characterization of the bactericidal effect of dietary sphingosine and its activity under intestinal...
TRANSCRIPT

www.elsevier.com/locate/ijfoodmicro
International Journal of Food Mic
Characterization of the bactericidal effect of dietary sphingosine
and its activity under intestinal conditions
Sam Possemiers a, John Van Camp b, Selin Bolca a, Willy Verstraete a,*
aLaboratory of Microbial Ecology and Technology (LabMET), Ghent University, Coupure Links 653, B-9000 Ghent, BelgiumbLaboratory of Food Science and Human Nutrition, Ghent University, Belgium
Received 10 November 2004; received in revised form 12 May 2005; accepted 12 May 2005
Abstract
Sphingosine is known as a natural antimicrobial agent, protecting the human skin from bacterial colonization and possibly
affecting the intestinal microbial community after ingestion. In this study we further investigated the antibacterial spectrum of
dietary d-eythro-sphingosine in saline towards three intestinal pathogens and to the health promoting lactobacilli and
bifidobacteria. The degree of bactericidal effect was studied using plate counts and Live/Dead analysis combined with flow
cytometry. To assess activity under complex intestinal conditions, sphingosine was dosed to the Simulator of the Human
Intestinal Microbial Ecosystem (SHIME) for a period of 11 days. Finally, we tried to elucidate the factors influencing the
activity and the mode of action of sphingosine. In all performed experiments, high correlation occurred between plate counts
and Live/Dead analysis. In saline a strong antibacterial effect was seen to all tested species, Gram-negative and Gram-positive,
and sphingosine not selectively acted against pathogens, as health promoting bacteria were also affected. Under simulated
intestinal conditions however, no shifts in bacterial concentrations were detected. Experiments with individual medium
components thought that the effect of sphingosine is very easily neutralized by BSA, stearic acid and surfactants. Based on
our results, d-erythro-sphingosine would only be active when protonated and its mode of action would imply electrostatic
attraction to the bacteria and disruption of membrane integrity. In conclusion, the application of sphingosine is limited to
specific environments, as activity was very sensitive to inhibition. Yet, because of its broad spectrum membrane disrupting
activity, it could be very useful under controlled conditions.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Sphingosine; Antibacterial; Mode of action; SHIME; Intestine; Sphingolipids
0168-1605/$ - see front matter D 2005 Elsevier B.V. All rights reserved.
doi:10.1016/j.ijfoodmicro.2005.05.007
* Corresponding author. Tel.: +32 9 264 59 76; fax: +32 9 264 62
48.
E-mail address: [email protected] (W. Verstraete).
1. Introduction
Sphingolipids are estimated to constitute 0.01 to
0.02% of the human diet with a yearly intake per
capita of about 115 g, with dairy products being the
primary source of uptake (Vesper et al., 1999).
robiology 105 (2005) 59–70

S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–7060
Despite this low consumption, sphingolipids and their
metabolites, ceramide and sphingosine (SPH), are
highly bioactive molecules with multiple beneficial
effects on human health, e.g. cancer inhibition
(Schmelz et al., 2000) and the inhibition of cholesterol
absorption (Eckhardt et al., 2001; Jiang et al., 2001).
Another possible effect would be the protective capa-
cities of sphingolipids against bacterial toxins and
infection by bacteria or viruses. Food sphingolipids
can compete for cellular binding sites on the intestinal
mucosa (Bibel et al., 1992b; Fantini et al., 1997).
Because microbial adherence is often the first step
in infection (Ofek et al., 2003), this competition
could protect against food borne pathogens. Infants
consuming milk supplemented with gangliosides, a
specific group of sphingolipids, had significantly
lower E. coli and higher Bifidobacterium sp. counts
in their faeces (Rueda et al., 1998). Beside this, SPH
could also have a direct bactericidal effect. Free SPH
acts as a natural antimicrobial barrier of the human
skin (Arikawa et al., 2002; Bibel et al., 1993, 1995)
and has been shown to strongly decrease the concen-
tration of several food borne pathogens at low con-
centration (Sprong et al., 2001, 2002).
d-erythro-sphingosine (SPH) is the most prevalent
sphingoid base in mammalian tissues and dairy pro-
ducts. The amino group of this component is usually
substituted with long-chain fatty acids to produce
ceramides. The addition of a polar phosphocholine
produces sphingomyelin, the main sphingolipid pre-
sent in foods (Vesper et al., 1999). During intestinal
passage, sphingomyelin is partially digested to cera-
mide and SPH by human sphingomyelinases and
ceramidases (Liu et al., 2000; Lundgren et al., 2001;
Nilsson and Duan, 1999) and colon bacteria could
also be able to transform sphingolipids (Nyberg et
al., 1997). Therefore, SPH can be present in the
large intestine and exert bactericidal effects and
protect against intestinal pathogens. Sprong et al.
(2001) showed that 25 AM SPH tested in saline,
minimally gave a 4 log reduction of the Escherichia
coli, Salmonella enteritidis, Campylobacter jejuni,
and Listeria monocytogenes concentrations, whereas
sphingomyelin and ceramide showed no antibacterial
effects. However, the colon harbours up to 1014
microorganisms from approximately 400 different
species, which strongly influence human health (Mac-
Farlane and Cummings, 1999). If SPH does not act
selectively towards pathogens, but also kills beneficial
intestinal bacteria like lactobacilli and bifidobacteria,
this could negatively influence human health.
Up to now, bactericidal activity of SPH has only
been tested in saline (Sprong et al., 2001). Under
intestinal conditions however, the environment is
very complex with changing conditions in the different
parts of the colon (Macfarlane et al., 1992). It is known
that environmental conditions strongly influence the
efficacy of an antibacterial component (Bessems,
1998). Another important factor influencing activity
towards different bacterial species is the mode of
action of SPH. Next to a possible detergent activity,
interference with a protein-kinase system analogous to
mammalian cells is proposed (Bibel et al., 1992a), but
no clear evidence in this direction was obtained.
Therefore, bactericidal activity of SPH further has to
be characterised in terms of its mode of action and
activity under the physiological conditions of the intes-
tine. In this work, selectivity of action towards three
pathogenic strains, being E. coli, Enterobacter aero-
genes BE1 and Clostridium perfringens, was tested by
comparing results with experiments using lactobacilli
and bifidobacteria. This was done using plate counts as
well as with Live/Dead analysis combined with flow
cytometry, allowing rapid screening of the degree of
bactericidal effect. In addition, a modification of the
Simulator of the Human Intestinal Microbial Ecosys-
tem (SHIME) (De Boever et al., 2000; Molly et al.,
1993) was used to assess the efficacy under more
relevant conditions and the influence of different
environmental conditions. This way, a more clear
view of the mode of action and activity of dietary
SPH as a natural antibacterial agent was obtained.
2. Materials and methods
2.1. Bacterial strains and growth conditions
As examples of pathogenic strains, a multidrug
resistant clinical isolate E. coli ESBL112 (LMG
22096), E. aerogenes BE1 (LMG 22092) (De Gheldre
et al., 2001) and C. perfringens (LMG 11264) were
used to investigate the spectrum of bactericidal activ-
ity of SPH and Lactobacillus acidophilus (LMG
13550), Lactobacillus amylovorus (LMG 18197),
Bifidobacterium longum (LMG 11047) and Bifido-

S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–70 61
bacterium bifidum (LMG 11582) were used as exam-
ples of health promoting strains. All strains were
obtained from the Belgian Coordinated Cultures of
Microorganisms (BCCMk/LMG, Ghent, Belgium).
The culture conditions of the different strains are
described in Table 1. Aerobic experiments with liquid
cultures were performed in test tubes, whereas anae-
robic liquid incubations were performed in flasks,
sealed with rubber tops and anaerobiosis was obtained
by flushing the flasks with N2 during 20 cycles of 2
min 700 mbar overpressure and 900 mbar underpres-
sure. Anaerobic plate incubations were performed in
jars, containing AnaeroGenk bags (Oxoid, Hamp-
shire, UK) to remove oxygen.
2.2. Antimicrobial susceptibility testing
2.2.1. In vitro assay
d-erythro-sphingosine was obtained from AvantiRPolar lipids, Inc. (Alabaster, Alabama). Stock solu-
tions (4 mM) were prepared in absolute ethanol and
stored at �20 8C. Bactericidal activity was tested in
saline (8.5 g/l NaCl) at pH 7. Incubates contained
3.8% (v/v) ethanol but this did not affect bactericidal
activity of SPH (data not shown).
The activity of SPH was tested in triplicate at
concentrations of 0, 25, 50, 100 and 150 AM. Bacter-
ial cultures were tenfold diluted in saline to avoid
matrix effects. 300 Al diluted culture (F106 CFU/
ml) was then transferred to an eppendorf tube contain-
ing 662 Al saline. Finally, SPH stock solution was
added together with absolute EtOH so that to every
tube a total volume of 38 Al was dosed. Samples were
vortexed and incubated for 2 h at 37 8C under aerobic
or anaerobic conditions, depending on the growth
conditions for the different strains (Table 1).
Table 1
Bacterial strains tested for susceptibility to sphingosine (SPH), the cultu
Bacteria Broth
E. coli Luria-Bertani (A)
E. aerogenes BE1
C. perfringens Reinforced Clostridial Medium (AN)
L. acidophilus De Man Rogosa Sharpe (AN)
L. amylovorus
B. bifidum De Man Rogosa Sharpe+0.5 g/l L-cystein-HCl
B. longum
Cultures were incubated aerobically (A) or anaerobically after flushing w
2.2.2. Plate counts
After incubation, tenfold dilution series of the dif-
ferent samples were prepared in saline and plated in
triplicate. Plates were incubated at 37 8C under con-
ditions as described in Table 1. Results were presented
as mean logarithmic decrease in bacterial concentra-
tion in function of the concentration of SPH. To
achieve this, logarithmic bacterial concentrations
were subtracted from the concentration in the control
samples without SPH. Mean logarithmic decrease in
bacterial concentration and standard deviation were
calculated for the different incubates and plotted.
2.2.3. Live/Dead analysis
For Live/Dead analysis, a Cyan flow cytometer
(DakoCytomation, Glostrup, Denmark) was used,
equipped with a 50 mW solid state blue Sapphire
laser (488 nm) as the excitation light source. The
Live/DeadR BacLightk bacterial viability kit (L-
13152, Molecular Probes, Eugene, Oregon) was
applied for staining the bacteria. A stock solution of
the SYTO 9 and propidium iodide nucleic acid stains
was prepared by diluting both stains in 5 ml of filter
sterilized sheath fluid (DakoCytomation, Glostrup,
Denmark). From this, a staining solution was made
by diluting the stock solution 1/10.
After incubation, the same samples as used for
plating were tenfold diluted in filter sterilised saline.
Aliquots of 500 Al cell suspensions and 500 Al stain-ing solution were mixed and incubated for 20 min at
room temperature in the dark prior to analysis. Live
(+injured) and dead stained cells were discriminated
based on a FL1 (530/40 nm) vs. FL3 (613/20 nm)
excitation plot. Results were presented as mean loga-
rithmic decrease in function of the concentration of
SPH. For this, the ratio of live to dead cells was
re conditions for liquid cultures and media used for plate counts
Agar Time (h)
Luria-Bertani (A) 24
Tryptose Sulfite Cycloserine agar (AN) 72
Rogosa agar (AN) 72
(AN) Raffinose-Bifidobacterium (AN) 24
ith N2 (AN).

S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–7062
calculated and divided by the ratio of the control
sample without SPH. After logarithmic transforma-
tion, mean logarithmic decrease in Live/Dead ratio
and standard deviation were calculated for the differ-
ent incubates and plotted.
2.3. Simulator of the human intestinal microbial
ecosystem (SHIME)
The reactor setup was based on the adult SHIME
(De Boever et al., 2000; Molly et al., 1993) with this
difference that only the first colon reactor was used.
The SHIME was designed to maintain a microbial
community that is representative for that in the
human intestine. In this setup, the SHIME consisted
of three reactors that simulate the stomach and duo-
denum (reactor 1), jejunum and ileum (reactor 2) and
the ascending colon (reactor 3). Reactor design, inocu-
lum preparation, reactor startup and feeding scheme
were previously described by De Boever et al. (2000).
After a microbial community stabilization period of 3
weeks, the content of reactor 3 was split in two sepa-
rate reactors (250 ml), the experiment and control
reactor, to ensure identical microbial communities at
the beginning of the experiment. For a period of 11
days, SPH (AvantiR Polar Lipids, Alabaster, Alabama,
USA) diluted in 100% ethanol was daily added to the
experiment reactor in a final concentration of 50 AMand an equal amount of 100% ethanol was added to the
control reactor. On days 1, 3, 6, 8, 10 and 11 samples
were taken from the experiment and control reactor
and aerobic mesophilic bacteria, anaerobic mesophilic
bacteria, coliforms, Staphylococcus spp., Enterococ-
cus spp. and Clostridium spp. were plated (Table 2).
2.3.1. Effect of pH
To evaluate the influence of pH on bactericidal
activity of SPH, experiments were conducted with
Table 2
Analysed bacterial groups from the SHIME microbial community, to
Bacterial group Medium
Aerobic mesophilic bacteria Brain Heart Infusion agar
Anaerobic mesophilic bacteria Brain Heart Infusion agar+
Coliforms Mc. Conkey agar
Staphylococcus spp. Mannitol Salt agar
Enterococcus spp. Enterococcus agar
Clostridium spp. Tryptose Sulfite Cycloserin
E. coli (F106 CFU/ml) in buffered saline at a con-
centration of 25 AM SPH. With a 0.1 M phosphate, a
0.1 M BIS-TRIS and a 0.1 M TRIS buffer, pH was
respectively set to 5.5, 6, 7 and 8; to pH 5.5, 6 and 7
and to pH 7.5, 8 and 9. After 2 h of incubation, the
logarithmic effect of SPH was calculated as the
inverse value of the logarithmic decrease, as described
above, obtained by plating and Live/Dead analysis.
2.3.2. Effect of different medium components
To evaluate whether the medium in which the
experiment is conducted, influences the degree of
bactericidal capacity of SPH, an experiment was con-
ducted with E. coli (F106 CFU/ml) in Luria-Bertani
broth at a concentration of 25 AM SPH.
Saline containing different concentrations of glu-
cose, lactose, Tween 20, bovine serum albumin, stea-
ric acid or sphingomyelin was used for experiments to
examine the effect of individual medium components.
Sphingomyelin was obtained from AvantiR Polar
lipids, Inc. (Alabaster, Alabama), the other com-
pounds were purchased from Sigma-Aldrich (St.
Louis, MO). E. coli (F106 CFU/ml) was used as
test organism at a concentration of 25 AM SPH.
After 2 h incubation, the logarithmic effect of SPH
was calculated as the inverse value of the logarithmic
decrease, as described above, obtained by plating and
Live/Dead analysis.
2.3.3. Haemolytic activity
To 165 Al phosphate-buffered saline, 5 Al stock
solution (4 mM in absolute EtOH) of SPH or sphin-
gomyelin diluted in 100% ethanol was added in a final
concentration of 5, 10, 25, 50, 100, 200, 400, 600 and
800 AM SPH or sphingomyelin. After incubation for 1
min at 37 8C in a shaking water bath, 30 Al of redblood cells of sheep origin (10% suspension, ICN,
Brussels, Belgium) were added. Simultaneously, red
gether with the isolation media and incubation conditions used
Condition Time (h)
Aerobic 24
0.5 g l�1 cystein Anaerobic 72
Aerobic 24
Aerobic 48
Aerobic 48
e agar Anaerobic 72
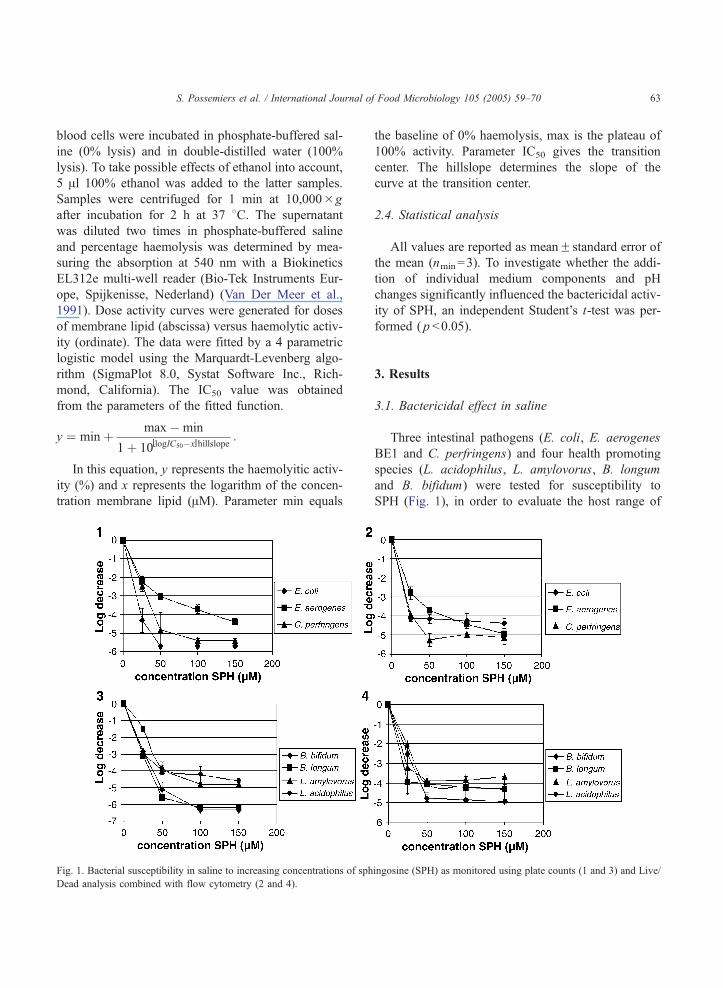
S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–70 63
blood cells were incubated in phosphate-buffered sal-
ine (0% lysis) and in double-distilled water (100%
lysis). To take possible effects of ethanol into account,
5 Al 100% ethanol was added to the latter samples.
Samples were centrifuged for 1 min at 10,000�g
after incubation for 2 h at 37 8C. The supernatant
was diluted two times in phosphate-buffered saline
and percentage haemolysis was determined by mea-
suring the absorption at 540 nm with a Biokinetics
EL312e multi-well reader (Bio-Tek Instruments Eur-
ope, Spijkenisse, Nederland) (Van Der Meer et al.,
1991). Dose activity curves were generated for doses
of membrane lipid (abscissa) versus haemolytic activ-
ity (ordinate). The data were fitted by a 4 parametric
logistic model using the Marquardt-Levenberg algo-
rithm (SigmaPlot 8.0, Systat Software Inc., Rich-
mond, California). The IC50 value was obtained
from the parameters of the fitted function.
y ¼ minþ max�min
1þ 10tlogIC50�xbhillslope:
In this equation, y represents the haemolyitic activ-
ity (%) and x represents the logarithm of the concen-
tration membrane lipid (AM). Parameter min equals
Fig. 1. Bacterial susceptibility in saline to increasing concentrations of sph
Dead analysis combined with flow cytometry (2 and 4).
the baseline of 0% haemolysis, max is the plateau of
100% activity. Parameter IC50 gives the transition
center. The hillslope determines the slope of the
curve at the transition center.
2.4. Statistical analysis
All values are reported as meanF standard error of
the mean (nmin=3). To investigate whether the addi-
tion of individual medium components and pH
changes significantly influenced the bactericidal activ-
ity of SPH, an independent Student’s t-test was per-
formed ( p b0.05).
3. Results
3.1. Bactericidal effect in saline
Three intestinal pathogens (E. coli, E. aerogenes
BE1 and C. perfringens) and four health promoting
species (L. acidophilus, L. amylovorus, B. longum
and B. bifidum) were tested for susceptibility to
SPH (Fig. 1), in order to evaluate the host range of
ingosine (SPH) as monitored using plate counts (1 and 3) and Live/

Fig. 2. Bactericidal activity of sphingosine (SPH) towards E. coli in
saline (E) and Luria-Bertani broth (n) as monitored using plate
counts and Live/Dead analysis combined with flow cytometry.
S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–7064
the antibacterial agent and the selectivity towards
pathogens. Plate counts and Live/Dead analysis gave
similar results. All tested strains were strongly
reduced (N4 log units) at 150 AM. In graph 3, the
apparent difference between the tested lactobacilli and
bifidobacteria is due to a difference in detection limit
rather than a difference in sensitivity. Lactobacilli are
spread on top of the plates (100 Al/plate), giving a
detection limit of 100 CFU/ml; for bifidobacteria
pouring plates are used (1 ml/plate) with a detection
limit of 10 CFU/ml. At 25 AM E. coli was reduced the
most and L. acidophilus the least, but in general no
real differences in the effect of SPH were noted. This
means SPH was active against Gram-positive as well
as Gram-negative bacteria, to pathogens as well as
health promoting bacteria. At equivalent concentra-
tions, neither ceramide nor sphingomyelin exerted
antibacterial activity against the tested species (data
not shown).
3.2. Antibacterial effect under complex conditions
When the bactericidal effect of SPH on E. coli in
saline was compared with the activity in Luria-Bertani
broth (Fig. 2), inhibition of the activity clearly
occurred in Luria-Bertani broth. While a 4 log reduc-
tion was seen in saline at 25 AM using plate counts as
well as Live/Dead analysis, almost no decrease in
bacterial concentration occurred in Luria-Bertani
broth.
To assess the antibacterial activity of SPH in com-
plex environments like the intestinal tract, a final con-
centration of 50 AM SPH was dosed to the experiment
reactor of the modified SHIME system for a period of
11 days. However, no difference in the evolution of the
concentrations of the aerobic mesophilic bacteria,
anaerobic mesophilic bacteria, coliforms, Staphylo-
coccus spp., Enterococcus spp. and Clostridium spp.
were detected between the experiment and control
reactor, indicating that SPH exerted no bactericidal
activity under simulated intestinal conditions.
3.3. Influence of individual medium components
To assess the origin of this inhibitory effect of
environmental conditions, the influence of specific
medium components was investigated. Different con-
centrations of individual medium components were
added to saline and the experiment was repeated (Fig.
3). Even at a concentration of 25 g/l, glucose and
lactose did not inhibit the capacity of SPH to decrease
bacterial concentrations, indicating that medium car-
bohydrates do not cause the noted matrix effects.
Proteins on the other hand, interact with SPH, when
supplied as bovine serum albumin (BSA), and dis-
played a concentration dependent inhibitory effect. At
5 g/l, BSA already decreased the effect by 50% and at
25 g/l no activity of SPH was seen anymore. Sphin-
gomyelin, a membrane lipid and containing the SPH
backbone, also influenced the activity of SPH at low
concentration. At 120 AM sphingomyelin, SPH only
showed a minor activity. Stearic acid and the surfac-
tant Tween 20 finally, showed no inhibitory effect
until a threshold concentration of respectively 14
AM and 0.4 mM was reached.
Live/Dead analysis proved to be a good alternative
to plate counts to monitor the antibacterial effect of
SPH. Both techniques showed the same trends
describing the effect of individual medium compo-
nents. This was confirmed after calculating the overall
correlation between the two techniques for these
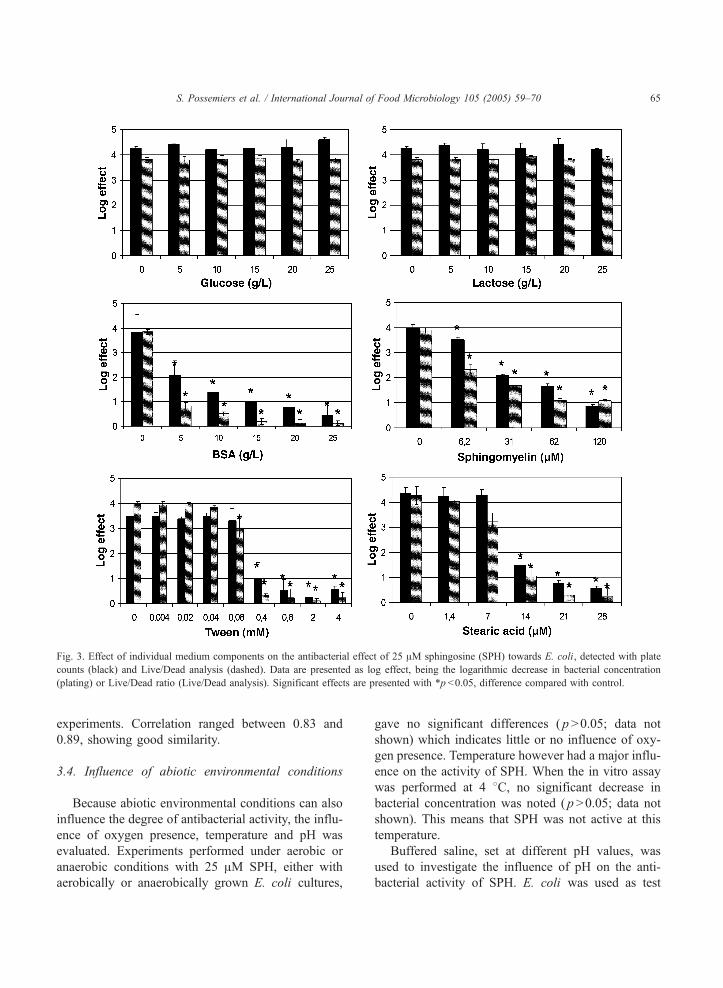
Fig. 3. Effect of individual medium components on the antibacterial effect of 25 AM sphingosine (SPH) towards E. coli, detected with plate
counts (black) and Live/Dead analysis (dashed). Data are presented as log effect, being the logarithmic decrease in bacterial concentration
(plating) or Live/Dead ratio (Live/Dead analysis). Significant effects are presented with *p b0.05, difference compared with control.
S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–70 65
experiments. Correlation ranged between 0.83 and
0.89, showing good similarity.
3.4. Influence of abiotic environmental conditions
Because abiotic environmental conditions can also
influence the degree of antibacterial activity, the influ-
ence of oxygen presence, temperature and pH was
evaluated. Experiments performed under aerobic or
anaerobic conditions with 25 AM SPH, either with
aerobically or anaerobically grown E. coli cultures,
gave no significant differences ( p N0.05; data not
shown) which indicates little or no influence of oxy-
gen presence. Temperature however had a major influ-
ence on the activity of SPH. When the in vitro assay
was performed at 4 8C, no significant decrease in
bacterial concentration was noted ( p N0.05; data not
shown). This means that SPH was not active at this
temperature.
Buffered saline, set at different pH values, was
used to investigate the influence of pH on the anti-
bacterial activity of SPH. E. coli was used as test

0
20
40
60
80
100
5 10 25 50 100 200 400 600 800
Concentration ( M)
% ly
sis
SPH
SM
Fig. 5. Haemolytic activity of sphingosine (SPH) and sphingomye-
lin in phosphate-buffered saline after 2 h incubation at 37 8C. ForSPH, a 50% inhibitory concentration (IC50) of 174 AM was calcu-
lated, while for sphingomyelin the IC50 was never reached within
the range of tested concentrations.
S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–7066
organism and was incubated at 37 8C for 2 h with 25
AM SPH (Fig. 4). Using BIS-TRIS and TRIS buffers,
high and constant bactericidal activity was seen at the
normal pH range of the large intestine (5.5–7), but at
pH 8 and 9 the effect rapidly decreased with one log
unit per pH unit. However, when a phosphate buffer
was used to set the pH from 5.5 to 8, almost no
activity of SPH was seen in comparison with the
other buffers, indicating a negative effect of phosphate
on the antibacterial activity of SPH.
3.5. Haemolytic activity of sphingosine
To investigate whether the antibacterial activity of
SPH is based on membrane disruption, the haemolytic
activity of different concentrations of SPH and sphin-
gomyelin was tested in saline under the same incuba-
tion conditions as the in vitro assay to monitor
bactericidal activity. For 2 h, red blood cells were
Live/Dead
0
1
2
3
4
5
5,5 6 9pH
Lo
g e
ffec
t
BIS-TRIS
TRIS
Phosphate
Plating
0
1
2
3
4
5
Lo
g e
ffec
t
**
**
7 8
5,5 6 9pH7 8
BIS-TRIS
TRIS
Phosphate
Fig. 4. Plate counts and Live/Dead analysis of the activity of 25 AMSPH towards E. coli in pH-buffered saline using a BIS-TRIS, TRIS
or phosphate buffer. Data are presented as log effect, being the
logarithmic decrease in bacterial concentration (plating) or Live/
Dead ratio (Live/Dead analysis). With the phosphate buffer only
very small effects were noted. Using the TRIS-based buffers, a
significant decrease in effect was seen at high pH (*p b0.05,
difference compared with saline at pH 7).
incubated with increasing concentrations of SPH or
sphingomyelin (Fig. 5). The baseline of 0% lysis and
the maximum of 100% lysis were set using PBS buffer
and double distilled water respectively, with 5 Alethanol added as control. While SPH induced strong
haemolysis at increasing concentrations, sphingomye-
lin showed only minor activity at very high concentra-
tion. As calculated with SigmaPlot 8.0, the IC50 of
SPH was 174 AM. While 800 AM SPH induced 100%
lysis, sphingomyelin did not even give 20% lysis at
this concentration and the IC50 was never reached.
4. Discussion
The intestinal microbial community plays a signif-
icant role in processes as food digestion, bioconver-
sion of endogenous or exogenous compounds,
immunomodulation, and prevention from infection
by intestinal pathogens (Gibson and Roberfroid,
1995; Hart et al., 2002). Therefore, strategies improv-
ing the composition towards beneficial bacteria have
gained much attention. One way of reaching this,
could be preventing the colonisation of the gut by
harmful bacteria like food pathogens. Sprong et al.
(2001, 2002) proposed bovine fat milk components to
be important for this purpose. Based on the strong
bactericidal activity towards pathogens in saline, they
stated that sphingolipids like SPH may enhance the
resistance to certain food-borne intestinal infections.
This strong activity was in good correlation with our

Fig. 6. Structure of sphingosine (SPH). For bactericidal activity
SPH has to be positively charged.
S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–70 67
results. In saline, all tested strains, Gram-positives and
Gram-negatives, were very susceptible to SPH after a
short incubation period. However, when tested under
more realistic matrix conditions, no bactericidal activ-
ity was detected. Tests with individual medium com-
ponents showed that SPH is very sensitive to
inhibition and therefore not active in complex
matrices like in the intestine. Based on the combina-
tion of our results, the mode of action of SPH would
include electrostatic attraction and disruption of the
bacterial membrane.
The first reports on the importance of SPH as an
antimicrobial component originate from skin research.
Based on in vitro and in vivo research, Bibel et al.
(1992a, 1993) stated that SPH could have an impor-
tant barrier function protecting the human skin from
colonisation by pathogens. SPH would act as a broad
spectrum antimicrobial agent with antibacterial as
well as antifungal activity. Antibacterial activity was
mainly restricted to Gram-positive bacteria, with no
effect towards E. coli (Bibel et al., 1992b). Because
sphingolipids are present in many foods, including
soy and dairy products (Ahn and Schroeder, 2002;
Molkentin, 2000), and because intestinal metabolism
of sphingomyelin towards SPH has been shown in
vitro and in vivo (Liu et al., 2000; Nyberg et al.,
1997), SPH could also be important to protect against
intestinal colonisation by pathogens. Therefore,
Sprong et al. (2001) investigated the susceptibility
of a number of food pathogens to increasing concen-
trations of SPH. Yet, in contrast with the results of
Bibel et al. (1992a), Gram-positive as well as Gram-
negative bacteria, including E. coli, were sensitive to
low concentrations. Our results are in good concor-
dance with the results obtained by Sprong et al.
(2001), with a similar decrease in concentration of
E. coli when tested in saline at 25 AM SPH (F4 log
CFU/ml). This strain even was the most sensitive of
all tested strains. Any differences in species sensitivity
may be due to differences in cell wall composition,
like the outer membrane, because SPH needs to reach
the cytoplasmic membrane to exert its activity (Gia-
cometti et al., 1999; Ishikawa et al., 2002). In general,
in these experiments only small differences in sensi-
tivity occurred, with no selectivity towards Gram-
positive or Gram-negative bacteria. Important to
notice is that SPH was also active against health
promoting bacteria like lactobacilli and bifidobacteria.
Bibel et al. (1992a) formulated the hypothesis that
a protein-kinase system analogous to mammalian cells
is involved in the antibacterial activity of SPH. This
was based on the fact that they observed calcium
dependency, which could imply enzyme mediation.
However, neither in our experiments nor in the experi-
ments of Sprong et al. (2001), any necessity of cal-
cium was detected. On the contrary, the following
arguments indicate the mode of action is primary
based on disruption of cell membrane integrity.
Firstly, the activity of SPH rapidly decreases above
pH 8 (Fig. 4). Because the amino group of SPH has a
pKa of 7.99, with deprotonation at higher pH, the pH
effect indicates this group has to be protonated for
activity (Fig. 6). Being positively charged, SPH shows
high structural similarity with quaternary ammonium
compounds (QACs). The mode of action of the latter
is based on adsorption to the cell surface, diffusion
through the cell wall and disruption of the cytoplasmic
membrane. This probably leads to secondary leakage
of metabolites and enzymes, killing the bacteria (Mer-
ianos, 2001; Paulus, 1993). Next to QACs, high
structural similarity occurs with cationic peptides,
which also act on bacterial membranes (Giacometti
et al., 1999). Secondly, in this research we used the
Live/DeadR BacLight bacterial viability kit which has
been extensively used to study different topics, such
as permeabilization of cheese starters using mutano-
lysin (Bunthof et al., 2001), disinfectant activity of
quaternary ammonium components (Langsrud and
Sundheim, 1996) and antibiotics like phosphomycin
(Jacobsen et al., 1997). All compounds applied in
these studies have in common that their mode of
action implies membrane disruption and the fact that
the kit was shown to be efficient to study the activity
of SPH, could imply a similar mode of action. Thirdly,
SPH exhibited a very rapid antimicrobial effect. When
E. coli was incubated with 25 AM SPH for 20 min,
already a 3.5 log reduction was noted (data not
shown), which indicated a rapid attack of the bacterial
,

S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–7068
integrity rather than influencing an enzymatic system.
This conclusion was also stated for QACs (Ahlstrom
et al., 1999; Russell et al., 1999) and cationic peptides
(Giacometti et al., 1999). Finally, SPH has a high
surface activity. To test membrane damaging activity,
a haemolysis test is often performed (Gandhi and
Cherian, 2000; Soderlind et al., 2003; Takechi et al.,
2003). Any lysis of the erythrocytes used in this test
can be attributed to membrane damaging effects
because these cells lack organelles and metabolic
systems (De Boever and Verstraete, 1999). The low
IC50 of SPH (174 AM) in comparison with for
instance bile salts (IC50~1–5 mM) (De Boever and
Verstraete, 1999) here also suggests membrane dis-
rupting activity. These arguments lead to the hypoth-
esis that the positively charged SPH is attracted to the
bacterial cell, which is negatively charged at physio-
logical pH, by electrostatic attraction and hydrophobic
interactions and after reaching the cytoplasmic mem-
brane, the long hydrophobic chain penetrates into the
lipid bilayer, forming channels. The reduced activity
at 4 8C can then be due to the lower membrane
fluidity and the concomitant inability to perforate
the lipid bilayer.
Despite the strong activity of SPH in saline, the
hypothesis as proposed by Sprong et al. (2001), being
a bactericidal effect of SPH in the human intestine,
does not seem to hold. When tested in a matrix
simulating the intestinal conditions, no effect of 50
AM SPH was detected, which suggests a lack of
activity under intestinal conditions. Experiments
with individual medium components showed a very
high sensitivity of SPH to the presence of matrix
components. This is also seen for other antibacterial
compounds like QACs, and based on the structure and
the proposed mode of action of SPH, most of this
inhibition can be explained. BSA molecules are glob-
ular proteins with hydrophilic and hydrophobic
domains, having a net negative charge at pH 7. In
case of QACs, a rapid interaction with the hydrophilic
regions occurs, followed by hydrophobic interactions
of the tail structure (Ahlstrom et al., 1999; Merianos,
2001). The same interaction could happen between
SPH and BSA or in general with negatively charged
structures, inhibiting the attraction to the bacteria and
subsequent bactericidal activity. The importance of
this effect is confirmed by the fact that, almost no
activity of SPH was seen using a negatively charged
phosphate buffer, whereas in a positively charged
buffer the activity only decreased above pH 8. The
latter could also be an explanation for the fact that
Bibel et al. (1992a) observed a necessity of Ca2+. This
ion could neutralize the negative charges of the pep-
tone buffer and thereby reduce the inhibitory effects
rather than the fact it would be necessary for the
bactericidal activity of SPH. Non-ionic and anionic
surfactants are known to have antagonistic effects on
the activity of cationic surface active agents, when
dosed in concentrations higher than the critical micelle
concentration (CMC), by inclusion of active ingredi-
ents in the micelles or by complex formation (Paulus,
1993). As the CMC of Tween 20 is about 0.06 mM,
this explains the strong inhibitive effect from a con-
centration of 0.08 mM. Stearate also forms micelles
with negatively charged surfaces, thereby attracting
SPH. Again, the effect starts above the CMC of
stearate (F6 AM) (Cawthern et al., 1997). Finally,
negatively charged phospholipids like sphingomyelin
are known to inhibit the activity of QACs (Merianos,
2001), which was also the case for SPH.
As it is now accepted that foods like dairy products
contain a number of health promoting components,
there is an increasing interest in the relation between
nutrition and human health (Molkentin, 2000; Wal-
zem et al., 2002). In this context, sphingolipids are
very important, having a number of positive effects
like the prevention of cancer and hypercholesterole-
mia and the inhibition of bacterial adhesion (Vesper et
al., 1999). Because SPH has been shown to exert
strong and rapid antimicrobial effects towards a
large variety of species, it could act as a natural
antimicrobial agent. As no activity was seen in the
complex chemical matrix of the SHIME, the combi-
nation of the factors interacting with SPH, as men-
tioned above, limit the application of SPH as an active
antimicrobial compound in the human intestine, espe-
cially because colonic concentrations of SPH are
shown to be in the range of only a few AM (Nyberg
et al., 1997). However, many arguments favor further
research on this topic. For instance, the potential for
SPH as topical antibiotic should be further explored as
antibacterial activity on the human skin has been
shown. SPH could also be very useful as protective
coating material on body implants or as preservative
in defined preparations like pharmaceuticals. Further-
more, in this research we tested the most commonly

S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–70 69
present SPH in dairy products, but over 60 different
backbones are known, such as phytosphingosine or
sphinganine (Karlsson, 1970). Their little structural
differences could lead to different activities or stabi-
lity. Therefore, the applicability of sphingosines as
antibacterial agents needs further investigation.
Acknowledgements
This work was supported by a PhD grant (aspirant)
for Sam Possemiers from the Fund for Scientific
Research-Flanders (Fonds voor Wetenschappelijk
Onderzoek (FWO) Vlaanderen).
The authors gratefully thank Els Jolie for technical
assistance and Nico Boon, Karel Decroos, Tom Van
de Wiele and Kristof Verthe for critically reviewing
this manuscript. Special thanks go to Hans Nelis for
intellectual assistance about surface active agents.
References
Ahlstrom, B., Thompson, R.A., Edebo, L., 1999. Loss of bacter-
icidal capacity of long-chain quaternary ammonium compounds
with protein at lowered temperature. APMIS 107, 606–614.
Ahn, E.H., Schroeder, J.J., 2002. Bioactive sphingolipids are con-
stituents of soy and dairy products. Journal of Food Science 67,
522–524.
Arikawa, J., Ishibashi, M., Kawashima, M., Takagi, Y., Ichikawa,
Y., Imokawa, G., 2002. Decreased levels of sphingosine, a
natural antimicrobial agent, may be associated with vulnerability
of the stratum corneum from patients with atopic dermatitis to
colonization by Staphylococcus aureus. Journal of Investigative
Dermatology 119, 433–439.
Bessems, E., 1998. The effect of practical conditions on the efficacy
of disinfectants. International Biodeterioration & Biodegrada-
tion 41, 177–183.
Bibel, D.J., Aly, R., Shinefield, H.R., 1992a. Antimicrobial activ-
ity of sphingosines. Journal of Investigative Dermatology 98,
269–273.
Bibel, D.J., Aly, R., Shinefield, H.R., 1992b. Inhibition of microbial
adherence by sphinganine. Canadian Journal of Microbiology
38, 983–985.
Bibel, D.J., Aly, R., Shah, S., Shinefield, H.R., 1993. Sphingo-
sines—antimicrobial barriers of the skin. Acta Dermato-Vener-
eologica 73, 407–411.
Bibel, D.J., Aly, R., Shinefield, H.R., 1995. Topical sphingolipids in
antisepsis and antifungal therapy. Clinical and Experimental
Dermatology 20, 395–400.
Bunthof, C.J., van Schalkwijk, S., Meijer, W., Abee, T., Hugenholtz,
J., 2001. Fluorescent method for monitoring cheese starter
permeabilization and lysis. Applied and Environmental Micro-
biology 67, 4264–4271.
Cawthern, K.M., Narayan, M., Chaudhuri, D., Permyakov, E.A.,
Berliner, L.J., 1997. Interactions of alpha-lactalbumin with fatty
acids and spin label analogs. Journal of Biological Chemistry
272, 30812–30816.
De Boever, P., Verstraete, W., 1999. Bile salt deconjugation by
Lactobacillus plantarum 80 and its implication for bacterial
toxicity. Journal of Applied Microbiology 87, 345–352.
De Boever, P., Deplancke, B., Verstraete, W., 2000. Fermentation by
gut microbiota cultured in a simulator of the human intestinal
microbial ecosystem is improved by supplementing a soygerm
powder. Journal of Nutrition 130, 2599–2606.
De Gheldre, Y., Struelens, M.J., Glupczynski, Y., De Mol, P., Maes,
N., Nonhoff, C., Chetoui, H., Sion, C., Ronveaux, O., Vanee-
choutte, M., 2001. National epidemiologic surveys of Entero-
bacter aerogenes in Belgian hospitals from 1996 to 1998.
Journal of Clinical Microbiology 39, 889–896.
Eckhardt, E.R., Wang, D.Q.H., Donovan, J.M., Carey, M.C., 2001.
Dietary sphingomyelin (SM) significantly inhibits intestinal
cholesterol (Ch) absorption by lowering thermodynamic activity
(TA) of Ch in bile salt (BS) mixed micellar solution. Gastro-
enterology 120, A679–A680.
Fantini, J., Hammache, D., Delezay, O., Yahi, N., AndreBarres, C.,
RicoLattes, I., Lattes, A., 1997. Synthetic soluble analogs of
galactosylceramide (GalCer) bind to the V3 domain of HIV-1
gp120 and inhibit HIV-1-induced fusion and entry. Journal of
Biological Chemistry 272, 7245–7252.
Gandhi, V.M., Cherian, K.M., 2000. Red cell haemolysis test as an
in vitro approach for the assessment of toxicity of karanja oil.
Toxicology in Vitro 14, 513–516.
Giacometti, A., Cirioni, O., Barchiesi, F., Del Prete, M.S., Scalise,
G., 1999. Antimicrobial activity of polycationic peptides. Pep-
tides 20, 1265–1273.
Gibson, G.R., Roberfroid, M.B., 1995. Dietary modulation of the
human colonic microbiota-introducing the concept of prebiotics.
Journal of Nutrition 125, 1401–1412.
Hart, A.L., Stagg, A.J., Frame, M., Graffner, H., Glise, H., Falk, P.,
Kamm, M.A., 2002. Review article: the role of the gut flora in
health and disease, and its modification as therapy. Alimentary
Pharmacology & Therapeutics 16, 1383–1393.
Ishikawa, S., Matsumura, Y., Katoh-Kubo, K., Tsuchido, T., 2002.
Antibacterial activity of surfactants against Escherichia coli
cells is influenced by carbon source and anaerobiosis. Journal
of Applied Microbiology 93, 302–309.
Jacobsen, C.N., Rasmussen, J., Jakobsen, M., 1997. Viability stain-
ing and flow cytometric detection of Listeria monocytogenes.
Journal of Microbiological Methods 28, 35–43.
Jiang, Y.Z., Noh, S.K., Koo, S.I., 2001. Egg phosphatidylcholine
decreases the lymphatic absorption of cholesterol in rats. Journal
of Nutrition 131, 2358–2363.
Karlsson, K.-A., 1970. On the chemistry and occurrence of sphin-
golipid longchain bases. Lipids 5, 6–43.
Langsrud, S., Sundheim, G., 1996. Flow cytometry for rapid
assessment of viability after exposure to a quaternary
ammonium compound. Journal of Applied Bacteriology 81,
411–418.
Liu, J.J., Nilsson, A., Duan, R.D., 2000. Effects of phospholipids on
sphingomyelin hydrolysis induced by intestinal alkaline sphin-

S. Possemiers et al. / International Journal of Food Microbiology 105 (2005) 59–7070
gomyelinase: an in vitro study. Journal of Nutritional Biochem-
istry 11, 192–197.
Lundgren, P., Nilsson, A., Duan, R.D., 2001. Distribution and
properties of neutral ceramidase activity in rat intestinal tract.
Digestive Diseases and Sciences 46, 765–772.
MacFarlane, G.T., Cummings, J.H., 1999. Probiotics and prebiotics:
can regulating the activities of intestinal bacteria benefit health?
British Medical Journal 318, 999–1003.
Macfarlane, G.T., Gibson, G.R., Cummings, J.H., 1992. Compar-
ison of fermentation reactions in different regions of the human
colon. Journal of Applied Bacteriology 72, 57–64.
Merianos, J.J., 2001. Surface-active agents. In: Block, S.S. (Ed.),
Disinfection, Sterilization, and Preservation. Lippincott, Wil-
liams and Wilkins, Philadelphia, pp. 283–320.
Molkentin, J., 2000. Occurrence and biochemical characteristics of
natural bioactive substances in bovine milk lipids. British Jour-
nal of Nutrition 84, S47–S53.
Molly, K., Woestyne, M.V., Verstraete, W., 1993. Development of a
5-step multichamber reactor as a simulation of the human
intestinal microbial ecosystem. Applied Microbiology and Bio-
technology 39, 254–258.
Nilsson, A., Duan, R.D., 1999. Alkaline sphingomyelinases and
ceramidases of the gastrointestinal tract. Chemistry and Physics
of Lipids 102, 97–105.
Nyberg, L., Nilsson, A., Lundgren, P., Duan, R.D., 1997. Localiza-
tion and capacity of sphingomyelin digestion in the rat intestinal
tract. Journal of Nutritional Biochemistry 8, 112–118.
Ofek, I., Hasy, D.L., Sharon, N., 2003. Anti-adhesion therapy of
bacterial diseases: prospects and problems. FEMS Immunology
and Medical Microbiology 38, 181–191.
Paulus, W., 1993. Microbiocides for the protection of materials.
Chapman and Hall, London. 496 pp.
Rueda, R., Sabatel, J.L., Maldonaldo, J., Molina-Font, J.A., Gil, A.,
1998. Addition of gangliosides to an adapted milk formula
modifies levels of fecal Escherichia coli in preterm newborn
infants. Journal of Pediatrics 133, 90–94.
Russell, A.D., Hugo, W.B., Ayliffe, G.A.J., 1999. Principles and
Practice of Disinfection, Preservation and Sterilization. Black-
well Science, London. 826 pp.
Schmelz, E.M., Sullards, M.C., Dillehay, D.L., Merrill, A.H., 2000.
Colonic cell proliferation and aberrant crypt foci formation are
inhibited by dairy glycosphingolipids in 1,2-dimethylhydrazine-
treated CF1 mice. Journal of Nutrition 130, 522–527.
Soderlind, E., Wollbratt, M., von Corswant, C., 2003. The usefulness
of sugar surfactants as solubilizing agents in parenteral formula-
tions. International Journal of Pharmaceutics 252, 61–71.
Sprong, R.C., Hulstein, M.F.E., Van der Meer, R., 2001. Bacterici-
dal activities of milk lipids. Antimicrobial Agents and Che-
motherapy 45, 1298–1301.
Sprong, R.C., Hulstein, M.F.E., van der Meer, R., 2002. Bovine
milk fat components inhibit food-borne pathogens. International
Dairy Journal 12, 209–215.
Takechi, M., Doi, K., Wakayama, Y., 2003. Biological activities of
synthetic saponins and cardiac glycosides. Phytotherapy
Research 17, 83–85.
Van Der Meer, R., Termont, D., De Vries, H.T., 1991. Differential
effects of calcium ions and calcium phosphate on cytotoxicity of
bile acids. American Journal of Physiology 260, G142–G147.
Vesper, H., Schmelz, E.M., Nikolova-Karakashian, M.N., Dillehay,
D.L., Lynch, D.V., Merrill, A.H., 1999. Sphingolipids in food
and the emerging importance of sphingolipids to nutrition.
Journal of Nutrition 129, 1239–1250.
Walzem, R.L., Dillard, C.J., German, J.B., 2002. Whey compo-
nents: Millennia of evolution create functionalities for mamma-
lian nutrition: What we know and what we may be overlooking.
Critical Reviews in Food Science and Nutrition 42, 353–375.