cell contact-dependent pmn hla-dr and cd69 membrane expression induced by autologous...
TRANSCRIPT

Inflammation, Vol. 26, No. 4, August 2002 ( 2002)
0360-3997/ 02/ 0800-0143/ 0 2002 Plenum Publishing Corporation
143
Cell Contact-Dependent PMN HLA-DR and CD69Membrane Expression Induced by AutologousMono-Lymphocytes and Cell Lines
A. Vella,1,3 S. Sartoris,1 L. Bambara,2 R. Ortolani,1 A. Carletto,2 D. Biasi,2 E. Stefani,1 andG. Tridente1
Abstract—Polymorphonuclear granulocytes (PMN) are commonly considered short-lived cellsplaying an efficient role in primary host defense via phagocytosis and release of cytotoxic com-pounds and inflammatory cytokines.
Purified PMN do not express HLA-DR and CD69 molecules on cell surface, but they can beinduced to do so by co-culture with peripheral blood derived mono-lymphocytes. De novo cell-surface expression of HLA-DR was also induced in PMN by co-culture with cell lines of lymphoidphenotype, but not with cell lines of myeloid phenotype. CD69 expression was not induced by co-culture with any of the cell lines used in the present study. In addition, we have observed inductionof HLA-DR surface expression on PMN by culture in presence of culture supernatant of one ofthe cell lines of lymphoid origin, RPMI-8866.
Quantitative analysis of HLA-DR and CD69 expression in stimulated PMN allowed us to dividePMN donors in two main groups, one with low expression and the other with high expression ofthe two molecules.
HLA-DR surface expression was not altered by treatment with CHX and BFA, and RT-PCRanalysis of total RNA from resting and stimulated PMN with RPMI-8866 supernatant did notdetect the presence of any specific HLA-DR and CIITA transcript. Flow-cytometry and fluores-cence microscopy analysis of resting PMN revealed the presence of HLA-DR molecules localizedin intracellular vesicular-tubular structures.
These data show that a reservoir of HLA-DR molecules is stored in the cytoplasm of humanresting PMN and can be released to reach cell surface by a mobilization mechanism induced bycell surface interactions with selected cell types and sometimes with molecules released in culturesupernatants.
KEY WORDS: HLA-DR, CD69, immune presentation, chronic, acute, inflammation
1 Department of Pathology, Immunology Section, University of Verona,Italy.
2 Department of Clinical and Experimental Medicine, University ofVerona, Italy.
3 To whom correspondence should be addressed at the Department ofPathology, Immunology Section University of Verona, Piazzale Anto-nio Scuro, 10 37100 Verona, Italy. Fax: 0039 045 8202459; E-mail:[email protected].
INTRODUCTION
MHC class II (MHC-II) molecules present antigenic pep-tides to cell receptor of CD4+ T helper cells, trigger-ing the cascade of events leading to both cellular andhumoral immune effector functions (1).
The regulation of MHC-II expression occurs pri-

Vella, Sartoris, Bambara, Ortolani, Carletto, Biasi, Stefani, and Tridente144
marily at the transcriptional level and the Class II Trans-Activator (CIITA) factor has been shown to play a piv-otal role in both constitutive and inducible expression(2). During human granulopoiesis, HLA-DR moleculesare expressed on the cell surface of PMN precursors inthe bone marrow, and HLA-DR-specific transcripts canbe detected (3). On the contrary, mature resting PMN donot express HLA-DR molecules on cell surface, but theycan be induced to do so in vitro by prolonged incubationwith IFN-g, IL-3 and GM-CSF (4).
CD69, also known as very early activation antigen,is a member of the natural killer (NK) cell gene complexfamily of signal transducing receptors. CD69 is as a type IItransmembrane glycoprotein with a C-type lectin bindingdomain in the extracellular portion of the molecule. CD69expression is induced in vitro on cells of most hematopoi-etic lineages, including T and B lymphocytes, NK cells,murine macrophages and eosinophils, while it is con-stitutively expressed on human monocytes, thymocytes,platelets and epidermal Langerhans cells. Although a spe-cific ligand for CD69 has not been identified yet, its widecellular distribution and the induction of intracellular sig-nals upon crosslinking suggests a role for the receptor inthe biology of hematopoietic cells (5). CD69 is expressedon cell surface of bone marrow-derived myeloid precur-sors, while it is located in the cytoplasm in mature, resting,neutrophils (5). However, intracellular CD69 moleculesare rapidly mobilized to the cell surface upon activationby PMA or fMLP. This translocation is independent fromprotein synthesis, being not inhibited by cycloheximide:CD69 molecules are probably stored in a trans-Golgistructure since their expression is not affected by brefeldinA, a drug that blocks molecular trafficking from ER toGolgi vesicles (6).
The present report describes a new model of invitro induction of HLA-DR and CD69 expression oncell surface of peripheral blood-derived PMN. Evidenceis provided that HLA-DR expression can be inducedin absence of de novo transcription and translation ofboth CIITA and HLA-DR. It is tempting to speculatethat, similarly to CD69, an intracellular pool of HLA-DR molecules is present in mature, resting PMN. Thesemolecules can be rapidly targeted to the cell surface inresponse to cell contact stimuli.
MATERIALS AND METHODS
PMN and mono-lymphocytes were isolated fromperipheral blood by density centrifugation using discon-
tinuous Percoll gradients (Pharmacia Upsala Sweden)from 30 healthy donors followed by hypotonic lysis oferythrocytes, as described (7). The human cell lines usedin this work were: RPMI-8866, a B cell line derivedfrom a chronic myelogenous leukemia; RAJI, a B cellline derived from a human B cell lymphoma; H9, a lym-phoblastic cell line derived from a human acute T celllymphoma; JURKAT, a lymphoblastic cell line derivedfrom a human acute T cell leukemia; U937, a mono-cytic cell line derived from a human histiocytic lym-phoma; THP-1, a monocytic cell line derived from ahuman acute monocytic leukemia; HL-60, a human acutepromyelocytic leukemia. PMN were co-cultured at ratiosof 1 : 0.1, 1 : 0.5, 1 : 1, 1 : 2, with each cell line andat 1 : 1 ratio with autologous mono-lymphocytes. Cellswere maintained in RPMI 1640 supplemented with 10%FBS, 2 mM L-glutamine at 378C at 5% CO2.
To inhibit protein synthesis and intracellular vesic-ular transport to Golgi, cycloheximide (CHX) andBrefeldin A (BFA) (SIGMA Italia) were added to PMNat a concentration of 1–10 mg/ ml for 30 min before co-culture.
Monoclonal antibodies specific for human CD45,HLA-DR and CD69, directly conjugated with fluoro-cromes, and isotype control irrelevant antibodies werepurchased from Immunotech (Italy). The D1-12 anti-HLA-DR monoclonal antibody was kindly provided byProf. R. S. Accolla (University of Insubria, Varese,Italy) (8). PMN freshly purified or co-cultured withmono-lymphocytes or cell lines were analyzed by directimmunofluorescence at the EPICS XL-MCL CYTOME-TER (COULTER). PMN were gated by Forward andSide Scatter (FS, SS) or by CD45/ SS. Values for surfacemolecule expression were obtained as the percentage ofpositive cells and mean fluorescence intensity (MFI).
The presence of intracellular HLA-DR moleculesin resting PMN was investigated by Flow-Cytometryand microscope analysis. PMN fixed in 0,4% paraben-zoquinone (PBQ) (Sigma) were permeabilized by incu-bation in 0,74% n-octyl-b-D-glucopyranoside (OG)(Sigma) (9) and then stained with FITC-D1-12 or controlantibody for one hour at 48C. Total RNA was extractedfrom PMN resting or stimulated with RPMI-8866 super-natant according to established procedures (10). The pres-ence of CIITA, DR-a and DR-b specific transcripts wasassessed by RT-PCR using the GeneAmp RT-PCR kit(Applyed Biosystem, Italy), according to the manufac-turer’s recommendations. Primers specific for CIITA,DR-a, DR-b, b-actin and RT-PCR conditions were asdescribed by Chang et al. (11).
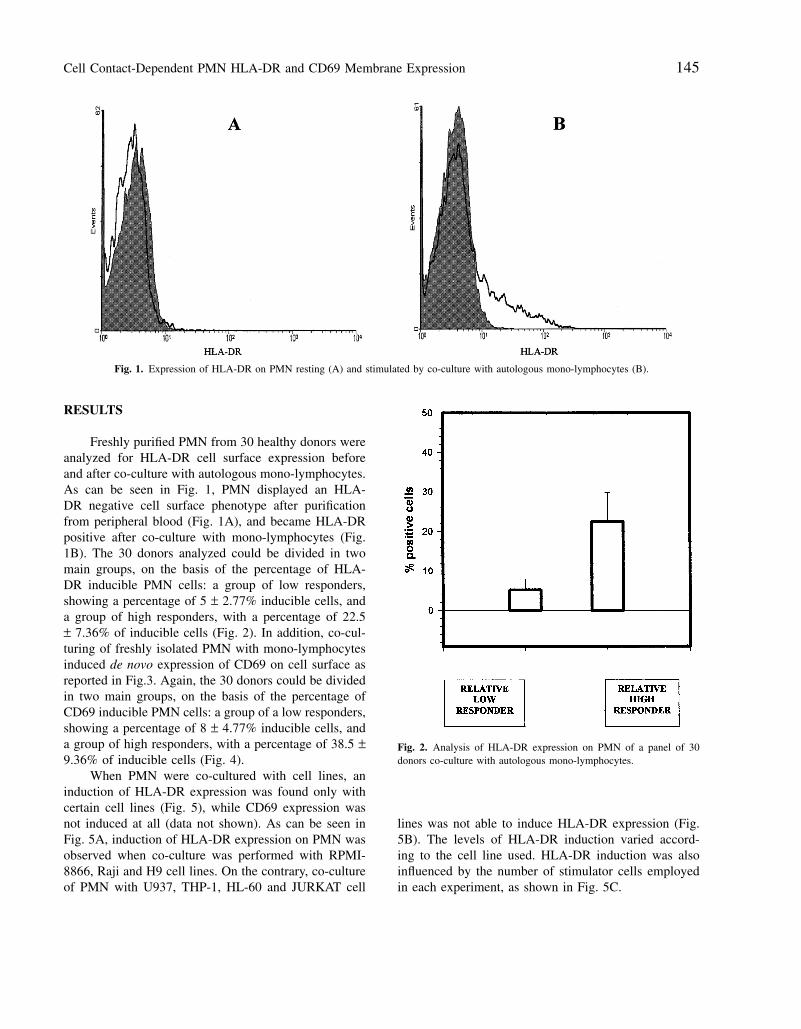
Cell Contact-Dependent PMN HLA-DR and CD69 Membrane Expression 145
Fig. 1. Expression of HLA-DR on PMN resting (A) and stimulated by co-culture with autologous mono-lymphocytes (B).
RESULTS
Freshly purified PMN from 30 healthy donors wereanalyzed for HLA-DR cell surface expression beforeand after co-culture with autologous mono-lymphocytes.As can be seen in Fig. 1, PMN displayed an HLA-DR negative cell surface phenotype after purificationfrom peripheral blood (Fig. 1A), and became HLA-DRpositive after co-culture with mono-lymphocytes (Fig.1B). The 30 donors analyzed could be divided in twomain groups, on the basis of the percentage of HLA-DR inducible PMN cells: a group of low responders,showing a percentage of 5 ± 2.77% inducible cells, anda group of high responders, with a percentage of 22.5± 7.36% of inducible cells (Fig. 2). In addition, co-cul-turing of freshly isolated PMN with mono-lymphocytesinduced de novo expression of CD69 on cell surface asreported in Fig.3. Again, the 30 donors could be dividedin two main groups, on the basis of the percentage ofCD69 inducible PMN cells: a group of a low responders,showing a percentage of 8 ± 4.77% inducible cells, anda group of high responders, with a percentage of 38.5 ±9.36% of inducible cells (Fig. 4).
When PMN were co-cultured with cell lines, aninduction of HLA-DR expression was found only withcertain cell lines (Fig. 5), while CD69 expression wasnot induced at all (data not shown). As can be seen inFig. 5A, induction of HLA-DR expression on PMN wasobserved when co-culture was performed with RPMI-8866, Raji and H9 cell lines. On the contrary, co-cultureof PMN with U937, THP-1, HL-60 and JURKAT cell
Fig. 2. Analysis of HLA-DR expression on PMN of a panel of 30donors co-culture with autologous mono-lymphocytes.
lines was not able to induce HLA-DR expression (Fig.5B). The levels of HLA-DR induction varied accord-ing to the cell line used. HLA-DR induction was alsoinfluenced by the number of stimulator cells employedin each experiment, as shown in Fig. 5C.
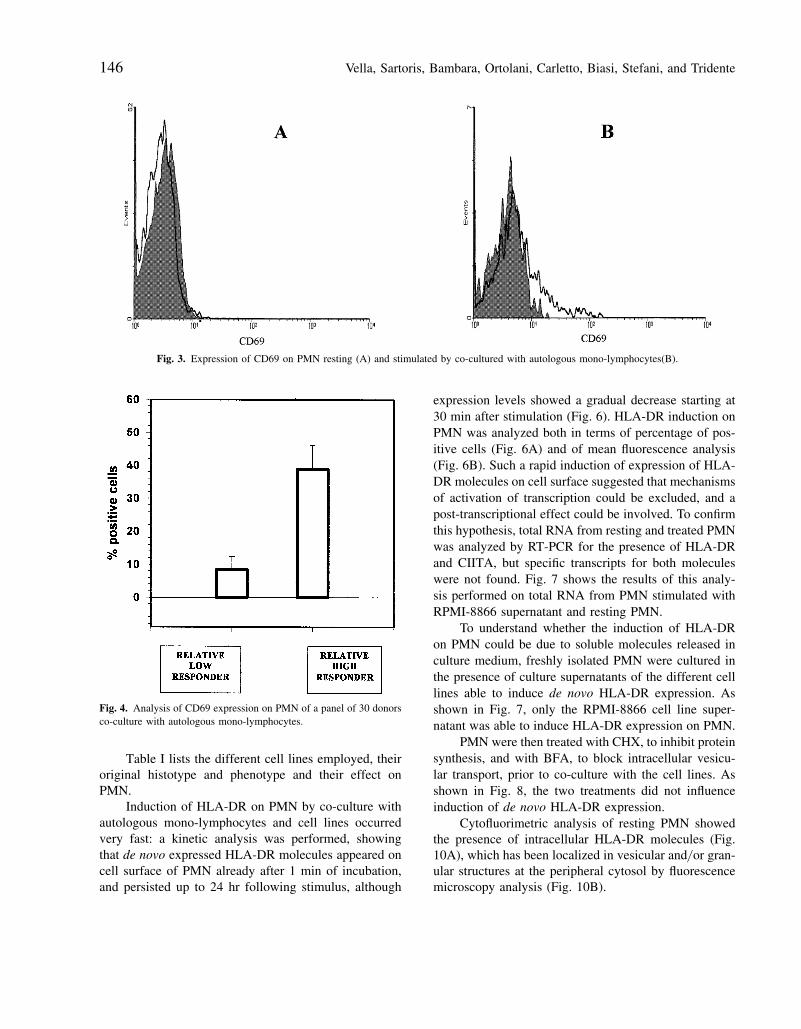
Vella, Sartoris, Bambara, Ortolani, Carletto, Biasi, Stefani, and Tridente146
Fig. 3. Expression of CD69 on PMN resting (A) and stimulated by co-cultured with autologous mono-lymphocytes(B).
Fig. 4. Analysis of CD69 expression on PMN of a panel of 30 donorsco-culture with autologous mono-lymphocytes.
Table I lists the different cell lines employed, theiroriginal histotype and phenotype and their effect onPMN.
Induction of HLA-DR on PMN by co-culture withautologous mono-lymphocytes and cell lines occurredvery fast: a kinetic analysis was performed, showingthat de novo expressed HLA-DR molecules appeared oncell surface of PMN already after 1 min of incubation,and persisted up to 24 hr following stimulus, although
expression levels showed a gradual decrease starting at30 min after stimulation (Fig. 6). HLA-DR induction onPMN was analyzed both in terms of percentage of pos-itive cells (Fig. 6A) and of mean fluorescence analysis(Fig. 6B). Such a rapid induction of expression of HLA-DR molecules on cell surface suggested that mechanismsof activation of transcription could be excluded, and apost-transcriptional effect could be involved. To confirmthis hypothesis, total RNA from resting and treated PMNwas analyzed by RT-PCR for the presence of HLA-DRand CIITA, but specific transcripts for both moleculeswere not found. Fig. 7 shows the results of this analy-sis performed on total RNA from PMN stimulated withRPMI-8866 supernatant and resting PMN.
To understand whether the induction of HLA-DRon PMN could be due to soluble molecules released inculture medium, freshly isolated PMN were cultured inthe presence of culture supernatants of the different celllines able to induce de novo HLA-DR expression. Asshown in Fig. 7, only the RPMI-8866 cell line super-natant was able to induce HLA-DR expression on PMN.
PMN were then treated with CHX, to inhibit proteinsynthesis, and with BFA, to block intracellular vesicu-lar transport, prior to co-culture with the cell lines. Asshown in Fig. 8, the two treatments did not influenceinduction of de novo HLA-DR expression.
Cytofluorimetric analysis of resting PMN showedthe presence of intracellular HLA-DR molecules (Fig.10A), which has been localized in vesicular and/ or gran-ular structures at the peripheral cytosol by fluorescencemicroscopy analysis (Fig. 10B).
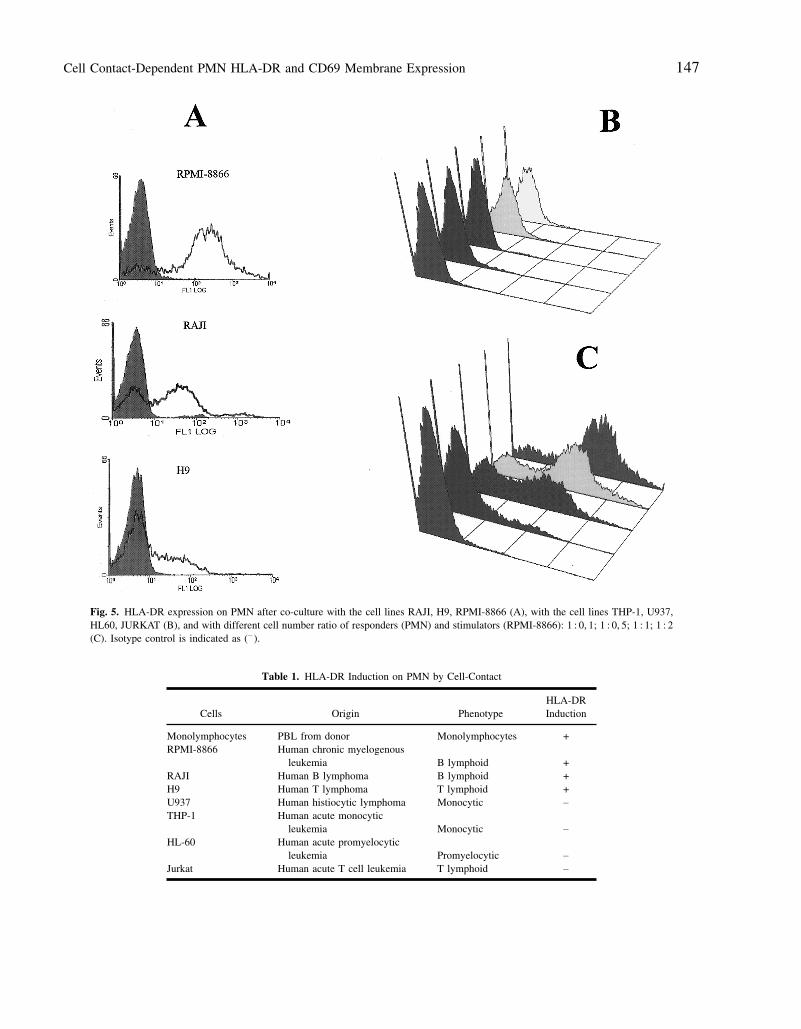
Cell Contact-Dependent PMN HLA-DR and CD69 Membrane Expression 147
Fig. 5. HLA-DR expression on PMN after co-culture with the cell lines RAJI, H9, RPMI-8866 (A), with the cell lines THP-1, U937,HL60, JURKAT (B), and with different cell number ratio of responders (PMN) and stimulators (RPMI-8866): 1 : 0, 1; 1 : 0, 5; 1 : 1; 1 : 2(C). Isotype control is indicated as (− ).
Table 1. HLA-DR Induction on PMN by Cell-Contact
HLA-DRCells Origin Phenotype Induction
Monolymphocytes PBL from donor Monolymphocytes +RPMI-8866 Human chronic myelogenous
leukemia B lymphoid +RAJI Human B lymphoma B lymphoid +H9 Human T lymphoma T lymphoid +U937 Human histiocytic lymphoma Monocytic −
THP-1 Human acute monocyticleukemia Monocytic −
HL-60 Human acute promyelocyticleukemia Promyelocytic −
Jurkat Human acute T cell leukemia T lymphoid −
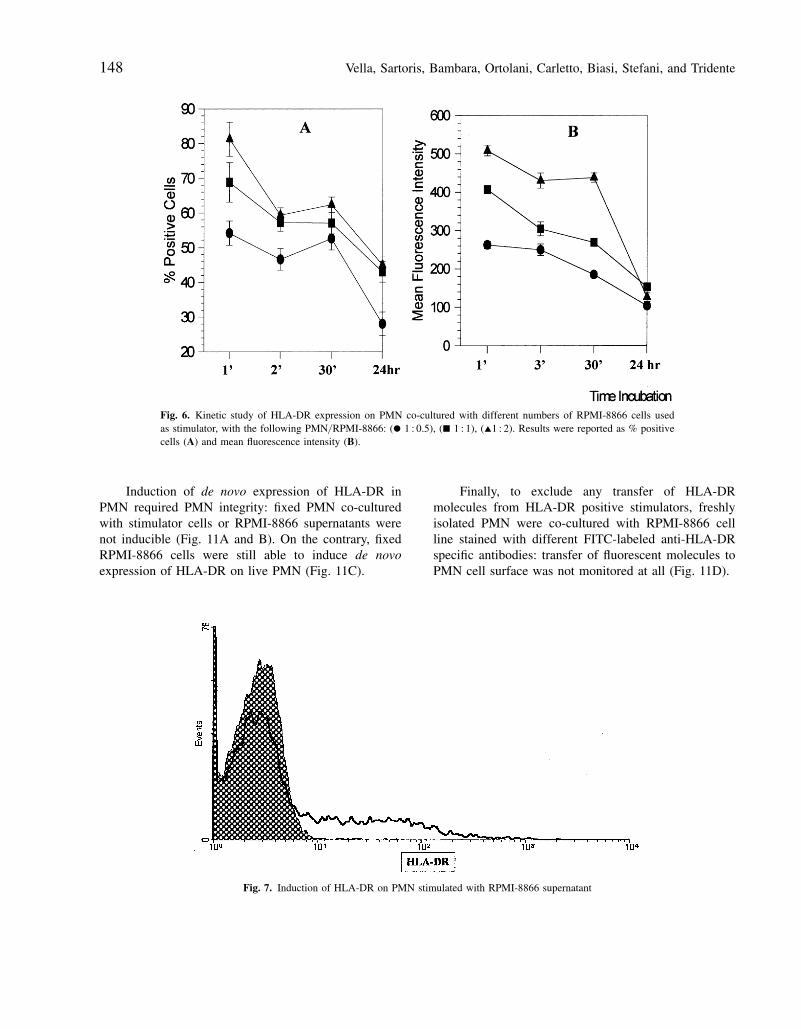
Vella, Sartoris, Bambara, Ortolani, Carletto, Biasi, Stefani, and Tridente148
Fig. 6. Kinetic study of HLA-DR expression on PMN co-cultured with different numbers of RPMI-8866 cells usedas stimulator, with the following PMN/ RPMI-8866: (O 1 : 0.5), (Q 1 : 1), (I1 : 2). Results were reported as % positivecells (A) and mean fluorescence intensity (B).
Induction of de novo expression of HLA-DR inPMN required PMN integrity: fixed PMN co-culturedwith stimulator cells or RPMI-8866 supernatants werenot inducible (Fig. 11A and B). On the contrary, fixedRPMI-8866 cells were still able to induce de novoexpression of HLA-DR on live PMN (Fig. 11C).
Finally, to exclude any transfer of HLA-DRmolecules from HLA-DR positive stimulators, freshlyisolated PMN were co-cultured with RPMI-8866 cellline stained with different FITC-labeled anti-HLA-DRspecific antibodies: transfer of fluorescent molecules toPMN cell surface was not monitored at all (Fig. 11D).
Fig. 7. Induction of HLA-DR on PMN stimulated with RPMI-8866 supernatant
Cell Contact-Dependent PMN HLA-DR and CD69 Membrane Expression 149
Fig. 8. Induction of HLA-DR on PMN treated with CHX (− )or BFA(− ), before co-culture with RPMI-8866cell line. Untreated cells are indicated as (− ) The fitc-isotype control is indicated as (− ).
Fig. 9. RT-PCR analysis for presence of specific HLA-DR and CIITA transcripts on total RNA extracted from PMN restingand stimulated with RPMI-8866 supernatant.
Vella, Sartoris, Bambara, Ortolani, Carletto, Biasi, Stefani, and Tridente150
Fig. 10. Detection of intracellular HLA-DR molecules by immunofluorescence on freshly PMN. Flow-Cytometry (A) andmicroscopic analysis (B).
Fig. 11. Expression of HLA-DR molecules on PMN surface in different experimental conditions: fixed PMN cultured with freshlyRPMI-8866 cell line (A) or RPMI-8866 supernatant (B); fixed RPMI-8866 cell line co-culture with freshly PMN (C); RPMI-8866cell line stained with anti-HLA-DR-FITC before co-culture with freshly PMN (D).

Cell Contact-Dependent PMN HLA-DR and CD69 Membrane Expression 151
DISCUSSION
This report shows that an intracellular pool of HLA-DR molecules is present in the cytoplasm of restingmature PMN which can be rapidly transported to cellsurface in response to a cell-contact stimulus with donor-derived autologous mono-lymphocytes or certain celllines of lymphoid origin, as suggested by the experi-ments shown in Fig. 11. On the contrary, cell lines ofmyeloid origin and the T cell leukemia JURKAT, with aT cell immature phenotype, did not induce HLA-DR onPMN by cell-contact.
The induction of HLA-DR cell surface expression isa very rapid effect, requiring a few min of contact betweencell surfaces, and independent on de novo gene activationand protein synthesis, as RT-PCR analysis of HLA-DRand CIITA transcripts and experiments with CHX haveconfirmed. Ineffectiveness of BFA treatment and detec-tion of a pool of intracellular HLA-DR molecules on rest-ing PMN suggest that cell contact transduces a mobiliza-tion signal of these intracellular molecules located in apost-Golgi compartment to cell surface. This effect ofmobilization of HLA-DR molecules has been observedalso treating PMN with the culture supernatant of RPMI-8866, the cell line inducing by cell-contact HLA-DRexpression levels on PMN higher than those found withRaji and H9. We wish to speculate that still unknownsurface molecule(s) expressed on cell lines of lymphoidbut not myeloid phenotype could be responsible for thesignal of rapid HLA-DR mobilization to cell surface, byinteracting to specific counter-receptor(s) expressed onthe cell surface of resting PMN. This hypothetical surfacemolecule(s), expressed on RPMI-8866 at levels higherthan on the other cell lines analyzed, could be more sub-ject to shedding in RPMI-8866 than in other cell lines andpresent in the culture supernatant.
It has been reported that several cell types cansecrete small membrane vesicles called exosomes as aconsequence of fusion of multivesicular late endosomes/lysosomes with the plasma membrane. Antigen present-ing cells such as B lymphocytes and dendritic cells secreteexosomes containing MHC-I and -II molecules, able toactivate T cell proliferation in vitro (12). It was then nec-essary to exclude that the HLA-DR molecules presentthe novo on stimulated PMN could be due to aspecificexternal binding or to hypothetical unknown ligands. Co-culture of PMN with RPMI-8866 stained with fluores-cent anti-HLA-DR antibodies did not result in any trans-fer of fluorescence to PMN, confirming that HLA-DR
molecules induced on PMN surface by cell contact stimu-lus derive from proper PMN intracellular pool.
The best known function of HLA-DR moleculesis presentation of antigenic peptides to specific T lym-phocytes, and it has been recently demonstrated thatPMN are able to function as antigen presenting cellsand to induce T lymphocytes proliferation in a MHC-IIrestricted manner (13).
De novo expression of HLA-DR on PMN cell-sur-face probably depends by an activated status of PMN,in view of the fact that, in the experimental conditionsdescribed, induction of the expression of CD69 activa-tion molecule was also found. PMN, as well as othercells of the hematopoietic lineage contain an intracellu-lar pool of CD69 molecules that are redistributed on cellsurface in response to some stimuli (6).
CD69 is expressed by various cell types in theiractivated status: neutrophils, lymphocytes and platelets;its activation induces Ca2+ ion influx through cell mem-brane and lysozyme release in PMA-activated neu-trophils, suggesting a role of this molecule in the regu-lation of granule exocytosis, probably through a Ca(2+)-dependent mechanism (6).
Differently from HLA-DR, we have found thatCD69 is induced on PMN only by cell contact withperipheral blood-derived mono-lymphocytes, and notwith cell lines of lymphoid or myeloid origin. An intensecross-talk between PMN and mono-lymphocytes medi-ated by cell membrane contact could occur in vivo, andits deregulation could sustain pathological conditions,as suggested by a correlation of CD69 expression withchronic inflammatory diseases (5). MHC-II expressingPMN have been recently associated to the Wegener’sgranulomatosis inflammatory disease (14).
PMN are the first line of defense in acute phaseresponse whereas macrophage are effective in sustainingchronic phase response and our model could be a sim-ple way to reproduce in vitro some of the cellular alter-ations occurring during leukocyte migration to inflamedsites. The results described in the present report suggestthat an MHC-II-mediated antigen presentation by PMN,activated by cell contact interaction with the endothelialbarrier and/ or the stromal tissue at inflamed sites, couldplay a role in the conversion from acute phase responseto chronic phase occurring at the onset of inflammatorydiseases.
Acknowledgements—The authors wish to thank Mrs. N. Bruttiand Mrs. F. Poli for their technical assistance. This work was

Vella, Sartoris, Bambara, Ortolani, Carletto, Biasi, Stefani, and Tridente152
supported by a grant from Progetto Sanita, Fondazione Cassadi Risparmio di Verona, Vicenza, Belluno ed Ancona.
REFERENCES
1. Germain, R. N. 1994. MHC dependent antigen processing andpeptide presentation: providing ligands for lymphocyte activation.Cell. 76:287.
2. Mach, B., V. Steimle, E. Martinez-Soria, and W. Reith. 1996. Reg-ulation of MHC class II genes: lessons from a disease. Annu. Rev.Immunol. 14:301.
3. Amtruda, T. T., R. Bohman, J. Ranyard, and H. P. Koeffler. 1987.Pattern of expression of HLA-DR and HLA-DQ antigens andmRNA in myeloid differentiation. Blood. Apr; 69(4):1225–1236.
4. Gosselin, E. J., K. Wardwell, W. F. Rigby, and P. M. Guyre.1993. Induction of MHC class II on human polymorphonuclearneutrophils by granulocyte/ macrophage colony-stimulating factor,IFN-gamma, and IL-3. J. Immunol. Aug 1;151(3):1482–1490.
5. Marzio, R., J. Mauel, and S. Betz-Corradin. 1999. CD69 and regu-lation of the immune function. Immunopharmacol. Immunotoxicol.Aug;21(3):565–582.
6. Gavioli, R, A. Risso, D. Smilovich, I. Baldissarro, M. C. Capra, A.Bargellesi, and M. E. Cosulich. 1992. CD69 molecule in humanneutrophils: its expression and role in signal-transducing mecha-nisms. Cell Immunol. Jun;142(1):186–196
7. Bellavite, P., S. Chirumbolo, C. Mansoldo, G. Gandini, and P. Dri.1992. Simultaneous assay for oxidative metabolism and adhesion
of human neutrophils: evidence for correlations and dissociationsof the two responses. J. Leukoc. Biol. Apr;51(4):329–335.
8. Accolla, R. S., N. Gross, S. Carrel, and G. Corte. Distinct formsof both and subunits are present in the human Ia molecular pool.Proc. Nat. Acad. Sci. USA. 78:4549–4551.
9. Krug, N., A. M. Thurau, P. Lackie, J. Baier, G. Schultze-Wern-inghaus, C. H. Rieger, and U. J. Schauer. 1996. A flow cytometricmethod for the detection of intracellular basic proteins in unsep-arated peripheral blood and bone marrow eosinophils. Immunol.Methods. Apr 19;190(2):245-254.
10. Maniatis, T., E. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor: Cold SpringHarbor Laboratory.
11. Chang, C. H. and R. A. Flavell. 1995. Class II transactivator regu-lates the expression of multiple genes involved in antigen presen-tation. J. Exp. Med. Feb 1;181(2):765–767.
12. Denzer, K., M. J. Kleijmeer, H. F. Heijnen, W. Stoorvogel, and H.J. Geuze. 2000. Exosome: from internal vesicle of the multivesic-ular body to intercellular signaling device. J. Cell Sci. Oct;113 Pt19:3365–3374.
13. Radsak, M., C. Iking-Konert, S. Stegmaier, K. Andrassy, and G.M. Hansch. 2000. Polymorphonuclear neutrophils as accessorycells for T-cell activation: major histocompatibility complex classII restricted antigen-dependent induction of T-cell proliferation.Immunology. Dec;101(4):521–530
14. Hansch, G. M., M. Radsak, C. Wagner, B. Reis, A. Koch,A. Breitbart, and K. Andrassy. 1999. Expression of major his-tocompatibility class II antigens on polymorphonuclear neu-trophils in patients with Wegener’s granulomatosis. Kidney Int.May;55(5):1811–1818.