cardioprotective effect of apelin-13 on cardiac performance
TRANSCRIPT

Circulation JournalOfficial Journal of the Japanese Circulation Societyhttp://www.j-circ.or.jp
pelin is a recently described ligand for the G-protein-coupled receptor APJ (APLNR).1,2 The apelin and APJ constitute a signaling pathway implicated in
numerous cardiac and vascular functions. In addition, apelin is synthesized as a pre-propeptide consisting of 77 amino acids with shorter biologically active forms.3 Given the cell and developmental specific patterns of expression of apelin and APJ in vascular and cardiac structures and the initial studies in developmental model organisms, it is likely that this pathway has a fundamental role in the embryogenesis of the cardiovas-cular system.4 In the adult cardiovascular system, both apelin and APJ are expressed in the endothelium of heart, kidney, and lung, and APJ is expressed by myocardial cells and some vas-cular smooth muscle cells.5,6 Recent studies have suggested that the apelin/APJ pathway is a pivotal regulator of cardiovas-cular function. Apelin has been shown to have a positive effect on contractility, and experiments using myocardial injury models have suggested that apelin also has a positive inotropic effect on failing myocardium.7,8 Apelin causes endothelium-
dependent vasorelaxation by triggering the release of nitric oxide (NO), and this effect can be almost completely abolished in the presence of endothelial NO synthase (eNOS) inhibitor, suggesting that the apelin/APJ pathway exerts a vasorelaxation effect via activation of serine/threonine Akt and eNOS phos-phorylation.9–11 Moreover, previous studies have demonstrated that treatment with the NOS inhibitor, L-NG-nitroarginine methyl ester (L-NAME), enhances the expression of NAD(P)H oxidase subunit, which activates the production of reactive oxygen species (ROS).12
Recently, a reduction in myocardial apelin/APJ expression has been reported in experimental models of heart failure (HF). In a pressure overload-induced HF model, apelin knock-out (KO) mice developed severely impaired heart contractil-ity.13 Moreover, Iwanaga et al reported that the cardiac apelin/APJ pathway is markedly downregulated in Dahl salt-sensitive hypertensive (DS) rats with HF.14 Recently, we also showed a similar reduction in cardiac apelin/APJ protein expression seen and these suppressive expressions were restored by
A
Received July 3, 2011; revised manuscript received September 22, 2011; accepted September 26, 2011; released online November 12, 2011 Time for primary review: 19 days
Department of Hypertension and Cardiorenal Medicine, Dokkyo Medical University School of Medicine, Tochigi, JapanMailing address: Naohiko Kobayashi, MD, PhD, Department of Hypertension and Cardiorenal Medicine, Dokkyo Medical University
School of Medicine, Mibu, Tochigi 321-0293, Japan. E-mail: [email protected] doi: 10.1253/circj.CJ-11-0689All rights are reserved to the Japanese Circulation Society. For permissions, please e-mail: [email protected]
Cardioprotective Effect of Apelin-13 on Cardiac Performance and Remodeling
in End-Stage Heart FailureWataru Koguchi, MD; Naohiko Kobayashi, MD, PhD; Hiroshi Takeshima, MD; Mayuko Ishikawa, MD;
Fumihiro Sugiyama, MD; Toshihiko Ishimitsu, MD, PhD
Background: Apelin and its cognate G protein-coupled receptor, APJ, constitute a signaling pathway with a positive inotropic effect on cardiac function. Recently, we and other investigators demonstrated that a reduction in myocardial apelin/APJ expression might play a critical role in experimental models of end-stage heart failure (HF). Therefore, we evaluated whether exogenous apelin infusion restores apelin/APJ expression and improves cardiac function in the failing heart of Dahl salt-sensitive hypertensive (DS) rats.
Methods and Results: High salt-loaded DS rats were treated with vehicle and pyroglutamylated apelin-13 (Pyr-AP13; 200 μg · kg–1 · day–1, IP) from the age of 11 to 18 weeks. Decreased end-systolic elastance and percent frac-tional shortening in failing rats was significantly ameliorated by Pyr-AP13. Pyr-AP13 effectively inhibited vascular lesion formation and suppressed expression of inflammation factors such as tumor necrosis factor-α and interleukin-1β protein. Downregulation of apelin and APJ expression, and phosphorylation of endothelial nitric oxide synthase at Ser1177 and Akt at Ser473 in failing rats was significantly increased by Pyr-AP13. Upregulation of NAD(P)H oxidase p22phox, p47phox, and gp91phox in DS rats was significantly suppressed by Pyr-AP13.
Conclusions: Exogenous apelin-13 may ameliorate cardiac dysfunction and remodeling and restore apelin/APJ expression in DS rats with end-stage HF. Thus, apelin-13 may have significant therapeutic potential for end-stage HF.
Key Words: Apelin; Cardiac function; Heart failure; Remodeling
Advance Publication by J-STAGE

KOGUCHI W et al.
angiotensin II (Ang II) type 1 receptor (AT1R) blocker in end-stage HF with severe left ventricular (LV) dysfunction.15 How-ever, the effect of exogenous apelin on the apelin/APJ pathway in end-stage HF remain unknown. Accordingly, we investi-gated whether exogenous apelin-13 restores expression of apelin and APJ and improves cardiac function and cardiovas-cular remodeling in DS rats with end-stage severe HF.
MethodsAll experimental procedures and protocols in this study were in accordance with the Dokkyo Medical University School of Medicine institutional guidelines for animal research and with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Animal Models and Experimental DesignsMale, inbred DS rats and Dahl salt-resistant (DR) rats (Eisai, Tokyo, Japan) were weaned and fed a diet containing 0.3% NaCl until the age of 6 weeks. Thereafter, they were fed a diet containing 8% NaCl until the age of 18 weeks. Systolic blood pressure (SBP) was measured by the tail-cuff method at the start of the 8% NaCl diet and at 1-week intervals thereafter. Transthoracic echocardiography evaluating the LV end-dia-stolic diameter (LVEDD), and percent fractional shortening (%FS) were performed at 18 weeks, as described previously.16–18 At the age 11 weeks, when LV hypertrophy developed, DS rats were randomly divided into 2 groups: treated with vehicle (DSHF-V), and automatic infusion of pyroglutamylated ape-lin-13 (Pyr-AP13, 200 μg · kg–1 · day–1; DSHF-AP) via an Alzet osmotic minipump implanted intraperitoneally in the abdomi-nal cavity followed by closure of the abdominal wall in layers. This common posttranslational modification preserves bio-logical activity by rendering the peptide more resistant to enzymatic cleavage. Therefore, we used Pyr-AP13 for this chronic study.19 Age-matched male DR rats served as a con-trol group (DR-C).
LV Pressure-Volume RelationThe chest was opened via a midline sternotomy, and the peri-cardium was dissected to expose the heart. The LV end-sys-tolic pressure – volume relationship (contractility: end-systolic elastance (Ees)) was modified for the conductance catheter technique as described previously.20–22 Briefly, the conductance catheter was inserted into the LV through the apex and pushed until the distal tip was placed in the ascending aorta along the longitudinal axis of the LV. A 3F catheter-tip micromanometer (SPR-524, Millar instruments) was also inserted into the LV from the apex. To change the preload, a snare was placed around the inferior vena cava. We recorded conductance vol-ume and LV pressure simultaneously during gradual inferior vena cava occlusion. Electrical signals were digitized through an analog-to-digital converter (AD12-8, Contec) at a sampling frequency of 1,000 Hz with 12-bit resolution and stored on a personal computer (Dynabook SS 330, Toshiba). The points of the end-systolic pressure – volume relation were determined by an iterative technique reported previously.23 In several con-secutive pressure – volume loops, the points of each cardiac cycle with maximum pressure-to-volume ratio were first deter-mined. Linear regression of these points with expression:
Pes=Ees(Ves – V0)yielded estimates for the slope, or Ees, and the volume-axis intercept (V0), where Pes and Ves are end-systolic pressure and volume, respectively.
Western Blot AnalysisApelin, APJ, Akt, NAD(P)H oxidase p22phox, p47phox, gp91phox, tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), mono-cyte chemoattractant protein-1 (MCP-1), angiotensin-converting enzyme (ACE), AT1R, and nuclear factor-κ B (NF-κB) protein expressions were measured as described previously.21,22,24 LV was homogenized (25% W/vol) in 10 mmol/L HEPES buffer, pH 7.4, containing 320 mmol/L sucrose, 1 mmol/L EDTA, 1 mmol/L DTT, 10 μg/ml leupeptin, and 2 μg/ml aprotinin at 0°C to 4°C with a polytron homogenizer. Protein concentrations were determined with bovine serum albumin as a standard protein. Equal amounts of protein were loaded in each lane of SDS-PAGE using 13% gels. The proteins in the gels were trans-ferred electrophoretically to PVDF sheets for 1 h at 2 mA/cm2. The sheets were immunoblotted with an anti-apelin, anti-APJ, anti-Akt, anti-NAD(P)H oxidase subunits, anti-TNF-α, anti-IL-1β, and anti-MCP-1 antibody (Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA) in a buffer containing 10 mmol/L Tris-HCl, pH 7.5, 100 mmol/L NaCl, 0.1% Tween 20, and 5% skim milk. The proteins transferred to the sheets were detected using the ECL immunoblotting detection system (Amersham Life Science Inc).
Phosphorylation of eNOS at Ser1177, Akt at Ser473, and NF-κB at Ser536
Phosphorylation of eNOS at Ser1177, Akt at Ser473, and NF-κB at Ser536 was measured in detail as described previously.22,24,25
Detection of Superoxide Anion in the LVHistological detection of superoxide anion in the LV was performed using dihydroethidium (DHE) as described previ-ously.20,22,24
Nitrite ProductionNitrite production was measured as described in detail previ-ously.26,27 The LV was used for the assay of nitrite produc-tion within 24 h. Three 50-μm sections of myocardium from each rat were cut on a vibratome and incubated in a buffer (pH 7.2) containing 25 mmol/L N-2-hydroxyethylpiperazine-N’-2-ethanesulfonic acid (HEPES) (Sigma, St Louis, MO, USA); 140 mmol/L NaCl, 5.4 mmol/L KCl, 1.8 mmol/L CaCl2, 1 mmol/L MgCl2, and 5 mmol/L glucose for 48 h at 37°C. The supernatant was used for the assay of NO2− production, and the amount of NO2− was corrected via the protein amount measured using the Bradford method (Bio-Rad, Richmond, CA, USA). Nitrite was measured with an autoanalyzer (TCI-NOX 1000 m; Tokyo Kasei Kogyo, Tokyo, Japan) using the Griess method.
Histologic Examination and Evaluation of Cardiovascular RemodelingHistologic examination was performed as described in detail previously.15–17,20–22,24 The wall-to-lumen ratio (the area of the vessel wall divided by the area of the total blood vessel lumen) was determined. The area of fibrosis immediately surrounding the blood vessels was calculated, and perivascular fibrosis was determined as the ratio of the area of fibrosis surrounding the vessel wall to the total area of the vessel. To assess the area of myocardial interstitial fibrosis, the area of pathological colla-gen deposition was measured in the microscopic field of each Masson’s trichrome-stained section. The ratio of the total area of fibrosis within the LV myocardial sample to the total area of the LV myocardium in each heart was calculated and used for analysis.
Advance Publication by J-STAGE

Effect of Apelin-13 in CHF
Statistical AnalysisAll values are expressed as mean ± SEM. Mean values were compared among the 3 groups by ANOVA and the Bonferroni post hoc test for multiple comparisons. A value of P<0.05 was considered statistically significant.
ResultsPhysiological Profiles After 7 Weeks’ Treatment With Pyr-AP13 in DS RatsBody weight (BW), SBP, LV weight (LVW) to BW ratio, and heart rate (HR) in the 3 groups are presented in Table. BW was significantly lower in DSHF-V than in DR-C rats. Long-term Pyr-AP13 therapy in DS rats significantly increased BW. In contrast, DSHF-V rats had a higher LVW/BW than did DR-C rats. Long-term Pyr-AP13 therapy in DS rats significantly decreased LVW/BW.
Time-related changes in SBP among the 3 groups are shown in Figure 1. Before feeding with 8% NaCl diet, SBP was 114±3 mmHg in DR-C, 117±4 mmHg in DSHF-V and 118±4 mmHg in DSHF-AP. As shown in Figure 1 and Table, SBP in DSHF-V and DSHF-AP rats was similar and signifi-cantly higher than that in DR-C rats. HR was significantly higher in DSHF-V than in DR-C rats. Long-term Pyr-AP13 therapy in DS rats significantly decreased HR (Table, Figure 1).
LV Ees in End-Systolic Pressure-Volume Relation and Cardiac Function for LVEDD and %FSLVEDD was significantly higher in DSHF-V than in DR-C rats. Long-term Pyr-AP13 therapy in DS rats significantly decreased LVEDD. In contrast, DSHF-V rats had lower Ees and %FS than did DR-C rats. Long-term Pyr-AP13 therapy in DS rats significantly improved Ees and %FS (Table).
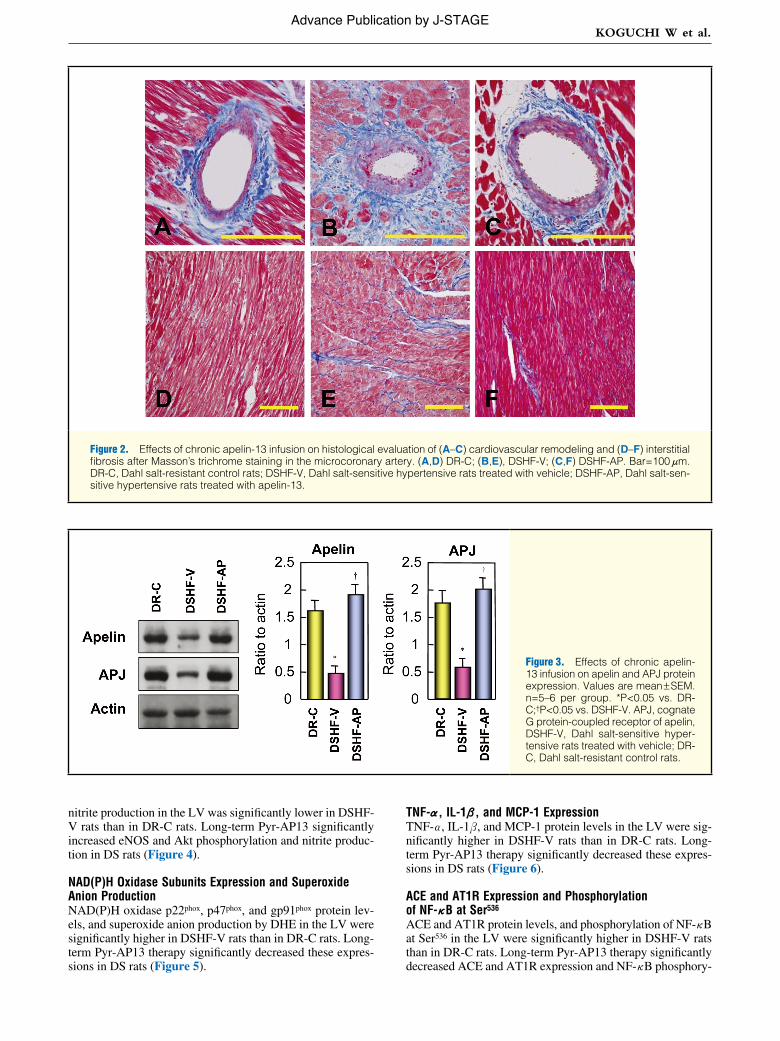
Cardiovascular RemodelingThe wall-to-lumen ratio, perivascular fibrosis, and area of interstitial fibrosis of the heart in the 3 groups are shown in Figure 2F and Table (n=10 tissue sections per group). The wall-to-lumen ratio was significantly increased in DSHF-V rats compared with DR-C. Long-term Pyr-AP13 treatment caused significant amelioration of this ratio. Perivascular and intersti-tial fibrosis was significantly greater in DSHF-V rats than in DR-C rats. Long-term Pyr-AP13 treatment also caused signifi-
cant improvement in perivascular and interstitial fibrosis.
Apelin and APJ ExpressionApelin and APJ protein expressions in the LV of DS rats were significantly lower in DSHF-V rats than in DR-C rats. Long-term Pyr-AP13 significantly increased these expressions in DS rats (Figure 3).
Phosphorylation of eNOS at Ser1177 and Akt at Ser473 and Nitrite ProductionPhosphorylation of eNOS at Ser1177 and Akt at Ser473 and
Table. General Characteristics and Cardiac Function and Remodeling in DS Rats and Apelin-Treated DS Rats
Parameter DR-C DSHF-V DSHF-AP
N 10 9 8
BW, g 474±6 296±5* 391±10*,†
SBP, mmHg 130±3 249±6* 241±7*
LVW/BW, mg/g 1.92±0.04 4.49±0.09* 2.74±0.07*,†
Lung weight/BW, mg/g 3.2±0.1 12.6±1.4* 5.5±0.5*,†
HR, beats/min 409±11 487±13* 414±12†
Ees, mmHg/ml 2,393±201 1,064±94* 2,301±113†
%FS 53.9±0.8 26.1±1.1* 45.4±0.9*,†
LVEDD, mm 7.2±0.1 9.4±0.3* 7.5±0.2†
Wall-to-lumen ratio 0.13±0.01 0.40±0.03* 0.26±0.02*,†
Perivascular fibrosis 0.22±0.02 0.79±0.05* 0.51±0.04*,†
Area of interstitial fibrosis, % 2.0±0.2 13.1±0.8* 6.1±0.5*,†
Data are expressed as mean ± SEM.*P<0.01 vs. DR-C; †P<0.01 vs. DSHF-V.DS, Dahl salt-sensitive hypertensive; DR-C, Dahl salt-resistant control rats; DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; AP, apelin-13; BW, body weight; SBP, systolic blood pressure; LVW, left ventricular weight; HR, heart rate; Ees, contractility; FS, fractional shortening; LVEDD, left ventricular end-diastolic diameter.
Figure 1. Time-related changes in systolic blood pressure in the DR-C, DSHF-V, and DSHF-AP groups. DR-C, Dahl salt-resistant rats; DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; DSHF-AP, Dahl salt-sensitive hypertensive rats treated with apelin-13. Values are mean ± SEM. **P<0.01 vs. DR-C.
Advance Publication by J-STAGE
KOGUCHI W et al.
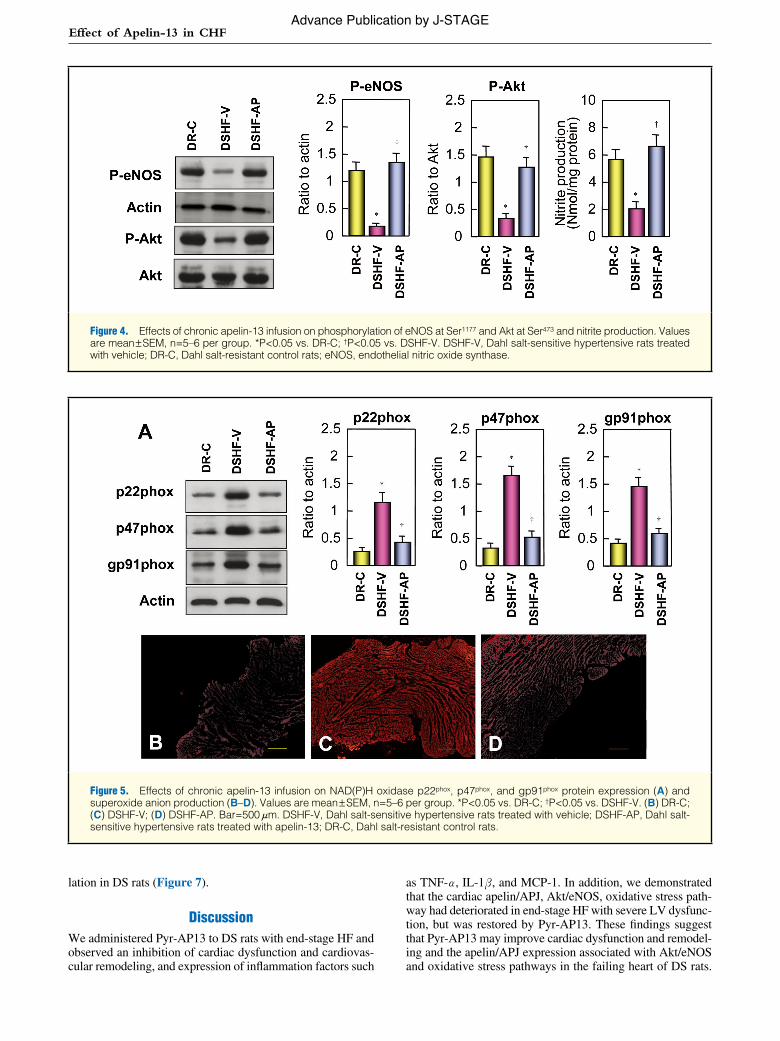
nitrite production in the LV was significantly lower in DSHF-V rats than in DR-C rats. Long-term Pyr-AP13 significantly increased eNOS and Akt phosphorylation and nitrite produc-tion in DS rats (Figure 4).
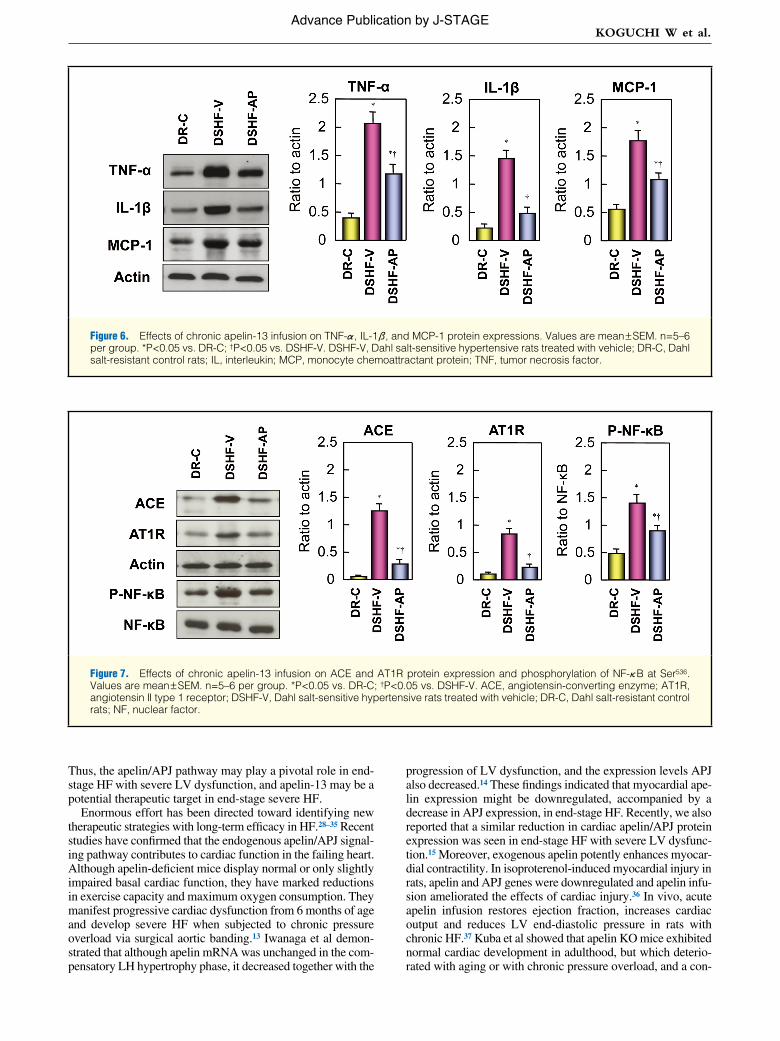
NAD(P)H Oxidase Subunits Expression and Superoxide Anion ProductionNAD(P)H oxidase p22phox, p47phox, and gp91phox protein lev-els, and superoxide anion production by DHE in the LV were significantly higher in DSHF-V rats than in DR-C rats. Long-term Pyr-AP13 therapy significantly decreased these expres-sions in DS rats (Figure 5).
TNF-α, IL-1β, and MCP-1 ExpressionTNF-α, IL-1β, and MCP-1 protein levels in the LV were sig-nificantly higher in DSHF-V rats than in DR-C rats. Long-term Pyr-AP13 therapy significantly decreased these expres-sions in DS rats (Figure 6).
ACE and AT1R Expression and Phosphorylation of NF-κB at Ser536
ACE and AT1R protein levels, and phosphorylation of NF-κB at Ser536 in the LV were significantly higher in DSHF-V rats than in DR-C rats. Long-term Pyr-AP13 therapy significantly decreased ACE and AT1R expression and NF-κB phosphory-
Figure 2. Effects of chronic apelin-13 infusion on histological evaluation of (A–C) cardiovascular remodeling and (D–F) interstitial fibrosis after Masson’s trichrome staining in the microcoronary artery. (A,D) DR-C; (B,E), DSHF-V; (C,F) DSHF-AP. Bar=100 μm. DR-C, Dahl salt-resistant control rats; DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; DSHF-AP, Dahl salt-sen-sitive hypertensive rats treated with apelin-13.
Figure 3. Effects of chronic apelin-13 infusion on apelin and APJ protein expression. Values are mean ± SEM. n=5–6 per group. *P<0.05 vs. DR-C;†P<0.05 vs. DSHF-V. APJ, cognate G protein-coupled receptor of apelin, DSHF-V, Dahl salt-sensitive hyper-tensive rats treated with vehicle; DR-C, Dahl salt-resistant control rats.
Advance Publication by J-STAGE
Effect of Apelin-13 in CHF
lation in DS rats (Figure 7).
DiscussionWe administered Pyr-AP13 to DS rats with end-stage HF and observed an inhibition of cardiac dysfunction and cardiovas-cular remodeling, and expression of inflammation factors such
as TNF-α, IL-1β, and MCP-1. In addition, we demonstrated that the cardiac apelin/APJ, Akt/eNOS, oxidative stress path-way had deteriorated in end-stage HF with severe LV dysfunc-tion, but was restored by Pyr-AP13. These findings suggest that Pyr-AP13 may improve cardiac dysfunction and remodel-ing and the apelin/APJ expression associated with Akt/eNOS and oxidative stress pathways in the failing heart of DS rats.
Figure 4. Effects of chronic apelin-13 infusion on phosphorylation of eNOS at Ser1177 and Akt at Ser473 and nitrite production. Values are mean ± SEM, n=5–6 per group. *P<0.05 vs. DR-C; †P<0.05 vs. DSHF-V. DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; DR-C, Dahl salt-resistant control rats; eNOS, endothelial nitric oxide synthase.
Figure 5. Effects of chronic apelin-13 infusion on NAD(P)H oxidase p22phox, p47phox, and gp91phox protein expression (A) and superoxide anion production (B–D). Values are mean ± SEM, n=5–6 per group. *P<0.05 vs. DR-C; †P<0.05 vs. DSHF-V. (B) DR-C; (C) DSHF-V; (D) DSHF-AP. Bar=500 μm. DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; DSHF-AP, Dahl salt-sensitive hypertensive rats treated with apelin-13; DR-C, Dahl salt-resistant control rats.
Advance Publication by J-STAGE
KOGUCHI W et al.
Thus, the apelin/APJ pathway may play a pivotal role in end-stage HF with severe LV dysfunction, and apelin-13 may be a potential therapeutic target in end-stage severe HF.
Enormous effort has been directed toward identifying new therapeutic strategies with long-term efficacy in HF.28–35 Recent studies have confirmed that the endogenous apelin/APJ signal-ing pathway contributes to cardiac function in the failing heart. Although apelin-deficient mice display normal or only slightly impaired basal cardiac function, they have marked reductions in exercise capacity and maximum oxygen consumption. They manifest progressive cardiac dysfunction from 6 months of age and develop severe HF when subjected to chronic pressure overload via surgical aortic banding.13 Iwanaga et al demon-strated that although apelin mRNA was unchanged in the com-pensatory LH hypertrophy phase, it decreased together with the
progression of LV dysfunction, and the expression levels APJ also decreased.14 These findings indicated that myocardial ape-lin expression might be downregulated, accompanied by a decrease in APJ expression, in end-stage HF. Recently, we also reported that a similar reduction in cardiac apelin/APJ protein expression was seen in end-stage HF with severe LV dysfunc-tion.15 Moreover, exogenous apelin potently enhances myocar-dial contractility. In isoproterenol-induced myocardial injury in rats, apelin and APJ genes were downregulated and apelin infu-sion ameliorated the effects of cardiac injury.36 In vivo, acute apelin infusion restores ejection fraction, increases cardiac output and reduces LV end-diastolic pressure in rats with chronic HF.37 Kuba et al showed that apelin KO mice exhibited normal cardiac development in adulthood, but which deterio-rated with aging or with chronic pressure overload, and a con-
Figure 6. Effects of chronic apelin-13 infusion on TNF-α, IL-1β, and MCP-1 protein expressions. Values are mean ± SEM. n=5–6 per group. *P<0.05 vs. DR-C; †P<0.05 vs. DSHF-V. DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; DR-C, Dahl salt-resistant control rats; IL, interleukin; MCP, monocyte chemoattractant protein; TNF, tumor necrosis factor.
Figure 7. Effects of chronic apelin-13 infusion on ACE and AT1R protein expression and phosphorylation of NF-κB at Ser536. Values are mean ± SEM. n=5–6 per group. *P<0.05 vs. DR-C; †P<0.05 vs. DSHF-V. ACE, angiotensin-converting enzyme; AT1R, angiotensin II type 1 receptor; DSHF-V, Dahl salt-sensitive hypertensive rats treated with vehicle; DR-C, Dahl salt-resistant control rats; NF, nuclear factor.
Advance Publication by J-STAGE

Effect of Apelin-13 in CHF
tinuous infusion of apelin-13 in these elderly mice reversed the decreased contractility because of aging.13 In addition, Ashley et al reported that acute administration of apelin in vivo caused a reduction in LV end-diastolic area and an increase in LV elastance, whereas chronic apelin infusion increased cardiac output without causing hypertrophy.7 In a recent important report, Japp et al38 demonstrated that short-term apelin infusion caused peripheral and coronary vasodilatation, reduced cardiac preload and afterload, and increased cardiac output in vivo in humans. These important hemodynamic effects appeared to be preserved in patients with HF. They concluded that APJ ago-nism had potential therapeutic benefits in patients with HF maintained on optimal contemporary medical therapy. In the present study, we showed that apelin-13 infusion ameliorated cardiac function and cardiovascular remodeling, and restored apelin/APJ expression in the LV of DS rats with end-stage HF. These findings suggest that the apelin/APJ pathway is main-tained or even augmented in conditions of chronic pressure overload and the early stages of HF, but is substantially down-regulated in severe HF, and that the beneficial effects of apelin-13 infusion may at least in part be explained by restoration of the apelin/APJ signaling pathway in end-stage HF with severe LV dysfunction.
The vasodilatory response to apelin is endothelium-depen-dent because vasoconstriction occurs in endothelium-denuded vessels.39 This endothelium-dependent vasodilatation appears to be mediated predominantly through eNOS-dependent path-ways.9 The current study found diminished phosphorylation of Akt and eNOS in aortic tissues form diabetic db/db mice. In addition, an abnormal Akt/eNOS phosphorylation pathway might serve to dampen endothelial NO signaling in aortic rings, thereby leading to abnormal vasoactive responses to Ang II and acetylcholine. In aortic rings from db/db mice, apelin treatment significantly enhanced phosphorylation of Akt and eNOS, which is consistent with previously published find-ings.10,40 Apelin can facilitate the activity of eNOS in endothe-lial cells and APJ is crucially involved in apelin-induced phos-phorylation of eNOS, which promotes the release of NO.9,10 In addition, the apelin/APJ pathway mediated amelioration of atherosclerosis might be related to increased NO production, and the NOS inhibitor, L-NAME, blocks apelin-mediated ame-lioration of Ang II-exacerbated vascular disease in the ApoE-KO model.41 These results suggest that chronic apelin infusion potentiates the activity of the apelin/APJ signaling pathway, which may, at least in part, contribute to increased Akt/eNOS phosphorylation. On the other hand, ROS act as intercellular and intracellular messengers that play an important patho-physiologic role in vascular biology.42 Increased oxidative stress resulting from enhanced production of ROS and/or inad-equate mechanisms of antioxidant defense has been recognized as a critical mechanism in the initiation of cardiac dysfunction and the transition from hypertrophy to HF.43 Moreover, numer-ous studies have reported that the failing heart is subjected to increased oxidative stress, and antioxidant strategies have been proposed to prevent the progression and development of chronic HF.44,45 Recently, Foussal et al46 showed that chronic treatment of mice with apelin attenuated pressure-overload-induced LV hypertrophy, and that the prevention of hypertro-phy by apelin was associated with increased myocardial cata-lase activity and decreased plasma lipid hydroperoxide, as an index of oxidative stress. Apelin preserved cardiac function by inhibiting oxidative stress and stimulating catalase activity, supporting the premise that apelin is a potent regulator of car-diomyocyte antioxidant reserve against oxidative stress during hypertrophic remodeling of the heart. In the present study, we
demonstrated that apelin significantly prevented the progres-sion of cardiac dysfunction and cardiovascular remodeling in HF, and inhibited NAD(P)H oxidase subunit expression and ROS production. These findings suggest that chronic apelin infusion may play a pivotal role in activation of the antioxidant system in DS rats with end-stage severe HF.
In the present study, we showed that long-term apelin-13 infusion ameliorated LV weight, cardiovascular remodeling, and cardiac function, and inhibited inflammation and oxidative stress, and restored apelin/APJ expression. However, the under-lying cardioprotective mechanism of apelin-13 is unclear. The apelin/APJ pathway has opposing actions to the renin – angio-tensin system (RAS) in a number of physiologic and patho-physiologic settings.5,9 There is some evidence for direct coun-terregulation of the RAS by the apelin/APJ pathway. Ishida et al10 have demonstrated differences in blood pressure response in mice lacking both AT1R and APJ compared with mice lacking only AT1R. In addition, Ang II stimulation produces superoxide via AT1R and activation of NAD(P)H oxidases in endothelial and vascular smooth muscle cells. Moreover, chronic activation of NF-κB may induce expression of inflam-matory cytokines and produce detrimental consequences. Therefore, to evaluate the mechanism, we performed experi-ments to determine whether long-term apelin-13 infusion inhibited ACE and AT1R expression and NF-κB phosphory-lation. We found that ACE and AT1R protein expressions and NF-κB phosphorylation were upregulated in DS rats, and apelin-13 infusion suppressed these changes. These findings suggest that the cardioprotective mechanism of apelin-13 may in part be explained by inhibition of the RAS and NF-κB signaling pathway.
ConclusionsWe administered long-term apelin infusion DS rats with severe end-stage HF and observed an inhibition of both cardiovascu-lar remodeling and cardiac dysfunction. Moreover, we showed that the cardiac apelin/APJ pathway is suppressed in end-stage HF with severe LV dysfunction, and that this signaling path-way is restored by apelin infusion. In addition, apelin infusion stimulated Akt/eNOS phosphorylation, and inhibited expres-sion of NAD(P)H oxidase subunits and ROS production. These findings suggest that the apelin/APJ pathway may play a critical role in the pathophysiology of HF, and that apelin infusion may be a therapeutic strategy for severe HF.
AcknowledgmentsWe thank Yasuko Mamada, Keiko Fukuda, Hisato Hirata, Yoshifumi Machida, Noriko Suzuki, Fumie Yokotsuka, Kyoko Tabei, and Machiko Sakata for technical assistance.
DisclosureAll the authors declared no conflicts of interest.
References 1. Lee DK, Cheng R, Nguyen T, Fan T, Kariyawasam AP, Liu Y, et al.
Characterization of apelin, the ligand for the APJ receptor. J Neuro-chem 2000; 74: 34 – 41.
2. Habata Y, Fujii R, Hosoya M, Fukusumi S, Kawamata Y, Hinuma S, et al. Apelin, the natural ligand of the orphan receptor APJ, is abundantly secreted in the colostrum. Biochim Biophys Acta 1999; 1452: 25 – 35.
3. Tatemoto K, Hosoya M, Habata Y, Fujii R, Kakegawa T, Zou MX, et al. Isolation and characterization of a novel endogenous peptide ligand for the human APJ receptor. Biochem Biophys Res Commun 1998; 251: 471 – 476.
4. Kasai A, Shintani N, Oda M, Kakuda M, Hashimoto H, Matsuda T,
Advance Publication by J-STAGE

KOGUCHI W et al.
et al. Apelin is a novel angiogenic factor in retinal endothelial cells. Biochem Biophys Res Commun 2004; 325: 395 – 400.
5. Chen MM, Ashley EA, Deng DX, Tsalenko A, Deng A, Tabibiazar R, et al. Novel role for the potent endogenous inotrope apelin in human cardiac dysfunction. Circulation 2003; 108: 1432 – 1439.
6. Kleinz MJ, Davenport AP. Immunocytochemical localization of the endogenous vasoactive peptide apelin to human vascular and endo-cardial endothelial cells. Regul Pept 2004; 118: 119 – 125.
7. Ashley EA, Powers J, Chen M, Kundu R, Finsterbach T, Caffarelli A, et al. The endogenous peptide apelin potently improves cardiac contractility and reduces cardiac loading in vivo. Cardiovasc Res 2005; 65: 73 – 82.
8. Szokodi I, Tavi P, Földes G, Voutilainen-Myllylä S, Ilves M, Tokola H, et al. Apelin, the novel endogenous ligand of the orphan receptor APJ, regulates cardiac contractility. Circ Res 2002; 91: 434 – 440.
9. Tatemoto K, Takayama K, Zou MX, Kumaki I, Zhang W, Kumano K, et al. The novel peptide apelin lowers blood pressure via a nitric oxide-dependent mechanism. Regul Pept 2001; 99: 87 – 92.
10. Ishida J, Hashimoto T, Hashimoto Y, Nishiwaki S, Iguchi T, Harada S, et al. Regulatory roles for APJ, a seven-transmembrane receptor related to angiotensin-type 1 receptor in blood pressure in vivo. J Biol Chem 2004; 279: 26274 – 26279.
11. Zhong JC, Yu XY, Huang Y, Yung LM, Lau CW, Lin SG. Apelin modulates aortic vascular tone via endothelial nitric oxide synthase phosphorylation pathway in diabetic mice. Cardiovasc Res 2007; 74: 388 – 395.
12. Harrison CB, Drummond GR, Sobey CG, Selemidis S. Evidence that nitric oxide inhibits vascular inflammation and superoxide produc-tion via a p47phox-dependent mechanism in mice. Clin Exp Phar-macol Physiol 2010; 37: 429 – 434.
13. Kuba K, Zhang L, Imai Y, Arab S, Chen M, Maekawa Y, et al. Impaired heart contractility in Apelin gene-deficient mice associated with aging and pressure overload. Circ Res 2007; 101: e32 – e42.
14. Iwanaga Y, Kihara Y, Takenaka H, Kita T. Down-regulation of car-diac apelin system in hypertrophied and failing hearts: Possible role of angiotensin II-angiotensin type 1 receptor system. J Mol Cell Car-diol 2006; 41: 798 – 806.
15. Fukushima H, Kobayashi N, Takeshima H, Koguchi W, Ishimitsu T. Effects of olmesartan on Apelin/APJ and Akt/endothelial nitric oxide synthase pathway in Dahl rats with end-stage heart failure. J Cardio-vasc Pharmacol 2010; 55: 83 – 88.
16. Kobayashi N, Nishikimi T, Horinaka S, Ishimitsu T, Matsuoka H. Effects of TCV-116 on expression of NOS and adrenomedullin in failing heart of Dahl salt-sensitive rats. Atherosclerosis 2001; 156: 255 – 265.
17. Kobayashi N, Horinaka S, Mita S, Yoshida K, Honda T, Kobayashi T, et al. Aminoguanidine inhibits mitogen-activated protein kinase and improves cardiac performance and cardiovascular remodeling in failing hearts of salt-sensitive hypertensive rats. J Hypertens 2002; 20: 2475 – 2485.
18. Kobayashi N, Horinaka S, Mita S, Nakano S, Honda T, Yoshida K, et al. Critical role of Rho-kinase pathway for cardiac performance and remodeling in failing rat hearts. Cardiovasc Res 2002; 55: 757 – 767.
19. Falcão-Pires I, Gonçalves N, Henriques-Coelho T, Moreira-Gonçalves D, Roncon-Albuquerque R Jr, Leite-Moreira AF. Apelin decreases myocardial injury and improves right ventricular function in mono-crotaline-induced pulmonary hypertension. Am J Physiol Heart Circ Physiol 2009; 296: H2007 – H2014.
20. Kobayashi N, Yoshida K, Nakano S, Ohno T, Honda T, Tsubokou Y, et al. Cardioprotective mechanisms of eplerenone on cardiac perfor-mance and remodeling in failing rat hearts. Hypertension 2006; 47: 671 – 679.
21. Ohno T, Kobayashi N, Yoshida K, Fukushima H, Matsuoka H. Car-dioprotective effect of benidipine on cardiac performance and remod-eling in failing rat hearts. Am J Hypertens 2008; 21: 224 – 230.
22. Kobayashi N, Takeshima H, Fukushima H, Koguchi W, Mamada Y, Hirata H, et al. Cardioprotective effects of pitavastatin on cardiac performance and remodeling in failing rat hearts. Am J Hypertens 2009; 22: 176 – 182.
23. Ito H, Takaki M, Yamaguchi H, Tachibana H, Suga H. Left ventricu-lar volumetric conductance catheter for rats. Am J Physiol 1996; 270: H1509 – H1514.
24. Kobayashi N, Ohno T, Yoshida K, Fukushima H, Mamada Y, Nomura M, et al. Cardioprotective mechanism of telmisartan via PPAR-γ-eNOS pathway in Dahl salt-sensitive hypertensive rats. Am J Hypertens 2008; 21: 576 – 581.
25. Kobayashi N, Hara K, Tojo A, Onozato ML, Honda T, Yoshida K, et al. Eplerenone shows renoprotective effect by reducing LOX-1-mediated adhesion molecule, PKCε-MAPK-p90RSK, and Rho-kinase pathway. Hypertension 2005; 45: 538 – 544.
26. Kobayashi N, Mori Y, Nakano S, Tsubokou Y, Kobayashi T, Shirataki H, et al. TCV-116 stimulates eNOS and caveolin-1 expression and improves coronary microvascular remodeling in normotensive and angiotensin II-induced hypertensive rats. Atherosclerosis 2001; 158: 359 – 368.
27. Kobayashi N, Mori Y, Nakano S, Tsubokou Y, Shirataki H, Matsuoka H. Celiprolol stimulates endothelial nitric oxide synthase expression and improves myocardial remodeling in deoxycorticosterone acetate-salt hypertensive rats. J Hypertens 2001; 19: 795 – 801.
28. Nishi I, Noguchi T, Iwanaga Y, Furuichi S, Aihara N, Takaki H, et al. Effects of exercise training in patients with chronic heart failure and advanced left ventricular systolic dysfunction receiving β-block-ers. Circ J 2011; 75: 1649 – 1655.
29. Mano Y, Anzai T, Kaneko H, Nagatomo Y, Nagai T, Anzai A, et al. Overexpression of human C-reactive protein exacerbates left ventricu-lar remodeling in diabetic cardiomyopathy. Circ J 2011; 75: 1717 – 1727.
30. Shibuya M, Miura T, Fukagawa Y, Akashi S, Oda T, Kawamura S, et al. Tongue muscle-derived stem cells express connexin 43 and improve cardiac remodeling and survival after myocardial infarction in mice. Circ J 2010; 74: 1219 – 1226.
31. Tang XL, Rokosh DG, Guo Y, Bolli R. Cardiac progenitor cells and bone marrow-derived very small embryonic-like stem cells for car-diac repair after myocardial infarction. Circ J 2010; 74: 390 – 404.
32. Isobe K, Kuba K, Maejima Y, Suzuki J, Kubota S, Isobe M. Inhibi-tion of endostatin/collagen XVIII deteriorates left ventricular remod-eling and heart failure in rat myocardial infarction model. Circ J 2010; 74: 109 – 119.
33. Matsumiya G, Saitoh S, Sakata Y, Sawa Y. Myocardial recovery by mechanical unloading with left ventricular assist system. Circ J 2009; 73: 1386 – 1392.
34. Hiramitsu S, Miyagishima K, Kimura H, Mori K, Shiino K, Yamada A, et al. Management of severe heart failure. Circ J 2009; 73(Suppl A): A-36 – A-41.
35. Keida T, Ohira H, Fujita M, Chinen T, Nakamura K, Kato T, et al. Quantitative assessment of dyssynchrony using ECG-gated SPECT myocardial perfusion imaging prior to and following cardiac resyn-chronization therapy. Circ J 2009; 73: 1550 – 1553.
36. Jia YX, Pan CS, Zhang J, Geng B, Zhao J, Gerns H, et al. Apelin protects myocardial injury induced by isoproterenol in rats. Regul Pept 2006; 133: 147 – 154.
37. Berry MF, Pirolli TJ, Jayasankar V, Burdick J, Morine KJ, Gardner TJ, et al. Apelin has in vivo inotropic effects on normal and failing hearts. Circulation 2004; 110: 187 – 193.
38. Japp AG, Cruden NL, Barnes G, van Gemeren N, Mathews J, Adamson J, et al. Acute cardiovascular effects of apelin in humans: Potential role in patients with chronic heart failure. Circulation 2010; 121: 1818 – 1827.
39. Maguire JJ, Kleinz MJ, Pitkin SL, Davenport AP. [Pyr1]apelin-13 identified as the predominant apelin isoform in the human heart: Vasoactive mechanisms and inotropic action in disease. Hypertension 2009; 54: 598 – 604.
40. Masri B, Morin N, Pedebernade L, Knibiehler B, Audigier Y. The apelin receptor is coupled to Gi1 or Gi2 protein and is differentially desensitized by apelin fragments. J Biol Chem 2006; 281: 18317 – 18326.
41. Chun HJ, Ali ZA, Kojima Y, Kundu RK, Sheikh AY, Agrawal R, et al. Apelin signaling antagonizes Ang II effects in mouse models of atherosclerosis. J Clin Invest 2008; 118: 3343 – 3354.
42. Touyz RM, Schiffrin EL. Reactive oxygen species in vascular biol-ogy: Implications in hypertension. Histochem Cell Biol 2004; 122: 339 – 352.
43. Dhalla AK, Hill MF, Singal PK. Role of oxidative stress in transition of hypertrophy to heart failure. J Am Coll Cardiol 1996; 28: 506 – 514.
44. Nakagami H, Takemoto M, Liao JK. NADPH oxidase-derived superoxide anion mediates angiotensin II-induced cardiac hypertro-phy. J Mol Cell Cardiol 2003; 35: 851 – 859.
45. Day BJ. Catalase and glutathione peroxidase mimics. Biochem Phar-macol 2009; 77: 285 – 296.
46. Foussal C, Lairez O, Calise D, Pathak A, Guilbeau-Frugier C, Valet P, et al. Activation of catalase by apelin prevents oxidative stress-linked cardiac hypertrophy. FEBS Lett 2010; 584: 2363 – 2370.
Advance Publication by J-STAGE