characterisation of porcine bone marrow progenitor cells identified by the anti-c-kit (cd117)...
TRANSCRIPT

Journal of Immunological Methods 321 (2007) 70–79www.elsevier.com/locate/jim
Research paper
Characterisation of porcine bone marrow progenitor cells identifiedby the anti-c-kit (CD117) monoclonal antibody 2B8/BM
C. Pérez a,1, S. Moreno a,1, A. Summerfield b, N. Domenech c, B. Alvarez a,1, C. Correa d,F. Alonso a,1, A. Ezquerra a,1, J. Domínguez a,1,2, C. Revilla a,⁎,2
a Dpto. Biotecnología, Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), 28040, Madrid, Spainb Institute of Virology and Immunoprophylaxis, Mittelhäusern, Switzerlandc Unidad de Investigación, CHU Juan Canalejo, 15006, La Coruña, Spain
d Servicio de Cirugia Experimental, Dpto de Investigación, Hospital Ramón y Cajal, 28034, Madrid, Spain
Received 6 November 2006; received in revised form 4 January 2007; accepted 7 January 2007Available online 5 February 2007
Abstract
c-kit (CD117) plays an important role in the early stages of haematopoiesis. Previous studies of porcine haematopoietic stem cellshave relied for their identification on the use of the c-kit ligand stem cell factor. Here, we describe a newmAb, 2B8/BM, that recognizes a155-kDa protein expressed on a small subset (2–8%) of bone marrow haematopoietic cells. 2B8/BM+ cells have a blast appearance, andare mostly negative for lineage-specific markers or express low levels of CD172a or SLA-II. In in vitro colony-forming unit assays thesecells were able to give rise to erythroid and myeloid colonies. Altogether these data suggested that the 2B8/BM antigen might be theporcine orthologue of the human c-kit. This specificity was confirmed by the binding of mAb 2B8/BM to CHO cells transfected with aplasmid encoding the porcine c-kit ectodomain. This antibody can facilitate the isolation and enrichment of porcine stem cells to be usedin procedures aimed to induce xenograft tolerance or to test their potential to repair damaged tissues and organs.© 2007 Elsevier B.V. All rights reserved.
Keywords: c-kit; CD117; Porcine; Haematopoietic stem cells
1. Introduction
The pig is a useful animal model for biomedicalresearch in allo-and xeno-transplantation as well as inregenerative medicine. Organ size and physiological
Abbreviations: GVHD, graft-versus-host disease; HSC, haemato-poietic stem cells; BMC, bone marrow cells; FSC, forward scatter;SSC, side scatter.⁎ Corresponding author. Departamento de Biotecnología, INIA, Ctra
de la Coruña, km 7, 28040 Madrid, Spain. Tel.: +34 91 347 3758; fax:+34 91 357 2293.
E-mail address: [email protected] (C. Revilla).1 Tel.: +34 91 347 3758; fax: +34 91 357 2293.2 These two authors share the senior authorship.
0022-1759/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jim.2007.01.003
similarities between humans and pigs make these animalssuitable organ donors for humans (Sachs, 1994; Cooperet al., 2002). However, organ transplantation acrossspecies is prone to vigorous immunologic rejection. Trans-plantation of whole bone marrow cells or selected subsetshas been a successful strategy to achieve chimerism andinduction of tolerance in different allo-or xeno-transplan-tation studies (de Vries–van der Zwan et al., 1998; Abeet al., 2002). Nevertheless, in spite of promising results,this approach remains limited in part because certain bonemarrow cell subpopulations may cause graft-versus-hostdisease (GVHD) or engraftment failure.
Haematopoietic stemcells (HSC) are themost primitivecells that establish a fully self-renewing population and

71C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
retain multilineage haematopoietic differentiation poten-tial. Unlike whole bone marrow, HSC-enriched popula-tions do not contain mature lymphocytes capable ofinducing GVHD. In the adult bone marrow mosthaematopoietic progenitors express c-kit (CD117), amember of a family of cell-surface receptors with tyrosinekinase activity. Purified bone marrow-derived c-kit+ cellshave been used in mice to induce stable multilineagemixed chimerism and permanent acceptance of donor-specific grafts without any clinical signs of GVHD (deVries–van der Zwan et al., 1998). Human HSC with long-term engrafting abilities are also almost exclusively in thec-kit+ population of CD34+ bone marrow cells (Kawa-shima et al., 1996).
On the other hand, the recent demonstration of thecapacity of bone marrow HSC to differentiate intononhaematopoietic lineages such as hepatocytes, skel-etal muscle cells, neurons or epithelial cells of the lung,gastrointestinal tract and skin, has raised hopes as totheir therapeutic potential in regenerative medicine(Dalakas et al., 2005).
Characterisation of porcine haematopoietic progen-itor cells has been hampered by the lack of specificreagents. Swine haematopoietic c-kit+ cells have beenanalysed and sorted by using recombinant porcine stemcell factor (rpSCF), the ligand of the c-kit receptor(Summerfield et al., 2003; Le Gern et al., 2003). Thisapproach has shown that porcine c-kit+ but not c-kit−
bone marrow cells are capable of reconstituting long-term haematopoiesis in NOD/SCID mice expressingporcine cytokine transgenes. However, binding of SCFto c-kit receptor leads to its internalisation what couldmake difficult the characterisation and isolation of c-kit+
cells (Miyazawa et al., 1994; Yee et al., 1994).Here, we describe a new mAb, named 2B8/BM, that
recognises a population of bone marrow progenitor cells.The molecular size of the antigen and the phenotype anddifferentiation abilities of 2B8/BM+ cells suggested thatthis mAb reacted with the porcine c-kit ortologue.Confirmation of the porcine c-kit specificity was obtainedby its reactivity with CHO transfectants expressing the c-kit ectodomain. This antibody may facilitate the isolationand enrichment of porcine c-kit positive bone marrowstem cells for testing their potential in the treatment of avariety of clinical conditions.
2. Materials and methods
2.1. Animals and cells
Three-month-old Large-White outbred pigs wereused to obtain bone marrow cells by perfusing ribs or
the sternum with PBS plus EDTA at 4 °C. Erythrocytesand non-viable cells were eliminated by ficoll gradientcentrifugation (at 350 ×g for 15 min on lymphocyteseparation medium at 1.077 g/ml). Cells were washedtwice with PBS plus EDTA and resuspended in RPMI tofurther analysis.
Blood samples were obtained from 14-to 20-month-old Large-White outbred pigs. Peripheral blood mono-nuclear cells were isolated on Percoll discontinuousgradients after blood sedimentation in dextran, aspreviously described (Gonzalez et al., 1990). Granulo-cytes were recovered from the lower Percoll phase, afterlysis of residual erythrocytes by hypotonic treatment.Platelets were obtained from the platelet-rich plasmafraction resulting from the centrifugation of 50 ml ofnormal swine blood for 30 min at 350 g.
The reported experiments have been executed in fullcompliance with guidelines by the ethical committee ofthe Institute.
2.2. Monoclonal antibody production
Hybridoma secreting mAb 2B8/BM was derivedfrom a fusion of Sp2/0 myeloma cells with spleen cellsfrom a BALB/c mouse, immunized with porcine bonemarrow cells (BMC) according to current protocols(Kohler and Milstein, 1975). MAb isotype wasdetermined by ELISA, using rat anti-mouse heavy andlight chains antisera and a peroxidase-conjugated rabbitanti-rat Ig (BD Pharmingen, CA, USA). 2B8/BM mAbis isotype IgG1 κ.
mAbs to porcine SWC8 (MIL3, IgM) and to CD11R1(MIL4, IgG1) were kindly provided by K. Haverson(Bristol University, UK); to CD2 (MSA4; IgG2a), CD4(72–12–4; IgG2b) and CD8α (76–2-11; IgG2a) by J.Lunney (ARS USDA, Beltsville, USA); to CD3 (BB23–8E; IgG2b) by M. Pescovitz (Indiana University, USA).mAb toCD16 (G7, IgG1)was a gift fromY.B.Kim (FinchUniversity ofHealth Sciences, IL, USA).mAbs to porcineCD172a (BA1C11, IgG1), SLAI (BL6H4; IgG1), CD45(2A5, IgG1), CD18 (BA3H2, IgG1), CD11a (BL1H8,IgG2b), CD11R3 (BL3F1, IgG1), CD45RA (6E3/7;IgG1), CD163 (2A10/11; IgG1), CD5 (1H6/8; IgG2a),SLAIIDR (1F12; IgG2b) and to neutrophils (6D10;IgG2a; Perez et al., 2007) were produced in our labo-ratory. MAbs to CD21 (B-Ly4; IgG1) and to CD11R2(S-Hcl3, IgG2b) were purchased from BD Pharmingen(CA, USA).
For two-color immunofluorescence assays, mAbswere purified by affinity chromatography on Protein A-Sepharose CL4B, and labeled with biotin followingstandard protocols.

72 C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
2.3. Isolation of 2B8/BM+ cells and cytological analysis
BMC were incubated with 2B8/BM hybridomasupernatant for 45 min on ice. Then cells were washedwith PBS containing 5% FCS and 2 mM EDTA (MACSbuffer) and incubated with goat anti-mouse IgGmagnetic microbeads (Myltenyi Biotec, Germany) for15 min on ice. After washing with MACS buffer, cellswere passed through a MACS separation column (VS+
positive selection) and magnetically labeled cells (2B8/BM+) and the effluent negative fraction (2B8/BM−
cells) were collected.Isolated 2B8/BM+ cells were centrifuged at 80 ×g
during 4 min in a Shandon centrifuge (Cytospin 3).Cytospin preparations were fixed by treatment with coldmethanol for 10 min, and then stained with May-Grünwald/Giemsa dye. Analyses were carried out in aLeica DMLB microscope (Germany).
2.4. Immunohistochemical analyses
Femur samples were fixed and decalcified in a solutionof 1% acetic acid in 10% buffered formalin for at least48 h, and embedded in paraffin wax. Serial 4 μm sectionswere cut, placed on silane coated slides, and allowed todry at room temperature. Slides were dewaxed in xyleneand rehydrated through graded alcohols to distilled water.Slides were then subjected to heat-mediated antigenretrieval treatment and stained according to previousdescribed protocols (Domenech et al., 2003).
2.5. Progenitor cell assay
The colony-forming activity of unfractionated BMC,2B8/BM+ and 2B8/BM− cells (isolated as describedabove) was determined by culture in methylcellulosemedium supplemented with 10% FBS (MethoCult;StemCell Technologies, Meylan, France), penicillin(100 U/ml), streptomycin 100 μg/ml (both Invitrogen),erythropoietin (3 U/ml; Roche), rpSCF (20 U/ml), andrpGM-CSF (20 U/ml). Bone marrow cells were plated at25,000 cells and 2B8/BM+ and 2B8/BM− cells at 10,000cells per 35 ml petri dish (StemCell Technologies) intriplicate cultures, and cultured for 9 days at 39 °C. Then,colonieswere identifiedmorphologically bymicroscopy asburst-forming-units-erythroid (BFU-E) or colony-forming-units granulocyte-macrophage (CFU-GM) and counted.
2.6. Flow cytometry
For single-color staining, cells (5×105) were incubat-edwith 75μl hybridoma supernatant ofmAb for 30min at
4 °C. After two washes in PBS containing 0.1% BSA and0.01% sodium azide (washing buffer), cells wereincubated with FITC-conjugated rabbit F(ab′)2 anti-mouse Ig (Dako, Denmark). Then, they were washedand fixed in 0.1% formaldehyde prior to analysis on aFACScalibur cytometer (Becton Dickinson, USA). Irrel-evant unlabeled isotype-matched mAbs were used asnegative controls.
For two-color staining, after incubation with FITC-conjugated rabbit F(ab′)2 anti-mouse Ig (Dako, Den-mark), cells were washed and free binding sites blockedwith 5% of normal mouse serum for 15 min. Then,biotinylated mAbs were added and detected withstreptavidin-phycoerythrin (PE) (BD Pharmingen, CA,USA). Subsequently, cells were washed and fixed in0.1% formaldehyde prior to analysis on a FACScaliburcytometer (Becton Dickinson, USA). Irrelevant isotype-matched mAbs, unlabeled or labeled with the differentfluorochromes were used as negative controls.
In cross-blocking experiments SWC8+ granulocyticbone marrow cells were removed by magnetic immu-nobead subtraction. To this end, cells were incubatedwith anti-SWC8 mAb, followed by anti-mouse IgM-biotin (Southern Biotechnology, Birminghan, AL, USA)and streptavidin-microbeads (Myltenyi Biotec, Ger-many). After washing with MACS buffer, cells werepassed through a MACS separation column (MyltenyiBiotec, Germany) and the effluent negative fraction(SWC8− cells) was collected. Cells were then incubatedwith 2B8/BM mAb followed by rpSCF as describedbefore (Summerfield et al., 2003).
2.7. Immunoprecipitation and Western blotting analyses
Cells (5×107) were washed three times in phosphatebuffered saline (PBS) and resuspended in 5 ml of PBS.Sulfo-NHS-biotin (Pierce, USA) (0.4 mg/ml, finalconcentration) was added to the cells and incubated for15min at 4 °C. After washing three times with PBS, cellswere lysed with 0.5 ml of a lysis buffer consisting of10 mM Tris–HCl pH 7.4, 150 mM NaCl, 1 mM EDTA,1% NP40, 0.6 U/ml aprotinin and 3 mM phenylmethyl-sulphonylfluoride. The lysate was precleared twice with50 μl of a 25% (v/v) suspension of protein G-Sepharose(Pharmacia, Sweden) in lysis buffer. Hybridoma super-natant (0.3 ml) was added to 0.1 ml of lysate andincubated for 2 h at room temperature. Then, 40 μl of25% (v/v) suspension of protein G-Sepharose wereadded and incubated for 1 h with gentle mixing. Beadswere washed three times with lysis buffer, boiled inelectrophoresis sample buffer (0.062 M Tris–HCl pH6.8, 2% SDS, 10% glycerol, 0.7 M 2-mercaptoethanol,

73C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
0.001% bromophenol blue) and supernatants run on a10%SDS-PAGE and transferred to nitrocellulose. Filterswere incubated with streptavidin–peroxidase (Pierce,USA) and bands visualized with the ECL detection assayfollowing the recommendations of the manufacturer(Amersham, England).
For Western blotting, cell lysates were run on a 10%SDS-PAGE under non-reducing conditions and thentransferred to nitrocellulose. After blocking, filters wereincubated with hybridoma supernatants for 1 h at roomtemperature, followed by 1 h of incubation with aperoxidase-labeled rabbit anti-mouse Ig (Dako, Den-mark). Peroxidase activity was visualized using the ECLdetection assay as described above.
2.8. Cloning of porcine c-kit ectodomain cDNA
RNAwas isolated from 2B8/BM+ cells using TriPureReagent (Roche Diagnostics GmbH, Germany) followingthe manufacturer's instructions. First-strand cDNA wasobtained from5μg of total RNA (previously denatured byheating for 2 min at 65 °C and immediately placed on ice)in a total volume of 50 μl of buffer containing 50 mMTris–HCl (pH 8.6), 75 mM KCl, 10 mMMgCl2, 10 mMDTT, 50 μM each dNTPs (Epicentre, WI, USA), 0.5 mMoligo(dT), 20 U of RNasin ribonuclease inhibitor(Promega, WI, USA), and 12.5 U of Moloney murineleukaemia virus RT (Epicentre, WI, USA). The reactionmixtures were incubated for 1 h at 37 °C. Based on a
Fig. 1. Flow cytometric analysis of 2B8/BMmAb reactivity in bloodA) and bonB). Cells were stained with 2B8/BMmAb followed by rabbit F(ab′)2 anti-mous(open histogram). 30,000 blood and 50,000 bone marrow cells were acquired.
partial sequence of porcine c-kit gene (GeneBankaccession no. AJ223228), two primers were designedfor PCR amplification of the ectodomain (ECD) codingsequence: c-kitExF (GAGAGCAGGAACGTGGAAAG)and c-kitExR (GGTGGGCATGGATTTGTTC). Thetemplate cDNA from 2B8/BM+ cells was amplifiedusing BD AdvantageTM 2 PCR Enzyme System asrecommended by the manufacturer (BD BioscienceClontech, CA, USA) with 0.5 mM of MgCl2. Afterdenaturation of cDNA at 94 °C for 5 min, amplificationwas performed for 40 cycles at 94 °C for 45 s, 51 °C for45 s and 72 °C for 2 min followed by an extra-extensiontime of 10 min at 72 °C. After agarose-gel electrophoresisof PCR-amplified products, a band of approximately1592 bp was purified from the gel and cloned into thepcDNA3.1/CT-GFP-TOPO (Invitrogen, CA, USA) fol-lowing the manufacturer's instructions. Several cloneswere sequenced to confirm the orientation and theintegrity of the inserts. Four silent point mutations weredetected in all clones sequenced compared with theporcine c-kit reference sequence (GeneBank accessionno. AJ223228).
2.9. Transient expression of c-kit–GFP constructs andanalysis by flow cytometry
CHO cells were transfected with plasmid pECD-c-kit-GFP, encoding ECD-c-kit-GFP fusion protein, by usingthe LipofectAMINE PLUS Reagent (Invitrogen, CA,
emarrow cells B). FSC/SSC dot-plot of 2B8/BM+ gated cells is showed ine Ig FITC. Irrelevant isotype-matched mAb was used as negative controlResults are representative of three independent experiments.

Fig. 2. A) Immunohistochemical analysis of 2B8/BM+ cells (arrows)distribution on formalin-fixed, paraffin-embedded sections of bonemarrow. Original magnification: ×400. B) Morphology of 2B8/BM+
cells. Cells were isolated by using magnetic microbeads, spun ontoslides and stained with May-Grünwald/Giemsa. Original magnifica-tion: ×1000.
74 C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
USA) accordingly to the manufacturer's instructions.Briefly, the day prior to transfection 3×105 CHO cellswere seeded in M6 plates (NUNC, Denmark) and grownin Dulbecco's modified Eagle's minimal essentialmedium (DEMEM) (BioWhittaker, Belgium) supple-mentedwith 30 μg/ml gentamicin, 2 mML-glutamine and10% foetal bovine serum (FBS) (BioWhittaker, Belgium).Immediately before transfection, cells were rinsed withmedium without serum and antibiotic (transfectionmedium, TM). For each well, 1 μg of plasmid DNAand 6 μl of PLUS reagent weremixed, diluted in 500 μl ofTM and incubated for 15 min at room temperature. Afterthat, 4 μl of Lipofectamine diluted in 500 μl of TM, wasadded to the transfectionmixture. After 15min incubationat room temperatute, the DNA/LipofectAMINE PLUScomplexes were added to the cells. Cells were incubatedat 37 °C in a 5% CO2 atmosphere for 3 h. Then 1 ml offresh growth medium was added and cells cultured for24 h. Monensin (2 μg/ml), a protein secretion inhibitor,was added 4 h before harvesting. Cells were permeabi-lized with FACS™ Permeabilizing Solution 2 (BDBiosciences Immunocytometry Systems, San Jose, CA),and incubated for 30 min with 100 μl of 2B8/BMhybridoma supernatant. After three washes in PBScontaining 0.1% BSA and 0.01% sodium azide, cellswere incubated for 30 min with PE-conjugated rabbit F(ab′)2 anti-mouse Ig (Dako, Denmark). Cells werewashed again and analysed on a FACScalibur flowcytometer (Becton Dickinson, USA). Non-transfectedcells and transfected cells stained with an irrelevantisotype-matched mAb were used as negative controls.
3. Results
3.1. Phenotypic and morphological characterisation of2B8/BM+ cells
Mab 2B8/BM reacted with a minor subset of adultBMC (2–8%), while it did not stain peripheral bloodlymphocytes, monocytes, granulocytes, platelets orerythrocytes (Fig. 1A, B). The majority of the 2B8/BM+ cells exhibited a FSCmedium/highSSClow profile butthere was a small population of FSChighSSChigh cells(Fig. 1B).
On formalin-fixed, paraffin-embedded bone marrowsections, mAb 2B8/BM labeled medium-size cells withround or oval shape, located within haematopoieticniches scattered among cells of different lineages(Fig. 2A). Morphologically, in cytospin preparationsstained with May-Grünwald/Giemsa, 2B8/BM+ cellsisolated by immunomagnetic positive selectionappeared as a heterogeneous population of medium to
large-size cells, of blastic appearance, with blue cyto-plasm and round nucleus having fine or slightlycondensed chromatin and one to three prominent nu-cleoli (Fig. 2B).
Further phenotypic characterisation by two-colorflow cytometry showed that over 80% of 2B8/BM+
cells were positive for SLA-I, CD45 and CD18 (Fig. 3).Approximately 40–50% of them expressed SLA-II,CD11a and CD172a at low levels, and CD11R3 atintermediate levels. A low expression of CD16 was alsodetected in less than 25% of these cells. As expected forprogenitor BMC, 2B8/BM+ cells did not expresslineage markers such as CD2, CD3, CD21, CD4,CD5, CD8α, CD163 or 6D10 (Fig. 3 and data not
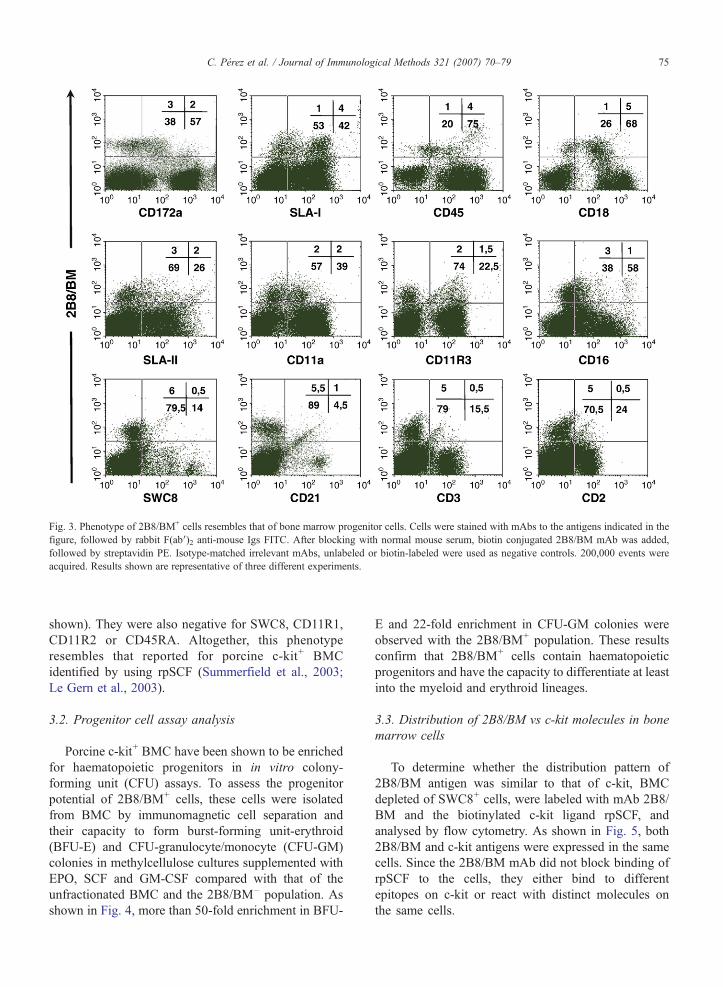
Fig. 3. Phenotype of 2B8/BM+ cells resembles that of bone marrow progenitor cells. Cells were stained with mAbs to the antigens indicated in thefigure, followed by rabbit F(ab′)2 anti-mouse Igs FITC. After blocking with normal mouse serum, biotin conjugated 2B8/BM mAb was added,followed by streptavidin PE. Isotype-matched irrelevant mAbs, unlabeled or biotin-labeled were used as negative controls. 200,000 events wereacquired. Results shown are representative of three different experiments.
75C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
shown). They were also negative for SWC8, CD11R1,CD11R2 or CD45RA. Altogether, this phenotyperesembles that reported for porcine c-kit+ BMCidentified by using rpSCF (Summerfield et al., 2003;Le Gern et al., 2003).
3.2. Progenitor cell assay analysis
Porcine c-kit+ BMC have been shown to be enrichedfor haematopoietic progenitors in in vitro colony-forming unit (CFU) assays. To assess the progenitorpotential of 2B8/BM+ cells, these cells were isolatedfrom BMC by immunomagnetic cell separation andtheir capacity to form burst-forming unit-erythroid(BFU-E) and CFU-granulocyte/monocyte (CFU-GM)colonies in methylcellulose cultures supplemented withEPO, SCF and GM-CSF compared with that of theunfractionated BMC and the 2B8/BM– population. Asshown in Fig. 4, more than 50-fold enrichment in BFU-
E and 22-fold enrichment in CFU-GM colonies wereobserved with the 2B8/BM+ population. These resultsconfirm that 2B8/BM+ cells contain haematopoieticprogenitors and have the capacity to differentiate at leastinto the myeloid and erythroid lineages.
3.3. Distribution of 2B8/BM vs c-kit molecules in bonemarrow cells
To determine whether the distribution pattern of2B8/BM antigen was similar to that of c-kit, BMCdepleted of SWC8+ cells, were labeled with mAb 2B8/BM and the biotinylated c-kit ligand rpSCF, andanalysed by flow cytometry. As shown in Fig. 5, both2B8/BM and c-kit antigens were expressed in the samecells. Since the 2B8/BM mAb did not block binding ofrpSCF to the cells, they either bind to differentepitopes on c-kit or react with distinct molecules onthe same cells.
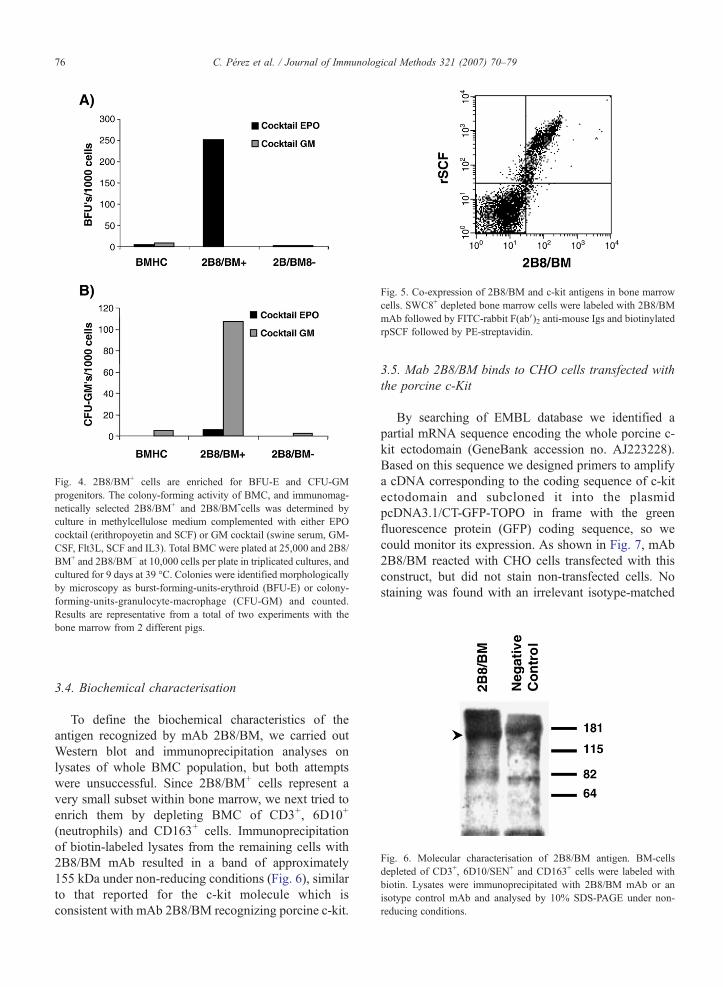
Fig. 4. 2B8/BM+ cells are enriched for BFU-E and CFU-GMprogenitors. The colony-forming activity of BMC, and immunomag-netically selected 2B8/BM+ and 2B8/BM-cells was determined byculture in methylcellulose medium complemented with either EPOcocktail (erithropoyetin and SCF) or GM cocktail (swine serum, GM-CSF, Flt3L, SCF and IL3). Total BMC were plated at 25,000 and 2B8/BM+ and 2B8/BM− at 10,000 cells per plate in triplicated cultures, andcultured for 9 days at 39 °C. Colonies were identified morphologicallyby microscopy as burst-forming-units-erythroid (BFU-E) or colony-forming-units-granulocyte-macrophage (CFU-GM) and counted.Results are representative from a total of two experiments with thebone marrow from 2 different pigs.
Fig. 5. Co-expression of 2B8/BM and c-kit antigens in bone marrowcells. SWC8+ depleted bone marrow cells were labeled with 2B8/BMmAb followed by FITC-rabbit F(abV)2 anti-mouse Igs and biotinylatedrpSCF followed by PE-streptavidin.
Fig. 6. Molecular characterisation of 2B8/BM antigen. BM-cellsdepleted of CD3+, 6D10/SEN+ and CD163+ cells were labeled withbiotin. Lysates were immunoprecipitated with 2B8/BM mAb or anisotype control mAb and analysed by 10% SDS-PAGE under non-reducing conditions.
76 C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
3.4. Biochemical characterisation
To define the biochemical characteristics of theantigen recognized by mAb 2B8/BM, we carried outWestern blot and immunoprecipitation analyses onlysates of whole BMC population, but both attemptswere unsuccessful. Since 2B8/BM+ cells represent avery small subset within bone marrow, we next tried toenrich them by depleting BMC of CD3+, 6D10+
(neutrophils) and CD163+ cells. Immunoprecipitationof biotin-labeled lysates from the remaining cells with2B8/BM mAb resulted in a band of approximately155 kDa under non-reducing conditions (Fig. 6), similarto that reported for the c-kit molecule which isconsistent with mAb 2B8/BM recognizing porcine c-kit.
3.5. Mab 2B8/BM binds to CHO cells transfected withthe porcine c-Kit
By searching of EMBL database we identified apartial mRNA sequence encoding the whole porcine c-kit ectodomain (GeneBank accession no. AJ223228).Based on this sequence we designed primers to amplifya cDNA corresponding to the coding sequence of c-kitectodomain and subcloned it into the plasmidpcDNA3.1/CT-GFP-TOPO in frame with the greenfluorescence protein (GFP) coding sequence, so wecould monitor its expression. As shown in Fig. 7, mAb2B8/BM reacted with CHO cells transfected with thisconstruct, but did not stain non-transfected cells. Nostaining was found with an irrelevant isotype-matched
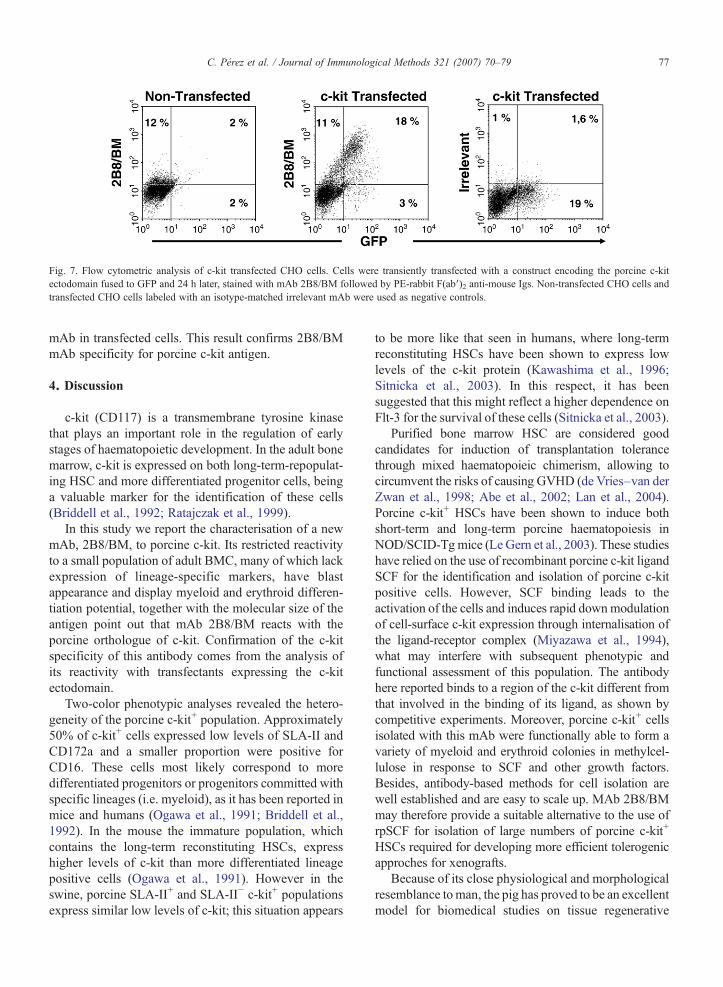
Fig. 7. Flow cytometric analysis of c-kit transfected CHO cells. Cells were transiently transfected with a construct encoding the porcine c-kitectodomain fused to GFP and 24 h later, stained with mAb 2B8/BM followed by PE-rabbit F(ab′)2 anti-mouse Igs. Non-transfected CHO cells andtransfected CHO cells labeled with an isotype-matched irrelevant mAb were used as negative controls.
77C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
mAb in transfected cells. This result confirms 2B8/BMmAb specificity for porcine c-kit antigen.
4. Discussion
c-kit (CD117) is a transmembrane tyrosine kinasethat plays an important role in the regulation of earlystages of haematopoietic development. In the adult bonemarrow, c-kit is expressed on both long-term-repopulat-ing HSC and more differentiated progenitor cells, beinga valuable marker for the identification of these cells(Briddell et al., 1992; Ratajczak et al., 1999).
In this study we report the characterisation of a newmAb, 2B8/BM, to porcine c-kit. Its restricted reactivityto a small population of adult BMC, many of which lackexpression of lineage-specific markers, have blastappearance and display myeloid and erythroid differen-tiation potential, together with the molecular size of theantigen point out that mAb 2B8/BM reacts with theporcine orthologue of c-kit. Confirmation of the c-kitspecificity of this antibody comes from the analysis ofits reactivity with transfectants expressing the c-kitectodomain.
Two-color phenotypic analyses revealed the hetero-geneity of the porcine c-kit+ population. Approximately50% of c-kit+ cells expressed low levels of SLA-II andCD172a and a smaller proportion were positive forCD16. These cells most likely correspond to moredifferentiated progenitors or progenitors committed withspecific lineages (i.e. myeloid), as it has been reported inmice and humans (Ogawa et al., 1991; Briddell et al.,1992). In the mouse the immature population, whichcontains the long-term reconstituting HSCs, expresshigher levels of c-kit than more differentiated lineagepositive cells (Ogawa et al., 1991). However in theswine, porcine SLA-II+ and SLA-II− c-kit+ populationsexpress similar low levels of c-kit; this situation appears
to be more like that seen in humans, where long-termreconstituting HSCs have been shown to express lowlevels of the c-kit protein (Kawashima et al., 1996;Sitnicka et al., 2003). In this respect, it has beensuggested that this might reflect a higher dependence onFlt-3 for the survival of these cells (Sitnicka et al., 2003).
Purified bone marrow HSC are considered goodcandidates for induction of transplantation tolerancethrough mixed haematopoieic chimerism, allowing tocircumvent the risks of causing GVHD (de Vries–van derZwan et al., 1998; Abe et al., 2002; Lan et al., 2004).Porcine c-kit+ HSCs have been shown to induce bothshort-term and long-term porcine haematopoiesis inNOD/SCID-Tgmice (Le Gern et al., 2003). These studieshave relied on the use of recombinant porcine c-kit ligandSCF for the identification and isolation of porcine c-kitpositive cells. However, SCF binding leads to theactivation of the cells and induces rapid downmodulationof cell-surface c-kit expression through internalisation ofthe ligand-receptor complex (Miyazawa et al., 1994),what may interfere with subsequent phenotypic andfunctional assessment of this population. The antibodyhere reported binds to a region of the c-kit different fromthat involved in the binding of its ligand, as shown bycompetitive experiments. Moreover, porcine c-kit+ cellsisolated with this mAb were functionally able to form avariety of myeloid and erythroid colonies in methylcel-lulose in response to SCF and other growth factors.Besides, antibody-based methods for cell isolation arewell established and are easy to scale up. MAb 2B8/BMmay therefore provide a suitable alternative to the use ofrpSCF for isolation of large numbers of porcine c-kit+
HSCs required for developing more efficient tolerogenicapproches for xenografts.
Because of its close physiological and morphologicalresemblance toman, the pig has proved to be an excellentmodel for biomedical studies on tissue regenerative

78 C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
therapies (Vodicka et al., 2005). Besides on bone marrowstem cells, c-kit is expressed on other tissue-specificstem cells (i.e. cardiac and neural stem cells) (Leri et al.,2005; Limana et al., 2005; Erlandsson et al., 2004; Daset al., 2004; Sun et al., 2004) and therefore this antibodymay also be useful for investigating the role of these cellsin tissue regeneration. Indeed, the reactivity of mAb2B8/BM on formalin-fixed paraffin-embedded tissueswill facilitate a detailed knowledge of the distribution ofc-kit positive populations in different tissues both inhealthy and pathological conditions and may beparticularly useful for studying the fate of transplantedstem cells in recipient tissues/organs and themechanismsby which damaged tissues are regenerated (Kajsturaet al., 2005; Amado et al., 2005).
Another cell type in which c-kit is expressed inhumans is the mast cell. Although in the present studywe have not investigated the reactivity of mAb 2B8/BMwith porcine mast cells, this antibody may be a usefultool for studies on porcine mast cell differentiation andfunction.
In summary, we have developed and characterised anew monoclonal antibody to the porcine c-kit. Theavailability of this antibody will contribute to a betterknowlegde of the early developmental stages of porcinehaematopoiesis and will facilitate the isolation ofporcine stem cells for studies aimed to explore novelstrategies for improving xenograft tolerance or test theirtherapeutic potential in repairing damaged tissues.
Acknowledgements
1. This work was supported by CICYT grant AGL2004–04074 and INIA grant CPE03–021-C4–2.
2. C.P. was a recipient of a I.N.I.A. predoctoral fellowshipfrom the Ministerio de Educación y Ciencia.
3. C.R. was a recipient of a Ramón y Cajal contractfrom the Ministerio de Educación y Ciencia.
References
Abe, M., Qi, J., Sykes, M., Yang, Y.G., 2002. Mixed chimerisminduces donor-specific T-cell tolerance across a highly disparatexenogeneic barrier. Blood 99, 3823.
Amado, L.C., Saliaris, A.P., Schuleri, K.H., St. John, M., Xie, J.S.,Cattaneo, S., Durand, D.J., Fitton, T., Kuang, J.Q., Stewart, G.,Lehrke, S., Baumgartner, W.W., Martin, B.J., Heldman, A.W.,Hare, J.M., 2005. Cardiac repair with intramyocardial injection ofallogeneic mesenchymal stem cells after myocardial infarction.PNAS 102, 11474.
Briddell, R.A., Broudy, V.C., Bruno, E., Brandt, J.E., Srour, E.F.,Hoffman, R., 1992. Further phenotypic characterization andisolation of human hematopoietic progenitor cells using amonoclonal antibody to the c-kit receptor. Blood 79, 3159.
Cooper, D.K., Gollackner, B., Sachs, D.H., 2002. Will the pig solve thetransplantation backlog? Annu. Rev. Med. 53, 133.
Dalakas, E., Newsome, P.N., Harrison, D.J., Plevris, J.N., 2005.Hematopoietic stem cell trafficking in liver injury. FASEB J. 19,1225.
Das, A.V., James, J., Zhao, X., Rahnenfurher, J., Ahmad, I., 2004.Identification of c-Kit receptor as a regulator of adult neural stemcells in the mammalian eye: interactions with Notch signaling.Dev. Biol. 273, 87.
de Vries-van der Zwan, A., Van der Pol, M.A., Besseling, A.C., deWaal, L.P., Boog, C.J.P., 1998. Haematopoietic stem cells caninduce specific skin graft acceptance across ful MHC barriers.Bone Marrow Transplant. 22, 91.
Domenech, N., Rodríguez-Carreño, M.P., Filgueira, P., Alvarez, B.,Chamorro, S., Domínguez, J., 2003. Identification of porcinemacrophages with monoclonal antibodies in formalin-fixed,paraffin-embedded tissues. Vet. Immunol. Immunopathol. 94,77.
Erlandsson, A., Larsson, J., Forsberg-Nilsson, K., 2004. Stem cellfactor is a chemoattractant and a survival factor for CNS stem cells.Exp. Cell Res. 301, 201.
Gonzalez, S., Mendoza, C., Sánchez-Vizcaino, J.M., Alonso, F., 1990.Inhibitory effect of African swine fever virus on lectin-dependentswine lymphocyte proliferation. Vet. Immunol. Immunopathol. 26,71.
Kajstura, J., Rota, M., Whang, B., Cascapera, S., Hosoda, T., Bearzi,C., Nurzynska, D., Kasahara, H., Zias, E., Bonafe, M., Nadal-Ginard, B., Torella, D., Nascimbene, A., Quaini, F., Urbanek, K.,Leri, A., Anversa, P., 2005. Bone marrow cells differentiate incardiac cell lineages after infarction independently of cell fusion.Circ. Res. 96, 127.
Kawashima, I., Zanjani, E.D., Almaida-Porada, G., Flake, A.W., Zeng,H., Ogawa, M., 1996. CD34+ human marrow cells that expresslow levels of kit protein are enriched for long-term marrow-engrafting cells. Blood 87, 4136.
Kohler, G., Milstein, C., 1975. Continuous cultures of fused cellssecreting antibody of predefined specificity. Nature 256, 495.
Lan, P., Wang, L., Diouf, B., Eguchi, H., Su, H., Bronson, R., Sachs,D.H., Sykes, M., Yang, Y.-G., 2004. Induction of human T-celltolerance to porcine xenoantigens through mixed hematopoieticchimerism. Blood 103, 3964.
Le Gern, A.C., Giovino, M.A., Abe, M., Theodore, P.R., Qi, J., Down,J.D., Sachs, D.H., Sykes, M., Yang, Y.-G., 2003. Stem cell activityof porcine c-kit+ hematopoietic cells. Exp. Hematol. 31, 833.
Leri, A., Kajstura, J., Anversa, P., 2005. Cardiac stem cells andmechanisms of myocardial regeneration. Physiol. Rev. 85, 1373.
Limana, F., Germani, A., Zacheo, A., Kajstura, J., DiCarlo, A.,Borsellino, G., Leoni, O., Palumbo, R., Battistini, L., Rastaldo, R.,Muller, S., Pompilio, G., Anversa, P., Bianchi, M.E., Capogrossi,M.C., 2005. Exogenous high-mobility group box 1 protein inducesmyocardial regeneration after infarction via enhanced cardiac c-kit+
cell proliferation and differentiation. Circ. Res. 97, 73.Miyazawa, K., Toyama, K., Gotoh, A., Hendrie, P.C., Mantel, C.,
Broxmeyer, H.E., 1994. Ligand-dependent polyubiquitination ofc-kit gene product: a posible mechanism of receptor downmodulation in M07 e cells. Blood 83, 137.
Ogawa, M., Matsuzaki, Y., Nishikawa, S., Hayashi, S., Kunisada, T.,Sudo, T., Kina, T., Nakauchi, H., Nishikawa, S., 1991. Expressionand function of c-kit in hemopoietic progenitor cells. J. Exp. Med.174, 63.
Perez, C., Revilla, C., Alvarez, B., Chamorro, S., Correa, C.,Doménech, N., Alonso, F., Ezquerra, A., Domínguez, J., 2007.

79C. Pérez et al. / Journal of Immunological Methods 321 (2007) 70–79
Phenotypic and functional characterization of porcine granulocytedevelopmental stages using two new markers. Dev. Comp.Immunol. 31, 296.
Ratajczak, J., Machalinski, B., Majka, M., Kijowski, J., Marlicz, W.,Rozmyslowicz, T., Ostrowski, M., Ratajczak, M.Z., 1999.Evidence that human haematopoietic stem cells (HSC) do notreside within the CD34+KIT-cell population. Ann. Transplant. 4,22.
Sachs, D.H., 1994. The pig as a potential xenograft donor. Vet.Immunol. Immunopathol. 43, 185.
Sitnicka, E., Buza-Vidas, N., Larsson, S., Nygren, J.M., Liuba, K.,Jacobsen, S.E., 2003. Human CD34+ hematopoietic stem cellscapable of multilineage engrafting NOD/SCID mice express flt3:distinct flt3 and c-kit expresión and response patterns on mouseand candidate human hematopoietic stem cells. Blood 102, 881.
Summerfield, A., Horn, M.P., Lozano, G., Carrasco, C.P., Atze, K.,McCullough, K., 2003. C-kit positive porcine bone marrowprogenitor cells identified and enriched using recombinant stemcell factor. J. Immunol. Methods 280, 113.
Sun, L., Lee, J., Fine, H.A., 2004. Neuronally expressed stem cellfactor induces neural stem cell migration to areas of brain injury.J. Clin. Invest. 113, 1364.
Vodicka, P., Smetana, K., Dvorankova, B., Emerick, T., Xu, Y.Z.,Ourednik, J., Ourednik, V., Motlik, J., 2005. The miniature pig asan animal in biomedical research. Ann. N.Y. Acad. Sci. 1049, 161.
Yee, N.S., Hsiau, C.W., Serve, H., Vosseller, K., Besmer, P., 1994.Mechanism of down-regulation of c-kit receptor. Roles of receptortyrosine kinase, phosphatidylinositol 3′-kinase, and protein kinaseC. J. Biol. Chem. 269, 31991.