characterisation of α3β1 and αvβ3 integrin n-oligosaccharides in metastatic melanoma wm9 and...
TRANSCRIPT

Biochimica et Biophysica Acta 1780 (2008) 1421–1431
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbagen
Characterisation of α3β1 and αvβ3 integrin N-oligosaccharides in metastaticmelanoma WM9 and WM239 cell lines
Marcelina E. Kremser a,⁎, Małgorzata Przybyło a, Dorota Hoja-Łukowicz a, Ewa Pocheć a, Angela Amoresano b,Andrea Carpentieri b, Monika Bubka a, Anna Lityńska a
a Department of Glycoconjugate Biochemistry, Institute of Zoology, Jagiellonian University, ul. Ingardena 6, 30-060 Krakow, Polandb Dipartimento di Chimica e Biochimica, Complesso Universitario Monte S'Angelo, Napoli, Italy
⁎ Corresponding author. Fax: +48 012 634 37 16.E-mail address: [email protected] (M.E. Kremser
0304-4165/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.bbagen.2008.07.011
a b s t r a c t
a r t i c l e i n f oArticle history:
It is well documented that Received 23 April 2008Received in revised form 11 July 2008Accepted 22 July 2008Available online 29 July 2008Keywords:α3β1 integrinαVβ3 integrinN-oligosaccharideMALDI MSLectin
glycan synthesis is altered in some pathological processes, including cancer. Themost frequently observed alterations during tumourigenesis are extensive expression of β1,6-branchedcomplex type N-glycans, the presence of poly-N-acetyllactosamine structures, and high sialylation of cellsurface glycoproteins. This study investigated two integrins, α3β1 and αvβ3, whose expression is closelyrelated to cancer progression. Their oligosaccharide structures in two metastatic melanoma cell lines(WM9, WM239) were analysed with the use of matrix-assisted laser desorption ionisation massspectrometry. Both examined integrins possessed heavily sialylated and fucosylated glycans, with β1,6-branches and short polylactosamine chains. In WM9 cells, α3β1 integrin was more variously glycosylatedthan αvβ3; in WM239 cells the situation was the reverse. Functional studies (wound healing and ELISAintegrin binding assays) revealed that the N-oligosaccharide component of the tested integrins influencedmelanoma cell migration on vitronectin and α3β1 integrin binding to laminin-5. Additionally, morevariously glycosylated integrins exerted a stronger influence on these parameters. To the best of ourknowledge, this is the first report concerning structural characterisation of αvβ3 integrin glycans inmelanoma or in any cancer cells.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Over half of the proteins present in living organisms areglycosylated [1], and many of them are expressed in cell mem-branes. For all glycoproteins this modification is a major determi-nant of their activity, folding and/or stability [2]. It is welldocumented in the literature that glycan synthesis is altered insome pathological processes, including cancer [3–5]; the mostfrequently observed alterations during tumourigenesis are extensiveexpression of β1,6-branched tri- and tetraantennary complex typeN-glycans [6–9], the presence of poly-N-acetyllactosamine struc-tures, and high sialylation of cell surface glycoproteins. In the case ofthe cell surface receptors, the changed glycosylation profile mayaffect their presence in the cell membrane [10]. Carbohydrates havebeen shown to interact with proteins during recognition events, soany changes in the structure of the sugar component may influencethe glycoprotein's ligand binding ability [11]. Hence the commonobservation is that altered glycosylation modifies migration and
).
l rights reserved.
adhesive properties and supports the metastatic potential of cancercells.
Integrins, a large family of cell membrane receptors, are involved inimportant processes such as cell–cell and cell–extracellular matrix(ECM) adhesion. They are built of two subunits:α and β. Both subunitsare glycosylated and possess multiple potential N-glycosylation sites[12]. It is reported that an increase of the metastatic potential oftumour cells is accompanied by changes in the expression level ofseveral adhesive molecules, including integrins. Altered integrinexpression is associated with tissue invasion and metastasis in manytypes of cancer, but there is no universal pattern of integrin expressionin cancer cells [13,14]. In melanoma, α3β1 and αvβ3 integrins arerecognised as specific markers of tumour progression, and αvβ3 inparticular is used as a marker to distinguish the radial growth phase(RGP) from the vertical growth phase (VGP) [15–17].
The aim of this study was to analyse glycan pools of α3β1 and αvβ3
integrins purified from two human melanoma cell lines of differentorigin: WM9 from lymph node metastasis, and WM239 from skinmetastasis. Moreover, the biological relevance of these findings wasexamined by ELISA integrin binding and wound healing assays todetermine whether the oligosaccharides attached to these integrinsmodified their interaction with primary ligands — laminin-5 andvitronectin. To the best of our knowledge this is the first report of a
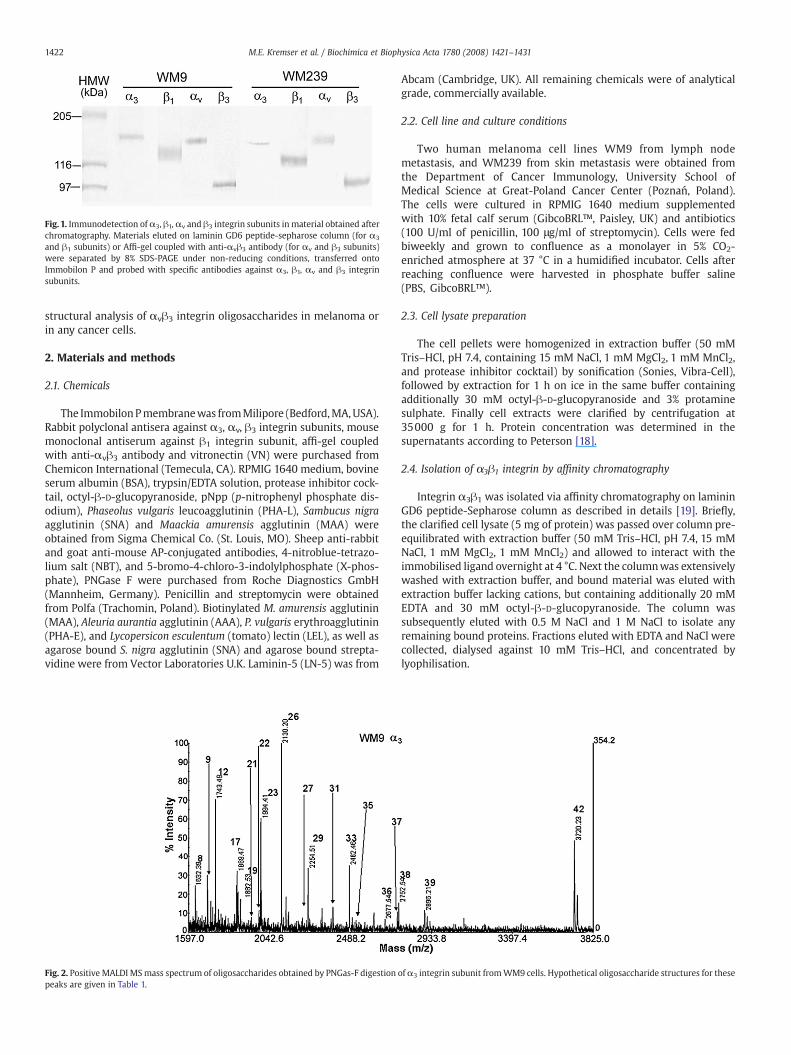
Fig. 1. Immunodetection of α3, β1,αv and β3 integrin subunits in material obtained afterchromatography. Materials eluted on laminin GD6 peptide-sepharose column (for α3
and β1 subunits) or Affi-gel coupled with anti-αvβ3 antibody (for αv and β3 subunits)were separated by 8% SDS-PAGE under non-reducing conditions, transferred ontoImmobilon P and probed with specific antibodies against α3, β1, αv and β3 integrinsubunits.
1422 M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
structural analysis of αvβ3 integrin oligosaccharides in melanoma orin any cancer cells.
2. Materials and methods
2.1. Chemicals
The ImmobilonPmembranewas fromMilipore (Bedford,MA,USA).Rabbit polyclonal antisera against α3, αv, β3 integrin subunits, mousemonoclonal antiserum against β1 integrin subunit, affi-gel coupledwith anti-αvβ3 antibody and vitronectin (VN) were purchased fromChemicon International (Temecula, CA). RPMIG 1640 medium, bovineserum albumin (BSA), trypsin/EDTA solution, protease inhibitor cock-tail, octyl-β-D-glucopyranoside, pNpp (p-nitrophenyl phosphate dis-odium), Phaseolus vulgaris leucoagglutinin (PHA-L), Sambucus nigraagglutinin (SNA) and Maackia amurensis agglutinin (MAA) wereobtained from Sigma Chemical Co. (St. Louis, MO). Sheep anti-rabbitand goat anti-mouse AP-conjugated antibodies, 4-nitroblue-tetrazo-lium salt (NBT), and 5-bromo-4-chloro-3-indolylphosphate (X-phos-phate), PNGase F were purchased from Roche Diagnostics GmbH(Mannheim, Germany). Penicillin and streptomycin were obtainedfrom Polfa (Trachomin, Poland). Biotinylated M. amurensis agglutinin(MAA), Aleuria aurantia agglutinin (AAA), P. vulgaris erythroagglutinin(PHA-E), and Lycopersicon esculentum (tomato) lectin (LEL), as well asagarose bound S. nigra agglutinin (SNA) and agarose bound strepta-vidine were from Vector Laboratories U.K. Laminin-5 (LN-5) was from
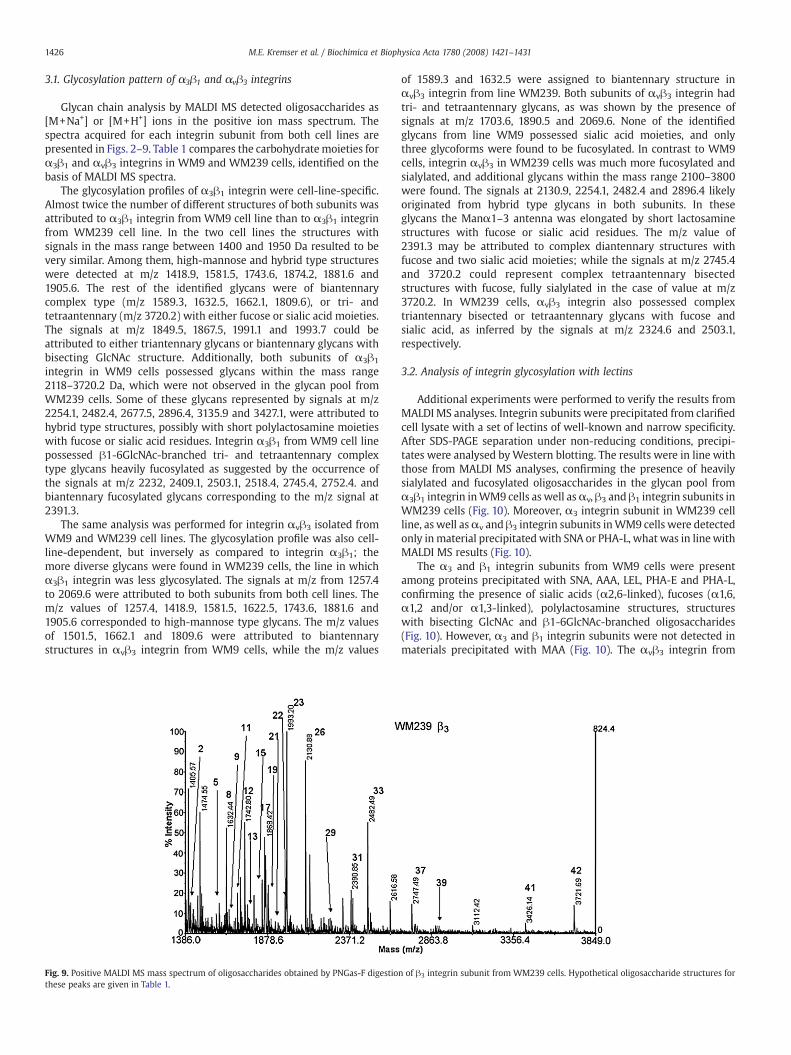
Fig. 2. Positive MALDI MSmass spectrum of oligosaccharides obtained by PNGas-F digestionpeaks are given in Table 1.
Abcam (Cambridge, UK). All remaining chemicals were of analyticalgrade, commercially available.
2.2. Cell line and culture conditions
Two human melanoma cell lines WM9 from lymph nodemetastasis, and WM239 from skin metastasis were obtained fromthe Department of Cancer Immunology, University School ofMedical Science at Great-Poland Cancer Center (Poznań, Poland).The cells were cultured in RPMIG 1640 medium supplementedwith 10% fetal calf serum (GibcoBRL™, Paisley, UK) and antibiotics(100 U/ml of penicillin, 100 μg/ml of streptomycin). Cells were fedbiweekly and grown to confluence as a monolayer in 5% CO2-enriched atmosphere at 37 °C in a humidified incubator. Cells afterreaching confluence were harvested in phosphate buffer saline(PBS, GibcoBRL™).
2.3. Cell lysate preparation
The cell pellets were homogenized in extraction buffer (50 mMTris–HCl, pH 7.4, containing 15 mM NaCl, 1 mM MgCl2, 1 mM MnCl2,and protease inhibitor cocktail) by sonification (Sonies, Vibra-Cell),followed by extraction for 1 h on ice in the same buffer containingadditionally 30 mM octyl-β-D-glucopyranoside and 3% protaminesulphate. Finally cell extracts were clarified by centrifugation at35000 g for 1 h. Protein concentration was determined in thesupernatants according to Peterson [18].
2.4. Isolation of α3β1 integrin by affinity chromatography
Integrin α3β1 was isolated via affinity chromatography on lamininGD6 peptide-Sepharose column as described in details [19]. Briefly,the clarified cell lysate (5 mg of protein) was passed over column pre-equilibrated with extraction buffer (50 mM Tris–HCl, pH 7.4, 15 mMNaCl, 1 mM MgCl2, 1 mM MnCl2) and allowed to interact with theimmobilised ligand overnight at 4 °C. Next the columnwas extensivelywashed with extraction buffer, and bound material was eluted withextraction buffer lacking cations, but containing additionally 20 mMEDTA and 30 mM octyl-β-D-glucopyranoside. The column wassubsequently eluted with 0.5 M NaCl and 1 M NaCl to isolate anyremaining bound proteins. Fractions eluted with EDTA and NaCl werecollected, dialysed against 10 mM Tris–HCl, and concentrated bylyophilisation.
ofα3 integrin subunit fromWM9 cells. Hypothetical oligosaccharide structures for these

Fig. 3. Positive MALDI MSmass spectrum of oligosaccharides obtained by PNGas-F digestion of β1 integrin subunit fromWM9 cells. Hypothetical oligosaccharide structures for thesepeaks are given in Table 1.
1423M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
2.5. Purification of αvβ3 integrin by immunoaffinity chromatography
The αvβ3 integrin receptor was isolated with a use of Affi-gelcoupled with anti-αvβ3 antibody (Chemicon) as described in details[20]. Briefly, the gel was per-equilibrated with PBS, pH 7.6, containing0.1% NP-40, 1 mM CaCl2. The clarified cell lysate (300 μg of protein)was incubated with the gel overnight at 4 °C. Next the gel was washedwith two washing buffers: PBS, pH 7.6, containing 0.1% NP-40, 1 mMCaCl2; and 10 mM NaHOAc, pH 4.5, containing 0.1% NP-40, 2 mMCaCl2. The bound material was eluted with a use of 10 mM NaHOAc,pH 3.0 containing 0.1% NP-40, 2 mM CaCl2. Collected factions wereneutralized in 3 M Tris–HCl, pH 8.8, and then dialysed against 5 mMNaH2PO4x2H2O, pH 7.6, and concentrated by lyophilisation.
2.6. Precipitation with lectins
The clarified cell lysate of WM9 (300 μg of protein) and WM239(500 μg of protein) cells were incubated with 10 μl of biotinylated M.amurensis agglutinin (MAA), 8 μl of biotinylated A. aurantia agglutinin
Fig. 4. Positive MALDI MSmass spectrum of oligosaccharides obtained by PNGas-F digestion opeaks are given in Table 1.
(AAA), 6 μl of biotinylated P. vulgaris erythroagglutinin (PHA-E), 30 μlof agarose bound P. vulgaris leucoagglutinin (PHA-L), 6 μl ofbiotinylated L. esculentum (tomato) lectin (LEL), and 16.5 μl agarosebound S. nigra agglutinin (SNA), overnight at 4 °C on an orbital rotator.Then the samples (with the exception of these precipitated with SNAand PHA-L) were mixed with streptavidine bound agarose andincubated for another 2 h at 4 °C on an orbital rotator. After thatprecipitated materials were washed three times with buffer 10 mMHEPES, pH 7.5, and 150 mM NaCl (containing additionally 1 mM CaCl2and 1 mM MgCl2 in case of precipitation with PHA-E and PHA-L), andonce in PBS. The precipitated proteins were eluted by boiling theagarose bound complexes for 10min in SDS-PAGE sample buffer in theabsence of reductant.
2.7. SDS-PAGE and immunodetection of integrin subunits
Eluted from affinity chromatography column or antibody-boundmaterials as well as the samples after precipitation with lectins, wereseparated by 8% SDS-PAGE under non-reducing condition according to
f αv integrin subunit fromWM9 cells. Hypothetical oligosaccharide structures for these

Fig. 5. Positive MALDI MSmass spectrum of oligosaccharides obtained by PNGas-F digestion of β3 integrin subunit fromWM9 cells. Hypothetical oligosaccharide structures for thesepeaks are given in Table 1.
1424 M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
Laemmli [21]. Next the proteins were electrotransferred to anImmobilon P membrane for 18 h at constant amperage 0.1 A withcooling. The blots were blocked in TBS/Tween (0.02 M Tris–HCl, pH7.6, containing 150 mM NaCl and 0.1% Tween 20), with 1% BSA.Afterwards, membranes were sequentially incubated with specificantibodies diluted in TBS/Tween with 1% BSA (1:2000 for antibodyanti-α3, anti-αv, anti-β3, anti-β1 integrin subunits) for 1 h. After triplewash with TBS/Tween, blots were incubated with alkaline phospha-tase coupled sheep anti-rabbit or goat anti-mouse Ig (1:2000 workingdilution in TBS/Tween with 1% BSA) for 1 h. The subunits werelocalised with the use of 4-nitroblue-tetrazolium salt/5-bromo-4-chloro-3-indolylophosphate solution (NBT/X-phosphate).
2.8. Glycan chain analysis
Protein bands corresponding to α3, αv, β1 and β3 subunits wereexcised from Immobilon P sheet. Further analysis (protein alkylation,in situ digestion, sugar extraction and microcolumn clean-up ofsugars) was carried out as previously described [22].
Matrix-assisted laser desorption ionisation (MALDI) mass spectrawere recorded by using a Voyager DE-STR MALDI-TOF mass spectro-
Fig. 6. Positive MALDI MS mass spectrum of oligosaccharides obtained by PNGas-F digestionthese peaks are given in Table 1.
meter (Applied Biosystem) equipped with the delayed extraction andoperating in the reflectron mode. The MALDI matrix was prepared bydissolving 25 mg of DHB in 1 ml of acetonitrile/0.2% trifluoroaceticacid (70:30 v/v). Typically, 1 μl of matrix was applied to the metallicsample plate and 1 μl of analyte was then added. Acceleration andreflector voltages were set up as follow: target voltage at 20 kV, firstgrid at 95% of target voltage, delayed extraction at 600 ns to obtainthe best signal-to-noise ratios and the best possible isotopicresolution with multipoint external calibration using peptide mixturepurchased from Applied Biosystems. Each spectrum represents thesum of 1500 laser pulses from randomly chosen spots per sampleposition. Raw data were analysed using the computer softwareprovided by the manufacturers and are reported as monoisotopicmasses.
2.9. Modified ELISA integrin binding assay
The ELISA assay was performed as described previously [23].Briefly, the material purified on laminin GD6 peptide-Sepharosecolumn (PNGase F-treated or non-treated) was added into microtiterwells coated with LN-5 (2.8 μg/ml). The ELISA was developed with
of α3 integrin subunit from WM239 cells. Hypothetical oligosaccharide structures for

Fig. 7. Positive MALDI MS mass spectrum of oligosaccharides obtained by PNGas-F digestion of β1 integrin subunit from WM239 cells. Hypothetical oligosaccharide structures forthese peaks are given in Table 1.
1425M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
10 mM pNpp and the absorbance was measured at 405 nm after 18 hof incubation. Each value was determined in triplicate in threeindependent experiments.
2.10. Wound healing assay
In vitro scrape wound healing assays on VN-coated (0.7 μg/ml)surface were performed as described previously [23]. In someexperiments, wound healing in culture medium containing PHA-L(25 μg/ml), SNA (50 μg/ml) or MAA (25 μg/ml) was examined. Theapplied doses of lectins had no effect on the viability or growth rate ofthe tested cells as demonstrated by trypan blue exclusion and 3[4,5-dimethylthiazol-2-yl]-2,5diphenyltetrazolium bromide (MTT) tests(data not shown). Migration of cells into wounded areas was observedin an inverted microscope and photographed. The average extent ofwound closure was quantified bymultiple measurements of thewidthof the wound space for each of these cases. Values are expressed asmean ± standard deviation of three separate experiments.
Fig. 8. Positive MALDI MS mass spectrum of oligosaccharides obtained by PNGas-F digestionthese peaks are given in Table 1.
2.11. Statistics
The significance of the differences between mean values wascomputed using Duncan's new multiple range test and P values lowerthan 0.05 were considered significant.
3. Results
In order to purify the integrins from human melanoma WM9 andWM239 cell lines, affinity chromatography on GD-6 peptide (forα3β1)and immunoaffinity chromatography with immobilised antibodiesagainst αvβ3 integrin were performed. The presence of isolatedproteins in the collected material was confirmed by SDS-PAGE undernon-reducing conditions followed by Western blotting with specificantibodies (Fig. 1). Themolecular weights of integrin subunits inWM9andWM239, respectively, were as follows: α3 subunit—154.8 kDa and152.9 kDa; β1 subunit—128.9 kDa and 125.6 kDa; αv subunit—148.8 kDa and 148.9 kDa; and β3 subunit—99.5 kDa and 101.4 kDa.
of αv integrin subunit from WM239 cells. Hypothetical oligosaccharide structures for
1426 M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
3.1. Glycosylation pattern of α3β1 and αvβ3 integrins
Glycan chain analysis by MALDI MS detected oligosaccharides as[M+Na+] or [M+H+] ions in the positive ion mass spectrum. Thespectra acquired for each integrin subunit from both cell lines arepresented in Figs. 2–9. Table 1 compares the carbohydratemoieties forα3β1 and αvβ3 integrins in WM9 and WM239 cells, identified on thebasis of MALDI MS spectra.
The glycosylation profiles of α3β1 integrin were cell-line-specific.Almost twice the number of different structures of both subunits wasattributed to α3β1 integrin from WM9 cell line than to α3β1 integrinfrom WM239 cell line. In the two cell lines the structures withsignals in the mass range between 1400 and 1950 Da resulted to bevery similar. Among them, high-mannose and hybrid type structureswere detected at m/z 1418.9, 1581.5, 1743.6, 1874.2, 1881.6 and1905.6. The rest of the identified glycans were of biantennarycomplex type (m/z 1589.3, 1632.5, 1662.1, 1809.6), or tri- andtetraantennary (m/z 3720.2) with either fucose or sialic acid moieties.The signals at m/z 1849.5, 1867.5, 1991.1 and 1993.7 could beattributed to either triantennary glycans or biantennary glycans withbisecting GlcNAc structure. Additionally, both subunits of α3β1
integrin in WM9 cells possessed glycans within the mass range2118–3720.2 Da, which were not observed in the glycan pool fromWM239 cells. Some of these glycans represented by signals at m/z2254.1, 2482.4, 2677.5, 2896.4, 3135.9 and 3427.1, were attributed tohybrid type structures, possibly with short polylactosamine moietieswith fucose or sialic acid residues. Integrin α3β1 from WM9 cell linepossessed β1-6GlcNAc-branched tri- and tetraantennary complextype glycans heavily fucosylated as suggested by the occurrence ofthe signals at m/z 2232, 2409.1, 2503.1, 2518.4, 2745.4, 2752.4. andbiantennary fucosylated glycans corresponding to the m/z signal at2391.3.
The same analysis was performed for integrin αvβ3 isolated fromWM9 and WM239 cell lines. The glycosylation profile was also cell-line-dependent, but inversely as compared to integrin α3β1; themore diverse glycans were found in WM239 cells, the line in whichα3β1 integrin was less glycosylated. The signals at m/z from 1257.4to 2069.6 were attributed to both subunits from both cell lines. Them/z values of 1257.4, 1418.9, 1581.5, 1622.5, 1743.6, 1881.6 and1905.6 corresponded to high-mannose type glycans. The m/z valuesof 1501.5, 1662.1 and 1809.6 were attributed to biantennarystructures in αvβ3 integrin from WM9 cells, while the m/z values
Fig. 9. Positive MALDI MS mass spectrum of oligosaccharides obtained by PNGas-F digestiothese peaks are given in Table 1.
of 1589.3 and 1632.5 were assigned to biantennary structure inαvβ3 integrin from line WM239. Both subunits of αvβ3 integrin hadtri- and tetraantennary glycans, as was shown by the presence ofsignals at m/z 1703.6, 1890.5 and 2069.6. None of the identifiedglycans from line WM9 possessed sialic acid moieties, and onlythree glycoforms were found to be fucosylated. In contrast to WM9cells, integrin αvβ3 in WM239 cells was much more fucosylated andsialylated, and additional glycans within the mass range 2100–3800were found. The signals at 2130.9, 2254.1, 2482.4 and 2896.4 likelyoriginated from hybrid type glycans in both subunits. In theseglycans the Manα1–3 antenna was elongated by short lactosaminestructures with fucose or sialic acid residues. The m/z value of2391.3 may be attributed to complex diantennary structures withfucose and two sialic acid moieties; while the signals at m/z 2745.4and 3720.2 could represent complex tetraantennary bisectedstructures with fucose, fully sialylated in the case of value at m/z3720.2. In WM239 cells, αvβ3 integrin also possessed complextriantennary bisected or tetraantennary glycans with fucose andsialic acid, as inferred by the signals at m/z 2324.6 and 2503.1,respectively.
3.2. Analysis of integrin glycosylation with lectins
Additional experiments were performed to verify the results fromMALDI MS analyses. Integrin subunits were precipitated from clarifiedcell lysate with a set of lectins of well-known and narrow specificity.After SDS-PAGE separation under non-reducing conditions, precipi-tates were analysed byWestern blotting. The results were in line withthose from MALDI MS analyses, confirming the presence of heavilysialylated and fucosylated oligosaccharides in the glycan pool fromα3β1 integrin inWM9 cells as well asαv, β3 and β1 integrin subunits inWM239 cells (Fig. 10). Moreover, α3 integrin subunit in WM239 cellline, as well asαv and β3 integrin subunits inWM9 cells were detectedonly inmaterial precipitated with SNA or PHA-L, what was in linewithMALDI MS results (Fig. 10).
The α3 and β1 integrin subunits from WM9 cells were presentamong proteins precipitated with SNA, AAA, LEL, PHA-E and PHA-L,confirming the presence of sialic acids (α2,6-linked), fucoses (α1,6,α1,2 and/or α1,3-linked), polylactosamine structures, structureswith bisecting GlcNAc and β1-6GlcNAc-branched oligosaccharides(Fig. 10). However, α3 and β1 integrin subunits were not detected inmaterials precipitated with MAA (Fig. 10). The αvβ3 integrin from
n of β3 integrin subunit from WM239 cells. Hypothetical oligosaccharide structures for
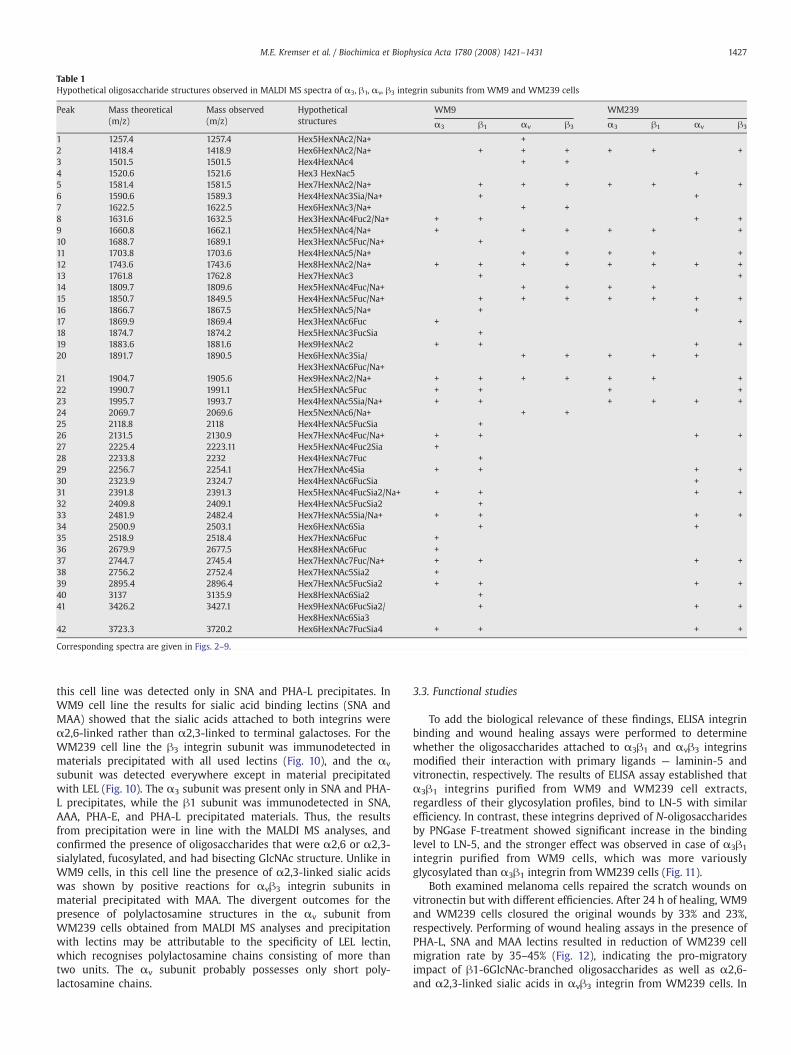
Table 1Hypothetical oligosaccharide structures observed in MALDI MS spectra of α3, β1, αv, β3 integrin subunits from WM9 and WM239 cells
Peak Mass theoretical(m/z)
Mass observed(m/z)
Hypotheticalstructures
WM9 WM239
α3 β1 αv β3 α3 β1 αv β3
1 1257.4 1257.4 Hex5HexNAc2/Na+ +2 1418.4 1418.9 Hex6HexNAc2/Na+ + + + + + +3 1501.5 1501.5 Hex4HexNAc4 + +4 1520.6 1521.6 Hex3 HexNac5 +5 1581.4 1581.5 Hex7HexNAc2/Na+ + + + + + +6 1590.6 1589.3 Hex4HexNAc3Sia/Na+ + +7 1622.5 1622.5 Hex6HexNAc3/Na+ + +8 1631.6 1632.5 Hex3HexNAc4Fuc2/Na+ + + + +9 1660.8 1662.1 Hex5HexNAc4/Na+ + + + + + +10 1688.7 1689.1 Hex3HexNAc5Fuc/Na+ +11 1703.8 1703.6 Hex4HexNAc5/Na+ + + + + +12 1743.6 1743.6 Hex8HexNAc2/Na+ + + + + + + + +13 1761.8 1762.8 Hex7HexNAc3 + +14 1809.7 1809.6 Hex5HexNAc4Fuc/Na+ + + + +15 1850.7 1849.5 Hex4HexNAc5Fuc/Na+ + + + + + + +16 1866.7 1867.5 Hex5HexNAc5/Na+ + +17 1869.9 1869.4 Hex3HexNAc6Fuc + +18 1874.7 1874.2 Hex5HexNAc3FucSia +19 1883.6 1881.6 Hex9HexNAc2 + + + +20 1891.7 1890.5 Hex6HexNAc3Sia/
Hex3HexNAc6Fuc/Na++ + + + +
21 1904.7 1905.6 Hex9HexNAc2/Na+ + + + + + + +22 1990.7 1991.1 Hex5HexNAc5Fuc + + + +23 1995.7 1993.7 Hex4HexNAc5Sia/Na+ + + + + + +24 2069.7 2069.6 Hex5NexNAc6/Na+ + +25 2118.8 2118 Hex4HexNAc5FucSia +26 2131.5 2130.9 Hex7HexNAc4Fuc/Na+ + + + +27 2225.4 2223.11 Hex5HexNAc4Fuc2Sia +28 2233.8 2232 Hex4HexNAc7Fuc +29 2256.7 2254.1 Hex7HexNAc4Sia + + + +30 2323.9 2324.7 Hex4HexNAc6FucSia +31 2391.8 2391.3 Hex5HexNAc4FucSia2/Na+ + + + +32 2409.8 2409.1 Hex4HexNAc5FucSia2 +33 2481.9 2482.4 Hex7HexNAc5Sia/Na+ + + + +34 2500.9 2503.1 Hex6HexNAc6Sia + +35 2518.9 2518.4 Hex7HexNAc6Fuc +36 2679.9 2677.5 Hex8HexNAc6Fuc +37 2744.7 2745.4 Hex7HexNAc7Fuc/Na+ + + + +38 2756.2 2752.4 Hex7HexNAc5Sia2 +39 2895.4 2896.4 Hex7HexNAc5FucSia2 + + + +40 3137 3135.9 Hex8HexNAc6Sia2 +41 3426.2 3427.1 Hex9HexNAc6FucSia2/
Hex8HexNAc6Sia3+ + +
42 3723.3 3720.2 Hex6HexNAc7FucSia4 + + + +
Corresponding spectra are given in Figs. 2–9.
1427M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
this cell line was detected only in SNA and PHA-L precipitates. InWM9 cell line the results for sialic acid binding lectins (SNA andMAA) showed that the sialic acids attached to both integrins wereα2,6-linked rather than α2,3-linked to terminal galactoses. For theWM239 cell line the β3 integrin subunit was immunodetected inmaterials precipitated with all used lectins (Fig. 10), and the αv
subunit was detected everywhere except in material precipitatedwith LEL (Fig. 10). The α3 subunit was present only in SNA and PHA-L precipitates, while the β1 subunit was immunodetected in SNA,AAA, PHA-E, and PHA-L precipitated materials. Thus, the resultsfrom precipitation were in line with the MALDI MS analyses, andconfirmed the presence of oligosaccharides that were α2,6 or α2,3-sialylated, fucosylated, and had bisecting GlcNAc structure. Unlike inWM9 cells, in this cell line the presence of α2,3-linked sialic acidswas shown by positive reactions for αvβ3 integrin subunits inmaterial precipitated with MAA. The divergent outcomes for thepresence of polylactosamine structures in the αv subunit fromWM239 cells obtained from MALDI MS analyses and precipitationwith lectins may be attributable to the specificity of LEL lectin,which recognises polylactosamine chains consisting of more thantwo units. The αv subunit probably possesses only short poly-lactosamine chains.
3.3. Functional studies
To add the biological relevance of these findings, ELISA integrinbinding and wound healing assays were performed to determinewhether the oligosaccharides attached to α3β1 and αvβ3 integrinsmodified their interaction with primary ligands — laminin-5 andvitronectin, respectively. The results of ELISA assay established thatα3β1 integrins purified from WM9 and WM239 cell extracts,regardless of their glycosylation profiles, bind to LN-5 with similarefficiency. In contrast, these integrins deprived of N-oligosaccharidesby PNGase F-treatment showed significant increase in the bindinglevel to LN-5, and the stronger effect was observed in case of α3β1
integrin purified from WM9 cells, which was more variouslyglycosylated than α3β1 integrin from WM239 cells (Fig. 11).
Both examined melanoma cells repaired the scratch wounds onvitronectin but with different efficiencies. After 24 h of healing, WM9and WM239 cells closured the original wounds by 33% and 23%,respectively. Performing of wound healing assays in the presence ofPHA-L, SNA and MAA lectins resulted in reduction of WM239 cellmigration rate by 35–45% (Fig. 12), indicating the pro-migratoryimpact of β1-6GlcNAc-branched oligosaccharides as well as α2,6-and α2,3-linked sialic acids in αvβ3 integrin from WM239 cells. In

Fig. 10. Immunodetection of α3, β1, αv and β3 integrin subunits in materials precipitated with the following lectins: Sambucus nigra agglutinin (SNA), Maackia amurensis agglutinin(MAA), Aleuria aurantia agglutinin (AAA), Phaseolus vulgaris erythroagglutinin (PHA-E), Lycopersicon esculentum (tomato) lectin (LEL) and P. vulgaris leucoagglutinin (PHA-L).Materials obtained after lectin precipitation were separated by 8% SDS-PAGE under non-reducing conditions, transferred onto Immobilon P and probed with specific antibodiesagainst α3, β1, αv and β3 integrin subunits.
Fig. 11. Ligand binding specificity of purified PNGase F-treated α3β1 integrin fromWM9and WM239 cells. Integrin binding to laminin-5 (LN-5) was quantitated using modifiedELISA test as described in Materials and methods. ⁎Pb0.05.
1428 M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
contrast, although the presence of PHA-L also diminished themigration rate of WM9 cells into the scratch wounds by 35%, thepresence of SNA and MAA lectins had no influence on this process(Fig. 12).
4. Discussion
In the present study we showed that α3β1 and αvβ3 integrinspurified from two humanmelanoma cell lines derived from particularmetastasis sites differ in their glycosylation profiles. Both subunits ofα3β1 integrin in WM9 cells originating from lymph node metastasisshowed more diverse glycan types than did WM239 cells originatingfrom skin metastasis. In the case of αvβ3 integrin it was just thereverse: the glycan pool from WM239 cells was much more diversethan in WM9 cells. Nevertheless, both integrins showed the presenceof tumour-associated glycans regardless of the differences in glyco-sylation profiles between these integrins in the two cell lines.
The α3β1 and αvβ3 integrin subunits from both metastatic celllines displayed tri- and tetraantennary oligosaccharides, but thequantity of these glycans was cell-line-dependent. Integrin α3β1
possessed more β1,6-branched structures in WM9 cells than inWM239 cells, unlikeαvβ3 integrinwhich hadmore of these glycans inWM239 cells. In our previous studies we used tandem mass spectro-metry analysis to identify proteins bearing β1,6-branched oligosac-charides in primary WM35 and three metastatic melanoma cell lines(WM9, WM239, A375) [24,25]. In metastatic cell lines, among theidentified proteins the largest group consisted of integrin subunitsincluding α3β1 and αvβ3, but the α3 subunit was shown not be asubstrate for GnT-V (N-acetylglucosaminyltransferase V) in primarymelanoma WM35 cells. These results are in good agreement with apreviously performed structural analysis of α3β1 integrin glycans inWM35 and A375 cells [26] and with the present analysis of α3β1
integrin glycans in WM9 and WM239 cells. They are also in line withevidence that the presence of β1,6-branched oligosaccharide struc-tures influences tumour cell adhesion and seems critical to theestablishment of metastasis [27–30].
Increased β1,6-branching leads to increased numbers of Galβ1-4GlcNAc structures to which sialic acid can be attached, resulting inthe stronger cell surface sialylation that is common in varioustumours. Due to its terminal location and negative charge, sialic acidcould be involved in the regulation of binding to the ECM. However,the correlation between the cell surface sialylation and metastatic
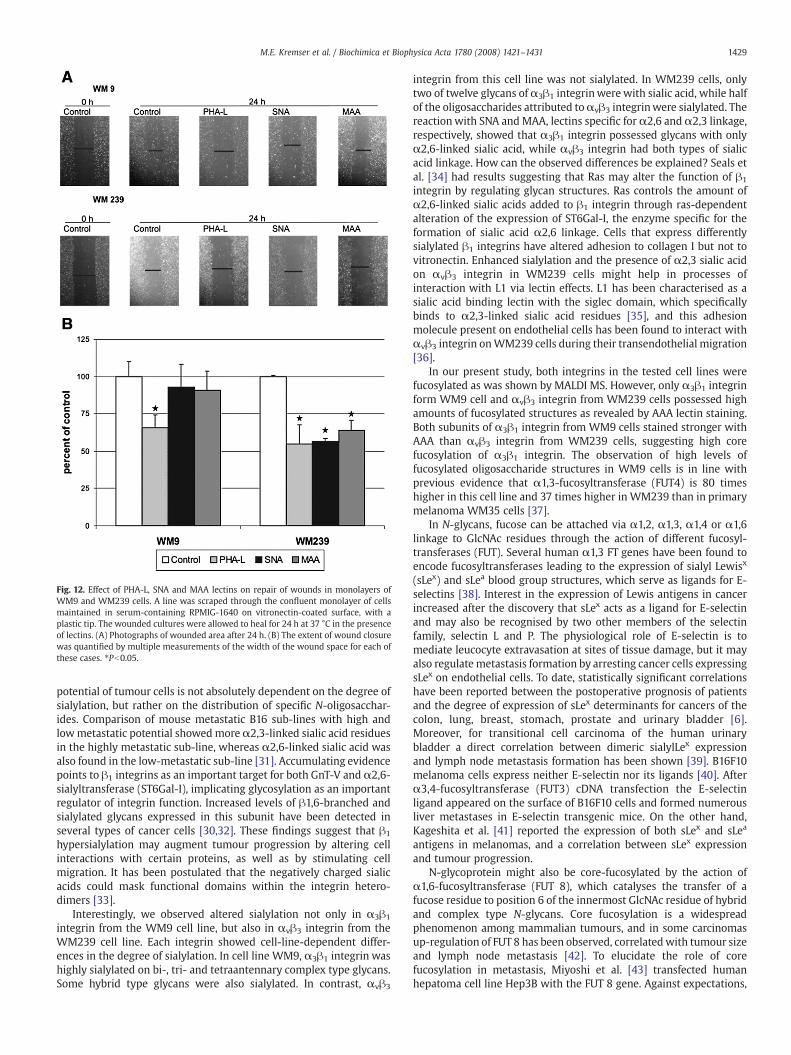
Fig. 12. Effect of PHA-L, SNA and MAA lectins on repair of wounds in monolayers ofWM9 and WM239 cells. A line was scraped through the confluent monolayer of cellsmaintained in serum-containing RPMIG-1640 on vitronectin-coated surface, with aplastic tip. The wounded cultures were allowed to heal for 24 h at 37 °C in the presenceof lectins. (A) Photographs of wounded area after 24 h. (B) The extent of wound closurewas quantified by multiple measurements of the width of the wound space for each ofthese cases. ⁎Pb0.05.
1429M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
potential of tumour cells is not absolutely dependent on the degree ofsialylation, but rather on the distribution of specific N-oligosacchar-ides. Comparison of mouse metastatic B16 sub-lines with high andlowmetastatic potential showedmoreα2,3-linked sialic acid residuesin the highly metastatic sub-line, whereas α2,6-linked sialic acid wasalso found in the low-metastatic sub-line [31]. Accumulating evidencepoints to β1 integrins as an important target for both GnT-V and α2,6-sialyltransferase (ST6Gal-I), implicating glycosylation as an importantregulator of integrin function. Increased levels of β1,6-branched andsialylated glycans expressed in this subunit have been detected inseveral types of cancer cells [30,32]. These findings suggest that β1
hypersialylation may augment tumour progression by altering cellinteractions with certain proteins, as well as by stimulating cellmigration. It has been postulated that the negatively charged sialicacids could mask functional domains within the integrin hetero-dimers [33].
Interestingly, we observed altered sialylation not only in α3β1
integrin from the WM9 cell line, but also in αvβ3 integrin from theWM239 cell line. Each integrin showed cell-line-dependent differ-ences in the degree of sialylation. In cell line WM9, α3β1 integrin washighly sialylated on bi-, tri- and tetraantennary complex type glycans.Some hybrid type glycans were also sialylated. In contrast, αvβ3
integrin from this cell line was not sialylated. In WM239 cells, onlytwo of twelve glycans of α3β1 integrinwerewith sialic acid, while halfof the oligosaccharides attributed toαvβ3 integrinwere sialylated. Thereactionwith SNA andMAA, lectins specific for α2,6 and α2,3 linkage,respectively, showed that α3β1 integrin possessed glycans with onlyα2,6-linked sialic acid, while αvβ3 integrin had both types of sialicacid linkage. How can the observed differences be explained? Seals etal. [34] had results suggesting that Ras may alter the function of β1
integrin by regulating glycan structures. Ras controls the amount ofα2,6-linked sialic acids added to β1 integrin through ras-dependentalteration of the expression of ST6Gal-I, the enzyme specific for theformation of sialic acid α2,6 linkage. Cells that express differentlysialylated β1 integrins have altered adhesion to collagen I but not tovitronectin. Enhanced sialylation and the presence of α2,3 sialic acidon αvβ3 integrin in WM239 cells might help in processes ofinteraction with L1 via lectin effects. L1 has been characterised as asialic acid binding lectin with the siglec domain, which specificallybinds to α2,3-linked sialic acid residues [35], and this adhesionmolecule present on endothelial cells has been found to interact withαvβ3 integrin onWM239 cells during their transendothelial migration[36].
In our present study, both integrins in the tested cell lines werefucosylated as was shown by MALDI MS. However, only α3β1 integrinform WM9 cell and αvβ3 integrin from WM239 cells possessed highamounts of fucosylated structures as revealed by AAA lectin staining.Both subunits of α3β1 integrin from WM9 cells stained stronger withAAA than αvβ3 integrin from WM239 cells, suggesting high corefucosylation of α3β1 integrin. The observation of high levels offucosylated oligosaccharide structures in WM9 cells is in line withprevious evidence that α1,3-fucosyltransferase (FUT4) is 80 timeshigher in this cell line and 37 times higher in WM239 than in primarymelanoma WM35 cells [37].
In N-glycans, fucose can be attached via α1,2, α1,3, α1,4 or α1,6linkage to GlcNAc residues through the action of different fucosyl-transferases (FUT). Several human α1,3 FT genes have been found toencode fucosyltransferases leading to the expression of sialyl Lewisx
(sLex) and sLea blood group structures, which serve as ligands for E-selectins [38]. Interest in the expression of Lewis antigens in cancerincreased after the discovery that sLex acts as a ligand for E-selectinand may also be recognised by two other members of the selectinfamily, selectin L and P. The physiological role of E-selectin is tomediate leucocyte extravasation at sites of tissue damage, but it mayalso regulate metastasis formation by arresting cancer cells expressingsLex on endothelial cells. To date, statistically significant correlationshave been reported between the postoperative prognosis of patientsand the degree of expression of sLex determinants for cancers of thecolon, lung, breast, stomach, prostate and urinary bladder [6].Moreover, for transitional cell carcinoma of the human urinarybladder a direct correlation between dimeric sialylLex expressionand lymph node metastasis formation has been shown [39]. B16F10melanoma cells express neither E-selectin nor its ligands [40]. Afterα3,4-fucosyltransferase (FUT3) cDNA transfection the E-selectinligand appeared on the surface of B16F10 cells and formed numerousliver metastases in E-selectin transgenic mice. On the other hand,Kageshita et al. [41] reported the expression of both sLex and sLea
antigens in melanomas, and a correlation between sLex expressionand tumour progression.
N-glycoprotein might also be core-fucosylated by the action ofα1,6-fucosyltransferase (FUT 8), which catalyses the transfer of afucose residue to position 6 of the innermost GlcNAc residue of hybridand complex type N-glycans. Core fucosylation is a widespreadphenomenon among mammalian tumours, and in some carcinomasup-regulation of FUT 8 has been observed, correlatedwith tumour sizeand lymph node metastasis [42]. To elucidate the role of corefucosylation in metastasis, Miyoshi et al. [43] transfected humanhepatoma cell line Hep3B with the FUT 8 gene. Against expectations,

1430 M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
tumour formation in the liver was suppressed after injection oftransfected cells to athymic mice. In the same study, attachment ofFUT 8 transfectants to a fibronectin-coated dish was decreased versusthe controls because α5β1 integrin was more strongly core-fucosy-lated in these cells. Two-dimensional electrophoresis followed byreactionwith LCA (Lens culinaris agglutinin), a lectin which recognisesα1,6-linked fucose, showed fucosylation of several glycoproteins.Among themwasα5β1 integrin but alsoαvβ3 integrin, suggesting thatit could also be a substrate for FUT 8. Integrins α2β1 and α3β1 wereexpressed in these cells at extremely low levels [43]. In view of thosefindings, Zhao et al. [42] investigated the effect of core fucosylation onα3β1 integrin in embryonic fibroblasts from FUT 8−/− mice (FUT 8knockout mice). In FUT 8−/− cells, core fucosylation of α3β1 integrinwas totally abolished, and this was associated with deficientmigrationon laminin-5. Moreover, integrin-mediated cell signalling wasreduced in FUT 8−/− cells. These results suggested that corefucosylation is essential to the functioning of α3β1 integrin. Suchdata strongly suggest that both integrins analysed in our present studymay be substrates for FUT 8, and that core fucosylation has an impacton the functioning of α3β1 integrin. Why core fucosylation modifiesα3β1 integrin functioning is not known, but some possible mechan-isms have been proposed [42]. FUT 8 may affect the cross-talkbetween growth factor receptors and integrin, or may cause a changein the conformation of integrin. It is reasonable to suppose that corefucosylation contributes to conformation stability and normal activityof α3β1 integrin with its ligand.
In this study the signals at m/z 2130.9 and 2677.5 as well as 3135.9and 3427.1 corresponded to short GlcNAc-Gal repeats in both subunitsof integrin α3β1 isolated from WM9 cells. This outcome is inagreement with previous results for α3β1 integrin from humancolon carcinoma [44] and human metastatic melanoma cells [26]. Incontrast, in WM239 cells the signals at m/z 2254.1, 2482.4, 2896.4 and3427.1, corresponding to N-acetyl-lactosamine (lacNAc), were attrib-uted to both subunits of integrin αvβ3. The presence of lacNAc orpolylacNAc moieties is another cancer-associated modification ofglycans. PolylacNAc moieties consist of repeating Galβ1-4GlcNAcβ1-3disaccharide units, which are preferentially added to β1,6GlcNAc-linked antennae to complex type N-glycans [45] by repeating action oftwo transferases, β1,3GlcNAcT and β1,4GalT. β1,4GalT activity iswidely expressed and appears to be present in excess in most tissues.Hence the rate-limiting step in polylacNAc biosynthesis has beensuggested to be either β1,3GlcNAcT- or GnT-V-catalysed reactions[46]. PolylacNAc glycosylation is added preferentially to selectproteins, that is, LAMPs [29,45], but also integrin subunits[23,26,44]. Krishman et al. [29] have shown that altered melanomacell surface glycosylation due to the presence of LAMP withpolylacNAc moieties mediates organ-specific metastasis via lectinreceptors on the lung vascular endothelium.
It is well established that the presence of β1,6GlcNAc-branchedglycanwith a polylactosamine extensionmay play a role in controllingthe residence time of receptors on the cell surface and in regulatingtheir activity and stability [4,47]. Polylactosamine chains can bendback toward the protein backbone and interact with it [32]. Forintegrins this interaction may affect protein activity, because some ofthem are present in the cell membrane in bent conformation, and a“switchblade” model for their affinity switching is proposed. It is afeature of αvβ3 integrin activity, and therefore the presence ofpolylactosamine chains may keep this receptor in the extended,active state [48]. In the present study, integrinαvβ3 in theWM239 cellline possessed glycans with lacNAc moieties which might modulatethe conformation of this integrin. In functional studies we confirmedthat the interaction ofα3β1 integrinwith its primary ligand— LN-5 (inELISA test) wasmodified by the presence of N-linked oligosaccharides.Moreover the migration properties of WM239 cells on vitronectinwere modulated by the presence of sialic acids and β1-6 branchedcomplex type oligosaccharides as reveal the wound healing assay,
while WM9 cell migration was promoted only by β1-6 branched N-oligosaccharides.
The most important observation in our present study was the cell-specific glycosylation of α3β1 and αvβ3 integrins in metastaticmelanoma WM9 (lymph node) and WM239 (skin) cells. Thisobservation is very interesting in view of the importance of theseintegrins in melanoma progression [49–52]. The potential role ofaltered glycosylation in modulating integrin function has long beenlargely ignored despite the observation that cells express variantintegrin glycoforms when undergoing significant phenotypic changes[32]. Now the glycans of cell surface receptors such as integrinreceptors are thought to be a common means of adhesion andrecognition between cells and between cells and ECM [4,12]. Earlierstudies mainly concerning β1 integrins have shown a causal relation-ship between variant glycosylation and altered integrin activity [32].
Among the integrins that undergo changes in glycosylationprofiles, α3β1 integrin has been intensively investigated[19,23,26,44]. Almost nothing is known about glycosylation of αvβ3
integrin, despite the observation of αvβ3 integrin's higher molecularmass in GnT-V transfectant Mv1Lu mink lung epithelial cells, whichsuggested that this integrin could be a substrate for GnT-V [53]. On theother hand, it is well documented that αvβ3 integrin supportsadhesive and invasive tumour cell functioning, critical duringmelanoma progression. This integrin is expressed when melanomacells switch from radial growth phase (RGP) to vertical growth phase(VGP), and thus is correlated with cancer progression [16,17].
This work on α3β1 and αvβ3 integrin N-oligosaccharides in WM9and WM239 cells showed remarkable differences between the N-glycan pools of these integrins in a particular cell line, and theoccurrence of various N-oligosaccharides in a particular integrin ofWM9 andWM239 cells. In our study the characterisation of the glycanpool of αvβ3 integrin in melanoma cells showed that it is a carrier ofcancer-related oligosaccharides. Since the examined cell lines comefrom different sites of metastasis, our observations might suggest thatglycosylation of a particular protein can be modified under theinfluence of surrounding tissue. However, it is more likely that alteredoligosaccharides of the cancer cell take part in forming the metastasis.The origin of this phenotype and its biological significance are as yetunclear and will require considerable further study.
Acknowledgments
This work was supported by the Polish State Committee forScientific Research (PB/0939/P05/2004/26) and by Jagiellonian Uni-versity (BW/12a/IZ/2003).
References
[1] R. Apweiler, H. Hermjakob, N. Sharon, On the frequency of protein glycosylationdeduced from analysis of the SWISS-PROT database, Biochim. Biophys. Acta 1473(1999) 4–8.
[2] C.J. Bosques, S.M. Tschampel, R.J. Woods, B. Imperiali, Effects of glycosylation onpeptide conformation: a synergistic experimental and computational study, J. Am.Chem. Soc. 126 (2004) 8421–8425.
[3] J.W. Dennis, M. Granovsky, C.E. Warren, Glycoprotein glycosylation and cancerprogression, Biochim. Biophys. Acta 1473 (1999) 21–34.
[4] J.W. Dennis, M. Granovsky, C.E. Warren, Protein glycosylation in development anddisease, Bioessays 21 (1999) 412–421.
[5] J. Gu, N. Taniguchi, Regulation of integrin functions by N-glycans, Glycoconj. J. 21(2004) 9–15.
[6] F. Dall'Olio, Protein glycosylation in cancer biology: an overview, J. Clin. Pathol.Mol. Pathol. 49 (1996) M126–M135.
[7] H.B. Guo, I. Lee, M. Kamar, S.K. Akiyama, M. Pierce, Aberrant N-glycosylation of β1
integrin causes reduced α5β1 integrin clustering and stimulates cell migration,Cancer Res. 62 (2002) 6837–6845.
[8] S. Hakomori, Glycosylation defining cancer malignancy: newwine in an old bottle,Proc. Natl. Acad. Sci. U. S. A. 99 (2002) 10231–10233.
[9] P. Laidler, A. Lityńska, D. Hoja-Łukowicz, M. Łabędź, M. Przybyło, D. Ciołczyk-Wierzbicka, E. Pocheć, E. Trębacz, E. Kremser, Characterization of glycosylationand adherent properties of melanoma cell lines, Cancer Immun. Immunother. 55(2006) 112–118.

1431M.E. Kremser et al. / Biochimica et Biophysica Acta 1780 (2008) 1421–1431
[10] I. Lee, M.A. Skinner, H.B. Guo, A. Sujan, M. Pierce, Expression of the vacuolar H+-ATPase 16-kDa subunit result in the Triton X-100-insoluble aggregation of β1
integrin and reduction of its cell surface expression, J. Biol. Chem. 279 (2004)53007–53014.
[11] A. Lityńska, M. Przybyło, E. Pocheć, P. Laidler, Adhesion properties of humanbladder cell lines with extracellular matrix components: the role of integrins andglycosylation, Acta Biochim. Pol. 49 (2002) 643–650.
[12] R. Chammas, S.S. Veiga, L.R. Travassos, R.R. Brentani, Functionally distinct roles forglycosylation of α and β integrin chains in cell–matrix interactions, Proc. Natl.Acad. Sci. U. S. A. 90 (1993) 1795–1799.
[13] G.J. Mizejewski, Role of integrins in cancer: survey of expression patterns, Proc.Soc. Exp. Biol. Med. 222 (1999) 124–138.
[14] B. Wehrle-Haller, B. Imhof, Integrin-dependent pathologies, J. Pathol. 200 (2003)481–487.
[15] A. Melchiori, R. Mortarini, S. Carlone, P.C. Morchisio, A. Anichini, D.M. Noonan, A.Albini, The α3β1 integrin is involved in melanoma cell migration and invasion,Exp. Cell Res. 219 (1995) 233–242.
[16] A. Koistinen, M. Ahonen, V.M. Kähäri, J. Heino,αv integrin promotes in vitro and invivo survival of cells in metastatic melanoma, Int. J. Cancer 112 (2004) 61–70.
[17] K.D. Cowden Dahl, S.E. Robertson, V.M. Weaver, M.C. Simson, Hypoxia-induciblefactor regulates αvβ3 integrin cell surface expression, Mol. Biol. Cell 16 (2005)1901–1912.
[18] G.L. Peterson, A symplification of the protein assaymethod of Lowry et al. which ismore generally applicable, Anal. Biochem. 83 (1977) 346–356.
[19] A. Lityńska, E. Pocheć, D. Hoja-Łukowicz, E. Kremser, P. Laidler, A. Amoresano, C.Monti, The structure of the oligosaccharides of α3β1 integrin from human ureterepithelium (HCV29) cell line, Acta Biochim. Pol. 49 (2002) 491–500.
[20] J.W. Smith, D.A. Cheresh, The Arg-Gly-Asp binding domain of the vitronectinreceptor, J. Biol. Chem. 263 (1988) 18726–18731.
[21] U.K. Laemmli, Cleavage of structural proteins during the assembly of the head ofbacteriophage T4, Nature 227 (1970) 690-685.
[22] D. Hoja-Łukowicz, D. Ciołczyk, J. Bergquist, A. Lityńska, P. Laidler, High-mannose-type oligosaccharides from human placental arylsulfatase A are core fucosylatedas confirmed by MALDI MS, Glycobiology 10 (2000) 551–557.
[23] E. Pocheć, A. Lityńska, M. Bubka, A. Amoresano, A. Casbarra, Characterization ofthe oligosaccharide component of α3β1 integrin from human bladder carcinomacell line T24 and its role in adhesion and migration, Eur. J. Cell. Biol. 85 (2006)47–57.
[24] D. Ochwat, D. Hoja-Łukowicz, A. Lityńska, N-glycoproteins bearing β1-6 branchedoligosaccharides from the A375 human melanoma cell line analysed by tandemmass spectrometry, Melanoma Res. 14 (2004) 479–485.
[25] M. Przybyło, D. Martuszewska, E. Pocheć, D. Hoja-Łukowicz, A. Lityńska,Identification of proteins bearing β1-6 branched N-glycans in human melanomacell lines from different progression stages by tandem mass spectrometryanalysis, Biochim. Biophis. Acta 1770 (2007) 1427–1435.
[26] E. Pocheć, A. Lityńska, A. Amoresano, A. Casbarra, Glycosylation profile ofintegrin α3β1 changes with melanoma progression, Biochim. Biophys. Acta 7(2003) 1–3.
[27] H. Yamamoto, J. Swoger, S. Greene, T. Saito, J. Hurh, C. Sweeley, J. Leestma, E.Mkrdichian, L. Cerullo, A. Nishikawa, Y. Ihara, N. Taniguchi, J.R. Moskal, β1,6-N-acetylglucosamine-bearing N-glycans in human gliomas: implications for a role inregulating invasivity, Cancer Res. 60 (2000) 134–142.
[28] T. Handerson, J.M. Pawelek, β1,6-branched oligosaccharides and coarse vesicles: acommon, pervasive phenotype in melanoma and other human cancers, CancerRes. 63 (2003) 5363–5369.
[29] V. Krishnan, S.M. Bane, P.D. Kawle, K.N. Naresh, R.D. Kalraiya, Altered melanomacell surface glycosylation mediates organ specific adhesion and metastasis vialectin receptors on the lung vascular endothelium, Clin. Exp. Metastasis 22 (2005)11–24.
[30] B.V.V.G. Reddy, R.D. Kalraiya, Sialilated β1,6 branched N-oligosaccharidesmodulated adhesion, chemotaxis and motility of melanoma cells: effect oninvasion and spontaneous metastasis properties, Biochim. Biophys. Acta 1760(2006) 1393–1402.
[31] A. Passaniti, G.W. Hart, Cell sialilation and tumor metastasis: metastatic potentialof B16 melanoma variants correlates with their relative numbers of specificpenultimate oligosaccharide structures, J. Biol. Chem. 263 (1988) 7591–7603.
[32] S.L. Bellis, Variant glycosylation: an underappreciated regulatory mechanism forβ1 integrins, Biochim. Biophys. Acta 1663 (2004) 52–60.
[33] E.C. Seales, G.A. Jurado, B.A. Brunson, J.K. Wakefield, A.R. Frost, S.L. Bellis,Hypersialilation of β1 integrins, observed in colon adenocarcinoma, maycontribute to cancer progression by up-regulating cell motility, Cancer Res. 65(2005) 4645–4652.
[34] E.C. Seales, G.A. Jurado, A. Singhal, S.L. Bellis, Ras oncogene directs expression of adifferentially sialylated, functionally altered β1 integrin, Oncogene 22 (2003)7137–7145.
[35] A. Thies, M. Schachner, I. Moll, J. Berger, H.J. Schulze, G. Brunner, U. Schumacher,Overexpression of the cell adhesion molecule L1 is associated with metastasis incutaneous malignant melanoma, Eur. J. Cancer 38 (2002) 1708–1716.
[36] E.B. Voura, R.A. Ramjeesingh, A.M.P. Montgomery, C.H. Siu, Involvement ofintegrin αvβ3 and cell adhesion molecule L1 in transendothelial migration ofmelanoma cells, Mol. Biol. Cell. 12 (2001) 2699–2710.
[37] D. Ciołczyk-Wierzbicka, M. Bodzioch, D. Gil, D. Żmudzińska, A. Dembińska-Kieć, P.Laidler, Expression of fucosylated glycans in humanmelanoma cell lines mediatedcell adhesion, Med. Chem. 3 (2007) 418–424.
[38] K. Yago, K. Zenita, H. Ginya, M. Sawada, K. Ohmori, M. Okuma, R. Kannagi, J.B.Lowe, Expression of α-(1,3)-fucosyltransferases which synthetize sialyl Lex andsialyl Lea, the carbohydrate ligands for E- and P-selectins, in human malignant celllines, Cancer Res. 53 (1993) 5559–5565.
[39] T. Matsusako, H. Muramatsu, T. Shirahama, T. Muramatsu, Y. Ohi, Expression of acarbohydrate signal, sialyl-dimeric Lex antigen, is associated with metastaticpotential of transitional cell carcinoma of the human urinary bladder, Biochem.Biophys. Res. Commun. 181 (1991) 1218–1222.
[40] L. Biancone, M. Araki, K. Araki, P. Vassalli, I. Stamenkovic, Redirection of tumormetastasis by expression of E-selectin in vivo, J. Exp. Med. 183 (1996) 581–587.
[41] T. Kageshita, S. Hirai, T. Kimura, N. Hanai, S. Ohta, T. Ono, Association betweensialyl Lewisa expression and tumor progression in melanoma, Cancer Res. 55(1995) 748–751.
[42] Y. Zhao, S. Itoh, X. Wang, T. Isaji, E. Miyoshi, Y. Kariya, K. Miyazaki, N. Kawasaki, N.Taniguchi, J. Gu, Deletion of core fucosylation on α3β1 integrin down-regulates itsfunctions, J. Biol. Chem. 281 (2006) 38343–38350.
[43] E. Miyoshi, K. Noda, Y. Yamaguchi, S. Inoue, Y. Ikeda, W. Wang, J.H. Ko, N. Uozumi,W. Li, N. Taniguchi, The α1-6-fucosyltransferase gene and its biologicalsignificance, Biochim. Biophys. Acta 1473 (1999) 9–20.
[44] N.L. Prokopishyn, W. Puzon-McLaughlin, Y. Takada, S. Laferté, Integrin α3β1
expressed by human colon cancer cells is a major carrier of oncodevelopmentalcarbohydrate epitopes, J. Cell Biochem. 72 (1999) 189–209.
[45] I.R. Nabi, J.W. Dennis, The extend of polylactosamine glycosylation of MDCKLAMP-2 is determined by its Golgi residence time, Glycobiology 8 (1998) 947–953.
[46] S. Yousefi, E. Higgins, Z. Daoling, A. Pollex-Krüger, O. Hindsgaul, J.W. Dennis,Increased UDP-GclNAc:Gal β1-3GalNAc-R (GlcNAc to GalNAc) β1-6-N-acetylglucosaminyltransferase activity inmetastatic murine tumor cell lines. Controlof polylactosamine synthesis, J. Biol. Chem. 266 (1991) 1772–1782.
[47] H.B. Guo, M. Randolph, M. Pierce, Inhibition of a specific N-glycosylation activityresults in attenuation of breast carcinoma cell invasiveness-related phenotypes:inhibition of epidermal growth factor-induced dephosphorylation of focaladhesion kinase, J. Biol. Chem. 282 (2007) 22150–22162.
[48] B.D. Adair, J.P. Xiong, C. Maddock, S.L. Goodman, M.A. Arnaout, M. Yeager, Threedimensional EM structure of the ectodomain of integrin αvβ3 in a complex withfibronectin, J. Cell Biol. 168 (2005) 1109–1118.
[49] P.G. Natali, M.R. Nicotra, A. Bartolazzi, R. Cavaliere, A. Bigotti, Integrin expressionin cutaneous malignant melanoma: association of the α3β1 heterodimer withtumor, Int. J. Cancer 54 (1993) 68–72.
[50] T. Tsuji, Y. Kawada, M. Kai-Murozono, S. Komatsu, S.A. Han, K. Takeuchi, H.Mizushima, K. Miyazaki, T. Irimura, Regulation of melanoma cell migration andinvasion by laminin-5 and α3β1 integrin (VLA-3), Clin. Exp. Metastasis 19 (2002)127–134.
[51] T. Tsuji, Physiological and pathological roles of α3β1 integrin, 200 (2004) 115–132.[52] H. Takatsuki, S. Komatsu, R. Sano, Y. Takada, T. Tsuji, Adhesion of gastric carcinoma
cells to peritoneum mediated by α3β1, Cancer Res. 64 (2004) 6065-6050.[53] M. Demetriou, I.R. Nabi, M. Coppolino, S. Dedhar, J.W. Dennis, Reduced contact-
inhibition and substratum adhesion in epithelial cells expressing GlcNAc-transferase V, J. Cell Biol. 130 (1995) 383–392.