characterization of a cdna encoding a cysteine-rich cell
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, Sept. 1990, p. 4506-45170270-7306/90/094506-12$02.00/0Copyright C 1990, American Society for Microbiology
Characterization of a cDNA Encoding a Cysteine-Rich CellSurface Protein Located in the Flagellar Pocket of
the Protozoan Trypanosoma bruceiMARY GWO-SHU LEE,'t BERNARD E. BIHAIN,1' DAVID G. RUSSELL,2
RICHARD J. DECKELBAUM,3 AND LEX H. T. VAN DER PLOEG1*Department of Genetics and Development' and Department of Pediatrics,3 Columbia University,
701 West 168th Street, New York, New York 10032, and Department of Pathology,New York University Medical Center, New York, New York 100162
Received 13 March 1990/Accepted 13 June 1990
We have characterized a cDNA encoding a cysteine-rich, acidic integral membrane protein (CRAM) of theparasitic protozoa Trypanosoma brucei and Trypanosoma equiperdum. Unlike other membrane proteins of T.brucei, which are distributed throughout the cell surface, CRAM is concentrated in the flagellar pocket, aninvagination of the cell surface of the trypanosome where endocytosis has been documented. Accordingly,CRAM also locates to vesicles located underneath the pocket, providing evidence of its internalization. CRAMhas a predicted molecular mass of 130 kilodaltons and has a signal peptide, a transmembrane domain, and a41-amino-acid cytoplasmic extension. A characteristic feature of CRAM is a large extracellular domain with aroughly 66-fold acidic, cysteine-rich 12-amino-acid repeat. CRAM is conserved among different protozoanspecies, including Trypanosoma cruzi, and CRAM has structural similarities with eucaryotic cell surfacereceptors. The most striking homology ofCRAM is to the human low-density-lipoprotein receptor. We proposethat CRAM functions as a cell surface receptor of different trypanosome species.
The protozoan parasite Trypanosoma brucei, causativeagent of sleeping sickness, survives in blood by undergoingantigenic variation of its variant cell surface glycoprotein(VSG) coat. The parasite is transmitted by the tsetse flythrough ingestion in a bloodmeal and transfer with the tsetsefly saliva. In the tsetse fly, the bloodstream form trypano-somes differentiate into insect (procyclic) form trypano-somes, resulting in the loss of the protective VSG coat andthe expression of a different and small family of cell surfaceproteins, the procyclic acidic repetitive proteins (or procy-clins; 6, 7, 39, 44, 45, 55, 59). Both VSG and the procyclicacidic repetitive proteins are extracellular proteins anchoredto the lipid bilayer by a covalently attached lipid moiety (12,15, 43). Proteins with transmembrane domains have not beencharacterized in trypanosomes thus far.
Previous studies led to the proposal that a low-density-lipoprotein (LDL) receptor, a transferrin receptor, a nucle-oside transport receptor, and a growth factor receptor existin T. brucei (13, 14, 26; G. Hide, A. Gray, C. M. Harrison,and A. Tait, Mol. Biochem. Parasitol., in press). A densenetwork of microtubules below the plasma membrane pre-sumably prevents the uptake of cell surface receptors andtheir ligands through endocytosis. However, the flagellarpocket, a deep invagination of the plasma membrane wherethe flagellum extends from the cell, is not lined with thissubpellicular microtubule network. The flagellar pocket isbelieved to be the site of uptake of nutrients and macromol-ecules, possibly via receptor-mediated endocytosis (13, 14,32, 40, 57, 58, 59, 62). Here we characterize a conservedtransmembrane protein localized in the flagellar pocket of T.
* Corresponding author.t Present address: Division of Tropical Medicine, School of
Public Health, Columbia University, New York, NY 10032.t Present address: Department of Physiology, Louisiana State
University Medical Center, New Orleans, LA 70112.
brucei. The identification of this protein will enhance ourunderstanding of protein transport and endocytosis in T.brucei.
MATERIALS AND METHODSProtozoan strains and culture conditions. T. brucei 427
bloodstream form variant 118 clone 1 (34) and T. brucei 427procyclic trypanosomes (initially obtained from R. Brun)were used. Procyclic trypanosomes have been maintained inSDM79 medium at 24°C as described by Brun and Schonen-berger (9). Procyclic T. brucei TREU 667 BUT 55 was usedfor the construction of a cDNA library in the bacteriophagevector lambda gtlO (kindly provided by C. Clayton; de-scribed by Mowatt and Clayton [39]). The Trypanosomaequiperdum strain used was Botat 1/78 (37). Trypanosomalewisi was grown in rats for 7 days to peak parasitemia asdescribed previously (20). Leptomonas seymourii (ATCC30220) was cultured by the method of Bone and Steinert (5).Crithidia fasciculata (wild type, originally derived from G.Hill) was cultured by the method of Le Trant et al. (35).Leishmania major (WR300; 21) was cultured in Schneider'smedium (GIBCO) with the addition of 10% fetal calf serum.
T. brucei variant 118 clone 1 was grown in white maleWistar rats and purified from blood elements as described byFairlamb et al. (18) and then used for DNA and RNAisolation. Procyclic trypanosomes were grown to late-logphase (cell density 0.5 x 107 to 1 x 107 per ml), after whichthe cells were spun down and washed with PSG buffer (50mM sodium phosphate [pH 7.3], 137 mM NaCl, 1.0%glucose), and then DNA and RNA were isolated.DNA isolation and Southern analysis. CRAM (cysteine-
rich, acidic integral membrane protein) RNA was identifiedon the basis of its artificial hybridization with a syntheticchloramphenicol acetyltransferase oligonucleotide in primerextensions with trypanosome poly(A) mRNA. Since theprimer extension pattern indicated that the oligonucleotide
4506
Vol. 10, No. 9

FLAGELLAR POCKET PROTEIN OF T. BRUCEI 4507
detected an mRNA with an internal repeat, we purifiedcDNAs for further analysis of the gene. CRAM cDNAclones in phage lambda gtlO were purified to homogeneity,and the inserts were subcloned in the plasmid pUC18.Nuclear DNA was isolated from trypanosomes as describedby Van der Ploeg et al. (56) and subsequently digested withrestriction endonucleases. The DNA was separated on a0.8% agarose gel and transferred onto nitrocellulose filters(49). After hybridization with a nick-translated 32P-labeledprobe, the nitrocellulose filters were washed at final posthy-bridizational stringencies as indicated in the figures.RNA isolation and Northern (RNA) analyses. All RNA
samples were isolated by guanidine thiocyanate lysis (11)and purified by centrifugation through CsCl cushions. North-ern analysis of RNA was carried out by separation of RNAsamples in a 1% formaldehyde agarose gel as described byBoetdker (4). Northern blots were hybridized with 32P-labeled probes. Following hybridization, filters were washedto a final stringency of 0.1x SSC (lx SSC is 0.15 M NaClplus 0.015 M sodium citrate) and 0.1% sodium dodecylsulfate (SDS) at 65°C.DNA sequence analysis. Sequence analysis was performed
by the dideoxy-chain termination method (47). SuitableDNA fragments generated by cleavage with restriction en-zymes and Bal 31 exonuclease were subsequently subclonedinto bacteriophages M13, mpl8, and mpl9 and then used astemplates for sequencing. Both strands were sequenced,except in the repeat region, where it was difficult to deter-mine the overlap of clones due to the presence of overallidentical repeats. We generated Bal 31 deletion libraries ofthe CRAM.1 cDNA in the phage M13, covering the 2,360-base-pair (bp)-repeat region. Sequence analysis of 60 dif-ferent Bal 31 deletion clones identified a consensus sequencefor the 36-bp repeats. It was possible to determine therelative order of only 7 of the repeats flanking the NH2-terminal single-copy sequence and 15 of the repeats flankingthe COOH-terminal single-copy sequence. The order of theother, internally located, repeats could not be determined,since the similarity of the repeats made the alignment of theBal 31 clones within the repeat array impossible. The ap-proximate number of repeats was deduced from the size ofthe repeat region divided by 36.
Generation of fusion proteins and antiserum. VariouscDNA fragments encoding CRAM were subcloned into theexpression vector pATH, which contains the Escherichiacoli thranilate synthetase (trpE) gene (48). The fusion geneconstructs were made by using either restriction enzyme-generated fragments or Bal 31 deletion clones. The DNAsequences of the resulting recombinant constructs confirmedthat the fusion proteins were translated in the correct readingframes. The resulting proteins contain the TrpE proteinfused to different regions of CRAM. Fusion protein wasproduced in E. coli HB101. The lysates (generated by boilingcells in SDS sample buffer [31]) were analyzed in a 7.5%SDS-polyacrylamide gel. Following staining with Coomassiebrilliant blue and destaining, bands corresponding to thefusion proteins were excised, rinsed in phosphate-bufferedsaline (PBS), and used for immunization of Wistar rats orNew Zealand White rabbits. Animals were injected subcu-taneously with the minced acrylamide gel slices containingapproximately 0.5 mg of fusion protein macerated withadjuvant (either Freund complete adjuvant or 10% aluminumsulfate). Booster injections were given at 2-week intervals.Six weeks after the first immunization, animals were bled,yielding sera with titers ranging from 1/250 to 1/1,000.FACS analysis and light microscopic analysis of trypano-
somes. Flow cytometry was performed on live insect formtrypanosomes. Trypanosomes (2 x 107) were washed withPBS supplemented with 10% fetal calf serum. Cells wereincubated with the different antisera for 1 h at 0°C in PBSwith 10% fetal calf serum. Following this incubation, cellswere washed three times with PBS-10% fetal calf serum at0°C and then incubated with fluorescein isothiocyanate(FITC)-conjugated goat anti-rat immunoglobulin G (IgG) at0°C for 1 h (Cappel Laboratories, Inc.). Cells were againwashed three times and suspended in PBS for fluorescence-activated cell sorter (FACS) analysis (FACS IV; BectonDickinson and Co.).
Procyclic trypanosomes were harvested by centrifugationfor 5 min at 400 x g and washed once with PBS. Then, cellswere suspended in 3.7% formaldehyde in PBS, pH 7.4, for 10min. Following fixation, cells were spun down, resuspendedin fresh PBS, sedimented onto slides by using a cytospincentrifuge (8 min at 1,000 rpm; Shandon Instruments, Inc.)and then fixed in cold methanol for 5 min. After rehydrationin PBS for 5 min, slides were incubated with antibodies.Following the reaction with the first antibody, cells wereincubated with the FITC-conjugated goat anti-rabbit IgG(Cappel). After the last wash, 4,6-diamidino-2-phenylindolewas applied in water at a concentration of 1 ,ug/ml for 5 minat room temperature, after which the slides were brieflyrinsed with water. The cells were mounted under a cover slipwith Elvanol. Cells were viewed and photographed under aZeiss immunofluorescence microscope by using a 100xobjective (Ph3-Neofluar).Immunoelectron microscopy. Procyclic and bloodstream
forms of T. brucei were fixed in 4% formaldehyde and 0.5%gluteraldehyde in 200 mM PIPES [piperazine-N,N'-bis(2-ethanesulfonic acid)}-2 mM MgCl2, pH 7.0, for 60 min atroom temperature. The cells were then dehydrated throughan alcohol series at decreasing temperature to -30°C andembedded in Lowicryl according to the instructions of themanufacturer. Thin sections were cut, blocked in 50 mMglycine in PBS, and incubated in affinity-purified rabbitanti-Pl-55 IgG in PBS with 0.1% Tween 20. After beingwashed, the bound antibody was visualized with 15-nm-diameter gold particles coupled with goat anti-rabbit IgG(Amersham Corp.). The grids were washed, fixed in 2%gluteraldehyde in PBS, and contrasted in Reynold's lead anduranyl acetate.
Immunoblotting analysis. Total cell lysates of 2 x 107trypanosomes were size separated in a 7.5% polyacrylamidegel and electrophoretically transferred onto nitrocellulosefilters. The different panels were next treated with thedifferent antibodies (31, 53). The nitrocellulose filters werefirst blocked with 3% nonfat milk in PBS. Antibody reactionswere done in PBS with 0.3% Tween 20. Following the firstantibody reaction, filters were treated with horseradishperoxidase-labeled goat anti-rabbit IgG (Sigma ChemicalCo.) and the location of the peroxidase was detected byusing diaminobenzidine (0.5 mg/ml) in the presence of hy-drogen peroxide (0.003%).
RESULTS
Expression of CRAM in insect and bloodstream form try-panosomes. Several different techniques, focused on theidentification of conserved cell surface proteins of parasiticprotozoa, have been used in our laboratory to identify genesencoding stable (nonvariant) cell surface proteins of T.brucei (M. G.-S. Lee and L. H. T. Van der Ploeg, unpub-lished data). One cDNA clone was obtained through a
VOL. 10, 1990

4508 LEE ET AL. MOL. CELL. BIOL.
f 36 bp Hph repeat Hp_urIs--I 1 I t n r
TAA Poly-ACRAM.
- .2.3
500 bp
MINI-SXON ACACTTTCCCTACTATATTG TTCC 29COIAGACAAGTTAACCCTATTCTTAAACACGGCAATCTCCCCCAAATACTAC 80
ATTCCTTTCTTTATCACCCTCTTATTCCGCCAGCACCGCCTGTGTCCTCCC 131ATC GCC AAC GAG GCA CGT CCA ATC TTT CAA CAA TCA AAT 170Mat Gly Ann Clu Ala Cly Pro Ile Ph Clu Clu Sor han
GCT GAG CTA CCC ACA CCC CCT CCT CAT GCC CTA CAT CAT 209
Ala Glu Val Ala Thr Pro Pro Ala Asp Ala Va1 EtH Asp
CAT TTC TTC TTC GAC TAC AAC AAC CCT ACG CCT TAC GCC 248Asp Ph. Ph. Ph. Asp Tyr Lys Asn Ala Thr Cly Tyr Ala
I 3
FIG. 1. Differential expression of the CRAM gene. Total RNA(20 ,ug) from bloodstream form (variant 118 clone 1, lane B) andprocyclic (insect form, stock 427 derived; lane P) trypanosomeswere size separated in a 1% formaldehyde agarose gel and trans-ferred to nitrocellulose filters. This filter was probed with 32P-labeledCRAM.1 cDNA and subsequently stripped and reprobed with anoligonucleotide complementary to the 28S rRNA sequence. Follow-ing hybridization, the filters were washed to a final stringency of0.1x SSC (pH 6.15) and 0.1% SDS at 65°C (left-hand panel) or 3.0xSSC and 0.1% SDS at 40°C (right-hand panel). Molecular sizes ofmarker (in kilobase pairs) are indicated at the left.
different route and was isolated because of its artificialcross-hybridization with a synthetic oligonucleotide, identi-fying an mRNA that encoded an internal repeat in its readingframe. We choose to analyze this mRNA, since severalprotozoan genes encoding repeated sequences in their read-ing frames encode cell surface proteins (28). We isolatedthree overlapping cDNA clones (CRAM.1 to CRAM.3)which encoded the repeat in the mRNA (see Materials andMethods for details). These CRAM cDNAs detected a3,400-nucleotide (nt) mRNA in insect form (Fig. 1, lane P)and bloodstream form (Fig. 1, lane B) trypanosomes. Thehybridization signal was approximately fivefold weaker withRNA derived from bloodstream form trypanosomes. Controlhybridizations of this Northern panel, to identify the try-panosome 28S rRNA, showed that equal amounts of totalRNA were loaded in each lane (Fig. 1, right panel). The levelof expression of CRAM in bloodstream form trypanosomesis variable, ranges from 2- to 10-fold less than in insect formtrypanosomes, and is dependent on the batch of bloodstreamform trypanosomes (Fig. 1 and data not shown).
Predicted amino acid sequence of CRAM. The three over-lapping cDNA clones (CRAM.1 to CRAM.3) were analyzedand were shown to comprise almost the entire 3,400-nt
CAT CAT TCC AAC ATC ACA CWC CAC TCC AAT CGA ACT 284Asp Asp Cys Asn Ile Thr Cly Aap Cys Agn Clu Thr
AspHp
GAT CAT TCT CAC ATC ACA I TCC AAT CAA ACTCAT CAT TGC AAC ATC ACA CCT CAC TCC AAT GAA ACCCAT GAT TCC AAC ATC ACA CCS CAC TCC hAT CAA ACTCAT CAT TCC AAC ATC ACA CCT CAC TCC AAT CAA ACTCAT C-AT TOC AAC ATC ACA GCT CAC TCC AAT CAA ACTCAT CAT TCC GAG ATC ACA CCS CAC TGC AAT CAA ACTCAT CAT TCC AAC ATC ACA CGT CAC TCC PAT CAA ACC CONSKNSUS.. ... .. T C ...T.... ... ... .. ... ... T SEQUENCE
C-AT CAT TGC AAC ATC ACA GCT CAC T0C AAT CAA ACTGAT CAT TCT AG ATC ACA CCT CAC TCC AT GAA AGTCAT CAT TCC CAC ATC ACA CCT CAC TOC AAT CAA ACCCAT CAT TOT AAC ATC AGA GCT CAC TCC AAT GAA ACTGAT CAT TGC CAC ATC AC-A CGT CAC TCC AAT CAA ACCCAT CAT TGC AAC ATC ACA GOT CAC TCC AAT GAA ACCCAT CAT T0C AAC ATC ACA CCT CA:C TCC AAT GAA ACCCAT CAT TC:C hAG ATC ACA OCT CAC TCC AAT CAA ACCGAT CAT TOT AAC ATC ACA CGT CAC TCC AAT CAA ACTGAT CAT TOT AAC ATC ACA OCT CAC TCC JAT CAA ACTGAT CAT TOT AAC ATC ACA CCCT &AC TCC AAT GAA ACC
CAT CALT TCC CAC ATC ACA OCT CAC TGC AAT CAA ACTOAT CT TGT AAC ATC ACA CGT CAC TGC AAT CAA ACT
CAT CAT TGT AAC ATC ACA OCT GAC TGC AAT CAA ACC
CAT CAT TCC AAC ATC ACA GCT CAC TCC AAT CAA ACT 2624
GAG GTA AGC CAT GCA CCC GAC CCC ACT CAC CCG ATC TTCGlU Val Sr Asp Ala Al. Asp Cly Thr Asp Cly Mot Ph.
HTTC AAA OCT TCC TCT TCC TTA hAG COT OTT CCA CTC TOTLou Lye S-r Ser S-r S9r Lou Lys Lou Val Ala Lou Cys
CAT CCAI TCCCT At :Ar CGAT TM CCC AG TCG AATAsp Cly Cys Pro Thr Clu Asp S9r Pro Lys S&r Ser Asn
tCA AAC CCC AAA CCI TCC AGC COC AGT CCC CCA TTG CTG TRANS-Ala Lys Gly Lys Gly Ser S9r Vsl Ser Ala Cly Leu Lou MEIMEANE
COT TTG CCT OCT 0CC ACG TTT TTC GTA CTC CC CTC0CCLou Lou Ala Cly Ser Thr Ph. Lou Va1 Leu Ala Val Cly
CT TCCG CA CTT TTA TTT TTC CCG ACX CAA CCC CAC AACLou Ser Ala Va1 Lou Phe Leu Cly Arg Clu Arg Cln Asn
CCC CGT CTC ATA I| CAC AAC GAC CTT ATC ATC GAG GAGAla Val Va1 I1 jCy. Thr Agn :lu Va1 Met Net Clu Clu
CTG CCC OCA 0SC]TTA TCT CAT CCA TCT TTT CCC CTC CCC 2936Val Pro Arg [Caj Lou Ser Amp Ala Ser Phe A1a Va1 Pro
CTA ACT CAA TCC TCA CAT CIA CCG ACA CCC TAAACCTATAC 2977Val Thr Gln Ser Sar Asp Clu Ala Ar; Pro ---
GTACTCATTGAGATTTAACAGTTCTGCATCTACAGAACAGGACGTGAACAC
AGCGCTTCTCCCCTTC:TACTCGAGTTGGCACTTATTGCATTTCTGTTTAG;ATGTCTTCGTAG;TATTAGCACTACATTTATGTAC;AGTGC CACGTATGAAT
TGATCTTCGT. AGCAATTTCAACTGTTCCGCAATAA.AAAAAAAAAAAA............AAAAIAAAAAAAAAAIAAJIL
AAAA CRAM .1
AAAAAA CRAM. 3
(:RRA N
B F)
28 SrR \AB P
4.4 -
mino- 1Gexon
2 4
1.4 ..
r

FLAGELLAR POCKET PROTEIN OF T. BRUCEI 4509
CRAM mRNA. The CRAM.1 cDNA contained a short 22-ntpoly(A) tail, while CRAM.2 encoded 20 nt of the common5'-capped 39-nt miniexon (Fig. 2). The CRAM.1 cDNA isaltnost full length and lacks only 135 nt from the 5' end of themRNA. The cDNA clones contained a large, roughly 2,400-bp, internal region lacking sites for most restriction en-zymes. Digestion of this region with the restriction enzymeHphI released a unique 36-bp repeat fragment, as deter-mined by polyacrylamide gel electrophoresis (see HphIphysical map in Fig. 2). Thus, we expected that this regionencoded a repetitive amino acid sequence.The DNA nucleotide sequence of the CRAM cDNAs
revealed a large open reading frame encoding a predictedpolypeptide of 945 amino acids with a conserved 12-amino-acid repeat (Fig. 2; see Materials and Methods for details).We could not be certain that we had sequenced all the 36-bprepeats, due to their similarity (see Materials and Methodsfor details), and we considered the possibility that a singlerepeat could contain a stop codon resulting in a shorterprotein. We addressed this possibility by expressing theCRAM protein in E. coli by using pATH expression vectors(Fig. 3; 48). The predicted fusion proteins, with the followingmolecular masses, could be identified: 150 kilodaltons (kDa),containing the N terminus, the repeat array, and 22 aminoacids following the repeats (lane B); 80 kDa, containing partof the repeat array (lane C); 50 kDa, containing the nonre-petitive COOH-terminal extension (lane D); and 165 kDa(entire gene) (lane A). The 165- and 150-kDa proteins werepresent only in minute quantities, due to their large size,which affects protein stability (data not shown); hence theyare only faintly visible in Fig. 3. The data show that thepredicted open reading frame of CRAM is correct. In thisreading frame, two of the base substitutions in the repeatsare third-base mutations which do not affect the amino acidsequence, while one additional base substitution changes anaspartic acid residue into asparagine.We assume that the third ATG of the cDNA is the
initiation codon, located 150 nt downstream of the predicted5' end of the miniexon, since it has the eucaryotic consensussequences of initiation codons (29). Two additional in-frameATGs exist upstream, which could give rise to a protein witha slightly longer NH2-terminal extension. The 945-amino-acid polypeptide has a putative signal sequence cleavage site(indicated with an arrow in the NH2 terminus of the protein
FIG. 2. Physical map and nucleotide sequence of the CRAMcDNAs. The physical map of CRAM is shown at the top. Abbrevi-ations: H, HindlIl; Hp, HphI. The presumed ATG translationinitiation codon and the TAA transcription termination codon areindicated. Black boxes indicate the vector sequences. The DNAnucleotide sequence and the predicted amino acid sequence ofCRAM are shown below. The nucleotide sequence of the cDNAclones was determined by the dideoxy-chain termination method(47). From 5' to 3', we find 20 nt of the miniexon, a 150-nt5'-untranslated extension, a 2,835-nt open reading frame, a 471-nt3'-untranslated extension, and a 22-nt poly(A) tail. The arrowdenotes a signal sequence cleavage site. The proposed N-linkedglycosylation sites (Asn X Thr) are marked by short thin lines. Theproposed 0-glycosylation sites in the region preceding the trans-membrane domain are indicated by broken lines. The 34 hydropho-bic amino acid residues of the potential transmembrane region are
marked with a solid line. Cysteines are boxed. The DNA sequenceof only 23 of the 66 repeats is presented, since it was not possible toalign the internally located repeats. The boxed region, labeledconsensus sequence, represents the unaligned 43 repeats. Twodifferent cDNAs contained slightly different poly(A) tails (CRAM.1and CRAM.3).
35 bp Hph reDeetA
HH'
1 65 kDe
hp 36 Up Hph repeet
TAA Poly-AHp H
B
36 bp Hph repeatC
D L.1 KBtA-ZZ,<Z:TAA IT*AA Poly-A
E /Z
E C B D C
1 50 kDo
80 kDe
50 kDe
35 kDo
B A E
_9 _-I -200
- 116
,V.00
_. -4_
.,.,ME.P
- ,-
_dmo
- 42
FIG. 3. Structure and expression of different CRAM fusionproteins in pATH vectors (a schematic representation of the dif-ferent plasmid constructs, labeled A through E, is shown at the top).The fusion gene constructs were made by using either restrictionenzyme-generated fragments or Bal 31 deletion clones ligated intopATH vectors (see Materials and Methods for details). The readingframes of the pATH constructs were checked by DNA sequenceanalysis. Proteins were expressed in E. coli and were size separatedby 7.5% reducing SDS-polyacrylamide gel electrophoresis. TheTrpE fusion protein products of 50, 80, 150, and 165 kDa are markedwith arrows. The far-right lane is a size standard. The hatched boxrepresents the pATH TrpE protein.
sequence in Fig. 2) (3, 60); encodes two cysteine residues per12-amino-acid repeat, with 5 amino acid residues separatingthe cysteines; and has a 114-amino-acid COOH-terminalnonrepetitive extension. Potential glycosylation sites are
present in the repeat as well as in the nonrepetitive extra-cellular extension (see next sections).The hydrophobicity plot of the predicted protein (Fig. 4)
indicates that it could be an integral membrane protein witha large hydrophilic extracellular domain and a transmem-brane domain (residues 870 through 904) devoid of chargedamino acids, followed by the charged amino acids Arg-Glu-Arg (42) and a 41-amino-acid cytoplasmic intracellular ex-
tension (Fig. 2). The potential transmembrane domain is 34amino acids, but the actual transmembrane domain is likelyto be shorter and could start at the amino acid sequence LLLLAGS.... The presence of the potential transmembranedomain and hydrophilic cytoplasmic extension strongly sug-gests that CRAM is an integral membrane protein.
In common with other trypanosome mRNAs, the large(471 nt) 3'-untranslated extension of the CRAM mRNA lacksthe AAUAAA consensus sequence for poly(A) addition.
Location of CRAM at the flagellar pocket. To investigate
kCDa
rz,ez7z772 ,& 70-M-M.
VOL. 10, 1990
--a
W44#0
-4QM6. lownew quomw I
WW,--W
4mmm VMW4.41mmommp 40.= 4.0
NW I

4510 LEE ET AL.
AMINO ACIDS: 100
-21% ^
_______- i.-1r~.-1J,*-"-,24~~~~~kI.V.'k,a=,,.i,
FIG. 4. Hydrophobicity plot of the CRAM amino acid sequence. A hydrophobicity plot was obtained from the predicted sequence ofCRAM by the method of Kyte and Doolittle (30). Only 6.5 of the 12-amino-acid repeats are shown. The numbers refer to amino acid positions.
whether CRAM mRNA encodes an integral membrane pro-tein, we raised antibodies against CRAM fusion proteins inrats and rabbits (Fig. 3 and data not shown). These antibod-ies were used to detect CRAM in Western immunoblots oftrypanosome lysates and on the cell surface of insect andbloodstream form trypanosomes by using indirect immuno-fluorescence labeling.
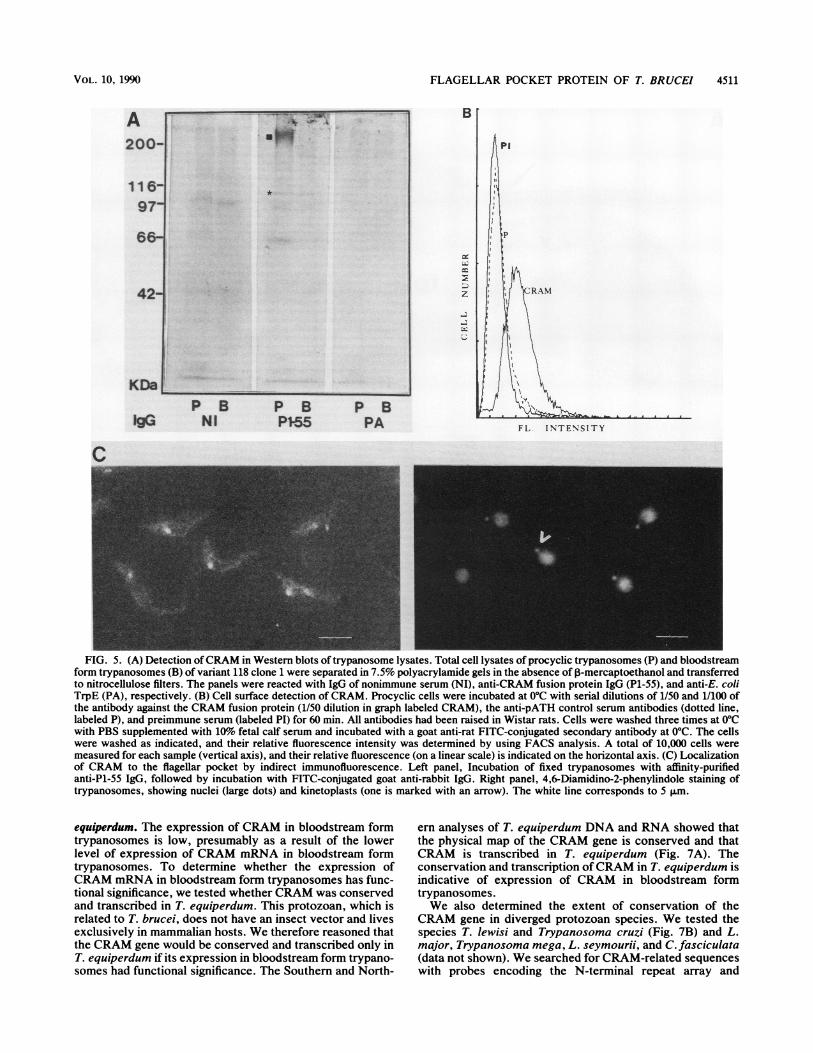
Antibodies were raised in a rabbit against a short fusionprotein (P1-55), containing 4.5 cysteine-rich repeats and 22amino acids of the nonrepetitive C-terminal extension (thenucleotide sequences of the constructs in the pATH expres-sion vectors were analyzed to confirm the reading frames,and we determined the amino acid composition of the pATH[P1-55] fusion protein; data not shown). Antibody waspurified by affinity absorption with the pATH and P1-55recombinant proteins. These affinity-purified antibodies de-tected only the CRAM recombinant proteins in Westernblots of E. coli lysates (data not shown). In Western blots ofinsect form trypanosome lysates, these antibodies specifi-cally detected a protein which migrated slightly slower thanthe 200-kDa molecular mass marker (Fig. 5A, marked with ablack square), while IgG of preimmune serum or anti-pATHantiserum gave no significant reactions. This CRAM proteinwas barely detectable in the lysates of bloodstream formtrypanosomes, and a shorter protein, which may be aproteolytic cleavage product of 100 kDa (marked with anasterisk in Fig. SA) is detectable in both insect and blood-stream form trypanosomes (Fig. SA and data not shown).The protein separation in Fig. 5A was performed in theabsence of the reducing agent P-mercaptoethanol. A slightreduction in the molecular weight of CRAM and the detec-tion of several proteolytic cleavage products occur in thepresence of reducing agents (data not shown). We concludethat this is due to the disruption of the disulfide bonds. Sincethe predicted molecular mass of the CRAM amino acidbackbone is only 130 kDa, the finding of a protein thatappeared larger is indicative of disulfide linkage and glyco-sylation (see next sections).
Figure SB summarizes the results of the FACS analysiswith antibodies made against fusion protein B (Fig. 3) withlive insect form trypanosomes. These antibodies indeeddetected epitopes on the cell surface of insect form trypano-somes. We compared the fluorescence intensities of the cellsby using (i) preimmune serum (labeled PI), (ii) immuneserum raised against the CRAM fusion protein (labeledCRAM), and (iii) a control antiserum raised against thepATH vector-derived protein only (dotted line labeled P).Sera at dilutions of 1/50 and 1/100 were tested, and bothdilutions gave identical results; a significant, although small,increase in FITC cell surface labeling was detected only withserum raised against the fusion protein (labeled CRAM; 1/SOdilution shown in Fig. SB). The relative amount of FITC cellsurface labeling is poor, indicating either a low concentrationof the protein, its rapid internalization, or an inefficientreaction between the antibodies and the CRAM protein in
live trypanosomes. The FITC cell surface labeling efficiencyvaried slightly in each experiment, giving a FITC stainingsignal roughly 10-fold over background, depending on thebatch of cells used.The CRAM protein was shown to be concentrated in the
flagellar pocket of formaldehyde-fixed trypanosomes whenindirect immunofluorescence was used (using the immuno-absorbed P1-55 CRAM antibodies; Fig. SC). Preimmune oranti-PATH antiserum never gave specific signals (Fig. 5Aand data not shown). The approximate position of theflagellar pocket was demonstrated indirectly by using theDNA-specific dye 4,6-diamidino-2-phenylindole, whichstains the kinetoplast DNA, located close to the flagellarpocket (indicated with arrow), and the trypanosome nucleus(Fig. SC, right panel). Superimposition of the fluorescenceand 4,6-diamidino-2-phenylindole images shows that CRAMis concentrated at or close to the kinetoplast DNA. Thisresult indicates that CRAM is a cell surface protein of theflagellar pocket.By using immunoelectron microscopy and P1-55 antise-
rum (raised against a short recombinant protein containing4.5 cysteine-rich repeats and 22 amino acids of the adjacentnonrepetitive extension), we confirmed the flagellar-pocketlocation of the CRAM protein (Fig. 6; 15-nm gold particlesidentify P1-55 antibodies). The analysis of a series of sec-tions (Fig. 6A through D) through different points along theflagellar pocket of several cells shows that the flagellarpocket is indeed specifically coated with the CRAM protein:the highest concentration of anti-P1-55 labeling was foundwithin the flagellar pocket of the procyclic-form trypano-somes, located close to the base of the flagellum (Fig. 6A).The gold particles were associated with either the membraneof the flagellum itself or with the pocket membrane (Fig. 6B,C, and D; panel D shows a dividing trypanosome with twoflagella). Significant amounts of label could not be seenlocated free in the lumen of the pocket. In most instances thelabel was in close proximity with the cellular membrane andlocated to the lumen side of the pocket, indicating that theprotein is membrane associated. The preferential location ofthe label to the lumen side of the pocket shows that theCRAM protein was most often oriented with its N terminusand repeat arrays exposed to the lumen of the flagellarpocket or the lumen of cytoplasmic vesicles where labelcould be found as well. These cytoplasmic vesicles wereabundant in specific regions close to the flagellar pocket, andCRAM-containing vesicles could not be found in otherregions of the cell. We assume that these vesicles result fromendocytosis of the CRAM protein. Bloodstream form try-panosomes of T. brucei stock 427-60, processed by the sameprocedures, showed little, if any, reactivity with the P1-55antiserum, supporting the data indicating that expression ofCRAM in long slender bloodstream form trypanosomes islow (Fig. 6E). From these data we infer that CRAM is a cellsurface protein located in the flagellar pocket.
Evolutionary conservation and expression of CRAM in T.
MOL. CELL. BIOL.
FLAGELLAR POCKET PROTEIN OF T. BRUCEI 4511
A200-
116-97-
42-
KDa.P B P BNI P155
P BPA
B
z
F L INTENSITY
FIG. 5. (A) Detection ofCRAM in Western blots of trypanosome lysates. Total cell lysates of procyclic trypanosomes (P) and bloodstreamform trypanosomes (B) of variant 118 clone 1 were separated in 7.5% polyacrylamide gels in the absence of ,B-mercaptoethanol and transferredto nitrocellulose filters. The panels were reacted with IgG of nonimmune serum (NI), anti-CRAM fusion protein IgG (P1-55), and anti-E. coliTrpE (PA), respectively. (B) Cell surface detection of CRAM. Procyclic cells were incubated at 0°C with serial dilutions of 1/50 and 1/100 ofthe antibody against the CRAM fusion protein (1/50 dilution in graph labeled CRAM), the anti-pATH control serum antibodies (dotted line,labeled P), and preimmune serum (labeled PI) for 60 min. All antibodies had been raised in Wistar rats. Cells were washed three times at 0°Cwith PBS supplemented with 10%o fetal calf serum and incubated with a goat anti-rat FITC-conjugated secondary antibody at 0°C. The cellswere washed as indicated, and their relative fluorescence intensity was determined by using FACS analysis. A total of 10,000 cells weremeasured for each sample (vertical axis), and their relative fluorescence (on a linear scale) is indicated on the horizontal axis. (C) Localizationof CRAM to the flagellar pocket by indirect immunofluorescence. Left panel, Incubation of fixed trypanosomes with affinity-purifiedanti-P1-55 IgG, followed by incubation with FITC-conjugated goat anti-rabbit IgG. Right panel, 4,6-Diamidino-2-phenylindole staining oftrypanosomes, showing nuclei (large dots) and kinetoplasts (one is marked with an arrow). The white line corresponds to 5 1lm.
equiperdum. The expression of CRAM in bloodstream formtrypanosomes is low, presumably as a result of the lowerlevel of expression of CRAM mRNA in bloodstream formtrypanosomes. To determine whether the expression ofCRAM mRNA in bloodstream form trypanosomes has func-tional significance, we tested whether CRAM was conservedand transcribed in T. equiperdum. This protozoan, which isrelated to T. brucei, does not have an insect vector and livesexclusively in mammalian hosts. We therefore reasoned thatthe CRAM gene would be conserved and transcribed only inT. equiperdum if its expression in bloodstream form trypano-somes had functional significance. The Southern and North-
em analyses of T. equiperdum DNA and RNA showed thatthe physical map of the CRAM gene is conserved and thatCRAM is transcribed in T. equiperdum (Fig. 7A). Theconservation and transcription ofCRAM in T. equiperdum isindicative of expression of CRAM in bloodstream formtrypanosomes.We also determined the extent of conservation of the
CRAM gene in diverged protozoan species. We tested thespecies T. lewisi and Trypanosoma cruzi (Fig. 7B) and L.major, Trypanosoma mega, L. seymourii, and C. fasciculata(data not shown). We searched for CRAM-related sequenceswith probes encoding the N-terminal repeat array and
-
a*
C
VOL. 10, 1990

4512 LEE ET AL.
A
.'
WB, . C
fp
ex tit^
Aw,,-
4trs
*s. ~ ~ ~
p 5rtv -
X., *
,.i.",
.k'sS s. a.:
*Ss
's ..
A.~~~~~~~~~~~~~~~S
fp4^
AS:~~~1.
A*-
MOL. CELL. BIOL.
,.jl .. IrjFN-.7 7 .,.r
a
B *. ..
Pi bt..t."
K;S

FLAGELLAR POCKET PROTEIN OF T. BRUCEI 4513
FIG. 6. Orientation of CRAM in the membrane and presence ofCRAM at the flagellar pocket. Immunoelectron microscopy revealed thelocation of CRAM at the flagellar pocket (fp), using gold-labeled anti-IgG for determination of the location of anti-CRAM antibodies.Antibodies, directed against the N-terminal CRAM repeat region, mainly localized to the lumen side of the flagellar pocket, the lumen sideof vesicles located underneath the pocket (v), and the base of the flagellum. (A through D), Sections through insect form trypanosomes; (E),a bloodstream form trypanosome. Symbols: fp, flagellar pocket; k, kinetoplast DNA; n, nucleus; v, vesicle; arrows point to the flagella.
COOH-terminal sequences separately. If identical restric-tion fragments can be detected in a particular restrictionenzyme digest with each probe separately, one significantlyreduces the possibility that the hybridization signals are dueto fortuitous cross-hybridization. The two probes, fromeither the NH2 terminus or the COOH terminus of theCRAM gene, detected multiple identical-sized bands in somerestriction enzyme digests of T. cruzi and T. lewisi DNAonly. This suggests that the hybridization signals defineCRAM gene homologs in T. cruzi and T. lewisi (Fig. 7B,compare the bands marked with an arrow in the left- andright-hand lanes). In the other protozoal species belonging todifferent genera, the hybridization signals were weaker anddiscrete cross-hybridizing restriction enzyme fragmentscould be detected only with probes from the C-terminalextension of the protein. The data thus indicate that CRAMis a conserved cysteine-rich protein of the genus Trypano-soma. The sequence conservation is strongest in the subge-nus Trypanozoon (T. brucei and T. equiperdum).The almost absolute sequence conservation in the CRAM
repeat in T. brucei may derive from gene conversion eventsin repeat arrays leading to the homogenization of repeatsequences (33; M. G.-S. Lee, R. I. Polvere, and L. H. T.Van der Ploeg, Mol. Biochem. Parasitol., in press).
DISCUSSION
We identified a cysteine-rich transmembrane protein(CRAM) located at the flagellar pocket of the protozoan T.brucei. The flagellar pocket is the only area of the cellsurface of the trypanosome proposed to function in (recep-tor-mediated) endocytosis (13, 14, 19, 32, 57, 58, 62). SinceCRAM also localized to the lumen of vesicles locatedunderneath the flageilar pocket, the protein is presumablyinternalized. The structure and hydrophilic cytoplasmic ex-tension of CRAM indicate that unlike other cell surfaceproteins of T. brucei examined so far, CRAM is not an-chored to the membrane through a lipid moiety but containsa transmembrane domain. It is of interest that the firsttransmembrane protein identified in these trypanosomes
A T.b T.eHP B PB
kb
2 3-9-4-
4-3-
2-3-2-0-
T.b T.ekb
7.5-
4.4-
2.4-
14-
B Tcruzi TIewisiC N C N Probe
kb
q-sc23- -_
9.4_ --_6.6-
4.3-5
2.3- _2.0-
0-
0.3-
FIG. 7. Southern and Northern blot analyses of the CRAM genes in T. brucei, T. equiperdum, T. lewisi, and T. cruzi. (A) Southern blotanalysis of T. brucei and T. equiperdum CRAM genes. Restriction enzyme-digested genomic DNA (2 ,ug) from T. brucei (labelled T.b) andT. equiperdum (labeled T.e) was size separated, transferred onto nitrocellulose, and hybridized with the entire 32P-labeled insert of theCRAM.1 cDNA clone. Identical restriction fragments are detected with PstI (P)- or BamHI (B)-digested DNA (H, HindIII). Posthybridiza-tional washing stringencies were at 65°C and 0.1x SSC. The Northern blot on the right-hand side, containing 20 ,ug of T. brucei insect formRNA (lane labeled T.b) and 20 ,ug of T. equiperdum bloodstream form RNA (lane labeled T.e) was hybridized with the entire CRAM.1 cDNA.(B) A 2-,ug portion of T. lewisi and T. cruzi nuclear DNA was digested with the restriction enzyme HindIII and hybridized with 32P-labeledprobes which covered the N-terminal extension and the repeat array (lanes labeled N) or the COOH-terminal single-copy sequence only (laneslabeled C). The arrowheads indicate identical-sized bands that hybridized with the NH2-terminal-repeat probe and the COOH-terminal probe,respectively. The blots were washed to a posthybridizational stringency of 3x SSC and 0.1% SDS at 55°C. Molecular size standards (inkilobases) are indicated at the left in both panels.
VOL. 10, 1990
_~ _

4514 LEE ET AL.
CONSENSUS SEQUENCE:
CRAM 40-832
B858
610
1e-r1RN40&rn-*C:i y AiIsd&RwQi auD&&-ARUDD-1kCiSK'tbAE3TC SR I KSGt WRN CIPQEWcrG DDNG EG C PP X SQDE E D-- G CRQF7c DDC GEDStvCP _CG PQLWA DDDIG DSE E _C-SAF - SSCX]_C RPD E2 SD-- GFCIHSQc EYD EED
CRAM Asp Gly Cys Pro Thr Glu Asp Ser Pro LysGAT GGA TGC CCT ACG GAG GAT TCC CCC AAG
*** *** *** ***
EGFR Glu Cly Cys Pro Thr Asn Gly Pro Lys
874 CRAM
626 EGFR
-NDIG EC LC
-ETD ITG D
DE
!2.34
:5678
1-22
Ser Ser Asn Ala Lys GlyTCC TCG AAT GCA AAG GGG
*** *** ... ***
Ile Pro Ser Ile Ala Thr Gly
Lys Gly Ser Ser Val Ser Ala GlY Lou leu Leu Leu Ala Gly Ser Thr PheAAA GGA TCG AGC GTC AGT GCG GGA TTG CTG CTT TTG GCT GGT TCC ACG TTTt Val G Ala L.u* Lo Lo Lou*Lou** * . . .*l **LouG Gl Lou Phe
Met Val Gly Ala Leu Leu Leu Leu Leu Val Ala Leu Gly Ile GlY LeaU Phe
891 CRAM Lou Val LeUTTG GTA CTG
645 EGFR Net//
Ala Val Glv Leu Ser Ala Val Leu Phe Leu Glv//GCG GTG GGG CTG TCG GCA GTT TTA TTT TTG GGA
FIG. 8. (A) Comparison of the CRAM repeat with the repetitive binding domain of the human LDL receptor. The alignment of repeats inthe NH2-terminal region of the human LDL receptor and the deduced consensus sequences are as described by Yamamoto et al. (64). Thebottom sequence gives the alignment of three 12-amino-acid repeats of the T. brucei CRAM gene with the human LDL receptor sequence.Homologous residues of the human and trypanosome repeats are boxed. The amino acid abbreviations are: A, Ala; C, Cys; D, Asp; E, Glu;F, Phe; G, Gly; H, His; I, Ile; K, Lys; L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; and W, Trp. (B) Homologybetween CRAM and the Drosophila EGF receptor homolog. The amino acids 858 through 891 of CRAM show homology with the amino acids610 through 645 of the Drosophila EGF receptor homolog. Identical residues are marked with asterisks, conservative amino acid changes are
marked with dots, and the transmembrane region of both proteins is underlined.
clusters uniquely to a single organelle, the flagellar pocket,while the other membrane proteins identified (VSG and theprocyclic acidic repetitive proteins) are lipid anchored anddistributed throughout the membrane (15). It is unclearwhich part of CRAM determines its specific positioning atthe flagellar pocket.Comparison of CRAM with cysteine-rich repeat proteins.
Arrays of regularly spaced cysteine-rich acidic repeats are a
common feature of cell surface receptors of higher eucary-otes. The repeats are grouped into class A and class Bcysteine-rich motifs, depending on the spacing of the cys-teines in the repeats (50-52). Class A cysteine-rich repeatmotifs are found in the human LDL (a cholesterol-carryinglipoprotein) receptor ApoE-binding domain, the LDL-recep-tor-related protein (LRP), the terminal complement compo-nents, and complement factor I (10, 17, 23, 25, 46, 51, 64).Class B motifs occur in the human LDL receptor, the LRP,and the epidermal growth factor (EGF) and nerve growthfactor receptors (22, 27, 36, 54). Several of these proteinsshare domains, as exemplified by the homology between thehuman LDL receptor, the LRP, and the EGF precursor (8,46, 64).When the CRAM protein was compared with several
proteins containing class A repeats, significant amino acidsequence and overall structural features were found to beconserved. Firstly, the consensus sequence of the receptorfor the human plasma LDL consists of class A repeats whichmatch the repetitive cysteine-rich acidic CRAM domain in10 residues, while the remaining 7 residues show conserva-tive changes (Fig. 2 and 8A; 64). In both proteins, cross-
linking of the cysteine residues could form short repetitiveamino acid loops with negatively charged residues facingoutward. In the human LDL receptor, these negativelycharged residues are believed to participate in the ionicinteraction involved in the binding of the receptor to itsligand (i.e., apoprotein B and apoprotein E; 1, 38, 63).CRAM has 22 of these 36-amino-acid repeats, comparedwith 8 repeat arrays in the human LDL receptor (Fig. 8A).
Secondly, a serine- and threonine-rich region with severalconsensus sequences for 0-linked glycosylation sites (indi-cated with dashed lines in Fig. 2) precedes the hydrophobictransmembrane domain in both CRAM and the human LDLreceptor. Thirdly, potential N-linked glycosylation sites (un-derlined in the first repeat in Fig. 2; 61) are found in thecysteine-rich repeats of CRAM and the LDL receptor. TheCRAM protein with a predicted molecular mass of 130 kDameasured about 200 kDa in Western blots and may thus beglycosylated. Finally, CRAM, the human LDL receptor, andthe LRP share amino acid sequence homology in the rela-tively short cytoplasmic domain responsible for internaliza-tion of the receptor in humans; residues 801 to 812 in thehuman LDL receptor, residues 4482 to 4493 of the LRP, andresidues 931 to 942 of CRAM are identical in 5 out of 11amino acids (16, 25).
Unlike the human LDL receptor, the CRAM protein lackshomology with the class B repeats of the EGF precursor andconsists of class A repeats only. Finally, the short 43-amino-acid extracellular domain of CRAM, flanking the transmem-brane region, has a 16-amino-acid stretch, 9 of which are
present in the Drosophila EGF receptor homolog (36) of the
A Um 4 A M^
LDL 1-38
291-322
n% xsiaue
'- 'F --'a8..GP.;ZO6 -ZECILDi.TWkI&Eaii
-r
I
MOL. CELL. BIOL.
A mtat+ Nrs
wall v Ia.1w a,. all So aw - -
rsTN E
.**T . ..... P. ...* I
i rNJ -NE -ID

FLAGELLAR POCKET PROTEIN OF T. BRUCEI 4515
human EGF receptor (Fig. 8B). This 16-amino-acid se-quence directly precedes the transmembrane domain of bothCRAM and EGF. This region of the Drosophila EGF recep-tor homolog is without a well-defined function.The only other known proteins with class A repeat motifs
are several of the complement components. The comple-ment components can be distinguished from the LDL recep-tor repeats and the CRAM repeats by the presence of aconserved positively charged segment in the center of eachrepeat (25). The repeats of the terminal complement compo-nents may serve a function similar to that of the LDLreceptor, namely in binding to ApoE-like regions of otherproteins of the complement cascade to stabilize formation ofthe complement complex (50).
Biological significance of CRAM. The conservation of theCRAM gene in several protozoan species, the cysteine-richand acidic nature of the protein, its structural homology tocell surface receptors, and its presence at the flagellar pocketand in vesicles underneath the pocket suggest that CRAM isa membrane receptor with a function in receptor-mediatedendocytosis.The function of CRAM is unclear. Several protozoan cell
surface proteins contain peptide repeats. However, most ofthese proteins evolve rapidly, are expressed in the blood-stream stages of the parasites, and are believed to serve afunction in immune evasion (28, 41). Since CRAM mRNAwas expressed in insect and bloodstream form trypano-somes, it is unlikely that CRAM is a protozoan repeatprotein with a function in immune evasion.CRAM may be a lipoprotein receptor. Coppens et al. (13,
14) had previously predicted the presence of an LDL recep-tor in the flagellar pocket of bloodstream form trypano-somes. Hajduk and co-workers (24) showed the efficientuptake of high-density lipoproteins by endocytosis in blood-stream form trypanosomes. Our unpublished work showedthe efficient uptake, by endocytosis, of high-density lipopro-teins in insect form T. brucei (F. T. Yen et al., unpublisheddata). We had inferred the presence of a high-density lipo-protein receptor in insect form trypanosomes from ourunpublished findings, showing that insect form trypano-somes require lipoprotein particles for optimal growth (F. T.Yen, B. E. Bihain, M. G.-S. Lee, and L. H. T. Van derPloeg, unpublished data; 2). It is unlikely that CRAM and theputative LDL-receptor protein identified by Coppens repre-sent the same protein. This conclusion is based on thefollowing observations. (i) The protein isolated from blood-stream form trypanosomes by Coppens et al. (13) measured86 kDa, while CRAM measures roughly 200 kDa. It is,however, possible that the protein identified by Coppens etal. (13) is a proteolytic cleavage product. (ii) More impor-tantly, Coppens et al. (13) studied the uptake of LDLlipoprotein particles in bloodstream form trypanosomes,while our studies have mainly focused on high-densitylipoprotein uptake in insect form trypanosomes.
It is assumed that the only cell surface protein of blood-stream form T. brucei is the highly antigenic and denselypacked VSG. Changing to the expression of a new VSGallows the parasite to survive immune attack through anti-genic variation. We have characterized an evolutionarilyconserved transmembrane protein which is uniquely locatedat the flagellar pocket in insect form trypanosomes. The levelof expression of CRAM in the long slender form of blood-stream T. brucei is low. However, since CRAM is conservedin T. equiperdum, we assume that CRAM expression hassignificance for bloodstream form trypanosomes. We aretherefore determining the levels of expression of CRAM in
the other developmental stages of bloodstream form try-panosomes (short stumpy bloodstream form and metacyclicform trypanosomes). If the flagellar pocket of metacyclic orbloodstream form trypanosomes can be shown to carryabundant levels of invariant cell surface proteins (40), com-parable to the CRAM protein described here, the pocketmight become the trypanosome's Achilles' heel, serving as apotential target for vaccination or chemotherapy.
ACKNOWLEDGMENTS
We thank Steven Brown, Hui-Min Chung, Sylvia Le Blancq,Gloria Rudenko, and Michael Weiden for critical reading of themanuscript; Tom Delaharo for FACS analysis; and Fred Opperdoesand Paul Michels for discussion and communicating unpublishedresults.
This work was supported by a grant from the John D. andCatherine T. MacArthur Foundation to L.H.T.V.D.P. and by PublicHealth Service grants Al 21784 (to L.H.T.V.D.P.) and HL 21006 (toR.J.D.) from the National Institutes of Health. L.H.T.V.D.P. is aBurroughs Wellcome Scholar in molecular parasitology.
LITERATURE CITED
1. Basu, S. K., J. L. Goldstein, R. G. W. Anderson, and M. S.Brown. 1976. Degradation of cationized low density lipoproteinsand regulation of cholesterol metabolism in homozygous famil-ial hypercholesterolemia fibroblast. Proc. Natl. Acad. Sci. USA73:3178-3182.
2. Black, S., and V. VandeWeerd. 1989. Serum lipoproteins arerequired for multiplication of Trypanosoma brucei brucei underaxenic culture conditions. Mol. Biochem. Parasitol. 37:65-72.
3. Blobel, G., and B. J. Dobberstein. 1975. Transfer of proteinsacross membranes: presence of proteolytically processed andunprocessed nascent immunoglobulin. J. Cell. Biol. 67:835-851.
4. Boetdker, H. 1971. Conformation independent molecular weightdetermination of RNA by gel-electrophoresis. Biochim.Biophys. Acta 240:448 453.
5. Bone, G. T., and M. Steinert. 1956. Isotopes incorporated in thenucleic acids of Trypanosoma mega. Nature (London) 178:308-309.
6. Borst, P. 1986. Discontinuous transcription and antigenic vari-ation in trypanosomes. Annu. Rev. Biochem. 55:701-732.
7. Brown, E. J., D. A. Evans, and K. Vickerman. 1973. Changes inoxidative metabolism and ultra-structure accompanying differ-entiation of the mitochondrion in Trypanosoma brucei. Int. J.Parasitol. 3:691-704.
8. Brown, M. S., and J. L. Goldstein. 1986. A receptor mediatedpathway for cholesterol homeostasis. Science 232:34-47.
9. Brun, R., and M. Schonenberger. 1979. Cultivation and in vitrocloning of procyclic culture form Trypanosoma brucei in asemidefined medium. Acta Trop. 36:289-292.
10. Catterall, C. F., A. Lyons, R. B. Sim, A. J. Say, and T. J. R.Harris. 1987. Characterization of the primary amino acid se-quence of human complement control factor I, from an analysisof cDNA clones. Biochem. J. 242:849-856.
11. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J.Rutter. 1979. Isolation of biologically active ribonucleic acidfrom sucrose enriched ribonuclease. Biochemistry 18:5294-5299.
12. Clayton, C. E., and M. Mowatt. 1989. The procyclic acidicrepetitive proteins of Trypanosoma brucei. J. Biol. Chem.264:15088-15093.
13. Coppens, I., P. Baudhuin, F. R. Opperdoes, and P. J. Courtoy.1988. Receptors for the host low density lipoproteins on thehemoflagellate Trypanosoma brucei: purification and involve-ment in the growth of the parasite. Proc. Natl. Acad. Sci. USA85:6753-6757.
14. Coppens, I., F. R. Opperdoes, P. J. Courtoy, and P. Baudhuin.1987. Receptor mediated endocytosis in the bloodstream formof Trypanosoma brucei. J. Protozool. 34:465-473.
15. Cross, G. A. M. 1987. Eukaryotic protein modification and
VOL. 10, 1990

4516 LEE ET AL.
membrane attachment via phosphatidylinositol. Cell 48:179-181.
16. Davis, G. C., I. R. Driel, D. W. Russel, M. S. Brown, and J. L.Goldstein. 1987. The low density lipoprotein receptor; identifi-cation of amino acids in cytoplasmic domain required for rapidendocytosis. J. Biol. Chem. 262:4075-4082.
17. DiScipio, R. G., D. N. Chakravarti, H. J. Muller-Eberhard, andG. H. Fey. 1988. The structure of human complement compo-nent C7 and the C5B-7 complex. J. Biol. Chem. 263:549-560.
18. Fairlamb, A. H., P. 0. Weislogel, J. H. J. Hoeijmakers, and P.Borst. 1978. Isolation and characterization of kinetoplast DNAfrom bloodstream form of Trypanosoma brucei. J. Cell. Biol.76:293-309.
19. Frevert, U., and E. Reinwald. 1988. Endocytosis and intracellu-lar occurrence of the variant surface glycoprotein in Trypano-soma congolense. J. Ultrastruct. Mol. Struct. Res. 99:137-149.
20. Giannini, S. H., and P. A. D'Alesandro. 1984. Isolation ofprotective antigens from Trypanosoma lewisi by using trypano-static (ablastic) immunoglobulin G from the surface coat. Infect.Immun. 43:617-621.
21. Giannini, S. H., M. Schittini, J. S. Keithly, S. E. Warburton,C. R. Cantor, and L. H. T. Van der Ploeg. 1986. Karyotypeanalysis of Leishmania species and its use in classification andclinical diagnosis. Science 232:761-765.
22. Gray, A., T. J. Dull, and A. Ullrich. 1983. Nucleotide sequenceof epidermal growth factor cDNA predicts a 128,000 molecularweight protein. Nature (London) 303:722-725.
23. Haeffiger, J., J. Tschopp, D. Nardelli, W. Wahli, H.-P. Kocher,M. Tosi, and K. K. Stanley. 1987. Complementary DNA cloningof complement C8B and its sequence homology to C9. Biochem-istry 26:3551-3556.
24. Hajduk, S. L., D. R. Moore, J. Vasudevacharya, H. Siqueira,A. F. Torri, E. M. Tytler, and J. D. Esko. 1989. Lysis ofTrypanosoma brucei by a toxic subspecies of human highdensity lipoprotein. J. Biol. Chem. 264:5210-5217.
25. Herz, J., U. Hamann, S. Rogne, 0. Myklebost, H. Gausepohl,and K. Stanley. 1988. Surface location and high affinity forcalcium of a 500-kd liver membrane protein closely related tothe LDL-receptor suggests a physiological role as lipoproteinreceptor. EMBO J. 7:4119-4127.
26. lovannisci, D. M., K. Kraut, L. Young, and B. Ullman. 1984.Genetic analysis of nucleoside transport in Leishmania dono-vani. Mol. Cell. Biol. 4:1013-1019.
27. Johnson, D., A. Lanahan, R. Buck, A. Sehgal, C. Morgan, E.Mercer, M. Bothwell, and M. Chao. 1986. Expression andstructure of the human NGF receptor. Cell 47:545-554.
28. Kemp, D. J., R. L. Coppel, H. D. Stahl, A. E. Bianco, L. M.Corcoran, P. McIntyre, C. J. Langford, J. M. Favaloro, P. E.Crewther, G. V. Brown, G. F. Mitchell, J. G. Culvenor, andR. F. Anders. 1986. Genes for antigens of Plasmodium falci-parum. Parasitology 91:S83-S108.
29. Kozak, M. 1986. Point mutations define a sequence flanking theAUG initiation codon that modulates translation by eukaryoticribosomes. Cell 44:283-292.
30. Kyte, J., and R. F. Doolittle. 1982. A simple method fordisplaying the hydrophobic character of a protein. J. Mol. Biol.157:105-132.
31. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
32. Langreth, S. G., and A. E. Balber. 1975. Protein uptake anddigestion in bloodstream and culture forms of Trypanosomabrucei. J. Protozool. 22:40-53.
33. Le Blancq, S. M., B. W. Swinkels, W. C. Gibson, and P. Borst.1988. Evidence for gene conversion between the phosphoglyc-erate kinase genes of Trypanosoma brucei. J. Mol. Biol. 200:439-447.
34. Lee, M. G.-S., and L. H. T. Van der Ploeg. 1987. Frequentindependent duplicative transpositions activate a single VSGgene. Mol. Cell. Biol. 7:357-364.
35. Le Trant, N., S. R. Meshnick, K. Kitchener, J. W. Eaton, and A.Cerami. 1983. Iron-containing superoxide dismutase fromCrithidia fasciculata. J. Biol. Chem. 258:125-130.
36. Livneh, E., L. Glazer, D. Segal, J. Schlessinger, and B.-Z. Shilo.1985. The drosophila EGF receptor gene homologue: conserva-tion of both hormone binding and kinase domains. Cell 40:599-607.
37. Longacre, S., U. Hibner, A. Raibaud, H. Eisen, T. Baltz, C.Giroud, and D. Baltz. 1983. DNA rearrangements and antigenicvariation in Trypanosoma equiperdum: multiple expression-linked sites in independent isolates of trypanosomes expressingthe same antigen. Mol. Cell. Biol. 3:399-409.
38. Mahley, R. W., T. L. Innerarity, R. E. Pitas, K. H. Weisgraber,J. H. Brown, and E. Cross. 1977. Inhibition of lipoproteinbinding to the cell surface receptors of fibroblasts followingselective modification of arginyl residues in arginine-rich and Bapoproteins. J. Biol. Chem. 252:7279-7287.
39. Mowatt, M., and C. E. Clayton. 1987. Developmental regulationof a novel repetitive protein of Trypanosoma brucei. Mol. Cell.Biol. 7:2838-2844.
40. Olenick, J. G., R. Wolf, R. K. Nauman, and J. McLaughlin.1988. A flagellar pocket membrane fraction from Trypanosomabrucei rhodesiense: immunogold localization and nonvariantimmunoprotection. Infect. Immun. 56:92-98.
41. Peterson, D. S., R. A. Wrightsman, and J. E. Manning. 1986.Cloning of a major surface-antigen gene of Trypanosoma cruziand identification of a nonapeptide repeat. Nature (London)322:566-568.
42. Porter, A. G., C. Barber, N. H. Carey, R. A. Hallewell, G.Threfall, and J. S. Emtage. 1979. Complete nucleotide sequenceof an influenza virus hemagglutinin gene clone DNA. Nature(London) 282:471-477.
43. Richardson, J. P., R. P. Beecroft, D. L. Tolson, M. K. Liu, andT. W. Pearson. 1988. Procyclin: an unusual immunodominantglycoprotein surface antigen from the procyclic stage of Africantrypanosomes. Mol. Biochem. Parasitol. 31:213-216.
44. Richardson, J. P., L. Jenni, R. P. Beecroft, and T. W. Pearson.1986. Procyclic tsetse fly midgut forms and culture forms ofAfrican trypanosomes share stage and species specific surfaceantigens identified by monoclonal antibodies. J. Immunol. 86:2259-2264.
45. Roditi, I., M. Carrington, and M. Turner. 1987. Expression of apolypeptide containing a dipeptide repeat is confined to theinsect form of Trypanosoma brucei. Nature (London) 325:272-274.
46. Russel, D. W., W. J. Schneider, T. Yamamoto, K. L. Luskey,M. S. Brown, and J. L. Goldstein. 1984. Domain map of theLDL receptor: sequence homology with the epidermal growthfactor precursor. Cell 37:577-585.
47. Sanger, F., A. R. Coulsen, B. G. Barrell, A. J. H. Smith, and B.Roe. 1980. Cloning in single stranded bacteriophage as an aid torapid sequencing. J. Mol. Biol. 143:161-178.
48. Spindler, K. R., D. S. E. Rosser, and A. J. Berk. 1984. Analysisof adenovirus transforming proteins from early regions 1A and1B with antisera to inducible fusion antigens produced inEscherichia coli. J. Virol. 49:132-141.
49. Southern, E. M. 1975. Detection of specific sequences amongDNA fragments separated by gel electrophoresis. J. Mol. Biol.98:503-517.
50. Stanley, K. K. 1988. The molecular mechanism of complementC9 insertion and polymerization in biological membranes. Curr.Top. Microbiol. Immunol. 140:49-65.
51. Stanley, K. K., H.-P. Kocher, J. P. Luzio, P. Jackson, and J.Tschopp. 1985. The sequence and topology of human comple-ment component C9. EMBO J. 4:375-382.
52. Stanley, K. K., M. Page, A. K. Campbell, and J. P. Luzio. 1986.A mechanism for the insertion of complement component C9into target membranes. Mol. Immunol. 23:451-458.
53. Tobin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
54. Ullrich, A., L. Coussens, J. S. Hayflick, T. J. Dull, A. Gray,A. W. Tam, J. Lee, Y. Yarden, T. A. Liberman, J. Schlessinger,J. Downward, E. L. V. Mayes, N. Whittle, M. D. Waterfield, andP. H. Seeburg. 1984. Human epidermal growth factor cDNA
MOL. CELL. BIOL.

FLAGELLAR POCKET PROTEIN OF T. BRUCEI 4517
sequence and aberrant expression of the amplified gene in A431epidermal carcinoma cells. Nature (London) 309:419-425.
55. Van der Ploeg, L. H. T. 1987. Control of variant surfaceglycoprotein switching in trypanosomes. Cell 51:159-161.
56. Van der Ploeg, L. H. T., D. Valerio, T. De Lange, A. Bernards,and P. Borst. 1982. An analysis of cosmid clones of nuclearDNA from Trypanosoma brucei shows that the genes for variantsurface glycoproteins are clustered in the genome. NucleicAcids Res. 10:593-609.
57. Vickerman, K. 1%9. On the surface coat and flagellar adhesionin trypanosomes. J. Cell. Sci. 5:163-193.
58. Vickerman, K. 1969. The fine structure of Trypanosoma congo-
lense in its bloodstream phase. J. Protozool. 16:54-69.59. Vickerman, K. 1985. Developmental cycles and biology of
pathogenic trypanosomes. Br. Med. Bull. 41:105-114.
60. Von Heijne, G. 1983. Patterns of amino acid near signal se-
quence cleavage sites. Eur. J. Biochem. 133:17-21.61. Ward, C. W., and T. A. Dopheide. 1979. Primary structure of
the Hong Kong (H3) heamagglutinin. Br. Med. Bull. 35:51-56.62. Webster, P., and D. J. Grab. 1988. Intracellular colocalization of
variant surface glycoprotein and transferrin-gold in Trypano-soma brucei. J. Cell Biol. 106:279-288.
63. Weisgraber, K. H., T. L. Innerarity, and R. W. Mahley. 1978.Role of the lysine residues of plasma lipoproteins in high affinitybinding to cell surface receptors on human fibroblasts. J. Biol.Chem. 253:9053-9062.
64. Yamamoto, T., C. G. Davis, M. S. Brown, W. J. Schneider,M. L. Casey, J. L. Goldstein, and D. W. Russell. 1984. Thehuman LDL receptor: a cysteine-rich protein with multiple Alusequences in its mRNA. Cell 39:27-38.
VOL. 10, 1990