cardiac and smooth muscle cell contribution to the formation of the murine pulmonary veins
TRANSCRIPT

Cardiac and Smooth Muscle Cell Contribution to theFormation of the Murine Pulmonary VeinsCATERINA MILLINO,1 FEDERICA SARINELLA,1 CECILIA TIVERON,2 ANTONELLO VILLA,3
SAVERIO SARTORE,1,4 AND SIMONETTA AUSONI1,4*1Department of Biomedical Sciences, Padua, Italy2Centro Ricerca Sperimentale-Istituto Regina Elena, Rome, Italy3Microscopy and Image Analysis, University of Milano, Bicocca-DIBIT, HSR4National Center Council Unit for Muscle Biology and Physiopathology, Padua, Italy
ABSTRACT Previous studies have demon-strated that the primordial pulmonary veinsoriginate as an outgrowth of the atrial cells andanastomosis with the pulmonary venous plexus.As a consequence of this embryologic origin thetunica media of these vessels is composed of car-diac cells that express atrial specific markers(Lyons et al. [1990] J Cell Biol 111:2427–2436;Jones et al. [1994] Dev Dyn 200:117–128). We usedtransgenic mice for the cardiac troponin I (cTNI)gene and smooth muscle (SM) myosin heavychain as differentiation markers, to analyze howcardiac and SM cells contribute to the formationand structural remodeling of the pulmonaryveins during development. We show here that thetunica media of the adult mouse pulmonary veinscontains an outer layer of cardiac cells and anintermediate SM cell compartment lining downon the inner endothelium. This structural orga-nization is well expressed in the intrapulmonaryveins from the beginning of vasculogenesis, withcardiac cells accumulating over preexistingroots of endothelial and SM cells and extendingto the third bifurcation of the pulmonarybranches without reaching the more distal tips ofthe vessels. On the other hand, SM cells, whichare widely distributed in the intrapulmonaryveins from the embryonic stage E16, accumulatealso in the extrapulmonary branches and reachthe posterior wall of the left atrium, includingthe orifices of the pulmonary veins. This eventtakes place around birth when the pulmonaryblood flow starts to function properly. A modelfor the development of the pulmonary veins ispresented, based upon our analysis. Dev Dyn2000;218:414–425. © 2000 Wiley-Liss, Inc.
Key words: pulmonary veins; pulmonary myo-cardium; smooth muscle; angiogene-sis; vasculogenesis; smooth musclemyosin heavy chain; a-smooth mus-cle actin; cardiac troponin I; trans-genic mice
INTRODUCTION
Pulmonary vascularization develops soon after theformation of the primitive lung bud that originatesfrom the foregut endoderm. Like in many other organs,blood vessels form by vasculogenesis, i.e., in situ differ-entiation of endothelial cells from hemangioblasts andassembling of primitive endothelial tubes (Risau andLemmon, 1988; Risau, 1997). Initially, this primitivenetwork is morphologically quite uniform and naivewith respect to positional and functional identity. How-ever, subsequently newly formed endothelial tubes pro-mote recruitment and differentiation of SM cells fromthe surrounding mesenchyme or from neural crest-derived cells (Rosenquist and Beall, 1990), a processthat presumably requires a cross-talk between the en-dothelial and the muscular compartments (Wang et al.,1998). A third event in the formation of the pulmonaryvascular system is the connection with the heart. Theformation of venous pole suggests that the main branchof the pulmonary vein grows from the heart and even-tually anastomoses with the pulmonary plexus. Thisevent takes place in the fifth week of human gestation(Larsen, 1997). In parallel, pulmonary veins becomecolonized by cardiac cells that contribute to the forma-tion of the tunica media of these vessels (Laudatscher,1968). Cardiac cells are present in the pulmonary veinsin many species (Rauschel, 1836): in large mammalstheir distribution is limited to the extrapulmonary seg-ments, but in rodents there are also intrapulmonaryextensions into the smallest veins (Kramer and Marks,1965). Favaro (1910) proposed the definition of “pulmo-nary myocardium” to identify this extension of cardiaccells into the vascular environment.
Colonization of the pulmonary veins by cardiac cells,initially shown by ultrastructural and electrophysio-logical studies, has been confirmed recently at the mo-lecular level with the demonstration that atrial myo-
Grant sponsor: Telethon; Grant numbers: 928 and B40; Grant spon-sor: MURST ex 60%; Grant sponsor: Regione Veneto 1998; Grantsponsor: Biomed Contract BMH4-CT98-4004.
*Correspondence to: Simonetta Ausoni, Department of BiomedicalSciences, University of Padova, Via G. Colombo, 3, 35121-Padova,Italy. E-mail: [email protected]
Received 7 October 1999; Accepted 13 March 2000
DEVELOPMENTAL DYNAMICS 218:414–425 (2000)
© 2000 WILEY-LISS, INC.

cardium markers such as a-myosin heavy chain (a-MHC), myosin light chain 1A (MLC1A) (Lyons et al.,1990; Jones et al., 1994), and atrial natriuretic factor(ANF) (Jones et al., 1994) are present in the pulmonaryveins. However, it remains unclear whether the pulmo-nary myocardium develops as an outgrowth fromproper atrial segment or from the sinus venosus of theheart.
The functional role of cardiac cells in pulmonaryveins has been associated with their ability to contractwith a rhythmic beat and a valve-like action. Thisproperty might augment resistance to backflow of bloodinto the lungs (Burch and Romney, 1954; Paes deAlmeida et al., 1975) without preventing the normalflow pattern of diastole. In the absence of this muscularcoat in pulmonary veins, pressure in lung capillariescould increase, thus, favouring edema formation.
The basic purpose of this study was to chart thedevelopment of the pulmonary veins by following thedistribution of SM and cardiac muscle cells and theirassemblage and organization during vessel formation.So far, the development of the pulmonary veins hasbeen only examined by looking either at the cardiaccompartment or at the general SM network of thelungs but never at both. To map cardiac cells, we tookadvantage of transgenic lines for the cardiac troponin I(cTNI) gene showing strong expression of the LacZreporter in the pulmonary veins. During development,cardiac cells accumulate around primitive venoustubes according to a caudocranial gradient and spreadout to the smallest vascular branches. The distributionof these cells follows roots of preexisting endothelialand SM cells. SM cells, localized between the cardiacand the endothelial compartments, express an SMmarker of differentiation and are more abundant in theintrapulmonary veins than in the extrapulmonaryones. After birth, the SM component of the extrapul-monary vessels increases significantly and persists inthe adult life. At the same time, the left atrium accu-mulates a-SM actin positive cells for partial incorpora-tion of the pulmonary vein trunk into the posteriorwall.
RESULTSCardiac Troponin I Transgene is a Marker ofthe Pulmonary Myocardium
In situ hybridization analysis with a specific probedemonstrated that the cardiac cTNI gene is highlyexpressed in the pulmonary veins of adult mice, thusreflecting the presence of the pulmonary myocardiumin these vessels (Fig. 1). To analyze more in detailcardiac cell distribution in the pulmonary veins weused transgenic mice carrying fragments of the cTNIpromoter linked to a LacZ reporter gene. Expression inthe pulmonary veins was found in nine independentlines and three of them were selected for further anal-yses. Lines LacZ21 (Di Lisi et al., 1998) and LacZ27contain 230 bp of the cTNI promoter, whereas lineLacZ60 contains 4000 bp of the promoter linked to the
same reporter gene. In Line LacZ21 and Line LacZ60,positive cells were first detected in the pulmonaryveins in E13.5 embryos (Fig. 2A,D), accumulated dur-ing development (Fig. 2B,E) and persisted in the adulttissues (Fig. 2C,F). LacZ staining in the pulmonaryveins was similar, although not overlapping, in theselines. In particular, we observed an abundance of pos-itive cells with wide distribution in large and smallpulmonary veins in Line LacZ21. On the other hand,the overall number of positive cells and the extension oftheir distribution was much lower in line LacZ60. Wealso observed that in Line LacZ27, which showed ex-pression of the transgene in the right atrium and a fewspots of reactivity in the left atrium (Fig. 2G,H), therewere no LacZ-positive cells in the pulmonary veins,neither in the adult stage (Fig. 2I ), nor during devel-opment (not shown). In all the transgenic lines exam-ined, LacZ staining in the vasculature was restricted tothe pulmonary and caval veins.
Tissue organization of the pulmonary veins was ex-amined in cTNI transgenic mice by using LacZ-positivenuclei to identify the pulmonary myocardium and an-tibodies specific to endothelial and SM cells as lineage
Fig. 1. Expression of the cTNI gene in the pulmonary veins. Lungcryosections of adult mice were incubated with a cTNI antisense probeand processed for in situ hybridization. pv, a large intrapulmonary vein.Scale bar 5 280 mm.
415DEVELOPMENT OF THE PULMONARY VEINS

markers. The results presented hereafter relate to lineLacZ21; however, line LacZ60 led to the same conclu-sions. Three distinct layers were identified in the adult
mouse (Fig. 3): an outer layer of cardiac cells, an innerlayer of endothelial cells identified for their expressionof PECAM (Fig. 3A), and a layer of SM cells expressing
Fig. 2. Pattern of cTNI transgene expression in the pulmonary veins.Transgenic lines LacZ21(A–C), LacZ60 (D–F), and LacZ27 (G–I) wereanalyzed by whole-mount staining of embryonic (E13.5), neonatal (N),and adult (Ad) tissues. Lines LacZ21 and LacZ27 were generated with a
short cTNI promoter region (-230/1126). Line LacZ60 was generatedwith a long promoter region (-4000/1126). Note the different stainingamong the lines in the pulmonary veins. pv, pulmonary veins; cv, cavalveins.
416 MILLINO ET AL.

a-SMA (Fig. 3B) and smooth muscle myosin (SMM)(Fig. 3C). We used a confocal microscope to test theexistence of three distinct cell layers inasmuch as theSM layer was not clearly distinguishable from theendothelium and because SM markers have beenfound to be expressed in the endothelium of formingvessels. Confocal analysis corroborated the results ofthe immunocytochemical study. Sections of adultlungs were double labeled either with the anti–a-SMA (Fig. 4A) and PECAM (Fig. 4B) or with anti–a-SMA (Fig. 4D) and anti-SMM (Fig. 4E). A merging ofthe images (Fig. 4C,F) showed that cells expressinga-SMA and SMM were distinct from PECAM-posi-tive endothelial cells and from the pulmonary myo-cardium.
Development of the Pulmonary Veins
We started our study in E12.5 embryos. At thisstage, the orifice of the pulmonary veins was localizedas a small cavity just at the border between the atrialwall and the sinus venosus (Fig. 5A). By the recon-struction of serial sections, we observed that LacZ-positive cardiac cells outgrew from this area for ap-proximately 40–50 mm. In E13 embryos, thepulmonary vein appeared as a lumen, with cardiaccells accumulating around a layer of a presumptiveendothelium (Fig. 5B). This lumen was localized near amain bronchial cavity, suggesting that the connectionbetween the pulmonary vein and the lung is achievedbetween E12.5 and E13. Cardiac cells were distributedaccording to a caudocranial gradient, being more abun-dant proximal to the heart and rare or completelyabsent at the distal tip of the vessel. No LacZ-positivecells were detected in the forming arteries (not shown).We asked ourselves whether at this stage SM cells hadalready been recruited within the tunica media of thepulmonary vein. To target SM cells, we used the anti-SMM antibody, because SM myosin heavy chain isexclusively expressed in fully differentiated SM cellsand because, varying from other markers, such asa-SM actin, SM22 or calponin, it is never expressed inthe developing myocardium (Miano et al., 1994). InE13.5 embryos, the forming pulmonary vein was de-void of SMM reactivity except for rare weakly reactingcells (Fig. 5D, arrowheads). In the embryos, the anti-SMM antiserum stained the SM of bronchi and esoph-agus but not the pulmonary parenchyma and the myo-cardium (Fig. 5D). Arterial SM cells were also darklystained (not shown). These results demonstrate that, inthe E13.5 embryos, the pulmonary vein branch con-necting to the lung is exclusively composed of cardiaccells and that these cells are already detectable pastthe hylus.
In E16 embryos, LacZ-positive cells were widely dis-tributed along the pulmonary veins up to the thirdbifurcation (Fig. 5E). At this stage, the anti-SMM an-tiserum gave strong reactivity in large (Fig. 5F) andsmall (Fig. 5G) intrapulmonary vessels. In addition, ituniformly stained airway as well as pulmonary arter-ies (not shown), thus confirming the specificity of thereaction. At this stage, SM cells were widely distrib-uted above the presumptive endothelium and reachedup to the distal ends of the vessels well before theaccumulation of the cardiac cells in the same areas(Fig. 5H,I).
In E16 embryos, we also observed that, varying fromthe intrapulmonary veins, the extrapulmonary seg-ments did not react with the anti-SMM antiserum (Fig.6A,B). The same difference between intra- and ex-trapulmonary vessels was observed in E18–E19 em-bryos (Fig. 6C,D) where very few SM-positive cellswere found in the extrapulmonary veins. Therefore, weconcluded that in the prenatal life SM cell distributionin the pulmonary veins was restricted to the intrapul-
Fig. 3. Tissue organization of the pulmonary veins. Lung cryosectionsof adult transgenic mice (line LacZ21) were stained for LacZ expressionto identify cardiac cells of the pulmonary myocardium, subsequentlyincubated with either anti-PECAM (A), or anti–a-SMA (B), or anti-SMM(C) and weakly counterstained with Mayer’s Hematoxylin. pm, pulmonarymyocardium; b, bronchi. Scale bar 5 130 mm C (applies to A–C).
417DEVELOPMENT OF THE PULMONARY VEINS

monary branches. This finding is in contrast to whatwas observed at the same stages in the caval veins,where SM cells populated the tunica media, forming acontinuous layer between cardiac and endothelial cells(Fig. 6E,F).
In 1-day-old neonatal mice (N), the SM component ofthe tunica media became more abundant in the in-trapulmonary veins and also accumulated in the ex-trapulmonary trunks (Fig. 6G,H). SMM expression inthe developing pulmonary veins is summarized in Ta-ble1.
Connection of the Pulmonary Veins to theHeart and Remodeling of the Left Atrium
According to a general view, during organogenesisthe muscle wall of the pulmonary veins becomes incor-porated into the posterior wall of the left atrium, andby a process of intussusception generates the SM wallof the definitive left atrium. We asked ourselves whenand how SM cells contributed to this process. Trans-
verse sections from the anterior to the posterior pro-spective of the heart were systematically analyzed forexpression of a-SMA that we chose as a marker of SMcells. We failed to detect SM cells in the wall of the leftatrium both in E16 (Fig. 7A) and in E18 embryos (notshown). This finding is in contrast to what was ob-served at the same stages in the wall of the rightatrium (Fig. 7B) where a layer of loosely connected SMcells were already visible. Background reactions in theatrial lumen were occasionally found where red bloodcells accumulated but were distinguishable from thesignal over the atrial wall. SM cells were first found inthe left atrium in 1-day-old neonatal mice (Fig. 7C,D).
DISCUSSIONStructure of Adult Pulmonary Veins
We have described the temporal and spatial distri-bution of SM and cardiac muscle cells during the de-velopment of the pulmonary veins in the mouse. Car-diac cells were mapped precisely by using cTNI
Fig. 4. A–F: Confocal image analysis of double-labeled cryosectionsfrom adult lung tissues of transgenic mice were stained with anti–a-SMA(FITC) (A) and anti-PECAM (TRITC) (B), or a-SMA (FITC) (D) and
anti-SMM (TRITC) (E). Merge of A and B is shown in C. Merge of D andE is shown in F. Colocalization is depicted in yellow. pm, pulmonarymyocardium. Scale bar 5 25 mm in F (applies to A–F).
418 MILLINO ET AL.
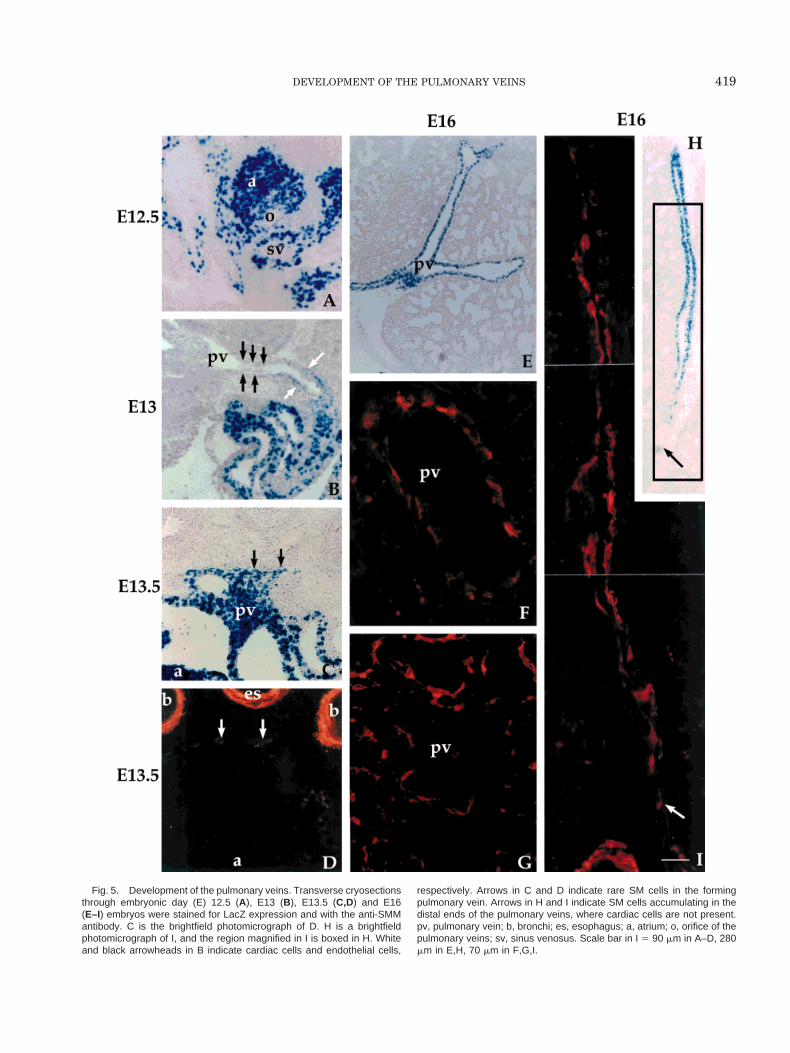
Fig. 5. Development of the pulmonary veins. Transverse cryosectionsthrough embryonic day (E) 12.5 (A), E13 (B), E13.5 (C,D) and E16(E–I) embryos were stained for LacZ expression and with the anti-SMMantibody. C is the brightfield photomicrograph of D. H is a brightfieldphotomicrograph of I, and the region magnified in I is boxed in H. Whiteand black arrowheads in B indicate cardiac cells and endothelial cells,
respectively. Arrows in C and D indicate rare SM cells in the formingpulmonary vein. Arrows in H and I indicate SM cells accumulating in thedistal ends of the pulmonary veins, where cardiac cells are not present.pv, pulmonary vein; b, bronchi; es, esophagus; a, atrium; o, orifice of thepulmonary veins; sv, sinus venosus. Scale bar in I 5 90 mm in A–D, 280mm in E,H, 70 mm in F,G,I.
419DEVELOPMENT OF THE PULMONARY VEINS

Fig. 6. Accumulation of smooth muscle (SM) cells in the extrapul-monary veins. Cryosections of embryos at different stages werestained for LacZ expression to identify cardiac cells, subsequentlyincubated with the anti-SMM to identify SM cells, and then weaklycounterstained with Mayer’s Hematoxylin. The reaction pattern of theextrapulmonary veins in embryonic day (E) 16 embryos (B), E18
embryos (D), and 1-day-old neonatal mice (H) is shown. F shows thereaction pattern of a caval vein in E16 embryos. A,C,E,G: are thebrightfield photomicrographs of B,D,F, and H, respectively. The regionboxed in E is magnified in F. epv, extrapulmonary veins; b, bronchi; cv,caval vein; ao, aorta. Scale bar in H 5 80 mm in A,B,G,H, 280 mm inC, 110 mm D, 220 mm in E, 60 mm in F.
420 MILLINO ET AL.

transgenic lines that exclusively express the LacZ re-porter gene in the myocardium and show strong label-ing in the pulmonary veins. In general, cardiac cells inthe pulmonary veins were found in all the lines withstrong expression in the atria. Lines with no or weakexpression in the left atrium, such as LacZ27, gave noreaction in the pulmonary veins, suggesting that leftatrium and pulmonary myocardium are part of thesame transcriptional domain. However, in LineLacZ60, expression was high in the atria but weak inthe pulmonary veins, raising the possibility that atrialcells and pulmonary myocardium may not be a homog-enous population with respect to their transcriptionalpotential.
The results presented in this study show that, inmice, the tunica media of the pulmonary veins is com-posed of both SM and cardiac cells that express distinctdifferentiation markers throughout the developmentalstages analyzed. Studies by electron microscopy (Kar-rer, 1959) have reported that, in mice, the entire tunicamedia of the pulmonary veins is of the striated typeand that SM cells are rarely present, if at all. In con-trast, we did find a widely distributed SM component,as provided by fluorescent microscopy analysis andconfocal image elaboration. Thus, the histologic struc-ture of the pulmonary veins in mice is similar to that ofrats (Ludatscher, 1968), with the exception that inmice SM cells are less abundant and more loosely con-nected. SM cells of the tunica media express both earlyas well as late markers of differentiation, such asa-SMA (Owens, 1995) and SMM (Miano et al., 1994),and are, in this respect, indistinguishable from SMcells of the arterial network or the bronchial plexus.
Development of Pulmonary Veins
The second part of this study deals with the devel-opment of the pulmonary veins. We show here that thedifferentiation of the venous and arterial systems inthe lungs proceeds in parallel, at least from stage E13onward. Therefore, the assumption that the venoussystem differentiates later than the arterial system(Woodcock-Mitchell et al., 1993) does not seem to ap-ply, at least in mice, to the pulmonary vascular net-work. A possible explanation for the contradictory re-sults on this matter is that in the pulmonary veins thecardiac and not the SM compartment is the major
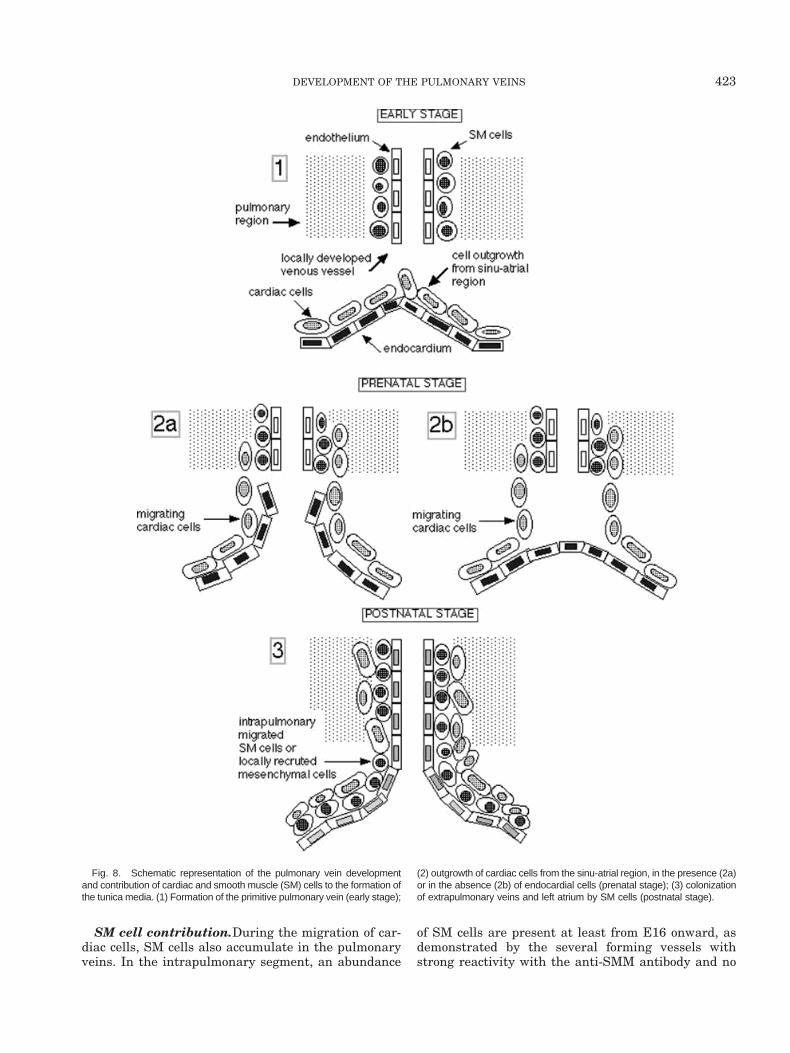
component of the tunica media. Cardiac cells accumu-late according to a caudocranial gradient, encircle thevessel around the endothelium, and within 2 days pop-ulate up to the third bifurcation of the pulmonaryveins. In our transgenic lines, LacZ positive cells werefirst detected in E12.5 embryos in the proximity of theleft atrium and in E13.5 embryos in the intrapulmo-nary region. Thus, it is during this narrow window oftime that the connection between the heart and thelung takes places. We hypothesize that two indepen-dent waves of cell growth generate the pulmonaryveins (Fig. 8). One wave starts from the pulmonaryparenchyma and consists of endothelial and SM cellsthat generate a primitive venous network in situ. Theother wave is the outgrowth of cardiac cells from thesinu-atrial region of the heart. Cardiac cells, perhapssupported by the endocardium, migrate toward thelungs, penetrate the parenchyma, and establish closecontacts with the primitive venous network. As a con-sequence of this process, the tunica media becomesenriched by cardiac cells that migrate along with thevessels up to the small branches. Around birth, SMcells, either as intrapulmonary migrating cells or aslocally recruited cells, populate the extrapulmonaryveins and progressively reach the left atrial wall.
Cardiac cell contribution.The precise cardiac re-gion from which the pulmonary veins originate is still amatter of debate, particularly because expression ofspecific molecular markers such as a-MHC andHNK-1, have provided contradictory results (Jones etal., 1994; DeRuiter et al., 1995; Tasaka et al., 1996). Ithas been suggested that in mice, as in human embryos(Los, 1968, 1978), the pulmonary veins arise from thesinus venosus and subsequently become incorporatedinto the left atrium (DeRuiter et al., 1995). We cannotbe conclusive on this matter because, in our study, itseems that the orifice of the primitive pulmonary trunkis located just at the border between the atrium and thesinus venosus at stage E12.5. A transgenic line withdifferential expression in atrial wall versus sinus ve-nosus could be helpful to clarify this point. Neverthe-less cell lineage experiments remain the approach ofchoice in this field, particularly because it seems thatthe sinus venosus does not only contribute to the sinusvenarum of the right atrium but also to the left atrium.Similarly, cell lineage studies will be necessary to es-tablish whether cardiac cell outgrowth is, at least inpart, supported by the endocardium or if it simplyfollows endothelial cells deriving as an outgrowth fromthe splanchnic plexus, as suggested by some authors(DeRuiter et al., 1997).
The expanding movement of cardiac cells in the pulmo-nary veins seems to be a predetermined phenomenon.LacZ-positive cells never go beyond well-defined bound-aries corresponding to the third bifurcation and neverreach the capillary plexus, whereas SM cells are moreabundant in the distal ends of the vessels. This selectivedistribution according to a caudocranial gradient hasbeen also described in the avian coronary veins
TABLE 1. SM Cell Distribution in the Tunica Media ofthe Murine Pulmonary Veins, as Revealed by
Expression of SMM Marker1
Stage Intrapulmonary vessels Extrapulmonary vesselsE13.5 n.i. 2E16 11 2E18–19 11 1/2N 111 111Adult 111 111
1E, embryonic day; N, 1-day old neonatal; n.i., not identified
421DEVELOPMENT OF THE PULMONARY VEINS

(Vrancken Peeters et al., 1997) where the proximal partof the media contains cardiac cells of atrial origin. Themedia located farther away appears to contain a mixedpopulation of cardiac and SM cells, the latter originatingfrom the epicardium (Vrancken Peeters et al., 1999). Asecond likewise intriguing property of migrating cardiaccells is the directional movement toward the venous butnot the arterial pole. It is likely, therefore, that endothe-lial cells, SM cells, or both, are a source of “signals” for the
recruitment of the cardiac cells. This finding would sug-gest that SM cells, endothelial cells, or both, from theveins differ from those of the arteries and that cardiaccells recognize their future position based upon thesedifferences. Possible candidate in this process might bethe ephrin/ephrin ligand system, which is known to bespecifically distributed in endothelial cells of arteries andveins from early stages of blood vessel formation. (Wanget al., 1998).
Fig. 7. Connection of the pulmonary veins to the left atrium. Theregion of the left atrium where the pulmonary veins become incorporatedwas examined by using the anti–a-SMA antibody as marker of smoothmuscle (SM) cells. Hearts from embryonic day (E) 16 embryos (A,B ) and1-day-old neonatal mice (C,D) were sectioned and examined by immu-nocytochemistry. Note that, in the E16 embryos, no reaction was found in
the left atrium (A), whereas positive cells were found in the right atrium(B). Arrowheads indicate the atrial regions populated by SM cells. Aster-isks in A and B indicate background reaction in the red blood cells.Overall pink staining of the sections is a result of counterstaining withMayer’s Hematoxylin. pv, pulmonary veins; ra, right atrium; la, left atrium.Scale bar in D 5 280 mm in A,B,C, 110 mm in D.
422 MILLINO ET AL.
SM cell contribution.During the migration of car-diac cells, SM cells also accumulate in the pulmonaryveins. In the intrapulmonary segment, an abundance
of SM cells are present at least from E16 onward, asdemonstrated by the several forming vessels withstrong reactivity with the anti-SMM antibody and no
Fig. 8. Schematic representation of the pulmonary vein developmentand contribution of cardiac and smooth muscle (SM) cells to the formation ofthe tunica media. (1) Formation of the primitive pulmonary vein (early stage);
(2) outgrowth of cardiac cells from the sinu-atrial region, in the presence (2a)or in the absence (2b) of endocardial cells (prenatal stage); (3) colonizationof extrapulmonary veins and left atrium by SM cells (postnatal stage).
423DEVELOPMENT OF THE PULMONARY VEINS

LacZ staining. Concerning the origin of SM cells in thisregion, it is generally assumed that they are recruitedlocally from the surrounding mesenchyme (Woodcock-Mitchell et al., 1993). Therefore, it is unlikely that SMcells of the pulmonary veins originate from the endo-thelium by a transdifferentiation process, as describedin other systems (DeRuiter et al., 1997). In fact, evenwith extensive confocal image analysis performed bothin the adult and in the embryonic stages, we neverobserved coexpression of endothelial and SM markerswithin the same cell. A conclusive demonstration of theembryologic origin of SM cells of the pulmonary veinswill require the identification of early markers of com-mitment and cell lineage tracing experiments.
In the extrapulmonary veins, SM cells accumulatesignificantly only around birth and may derive eitherfrom the migration from the pulmonary parenchyma orfrom the differentiation of local mesenchymal cells. Atthe same stage, an abundance of SM cells also reachesthe atrium. This finding may result from a partialincorporation of the pulmonary veins and an extensiveremodelling of the atrial wall, although no change inthe number of the pulmonary orifices was observedduring transition from late fetal stage (E20) to 1- to3-day-old neonatal mice (S. Ausoni, unpublished obser-vation). The putative mechanisms regulating the accu-mulation of SM cells in the extrapulmonary veinsmight be related to the dramatic functional changes inpulmonary circulation from the fetal to the postnatalpattern. Importantly, around birth, modifications ei-ther in the growth pattern or in the migration ability ofSM cells occur in two other vessels: the pulmonaryarteries (Stenmark and Mecham, 1997) and the ductusarteriosus (Slomp et al., 1997). To understand thesemorphologic events in detail, it will be helpful to clarifywhether SM cells possess peculiar responsivness to-ward factors released or available around birth.
EXPERIMENTAL PROCEDURESTransgenic Animals
By using promoter regions -230/1126 and -4000/1126, we generated cTNI transgenes. Line LacZ21 (DiLisi et al., 1998) and Line LacZ27 contained the shortpromoter region. Line LacZ60, containing the long pro-moter, was generated by insertion of the EcoRI/EcoNI(-4000/11) fragment into the -230/1126LacZ constructafter removal of the Xba/EcoNI (-230/11) segment.Transgenic mice with the long promoter region weregenerated by microinjection of the insert, purified fromvector sequences, into BDF2 (C57B1xDBA/2) one-cellfertilized eggs at a concentration of 500 copies/pl byusing standard techniques (Hogan et al., 1994). In-jected eggs were reimplanted into pseudopregnantBDF1 foster mothers. Transgenic animals were identi-fied by dot-blot and Southern blot analysis of genomicDNA (Sambrook et al., 1989) extracted from 3-week-oldmouse tail biopsies. The progeny of F1 and followinggenerations transgenic males crossed with normalBDF1 females were used for embryo analysis.
Tissue Preparation, Whole-Mount LacZ Stainingand Histologic Analysis
Embryos were dated taking 0.5 days postcoitum asthe day of vaginal plug. At the correct stages, they wereremoved from the uterus, washed in 13 phosphate-buffered saline (PBS), fixed in 4% p-formaldehyde for10 to 30 min and whole-mount stained with 5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside solution at37°C (Wilkinson, 1992) for a period ranging from 30min to several hours. For cryosections, embryos werefixed in 4% p-formaldehyde, treated in 30% sucrose inPBS overnight at 4°C, and embedded in OCT. Twelve-micrometer sections were stained for 15–30 min at37°C to detect b-galactosidase expression.
Immunocytochemistry
Antibodies used in this study were the following: ananti a-smooth actin monoclonal antibody directly con-jugated either with fluorescein isothiocyanate (FITC)(Sigma, clone.1A4) or with peroxidase (Dako, clone1A4); a polyclonal anti-SMM (Benzonana et al., 1988;Woodcock-Mitchell et al., 1993) and a rat monoclonalanti-mouse platelet endothelial cell adhesion molecule(PECAM) antibody (MEC 13.3, Pharmingen).
The anti-PECAM antibody was used according to theprocedure described by Vittet et al., (1997); the anti-SMM antiserum was diluted 1:2 and 1:3, as recom-mended. All the reactions were revealed with second-ary antibodies coupled with rhodamine isothiocyanate(TRITC) or peroxidase (Dako) after setting the appro-priate dilution. Sections were counterstained withMayer’s Hematoxylin, cover-slipped with Elvanol or100% glycerol, and photographed with either a LeitzOrthoplan or a Zeiss Axioplan microscope equippedwith darkfield and phase contrast optics. Immuno-staining was recorded and indicated on a scale of -, 1/-,11, 111, representing no, poor, strong, and intensereactivity, respectively.
Confocal microscopy was carried out on a Bio-RadMRC 1024 laser scanning confocal microscope (Bio-RadLaboratories) equipped with a Krypton/Argon laser. Toreduce bleed-through, double-label confocal images(both XY and XZ sections) were acquired sequentially.Noise reduction was achieved by “Kalman filtering”during acquisition. Micrographs were taken by usingeither a Focus Imagecorder Plus (Focus Graphics, Inc.)on Kodak film or a Professional Color Point II dyesublimation printer (Seiko).
In Situ Hybridization
In situ hybridization on cryosections was performedby using the rat cTNI antisense probe, (Ausoni et al.,1991). The procedure for preparing sense and antisense[35S]UTP probes has been previously described (Gorzaet al., 1993). In situ hybridization was performed oncryosections from adult samples according to Ausoni etal. (1991), with minor modifications. Sections were hy-bridized overnight at 52°C by using 2.5 3 106 cpm.
424 MILLINO ET AL.

Washing was done at 65°C in 50% formamide, 23 SSCand 100 mM dithiothreitol. Slides were processed forautoradiography by using Kodak-NTB-2 emulsion andexposed for 7 days. Microscope examination was per-formed with a Zeiss Axioplan microscope.
ACKNOWLEDGMENTS
We are grateful to Prof. Gabbiani and Dr. Benzonanafor the generous gift of the anti-SMM antibody and toDr. Elisabetta Ferrero for the anti-PECAM antibody.We thank Prof. S. Schiaffino for critical reading of themanuscript and Mrs. Sarah Hansen for editorial assis-tance. S.A. was funded by grants from Telethon,MURST, and Regione Veneto 1998.
NOTE ADDED IN PROOF
While this work was under submission, a scanningelectron microscopy and immunocytochemical study inrat embryos was published (Wenink AC, Symersky P,Ikeda T, DeRuiter MC, Poelmann RE, Gittenberger-de Groot AC. 2000. HNK-1 expression patterns in theembryonic rat heart distinguish between sinuatrial tis-sues and atrial myocardium. Anat Embryol 201:39–50)supporting further the hypothesis that in mammals, aspreviously shown in birds, the pulmonary veins draininto the sinus venosus.
REFERENCES
Ausoni S, De Nardi C, Moretti P, Gorza L, Schiaffino S. 1991. Devel-opmental expression of rat cardiac troponin I mRNA. Development112:1041–1051.
Benzonana G, Skalli O, Gabbiani G. 1988. Correlation between thedistribution of smooth muscle or nonmuscle myosins and a-smoothmuscle actin in normal and pathological soft tissues. Cell MotilCytoskeleton 11:260–274.
Burch GE, Romney RB. 1954. Functional anatomy and “throttlevalve” action of the pulmonary veins. Am Heart J 47:58–66.
DeRuiter MC, Gittenberger-de Groot AC, Wenink AC, Poelmann RE,Mentink MM. 1995. In normal development pulmonary veins areconnected to the sinus venosus segment in the left atrium. Anat Rec243:84–92
DeRuiter MC, Poelmann RE, VanMunsteren JC, Mironov V, Mark-wald RR, Gittenberger-de Groot AC. 1997. Embryonic endothelialcells transdifferentiate into mesenchymal cells expressing smoothmuscle actins in vivo and in vitro. Circ Res 80:444–451.
Di Lisi R, Millino C, Calabria E, Altruda F, Schiaffino S, Ausoni S.1998. Combinatorial cis-acting elements control tissue-specific ac-tivation of the cardiac troponin I gene in vitro and in vivo. J BiolChem 273:25371–25380.
Favaro G. 1910. Contributi all’istologia umana e comparata dei vasipolmonari. Int Monatschr Anat Physiol 27:375.
Gorza L, Ausoni S, Merciai N, Hastings KE, Schiaffino S. 1993.Regional differences in troponin I isoform switching during ratheart development. Dev Biol 156:253–264.
Hogan B, Beddington R, Costantini F, Lacy E. 1994. Manipulating themouse embryo: a laboratory manual. 2nd ed., Cold Spring Harbor,NY: Cold Spring Harbor Laboratory.
Jones WK, Sanchez A, Robbins J. 1994. Murine pulmonary myocar-dium: developmental analysis of cardiac gene expression. Dev Dyn200:117–128.
Karrer HE. 1959. The striated musculature of blood vessels. I. Gen-eral cell morphology. J Biophys Biochem Cytol 6:383–390.
Kramer AW, Marks LS. 1965. The occurrence of cardiac muscle in thepulmonary veins of rodents. J Morphol 117:135–150.
Larsen WJ. 1997. Human embryology. 2nd ed. New York: ChurchillLivingstone.
Laudatscher RM. 1968. Fine structure of the muscular wall of ratpulmonary veins. J Anat 103:345–357.
Lyons G, Schiaffino S, Sassoon D, Barton P, Buckingham M. 1990.Developmental regulation of myosin gene expression in mouse car-diac muscle. J Cell Biol 111:2427–2436.
Los JA. 1968. Embryology. In: Watson H, editor. Pediatric cardiology.London: Lloyd-Luke, Ldt.
Los JA. 1978. Cardiac septation and development of the aorta, pul-monary trunk, and pulmonary veins: previous work in the light ofrecent observations. Birth Defects Orig Artic Scr 14:109–138.
Miano JM, Cserjesi P, Ligon KL, Periasamy M, Olson EN. 1994.Smooth muscle myosin heavy chain exclusively marks the smoothmuscle lineage during mouse embryogenesis. Circ Res 75:803–812.
Owens GK 1995. Regulation and differentiation of vascular smoothmuscle cells. Physiol Rev 75:487–517.
Paes de Almeida O, Bohm GM, de Paula Carvalho M, Paes de Car-valho A. 1975. The cardiac muscle in the pulmonary vein of the rat:a morphological and electrophysiological study. J Morphol 145:409–433.
Rauschel F. 1836. De arteriarum et venaram structura. Breslau.Risau W, Lemmon V. 1988. Changes in the vascular extracellular
matrix during embryonic development. Dev Biol 125:441–450.Risau W. 1997. Mechanisms of angiogenesis. Nature 386:671–674.Rosenquist TH, Beall AC. 1990. Elastogenic cells in the developing
cardiovascular system: smooth muscle, nonmuscle and cardiac neu-ral crest. Ann N Y Acad Sci 558:106–119.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning: a lab-oratory manual, 2nd ed. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory.
Slomp J, Gittenberger-de Groot AC, Glukhova MA, Conny van Mun-steren J, Kockx MM, Schwartz SM, Koteliansky VE. 1997. Differ-entiation, dedifferentiation, and apoptosis of smooth muscle cellsduring the development of the human ductus arteriosus. Artererio-scler Thromb Vasc Biol 17:1003–1009.
Stenmark KR, Mecham RP. 1997. Cellular and molecular mecha-nisms of pulmonary vascular remodeling. Annu Rev Physiol 59:89–144.
Tasaka H, Krug EL, Markwald RR. 1996. Origin of the pulmonaryvenous orifice in the mouse and its relation to the morphogenesis ofthe sinus venosus, extracardiac mesenchyme (spina vestibuli), andatrium. Anat Rec 246:107–113.
Vittet D, Buchou T, Schweitzer A, Dejana E, Huber P. 1997. Targetednull-mutation in the vascular endothelial-cadherin gene impairsthe organization of vascular-like structures in embryoid bodies.Proc Natl Acad Sci USA 94:6273–6278.
Vrancken Peeters MP, Gittenberger-de Groot AC, Mentink MM, Hun-gerford JE, Little CD, Poelmann RE. 1997. The development of thecoronary vessels and their differentiation into arteries and veins inthe embryonic quail heart. Dev Dyn 208:338–348.
Vrancken Peeters MP, Gittenberger-de Groot AC, Mentink MM.,Poelman RE. 1999. Smooth muscle cells and fibroblasts of thecoronary arteries derive from epithelial-mesenchymal transforma-tion of the epicardium. Anat Embryol (Berl) 199:367–378.
Wang HU, Chen ZF, Anderson DJ. 1998. Molecular distinction andangiogenic interaction between embryonic arteries and veins re-vealed by ephrin-B2 and its receptor Eph-B4. Cell 93:741–753.
Wilkinson DG. 1992. In situ hybridization: a practical approach. In:Wilkinson DG, editor. Oxford: IRL Press. p 75–82.
Woodcock-Mitchell J, White S, Stirewalt W, Periasamy M, Mitchell J,Low RB. 1993. Myosin isoform expression in developing and remod-eling rat lung. Am J Respir Cell Mol Biol 8:617–625.
425DEVELOPMENT OF THE PULMONARY VEINS