chromium(iii) oxide nanoparticles induced remarkable oxidative stress and apoptosis on culture cells
TRANSCRIPT

Chromium(III) Oxide Nanoparticles Induced RemarkableOxidative Stress and Apoptosis on Culture Cells
Masanori Horie,1 Keiko Nishio,1 Shigehisa Endoh,2 Haruhisa Kato,3 Katsuhide Fujita,4
Arisa Miyauchi,2 Ayako Nakamura,3 Shinichi Kinugasa,3 Kazuhiro Yamamoto,5
Etsuo Niki,1 Yasukazu Yoshida,1 Hitoshi Iwahashi1
1Health Research Institute (HRI), National Institute of Advanced Industrial Science and Technology(AIST), 1-8-31 Midorigaoka, Ikeda, Osaka 563-8577, Japan
2Research Institute for Environmental Management Technology (EMTECH), AIST, PolymerStandards Section Japan (PSSJ), 16-1 Onogawa, Tsukuba, Ibaraki 305-8569, Japan
3National Metrology Institute of Japan (NMIJ), AIST, 1-1-1 Higashi, Tsukuba, Ibaraki 305-8565, Japan
4Research Institute of Science for Safety and Sustainability (RISS), AIST, 16-1 Onogawa, Tsukuba,Ibaraki 305-8569, Japan
5Research Institute of Instrumentation Frontier (RIIF), AIST, 1-1-1 Umezono, Tsukuba, Ibaraki 305-8565, Japan
Received 28 August 2010; revised 11 December 2010; accepted 18 December 2010
ABSTRACT: Chromium(III) oxide (Cr2O3) is used for industrial applications such as catalysts and pigments.In the classical form, namely the fine particle, Cr2O3 is insoluble and chemically stable. It is classified as alow-toxicity chromium compound. Recently, industrial application of nanoparticles (a new form composed ofsmall particles with a diameter of �100 nm, in at least one dimension) has been increasing. Cellular effectsinduced by Cr2O3 nanoparticles are not known. To shed light upon this, the release of soluble chromium fromCr2O3 nano- and fine-particles in culture medium was compared. Fine Cr2O3 particles were insoluble in theculture medium; on the contrary, Cr2O3 nanoparticles released soluble hexavalent chromium into the culturemedium. Cr2O3 nanoparticles showed severe cytotoxicity. The effect of Cr2O3 nanoparticles on cell viabilitywas higher than that of fine particles. Cr2O3 nanoparticles showed cytotoxicity equal to that of hexavalentchromium (K2Cr2O7). Human lung carcinoma A549 cells and human keratinocyte HaCaT cells showed anincrease in intracellular reactive oxygen species (ROS) level and activation of antioxidant defense systems onexposure to Cr2O3 nanoparticles. Exposure of Cr2O3 nanoparticles led to caspase-3 activation, showing thatthe decrease in cell viability by exposure to Cr2O3 nanoparticles was caused by apoptosis. Cellularresponses were stronger in the Cr2O3 nanoparticles-exposed cells than in fine Cr2O3- and CrCl3-exposedcells. Cellular uptake of Cr2O3 particles were observed in nano- and fine-particles. The cellular influence ofthe extracellular soluble trivalent chromium was lower than that of Cr2O3 nanoparticles. Cr2O3 nanoparticlesshowed cytotoxicity by hexavalent chromium released at outside and inside of cells. The cellular influencesof Cr2O3 nanoparticles matched those of hexavalent chromium. In conclusion, Cr2O3 nanoparticles have ahigh cytotoxic potential. # 2011 Wiley Periodicals, Inc. Environ Toxicol 00: 000–000, 2011.
Keywords: chromium oxide; nanoparticle; oxidative stress; hexavalent chromium; apoptosis
Correspondence to:M. Horie; e-mail: [email protected]
Contract grant sponsor: New Energy and Industrial Technology Devel-
opment Organization (NEDO); Contract grant number: P06041 (entitled
‘‘Evaluating risks associated with manufactured nanomaterials’’)
Published online in Wiley Online Library (wileyonlinelibrary.com).
DOI 10.1002/tox.20695
�C 2011 Wiley Periodicals, Inc.
1

INTRODUCTION
Among chromium oxide compounds, Cr2O3, trivalent chro-
mium (Cr(III)) is an important industrial material. It has
been used for catalysis, as abrading agents, and as pigments
(e.g., Viridian). Cr2O3 is chemically stable and insoluble in
acidic and basic solutions. Trivalent chromium is an essen-
tial element for human health and has a role in maintaining
the appropriate metabolism of carbohydrates and lipids by
enhancing insulin signaling (Vincent, 2000, 2004).
The toxicity of Cr2O3 is lower than that of hexavalent
chromium (Cr(VI)) compounds such as CrO3, which is
water soluble and a strong oxidant (Barceloux, 1999;
Hininger et al., 2007; Urrutia et al., 2008). There have been
many investigations focusing on the carcinogenicity of hex-
avalent chromium (De Flora and Wetterhahn, 1989;
Holmes et al., 2008). Although the mechanism of chro-
mate-induced lung cancer is incompletely understood, it is
thought that genomic instability is involved (Holmes et al.,
2008).
A carcinogenesis mechanism for hexavalent chromium
has been proposed (Holmes et al., 2008). Briefly, the cyto-
toxic effect of particulate hexavalent chromium such as
lead chromate (PbCrO4) is dependent on the particle solu-
bility. The particulate chromium internalizes to the cell by
phagocytosis, but the ‘‘particle’’ does not affect the cell
(Xie et al., 2004). The particulate hexavalent chromium dis-
solved in the extracellular fluid and soluble Cr(VI) enter the
cell by crossing the cell membrane via an anion transporter
(De Flora and Wetterhahn, 1989; Wise et al., 2004). The in-
tracellular Cr(VI), which enters via ion channels or is
released from internalized particulate hexavalent chro-
mium, is rapidly reduced to Cr(III) (De Flora and Wetter-
hahn, 1989). The reduction of hexavalent chromium to tri-
valent chromium produces reactive oxygen species (ROS),
resulting in oxidization of lipids and proteins. The lipid per-
oxide and oxidized proteins induce metabolic dysfunction
and chromosomal variation (Bagchi et al., 2002; Mattagaja-
singh et al., 2008). Compared with hexavalent chromium,
the cellular influences of trivalent chromium are less signif-
icant because trivalent chromium is more chemically stable
than hexavalent chromium. Additionally, extracellular
Cr(III) has more difficulty in passing through the cell mem-
brane than Cr(VI) (De Flora and Wetterhahn, 1989; Barce-
loux, 1999). Therefore, trivalent chromium compounds
such as Cr2O3 are less cytotoxic than hexavalent chromium
(Hininger et al., 2007).
Nevertheless, some of the biological effects of trivalent
chromium are known. Continuous exposure of human la-
ryngeal carcinoma Hep-2 cells to trivalent chromium
induced cell membrane injury and subsequent apoptosis
(Rudolf and Cervinka, 2003). It has been reported that tri-
valent chromium induced oxidative stress in the brain, liver,
and kidney of goldfish (Lushchak et al., 2009a,b). Cr(III) is
involved in the formation of a Cr-DNA adduct and inhibits
DNA replication (Zhitkovich et al., 2001; Zhitkovich,
2005). Although the biological effect of trivalent chromium
is lower than that of Cr(VI), attention to the toxic effect of
the former is necessary (Levina and Lay, 2008).
Similar to chromium compounds, the toxicity of nickel
compounds is affected by their solubility. In general,
among nickel compounds, nickel oxide (NiO) is classified
as having low toxicity because it is insoluble in water.
Additionally, black NiO has a higher solubility and toxicity
than green NiO (Takahashi et al., 1992).
Nanoparticles (particles with a diameter of �100 nm, in
at least one dimension) have larger surface area per weight
than fine particles (classical larger form). Therefore, the
chemical and physical reactivities of nanoparticle per
weight are larger than that of fine particles. For example,
although NiO is classified as insoluble material in water,
green NiO nanoparticles have higher solubility in water and
in culture media, and NiO nanoparticles show strong cyto-
toxicity than fine particles (Horie et al., 2009b). These
properties induce not only industrial benefits, but also novel
biological effects. Precise assessment of the biological
effects of nanoparticles is, therefore, important for the pre-
cise use of such materials. Understanding the properties of
the cellular response and the response mechanism induced
by nanoparticles are indispensable.
Details of properties of the soluble chromium release from
Cr2O3 nanoparticle in biological fluids and subsequent bio-
logical effects induction are not known. In this study, the bio-
logical effects induced by Cr2O3 nanoparticles were exam-
ined using cultured cells. In particular, we focused on the
soluble chromium release in culture medium and the induc-
tion of oxidative stress. The cellular responses of Cr2O3 nano-
particles were compared with those of fine Cr2O3 particles
and hexavalent chromium compound (K2Cr2O7).
MATERIALS AND METHODS
Cell Culture
Human keratinocyte HaCaT cells were purchased from the
German Cancer Research Center (DKFZ, Heidelberg, Ger-
many). Human lung carcinoma A549 cells were purchased
from the Riken BioResource Center (Tsukuba, Ibaraki, Ja-
pan). These cells were cultured in Dulbecco’s modified
Eagle’s medium (DMEM; Gibco, Invitrogen, Grand Island,
NY) supplemented with 10% heat-inactivated fetal bovine
serum (FBS; CELLect GOLD; MP Biomedicals, Solon,
OH), 100 units/mL of penicillin, 100 lg/mL of streptomy-
cin, and 250 ng/mL of amphotericin B (Nacalai Tesque,
Kyoto, Japan). In this study, this ‘‘DMEM cocktail’’ was
called ‘‘DMEM-FBS.’’
DMEM cultures were placed in a 75-cm2 flask (Corning,
Corning, NY) and incubated at 378C in an atmosphere of
5% CO2. For cellular examinations, cells were seeded in
2 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox
96-well or six-well plates (Corning) at 2 3 105 cells/mL
and incubated for 24 h. Subsequently, the culture medium
was removed, and cells were subjected to metal-oxide dis-
persion and incubated for a further 24 h.
Cr2O3 Particles and Preparation of Cr2O3
Dispersions in DMEM-FBS
Cr2O3 nanoparticles were purchased from Nanostructured &
Amorphous Materials (Houston, TX). According to the man-
ufacturer’s data sheet, the primary particle size was 60 nm
and was obtained from transmission electron microscopy
(TEM) images. Fine Cr2O3 particles and chromium(III) chlo-
ride (CrCl3) were purchased from Kanto Chemical Co. (To-
kyo, Japan). Potassium dichromate (K2Cr2O7) was purchased
fromWako Pure Chemical Industries (Osaka, Japan).
Specific surface area was measured by the Brunauer,
Emmett, and Teller (BET) method. Primary particle size
(Sauter mean diameter) was also estimated by the BET
method. For the cytotoxicity assays described in Figures 1–3,
Cr2O3 powder was dispersed directly in DMEM-FBS at
0.1–10 mg/mL without a dispersant. For some examina-
tions described in Figures 4–9, stable and uniform
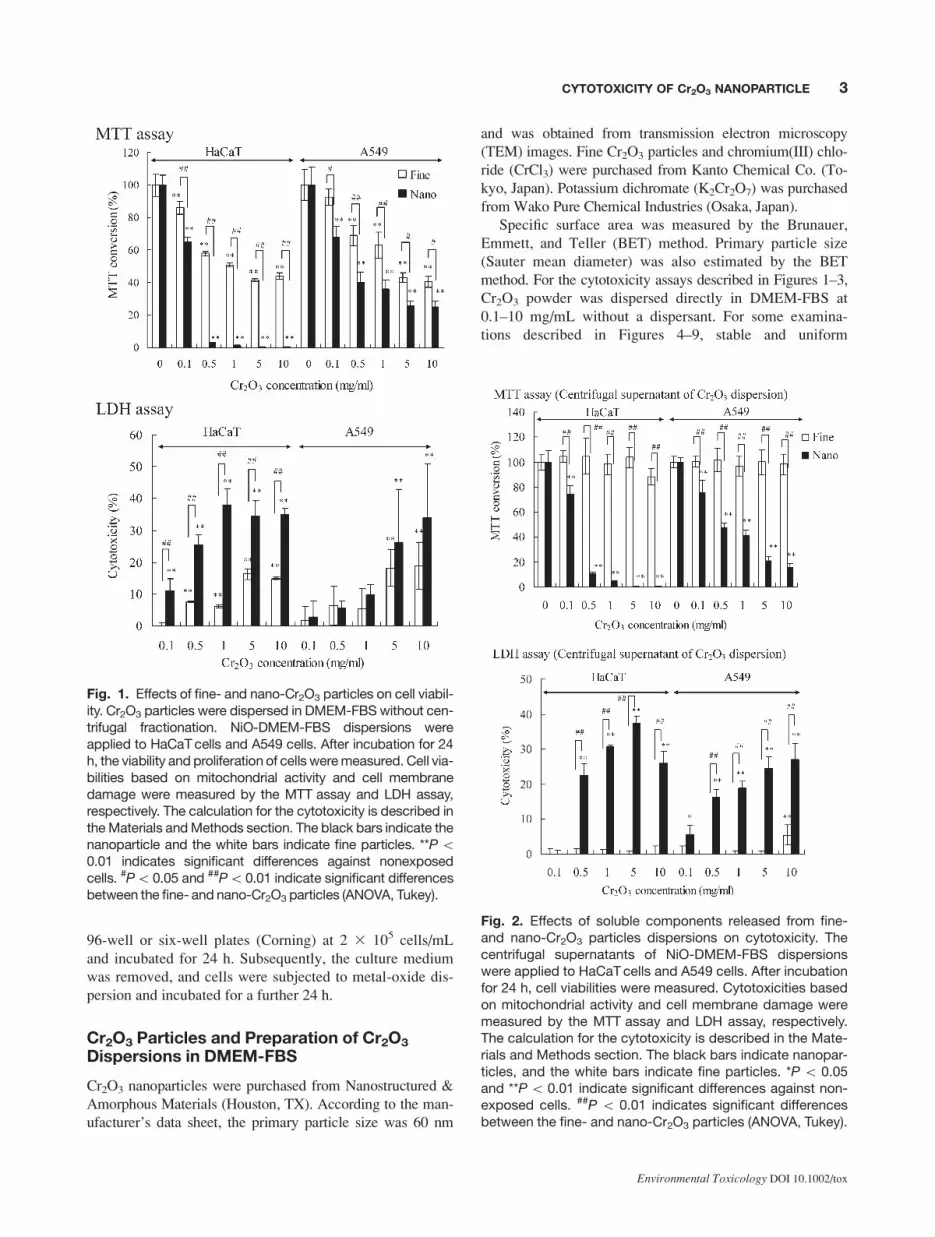
Fig. 1. Effects of fine- and nano-Cr2O3 particles on cell viabil-ity. Cr2O3 particles were dispersed in DMEM-FBS without cen-trifugal fractionation. NiO-DMEM-FBS dispersions wereapplied to HaCaTcells and A549 cells. After incubation for 24h, the viability and proliferation of cells weremeasured. Cell via-bilities based on mitochondrial activity and cell membranedamage were measured by the MTT assay and LDH assay,respectively. The calculation for the cytotoxicity is described intheMaterials andMethods section. The black bars indicate thenanoparticle and the white bars indicate fine particles. **P\0.01 indicates significant differences against nonexposedcells. #P\0.05 and ##P\0.01 indicate significant differencesbetween the fine- and nano-Cr2O3 particles (ANOVA, Tukey).
Fig. 2. Effects of soluble components released from fine-and nano-Cr2O3 particles dispersions on cytotoxicity. Thecentrifugal supernatants of NiO-DMEM-FBS dispersionswere applied to HaCaTcells and A549 cells. After incubationfor 24 h, cell viabilities were measured. Cytotoxicities basedon mitochondrial activity and cell membrane damage weremeasured by the MTT assay and LDH assay, respectively.The calculation for the cytotoxicity is described in the Mate-rials and Methods section. The black bars indicate nanopar-ticles, and the white bars indicate fine particles. *P \ 0.05and **P\ 0.01 indicate significant differences against non-exposed cells. ##P \ 0.01 indicates significant differencesbetween the fine- and nano-Cr2O3 particles (ANOVA, Tukey).
3CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox

dispersions were prepared using a previously described pre-
adsorption and centrifugation method (Horie et al., 2009a).
Nanoparticles induce artificial cellular influences by me-
dium depletion because they can adsorb protein (Casey et al.,
2008; Horie et al., 2009a). To prevent cell starvation caused
by adsorption of medium components onto the particle sur-
face, Cr2O3 particles were initially dispersed in FBS at 80
mg/mL. The dispersion was subsequently centrifuged at
16,000 3 g for 20 min. Precipitated Cr2O3 particles were
washed once with FBS-free DMEM and re-dispersed in an
equivalent volume of fresh DMEM-FBS. This dispersion of
nanoparticles in DMEM-FBS was centrifuged at 8000 3 gfor 20 min. After discarding the supernatant, the precipitate
was resuspended in an equal volume of fresh DMEM-FBS,
and the resulting Cr2O3 dispersion was centrifuged at 40003
g for 20 min. The supernatant was collected as a Cr2O3-
DMEM-FBS dispersion. The Cr2O3-DMEM-FBS dispersions
used in this study did not lead to medium depletion. Adsorp-
tion effects were verified by measuring the concentrations of
protein and calcium in the DMEM-FBS dispersions.
Characterization of Cr2O3-DMEM-FBSDispersions
In this study, we defined ‘‘secondary particles’’ as complex
aggregates of primary particles, proteins from FBS, and
other medium components such as Ca21. The Cr2O3-
DMEM-FBS dispersions prepared by the methods stated
above were divided into three parts and used for simultane-
ous biological examinations, chromium concentration
measurements, and particle-size measurements.
Secondary particle sizes in the Cr2O3-DMEM-FBS dis-
persions were measured by dynamic light scattering (DLS),
as described previously (Kato et al., 2009). Estimated diam-
eters of the secondary particles were calculated as the mean
of three measurements at different wavelengths taken with
the following devices: a DLS-7000 spectrophotometer (633
nm; Otsuka Electronics Co., Hirakata, Japan), an FPAR-
1000 fiberoptic particle analyzer (660 nm; Otsuka Electron-
ics Co.), and a Nanotrac machine (780 nm; Nikkiso Co.,
Tokyo, Japan). Undiluted dispersions were used in these
measurements. Measurements were carried out at 25.08C 60.18C with sample concentrations of 50–80 lg/mL. The
samples for particle-size measurements and cytotoxicity
assays were obtained 1 cm from the surface of the solutions
in static 15-mL tubes. Viscosities of the dispersions were
measured using an Ubbelohde viscometer number 0C
(Sibata Scientific Technology, Tokyo, Japan). DLS can be
used to calculate particle size but has low reliability in cal-
culating particle-size distribution.
The total concentration of chromium was measured by
X-ray fluorescence (XRF). Briefly, 13 mL of the Cr2O3-
DMEM-FBS dispersion was added to 13 mL of a standard
solution containing 0.1 mg/mL of iron (internal standard)
and mixed well. Next, 5 mL of the mixture was dried in an
oven at 2008C for 24 h. The dried sample was ground in an
agate mortar and subjected to XRF analyses using an
energy-dispersive XRF spectrometer (JSX-3201; Jeol, To-
kyo, Japan). The amount of chromium was estimated from
the molar ratio of chromium and the internal standard.
The concentration of soluble chromium was measured
by inductively coupled plasma atomic emission spectrome-
try (ICP-AES, SPS-4000; Seiko Instruments, Chiba, Japan)
in conjugation with ultrafiltration. To completely remove
particulate Cr2O3, the dispersion was kept standing for 24
h. Then, 10 mL of the dispersion was applied to the ultrafil-
tration membrane (molecular weight cutoff was 5000:
Vivaspin 20–50 k, GE Healthcare, Buckinghamshire, UK)
and centrifuged at 6,000 3 g for 50 min. After the mem-
brane was washed thrice with 5 mL of Milli-Q water, the
Fig. 3. Soluble chromium-releasing abilities of fine- andnano-Cr2O3 particles in DMEM. (A) Various amounts ofCr2O3 particles were dispersed in DMEM-FBS and kept for24 h. The dispersion was then separated to particle andsoluble chromium by ultrafiltration. The amount of chromiumin the filtrate (soluble chromium) was detected by ICP-massspectrometry (MS). The black circles indicate Cr2O3 nano-particles, and the white squares indicate fine Cr2O3 par-ticles. (B) Determination of oxidation state of soluble chro-mium released from Cr2O3 nanoparticles. The oxidationstate of soluble chromium was determined by diphenylcar-bazide color comparison method. The value was indicatedas K2Cr2O7 equivalence.
4 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox

Fig. 4. Comparison of the effect of Cr2O3 nanoparticles, Cr2O3 fine particles, CrCl3, andK2Cr2O7 on mitochondrial activity. (A) The influence of chromium compounds on cell via-bility. HaCaT cells and A549 cells were exposed to the Cr2O3 nanoparticle, the mixture ofCr2O3 fine particle and CrCl3, 1 mM CrCl3 solution, and 0.1 and 1 mM K2Cr2O7 solution for6 and 24 h. Mitochondrial activity was then measured by the MTT assay. The Cr2O3 nano-particles dispersion contained 88 lg/mL of Cr2O3 particles and 1 mM soluble chromium.The fine Cr2O3 dispersion contained 88 lg/mL of Cr2O3 particles and 1 mM CrCl3 solution*P\0.05 and **P\ 0.01 versus unexposed cells (ANOVA, Dunnett). (B) Effect of mediumchange on the cytotoxicities of Cr2O3 and K2Cr2O7. Cr2O3 nanoparticles and K2Cr2O7 so-lution was applied to HaCaT and A549 cells. Each dispersion included the same concen-tration of soluble chromium (�1 mM). The cells were exposed to the Cr2O3 nanoparticlesdispersion or K2Cr2O7 solution and then changed to a chromium-free culture medium after2 h. After incubation for additional 22 h, the mitochondrial activities were measured byMTT assay. The MTT conversion was estimated as relative values against the unexposedcells (100%). **P\0.01 versus unexposed cells (ANOVA, Dunnett).
5CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox

amount of chromium in the filtrate was measured by ICP-
AES. DLS was carried out on the filtrate, and light intensity
was not observed. This observation indicated that the par-
ticles were completely removed by ultrafiltration.
Hexavalent chromium concentration was determined by
diphenylcarbazide color comparison method using simpli-
fied assay kit ‘‘PACKTEST’’ (Kyoritsu Chemical-Check
Laboratory, Corp., Tokyo, Japan). To remove the particles,
Cr2O3 DMEM-FBS dispersion (10 mg/mL) was centrifuged
at 16,000 3 g for 20 min. Then, the supernatant or CrCl3solution was mixed with PACKTEST reagent. Subse-
quently, optical density at 540 nm (OD540) was measured.
K2Cr2O7 was used as standard of Cr(VI). The values were
indicated as K2Cr2O7 equivalence.
Measurement of the Cytotoxicity andColony-Forming Ability of Cells
For the cell viability assay, cells were seeded in a 96-well
plate (Corning) at 2 3 105 cells/well. They were incubated
for 24 h, and the culture medium was removed. Subse-
quently, Cr2O3-DMEM dispersion or chromium compound
solution was applied, and the cells were incubated for a fur-
ther 6 and 24 h. In some experiments, the cells were pre-
treated with N-acetylcysteine (NAC; Sigma-Aldrich, St.
Louis, MO). The cells were seeded at 1 3 105 cells/mL and
incubated for 24 h. Subsequently, the medium was changed
to fresh medium containing 2 mM NAC and incubated for
another 24 h. Then, the medium was removed and exposed
to the Cr2O3 dispersion or K2Cr2O7 solution containing 2
mM NAC and incubated for another 2 h. Then, the disper-
sion or solution was changed to fresh medium containing 2
mM NAC and incubated for another 22 h.
For the determination of mitochondrial activity, the 3-
(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bro-
mide (MTT) assay was used. Cells were incubated with 0.5
mg/mL MTT (Nacalai Tesque) at 378C for 2 h. Isopropyl
alcohol containing 40 mM HCl was added to the culture
medium (3:2 by volume). These components were mixed
by pipette until the formazan was completely dissolved.
The optical density of formazan was measured at OD570
using a Multiskan Ascent plate reader (Thermo Labsys-
tems, Helsinki, Finland). In the lactate dehydrogenase
(LDH) assays, LDH release was measured with a tetrazo-
lium salt using a Cytotoxicity Detection Kit PLUS (LDH)
(Roche Diagnostics GmbH, Mannheim, Germany) accord-
ing to the manufacturer’s protocol. The amount of forma-
zan salt formed was measured at 492 nm using the Multis-
kan Ascent plate reader. The maximum amount of LDH
released was determined by incubating the cells with a lysis
solution provided in the kit. Cytotoxicity was calculated
using the following equation:
Cytotoxicity ð%Þ ¼ ðexperimental value � low controlÞ=ðhigh control � low controlÞ3100
where the low control (refers to spontaneous LDH release)
was determined as the LDH released from untreated normal
cells and the high control (refers to the maximum release of
LDH) was determined as the LDH released from cells lysed
by surfactant treatment.
Colony-forming ability was examined by a clonogenic
assay based on the methods described by Herzog et al. (2007)
and Franken et al. (2006). Cells were seeded in six-well plates
(Corning) at 300 cells/well. Each well contained 2 mL of the
cell culture medium. Cells were allowed to attach for �14 h
before they were washed with phosphate-buffered saline
(PBS) and treated with 2 mL of centrifuged supernatant. Cells
were cultured over the time period needed by the control cells
to form colonies, whereby one colony was defined as �50
cells. Here, the cells were cultured for 7 days. After the cul-
ture procedure was completed, supernatants were removed
and cells were washed once with 2 mL of PBS. Cells were
then fixed by 100% methanol for 15 min. After the methanol
was removed, cells were stained by Giemsa’s staining solu-
tion (Nacalai Tesque) diluted 50 times with water for 15 min
and rinsed once with distilled water. The number of colonies
was then counted.
Measurement of Intracellular Oxidative Stress
Intracellular ROS levels were detected using 20,70-dichloro-fluorescin diacetate (DCFH-DA; Sigma-Aldrich). DCFH-
DA was dissolved in dimethyl sulfoxide (DMSO) at 5 mM
Fig. 5. Comparison of the effect of Cr2O3 nanoparticles,Cr2O3 fine particles, CrCl3, and K2Cr2O7 on colony-formingability. The Cr2O3 nanoparticles dispersion, the mixture offine Cr2O3 particles and CrCl3, 1 mM of CrCl3 solution, and0.1 and 1 mM K2Cr2O7 solution were applied to HaCaT cellsand A549 cells. Colony-forming ability was measured by aclonogenic assay. The black bars indicate HaCaT cells, andthe white bars indicate A549 cells. The Cr2O3 nanoparticlesdispersion contained 88 lg/mL of Cr2O3 particles and 1 mMsoluble chromium. The fine Cr2O3 dispersion contained 88lg/mL of Cr2O3 particles and 1 mM CrCl3 solution. **P \0.01 versus unexposed cells (ANOVA, Dunnett).
6 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox

as a stock solution and stored at 2208C; when used in an
experiment, it was diluted 500 times with serum-free me-
dium. Cells were exposed to Cr2O3 dispersion for 2, 6, 12,
and 24 h. The dispersion was then changed to serum-free
DMEM that included 10 lM of DCFH-DA and incubated
for 30 min at 378C. Cells were then washed once with PBS,
collected by 0.25% trypsinization, washed once again with
PBS, and resuspended in 500 lL of PBS. Cell samples in
PBS were excited with a 488-nm argon ion laser in a
Cytomics FC500 flow cytometry system, and the emission
of 20,70-dichlorofluorescein (DCF) was recorded at 525 nm.
Data were collected from at least 5000 gated events.
Fig. 6. Comparison of the effect of Cr2O3 nanoparticles, Cr2O3 fine particles, CrCl3, andK2Cr2O7 on the intracellular ROS level. (A) Effects of NAC on mitochondrial activity. Thecells were pretreated with 2 mM NAC for 24 h. Then, the cells were exposed to the Cr2O3
dispersion or K2Cr2O7 and changed to a chromium-free culture medium containing 2 mMNAC after 2 h. After incubation for additional 22 h, the mitochondrial activities were meas-ured by MTT assay. Significant differences are shown in the figure (ANOVA, Tukey). **Sig-nificantly higher than unexposed cells (P\ 0.01). (B) The Cr2O3 nanoparticle dispersion,the mixture of fine Cr2O3 particle and CrCl3, 1 mM of CrCl3 solution, and 0.1 and 1 mMK2Cr2O7 solution were applied to HaCaT cells and A549 cells. The intracellular ROS levelwas measured by the DCFH method after exposure for 2, 6, 12, and 24 h. The intracellularROS level is shown by a relative value to unexposed cells. The Cr2O3 nanoparticle disper-sion contained 88 lg/mL of Cr2O3 particles and 1 mM soluble chromium. The fine Cr2O3
dispersion contained 88 lg/mL of Cr2O3 particles and 1 mM CrCl3 solution. The intracellu-lar ROS level of HaCaT cells exposed to Cr2O3 nanoparticle dispersion for 24 h could notbe measured. Significant differences are shown in the figure (versus unexposed cells,ANOVA, Tukey). **Significantly higher than unexposed cells (P\0.01).
7CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox

Determination of Glutathione Content
Intracellular glutathione (GSH) content was determined
enzymatically using 5,50-dithiobis-(2-nitrobenzoic acid)
(DTNB), according to the method of Anderson (1985).
Treated cells (1.7 3 105 cells) were harvested by trypsini-
zation, washed with cold PBS, and resuspended in 190 lLof PBS. An equal volume of ice-cold 10% trichloroacetic
acid (TCA) was added to 150 lL of the cell suspension.
Then, cell samples were vortex-mixed for 1 min and centri-
fuged at 1000 3 g for 15 min. The treated soluble superna-
tant was mixed with 0.2 M phosphate buffer (pH 7.5) con-
taining 5 mM EDTA, 0.2 mM NADPH, and 1.0 U/mL GSH
reductase, and maintained at 378C. Absorbance at 412 nm
followed the addition of 1.0 mM DTNB. GSH content was
calculated using reduced GSH (Nacalai Tesque) as the
standard.
Caspase Activity Assay
To obtain total cell extracts, chromium compound-treated
cells were collected by 0.25% trypsinization, washed with
cold PBS, and resuspended on ice for 10 min in lysis buffer
(150 mM NaCl, 50 mM Tris–HCl, pH 7.4, 50 mM NaF, 5
mM EDTA, 0.5% Triton X-100, and 1 mM Na3VO4, along
with Complete; a protease inhibitor cocktail tablet (Roche
Diagnostics)). Nuclei and nonlysed cellular debris were
removed by centrifugation at 10,000 3 g for 1 min. The
protein concentration was determined using a bicinchoninic
acid (BCA) protein assay kit (Thermo Fisher Scientific,
Rockford, IL) with bovine serum albumin (BSA) as the
standard. Caspase enzyme activity was measured by cleav-
age of the Asp-Glu-Val-Asp (DEVD) peptide-conjugated
7-amino-4-trifluoromethyl coumarin (AFC), according to
the protocol outlined by the manufacturer of the caspase-3
fluorometric protease assay kit (Medical & Biological Lab-
oratories Co., Nagoya, Japan). Substrate cleavage, which
resulted in the release of AFC fluorescence (excitation, 400
nm; emission, 505 nm), was measured using a Fluoroskan
Ascent CF Plate Reader (Thermo Labsystems).
Western Blotting Analysis
Protein samples from cells were solubilized with sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-
PAGE) sample buffer (200 lL of 10% SDS, 60 lL of 1.0
M Tris–HCl (pH 6.8), 50 lL of 2-mercaptoethanol, 100 lLof glycerol, 1 mg of bromophenol blue, and 590 lL of dis-
tilled water) and heated at 1008C for 5 min. Proteins were
separated by 12.5% SDS-PAGE and transferred to a polyvi-
nylidene difluoride (PVDF) membrane using the iBlotTM
gel transfer system (Invitrogen). Blots were blocked for 1 h
at room temperature in fresh blocking buffer (0.1% Tween
20 in Tris-buffered saline, pH 7.4, containing 5% skimmed
milk). The membrane was incubated overnight at 48C with
anti-caspase-3 antibody (Cell Signaling Technology, Dan-
vers, MA), anti-poly(ADP-ribose) polymerase (PARP) anti-
body (Cell Signaling Technology), or anti-actin antibody
(Chemicon; Millipore Corp., Billerica, MA) in Tris-buf-
fered saline containing 0.1% Tween 20 (TBST). After
Fig. 7. Comparison of intracellular GSH level on Cr2O3 nanoparticles-, Cr2O3 fine particles-,CrCl3-, and K2Cr2O7-exposed cells The Cr2O3 nanoparticle dispersion, the mixture of fineCr2O3 particle and CrCl3, 1 mM of CrCl3 solution, and 0.1 and 1 mM K2Cr2O7 solution wereapplied to HaCaTcells and A549 cells. The intracellular GSH level was measured after 6 h ofexposure. The Cr2O3 nanoparticle dispersion contained 88 lg/mL of Cr2O3 particles and 1mM soluble chromium. The fine Cr2O3 dispersion contained 88 lg/mL of Cr2O3 particles and1 mM CrCl3 solution. Significant differences are shown in the figure (versus unexposed cells,ANOVA, Dunnett). *Significantly higher than unexposed cells (**P\0.01).
8 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox

incubation, the membrane was washed thrice for 5 min
with TBST and incubated for 1 h at room temperature with
peroxidase-conjugated anti-mouse IgG antibody (Jackson
Immuno Research Laboratories, West Grove, PA) or perox-
idase-conjugated anti-rabbit IgG antibody (Jackson
Immuno Research) and then washed again with TBST. Per-
oxidase-active species on the membrane were detected
using a Chemi-Lumi One Kit (Nacalai Tesque) and
Lumino-Image Analyzer LAS-3000 mini (Fujifilm Corp.,
Tokyo, Japan). Densitometric analysis was performed using
Multi Gauge Version 3.0 (Fujifilm Corp.).
Determination of DNA Damage
DNA damage by Cr2O3, CrCl3, and K2Cr2O7 was detected
by the comet assay (1988) using the CometAssayTM Silver
kit (Trevigen, Gaithersburg, MD) according to the protocol
outlined by the manufacturer. The kit included LMAgarose,
Fig. 8. Effect of Cr2O3 nanoparticles on caspase activityand PARP activation. The Cr2O3 nanoparticles dispersioncontained 88 lg/mL of Cr2O3 particles and 1 mM solublechromium. (A) Caspase enzyme activity. Cell lysate sampleswere taken after cells were exposed to the Cr2O3 nanopar-ticles dispersion for 2 and 6 h. The cell lysate was thenapplied to the caspase activity assay. Caspase activity wasmeasured using DEVD-AFC as the substrate. Caspase ac-tivity is shown as a relative value to unexposed cells. *P \0.05 (versus control, Dunnett, ANOVA). (B) Activated cas-pase-3 and PARP were detected by Western blotting. Lane1: untreated cells; lane 2: Cr2O3 nanoparticle-exposed cells.Relative densitometric intensity of each band showeduntreated cell as 1.0.
Fig. 9. Comparison of DNA damage to cells exposed to theCr2O3 nanoparticles, fine Cr2O3 particles, CrCl3, andK2Cr2O7. Cells were exposed to the Cr2O3 nanoparticlesdispersion, the mixture of fine Cr2O3 particles and CrCl3, 1mM of CrCl3 solution, and 0.1 and 1 mM K2Cr2O7 solutionfor 6 h, and the DNA damage was measured by the cometassay. The Cr2O3 nanoparticles dispersion contained 88 lg/mL of Cr2O3 particles and 1 mM soluble chromium. The fineCr2O3 dispersion contained 88 lg/mL of Cr2O3 particles and1 mM CrCl3 solution. Approximately 200 cells were counted,and DNA damage classified according to the classificationdescribed by Tayama and Nakagawa (2004). A typical imageat each class is shown under the graph.
9CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox

Fig. 10. Transmission electron microscopic observations of cells exposed to Cr2O3. (A–D)HaCaT cells exposed to the nano- and fine-Cr2O3 dispersion for 6 h. (E–H) A549 cellsexposed to the nano- (A, C, E, and G) and fine- (B, D, F, and H) Cr2O3 dispersion for 6 h.The bar in panels A, B, E, and F indicates 3.0 lm, and the bar in panels C, D, G, and H indi-cates 306 nm.
10 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox

CometSlide, and Lysis Solution. HaCaT cells were exposed
by a TiO2-DMEM-FBS dispersion for 24 h, harvested by
trypsinization, and prepared at 1 3 105 cells/mL with PBS.
Cells were mixed with molten LMAgarose at 1:10 (v/v),
and 75 lL of the mixture was immediately applied onto the
CometSlide. After the application of agarose gel, slides
were immersed in prechilled Lysis Solution and left on ice
for 30 min. After removing excess buffer from the slides,
slides was immersed in alkaline solution (300 mM NaOH,
1 mM EDTA, pH[13) and left for 20 min at room temper-
ature in the dark. Slides were washed with Tris-Borate-
EDTA (TBE) twice, and electrophoresis was carried out
using a Mupid submarine electrophoresis system (Advance
Co., Tokyo, Japan) with 50 V for 2 min. Slides were then
rinsed by dipping several times in dH2O. After the slides
were soaked in 70% ethanol for 5 min, electrophoresed
DNAs were stained by silver staining and counted under a
microscope. Determination of the level of DNA damage
was as described by Tayama and Nakagawa (2004).
TEM Observations
Cell specimens exposed to Cr2O3 were observed using a
transmission electron microscope. Briefly, cells were fixed
using 1.2% glutaraldehyde for 1 h at 48C and 1% OsO4 so-
lution for 1 h at 48C, dehydrated in ethanol, and embedded
in epoxy resin. The resulting sample was then cut into ultra-
thin sections suitable for TEM observations by diamond-
knife ultramicrotomy. TEM observations were then carried
out using an H-7000 machine (Hitachi, Tokyo, Japan). The
acceleration voltage in the TEM was 75 kV.
Statistical Analyses
Data are the mean 6 standard deviation (SD) for at least
three separate experiments. Statistical analyses were done
by the analysis of variance (ANOVA) using the Dunnett or
Tukey tests for multiple comparisons. The calculation
method has been described in each figure legend.
RESULTS
Comparison of Cytotoxicity Induced by Cr2O3
Fine and Nanoparticles
Specific surface area of Cr2O3 fine and nanoparticles were
2.63 and 43.41 m2/g, respectively. Primary particle size of
Cr2O3 fine and nanoparticles were estimated by BET results
were 440 and 26.5 nm, respectively.
Cr2O3 fine and nanoparticles were applied to A549 cells
and HaCaT cells at concentrations of 0.1–10 mg/mL. After
24 h exposure, cytotoxicities were detected by the MTT
assay and LDH assay. Cell viability was dose-dependently
decreased by Cr2O3 exposure in the MTT assay and LDH
assay (Fig. 1). The cytotoxicity of nanoparticles was stron-
ger than that of fine particles. HaCaT cells showed a greater
reduction in cell viability by Cr2O3 exposure than A549
cells. In particular, the cytotoxicity of nanoparticles was
higher than that for fine particles at a high concentration of
Cr2O3 (0.5 mg/mL). In NiO, if the particle size became
nano-scale (1–100 nm), solubility increased. The released
Ni21 affects cytotoxicity (Horie et al., 2009b). Some previ-
ous studies suggest that metal ion release from metal oxide
nanoparticles was involved in their cytotoxicity (Horie
et al., 2009b; Studer et al., 2010).
Therefore, next, the cellular influences of soluble compo-
nents from Cr2O3 particles were examined. The Cr2O3 me-
dium dispersion used in the examination shown in Figure 1
was centrifuged at 16,0003 g for 20 min to remove the parti-
cle. The supernatant was applied to the cells, and the influ-
ence on cell viability was examined (Fig. 2). The centrifugal
supernatant of the fine Cr2O3 dispersion did not show cyto-
toxicity. On the other hand, the centrifugal supernatant of
Cr2O3 nanoparticle dispersion had cytotoxic activity. These
results suggest that the soluble component released from
Cr2O3 is involved in the cytotoxicity induced by Cr2O3 nano-
particles. The specific gravity of fine and nanoparticles is dif-
ferent; so, the cytotoxic effect of Cr2O3 nanoparticles in the
supernatant may not be completely removed by centrifuga-
tion. The smaller nanoparticle might be remaining in the su-
pernatant. Therefore, further tests were carried out.
Release of Soluble Chromium from Cr2O3 Fineand Nanoparticles to the Culture Medium
The release of soluble chromium from fine and nanoparticles
in DMEM-FBS was measured (Fig. 3A). Fine Cr2O3 particles
were insoluble in DMEM-FBS. The soluble chromium
released from fine Cr2O3 particles was not detected by ICP-
AES. In contrast, the Cr2O3 nanoparticles dissolved in the
DMEM-FBS. The rate of soluble chromium from Cr2O3
nanoparticles against charged Cr2O3 particles was estimated
as 1–1.5%. When the initial concentration of Cr2O3 was 0.1,
0.5, 1.0, 5.0, and 10 mg/mL, the concentration of soluble
chromium from Cr2O3 nanoparticles was estimated to be
0.24, 0.98, 4.76, 19.49, and 51.23 lg/mL, respectively. Cr2O3
did not influence the salts (P, Na, and Ca) contained in the
medium. Concentration of P, Na, and Ca in the Cr2O3 nano-
particle dispersion tended to be higher than the concentration
of these elements in the fine Cr2O3 dispersion. Although the
concentration of these elements was slightly low when Cr2O3
concentration was high, there was no noted influence of
Cr2O3 concentration on the concentration of P, Na, and Ca.
Oxidation state of soluble chromium released from
Cr2O3 nanoparticles was determined (Fig. 3B). Cr2O3 nano-
particle-DMEM-FBS dispersion at a concentration of 10
mg/mL included �0.4 mM K2Cr2O7 equivalence. This
result indicated that Cr2O3 nanoparticle released hexavalent
chromium in medium.
11CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox

Influence of Released Soluble Chromium fromCr2O3 Particles on the Viability andProliferation of Cells
The cytotoxicity induced by Cr2O3 nanoparticles could
have been caused by released soluble Cr(VI). Therefore, we
compared the cellular influences of Cr2O3 nanoparticles,
fine Cr2O3 particles, CrCl3 as soluble Cr(III), and K2Cr2O7
as soluble Cr(VI).
The Cr2O3 nanoparticle-DMEM-FBS dispersion includ-
ing soluble chromium compound was prepared by centrifu-
gation. The secondary particle size of Cr2O3 in the DMEM-
FBS dispersion was measured by DLS. Secondary particle
size on the day after preparation of the dispersion was
295.1 and 124.6 nm as light intensity particle size and num-
ber average particle size, respectively. However, precipita-
tion of aggregate could be observed by the naked eye in the
Cr2O3 nanoparticle dispersion. The light intensity of DLS
measurement decreased with time. Therefore, the second-
ary particles of Cr2O3 nanoparticles in DMEM-FBS were
very unstable. The total chromium concentration in the
Cr2O3 nanoparticle-DMEM-FBS dispersion was 163.36 66.09 lg/mL (mean 6 SD). The concentrations of Cr2O3
particles and soluble chromium in the dispersion were
88.37 lg/mL (1.16 mM) and 75 lg/mL (0.99 mM), respec-
tively. Therefore, cellular influences induced by the Cr2O3
nanoparticle dispersion, the mixture of 88 lg/mL of fine
Cr2O3 particles and 1.0 mM CrCl3, 1.0 mM CrCl3 solution,
and 0.1 and 1.0 mM K2Cr2O7 solution were compared.
However, the sedimentation occurred in both Cr2O3 nano-
particle and Cr2O3 fine particle dispersion because these
dispersions were unstable. Because the particle accumu-
lated on the cell, the concentration of dispersion might not
reflect the cellular exposure concentration accurately.
Therefore, an accurate evaluation of concentration depend-
ency is not suitable.
The influence on cell viability was initially examined
(Fig. 4A). Each dispersion or solution was applied to
HaCaT cells and A549 cells and exposed for 6 and 24 h.
Cell viabilities were then measured by the MTT assay. Cell
viability was drastically reduced in cells exposed to the
Cr2O3 nanoparticle dispersion and K2Cr2O7 solution. The
exposure of the mixture of fine Cr2O3 particle and CrCl3led to a slight decrease in cell viability; cell viability after
24 h exposure was 80–90% in A549 cells. CrCl3 solution
did not induce a reduction in cell viability. Furthermore, we
examined the relationship between the extracellular soluble
chromium and cytotoxicity of Cr2O3 nanoparticles (Fig.
4B). The cells were exposed to the Cr2O3 nanoparticle dis-
persion or K2Cr2O7 solution and then changed to a chro-
mium-free culture medium after 2 h. After incubation for
additional 22 h, the mitochondrial activities were measured.
Both the Cr2O3 nanoparticles and the K2Cr2O7 solution
reduced mitochondrial activity. There were no significant
differences between the Cr2O3 nanoparticles- and the
K2Cr2O7 solution-exposed cells. This result indicates that
extracellular soluble chromium is critical for cytotoxicity
of Cr2O3 nanoparticle.
The influence on cell proliferation was examined by a
clonogenic assay (Fig. 5). An inhibitory effect on colony-
forming ability was shown in the Cr2O3 nanoparticle dis-
persion, the mixture of fine Cr2O3 particle and CrCl3, and
K2Cr2O7 solution.
Cellular Effects Induced by Cr2O3
Nanoparticles
The cellular influences induced by Cr2O3 nanoparticles
were examined. The decrease in mitochondrial activity was
reduced by pretreating the HaCaT cells with NAC (Fig.
6A). In A549 cells, NAC treatment did not significantly
affect the mitochondrial activity. However, pretreatment of
the A549 cells with NAC tended to reduce mitochondrial
damage. NAC is readily taken up by cells and, subse-
quently, acts as a source of cellular GSH. This result sug-
gests that oxidative stress is involved in the cytotoxicity of
Cr2O3 nanoparticles.
Levels of intracellular ROS in Cr2O3-, CrCl3-, or
K2Cr2O7-exposed cells over time were measured (Fig. 6B).
The intracellular ROS level was increased in cells exposed
to Cr2O3 nanoparticles and K2Cr2O7 solution. Exposure of
fine Cr2O3 particles and CrCl3 did not induce an increase in
the intracellular ROS level. The increase in intracellular
ROS level induced by exposure to Cr2O3 nanoparticles was
time-dependent. After cells were exposed to Cr2O3 nano-
particles for 6 h, intracellular ROS levels compared with
nonexposed cells were 11 and 7.8 times than that seen in
HaCaT cells and A549 cells, respectively. In Cr2O3 nano-
particles-exposed HaCaT cells, the intracellular ROS level
was increased to 27 times than that in nonexposed cells at
12 h exposure. Exposure of HaCaT cells to Cr2O3 nanopar-
ticles for 24 h led to cell death; thus, the intracellular ROS
level at 24 h exposure could not be measured. In A549
cells, intracellular ROS levels in Cr2O3 nanoparticles-
exposed cells at 12 and 24 h exposure were 10 and 16 times
of those of nonexposed cells, respectively. The induction of
intracellular ROS level by Cr2O3 nanoparticle was similar
to induction by K2Cr2O7.
The intracellular level of GSH was then measured
(Fig. 7). GSH level was reduced by exposure to Cr2O3 nano-
particles and K2Cr2O7. Compared with the Cr2O3 nanopar-
ticle, exposure of the fine Cr2O3 particle and CrCl3 did not
lead to GSH decrease in HaCaT cells. In A549 cells, expo-
sure of the CrCl3 led to a slight decrease in GSH after 12 h
exposure. Increase in the intracellular ROS level is related to
the induction of apoptosis (Annunziato et al., 2003). There-
fore, the caspase activity, which is apoptosis-related cysteine
peptidase activity, of Cr2O3-exposed cells was measured.
Caspase activity was increased in Cr2O3 nanoparticles-
12 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox

exposed cells after 6 h (Fig. 8A). Further, activation of cas-
pase-3, which is caspase at downstream of caspase-related ap-
optosis pathway, was detected by Western blotting (Fig. 8B).
Cleavage of PARP was also detected by Western blotting.
PARP cleavage is an important marker of caspase-3-mediated
apoptosis. The cleavage of 116-kDa PARP by caspase-3
forms 85-kDa fragments (Nicholson et al., 1995; Robertson
et al., 1997). PARP cleavage was detected in Cr2O3 nanopar-
ticles-exposed cells. DNA injury was detected by comet
assay (Fig. 9). Exposure to Cr2O3 nanoparticles and K2Cr2O7
solution induced considerable DNA injury. These results sug-
gest that exposure to Cr2O3 nanoparticles led to increased
levels of intracellular ROS and subsequent apoptosis and
DNA injury.
Cellular Uptake of Cr2O3 Particles
Cellular uptake of Cr2O3 particles was observed by TEM
because Cr2O3 nanoparticles showed stronger cytotoxicity
than fine Cr2O3 particles and CrCl3 (Fig. 10). Cellular
uptake of nanoparticles and fine particles was observed in
HaCaT cells and A549 cells. Internalized Cr2O3 formed
aggregates and was present in the cytosol. Entry of particles
to the nucleus as particles was not observed. Fine particles
tended to adhere to the cell surface.
DISCUSSION
In general, trivalent chromium has lower toxicity than hex-
avalent chromium. Hexavalent chromium shows toxicity
due to reduction from Cr(VI) to Cr(III) (e.g., formation of
Cr(V) by reduction of Cr(VI) generates the hydroxyl radical
(Shi and Dalal, 1989). Although Cr(VI) passes through the
cell membrane via an ion channel, Cr(III) permeability of
the cell membrane is lower than for Cr(VI) (De Flora and
Wetterhahn, 1989; Barceloux, 1999). Thus, the potential
cytotoxic activity of Cr(III) is lower than for Cr(VI) (Bie-
dermann and Landolph, 1990; Bagchi et al., 2002; Hininger
et al., 2007).
If Cr2O3 was insoluble, the cellular effects induced by
Cr2O3 would be limited, but Cr2O3 nanoparticles were solu-
ble in the cell culture medium. Cr2O3 nanoparticles used in
this study released hexavalent chromium in culture media.
Moreover, Cr2O3 nanoparticles showed remarkable cyto-
toxicity at a low concentration. In our study, cellular influ-
ences induced by Cr2O3 nanoparticles matched to that of
K2Cr2O7.
This study showed that extracellular soluble chromium
was critical for cytotoxicity of Cr2O3 nanoparticles. Extrac-
ellular Cr(VI) enters the cell via ion channel and shows
strong oxidizability outside and inside of cell (De Flora and
Wetterhahn, 1989). On the other hand, TEM observations
of Cr2O3-exposed cells showed cellular uptake of Cr2O3
nano- and fine-particles. Subordinately, internalized Cr2O3
particles will release soluble chromium inside the cell. This
intracellular soluble Cr(VI) release also is an important fac-
tor for Cr2O3 nanoparticles cytotoxicity. Additionally, in-
tracellular dissolution of Cr2O3 nanoparticles leads to a
high concentration of Cr(VI) inside the cell. Although the
cellular uptake of fine Cr2O3 particles was also observed,
the cellular effects induced by fine Cr2O3 particles were
small compared with the same weight of Cr2O3 nanopar-
ticles. We concluded that cellular uptake of Cr2O3 nanopar-
ticles and subsequent cytotoxicity was due to released
Cr(VI) inside and outside the cell. It has been reported that
cellular uptake of metal oxide nanoparticles and subsequent
release of metal ions induce cytotoxicity due to NiO (Horie
et al., 2009b). Cellular uptake of nanoparticles was also
observed in TiO2 particles (Horie et al., 2010). However,
taken into account that TiO2 nanoparticles were insoluble
in the medium, the cellular influence of TiO2 was lower
than for NiO and Cr2O3. These observations indicate that
the cellular uptake of nanoparticles is a prerequisite for its
cellular effects. However, pre-eminent cytotoxicity is not
induced only by cellular uptake. The increase in solubility
in the biological fluid such as a culture medium is one of
the important factors for the cytotoxicity of Cr2O3 nanopar-
ticles. Even though Cr2O3 is classified as trivalent chro-
mium, why Cr2O3 nanoparticles release hexavalent chro-
mium is still unclear. The composition or purity of Cr2O3
nanoparticles powder was not examined in this study, but it
can be said that at least the nanoparticles that were sold as
Cr2O3 released hexavalent chromium.
Understanding the properties of nanoparticle, such as
solubility and oxidation state, is essential for industrial use.
This fact is also important for environmental burden caused
by disposed Cr2O3 nanoparticles. In our examinations of
the cellular influences of Cr2O3, strong oxidative stress and
apoptosis were induced. On theoretical grounds, Cr(VI)
will be reduced to Cr(III) in the cell; hence, toxicity of
Cr(III) is lower than that of Cr(VI). Nevertheless, there are
some investigations that Cr(III) also showed cellular influ-
ences. Oral administration of Cr(III) induced an increase in
lipid peroxidation in urine, though; the increase in the lipid
peroxidation level was lower than for Cr(VI) (Bagchi et al.,
2002). Several investigations suggest that Cr(III) induces
an increase in intracellular ROS level and apoptosis (Bala-
murugan et al., 2002; Rudolf and Cervinka, 2003; Rudolf
and Cervinka, 2009). Oxidative stress induced by Cr(III)
has also been reported in vivo. Oxidative stress was
increased in the brain, liver, and kidney of goldfish exposed
to Cr(III) (Lushchak et al., 2009a,b). In these investigations,
soluble chromium compounds such as chromium chloride
and chromium acetate were used. Therefore, the toxicity
would be due mainly to extracellular soluble Cr(III). It is
important for the cytotoxicity that Cr2O3 nanoparticles
become source of Cr(VI) or Cr(III) inside of cells after cel-
lular uptake. This study showed that Cr2O3 fine particles
13CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox

did not dissolve in the culture medium. Hence, Cr2O3 solu-
bility was not sufficient for cytotoxicity. In contrast, Cr2O3
nanoparticles showed high solubility in the medium, and
cellular uptake of Cr2O3 nanoparticles was observed. The
extracellular and the intracellular dissolution of Cr2O3
nanoparticles induces drastic effects such as oxidative
stress, apoptosis, and DNA injury.
These cellular influences induced by Cr2O3 nanopar-
ticles were equivalent to cellular influences induced by
K2Cr2O7. This suggests that Cr2O3 nanoparticle has equiva-
lent risks as that of Cr(VI). Increased solubility, compared
with its larger form, is one of the features of metal oxide
nanoparticles, and this property is of great importance for
the toxicity of nanoparticles.
REFERENCES
Anderson ME. 1985. Determination of glutathione and glutathi-
one disulfide in biological samples. Methods Enzymol 113:
548–555.
Annunziato L, Amoroso S, Pannaccione A, Cataldi M, Pignataro
G, D’Alessio A, Sirabella R, Secondo A, Sibaud L, Di Renzo
GF. 2003. Apoptosis induced in neuronal cells by oxidative
stress: Role played by caspases and intracellular calcium ions.
Toxicol Lett 139:125–133.
Bagchi D, Stohs SJ, Downs BW, Bagchi M, Preuss HG. 2002. Cy-
totoxicity and oxidative mechanisms of different forms of chro-
mium. Toxicology 180:5–22.
Balamurugan K, Rajaram R, Ramasami T, Narayanan S. 2002.
Chromium(III)-induced apoptosis of lymphocytes: Death deci-
sion by ROS and Src-family tyrosine kinases. Free Radic Biol
Med 33:1622–1640.
Barceloux DG. 1999. Chromium. J Toxicol Clin Toxicol 37:
173–194.
Biedermann KA, Landolph JR. 1990. Role of valence state and
solubility of chromium compounds on induction of cytotoxicity,
mutagenesis, and anchorage independence in diploid human
fibroblasts. Cancer Res 50:7835–7842.
Casey A, Herzog E, Lyng FM, Byrne HJ, Chambers G, Davoren
M. 2008. Single walled carbon nanotubes induce indirect cyto-
toxicity by medium depletion in A549 lung cells. Toxicol Lett
179:78–84.
De Flora S, Wetterhahn KE. 1989. Mechanisms of chromium me-
tabolism and genotoxicity. Life Chem Rep 7:169–244.
Franken NA, Rodermond HM, Stap J, Haveman J, van Bree C.
2006. Clonogenic assay of cells in vitro. Nat Protoc 1:2315–
2319.
Herzog E, Casey A, Lyng FM, Chambers G, Byrne HJ, Davoren
M. 2007. A new approach to the toxicity testing of carbon-
based nanomaterials—The clonogenic assay. Toxicol Lett
174:49–60.
Hininger I, Benaraba R, Osman M, Faure H, Marie Roussel A,
Anderson RA. 2007. Safety of trivalent chromium complexes:
No evidence for DNA damage in human HaCaT keratinocytes.
Free Radic Biol Med 42:1759–1765.
Holmes AL, Wise SS, Wise JP Sr. 2008. Carcinogenicity of hexa-
valent chromium. Indian J Med Res 128:353–372.
Horie M, Nishio K, Fujita K, Endoh S, Miyauchi A, Saito Y, Iwa-
hashi H, Yamamoto K, Murayama H, Nakano H, Nanashima N,
Niki E, Yoshida Y. 2009a. Protein adsorption of ultrafine metal
oxide and its influence on cytotoxicity toward cultured cells.
Chem Res Toxicol 22:543–553.
Horie M, Nishio K, Fujita K, Kato H, Nakamura A, Kinugasa S,
Endoh S, Miyauchi A, Yamamoto K, Murayama H, Niki E,
Iwahashi H, Yoshida Y, Nakanishi J. 2009b. Ultrafine NiO par-
ticles induce cytotoxicity in vitro by cellular uptake and subse-
quent Ni(II) release. Chem Res Toxicol 22:1415–1426.
Horie M, Nishio K, Fujita K, Kato H, Endoh S, Suzuki M, Naka-
mura A, Miyauchi A, Kinugasa S, Yamamoto K, Iwahashi H,
Murayama H, Niki E, Yoshida Y. 2010. Cellular responses by
stable and uniform ultrafine titanium dioxide particles in cul-
ture-medium dispersions when secondary particle size was 100
nm or less. Toxicol In Vitro 24:1629–1638.
Kato H, Suzuki M, Fujita K, Horie M, Endoh S, Yoshida Y, Iwahashi
H, Takahashi K, Nakamura A, Kinugasa S. 2009. Reliable size deter-
mination of nanoparticles using dynamic light scattering method for
in vitro toxicology assessment. Toxicol InVitro 23:927–934.
Levina A, Lay PA. 2008. Chemical properties and toxicity of
chromium(III) nutritional supplements. Chem Res Toxicol
21:563–571.
Lushchak OV, Kubrak OI, Lozinsky OV, Storey JM, Storey KB,
Lushchak VI. 2009a. Chromium(III) induces oxidative stress in
goldfish liver and kidney. Aquat Toxicol 93:45–52.
Lushchak OV, Kubrak OI, Torous IM, Nazarchuk TY, Storey KB,
Lushchak VI. 2009b. Trivalent chromium induces oxidative
stress in goldfish brain. Chemospherea 75:56–62.
Mattagajasingh SN, Misra BR, Misra HP. 2008. Carcinogenic
chromium(VI)-induced protein oxidation and lipid peroxida-
tion: Implications in DNA-protein crosslinking. J Appl Toxicol
28:987–997.
Nicholson DW, Ali A, Thornberry NA, Vaillancourt JP, Ding CK,
Gallant M, Gareau Y, Griffin PR, Labelle M, Lazebnik YA,
Munday NA, Raju SM, Smulson ME, Yamin TT, Yu VL, Miller
DK. 1995. Identification and inhibition of the ICE/CED-3
protease necessary for mammalian apoptosis. Nature 376:37–
43.
Robertson NM, Zangrilli J, Fernandes-Alnemri T, Friesen PD, Lit-
wack G, Alnemri ES. 1997. Baculovirus P35 inhibits the gluco-
corticoid-mediated pathway of cell death. Cancer Res 57:43–47.
Rudolf E, Cervinka M. 2003. Chromium (III) produces distinct
type of cell death in cultured cells. Acta Med 46:139–146.
Rudolf E, Cervinka M. 2009. Trivalent chromium activates Rac-1
and Src and induces switch in the cell death mode in human
dermal fibroblasts. Toxicol Lett 188:236–242.
Shi XL, Dalal NS. 1989. Chromium (V) and hydroxyl radical for-
mation during the glutathione reductase-catalyzed reduction of
chromium (VI). Biochem Biophys Res Commun 163:627–634.
Studer AM, Limbach LK, Van Duc L, Krumeich F, Athanassiou
EK, Gerber LC, Moch H, Stark WJ. 2010. Nanoparticle cyto-
toxicity depends on intracellular solubility: Comparison of sta-
bilized copper metal and degradable copper oxide nanopar-
ticles. Toxicol Lett 197:169–174.
14 HORIE ET AL.
Environmental Toxicology DOI 10.1002/tox

Takahashi S, Yamada M, Kondo T, Sato H, Furuya K, Tanaka I.
1992. Cytotoxicity of nickel oxide particles in rat alveolar mac-
rophages cultured in vitro. J Toxicol Sci 17:243–251.
Tayama S, Nakagawa Y. 2004. Comet assay attempted with silver
staining method and manual microscopic analysis using CHO-
K1 cells. Ann Rep Tokyo Metr Inst PH 55:315–318.
Urrutia C, Rudolph A, Lermanda MP, Ahumada R. 2008. Assess-
ment of EDTA in chromium (III-VI) toxicity on marine intertidal crab
(Petrolisthes laevigatus). Bull Environ Contam Toxicol 80:526–528.
Vincent JB. 2000. The biochemistry of chromium. J Nutr
130:715–718.
Vincent JB. 2004. Recent developments in the biochemistry of
chromium(III). Biol Trace Elem Res 99:1–16.
Wise SS, Holmes AL, Ketterer ME, Hartsock WJ, Fomchenko E,
Katsifis S, Thompson WD, Wise JP Sr. 2004. Chromium is the
proximate clastogenic species for lead chromate-induced clasto-
genicity in human bronchial cells. Mutat Res 560:79–89.
Xie H, Holmes AL, Wise SS, Gordon N, Wise JP Sr. 2004. Lead
chromate-induced chromosome damage requires extracellular
dissolution to liberate chromium ions but does not require parti-
cle internalization or intracellular dissolution. Chem Res Toxi-
col 17:1362–1367.
Zhitkovich A. 2005. Importance of chromium-DNA adducts in
mutagenicity and toxicity of chromium(VI). Chem Res Toxicol
18:3–11.
Zhitkovich A, Song Y, Quievryn G, Voitkun VA. 2001. Non-oxi-
dative mechanisms are responsible for the induction of muta-
genesis by reduction of Cr(VI) with cysteine: Role of ternary
DNA adducts in Cr(III)-dependent mutagenesis. Biochemistry
40:549–560.
15CYTOTOXICITY OF Cr2O3 NANOPARTICLE
Environmental Toxicology DOI 10.1002/tox