characterization of rab-like4, the first identified rab-like protein from trypanosoma cruzi with...
TRANSCRIPT

www.elsevier.com/locate/ybbrc
Biochemical and Biophysical Research Communications 333 (2005) 808–817
BBRC
Characterization of RAB-like4, the first identified RAB-likeprotein from Trypanosoma cruzi with GTPase activity q
Fabiane Pereira Ramos a, Julia Rolao Araripe a, Turan Peter Urmenyi a,Rosane Silva a, Narcisa Leal Cunha e Silva c, Carlos Frederico Leite Fontes d,
Jose Franco da Silveira e, Edson Rondinelli a,b,*
a Laboratorio de Metabolismo Macromolecular Firmino Torres de Castro, Instituto de Biofısica Carlos Chagas Filho,
Universidade Federal do Rio de Janeiro, CCS, Cidade Universitaria, Rio de Janeiro 21949-900, Brazilb Departamento de Clınica Medica, Universidade Federal do Rio de Janeiro, Cidade Universitaria, Rio de Janeiro 21949-900, Brazil
c Laboratorio de Ultraestrutura Cellular Hertha Meyer, Instituto de Biofısica Carlos Chagas Filho,
Universidade Federal do Rio de Janeiro, CCS, Cidade Universitaria, Rio de Janeiro 21949-900, Brazild Laboratorio de Membranas Transportadoras, Instituto de Bioquımica Medica, Universidade Federal do Rio de Janeiro,
CCS, Cidade Universitaria, Rio de Janeiro 21941-590, Brazile Departamento de Microbiologia, Imunologia e Parasitologia, Escola Paulista de Medicina, UNIFESP, R. Botocatu, 862, Sao Paulo 04023-062, Brazil
Received 25 May 2005Available online 13 June 2005
Abstract
RAB proteins, which belong to the RAS superfamily, regulate exocytic and endocytic pathways of eukaryotic cells, controllingvesicle docking and fusion. Few RAB proteins have been identified in parasites. Molecular markers for cellular compartments areimportant to studies concerning about the protein traffic in Trypanosoma cruzi, the causal agent of Chagas disease. In this work, wedescribe the characterization of TcRABL4, the first RAB-like gene identified in T. cruzi (GenBank Accession No.: AY371275), pres-ent as a single-copy gene. TcRABL4 contains all five consensus RAB motifs but lacks cysteine residues at the C terminus, which areessential to isoprenylation, an absolute prerequisite for membrane association of these proteins. TcRABL4 is a functional GTPasethat is able to bind and hydrolyze GTP, and its gene is transcribed as a single 1.2 kb mRNA in epimastigotes. TcRABL4 appears tobe differentially regulated in the three cell forms of the parasite, and the protein is not associated to membranes, unlike other RABproteins. It is possible that TcRABL4 may be a member of a novel family of small GTPases.� 2005 Elsevier Inc. All rights reserved.
Keywords: GTPase; RAS superfamily; RAB-like; Trypanosomatids; Endocytic pathway
Trypanosoma cruzi, causative agent of Chagas� dis-ease, is a digenetic parasite which alternates in its biolog-ical cycle between vertebrate and invertebrate hosts[1,2]. It has replicative (epimastigotes and amastigotes)and non-replicative (trypomastigotes) forms. During
0006-291X/$ - see front matter � 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.bbrc.2005.05.183
q This work was supported by CNPq and FAPERJ. Note. Nucleo-tide sequence data reported in this paper are available in the GenBankdatabase under Accession No. AY371275 (TcRAB-like, completegenomic sequence).* Corresponding author. Fax: +55 21 2280 8293.E-mail address: [email protected] (E. Rondinelli).
its developmental stages, the parasite needs exogenousgrowth factors, taken by endocytosis, through the flage-lar pocket and cytostome. Although the T. cruzi endo-cytic pathway is poorly understood, it is known to behighly developed in epimastigotes. The cytostome, pres-ent in the cell�s anterior region, close to the flagellarpocket, is the preferential entry site of bovine albuminand transferrin [3], while the reservosomes are the endof the pathway [4].
To date, more than 60 distinct RAB proteins havebeen identified and are found in all eukaryotes, including

F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817 809
yeast, plants, and mammals, and most of them areexpressed ubiquitously, while some have a tissue-specificpattern of expression [5–7]. RAB family members exhib-it 30% of identity with the other RAS superfamily mem-bers, the most conserved regions being those directlyinvolved in GTP binding and hydrolysis (G1–G5motifs). The less conserved regions are present at theprotein�s N and C termini. The C terminus has one ortwo cysteine residues prone to isoprenylation, which iscarried out in all the members of the RAS superfamily,a crucial post-translational modification that enablesRAB proteins to target and associate to cell membranes[8–10].
Novel RAB proteins are being described, which dis-play features that diverge from the other family mem-bers. Some of them are being named RAB-likeproteins. Such differences may represent more recentevolutionary modifications of the RAB family, whereGTP/GDP binding motifs are present and assumingnew functions or more specialized ones [11].
Parasite Genome Projects made possible the identifi-cation of small GTPases in Trypanosoma brucei [12–15]and Leishmania major [16]. The identification of GTPas-es in T. cruzi [17] allowed the isolation and characteriza-tion of two T. cruzi RAB proteins. TcRAB11 isexpressed in epimastigotes and spheromastigotes and ispresent in reservosomes. Its mammalian homolog coun-terpart is localized on early endosomes [18]. TcRAB7was cloned and characterized in our laboratory [19],and the protein shown to be located at the Golgi com-plex [20]. The difference observed in the subcellularlocalization of TcRAB11 and TcRAB7, compared totheir orthologs in mammalian cells, emphasizes the needto better characterize and understand the T. cruzi endo-cytic compartments. A better understanding depends onthe development of new molecular markers for the endo-cytic pathway.
The present work describes the characterization ofTcRABL4, the first RAB-like gene identified in T. cruzi.TcRABL4 is differentially expressed in the three formsof the parasite, and its protein product exhibits a highGTP hydrolysis rate. The TcRABL4 protein also showspeculiar structural features when compared to otherRAB proteins, such as the absence of cysteine residuesat the C terminus. TcRABL4 may be an atypical RABprotein or could be a member of an uncharacterizedfamily of small GTPases, and is the first functionalRAB-like protein identified in T. cruzi.
Materials and methods
Cultivation of Trypanosoma cruzi. Epimastigotes of T. cruzi clonesCL Brener and DM28c were cultured at 29 �C in liver infusion tryptose(LIT) medium [21], supplemented with 10% heat-inactivated calf ser-um. Metacyclic trypomastigotes were obtained in vitro using M16medium (68.4 mM NaCl, 7.4 mM KCl, 56.4 mM Na2HPO4, 0.2%
glucose, 0.125% tryptose, 2.5% inactivated bovine serum, 0.001%hemin, 100 U/mL penicillin, and 100 mg/mL streptomycin) [22] andpurified according to [23]. The resulting cell populations contained atleast 90% of metacyclic trypomastigotes. Spheromastigote forms wereobtained in vitro as previously described [24].
Cloning of the TcRABL4 gene of Trypanosoma cruzi. A cDNAclone corresponding to expressed sequence tag (EST) TENF0549(GenBank Accession No. AA676030), generated from the T. cruzi
epimastigote normalized cDNA library [25,26], contained the partialTcRABL4 coding sequence and was used as a probe to isolate thecomplete coding sequence from a CL Brener genomic library [27]. TheTcRABL4 gene structure obtained was compared to preliminary CLBrener genomic sequence data accessed via http://Tcruzidb.org(Database version 3.0, data release of July 26 2004). Most of thegenomic data were provided by the TIGR-SBRI-KI SequencingConsortium (TSK-TSC) supported by NIH Grants AI45038, AI45061,and AI45039.
Nucleic acid manipulation and analysis. Plasmid DNA and genomicDNA were extracted according to standard protocols [28]. Sequencingwas performed using fluorescent cycle sequencing with the DYEnamicET Dye Terminator kit (Amersham Biosciences) in a MegaBACE 1000DNA Analysis System (Amersham Biosciences). Sequence alignmentswere performed using the BioEdit and ClustalX programs. Chromo-somal DNA was extracted and processed for pulse field gradient gelelectrophoresis (PFGE) as previously described [29]. Genomic DNAwas extracted using the phenol/chloroform method [28]. Total RNAwas extracted from exponentially growing epimastigotes using stan-dard guanidinium–isothiocyanate procedures [30,31]. Total RNA wasresolved on 1.5% agarose–formaldehyde denaturing gels and trans-ferred to nylon membranes. Hybridizations were carried out accordingto standard protocols [28]. Probes were labeled by random priming(Random Primer Labeling System kit, Gibco-BRL). Untranslated re-gions (UTRs) of TcRABL4 mRNA were amplified from epimastigotetotal RNA by RT-PCR according to standard procedures [28]. The 5 0
UTR was obtained using a sense primer directed to the T. cruzi mini-exon sequence (5 0 ggA Tgg AAT TCA gTT TCT gTA CTA TAT Tg3 0) and an antisense primer to a region immediately before the G3domain, (5 0gCC gCA TCT gCA gCg TgC gTg gCg ACC ATT gTC TC3 0), containing a EcoRI and a PstI restriction sites, respectively(underlined). The 3 0 UTR was obtained using the 3 0 RACE System(Invitrogen) and a sense primer directed to the TcRABL4 coding se-quence (5 0 CAU CAU CAU CAU ATg CAC TgA AgC ATg CgAACA g 3 0). The PCR conditions were: 25 cycles consisting of 94 �Cdenaturing for 1 min, 60 �C annealing for 1 min, and 72 �C extensionfor 2 min, followed by a 10 min-extension step. A sixth of the PCRproducts was resolved on a 2% agarose gel. The 5 0 UTR fragmentswere subcloned into pGEM4Z vector (Promega) and the 3 0UTRfragment was subcloned pAMP1 vector (Invitrogene). The 5 0 UTRclones were confirmed by sequencing.
Recombinant TcRABL4 and Western blot analysis. The TcRABL4coding sequence was amplified by PCR from genomic DNA usingoligos corresponding to the N-terminal (5 0 ACg CAT gAA TTC ATggCg TAT CTg CgC 3 0) and C-terminal (5 0gAA CTC gAg TTA CTTCAT CTg ggA AAg CTg 3 0) regions, containing the EcoRI and XhoIrestriction sites, respectively (underlined). Cycling conditions were:94 �C denaturing for 1 min, 60 �C annealing for 1 min, and 72 �Cextension for 2 min, followed by a 10 min extension step. The purified600 bp-PCR product was subcloned into pGEX 5X-1 plasmidexpression vector (Amersham Biosciences). After confirmation bysequencing, TcRABL4 fusion protein was superexpressed in E. coli
BL21 cells and purified following Bulk and Redipack GST PurificationModules kit (Amersham Biosciences). The expression product wasanalyzed by 12.5% SDS–PAGE [32]. A synthetic peptide (Y V D A YH K R I D Q L S Q M K) based on the predicted amino acid sequence(amino acid residues from 168 to 183) of TcRABL4 was synthesized(Bio synthesis) and used to raise a specific polyclonal serum in rabbits.Western blots were performed according to standard protocols as
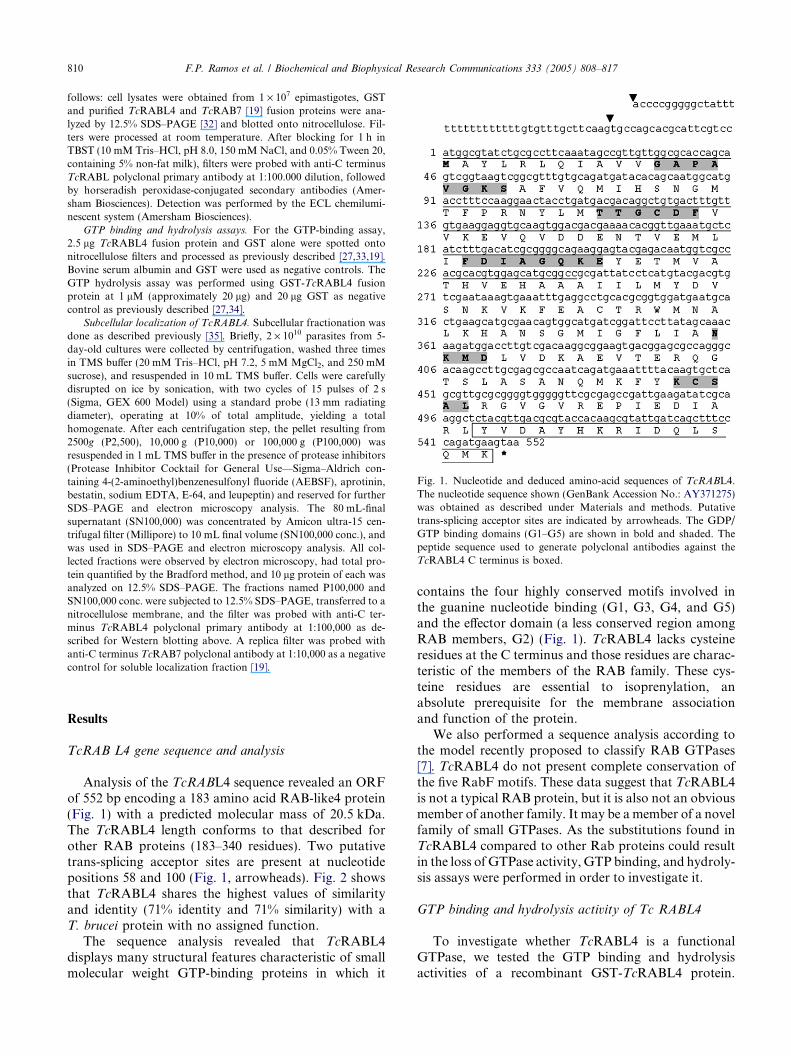
Fig. 1. Nucleotide and deduced amino-acid sequences of TcRABL4.The nucleotide sequence shown (GenBank Accession No.: AY371275)was obtained as described under Materials and methods. Putativetrans-splicing acceptor sites are indicated by arrowheads. The GDP/GTP binding domains (G1–G5) are shown in bold and shaded. Thepeptide sequence used to generate polyclonal antibodies against theTcRABL4 C terminus is boxed.
810 F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817
follows: cell lysates were obtained from 1 · 107 epimastigotes, GSTand purified TcRABL4 and TcRAB7 [19] fusion proteins were ana-lyzed by 12.5% SDS–PAGE [32] and blotted onto nitrocellulose. Fil-ters were processed at room temperature. After blocking for 1 h inTBST (10 mM Tris–HCl, pH 8.0, 150 mMNaCl, and 0.05% Tween 20,containing 5% non-fat milk), filters were probed with anti-C terminusTcRABL polyclonal primary antibody at 1:100.000 dilution, followedby horseradish peroxidase-conjugated secondary antibodies (Amer-sham Biosciences). Detection was performed by the ECL chemilumi-nescent system (Amersham Biosciences).
GTP binding and hydrolysis assays. For the GTP-binding assay,2.5 lg TcRABL4 fusion protein and GST alone were spotted ontonitrocellulose filters and processed as previously described [27,33,19].Bovine serum albumin and GST were used as negative controls. TheGTP hydrolysis assay was performed using GST-TcRABL4 fusionprotein at 1 lM (approximately 20 lg) and 20 lg GST as negativecontrol as previously described [27,34].
Subcellular localization of TcRABL4. Subcellular fractionation wasdone as described previously [35]. Briefly, 2 · 1010 parasites from 5-day-old cultures were collected by centrifugation, washed three timesin TMS buffer (20 mM Tris–HCl, pH 7.2, 5 mM MgCl2, and 250 mMsucrose), and resuspended in 10 mL TMS buffer. Cells were carefullydisrupted on ice by sonication, with two cycles of 15 pulses of 2 s(Sigma, GEX 600 Model) using a standard probe (13 mm radiatingdiameter), operating at 10% of total amplitude, yielding a totalhomogenate. After each centrifugation step, the pellet resulting from2500g (P2,500), 10,000 g (P10,000) or 100,000 g (P100,000) wasresuspended in 1 mL TMS buffer in the presence of protease inhibitors(Protease Inhibitor Cocktail for General Use—Sigma–Aldrich con-taining 4-(2-aminoethyl)benzenesulfonyl fluoride (AEBSF), aprotinin,bestatin, sodium EDTA, E-64, and leupeptin) and reserved for furtherSDS–PAGE and electron microscopy analysis. The 80 mL-finalsupernatant (SN100,000) was concentrated by Amicon ultra-15 cen-trifugal filter (Millipore) to 10 mL final volume (SN100,000 conc.), andwas used in SDS–PAGE and electron microscopy analysis. All col-lected fractions were observed by electron microscopy, had total pro-tein quantified by the Bradford method, and 10 lg protein of each wasanalyzed on 12.5% SDS–PAGE. The fractions named P100,000 andSN100,000 conc. were subjected to 12.5% SDS–PAGE, transferred to anitrocellulose membrane, and the filter was probed with anti-C ter-minus TcRABL4 polyclonal primary antibody at 1:100,000 as de-scribed for Western blotting above. A replica filter was probed withanti-C terminus TcRAB7 polyclonal antibody at 1:10,000 as a negativecontrol for soluble localization fraction [19].
Results
TcRAB L4 gene sequence and analysis
Analysis of the TcRABL4 sequence revealed an ORFof 552 bp encoding a 183 amino acid RAB-like4 protein(Fig. 1) with a predicted molecular mass of 20.5 kDa.The TcRABL4 length conforms to that described forother RAB proteins (183–340 residues). Two putativetrans-splicing acceptor sites are present at nucleotidepositions 58 and 100 (Fig. 1, arrowheads). Fig. 2 showsthat TcRABL4 shares the highest values of similarityand identity (71% identity and 71% similarity) with aT. brucei protein with no assigned function.
The sequence analysis revealed that TcRABL4displays many structural features characteristic of smallmolecular weight GTP-binding proteins in which it
contains the four highly conserved motifs involved inthe guanine nucleotide binding (G1, G3, G4, and G5)and the effector domain (a less conserved region amongRAB members, G2) (Fig. 1). TcRABL4 lacks cysteineresidues at the C terminus and those residues are charac-teristic of the members of the RAB family. These cys-teine residues are essential to isoprenylation, anabsolute prerequisite for the membrane associationand function of the protein.
We also performed a sequence analysis according tothe model recently proposed to classify RAB GTPases[7]. TcRABL4 do not present complete conservation ofthe five RabF motifs. These data suggest that TcRABL4is not a typical RAB protein, but it is also not an obviousmember of another family. It may be amember of a novelfamily of small GTPases. As the substitutions found inTcRABL4 compared to other Rab proteins could resultin the loss ofGTPase activity, GTP binding, and hydroly-sis assays were performed in order to investigate it.
GTP binding and hydrolysis activity of Tc RABL4
To investigate whether TcRABL4 is a functionalGTPase, we tested the GTP binding and hydrolysisactivities of a recombinant GST-TcRABL4 protein.

Fig. 2. Multiple alignment of T. cruzi TcRABL4 with orthologs from various species. The GDP/GTP binding domains (G1–G5) are indicated byhorizontal black bars. Degrees of identity are shown to the right of each sequence. TbGTPase, putative small GTP-binding protein of T. brucei(chromosome 3 RPCI93 from T. brucei genome database on http://www.tigr.org), HsRAB-like4, RAB-like4 of Homo sapiens (GenBank AccessionNo.: Q9BW83), MmRAB13, RAB13 of Mus musculus (GenBank Accession No.: NP_080953.1), HsRAB15, RAB15 of Homo sapiens (GenBankAccession No.: P59190), BbRAB1, RAB1 of Babesia bovis (GenBank Accession No.: AAP85297.1), TpRAB1, RAB1 of Theileria parva (GenBankAccession No.: AF107762_1), ScYPT1p, YPT1p of Saccharomyces cerevisiae (GenBank Accession No.: NP_116615.1), and HsRAB23, RAB23 ofHomo sapiens (GenBank Accession No.: AF498951_1. The sequence alignment was performed using the programs BioEdit and ClustalX. Theposition of identical aminoacids when compared to the T. cruzi sequence is represented by gray shading.
F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817 811
As can be seen in Fig. 3, the recombinant TcRABL4 isable to specifically bind GTP, since the GTP binding sig-nal showed by GST-TcRABL4 was significantly higherthan that of either GST alone or BSA and no signal
was detected with ATP (data not shown). In addition,the recombinant TcRABL4 was able to hydrolyzeGTP at 0.67 nmol mg protein�1 min�1 (Fig. 4A) andexhibited a 0.29% hydrolysis rate in 60 min with respect

Fig. 3. GTP-binding activity of TcRABL4. Recombinant TcRABL4(2.5 lg) was spotted onto nitrocellulose filters. Bovine serum albumin(BSA) and GST (2.5 lg) were used as negative controls. The filter wasincubated with [a�32P]GTP (3000 Ci/mmol, Amersham Biosciences)to a final concentration of 1 lCi/mL for 2 h at room temperature.After six 5-min washes in binding buffer, the filter was dried andautoradiographed.
Fig. 4. GTP hydrolysis activity of TcRABL4. One micromolar (20 lg)GST-TcRABL4 fusion protein and GST were incubated with 0.05 mM[c-32P]GTP (5000 Ci/mmol, Amersham Biosciences) at 37 �C. Twentymicrograms BSA and 20 lg GST were used as negative controls. Twohundred microliter aliquots were taken from the reaction after 5, 15,30, and 60 min of hydrolysis. The 32Pi content of a 0.04 mL sample wasdetermined by liquid scintillation counting. (A) Plot showing reactionvelocity in nmol mg�1 min�1 versus time in minutes; (B) Plot showingcpm versus hydrolysis time in minutes.
Fig. 5. Sizing of TcRABL4 transcript and the TcRABL4 mRNAexpression on epimastigotes, espheromastigotes, and trypomastigotescells. (A) Northern blot analysis of TcRABL4 transcripts. Total RNA(5 lg) from epimastigotes was separated on a formaldehyde–agarosegel, blotted to a nylon membrane and hybridized to the TcRABL4coding region. (B) RT-PCR analysis of TcRABL4 5 0 UTR ofepimastigote (1), metacyclic trypomastigote (2), and spheromastigote(3) forms. PCR reactions were performed with either non-diluted (a) orfive-times diluted (b) cDNA template. The negative control (�)corresponds to equivalent RT-PCR reactions without reverse trans-criptase. One hundred pb Ladder (Amersham Biosciences) was used asmolecular weight standard (MW).
812 F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817
to the 1.1 · 108 total cpm used in the assay (Fig. 4B). Weestimate that, in our assay, TcRABL4 hydrolyzed GTPat a rate of about 13.4 · 10�3 mol min�1 mol�1 of theprotein.
TcRAB L4 gene structure and expression
To verify the genomic organization of TcRABL4,genomic Southern blot analysis was performed and re-vealed that TcRABL4 is present as a single copy gene(data not shown). Preliminary genomic sequencedata from T. cruzi clone CL Brener, available athttp://Tcruzidb.org, are consistent with our results. Todetermine the chromosomal location of the TcRABL4locus, a PFGE analysis was performed. The two allelesof TcRABL4 are located at homologous chromosomesof approximately 1.43 Mb of size (chromosomal bandXIII) (data not shown).
To investigate TcRABL4 gene expression, Northernblot analysis was performed. A single TcRABL4 tran-script of about 1.2 kb was detected in epimastigotes(Fig. 5A), in agreement to what has been described forRAB mRNAs of other trypanosomatids [36,13,18,19,27]. The TcRABL4 transcript could not be observedby Northern blot analysis in metacyclic trypomastigoteor spheromastigote forms (data not shown). We there-fore have performed a RT-PCR assay to detect the 5 0
UTR of the mRNA in the three forms of the parasite.Fig. 5B shows amplified products of 346 and 396 bp inall parasite forms, corresponding to the two main spliceleader acceptor sites. A 250 bp-fragment was alsoobserved in epimastigotes, but sequencing revealed thatit does not contain TcRABL4 sequence and is due tononspecific amplification. The Northern blot andRT-PCR analyses suggest that TcRABL4 is develop-mentally regulated.

F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817 813
Analysis of TcRABL4 on subcellular fractions
To investigate the expression and subcellular localiza-tion of TcRABL4 in T. cruzi epimastigotes, a syntheticpeptide was generated based on the variable C-terminussequence and used to obtain specific polyclonal antibod-ies. Western blot analysis revealed a single 20 kDa poly-peptide in epimastigote cell lysates (Figs. 6A and C), inagreement with the predicted molecular weight ofTcRABL4. Recombinant GST-TcRABL4 was alsodetected as a 49 kDa polypeptide. GST alone and
Fig. 6. Specificity of the C-terminus TcRABL4 antiserum. Westernblot analysis of TcRABL4 antiserum generated by a peptide from theC-terminus. (A) Whole cell lysates of T. cruzi epimastigotes (1 · 107
cells) from CL Brener and DM28c were separated by 12.5% SDS–PAGE, transferred to nitrocellulose membranes, and probed with anti-TcRABL4 C-terminus antiserum. (B) Recombinant fusion proteinsGST-TcRABL4, GST-TcRAB7, and GST were separated by 12.5%SDS–PAGE, transferred to a nitrocellulose membrane, and probedwith anti-TcRABL4 C-terminus antiserum at 1:100,000. (C) Specificityof the TcRABL4 C-terminal: CL Brener and DM28c epimastigotewhole cell lysates (1 · 107 cells) were separated by 12.5% SDS–PAGE,transferred to nitrocellulose membranes, and probed with: (C) anti-TcRABL4 C-terminus antiserum, diluted 1:100,000, pre-incubatedovernight with no peptide. (D) Anti-TcRABL4 C-terminus antiserum,diluted 1:100,000, pre-incubated overnight with TcRABL4 C-terminuspeptide. (E) Anti-TcRABL4 C-terminus antiserum, diluted 1:100,000,pre-incubated overnight with TcRAB7 C-terminus peptide. Molecularweights in kilodaltons are shown to the left.
Fig. 7. Localization of TcRABL4 in subcellular fractions fromepimastigotes. Ten micrograms of protein from soluble (SN100,000)and microsomal (P100,000) fractions was separated on a 12.5% SDS–PAGE, transferred to a nitrocellulose membrane, and probed withanti-TcRABL4 C-terminus antiserum diluted 1:100,000. Molecularweight in kilodaltons is shown to the left.
GST-TcRAB7 were not recognized by the anti-TcRABL4 antiserum (Figs. 6B and C). In addition,pre-incubation of the antiserum with the TcRABL4 syn-thetic peptide showed that it successfully blocked thebinding of the anti-TcRABL4 antibodies (Fig. 6D). Anunrelated synthetic peptide, corresponding to TcRAB7,was unable to block antibody binding (Fig. 6E). Weconclude that the anti-TcRABL4 serum obtained is ableto specifically recognize the protein.
To determine if TcRABL4 is associated to membranesin epimastigotes, a subcellular fractionation was per-formed. Total protein extract was used as control of thecells before fractionation. Fraction SN100,000 corre-sponds to the soluble fraction (Fig. 7, lane 1). FractionP100,000 (Fig. 7, lane 2) corresponds to the microsomalcontent, containing all the cell membranes. The Westernblot analysis using anti-TcRABL4 antiserum shows thatTcRABL4 is mostly present in the soluble fractionSN100,000 (Fig. 7, lane 1). These results suggest thatTcRABL4 performs its function in the cytosol.
Discussion
TcRABL4 shows low levels of sequence similarity toother small GTPases. This analysis reinforces thehypothesis that those proteins act in different steps in

814 F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817
the endo and exocytic pathways [37]. RAB-like4 pro-teins were identified in silico in several genomes but theirfunction remains unclear. RAB-like4 fall slightly outsidethe main RAB cluster [38]. RAB23 and RAB15 areRABs exclusively or predominantly expressed in the cen-tral nervous system [39]. RAB15 is involved in the recy-cling pathway from endosomes to plasma membrane [6].Recent work showed the involvement of RAB23 in neu-ral patterning, highlighting the importance of RABs indevelopment [40]. Recently, a RAB23 homologous hasbeen identified in T. brucei as a nuclear-associatedRAB protein [41], but after sequence analysis it wasnot assigned as a TcRABL4 counterpart. There is noan evident link between mammalian RAB23 andRAB15 and their possible orthologs in trypanosomatids.Future functional studies will elucidate the specific roleof TcRABL4 in T. cruzi.
Despite being the most variable of the four domains,G1 showed full conservation in TcRABL4, as well asdomain G4. A threonine residue conserved among allmembers of RAS superfamily is also present inTcRABL4 effector domain G2, although it also containstwo substitutions from the consensus, isoleucine to thre-onine and isoleucine to cysteine. The G2 domain is theputative effector region of the RAB proteins, being in-volved in interactions with specific effector molecules[39]. A RAB-like protein from Entamoeba histolytica
was described to contain some substitutions in this do-main [42].
The highly conserved G3 domain of TcRABL4 con-tains two substitutions (tryptophan to phenylalanineand threonine to isoleucine), as well as an inserted lysineresidue. Although these modifications do not excludeTcRABL4 from the RAS superfamily, G2 is the mostconserved domain in the RAB family. The threonine res-idue of the G2 domain is highly conserved among mostsmall G-proteins, but a few members of RAB family(Dictyostelium RABE, mammalian RAB29, RAB32,and RAB39, and, most recently, a RAB protein fromDrosophila melanogaster) whose functions in vivo havenot been elucidated, also show this unique substitution[43–45]. The same occurs within region G5 and a glu-tamic acid to lysine substitution. A RAB-like proteinof E. histolytica also exhibits divergences in the G5 do-main, but still retains the ability to bind and hydrolyzeGTP [42].
The TcRABL4 sequence analysis, according to themodel recently proposed to classify RAB GTPases [7],showed that the protein presents regions that do notconform to the RabF motifs. The RAB family motif 1(RabF1, IGVDF) is not completely conserved inTcRABL4. The isoleucine and valine residues are substi-tuted in TcRABL4 by threonine and cysteine, respec-tively. However, it contains the glycine residue, whichis almost absolutely conserved in RAB, ARF, andRAN proteins. TcRABL4 also does not show conserva-
tion in RabF2 (KLQIW) having only the isoleucine con-served. The tryptophan residue is not RAB-specific,being conserved in all small GTPases except RAS.TcRABL4 does not contain any conserved residue inthe region corresponding to RabF3 (RFrSiT). The pro-tein contains only the alanine conserved in the last posi-tion from RabF4 (YYRGA). The RabF5 (LVYDIT) isnot completely represented in TcRABL4, which con-tains the residues leucine, tyrosine, and aspartic acid.TcRABL4 exhibits low overall identity to the rest ofthe RAB family (average identity 30%), which suggeststhat TcRABL4 is not a typical RAB protein. It maybe a member of a novel family of small GTPases.
Compared to TcRAB5 [27], TcRABL4 has a highGTPasic activity, of about twice that of TcRAB5. Ourdata are in agreement with what has been describedfor several human RAB proteins [46] and other GTPasessuch as YPT1, RHO, and RAS proteins [33,47].TcRABL4 has a higher intrinsic GTPase activity thanthat of RAS-related proteins, which is typically low,4.2 · 10�4 [48]. Mammalian RAB5 is among the fastestsmall GTPases with an intrinsic kcat of 2 · 10�1 [49],while RAS has a kcat of 2.8 · 10�2 [50]. Our resultsstrongly suggest that TcRABL4 is a functional GTPase.
The hypervariable tails of RAB proteins are crucialfor the specific membrane association of the proteins[8]. Some RABs (such as HsRAB8 and HsRAB23) havea single cysteine residue within a CAAX box (C, cys; A,aliphatic; X, any), a motif commonly observed in theRAS and RHO families [10]. Recent studies have ob-served some RABs with unusual C terminus. For exam-ple, ARA6 from Arabidopsis thaliana exhibits putativeN-terminal myristoylation and palmitoylation motifs,but no C-terminal prenylation motifs, and these motifsare also present in similar plant proteins [51]. Anotherpeculiar RAB protein is RAB24, which is thought tobe cytosolic [52]. Human RAB39 localizes to the Gol-gi-associated apparatus and does not contain the cys-teine residues, but a S-R-A C-terminal microbodytargeting signal was found, which may indicate that itssubcellular localization is special [53]. RAB-like4 pro-teins do not possess the cysteine residues and functionalstudies have not yet been performed. The unusual casesmay represent recent evolutions of the RAB familywhere motifs not normally present in this family arerecruited to provide for new functions [51].
TcRABL4 does not present any prenylatable cysteineresidues in the C terminus. Other GTPases assigned asRAB-like proteins, such as human RABL2A andRABL2B [54], similarly do not present any prenylatablecysteine residues in the C terminus. Sequence compari-son between RABL2A, RABL2B, and TcRABL4 tothe rest of the RAB family shows that TcRABL4 exhib-its a higher level of identity compared to the other pro-teins. Although most RAB proteins possess C-terminalprenylation signals, these are distinct from those found

F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817 815
in RAS and RHO GTPases. One RAB subfamily mem-ber, RAB35, has an adjacent polybasic motif. SeveralRABs have potential N-terminal myristoylation sites[38]. Computational analysis showed that TcRABL4lacks serine or tyrosine phosphorylation sites as wellas posttranslational modification signals. The absenceof C terminus cysteine residues is consistent withTcRABL4 being mostly present in the cytosol.
It is believed that interactions between RAS-like GTP-ases and effectors/regulators depend upon the switch re-gions. Since RAB GTPases play important roles invesicle trafficking, it is most likely that RAB-like proteinsregulate similar events in T. cruzi. Evolutionarily con-served RABs tend to be expressed in all cell and tissuetypes and regulate fundamental transport pathways,whereas less conserved family members, such asTcRABL4, may function in the many specialized path-ways found in different cell types. As trypanosomes be-came a separate branch during eukaryotes evolution,some features about its gene expression regulation arevery particular, as TcRABL4 seems to be. Future studiesshall focus on these variations in regard to the role ofGTPases in T. cruzi and other evolutionarily ancientorganisms.
The organization of TcRABL4 as a single copy genein the T. cruzi genome also agrees with what has beendescribed for RAB genes in other organisms [55] andfor other RAS superfamily genes identified in thisorganism: TcRAB11 [18], TcRAB7 [19], TcRAB5 [27],TcRHO1 [56], and TcARF1[57]. TcRABL4 is the firstRAB-like encoding gene identified in T. cruzi that hasshown particular features differing from those describedin the literature, not only for trypanosomes, but also forhigher eukaryotes.
Alterations on mRNA levels were also observed inRAB genes of other trypanosomatids. Until the demon-stration thatTbRAB11 gene expression is developmental-ly regulated [58], all GTPases identified in T. brucei werethought to be constitutively expressed [36,12,13,15]. Sub-sequently, other RAB genes were recognized as beingdevelopmentally regulated. The two RAB5 isoforms ofT. brucei are expressed in the bloodstream and procyclicforms of the parasite, even though endocytosis is not veryactive in the latter form. mRNA levels of TbRAB5A areconstitutive while TbRAB5B presents greater expressionin bloodstream forms, suggesting thatTbRAB5B is devel-opmentally regulated [13]. Other T. brucei RAB geneswere also shown to be developmentally regulated, suchas TbRAB31 [59] and TbRAB18 [60].
GTPase genes from T. cruzi have also been shown tobe developmentally regulated. TcRAB5 transcript wasshown recently to be less abundant in trypomastigotes[27]. Although TcRAB7 transcript levels remain unal-tered in epimastigotes and trypomastigotes, it is 5.7times less abundant in spheromastigotes [19]. TcARF1mRNA is abundant in epimastigotes, detectable in
spheromastigotes, and only barely detectable in trypom-astigotes [52]. It has been shown that non-proliferativetrypomastigotes exhibit lower protein content whencompared to proliferative epimastigotes forms [61].Epimastigotes present a high endocytic activity, whichis not the case for trypomastigotes that seem not to per-form receptor-mediated endocytosis. A close relation-ship exists among nutritional stress, endocytosis, andcell differentiation in epimastigotes [62]. Assuming thatTcRABL4 is a functional GTPase with a specific roleon the endocytic pathway of T. cruzi, its expressionwould be expected to be lower in trypomastigote forms,where the endocytic activity is lowest, but at the presentmoment we do not have any experimental evidence ofthis participation on endocytic pathway.
Once trypanosomatid small GTPases genes are betterunderstood, they will be used as potential endo and exo-cytic pathways markers, increasing the knowledge aboutT. cruzi biology. Considering the minimal set of RAB
genes that has been equated with the essential yeastRAB proteins [63] and that in T. brucei the RAB familywas shown to contain 16 members [64], it is plausiblethat T. cruzi also has a small number of them, which willmake easier the understanding of the parasite�s endo-cytic pathway. Characterization of RAB-like genes offersan opportunity to study the still poorly understoodmolecular events involved in vesicular traffic in theseorganisms. Ultrastructural and functional studies usingRABs as compartment-specific probes should yieldimportant new insights into the novel aspects of endo-cytic and exocytic pathways in this protozoan parasite.
Acknowledgments
The authors thank Claudio Nunes Pereira and CesarFelix Schmidt for technical assistance. Preliminary T.
cruzi clone CL Brener genomic data were accessed viahttp://TcruziDB.org. Most of the genomic data wereprovided by the TIGR-SBRI-KI Sequencing Consor-tium (TSK-TSC) supported by NIH Grants AI45038,AI45061, and AI45039. The work was supported bygrants from Conselho Nacional de DesenvolvimentoCientıfico e Tecnologico—CNPq, Fundacao Coorde-nacao de Aperfeicoamento de Pessoal de Nıvel Superior(CAPES), Fundacao Jose Bonifacio (FUJB), and Fun-dacao de Apoio a Pesquisa do Estado do Rio de Janeiro(FAPERJ), Brazil.
References
[1] A.H. Kollien, G.A. Schaub, The development of Trypanosoma
cruzi in Triatominae, Parasitol. Today 16 (9) (2000) 381–387.[2] C. Chagas, Nova tripanosomiaze humana, Mem. Inst. Oswaldo
Cruz 1 (1909) 159–218.

816 F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817
[3] I. Porto-Carreiro et al., Trypanosoma cruzi epimastigote endo-cytic pathway: cargo enters the cytostome and passes through anearly endosomal network before storage in reservosomes, Eur. J.Cell Biol. 79 (11) (2000) 858–869.
[4] W. De Souza, T.U. Carvalho, M. Benchimol, Trypanosoma cruzi:ultrastructural, cytochemical and freeze-fracture studies of proteinuptake, Exp. Parasitol. 45 (1978) 101–115.
[5] M. Zerial, H. McBride, Rab proteins as membrane organizers,Nat. Rev. Mol. Cell. Biol. 2 (2001) 107–117.
[6] N. Segev, Ypt and Rab GTPases: insight into functions throughnovel interactions, Curr. Opin. Cell Biol. 13 (2001) 500–511.
[7] J.B. Pereira-Leal, M.C. Seabra, Evolution of the Rab family ofsmall GTP-binding proteins, J. Mol. Biol. 313 (2000) 889–901.
[8] P. Chavrier et al., Hypervariable C-terminal domain of rabproteins acts as targeting signal, Nature (London) 353 (1991) 769–772.
[9] P. Brennwald, P. Novick, Interactions of three domains distin-guishing the Ras-related GTP-binding proteins Ypt1p and Sec4p,Nature (London) 365 (1993) 560–563.
[10] P.J. Casey, M.C. Seabra, Protein prenyltransferases, J. Biol.Chem. 271 (1996) 5289–5292.
[11] M.C. Seabra, E.H. Mules, A.N. Hume, Rab GTPases, intracel-lular traffic and disease, Trends Mol. Med. 8 (1) (2002) 23–30.
[12] H. Field, M.C. Field, Tandem duplication of Rab genes followedby sequence divergence and acquisition of distinct functions in T.
brucei, J. Biol. Chem. 272 (1997) 10498–10508.[13] H. Field et al., Complexity of trypanosomatid endocytosis
pathways revealed by Rab5 and Rab4 isoforms in Trypanosoma
brucei, J. Biol. Chem. 273 (1998) 32102–32110.[14] F. Bringaud et al., Conserved organization of genes in trypano-
somatids, Mol. Biochem. Parasitol. 94 (1998) 249–264.[15] H. Field et al., TbRab2p, a marker for the endoplasmic reticulum
of Trypanosoma brucei, localizes to the ERGIC in mammaliancells, J. Cell Sci. 112 (Pt. 2) (1999) 147–156.
[16] P.W. Denny et al., Leishmania RAB7: characterization of termi-nal endocytic stages in an intracellular parasite, Mol. Biochem.Parasitol. 123 (2002) 105.
[17] S.M. Mendonca et al., Identification of GTPase genes in theprotozoa parasites Trypanosoma cruzi and Leishmania amazonen-
sis, Biol. Res. 26 (1993) 3–9.[18] S.M. Mendonca et al., Characterization of Rab11 homologue in
Trypanosoma cruzi, Gene 243 (2000) 179–185.[19] S.T.G. Leal et al., Trypanosoma cruzi: cloning and characteriza-
tion of a RAB7 gene, Exp. Parasitol. 96 (2000) 1–9.[20] J.R. Araripe et al., TcRAB7 protein is localized at the Golgi
apparatus in epimastigotes, Biochem. Biophys. Res. Commun.321 (2) (2004) 397–402.
[21] E.P. Camargo, Growth and differentiation in Trypanosoma cruzi.Origin of metacyclic trypanosomes in liquid media, Rev. Inst.Med. Trop. Sao Paulo 6 (1964) 93–100.
[22] E. Chiari, Growth and differentiation of Trypanosoma cruzi
culture forms kept in laboratory for different periods of time, Rev.Inst. Med. Trop. Sao Paulo 16 (2) (1974) 81–87.
[23] S.N. al-Abbasy, T.M. Seed, J.P. Kreier, Isolation of the trypo-mastigote form of Trypanosoma cruzi from a mixture of thetrypomastigote and epimastigote forms of the parasite by use of aDEAE–cellulose column, J. Parasitol. 58 (1972) 631–632.
[24] E. Rondinelli, R. Silva, J.F. Carvalho, C.M. de Almeida Soares,E.F. de Carvalho, F.T. de Castro, Trypanosoma cruzi: an in vitrocycle of cell differentiation in axenic culture, Exp. Parasitol. 66(1988) 197–204.
[25] T.P. Urmenyi, Construction of a normalized cDNA library for theTrypanosoma cruzi genome project, J. Euk. Microbiol. 46 (5)(1999) 542–544.
[26] B.M. Porcel, L. Aslund, U. Pettersson, B. Andersson, Trypano-soma cruzi: a putative vacuolar ATP synthase subunit and aCAAX prenyl protease-encoding gene, as examples of gene
identification in genome projects, Exp. Parasitol. 95 (3) (2000)176–186.
[27] J.R. Araripe, F.P. Ramos, et al., Characterization of a RAB5homologue in Trypanosoma cruzi, Biochem. Biophys. Res. Com-mun. 329 (2) (2005) 638–645.
[28] J. Sambrook, E.F. Fritsch, T. Maniatis, Molecular Cloning: aLaboratory Manual, second ed., Cold Spring Habor LaboratoryPress, Cold Spring Habor, NY, 1989.
[29] Cano et al., Molecular karyotype of clone CL Brener chosen forthe Trypanosoma cruzi genome project, Mol. Biochem. Parasitol.71 (1995) 273–278.
[30] J.M. Chirgwin, A.E. Ptzybyla, R.J. MacDonald, W.J. Rutter,Isolation of biologically active ribonucleic acid from sourcesenriched in ribonuclease, Biochemistry 18 (24) (1979) 5294–5299.
[31] J.H. Han, C. Stratowa, W.J. Rutter, Isolation of full-lengthputative rat lysophospholipase cDNA using improved methodsfor mRNA isolation and cDNA cloning, Biochemistry 26 (6)(1987) 1617–1625.
[32] U.K. Laemmli, Cleavage of structural proteins during theassembly of the head of bacteriophage T4, Nature 227 (1970)680–685.
[33] P. Wagner, L. Hengst, D. Gallwitz, Ypt proteins in yeast,Methods Enzymol. 219 (1992) 369–387.
[34] C. Grubmeyer, H.S. Penefsky, The presence of the two hydrolyticsites on beef heart mitochondrial adenosine triphosphatase, J.Biol. Chem. 256 (8) (1981) 3718–3727.
[35] N.L. Cunha-e-Silva et al., Isolation and characterization of areservosome fraction from Trypanosoma cruzi, FEMS Microbiol.Lett. 214 (2002) 7–12.
[36] M.C. Field, J.C. Boothroyd, Trypanosoma brucei: molecularcloning of homologues of small GTP-binding proteins involvedin vesicle trafficking, Exp. Parasitol. 81 (1995) 313–320.
[37] C. Nuoffer, W.E. Balch, GTPases: multifunctional molecularswitches regulating vesicular traffic, Annu. Rev. Biochem. 63(1994) 949–990.
[38] J. Colicelli, Human RAS superfamily proteins and relatedGTPases, SciSTKE. 250 (2004) RE13.
[39] V.M. Olkkonen, H. Stenmark, Role of Rab GTPases in mem-brane traffic, Int. Rev. Cytol. 176 (1997) 1–85.
[40] J.T. Eggensschwiler, Rab23 is an essential negative regulator ofthe mouse Sonic hedgehog signalling pathway, Nature 412 (2001)194–198.
[41] V. Dhir, M.C. Field, TbRAB23: a nuclear-associated Rab proteinfrom Trypanosoma brucei, Mol. Biochem. Parasitol. 136 (2) (2004)297–301.
[42] M.A. Rodriguez et al., An Entamoeba histolytica Rab-likeencoding gene and protein: function and cellular location, Mol.Biochem. Parasitol. 108 (2000) 199–206.
[43] K. Fujikawa, A.K. Satoh, S. Kawamura, K. Ozaki, Molecularand functional characterization of a unique Rab protein, RAB-RP1, containing the WDIAGQE sequence in a GTPase motif,Zool. Sci. 19 (2002) 981–993.
[44] S. Jager, E. Stockert, E. Jager, A.O. Gure, M.J. Scanlan,Serological cloning of a melanocyte rab guanosine 50-triphos-phate-binding protein and a chromosome condensation proteinfrom a melanoma complementary DNA library, Cancer Res. 60(2000) 3584–3591.
[45] L. Norian, I.A. Dragoi, T. O�Halloran, Molecular characteriza-tion of RabE, a developmentally regulated Dictyostelium homo-log of mammalian rab GTPases, DNA Cell Biol. 18 (1999) 59–64.
[46] A. Zahraoui, N. Touchot, P. Chardin, A. Tavitian, The humanRab genes encode a family of GTP-binding proteins related toyeast YPT1 and SEC4 products involved in secretion, J. Biol.Chem. 264 (21) (1989) 12394–12401.
[47] P.S. Anderson, J.C. Lacal, Expression of the Aplysia californica
rho gene in Escherichia coli purification and characterization of itsencoded p21 product, Mol. Cell. Biol. 7 (10) (1987) 3620–3628.

F.P. Ramos et al. / Biochemical and Biophysical Research Communications 333 (2005) 808–817 817
[48] S.E. Neal et al., Kinetic analysis of the hydrolysis of GTP byp21N-ras. The basal GTPase mechanism, J. Biol. Chem. 263(1988) 19718–19722.
[49] S. Hoffenberg et al., Biochemical and functional characterizationof a recombinant GTPase, Rab5, and two of its mutants, J. Biol.Chem. 270 (1995) 5048–5056.
[50] I. John et al., C-terminal truncation of p21H preserves crucialkinetic and structural properties, J. Biol. Chem. 264 (1989) 13086–13092.
[51] J.B. Pereira-Leal, M.C. Seabra, The mammalian Rab familyof small GTPases: definition of family and subfamilysequence motifs suggests a mechanism for functional speci-ficity in the Ras superfamily, J. Mol. Biol. 301 (2001) 1077–1087.
[52] R.A. Erdman et al., Rab24 is an atypical member of the RabGTPase family. Deficient GTPase activity, GDP dissociationinhibitor interaction, and prenylation of Rab24 expressed incultured cells, J. Biol. Chem. 275 (2000) 3848–3856.
[53] T. Chen et al., Rab39, a novel Golgi-associated Rab GTPasefrom human dendritic cells involved in cellular endocytosis,Biochem. Biophys. Res. Commun. 303 (2003) 1114–1120.
[54] A.C.C. Wong et al., Two novel human RAB genes with nearidentical sequence each map to a telomere-associated region: thesubtelomeric region of 22q13.3 and the ancestral telomere band2q13, Genomics. 59 (1999) 326–334.
[55] M.C. Field, B.R.S. Ali, H. Field, GTPases in protozooanparasites: tools for cell biology and chemotherapy, Parasitol.Today 15 (9) (1999) 365–371.
[56] J. Nepomuceno-Silva et al., TcRho1, a farnesylated Rho familyhomologue from Trypanosoma cruzi, J. Biol. Chem. 276 (32)(2001) 29711–29718.
[57] A. De Sa-Freire et al., TcARF1: a Trypanosoma cruzi ADP-ribosylation factor, Parasitol. Res. 91 (2003) 166–170.
[58] T.R. Jeffries, G.W. Morgan, M.C. Field, A developmentallyregulated Rab11 homologue in Trypanosoma brucei is involved inrecycling processes, J. Cell Sci. 114 (2001) 2617–2626.
[59] H. Field et al., Cell-cycle and developmental regulation ofTbRAB31 localisation, a GTP-locked Rab protein from Trypan-
osoma brucei, Mol. Biochem. Parasitol. 106 (2000) 21–35, 2000TbRAB31.
[60] T.R. Jeffries, G.W. Morgan, M.C. Field, TbRAB18, a develop-mentally regulated Golgi GTPase from Trypanosoma brucei, Mol.Biochem. Parasitol. 121 (2002) 63–74.
[61] R.C. Figueiredo, M.J. Soares, The Golgi complexo of Trypano-soma cruzi epimastigote forms, J. Submicrosc. Cytol. Pathol. 27(1995) 209–215.
[62] R.C.B.Q. Figueiredo, D.S. Rosa, M.J. Soares, Differentiation ofTrypanosoma cruzi epimastigotes: metacyclogenesis and adhesionto substrate are triggered by nutritional stress, J. Parasitol. 86 (6)(2000) 1213–1218.
[63] T. Lazar, M. Gotte, D. Gallwitz, Vesicular transport: how manyYpt/Rab-GTPases make a eukaryotic cell? Trends Biochem. Sci.22 (1997) 468–472.
[64] J.P. Ackers, V. Dhir, M.C. Field, A bioinformatics analysis of theRAB genes of Trypanosoma brucei, Mol. Biochem. Parasitol. 141(2005) 89–97.