logs with and without sporocarps of F. pinicola by t-test, using logs in the beginning of decomposition (decay stages 1 and 2). We used Statistica 10 (StatS of t 2011) for univariate tests.
We analysed the possible influence of F. pinicola on the species composition of other fungi on logs using direct (constrained) or-dination method in Canoco for Windows 4.5 (ter Braak and Šmi lauer 2003), with de-fault options. We used the presence/absence of F. pinicola as the “environmental” variable. Because the species composition on individ-ual logs was highly variable (many zeros in species data) we decided to use the method based on the model of unimodal response of species, i.e. Canonical Correspondence Analysis, CCA. This method arranges species according to their optima in relation to envi-ronmental variables. In our case, the centroid representing single environmental variable (e.g. F. pinicola) is on the first axis and the distance between this environmental variable and a species corresponds to relative frequen-cy of that species on logs with F. pinicola. Be-cause standardisation by species (as well as by samples) is inherent in CCA, the effect of spe-cies with low frequency might introduce high
Table 1. The ratio of logs with Fomitopsis pinicola for each cause of tree death. Mean number of all spe-cies and the range of decay stages (the most frequent stage in bold) for logs belonging to each assessed cause are shown. Multiple causes represent all logs where more than one cause was assessed.
Cause of tree death
Mean number of species ± SEM*
Range of decay stages
Number of logs in decay stages 1
and 2
Number of logs
Number of logs with F. pinicola
Ratio (%)
Bark beetles 3.72 ± 0.35 1–2–4 28 39 32 82Wind 3.46 ± 1.15 1–2–3–4 6 11 4 36Butt rot 5.45 ± 0.61 2–3–5 4 20 2 10Competition 1.59 ± 0.24 2–4 11 22 1 5Unascertained 1.73 ± 0.24 2–4–5 5 74 3 4Multiple causes 36 96 32 33
*SEM – standard error of mean.
Fig. 1. A comparison of mean number of species on logs with and without sporocarps of Fomitopsis pinicola.
122
journal 33.indb 122 2013-04-30 10:05:41

Co-occurrence of wood-decaying fungi on Picea abies
levels of noise into the results. To avoid this, we omitted species with less than three occur-rences. Finally, (in addition to F. pinicola) 37 species (713 records) on 220 logs were used in the analysis. Since the species composition changes mainly with the progression in decay, we used the stage of decay as a covariable; and we used the volume of logs and the causes of tree death as covariables because they also in-fluence species composition (Pouska et al. 2011). We were interested in the variability among individual logs and we already knew that there are differences among plots (Pous-ka et al. 2010). Therefore, to separate out the effect of individual plots, we used the location of logs in plots as a set of covariables in CCA (one indicator variable per plot). We tested the significance using a Monte-Carlo permu-tation test with 999 permutations in blocks defined by covariables representing plots.
We additionally did the same procedure (analysis of covariance and CCA) with all species that occurred on more than 10 % of logs (Appendix 1).
3. RESULTS
Altogether, there were 61 species (819 re-cords) of wood-decaying fungi on 262 logs of natural origin. Two species, Dacrymyces stilla-tus Nees and Phellinus viticola (Schwein. in Fr.) Donk, were more frequent than F. pinicola (Appendix 1). We found the sporocarps of F. pinicola on 74 out of a total of 262 logs and we assessed all distinguished causes of tree death for these 74 logs. F. pinicola was most common on logs that originated from trees killed by bark beetles and these logs were the
most common among logs from recently dead trees; see also the ratio of logs with F. pinicola in each category of tree death (Table 1). Based on Kruskal-Wallis test, numbers of fungal spe-cies on logs differed between the causes of tree death (only logs with a single assessed type tested, P <0.0001, n = 166). Multiple compari-sons of mean ranks of species numbers showed that wind did not differ from any other cause; bark beetles and butt rot differed from compe-tition and unascertained. The volume of logs increased the number of species (linear regres-sion: R2 = 0.46, P <0.0001, n = 262).
The maximum number of species found on a single log was 16: Bertia moriformis (Tode) De Not., D. stillatus, Diplomitoporus lindbladii (Berk.) Gilbn. & Ryvarden, Gerro-nema chrysophyllum (Fr.) Singer, Hypholoma marginatum (Pers.: Fr.) J. Schröt., Lophium mytilinum (Pers.) Fr., Mycena purpureofusca (Peck) Sacc., Mycena rubromarginata (Fr.: Fr.) P. Kumm., Mycena viridimarginata P. Karst., Oligoporus caesius (Schrad.: Fr.) Gilb. & Ryvarden, Orbilia xanthostigma (Fr.: Fr.) Fr., P. viticola, Pholiota flammans (Batsch: Fr.) P. Kumm., Physisporinus vitreus (Pers.: Fr.) P. Karst., Pluteus pouzarianus Singer and Veluti-ceps abietina (Pers.: Fr.) Hjortstam & Tellería; this was one of the largest logs (7 m3), decay stage 3, without bark, broken from a snag that was affected by the white rot caused by Cli-macocystis borealis (Fr.) Kotl. & Pouzar (butt rot). The next highest number of species was 12, found on two different logs in decay stage 3, both hosted Antrodia serialis (Fr.) Donk, B. moriformis, F. pinicola, H. marginatum, P. viticola and Stropharia aeruginosa (M.A. Cur-tis: Fr.) Quél.
Table 2. The results of the analysis of covariance – the effects (significance level) of the presence/absence of the tested species, volume of logs, decay stage and decay stage squared on the number of the other species on logs are shown.
Species Effect of species Volume of logs Decay stage Decay stage squared
Antrodia serialis <0.0001 <0.0001 0.0001 <0.0001Dacrymyces stillatus <0.0001 <0.0001 <0.0001 <0.0001Fomitopsis pinicola 0.032 <0.0001 <0.0001 <0.0001Gymnopilus picreus 0.030 <0.0001 <0.0001 <0.0001Hypholoma marginatum <0.0001 <0.0001 0.0003 <0.0001Mycena viridimarginata 0.82 <0.0001 <0.0001 <0.0001Oligoporus caesius 0.008 <0.0001 <0.0001 <0.0001Phellinus nigrolimitatus 0.35 <0.0001 <0.0001 <0.0001Phellinus viticola <0.0001 <0.0001 0.001 0.0005
123
journal 33.indb 123 2013-04-30 10:05:41

Václav Pouska et al.
Logs in decay stages 1 and 2 with the sporocarps of F. pinicola had more species (mean 3.03, 40 logs) than all other logs in the same decay stages (mean 2.1, 50 logs), t-test (P = 0.018). Analysis of covariance with the volume of logs and the stage of decay in lin-ear and quadratic term as covariables showed significant effects of all covariables and the presence of F. pinicola on species numbers (Table 2), viz. logs with the sporocarps of F. pinicola were more species rich (Fig. 1).
Based on the analysis of covariance (Table 2), the presence of any of the following species significantly increased the number of the other species: A. serialis, D. stillatus, Gym-nopilus picreus (Pers.: Fr.) P. Karst., H. margin-atum, O. caesius and P. viticola. The presence of M. viridimarginata or Phellinus nigrolimita-tus (Romell) Bourdot & Galzin did not influ-ence the number of other species on logs.
The species composition on logs was influenced by F. pinicola. The first canoni-cal axis was significant in all cases, i.e. after including decay stage and volume as covari-ables (P = 0.001), causes of tree death as co-variables (P = 0.040), or everything together as covariables (P = 0.045). The results of CCA (Fig. 2, Appendix 2) show that some species were relatively common on logs with this polypore, e.g. G. picreus. Red-listed species Antrodiella citrinella Niemelä & Ryvarden and Camarops tubulina (Alb. & Schwein.: Fr.) Shear regularly co-occurred with F. pinicola.
When tested with all covariables in CCA (Appendix 2), A. serialis (P = 0.025), D. stilla-tus (P = 0.001), G. picreus (P = 0.024), H. mar-ginatum (P = 0.004) and P. viticola (P = 0.001) had significant influence on the species com-position, whereas M. viridimarginata, O. cae-sius (P = 0.063) and P. nigrolimitatus did not. Some of the species positively associated with F. pinicola were associated also with the other species: M. rubromarginata with D. stillatus; C. tubulina with D. stillatus, G. picreus or H. marginatum. Negative relationships seemed to have e.g. Trichaptum abietinum (J. Dicks.) Ryvarden with A. serialis or with P. viticola. Among the common species, positive rela-tionships had e.g. A. serialis with G. picreus or P. viticola.
4. DISCUSSION
F. pinicola can be a primary mortality agent of trees (L ännenpää et al. 2008), but we did not observe this effect. However, in accordance with our preliminary survey of snags, the occurrence of F. pinicola on logs was often associated with another mortality agent – bark beetles (see also Bače et al. 2012). Bark beetles can transfer F. pinicola and facilitate its entering into the wood through perforations in the bark. F. pinicola was isolated to culture from bark beetles in North America (Castel -lo et al. 1976, Pettey and Shaw 1986, L im et al. 2005) and it was found also on bark bee-
Fig. 2. CCA ordination diagram showing the affinity of some fungal species to Fomitopsis pinicola. Only species with the highest amount of variability explained by the first ordination axis are shown. Decay stage, volume and causes of tree death were set as covariables.
124
journal 33.indb 124 2013-04-30 10:05:41

Co-occurrence of wood-decaying fungi on Picea abies
tles from P. abies in Sweden (Persson et al. 2009, Persson et al. 2011). Among them, Ips typographus L., Pityogenes chalcographus L. and Crypturgus cinereus Hbst. were abun-dant bark beetles in the Bohemian Forest (Röder et al. 2010). The occurrence of F. pinicola sporocarps was positively associated with another bark beetle – Hylurgops palliatus Gyll. in high stumps in Sweden, but not with I. typographus or P. chalcographus (Wesl ien et al. 2011). H. palliatus was not abundant in the Bohemian Forest (Röder et al. 2010). Perhaps, the presence of basidiomycetes on bark beetles has been underestimated. Stud-ies dealing with bark-beetle associated fungi are usually not focused on basidiomycetes but it is interesting that the above cited investiga-tions done on media suitable for basidiomy-cetes or using cultivation independent tech-niques showed the presence of several species including F. pinicola.
F. pinicola most likely influences the oc-currence of other fungi on spruce logs. In contrast to the results of L indblad (1998), logs in the beginning of decomposition (our decay stages 1 and 2) with the sporocarps of F. pinicola had more species than other logs in the same decay stages. If all logs are included and the volume of log and the stage of decay are set as covariables, the logs with the spo-rocarps of F. pinicola still have more species than other logs. This is not in contradiction with the result of L indner et al. (2011) re-garding sporocarps, but their results based on culturing on media or sequencing ITS rDNA showed that logs inoculated with F. pinicola had fewer species than other logs. However, these results come from logs that were let to decompose six years and it is possible that the pattern would change later in more decom-posed wood. The presence of several other fruiting species also influenced species num-bers; the tested species had either positive or no influence on the number of other species; a negative influence was not detected. In ad-dition, the presence of F. pinicola or of several other tested species significantly influenced species composition. However, G. picreus had a significant influence on both species rich-ness and composition and it may not be a primary decayer. Likewise, it does not seem probable that D. stillatus would influence other more dominant species. We are aware
that our results cannot be interpreted as real causal relationships.
A possible reason behind the statisti-cal effect of a certain species’ presence on the richness and composition of other spe-cies could be their prevalent common oc-currence on logs that were more favourable than other logs. O vaskainen et al. (2010) found that several species occurred especially on large logs in an intermediate decay stage and thus differences in the overall suitability of logs generated positive correlations among the species. However, we used similar char-acteristics, i.e. the volume of logs and the stage of decay, as covariables. Species rich-ness (Kruskal-Wallis test result) and com-position (Pouska et al. 2011) are also cor-related with diverse causes of tree death (that can hardly be assessed fully). We used several covariables to partial out the possible effect of log properties and to reveal outcomes of interactions among the species. Neverthe-less, we may have missed some log properties that have similar effect on the fungi and their fruiting, and such effect can possibly simulate species interactions. The statistical effects do not provide a direct proof of causal relation-ships. Interactions of fungi are likely to play a key role in co-occurrence pattern, but it is hardly possible to unequivocally prove them without their manipulation (e.g. introduction of some species). Experiments of Fukami et al. (2010) showed that species diversity was influenced by a fungus that was inoculated in advance of the other fungal species, but this was done on small wood discs in vitro.
F. pinicola is not a strong competitor (Holmer and Stenl id 1997, Holmer et al. 1997, Iakovlev and Stenl id 2000) and our results indicate that its presence often in-creased the number of species on logs. This may be possible if F. pinicola does not occupy the logs entirely. On the basis of our obser-vation, if whole logs are brown-rotten, not many other species grow there. This is in con-cordance with the finding that the presence of F. pinicola was negatively correlated with several species (O vaskainen et al. 2010). Interactions of some fungi that rarely occur together may not necessarily be antagonis-tic, since they occupy different types of logs. It was shown that the occurrence of fungi is more or less associated with the origin of
125
journal 33.indb 125 2013-04-30 10:05:41

Václav Pouska et al.
dead wood, i.e. mortality type and the way the log fell down (Renval l 1995, Pouska et al. 2011). Regarding other tested species, P. viticola may be a strong competitor (Holmer and Stenl id 1997) but it also increased the number of species.
Regardless of the positive or negative in-fluence of F. pinicola on the richness of spe-cies growing in logs, its influence on species composition is very likely (e.g. Niemelä et al. 1995, O vaskainen et al. 2010, L ind-ner et al. 2011). Renval l (1995), Yl is irniö et al. (2009) and O vaskainen et al. (2010) found T. abietinum positively correlated with F. pinicola on logs. In contrast, Wesl ien et al. (2011) found a negative effect of T. abieti-num on F. pinicola on high stumps. We found T. abietinum relatively often on the same logs as F. pinicola (13 versus seven occurrences), but we did not find any relationship by the ordination analyses. T. abietinum, similarly as F. pinicola, prevalently occurs on logs from trees killed by bark beetles, but often on tree crowns (Renval l 1995, Pouska et al. 2011). Phlebia centrifuga P. Karst. (red-listed) was also reported as a successor (Renval l 1995), but it seems to have a negative relationship at our locality (Appendix 2; we found it with F. pinicola on only one out of seven logs). L indblad (1998) also found T. abietinum more often on logs with F. pinicola, but not P. centrifuga. O vaskainen et al. (2010) fur-ther found F. pinicola positively correlated with A. serialis and negatively with O. caesius, P. nigrolimitatus (red-listed) and P. viticola (see also Yl is irniö et al. 2009). Accordingly, our results of ordination analyses (Appendix 2) indicate that P. nigrolimitatus may have a negative relationship with F. pinicola. White rot-causing A. citrinella (red-listed) grows in brown-rotten wood following F. pinicola; it is a well known trait of this species (Ryvarden and Gi lber tson 1993, p. 152). Three out of the ten trees with A. citrinella most prob-ably died due to bark beetles, one due to butt rot, and two were windthrows. Red-listed C. tubulina grows on three species of trees – P. abies, A. alba and Fagus sylvatica L. (Holec 2005). Interestingly, all four finds of this fun-gus were on logs with both F. pinicola and A. citrinella. Regarding the other tested spe-cies, O vaskainen et al. (2010) also found a positive correlation between P. viticola and
A. serialis, and negative correlation between P. viticola and T. abietinum (also Yl is irniö et al. 2009).
Bäss ler and Mül ler (2010) concluded that A. citrinella dispersed rapidly in areas infested by bark beetles. Dead wood created by bark beetles favoured F. pinicola and con-sequently also A. citrinella. However, large pulses of dead wood can occur only in long time periods within a certain area and this may have only temporary positive effect on threatened fungi. The decomposition of dead wood originating from a disturbance leads to relatively low volume until new dead wood comes from next generation trees (e.g. Spies et al. 1988, Si itonen 2001, Kruys et al. 2002, Aakala 2010). Moreover, various causes of tree mortality are needed to provide the appropriate habitat for different red-listed species (Pouska et al. 2011). Therefore, dead wood of one type prevailing in a certain area may not be suitable for all threatened fungi (Bässler et al. 2012). Disturbances due to wind or bark beetles may influence the diver-sity of fungi both positively and negatively, depending on time and spatial scale.
ACKNOWLEDGEMENTS: We thank M. Kolařík for his opinion about bark-beetle medi-ated transport of fungi. This study was supported by the Ministry of Education (MSM6007665801). M. S. was supported by the National Agency for Agriculture Research (QI102A085).
5. REFERENCES
Aakala T. 2010 – Coarse woody debris in late-successional Picea abies forests in northern Europe: Variability in quantities and models of decay class dynamics – For. Ecol. Manage. 260: 770–779.
Abrahamsson M. , L indbladh M. , Rönn-berg J. 2008 – Influence of butt rot on beetle diversity in artificially created high-stumps of Norway spruce – For. Ecol. Manage. 255: 3396–3403.
Bače R . , Svoboda M. , Pouska V. , Janda P. , Č er venka J. 2012 – Natural regenera-tion in Central-European subalpine spruce forests: Which logs are suitable for seedling re-cruitment? – For. Ecol. Manage. 266: 254–262.
Bässler C. , Mül ler J. 2010 – Importance of natural disturbance for recovery of the rare polypore Antrodiella citrinella Niemelä & Ryvarden – Fungal Biol. 114: 129–133.
126
journal 33.indb 126 2013-04-30 10:05:41

Co-occurrence of wood-decaying fungi on Picea abies
Bässler C. , Mül ler J. , Svoboda M. , Lepšová A. , Hahn C. , Holzer H. , Pouska V. 2012 – Diversity of wood-decay-ing fungi under different disturbance regimes – a case study from spruce mountain forests – Biodivers. Conserv. 21: 33–49.
B oddy L. 2000 – Interspecific combative inter-actions between wood-decaying basidiomyce-tes – FEMS Microbiol. Ecol. 31: 185–194.
Castel lo J.D. , Shaw C.G. , Furniss M.M. 1976 – Isolation of Cryptoporus volvatus and Fomes pinicola from Dendroctonus pseudotsug-ae – Phytopathology, 66: 1431–1434.
Dighton J. , Thomas E.D. , L atter P.M. 1987 – Interactions between tree roots, mycor-rhizas, a saprotrophic fungus and the decom-position of organic substrates in a microcosm – Biol. Fertil. Soils, 4: 145–150.
Edman M. , Jönsson M. , Jonsson B.G. 2007 – Fungi and wind strongly influence the temporal availability of logs in an old-growth spruce forest – Ecol. Appl. 17: 482–490.
Fukami T. , Dickie I .A. , Wi lk ie J.P. , Pau-lus B.C. , Park D. , Rober ts A. , Buchan-an P.K. , Al len R .B. 2010 – Assembly his-tory dictates ecosystem functioning: evidence from wood decomposer communities – Ecol. Letters, 13: 675–684.
Fukasawa Y. , Osono T. , Takeda H. 2009 – Dynamics of physicochemical properties and occurrence of fungal fruit bodies during de-composition of coarse woody debris of Fagus crenata. – J. For. Res. 14: 20–29.
Hansen L. , Knudsen H. (eds . ) 1997 – Nor-dic Macromycetes, Vol. 3, Heterobasidioid,
Aphyl lophoroid and Gasteromycetoid Basidiomycetes – Nordsvamp, C open-hagen, 444 pp.
Hansen L. , Knudsen H. (eds . ) 2000 – Nor-dic Macromycetes, Vol. 1, Ascomycetes – Nor-dsvamp, Copenhagen, 309 pp.
Hei lmann-Clausen J. 2001 – A gradient anal-ysis of communities of macrofungi and slime moulds on decaying beech logs – Mycol. Res. 105: 575–596.
Høi land K. , B endiksen E. 1997 – Biodiver-sity of wood-inhabiting fungi in a boreal co-niferous forest in Sør-Trøndelag County, Cen-tral Norway – Nordic J. Bot. 16: 643–659.
Holec J. 2005 – Distribution and ecology of Camarops tubulina (Ascomycetes, Bolinia-ceae) in the Czech Republic and remarks on its European distribution – Czech Mycol. 57: 97–115.
Holec J. , B eran M. (eds . ) . 2006 – Červený seznam hub (makromycetů) České repub-liky [Red List of fungi (macromycetes) of the
Czech Republic] – Příroda, Praha 24: 1–282 (in Czech, English summary).
Holmer L . , Stenl id J. 1997 – Competitive hierarchies of wood decomposing basidiomy-cetes in artificial systems based on variable in-oculum sizes – Oikos, 79: 77–84.
Holmer L . , Renval l P. , Stenl id J. 1997 – Se-lective replacement between species of wood-rotting basidiomycetes, a laboratory study – Mycol. Res. 101: 714–720.
Horak E. 2005 – Röhrlinge und Blätterpilze in Europa. 6. Auflage – Elsevier, Spektrum Aka-demischer Verlag, München, 555 pp.
Iakovlev A. , Stenl id J. 2000 – Spatiotemporal patterns of laccase activity in interacting my-celia of wood-decaying basidiomycete fungi – Microb. Ecol. 39: 236–245.
Index Fungorum – URL http : / /www.in-dexfungorum.org .
Jonsel l M. , Nordlander G. 2002 – Insects in polypore fungi as indicator species: a compari-son between forest sites differing in amounts and continuity of dead wood – For. Ecol. Man-ag. 157: 101–118.
Jonsel l M. , Wesl ien J. 2003 – Felled or stand-ing retained wood – it makes a difference for saproxylic beetles – For. Ecol. Manage. 175: 425–435.
Jonsel l , M. , S chroeder, M. , Wesl ien J. 2005 – Saproxylic beetles in high stumps of spruce: Fungal flora important for determin-ing the species composition – Scand. J. For. Res. 20: 54–62.
Kauserud H., Colman J.E., Ryvarden L. 2008 – Re-lationship between basidiospore size, shape and life history characteristics: a comparison of polypores – Fungal Ecol. 1: 19–23.
Kot laba F. 1984 – Zeměpisné rozšíření a ekolo-gie chorošů (Polyporales s.l.) v Československu [Geographical distribution and ecology of polypores (Polyporales s. l.) in Czechoslova-kia] – Academia, Praha (in Czech, English summary), 194 pp.
Kruys N. , Jonsson B.G. , Ståhl G. 2002 – A stage-based matrix model for decay-class dynamics of woody debris – Ecol. Appl. 12: 773–781.
L ännenpää A. , Aakala T. , Kauhanen H. , Kuuluvainen T. 2008 – Tree mortality agents in pristine Norway spruce forests in northern Fennoscandia – Silva Fennica, 42: 151–163.
Lim Y.W., Kim J. J. , Lu M. , Breui l C. 2005 – Determining fungal diversity on Dendroc-tonus ponderosae and Ips pini affecting lodge-pole pine using cultural and molecular meth-ods – Fungal Divers. 19: 79–94.
127
journal 33.indb 127 2013-04-30 10:05:42

Václav Pouska et al.
Lindahl B. , Stenl id J. , Olsson S . , F in lay R . 1999 – Translocation of 32P between inter-acting mycelia of a wood-decomposing fungus and ectomycorrhizal fungi in microcosm sys-tems – New Phytol. 144: 183–193.
Lindblad I . 1998 – Wood-inhabiting fungi on fallen logs of Norway spruce: relations to for-est management and substrate quality – Nor-dic J. Bot. 18: 243–255.
Lindner D.L. , Vasait is R . , Kubar tová A. , Al lmér J. , Johannesson H. , Banik M.T. , Stenl id J. 2011 – Initial fungal colo-nizer affects mass loss and fungal community development in Picea abies logs 6 yr after in-oculation – Fungal Ecol. 4: 449–460.
Mül ler J. , Bußler H. , Goßner M. , Rette l-bach T. , Duel l i P. 2008 – The European spruce bark beetle Ips typographus in a nation-al park: from pest to keystone species – Bio-divers. Conserv. 17: 2979–3001.
Mül ler M.M., Varama M. , Heinonen J. , Hal laksela A.-M. 2002 – Influence of in-sects on the diversity of fungi in decaying spruce wood in managed and natural forests – For. Ecol. Manage. 166: 165–181.
Neuhäuslová Z. (ed.) 2001 – The map of po-tential natural vegetation of the Šumava Na-tional Park – Silva Gabreta, Supplementum 1.
Niemelä T. , Renval l P. , Pentt i lä R . 1995 – Interactions of fungi at late stages of wood decomposition – Annales Botanici Fennici, 32: 141–152.
Nordén B. 1997 – Genetic variation within and among populations of Fomitopsis pinicola (Ba-sidiomycetes) – Nordic J. Bot. 17: 319–329.
O vaskainen O. , Hottola J. , S i itonen J. 2010 – Modeling species co-occurrence by multivariate logistic regression generates new hypotheses on fungal interactions – Ecology, 91: 2514–2521.
Persson Y. , Ihrmark K. , Stenl id J. 2011 – Do bark beetles facilitate the establishment of rot fungi in Norway spruce? – Fungal Ecol. 4: 262–269.
Persson Y. , Vasait is R . , L ångström B. , Öhrn P. , Ihrmark K. , Stenl id J. 2009 – Fungi vectored by the bark beetle Ips typogra-phus following hibernation under the bark of standing trees and in the forest litter – Microb. Ecol. 58: 651–659.
Pettey T.M., Shaw C.G. 1986 – Isolation of Fomitop-sis pinicola from in-flight bark beetles (Coleop-tera: Scolytidae) – Can. J. Bot. 64: 1507–1509.
Pouska V. , Svoboda M. , Lepšová A. 2010 – The diversity of wood-decaying fungi in re-lation to changing site conditions in an old-growth mountain spruce forest, Central Eu-rope – Eur. J. For. Res. 129: 219–231.
Pouska V. , Lepš J. , Svoboda M. , Lepšová A. 2011 – How do log characteristics influ-ence the occurrence of wood fungi in a moun-tain spruce forest? – Fungal Ecol. 4: 201–209.
R aja la T. , Peltoniemi M. , Hantula J. , Mäkipää R . , Pennanen T. 2011 – RNA reveals a succession of active fungi during the decay of Norway spruce logs – Fungal Ecol. 4: 437–448.
Renval l P. 1995 – Community structure and dynamics of wood-rotting Basidiomycetes on decomposing conifer trunks in northern Fin-land – Karstenia, 35: 1–51.
Röder J. , Bäss ler C. , Brandl R . , D vořák L. , F loren A. , Goßner M.M., Gruppe A. , Jarzabek-Mül ler A. , Vojtěch O. , Wagner C. , Mül ler J. 2010 – Arthropod species richness in the Norway Spruce (Picea abies (L.) Karst.) canopy along an elevation gradient – For. Ecol. Manage. 259: 1513–1521.
Ryvarden L. , Gi lber tson R .L. 1993 – Eu-ropean Polypores. Part 1: 1–387. Abortiporus – Lindtneria – Synopsis Fungorum 6, Fungi-flora, Oslo.
Ryvarden L. , Gi lber tson R .L. 1994 – Eu-ropean Polypores. Part 2: 388–743. Meripilus – Tyromyces – Synopsis Fungorum 7, Fungi-flora, Oslo.
Si itonen J. 2001 – Forest management, coarse woody debris and saproxylic organisms: Fen-noscandian boreal forests as an example – Ecological Bulletins, 49: 11–41.
Sippola A.-L. , Renval l P. 1999 – Wood-decomposing fungi and seed tree cutting: A 40-year perspective – For. Ecol. Manag. 115: 183–201.
Spies T.A. , Frankl in J.F. , Thomas T.B. 1988 – Coarse woody debris in Douglas-fir forests of western Oregon and Washington – Ecology, 69: 1689–1702.
Spir in V.A. , Shirokov A.I . 2002 – The fea-tures of the wood-decay in the virgin Abies-Fir forests from Nizhny Novgorod region – Miko-logia i fitopatologia, 36: 25–31 (in Russian, English summary).
StatS of t , Inc . 2011. Statistica (data analysis software system), version 10. www.statsoft.com.
Svoboda M. , Pouska V. 2008 – Structure of a Central-European mountain spruce old-growth forest with respect to historical de-velopment – Forest Ecol. Manage. 255: 2177–2188.
Tedersoo L. , Kõlja lg U. , Hal lenberg N. , L arsson K.-H. 2003 – Fine scale distribu-tion of ectomycorrhizal fungi and roots across substrate layers including coarse woody debris in a mixed forest – New Phytol. 159: 153–165.
128
journal 33.indb 128 2013-04-30 10:05:42

Co-occurrence of wood-decaying fungi on Picea abies
ter Braak C.J.F. , Šmi lauer P. 2003 – Canoco 4.5. Biometris, Plant Research International, Wageningen, Netherlands.
Vasi l iauskas R . , Stenl id J. , Thomsen I .M. 1998 – Clonality and genetic variation in Am-ylostereum areolatum and A. chailletii from northern Europe – New Phytol. 139: 751–758.
Wesl ien J. , Djupström L.B. , S chroeder M. , Widenfa lk O. 2011 – Long-term prior-ity effects among insects and fungi colonizing decaying wood – J. Anim. Ecol. 80: 1155–1162.
Worra l l J. J. , Lee T.D. , Harr ington T.C. 2005 – Forest dynamics and agents that initiate and expand canopy gaps in Picea-Abies forests of Crawford Notch, New Hampshire, USA – J. Ecol. 93: 178–190.
Ylisirniö A.-L., Berglund H., Aakala T., Kuulu-vainen T., Kuparinen A.-M., Norokorpi Y., Hallikainen V., Mikkola K., Huhta E. 2009 – Spatial distribution of dead wood and the oc-currence of five saproxylic fungi in old-growth timberline forests in northern Finland – Scand. J. For. Res. 24: 527–540.
Received after revision May 2012
129
journal 33.indb 129 2013-04-30 10:05:42
Václav Pouska et al.
APPENDIX 1A list of species of wood-decaying fungi found on naturally fallen Picea abies logs in 12 plots on Troj-mezná Mt. Species without abbreviations were not included in the ordination analyses because they had less than three occurrences.Species Abbreviation Number of logsAmylostereum areolatum (Chaillet: Fr.) Boidin AmyAre 9Antrodia serialis (Fr.) Donk b, d AntSer 26Antrodia sinuosa (Fr.) P. Karst. d 1Antrodiella citrinella Niemelä & Ryvarden c AllaCit 10Armillaria spp. – rhizomorphs rhArm 8Ascocoryne cylichnium (Tul.) Korf. AscCyl 3Bertia moriformis (Tode) De Not. a BerMor 10Calocera viscosa (Pers.: Fr.) Fr. d CalVis 22Camarops pugillus (Schwein.) Shear 1Camarops tubulina (Alb. & Schwein.: Fr.) Shear c CamTub 4Climacocystis borealis (Fr.) Kotl. & Pouzar 1Clitocybe sp. 1 – ditopa aff. (Fr.: Fr.) Gillet 1Crepidotus kubickae Pilát e CreKub 16Dacrymyces sp. 1 d 1Dacrymyces stillatus Nees: Fr. a, b, d DacSti 118Diplomitoporus lindbladii (Berk.) Gilbn. & Ryvarden DipLin 3Exidia pithya (Alb. & Schwein.: Fr.) Fr. ExPith 5Fomitopsis pinicola (Swartz: Fr.) P. Karst. b, d FomPin 74Gerronema chrysophyllum (Fr.) Singer c 2Gloeophyllum odoratum (Wulf.: Fr.) Imaz. d 1Gloeophyllum sepiarium (Wulf.: Fr.) P. Karst. d GloeS 3Gymnopilus penetrans (Fr.: Fr.) Murrill 2Gymnopilus picreus (Pers.: Fr.) P. Karst. b GymPic 27Gymnopus acervatus (Fr.) Murrill 2Heterobasidion parviporum Niemelä & Korhonen a 2Hypholoma capnoides (Fr.: Fr.) P. Kumm. HyphC 8Hypholoma marginatum (Pers.: Fr.) J. Schröt. b HyphM 68Lachnum papyraceum (P. Karst.) P. Karst. 2Laetiporus montanus Černý ex Tomšovský & Jankovský a, d LaetMon 3Lentinellus castoreus (Fr.) Kühner & R. Maire c LentC 4Lophium mytilinum (Pers.) Fr. a 2Mycena maculata P. Karst. MycMac 3Mycena purpureofusca (Peck) Sacc. MycPurp 7Mycena rubromarginata (Fr.: Fr.) P. Kumm. MycRub 12Mycena silvae-nigrae Maas Geest. & Schwöbel 1Mycena stipata Maas Geest. & Schwöbel 1Mycena viridimarginata P. Karst. b MycVir 35Oligoporus caesius (Schrad.: Fr.) Gilb. & Ryvarden b, d OliCae 61Oligoporus fragilis (Fr.) Gilb. & Ryvarden d OliFra 4Oligoporus undosus (Peck) Gilb. & Ryvarden c, d 1Orbilia xanthostigma (Fr.: Fr.) Fr. OrbXan 9Phellinus nigrolimitatus (Romell) Bourdot & Galzin b, c PheNig 47Phellinus viticola (Schwein. in Fr.) Donk b PheVit 97Phlebia centrifuga P. Karst. c PhleC 7Phlebiella sp. 1 – christiansenii aff. (Parmasto) K.-H. Larss. & Hjort-stam 1
Pholiota flammans (Batsch: Fr.) P. Kumm. PhoFla 5
130
journal 33.indb 130 2013-04-30 10:05:42
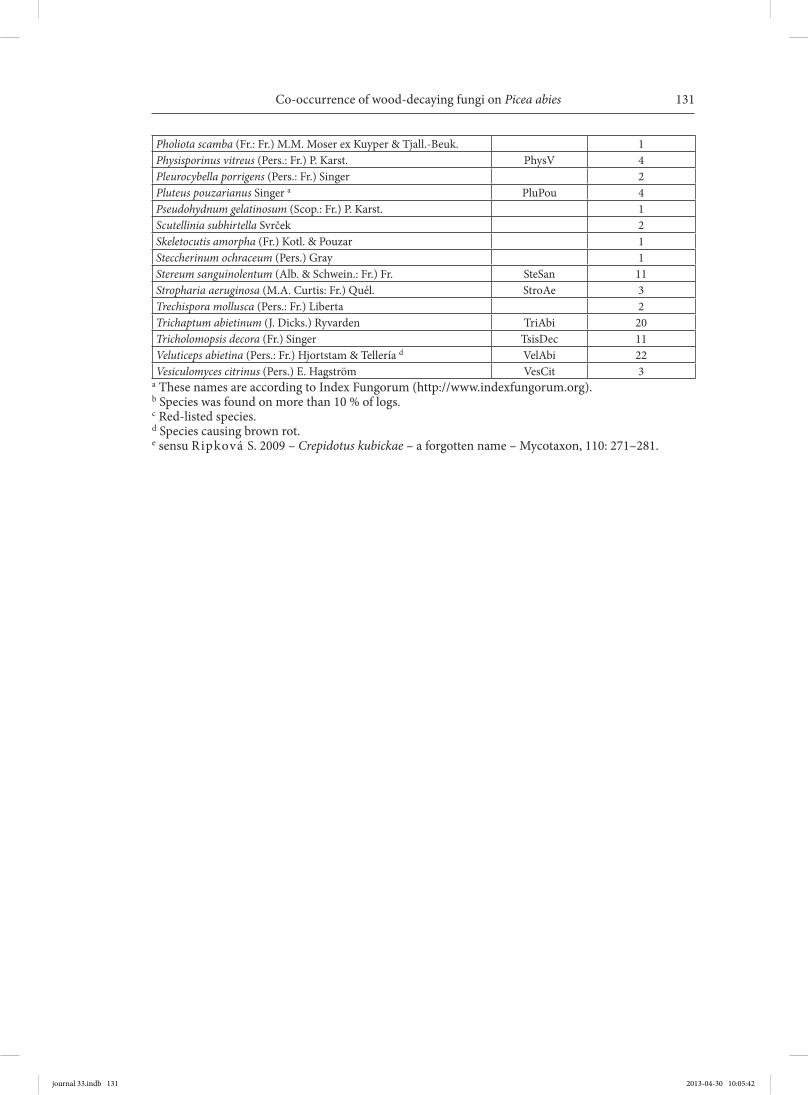
Co-occurrence of wood-decaying fungi on Picea abies
Pholiota scamba (Fr.: Fr.) M.M. Moser ex Kuyper & Tjall.-Beuk. 1Physisporinus vitreus (Pers.: Fr.) P. Karst. PhysV 4Pleurocybella porrigens (Pers.: Fr.) Singer 2Pluteus pouzarianus Singer a PluPou 4Pseudohydnum gelatinosum (Scop.: Fr.) P. Karst. 1Scutellinia subhirtella Svrček 2Skeletocutis amorpha (Fr.) Kotl. & Pouzar 1Steccherinum ochraceum (Pers.) Gray 1Stereum sanguinolentum (Alb. & Schwein.: Fr.) Fr. SteSan 11Stropharia aeruginosa (M.A. Curtis: Fr.) Quél. StroAe 3Trechispora mollusca (Pers.: Fr.) Liberta 2Trichaptum abietinum (J. Dicks.) Ryvarden TriAbi 20Tricholomopsis decora (Fr.) Singer TsisDec 11Veluticeps abietina (Pers.: Fr.) Hjortstam & Tellería d VelAbi 22Vesiculomyces citrinus (Pers.) E. Hagström VesCit 3
a These names are according to Index Fungorum (http://www.indexfungorum.org).b Species was found on more than 10 % of logs.c Red-listed species.d Species causing brown rot.e sensu Ripková S. 2009 – Crepidotus kubickae – a forgotten name – Mycotaxon, 110: 271–281.
131
journal 33.indb 131 2013-04-30 10:05:42

Václav Pouska et al.
APPENDIX 2
CCA ordination diagram showing the affinity of some fungal species to Fomitopsis pinicola. Decay stage and volume were set as covariables. Full names of species are in Appendix 1.
CCA ordination diagram showing the affinity of some fungal species to Fomitopsis pinicola. Causes of tree death were set as covariables.Below, CCA ordination diagrams showing the affinity of some fungal species to the species set as envi-ronmental variables. Only species with the highest amount of variability explained by the first ordina-tion axis are shown. Decay stage, volume and causes of tree death were set as covariables.
In addition to Antrodia serialis, 37 species (761 occurrences) were in the data matrix.
In addition to Dacrymyces stillatus, 37 species (668 occurrences) were in the data matrix.
132
journal 33.indb 132 2013-04-30 10:05:43

Co-occurrence of wood-decaying fungi on Picea abies
In addition to Gymnopilus picreus, 37 species (760 occurrences) were in the data matrix.
In addition to Hypholoma marginatum, 37 species (719 occurrences) were in the data matrix.
In addition to Phellinus viticola, 37 species (690 occurrences) were in the data matrix.
133
journal 33.indb 133 2013-04-30 10:05:43