cadherin-8 and n-cadherin differentially regulate pre- and postsynaptic development of the...
TRANSCRIPT

Cadherin-8 and N-cadherin Differentially Regulate Pre- andPostsynaptic Development of the Hippocampal Mossy Fiber Pathway
Iddil H. Bekirov, Vanja Nagy, Alexandra Svoronos,George W. Huntley, and Deanna L. Benson*
ABSTRACT: Cells sort into regions and groups in part by their selectivesurface expression of particular classic cadherins during development. Inthe nervous system, cadherin-based sorting can define axon tracts, restrictaxonal and dendritic arbors to particular regions or layers, and mayencode certain aspects of synapse specificity. The underlying model hasbeen that afferents and their targets hold in common the expression of aparticular cadherin, thereby providing a recognition code of homophiliccadherin binding. However, most neurons express multiple cadherins, andit is not clear whether multiple cadherins all act similarly in shaping neu-ral circuitry. Here we asked how two such cadherins, cadherin-8 and N-cadherin, influence the guidance and differentiation of hippocampal mossyfibers. Using organotypic hippocampal cultures, we find that cadherin-8regulates mossy fiber fasciculation and targeting, but has little effect onCA3 dendrites. In contrast, N-cadherin regulates mossy fiber fasciculation,but has little impact on axonal growth and targeting. However, N-cad-herin is essential for CA3 dendrite arborization. Both cadherins arerequired for formation of proper numbers of presynaptic terminals. Mech-anistically, such differential actions of these two cadherins could, intheory, reflect coupling to distinct intracellular binding partners. How-ever, we find that both cadherins bind b-catenin in dentate gyrus (DG).This suggests that cadherins may engage different intracellular signalingcascades downstream of b-catenin, coopt different extracellular bindingpartners, or target distinct subcellular domains. Together our findingsdemonstrate that cadherin-8 and N-cadherin are critical for generating themossy fiber pathway, but that each contributes differentially to afferentand target differentiation, thereby complementing one another in the as-sembly of a synaptic circuit. VVC 2007 Wiley-Liss, Inc.
KEY WORDS: axon guidance; synaptogenesis; DG; dendriticarborization; hippocampus
INTRODUCTION
Axons project accurately to their targets by sensing and responding to avariety of chemical cues. Diffusible and standing gradients of cues can
guide axons over relatively long distances, whileregionally restricted, more homogeneously distributedcues appear to more strongly influence guidance atintermediate ‘‘choice points’’ and to help stop growthwithin a target. Such regionally restricted cues caninfluence dendritic differentiation, highlighting thecollaboration that has to occur between ingrowingaxons and their simultaneously differentiating targets.
Classic cadherins show strict regional and laminardistribution patterns (Redies et al., 1993; Yamagataet al., 1995; Suzuki et al., 1997a; Huntley and Ben-son, 1999; Bekirov et al., 2002). Homophilic bindingbetween cadherins generates strong adhesion that sortscells into regions and groups early in development(Hatta and Takeichi, 1986; Espeseth et al., 1998;Price et al., 2002). As neurons differentiate and sendaxons to distant targets, it has been proposed thathomophilic cadherin adhesion between source and tar-get neurons may be important for ensuring accuracyin the development of connectivity. Cadherins canpromote axon extension (Neugebauer et al., 1988;Tomaselli et al., 1988; Friedlander et al., 1989; Bal-samo and Lilien, 1990; Drazba and Lemmon, 1990;Doherty et al., 1991; Shimoyama et al., 2000; Uttonet al., 2001; Andrews and Mastick, 2003; Marthienset al., 2005) and selective fasciculation (Wohrn et al.,1999; Treubert-Zimmermann et al., 2002), and canprovide short-range instructional cues (Andrews andMastick, 2003). Within a target, cadherins can influ-ence dendritic differentiation (Zhu and Luo, 2004).Additionally, cadherin binding across the synaptic cleftis essential for stabilizing young synapses (Togashiet al., 2002; Bozdagi et al., 2004; Elste and Benson,2006)—an action that would be expected to preventfurther extension. Consistent with this idea, N-cad-herin adhesion retains retinal-collicular and thalamo-cortical axons within their target layers (Inoue andSanes, 1997; Poskanzer et al., 2003), and in Drosoph-ila, appropriate photoreceptor-target interactionsrequire DN-cadherin (Lee et al., 2001). However,individual neurons express several cadherins, andmany of these are shared between source and target(Suzuki et al., 1997b; Miskevich et al., 1998; Wohrnet al., 1999; Bekirov et al., 2002; Gil et al., 2002). Itremains an unanswered question as to whether, andhow, such multiply expressed cadherins function inthe same pathway. The intracellular domains of classic
Fishberg Department of Neuroscience, Mount Sinai School of Medicine,New York, New YorkGrant sponsors: NINDS, NIMH, NIDA; Grant numbers: NS37731,NS34659, NS43083 (predoctoral training), MH075783, DA07135 (post-doctoral training); Grant sponsor: Irma T. Hirschl Career Scientist Award.Iddil H. Bekirov is currently at Section on Fundamental Neuroscience,NIMH, Bldg. 35/Rm1B-213, [35 Convent Dr., MSC 3728 (US mail)],Bethesda, MD 20892-3728, USA.*Correspondence to: Deanna L. Benson; Fishberg Department of Neuro-science, The Mount Sinai School of Medicine, 1425 Madison Avenue, Box1065, New York, NY 10029, USA. E-mail: [email protected] for publication 30 October 2007DOI 10.1002/hipo.20395Published online 6 December 2007 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 18:349–363 (2008)
VVC 2007 WILEY-LISS, INC.

cadherins are highly conserved and are presumed to engage thesame intracellular binding partners, making it difficult tounderstand how they might achieve a coherent functional out-put. Here, we sought to determine the individual actions ofcoexpressed cadherins on the generation of a well-defined ana-tomical pathway-the hippocampal mossy fiber pathway leadingfrom dentate gyrus (DG) to the CA3 region of hippocampus.Cadherin-8 and N-cadherin are two prototypical, classic cad-herins expressed early in development by DG granule neuronsand their target, area CA3 neurons (Redies and Takeichi, 1993;Korematsu and Redies, 1997; Bekirov, 2004). We find thatcadherin-8 and N-cadherin are both essential for the develop-ment of the mossy fiber projection. However, each cadherincontributes differently to establishing axon trajectory and targetregion, thus complementing one another in the formation ofthis synaptic circuit.
MATERIALS AND METHODS
Animals
Sprague Dawley rat pups were used for all of the experi-ments. The day of birth was designated as postnatal day (P) 0.The care and treatment of the rats adhered strictly to guidelinesestablished by both the National Institutes of Health andMount Sinai’s Institutional Animal Care and Use Committee.
Organotypic Hippocampal Cultures
Hippocampal slices were cultured on membranes asdescribed previously (Stoppini et al., 1991). After inducing an-esthesia by hypothermia, 5- to 6-day-old rat pups were decapi-tated and the hippocampus was dissected in sterile, ice cold D-(1)-glucose-enriched (6.5 g/L) Gey’s Balanced Salt Solution.Four-hundred micron thick slices were cut using a McIIwainTissue Chopper (Surrey, England), and those from the midsep-totemporal level of the hippocampus were transferred to mem-branes in culture inserts (0.4 lm pores; Millipore, MA) andplaced in 6 or 12-well plates containing a mixture of Eagle’sBasal Medium with Earle’s Salts (50%), Earle’s Balanced SaltSolution (25%), horse serum (25%), 50% D-(1)-glucose (1.0%),and 200 mM L-glutamine (0.5%). Slices were cultured at 378C,5% CO2. After 24 h, cultures were placed on a rocking platformfor the remainder of the culture period. A mixture of antimitoticinhibitors (1026 M of cytosine arabinoside, uridine, and fluoro-deoxyuridine, Sigma, St. Louis, MO) was added after 2 days, andmedia was changed every 2–3 days.
Cadherin Reagents
Synthetic peptides
The following peptide sequences, HLRAHAVDINGNQVEN(HAV), TLTAQAVDDWETNKPLE (QAV), and ARLQHDV-NANVHEING (scrambled control) were purchased from Mimo-
topes (Raleigh, NC). The HAV sequence was chosen because sev-eral studies have demonstrated that the tripeptide sequence, HAV,is contained within a region important for mediating adhesion(Blaschuk et al., 1990; Nose et al., 1990). Structural data demon-strate that at least four residues in the peptide sequence are partof the adhesive interface (Patel et al., 2006), and the flankingregions are sufficient to confer specificity (Blaschuk et al., 1990;Noe et al., 1999; Williams et al., 2000). The HAV motif in typeI cadherins is replaced with QAV in cadherin-8 and several othertype II cadherins (Shimoyama et al., 2000). Structural data indi-cate that the adhesive interface residues contained within thisregion are common to type I and type II cadherins (Patel et al.,2006). The flanking sequences correspond to rat cadherin-8(Williams et al., 2000). Cadherin peptides or a scrambled controlpeptide (SCR) were added to culture media starting at 0 days invitro (div) at a concentration of 200 lg/mL unless indicated oth-erwise (Doherty et al., 1991). Peptide was replenished whenmedia was changed, and slices were fixed after 5 days in cultureby immersion in 4% paraformaldehyde in phosphate bufferedsaline (PBS). Effects on the mossy fiber pathway were evaluatedusing DiI labeling or immunostaining as described later.
Cadherin-Fc chimeras
A cadherin-8-Fc chimeric DNA construct was generated byjoining the extracellular domain spanning EC1 through EC4 ofrat cadherin-8 to the Fc region of human IgG. The extracellu-lar domain of cadherin-8 was generated by PCR from pBS SKcontaining full-length cadherin-8 [a kind gift from Dr. S.T.Suzuki, Kido et al., 1998)] as a 1,657 bp XbaI-NotI restrictionfragment. This fragment was then inserted into a modifiedpCR3.1 vector [a kind gift from Dr. Martin Grumet, (Haspelet al., 2001)] cut with XbaI and NotI. The DNA was trans-formed into Top10F’ E. coli cells (Invitrogen, Carlsbad, CA).Plasmid DNA was purified from colonies using a standard plas-mid isolation protocol (Qiagen, Valencia, CA) and presence ofinsert was verified by restriction analysis and sequencing. Cad-herin-8-Fc plasmid DNA was transiently transfected in COS-7cells using Polyfect (Qiagen, Valencia, CA), and after 5 daysthe supernatant was collected and concentrated. N-cadherin-Fcwas similarly isolated from either HEK 293 cells that were sta-bly expressing N-cadherin-Fc (a gift from Dr. Takeshi Sakurai)or COS-7 cells that were transiently transfected. The identityand size of both Fc proteins was verified by Western blotting(data not shown). The Fc modification at the C-terminus doesnot affect adhesive function (Chappuis-Flament et al., 2001).
Immunohistochemistry
Cultures were fixed as described earlier and immunolabeledfor either MAP2 [monoclonal antibody AP-14; (Stearns andBinder, 1987)], b-III tubulin (monoclonal; Covance, Berkeley,CA) or SV2 (Developmental Studies Hybridoma Bank). Anti-body binding was visualized by incubation in fluorescently con-jugated secondary antibodies (Southern Biotech, Birmingham,AL). Laminar and regional borders were confirmed with thefluorescent Nissl stain TOTO-3 iodide (Molecular Probes,
350 BEKIROV ET AL.
Hippocampus

Eugene, OR). Images were acquired using a Zeiss LSM 510META confocal microscope (Zeiss, Thornwood, NY) with Ar-gon and k633 He-Ne lasers. To assay cell death, slice cultureswere maintained in a CO2 incubator for 30 min in culturemedia containing 1 lM of the membrane-impermeant dead-cell marker Sytox Green (Molecular Probes, Eugene, OR). Fluo-rescence of living slices was visualized directly through the cul-ture plate using an inverted fluorescence microscope (NikonDiaphot) and imaged using the same exposure parameters witha Hamamatsu ORCA-ER digital camera. Fluorescence intensitywas quantified in areas CA3 and CA1 using MetaMorph (Uni-versal Imaging, Dowlington, PA). To quantify SV2 labeling,images were acquired using a 1003/1.5 N.A. oil immersionobjective and a zoom factor of 2 on a Zeiss LSM 410 confocalmicroscope (Zeiss, Thornwood, NY) with an Ar/Kr laser andfilters k515–540 and k670–810. Twelve confocal images perslice (four images/lamina) were taken in strata lucidum, pyra-midale, and oriens and at least five slices were imaged per treat-ment condition. All images were obtained using the samebrightness and contrast settings. Puncta number, area, and in-tensity were determined using MetaMorph, and the data wereexported to Excel. Differences between groups were assessedusing unpaired t-tests; alpha was corrected for the parameterbeing compared and the peptide used (3 groups) (JMP soft-ware, SAS Institute, Cary, NC), and the data were graphed inExcel.
DiI Labeling
To assess mossy fiber trajectories in organotypic slices apulled glass capillary filament was used to place small crystalsof DiI (DiIC18(3); Molecular Probes, Eugene, OR) taken froma 0.5% DiI solution diluted in dimethylformamide accordingto previously described procedures (Poskanzer et al., 2003).Slice cultures were fixed for 12 h in 4% paraformaldehyde andcrystals were deposited under microscope guidance into thehilar region, where fibers converge as they exit the DG. Sliceswere then placed in PBS containing 0.2% sodium azide andincubated at 378C in the dark for 2–4 days. Labeling wasvisualized by using a Zeiss LSM 410 and a 403/0.9 N.A. oilobjective and photomontages were generated using Adobe Pho-toshop (Mountain View, CA). Maps of the trajectories weredrawn using a Zeiss Axiophot equipped with Neurolucida(MicroBrightField, Colchester, VT).
Dissociated Granule Cell Cutures
Postnatal day 6 (P6) rat pups were deeply anesthetized byhypothermia, and the hippocampi were dissected. A cut wasmade along the hippocampal fissure and the DG was excised.Dentate tissue was placed in balanced salt solution (BSS: 13Hank’s BSS; 0.3 M HEPES, pH 5 7.3; 13 penicillin/strepto-mycin), cut into small pieces and incubated in 0.25% trypsinand 0.1% DNase I at 378C for 30 min. Dissociated cells werefiltered through nylon mesh into a sterile conical tube contain-ing 3 mL 10% horse serum, pelleted, and resuspended in BSS.Sixty-three hundred granule cells per well were plated in 12-
well cell culture plates (Costar, Corning, Corning, NY)containing Neurobasal medium with 0.4 mM L-glutamine andB-27 supplement (Gibco-Invitrogen Corp., Carlsbad, CA) onglass coverslips coated with N-cadherin-Fc, cadherin-8 Fc, orhaving a confluent layer of L cells, N-cadherin L cells or cad-herin-8 L cells. To coat coverslips with Fc fusion proteins,poly-L-lysine coated coverslips were incubated with antihumanIgG antibody (Fc-specific monoclonal; Sigma, St. Louis, MO),followed by incubation with cadherin-8-Fc or N-cadherin-Fc.L-cells were plated directly on glass. Three hours after plating,synthetic peptides (QAV, HAV, or scrambled peptide) wereadded at a final concentration of 200 lg/mL (unless noted oth-erwise). Cultures were incubated overnight at 378C/5% CO2,rinsed in 0.01 M PBS and then fixed in 4% paraformaldehyde.Neurons were visualized under phase optics as well as fluores-cence microscopy using a Zeiss Axiophot microscope with a403/0.75 N.A. objective. Neurite lengths were traced usingNeurolucida software and analyzed using NeuroExplorer. Allindividual tree length data was imported into JMP for statisti-cal analysis. Means were compared using unpaired Student’s t-test or analysis of variance (ANOVA) and a post test (level ofsignificance, P < 0.05). P-values and details are provided inthe figure legends. Results were then graphed using Prism(GraphPad software).
Coimmunoprecipitations
Dentate gyri from postnatal day 6 rat pups were isolated andhomogenized in extraction buffer [1% digitonin in HEPESbuffer (25 mM HEPES, pH 7.4; 150 mM NaCl)] containing13 Complete protease inhibitor cocktail (Roche, Indianapolis,IN). One milligram of protein was preabsorbed with Protein GAgarose (Invitrogen) for 2 h at 48C. The spun-down superna-tant was incubated overnight at 48C with N-cadherin (BDTransduction Labs, San Jose, CA) or cadherin-8 (Santa CruzBiotechnology, Santa Cruz, CA). After adding beads, sampleswere incubated for 2 h at 48C. All IPs were washed twice in1% digitonin/HEPES, once in HEPES, and then analyzed by10% SDS-PAGE followed by Western blotting with anti-b-cat-enin antibody (monoclonal; BD Transduction Labs). Labelingwas visualized using ECL (Pierce, Rockland, IL).
RESULTS
Classic cadherins generally bind homophilically. In the firstpart of this study, we exploited this property to assess func-tional activity of N-cadherin and cadherin-8 in granule cellaxon outgrowth by culturing dissociated granule cells on cad-herin substrates. Additionally, this strategy also allowed us todetermine the effectiveness and specificity of peptide blockingreagents in a controlled environment. However, the develop-ment of connectivity in vivo occurs in a more complex, molec-ular environment. Thus, in the second part of this study, weassessed outgrowth and synapse formation in organotypic hip-
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 351
Hippocampus

pocampal slice cultures in order to determine the actions ofcadherins in the assembly of a circuit in situ.
N-Cadherin, But Not Cadherin-8, PromotesMossy Fiber Extension On Cadherin Substrates
DG granule cells express both N-cadherin and cadherin-8 dur-ing the first postnatal weeks when their axons are extending rap-idly (Amaral and Dent, 1981; Korematsu and Redies, 1997;Bekirov et al., 2002). To determine whether N-cadherin and cad-herin-8 are equally capable of promoting mossy fiber outgrowth,dentate granule neurons were dissociated at P5 or P6, plated onimmobilized N-cadherin or cadherin-8 substrates, and after 24 h,were fixed and immunostained for bIII-tubulin to label neuronalprocesses. Labeled neurites were traced using Neurolucida, and
axons were defined conventionally as processes that exceed theothers in length by at least 5 lm (Goslin and Banker, 1989b).At this stage of development, minor processes are commonlyquite short (20 lm or less) and the differences between thelengths of minor processes and axons are usually tens of microns(Goslin and Banker, 1989a; Goslin et al., 1990; Esch et al.,1999) (Fig. 1). On an N-cadherin substrate, the average axonlength is 90.7 lm (65.7), while when grown on cadherin-8,axons are on average 34% shorter (60.1 6 5) lm (Figs. 1A,Cand 2A). The reduced axon outgrowth on cadherin-8 did notresult from poor attachment, since cell counts indicate that simi-lar numbers of neurons attached to cadherin-8 and N-cadherinsubstrates (data not shown). Additionally, all of the neuritesappear to firmly adhere to cadherin-8 and do not float free.Thus, while both cadherin-8 and N-cadherin support cell binding
FIGURE 1. N-cadherin and cadherin-8 differentially promoteoutgrowth. Representative examples of Neurolucida drawings (A–D) or photomicrographs (E–G) of DG granule cells that were dis-sociated and plated on cadherin-Fc substrates (A–D) or cadherin-expressing L cells (E–G). The longest process was defined as theaxon according to convention (see text). N-cadherin promotes axo-nal growth (A) and this is reduced by HAV peptides (B). In con-
trast, neurons grown on cadherin-8 substrate have shorter axons(C) in comparison with those grown on N-cadherin, and axonsgrow longer in the presence of QAV peptides (D). On L cells, cad-herin-8-mediated outgrowth (G) is similar to L-cells alone (E), butaxons are shorter than those on N-cadherin L cells (F). Magnifica-tion bars 5 10 lm. Ncad, N-cadherin; cad8, cadherin-8.
352 BEKIROV ET AL.
Hippocampus

and axon extension, it is possible that cadherin-8 binding may bemore strongly adhesive than N-cadherin, attenuating the rateaxon extension. Alternatively, its actions may be modified by acoreceptor or by the differential activation of a signaling pathway(e.g., Rhee et al., 2002).
Although our data indicate that N-cadherin is a better sub-strate for granule cell axon extension than cadherin-8, it isunclear whether such rates of extension are faster or slower inconjunction with those on other substrates. To fully understandthe effects of these cadherins on outgrowth, we cultured disso-ciated granule cells on a more complex, cellular substrate. Wecompared axon extension on mouse L-cells [fibroblasts thatexpress fibronectin (Toda et al., 1994), but no native cadherins(Nose et al., 1988)] with that on L-cells stably expressing N-cadherin or cadherin-8. Axons growing on N-cadherin L-cellsare significantly longer than those on untransfected L-cells(Figs. 1E,F and 2B), while axons extending on cadherin-8 L-cells are slightly, but not significantly shorter than those grow-ing on untransfected L-cells (Figs. 1E,G and 2B).
If axon extension on cadherin-8 substrates reflects a strongeradhesive interaction, we would expect that increasing concentra-tions of cadherin-8 would decrease growth. However, when gran-ule cells were cultured on cadherin-8 concentrations ranging from1 to 500 lg/mL, axon length gradually, albeit modestly, increasedup to 100 lg/mL and decreased at 500 lg/mL (Fig. 2C). Thus,cadherin-8 supports slow mossy fiber extension at low concentra-tions and less extension at high concentrations, contrary to theaccelerated rates seen with other classic cadherins (Fig. 2A).
Peptides Interfere With Cadherin-DependentAxon Extension On Cadherin Substrates
The EC1 domain of classical cadherins is critical for mediatinghomophilic binding specificity (Nose et al., 1990) and adhesion
(Nose et al., 1990; Shapiro et al., 1995; Patel et al., 2006). A His-Ala-Val (HAV) sequence within EC1 is common to Type I classicalcadherins like N-cadherin, and several lines of experimental datashow that exogenous peptides containing this sequence flanked byamino acids specific for N-cadherin are highly effective at blockingN-cadherin mediated neurite outgrowth (Blaschuk et al., 1990;Williams et al., 2000). Consistent with this, we find that an HAVpeptide specifically inhibits granule cell axon extension on an N-cadherin substrate (Figs. 1B vs A and black bars in 3A).
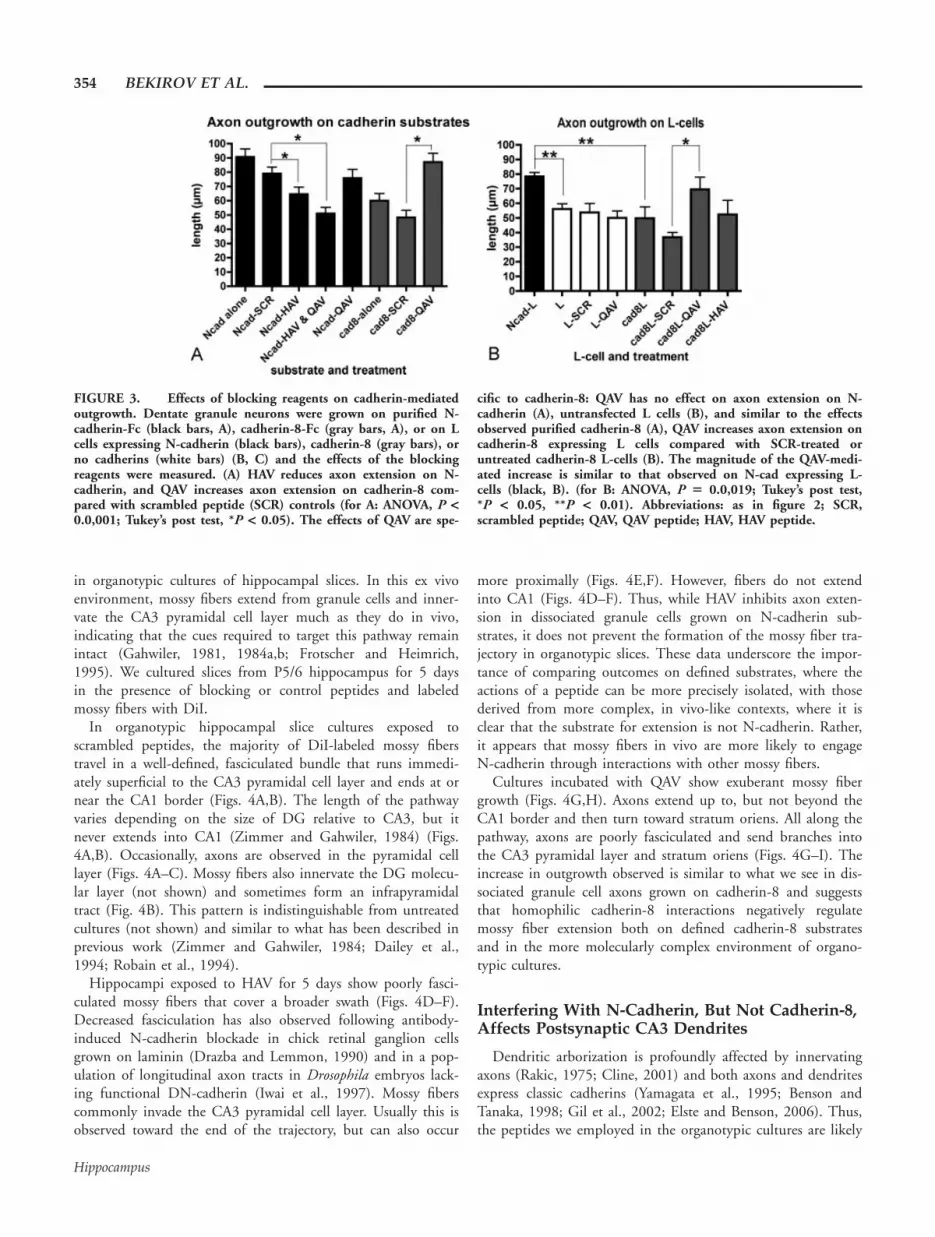
Type II cadherins-including cadherin-8-contain a glutamine (Q)in lieu of histadine in the homologous region. Thus, because HAVpeptides interfere with N-cadherin mediated outgrowth, wedesigned a QAV peptide that covers the homologous region incadherin-8, and similar to HAV peptides, covers several residuesinvolved in the cadherin-8 adhesive interface (Patel et al., 2006). Incontrast to the inhibitory effects on axon outgrowth observed withHAV peptides, we found that neurons exposed to the QAV peptideshow increased axon extension on both purified cadherin-8 sub-strates and cadherin-8 expressing L cells, suggesting that cadherin-8negatively regulates granule cell axon extension (Figs. 1D vs. C andgray bars in 3A,B). We verified that the QAV peptide does not sim-ply stimulate axon extension nonspecifically, since outgrowth onpurified N-cadherin or on untransfected L cells is not altered by ex-posure to QAV (black bars in Fig. 3A and white bars in 3B). Takentogether, these data indicate that regardless of whether extension ison purified cadherin substrates or on cadherin expressing L-cells,blocking N-cadherin with HAV peptide reduces axon growth whileblocking cadherin-8 with QAV peptide enhances axon growth.
N-Cadherin and Cadherin-8 Are Required forMossy Fiber Laminar and Regional Specificity
To determine the contributions of N-cadherin and cadherin-8 to mossy fiber targeting, we examined mossy fiber outgrowth
FIGURE 2. Quantitative differences between N-cadherin- andcadherin-8-mediated outgrowth. DG granule cells were dissociatedand plated on purified cadherin-Fc substrates (A, C), or on plainor transfected L-cells (B). (A) Total axon length for neurons grow-ing on cadherin-8 (gray) is significantly shorter than on N-cad-herin (black) (90.7 6 5.7 lm, n 5 40 vs. 60.1 6 5 lm, n 5 43,*P < 0.0,001, t-test). (B) On L-cells expressing cadherin-8 (gray),axonal growth is similar to untransfected L-cells (white) (49.9 66.1 lm, n 5 21 vs. 53.15 6 6 lm, n 5 40), while cells expressing
N-cadherin (black) support more outgrowth (78.4 6 6.1 lm, n 521) (ANOVA, P 5 0.023; Tukey’s post test, *P < 0.05, **P <0.002). Granule cells plated on a range of concentrations of cad-herin-8, show increased axon outgrowth up to 100 lg/mL(ANOVA, P 5 0.0,001; Tukey’s post test, *P < 0.05) (C). Ncad, N-cadherin-Fc substrate; cad-8, cadherin-8-Fc substrate; L, L cell;Ncad-L, N-cadherin expressing L cells; cad8-L, cadherin-8 express-ing L cells.
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 353
Hippocampus
in organotypic cultures of hippocampal slices. In this ex vivoenvironment, mossy fibers extend from granule cells and inner-vate the CA3 pyramidal cell layer much as they do in vivo,indicating that the cues required to target this pathway remainintact (Gahwiler, 1981, 1984a,b; Frotscher and Heimrich,1995). We cultured slices from P5/6 hippocampus for 5 daysin the presence of blocking or control peptides and labeledmossy fibers with DiI.
In organotypic hippocampal slice cultures exposed toscrambled peptides, the majority of DiI-labeled mossy fiberstravel in a well-defined, fasciculated bundle that runs immedi-ately superficial to the CA3 pyramidal cell layer and ends at ornear the CA1 border (Figs. 4A,B). The length of the pathwayvaries depending on the size of DG relative to CA3, but itnever extends into CA1 (Zimmer and Gahwiler, 1984) (Figs.4A,B). Occasionally, axons are observed in the pyramidal celllayer (Figs. 4A–C). Mossy fibers also innervate the DG molecu-lar layer (not shown) and sometimes form an infrapyramidaltract (Fig. 4B). This pattern is indistinguishable from untreatedcultures (not shown) and similar to what has been described inprevious work (Zimmer and Gahwiler, 1984; Dailey et al.,1994; Robain et al., 1994).
Hippocampi exposed to HAV for 5 days show poorly fasci-culated mossy fibers that cover a broader swath (Figs. 4D–F).Decreased fasciculation has also observed following antibody-induced N-cadherin blockade in chick retinal ganglion cellsgrown on laminin (Drazba and Lemmon, 1990) and in a pop-ulation of longitudinal axon tracts in Drosophila embryos lack-ing functional DN-cadherin (Iwai et al., 1997). Mossy fiberscommonly invade the CA3 pyramidal cell layer. Usually this isobserved toward the end of the trajectory, but can also occur
more proximally (Figs. 4E,F). However, fibers do not extendinto CA1 (Figs. 4D–F). Thus, while HAV inhibits axon exten-sion in dissociated granule cells grown on N-cadherin sub-strates, it does not prevent the formation of the mossy fiber tra-jectory in organotypic slices. These data underscore the impor-tance of comparing outcomes on defined substrates, where theactions of a peptide can be more precisely isolated, with thosederived from more complex, in vivo-like contexts, where it isclear that the substrate for extension is not N-cadherin. Rather,it appears that mossy fibers in vivo are more likely to engageN-cadherin through interactions with other mossy fibers.
Cultures incubated with QAV show exuberant mossy fibergrowth (Figs. 4G,H). Axons extend up to, but not beyond theCA1 border and then turn toward stratum oriens. All along thepathway, axons are poorly fasciculated and send branches intothe CA3 pyramidal layer and stratum oriens (Figs. 4G–I). Theincrease in outgrowth observed is similar to what we see in dis-sociated granule cell axons grown on cadherin-8 and suggeststhat homophilic cadherin-8 interactions negatively regulatemossy fiber extension both on defined cadherin-8 substratesand in the more molecularly complex environment of organo-typic cultures.
Interfering With N-Cadherin, But Not Cadherin-8,Affects Postsynaptic CA3 Dendrites
Dendritic arborization is profoundly affected by innervatingaxons (Rakic, 1975; Cline, 2001) and both axons and dendritesexpress classic cadherins (Yamagata et al., 1995; Benson andTanaka, 1998; Gil et al., 2002; Elste and Benson, 2006). Thus,the peptides we employed in the organotypic cultures are likely
FIGURE 3. Effects of blocking reagents on cadherin-mediatedoutgrowth. Dentate granule neurons were grown on purified N-cadherin-Fc (black bars, A), cadherin-8-Fc (gray bars, A), or on Lcells expressing N-cadherin (black bars), cadherin-8 (gray bars), orno cadherins (white bars) (B, C) and the effects of the blockingreagents were measured. (A) HAV reduces axon extension on N-cadherin, and QAV increases axon extension on cadherin-8 com-pared with scrambled peptide (SCR) controls (for A: ANOVA, P <0.0,001; Tukey’s post test, *P < 0.05). The effects of QAV are spe-
cific to cadherin-8: QAV has no effect on axon extension on N-cadherin (A), untransfected L cells (B), and similar to the effectsobserved purified cadherin-8 (A), QAV increases axon extension oncadherin-8 expressing L cells compared with SCR-treated oruntreated cadherin-8 L-cells (B). The magnitude of the QAV-medi-ated increase is similar to that observed on N-cad expressing L-cells (black, B). (for B: ANOVA, P 5 0.0,019; Tukey’s post test,*P < 0.05, **P < 0.01). Abbreviations: as in figure 2; SCR,scrambled peptide; QAV, QAV peptide; HAV, HAV peptide.
354 BEKIROV ET AL.
Hippocampus

to have had effects on postsynaptic target dendrites as well ason presynaptic axons. To assess dendritic differentiation in CA3in situ, hippocampal slices that were cultured in the presenceor absence of the cadherin peptides were immunostained forMAP2, a dendrite-specific marker. In comparison withuntreated slices (Fig. 5A), those exposed for 5 days to HAV(Fig. 5B) exhibited little MAP2-labeling in CA3 dendrites com-
pared with controls or to CA1 and the DG (Figs. 5A,B), butNissl-stained cell bodies were still detected in the pyramidalcell layer (Fig. 5C).
The HAV-induced loss of MAP2 staining could reflect sev-eral possibilities: HAV treatment could prevent dendrite out-growth, promote dendrite retraction, or initiate neuronal celldeath. The decreased labeling could also reflect a selective loss
FIGURE 4. Cadherin-8 and N-cadherin differentially affectmossy fiber outgrowth in situ. Confocal images and Neurolucidagenerated maps of DiI-labeled mossy fibers in P5/P6 hippocampalorganotypic slices that were cultured for 5 days. Low-magnifica-tion images are shown at left with higher magnification images ofstratum lucidum as insets (A, D, G). Maps of additional culturesunder similar conditions are shown in the middle column (B, E,H), and high-magnification images focused on the pyramidal celllayer (sp) are shown in the right hand column (C, F, I). Culturestreated with scrambled peptides (SCR) show hippocampal mossyfibers concentrated in stratum lucidum (sl) with only a few fibersinvading the pyramidal cell layer (sp, A–C). Higher magnificationinset of a more heavily labeled preparation more clearly reveals thetight, parallel organization of the fibers. Dotted lines delineate
layers, arrows indicate the CA3/CA1 border, and the injection sitein the dentate hilus is just out of the field of view in the upperleft (A). In the maps (B, E, H) a black dot indicates the injectionsite and the dotted circle, the intense glow surrounding the site.The principal cell layers are indicated by thick lines. Culturesincubated with HAV (D–F) show mossy fibers that more fre-quently invade the pyramidal cell layer (sp). The inset in D showsthe mossy fibers to be more splayed than in A. Following exposureto QAV peptide (G–I) mossy fibers show an even greater degree ofdisorganization and loss of laminar specificity. Fibers frequentlyappear overgrown and extend into stratum oriens (so). sl, stratumlucidum, sp, stratum pyramidale, so, stratum oriens. Magnificationbars 5 82 lm (A, D, G); 24.6 lm (C, F, I); 41 lm (insets).
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 355
Hippocampus
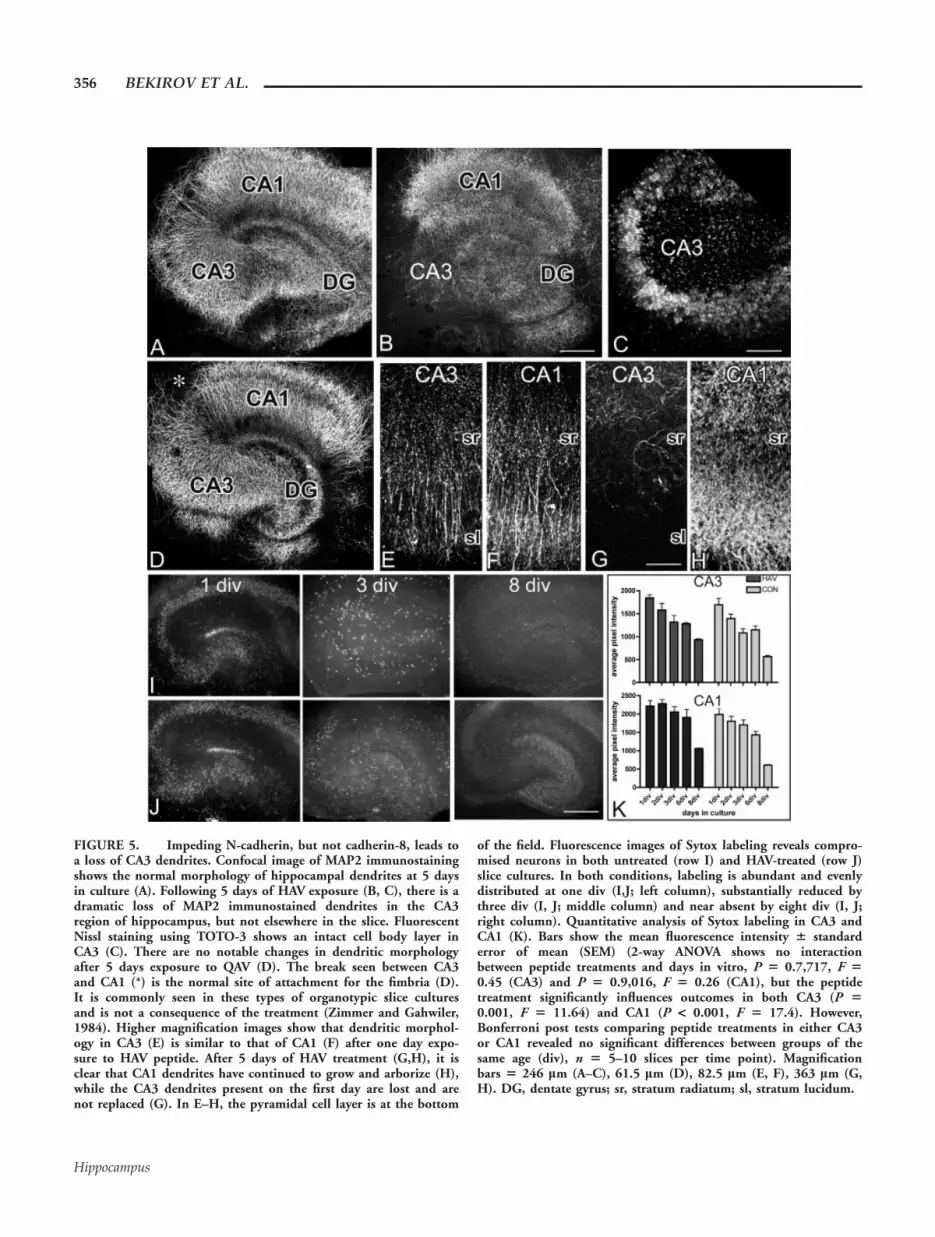
FIGURE 5. Impeding N-cadherin, but not cadherin-8, leads toa loss of CA3 dendrites. Confocal image of MAP2 immunostainingshows the normal morphology of hippocampal dendrites at 5 daysin culture (A). Following 5 days of HAV exposure (B, C), there is adramatic loss of MAP2 immunostained dendrites in the CA3region of hippocampus, but not elsewhere in the slice. FluorescentNissl staining using TOTO-3 shows an intact cell body layer inCA3 (C). There are no notable changes in dendritic morphologyafter 5 days exposure to QAV (D). The break seen between CA3and CA1 (*) is the normal site of attachment for the fimbria (D).It is commonly seen in these types of organotypic slice culturesand is not a consequence of the treatment (Zimmer and Gahwiler,1984). Higher magnification images show that dendritic morphol-ogy in CA3 (E) is similar to that of CA1 (F) after one day expo-sure to HAV peptide. After 5 days of HAV treatment (G,H), it isclear that CA1 dendrites have continued to grow and arborize (H),while the CA3 dendrites present on the first day are lost and arenot replaced (G). In E–H, the pyramidal cell layer is at the bottom
of the field. Fluorescence images of Sytox labeling reveals compro-mised neurons in both untreated (row I) and HAV-treated (row J)slice cultures. In both conditions, labeling is abundant and evenlydistributed at one div (I,J; left column), substantially reduced bythree div (I, J; middle column) and near absent by eight div (I, J;right column). Quantitative analysis of Sytox labeling in CA3 andCA1 (K). Bars show the mean fluorescence intensity 6 standarderror of mean (SEM) (2-way ANOVA shows no interactionbetween peptide treatments and days in vitro, P 5 0.7,717, F 50.45 (CA3) and P 5 0.9,016, F 5 0.26 (CA1), but the peptidetreatment significantly influences outcomes in both CA3 (P 50.001, F 5 11.64) and CA1 (P < 0.001, F 5 17.4). However,Bonferroni post tests comparing peptide treatments in either CA3or CA1 revealed no significant differences between groups of thesame age (div), n 5 5–10 slices per time point). Magnificationbars 5 246 lm (A–C), 61.5 lm (D), 82.5 lm (E, F), 363 lm (G,H). DG, dentate gyrus; sr, stratum radiatum; sl, stratum lucidum.
356 BEKIROV ET AL.
Hippocampus

of MAP2 in otherwise intact dendrites, but this is unlikely asstaining for tubulin also decreases in CA3 (data not shown). Todistinguish between an effect on dendritic outgrowth or retrac-tion, we monitored dendritic arbors over the course of 5 daysof HAV treatment. On day one, burgeoning dendritic arborscover the full extent of stratum radiatum in CA3 and CA1 incontrol and treated cultures (Figs. 5E,F). By the end of 3 days,labeled dendrites are reduced in CA3, and by day 5, nearlyabsent in HAV-treated cultures (Fig. 5G, and data not shown),a time at which DiI labeled mossy fibers show a relatively nor-mal trajectory (Figs. 4D,E). Over the same time course, CA3arbors in control cultures, and CA1 arbors in all cultures, showa dramatic increase in density (Figs. 5A,B,E,F,H). These datasuggest that HAV treatment prevents further dendritic elabora-tion and leads to dendritic retraction in CA3.
To determine whether this is a selective effect on dendritesor a cytotoxic effect on CA3 pyramidal neurons, we labeledHAV-treated and control cultures with Sytox, a high affinity,DNA binding stain that readily crosses compromised plasmamembrane in late apoptotic and necrotic cells and is excludedfrom healthy cells (Petersen and Dailey, 2004; Evans andCousin, 2006). At one day in culture, both control and HAV-treated cultures show extensive labeling (Figs. 5I–K) consistent
with the trauma that occurs on the surface during slicing(Pozzo Miller et al., 1994; Laake et al., 1999; Petersen andDailey, 2004). There is culture-to-culture variability in the re-gional intensity of the labeling patterns, but all principal layersare labeled consistently in both groups. By 3 days, Sytox label-ing is clearly decreased in control and HAV-treated cultures(Figs. 5I–K). Very little labeling is observed by 8 days in anyculture (Figs. 5I–K). Thus, some cell death occurs throughoutthe hippocampus similar to what has been reported previously(Pozzo Miller et al., 1994; Laake et al., 1999; Petersen andDailey, 2004), but the vast majority is early (1–2 days in cul-ture) and transient, similar between treatment groups, andshows no bias toward CA3 (Fig. 5K). Additionally, survival indissociated hippocampal pyramidal neurons exposed to HAVfor 2–7 days is indistinguishable from controls (DLB, unpub-lished data). Together these data suggest that N-cadherin adhe-sion is required to support and maintain the integrity of den-drites in CA3 and that the mistargeting we observe in themossy fibers (Figs. 4D–F) can be attributed, either partly or inwhole, to a loss of the target structure to which the axons nor-mally grow. Given the dramatic changes observed in CA3,mossy fiber targeting remains remarkably intact in the presenceof HAV peptide.
FIGURE 6. Synaptogenesis is inhibited when either N-cad-herin or cadherin-8 function is impeded. Confocal images of P5/6slices that were cultured for 5 days in the presence of the peptidesindicated and then fixed and immunostained for SV2 (A). Thehippocampal layers from which the images were taken are indi-cated by row labels: SL (stratum lucidum), SP (stratum pyrami-dale), and SO (stratum oriens). Images from HAV- and QAV-treated cultures show decreased labeling for SV2 throughout CA3relative to SCR control. The source of these differences was deter-mined using a morphometric analysis, and the data are summar-
ized in the bar graphs shown in B (HAV) and C (QAV). The num-ber of SV2-labeled puncta decreases after HAV treatment in alllamina compared with control (B). The size and intensity of thepuncta also decrease significantly in SL and SP. Following QAVtreatment (C), number, size, and intensity of labeled puncta aredecreased in SL. Puncta area and intensity are reduced in SP, andwhile puncta number and area are decreased in SO, intensity isslightly increased. Comparisons were made using unpaired t-testsand an alpha value that was corrected for layer and peptide. *P <0.05, **P < 0.005, ***P < 0.0,001. Magnification bar 5 12.3 lm.
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 357
Hippocampus

In contrast to the effects of HAV peptide on CA3 pyramidalneuron dendrites, MAP2 labeled dendrites in organotypic slicesthat have been exposed for 5 days to QAV appear very similarto untreated slices (Figs. 5A,D).
Cadherin-8 and N-Cadherin Are Required for theDevelopment of Presynaptic Terminals
Since mossy fiber targeting and CA3 pyramidal cell dendriticarbors are greatly changed following exposure to QAV or HAVpeptides, respectively, we asked how exposure to the peptidesimpacts the generation of synapses in CA3. To monitor synap-ses, cultures were immunostained for an integral synaptic vesi-cle protein, SV2 and the distribution of labeled clusters wasassessed (Voigt et al., 1993; Bozdagi et al., 2000). In HAV-treated cultures the numbers of SV2-labeled clusters arereduced in stratum lucidum as well as in the pyramidal celllayer and stratum oriens (Figs. 6A,B). In addition, size and in-tensity of individual clusters are reduced in stratum lucidumand the pyramidal cell layer as might be expected for vesiclesthat fail to form presynaptic aggregates and remain below thelimit of resolution. These data are consistent with a dramati-cally reduced target in CA3.
Exposure to QAV also reduces the number, size, and inten-sity of SV2-labeled clusters throughout most of CA3 (Figs.6A,C). Although pyramidal cell dendritic arbors remain intactin QAV-treated cultures, mossy fibers defasciculate and invadestrata pyramidale and oriens. Thus, QAV appears to prevent
the assembly of normal presynaptic vesicle aggregates in stra-tum lucidum despite multiple opportunities for en passant con-tact. This suggests that cadherin-8 binding or activation maybe important to stabilize developing synaptic contacts in stra-tum lucidum as well as in the pyramidal cell layer and stratumoriens, which also show a reduced cluster size (Figs. 6A,C).
Cadherin-8 and N-Cadherin Both Bind b-Cateninin DG Granule Cells
Interfering with either cadherin-8 or N-cadherin adhesionultimately reduces the number of presynaptic terminals formingin CA3. However, each cadherin appears to influence differ-ently either the pre- or the postsynaptic component of themossy fiber—CA3 synaptic circuit. In the organotypic cultures,interfering with N-cadherin binding primarily alters the organi-zation of the target CA3 pyramidal cell dendrites while interfer-ing with cadherin-8 binding alters the innervating mossy fibertrajectory. Since both cadherins are expressed by granule cells,we asked whether they might differentially recruit b-catenin.The highly conserved intracellular domains of all classic cadher-ins would be predicted to bind to b-catenin, which via a-cate-nin can firmly anchor cadherins to F-actin (Ozawa et al., 1989;Knudsen et al., 1995).
Immunoblots for cadherin-8 and N-cadherin indicate thatboth proteins are expressed in DG, and cadherin-8 levels areenriched relative to the cerebral cortex (Fig. 7A). Both cad-herin-8 and N-cadherin can efficiently pull down b-cateninfrom DG lysates (Fig. 7B). Cadherin-8 precipitates more b-cat-enin in the DG than in cerebral cortex, consistent with therelatively greater levels of cadherin-8 in DG (Figs. 7A,B). Incontrast, N-cadherin pulls down far more b-catenin in the DGthan in the cerebral cortex where expression levels are roughlyequivalent (Figs. 7A,B). This suggests that N-cadherin has astronger, more stable interaction with b-catenin in the DGthan in cerebral cortex, but could also reflect the increased lev-els of b-catenin in the DG. Although it is not possible to com-pare the relative levels of proteins pulled down in separateimmunoprecipitates, the results show clearly that b-catenin caninteract with both N-cadherin and cadherin-8 in DG. Thesedata suggest that the different functional outcomes we observefor N-cadherin and cadherin-8 may result from differences intheir subcellular distribution or from the recruitment and acti-vation of cis-binding partners that could activate different sig-naling cascades.
DISCUSSION
The selective expression of particular cadherins can sort cells,including neurons, into tissues, areas, or subregional domainsduring development. As neurons differentiate and form connec-tions, most express several cadherins simultaneously. How indi-vidual neurons utilize this diversity is poorly understood. Herewe asked how N-cadherin and cadherin-8, both coexpressed by
FIGURE 7. Cadherin-8 and N-cadherin bind b-catenin in DGgranule cells. (A) Whole cell lysates from P5/6 cerebral cortex andDG contain both cadherin-8 and N-cadherin. Cadherin-8 is moreabundant in DG and N-cadherin levels are equivalent in bothregions. (B) Immunoprecipitates of both cadherin-8 and N-cad-herin pull down b-catenin. Less b-catenin is pulled down in cere-bral cortex lysates than in DG by both cadherin-8 and N-cadherin.For cadherin-8, this likely reflects the different levels of cadherin-8in the lysates, but for N-cadherin, the interaction in DG appearsto be particularly strong. At the same time, b-catenin is expressedat higher levels in the DG than in cortex. Lysates incubated withequivalent levels of serum failed to show any specific reaction withb-catenin.
358 BEKIROV ET AL.
Hippocampus

DG granule cells and their targets, area CA3 neurons, influencethe development and targeting of the hippocampal mossy fiberpathway. Using organotypic hippocampal slice cultures, we findthat cadherin-8 is important for regulating mossy fiber growthand targeting, while N-cadherin is essential for maintaining theintegrity of CA3 pyramidal neurons. Both cadherins, however,contribute to stabilization of presynaptic boutons. Thus, eachcadherin appears to be recruited for distinct functions that dif-ferentially influence proper development of the presynapticinnervating pathway or the postsynaptic target dendrites,thereby acting complementing each other in the establishmentof a neural circuit.
Role of N-Cadherin in Mossy Fiber Targeting
In hippocampus, N-cadherin is distributed broadly in neu-rons and astrocytes (Redies and Takeichi, 1993; Wilby et al.,1999; Bekirov et al., 2002). Extending mossy fibers would beexpected to partner with N-cadherin on adjacent axons as wellas on astrocytes and dendrites in the neuropil. Thus, asexpected, HAV defasciculated mossy fibers in organotypic cul-tures. Premature defasciculation exposes fibers to inappropriatecues, and this is likely to be the cause of the increase in fibersinvading the pyramidal cell layer. Nevertheless, the effects onoutgrowth are modest, and the basic trajectory taken by thefibers is relatively normal. Fibers do not invade area CA3 stra-tum radiatum, rarely invade CA3 stratum oriens, and werenever observed anywhere in CA1. This is surprising, sinceHAV destroys the target CA3 dendrites in organotypic cultures,which are a likely source of homophilic binding interactions.Thus, mossy fiber guidance does not require interactionbetween appropriate pre- and postsynaptic targets. Although asimilar molecular disassociation has been observed in the devel-oping Drosophila olfactory system where N-cadherin influencesaxon and dendrite arborization independently (Hummel andZipursky, 2004; Zhu and Luo, 2004), our data show that nophysical association is required. The findings indicate that for atleast this pathway, guidance and synaptogenesis are separableevents.
Role of Cadherin-8 in Mossy Fiber Targeting
N-cadherin’s ability to promote axon extension has beenwell-documented in vitro for a variety of neuron types (Neuge-bauer et al., 1988; Tomaselli et al., 1988; Friedlander et al.,1989; Bixby and Zhang, 1990; Drazba and Lemmon, 1990;Doherty et al., 1991; Shimoyama et al., 2000; Utton et al.,2001). R-cadherin and cadherin-11 can also promote axonextension in vitro (Andrews and Mastick, 2003; Marthienset al., 2005). The high degree of structural similarity betweenclassic cadherins suggests that cadherin-8 should also promotegrowth. But we find that while dissociated granule cells adhereand extend axons on cadherin-8 substrates, axonal growth isnot enhanced as it is on N-cadherin. Exposure to QAV pep-tides, covering the domain analogous to the HAV domain inN-cadherin, increases mossy fiber growth on defined cadherin-8substrates and in organotypic cultures, but not when dissoci-
ated cells are plated on N-cadherin or untransfected L-cells.The simplest interpretation of these findings is that cadherin-8promotes stronger adhesion than N-cadherin in growing axonsand that QAV attenuates this, producing less controlled exten-sion. This idea is consistent with the distribution of cadherin-8in hippocampus: within CA3, cadherin-8 mRNA is expressedat low levels in proximal CA3, increases gradually, and reachesa peak near the CA1 border where mossy fibers end (Kore-matsu and Redies, 1997; Bekirov et al., 2002). However, disso-ciated granule cells show a slight increase in average axonlength as substrate concentrations are increased from 1 to 100lg/mL, suggesting that concentration dependent changes inadhesive strength are insufficient to account for our findings.Additionally, in vitro binding data suggest that cadherin-8mediated adhesion may be weaker than that mediated by N-cadherin (Kido et al., 1998). Although it is possible that theconcentrations tested are saturating and that the range observedin vivo is far below 1 lg/mL, it appears more likely that cad-herin-8 binding provokes a response via signaling intermediates,perhaps by recruiting a stop signal via b-catenin (Figs. 4 and7). Interestingly, b-catenin binding may be more important forstopping than for promoting growth as it also appears to bedispensable for N-cadherin mediated outgrowth in Xenopus reti-nal ganglion cells in vivo (Riehl et al., 1996).
Given that both N-cadherin and cadherin-8 bind b-cateninin DG, how might they differentially affect the mossy fiber tra-jectory? Several studies suggest that changes in additional juxta-membrane cadherin binding proteins can greatly alter strengthof adhesion by regulating lateral clustering (Yap et al., 1997,1998). Additionally, juxtamembrane-binding proteins, such asp120 catenin, delta catenin, and presenilin can bind and signalvia distinct sets of signaling partners (Hirano et al., 2003).However, it is not clear how cadherin-8 and N-cadherin, withtheir highly similar intracellular domains, could recruit differentjuxtamembrane partners. One possibility is that the cadherinsare signaling via distinct coreceptors or signaling complexes.The idea of a coreceptor is particularly appealing because cad-herin ectodomains are more likely to show binding characteris-tics specific to particular cadherins and could be used to distin-guish cadherin-8 and N-cadherin. In endothelial cells, interac-tions between the ectodomains of cadherin-5 and a coreceptor,the receptor protein tyrosine phosphatase (RPTP) VE-PTPmediate dephosphorylation of cadherin-5 and enhance thebarrier integrity of adherens junctions (Nawroth et al., 2002).In retinal ganglion cells, binding of the RPTP, PTPl, toN-cadherin regulates axon extension on an N-cadherin sub-strate (Burden-Gulley and Brady-Kalnay, 1999). Cadherininteraction with either of these RPTP partners is associatedwith stronger adhesion, and consistent with this, PTPl cancoimmunoprecipitate with an N-cadherin-b-catenin complex(Brady-Kalnay et al., 1995).
Proteoglycans may also regulate cadherin actions in mossyfibers (Balsamo and Lilien, 1990). Immunostaining for neuro-can reveals a sharp border of expression with high levels inCA3 stratum radiatum and low levels within stratum lucidum(Kurazono et al., 2001; Butler et al., 2004). Neurocan stimula-
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 359
Hippocampus

tion of retinal ganglion cells disassociates N-cadherin fromactin, presumably by binding a GPI-linked cell surface glycosyl-transferase that can form a complex with N-cadherin (Li et al.,2000). The phosphacan splice variant, RPTPb/f, may also reg-ulate N-cadherin adhesion (Meng et al., 2000), but it is largelyrestricted to astrocytes (Snyder et al., 1996; Butler et al.,2004), making it difficult to model a direct action with theneuronally restricted cadherin-8.
Recent work also suggests that N-cadherin can form a core-ceptor complex with Robo and that this inhibits N-cadherindependent axon extension in the presence of the Robo ligand,Slit (Rhee et al., 2002). The proposed N-cadherin-Robo com-plex provides an attractive mechanism to couple the mechanicsof outgrowth with axon guidance. Furthermore, one wouldanticipate such interactions to be regulated locally in a cell typespecific or regionally restricted manner. However, we have thusfar been unable to detect interactions between Robo familymembers and cadherins in DG (Bekirov, Svoronos, Benson,Kang, unpublished data).
Cadherins and Dendritic Arbors
Dendritic arbors of a variety of neurons require N-cadherinfor distinct stages of development. Xenopus retinal ganglion cellsexpressing a dominant negative N-cadherin fail to generatedendrites (Riehl et al., 1996), and chick horizontal cells showreduced arbor size (Tanabe et al., 2006). N-cadherin promotesdendritic outgrowth in hippocampal neurons (Esch et al., 2000),zebrafish amacrine cells require N-cadherin to arborize appropri-ately (Masai et al., 2003), and DN-cadherin restricts dendriticarbors that are generated by olfactory projection neurons in Dro-sophila (Zhu and Luo, 2004). Here we find that in CA3 of slicecultures, interfering with N-cadherin binding decreases MAP2-labeling over time suggesting that existing arbors retract and newgrowth is prevented, while MAP2-labeled dendrites in CA1 stra-tum radiatum and DG molecular layer appear comparatively nor-mal. Thus, it appears as though interactions across the HAVinterface trigger signals that are important for dendrite elabora-tion and maintenance and perhaps for CA3 pyramidal cell healthin general, since it seems unlikely that neurons will survive longwithout their dendrites. Since N-cadherin adhesion appears to beimportant for stabilizing synapses in developing hippocampalneurons (Togashi et al., 2002; Bozdagi et al., 2004), and recentwork supports a strong interrelationship between arbor and syn-apse stability (Meyer and Smith, 2006; Ruthazer et al., 2006), itmay be that dendrites retract in the absence of the stabilizinginfluence of synapses. Consistent with this, overall number andsize of presynaptic terminal puncta are reduced in CA3 in HAV-treated slices. However, QAV treatment produces similar changesin presynaptic terminal clusters without the devastating effect onCA3 neurons suggesting that synapse stability alone does notfully account for our observations.
Cadherins and Mossy Fiber Guidance
A variety of cell surface and cell adhesion proteins appears tocollectively promote mossy fiber fasciculation, as disruption of
any single one produces some defasciculation. This is particu-larly notable in proximal CA3. Mice deficient in CHL1 (closehomolog of L1) or NCAM, or the introduction of reagentsinterfering with LAMP or cadherin-8 adhesion produce defasci-culation in proximal CA3 (Pimenta et al., 1995; Cremer et al.,1997; Seki and Rutishauser, 1998; Montag-Sallaz et al., 2002).The defasciculated fibers inappropriately target the CA3 pyram-idal cell layer. Some innervation of the pyramidal cell layeroccurs normally over the course of development, but typicallynot to the degree observed following the manipulationsdescribed (Amaral, 1979; Amaral and Dent, 1981). The con-sistent direction of the misguided trajectories is likely to bedriven by the intact inhibitory extracellular matrix and repul-sive Sema6A activity in stratum radiatum (Wilson and Snow,2000; Butler et al., 2004; Suto et al., 2007), coupled with apermissive or even attractive environment generated locally bythe CA3 pyramidal cell layer (Steup et al., 2000). Distal CA3mistargeting occurs in mice lacking NCAM (Cremer et al.,1997; Seki and Rutishauser, 1998), following enzymatic re-moval of keratan sulfate glycosaminoglycans, or following expo-sure to anti-LAMP antibodies, HAV or QAV peptides (Pimentet al., 1995; Butler et al., 2004; present study). The targetingdefects suggest that keratan sulfate proteoglycans prevent mossyfibers from invading stratum radiatum (Butler et al., 2004),that LAMP and NCAM may be particularly important for pro-moting mossy fiber fasciculation or the retraction of aberrantprojections (Pimenta et al., 1995; Cremer et al., 1997; Sekiand Rutishauser, 1998), and cadherin-8 for stopping mossyfiber extension (Fig. 4). Together, these molecules help accountfor the high fidelity of mossy fiber guidance.
Acknowledgments
The authors thank Dr. S. T. Suzuki for generously providinghis cadherin-8 construct and stable cadherin-8 L-cell line, Dr.Ioana Carcea for her help with substrate strategies, and Dr.Caitlin Trasande for her work on the cell survival experiments.
REFERENCES
Amaral DG. 1979. Synaptic extensions from the mossy fibers of thefascia dentata. Anat Embryol (Berl) 155:241–251.
Amaral DG, Dent JA. 1981. Development of the mossy fibers of thedentate gyrus. I. A light and electron microscopic study of themossy fibers and their expansions. J Comp Neurol 195:51–86.
Andrews GL, Mastick GS. 2003. R-cadherin is a Pax6-regulated,growth-promoting cue for pioneer axons. J Neurosci 23:9873–9880.
Balsamo J, Lilien J. 1990. N-cadherin is stably associated with and isan acceptor for a cell surface N-acetylgalactosaminylphosphotrans-ferase. J Biol Chem 265:2923–2928.
Bekirov IH. 2004. The role of classic cadherins in the development ofhippocampal circuitry [Ph.D.]. New York: Mount Sinai School ofMedicine of New York University. 107 p.
Bekirov IH, Needleman LA, Zhang W, Benson DL. 2002. Identifica-tion and localization of multiple classic cadherins in developing ratlimbic system. Neuroscience 115:213–227.
360 BEKIROV ET AL.
Hippocampus

Benson DL, Tanaka H. 1998. N-cadherin redistribution during synap-togenesis in hippocampal neurons. J. Neurosci 18:6892–6904.
Bixby JL, Zhang R. 1990. Purified N-cadherin is a potent substratefor the rapid induction of neurite outgrowth. J Cell Biol 110:1253–1260.
Blaschuk OW, Sullivan R, David S, Pouliot Y. 1990. Identification ofa cadherin cell adhesion recognition sequence. Dev Biol 139:227–229.
Bozdagi O, Shan W, Tanaka H, Benson DL, Huntley GW. 2000.Increasing numbers of synaptic puncta during late-phase LTP: N-cadherin is synthesized, recruited to synaptic sites and required forpotentiation. Neuron 28:245–259.
Bozdagi O, Valcin M, Poskanzer K, Tanaka H, Benson DL. 2004.Temporally distinct demands for classic cadherins in synapse for-mation and maturation. Mol Cell Neurosci 27:509–521.
Brady-Kalnay SM, Rimm DL, Tonks NK. 1995. Receptor protein ty-rosine phosphatase PTPmu associates with cadherins and cateninsin vivo. J Cell Biol 130:977–986.
Burden-Gulley SM, Brady-Kalnay SM. 1999. PTPmu regulates N-cad-herin-dependent neurite outgrowth. J Cell Biol 144:1323–1336.
Butler CD, Schnetz SA, Yu EY, Davis JB, Temple K, Silver J, MaloufAT. 2004. Keratan sulfate proteoglycan phosphacan regulates mossyfiber outgrowth and regeneration. J Neurosci 24:462–473.
Chappuis-Flament S, Wong E, Hicks LD, Kay CM, Gumbiner BM.2001. Multiple cadherin extracellular repeats mediate homophilicbinding and adhesion. J Cell Biol 154:231–243.
Cline HT. 2001. Dendritic arbor development and synaptogenesis.Curr Opin Neurobiol 11:118–126.
Cremer H, Chazal G, Goridis C, Represa A. 1997. NCAM is essentialfor axonal growth and fasciculation in the hippocampus. Mol CellNeurosci 8:323–335.
Dailey ME, Buchanan J, Bergles DE, Smith SJ. 1994. Mossy fibergrowth and synaptogenesis in rat hippocampal slices in vitro. JNeurosci 14(3 Part 1):1060–1078.
Doherty P, Rowett LH, Moore SE, Mann DA, Walsh FS. 1991. Neu-rite outgrowth in response to transfected N-CAM and N-cadherinreveals fundamental differences in neuronal responsiveness toCAMs. Neuron 6:247–258.
Drazba J, Lemmon V. 1990. The role of cell adhesion molecules inneurite outgrowth on Muller cells. Dev Biol 138:82–93.
Elste AM, Benson DL. 2006. Structural basis for developmentallyregulated changes in cadherin function at synapses. J Comp Neurol495:324–335.
Esch T, Lemmon V, Banker G. 1999. Local presentation of substratemolecules directs axon specification by cultured hippocampal neu-rons. J Neurosci 19:6417–6426.
Esch T, Lemmon V, Banker G. 2000. Differential effects of NgCAMand N-cadherin on the development of axons and dendrites by cul-tured hippocampal neurons. J Neurocytol 29:215–223.
Espeseth A, Marnellos G, Kintner C. 1998. The role of F-cadherin inlocalizing cells during neural tube formation in Xenopus embryos.Development 125:301–312.
Evans GJ, Cousin MA. 2007. Simultaneous monitoring of three keyneuronal functions in primary neuronal cultures. J Neurosci Meth-ods 160:197–205.
Friedlander DR, Mege RM, Cunningham BA, Edelman GM. 1989.Cell sorting-out is modulated by both the specificity and amountof different cell adhesion molecules (CAMs) expressed on cell sur-faces. Proc Natl Acad Sci USA 86:7043–7047.
Frotscher M, Heimrich B. 1995. Lamina-specific synaptic connectionsof hippocampal neurons in vitro. J Neurobiol 26:350–359.
Gahwiler BH. 1981. Organotypic monolayer cultures of nervous tis-sue. J Neurosci Methods 4:329–342.
Gahwiler BH. 1984a. Development of the hippocampus in vitro: Celltypes, synapses and receptors. Neuroscience 11:751–760.
Gahwiler BH. 1984b. Slice cultures of cerebellar, hippocampal andhypothalamic tissue. Experientia 40:235–243.
Gil OD, Needleman L, Huntley GW. 2002. Developmental patternsof cadherin expression and localization in relation to compartmen-talized thalamocortical terminations in rat barrel cortex. J CompNeurol 453:372–388.
Goslin K, Banker G. 1989a. Experimental observations on the devel-opment of polarity by hippocampal neurons in culture. J Cell Biol108:1507–1516.
Goslin K, Banker G. 1989b. Experimental observations on the devel-opment of polarity by hippocampal neurons in culture. J Cell Biol108:1507–1516.
Goslin K, Schreyer DJ, Skene JHP, Banker G. 1990. Changes in thedistribution of GAP-43 during the development of neuronal polar-ity. J Neurosci 10:588–602.
Haspel J, Blanco C, Jacob J, Grumet M. 2001. System for cleavableFc fusion proteins using tobacco etch virus (TEV) protease. Bio-techniques 30:60–61,64–66.
Hatta K, Takeichi M. 1986. Expression of N-cadherin adhesion mole-cules associated with early morphogenetic events in chick develop-ment. Nature 320:447–449.
Hirano S, Suzuki ST, Redies CM. 2003. The cadherin superfamily inneural development: Diversity, function and interaction with othermolecules. Front Biosci 8:D306–D355.
Hummel T, Zipursky SL. 2004. Afferent induction of olfactory glo-meruli requires N-cadherin. Neuron 42:77–88.
Huntley GW, Benson DL. 1999. Neural (N)-cadherin at developingthalamocortical synapses provides an adhesion mechanism for theformation of somatopically organized connections. J Comp Neurol407:453–471.
Inoue A, Sanes JR. 1997. Lamina-specific connectivity in the brain:regulation by N-cadherin, neurotrophins, and glycoconjugates. Sci-ence 276:1428–1431.
Iwai Y, Usui T, Hirano S, Steward R, Takeichi M, Uemura T. 1997.Axon patterning requires DN-cadherin, a novel neuronal adhesionreceptor, in the Drosophila embryonic CNS. Neuron 19:77–89.
Kido M, Obata S, Tanihara H, Rochelle JM, Seldin MF, Taketani S,Suzuki ST. 1998. Molecular properties and chromosomal locationof cadherin-8. Genomics 48:186–194.
Knudsen KA, Peralta Soler A, Johnson KR, Wheelock MJ. 1995.Interaction of a-actinin with the cadherin/catenin cell-cell adhesioncomplex via a-catenin. J Cell Biol 130:67–77.
Korematsu K, Redies C. 1997. Expression of cadherin-8 mRNA inthe developing mouse central nervous system. J Comp Neurol387:291–306.
Kurazono S, Okamoto M, Sakiyama J, Mori S, Nakata Y, Fukuoka J,Amano S, Oohira A, Matsui H. 2001. Expression of brain specificchondroitin sulfate proteoglycans, neurocan and phosphacan, in thedeveloping and adult hippocampus of Ihara’s epileptic rats. BrainRes 898:36–48.
Laake JH, Haug FM, Wieloch T, Ottersen OP. 1999. A simple invitro model of ischemia based on hippocampal slice cultures andpropidium iodide fluorescence. Brain Res Brain Res Protoc 4:173–184.
Lee C, Herman T, Clandinin TR, Lee R, Zipursky SL. 2001. N-cad-herin regulates target specificity in the Drosophila visual system.Neuron 30:437–450.
Li H, Leung TC, Hoffman S, Balsamo J, Lilien J. 2000. Coordinateregulation of cadherin and integrin function by the chondroitinsulfate proteoglycan neurocan. J Cell Biol 149:1275–1288.
Marthiens V, Gavard J, Padilla F, Monnet C, Castellani V, LambertM, Mege RM. 2005. A novel function for cadherin-11 in the regu-lation of motor axon elongation and fasciculation. Mol Cell Neu-rosci 28:715–726.
Masai I, Lele Z, Yamaguchi M, Komori A, Nakata A, Nishiwaki Y,Wada H, Tanaka H, Nojima Y, Hammerschmidt M, Wilson SW,Okamoto H. 2003. N-cadherin mediates retinal lamination, main-tenance of forebrain compartments and patterning of retinal neu-rites. Development 130:2479–2494.
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 361
Hippocampus

Meng K, Rodriguez-Pena A, Dimitrov T, Chen W, Yamin M, NodaM, Deuel TF. 2000. Pleiotrophin signals increased tyrosine phos-phorylation of beta beta-catenin through inactivation of the intrin-sic catalytic activity of the receptor-type protein tyrosine phospha-tase beta/zeta. Proc Natl Acad Sci USA 97:2603–2608.
Meyer MP, Smith SJ. 2006. Evidence from in vivo imaging that syn-aptogenesis guides the growth and branching of axonal arbors bytwo distinct mechanisms. J Neurosci 26:3604–3614.
Miskevich F, Zhu Y, Ranscht B, Sanes JR. 1998. Expression of multi-ple cadherins and catenins in the chick optic tectum. Mol CellNeurosci 12:240–255.
Montag-Sallaz M, Schachner M, Montag D. 2002. Misguided axonalprojections, neural cell adhesion molecule 180 mRNA upregula-tion, and altered behavior in mice deficient for the close homologof L1. Mol Cell Biol 22:7967–7981.
Nawroth R, Poell G, Ranft A, Kloep S, Samulowitz U, Fachinger G,Golding M, Shima DT, Deutsch U, Vestweber D. 2002. VE-PTPand VE-cadherin ectodomains interact to facilitate regulation ofphosphorylation and cell contacts. Embo J 21:4885–4895.
Neugebauer KM, Tomaselli KJ, Lilien J, Reichardt LF. 1988. N-cad-herin, NCAM, and integrins promote retinal neurite outgrowth onastrocytes in vitro J Cell Biol 107:1177–1187.
Noe V, Willems J, Vandekerckhove J, Roy FV, Bruyneel E, Mareel M.1999. Inhibition of adhesion and induction of epithelial cellinvasion by HAV-containing E-cadherin-specific peptides. J CellSci 112 (Part 1):127–135.
Nose A, Nagafuchi A, Takeichi M. 1988. Expressed recombinant cad-herins mediate cell sorting in model systems. Cell 54:993–1001.
Nose A, Tsuji K, Takeichi M. 1990. Localization of specificitydetermining sites in cadherin cell adhesion molecules. Cell 61:147–155.
Ozawa M, Baribault H, Kemler R. 1989. The cytoplasmic domain ofthe cell adhesion molecule uvomorulin associates with three inde-pendent proteins structurally related in different species. Embo J8:1711–1717.
Patel SD, Ciatto C, Chen CP, Bahna F, Rajebhosale M, Arkus N,Schieren I, Jessell TM, Honig B, Price SR, Shapiro L. 2006. TypeII cadherin ectodomain structures: Implications for classical cad-herin specificity. Cell 124:1255–1268.
Petersen MA, Dailey ME. 2004. Diverse microglial motility behaviorsduring clearance of dead cells in hippocampal slices. Glia 46:195–206.
Pimenta AF, Zhukareva V, Barbe MF, Reinoso BS, Grimley C, HenzelW, Fischer I, Levitt P. 1995. The limbic system-associated mem-brane protein is an Ig superfamily member that mediates selectiveneuronal growth and axon targeting. Neuron 15:287–297.
Poskanzer K, Needleman LA, Bozdagi O, Huntley GW. 2003. N-cad-herin regulates ingrowth and laminar targeting of thalamocorticalaxons. J Neurosci 23:2294–2305.
Pozzo Miller LD, Mahanty NK, Connor JA, Landis DM. 1994. Spon-taneous pyramidal cell death in organotypic slice cultures from rathippocampus is prevented by glutamate receptor antagonists. Neu-roscience 63:471–487.
Price SR, De Marco Garcia NV, Ranscht B, Jessell TM. 2002. Regula-tion of motor neuron pool sorting by differential expression oftype II cadherins. Cell 109:205–216.
Rakic P. 1975. Role of cell interaction in development of dendriticpatterns. Adv Neurol 12:117–134.
Redies C, Engelhart K, Takeichi M. 1993. Differential expression ofN- and R-cadherin in functional neuronal systems and other struc-tures of the developing chicken brain. J Comp Neurol 333:398–416.
Redies C, Takeichi M. 1993. Expression of N-cadherin mRNA duringdevelopment of the mouse brain. Dev Dyn 197:26–39.
Rhee J, Mahfooz NS, Arregui C, Lilien J, Balsamo J, VanBerkum MF.2002. Activation of the repulsive receptor roundabout inhibits N-cadherin-mediated cell adhesion. Nat Cell Biol 4:798–805.
Riehl R, Johnson K, Bradley R, Grunwald GB, Cornel E, LilienbaumA, Holt CE. 1996. Cadherin function is required for axon out-growth in retinal ganglion cells in vivo. Neuron 17:837–848.
Robain O, Barbin G, Billette de Villemeur T, Jardin L, Jahchan T,Ben-Ari Y. 1994. Development of mossy fiber synapses in hippo-campal slice culture. Brain Res Dev Brain Res 80:244–250.
Ruthazer ES, Li J, Cline HT. 2006. Stabilization of axon branch dy-namics by synaptic maturation. J Neurosci 26:3594–3603.
Seki T, Rutishauser U. 1998. Removal of polysialic acid-neural cell ad-hesion molecule induces aberrant mossy fiber innervation and ec-topic synaptogenesis in the hippocampus. J Neurosci 18:3757–3766.
Shapiro L, Fannon AM, Kwong PD, Thompson A, Lehmann MS, Gru-bel G, Legrand JF, Als-Nielsen J, Colman DR, Hendrickson WA.1995. Structural basis of cell-cell adhesion by cadherins. Nature374:327–337.
Shimoyama Y, Tsujimoto G, Kitajima M, Natori M. 2000. Identifica-tion of three human type-II classic cadherins and frequent hetero-philic interactions between different subclasses of type-II classiccadherins. Biochem J 349 (Part 1):159–167.
Snyder SE, Li J, Schauwecker PE, McNeill TH, Salton SR. 1996.Comparison of RPTP zeta/beta, phosphacan, and trkB mRNAexpression in the developing and adult rat nervous system andinduction of RPTP zeta/beta and phosphacan mRNA followingbrain injury. Brain Res Mol Brain Res 40:79–96.
Stearns ME, Binder LI. 1987. Evidence that MAP-2 may be involvedin pigment granule transport in squirrel fish erythrophores. CellMotil Cytoskeleton 7:221–234.
Steup A, Lohrum M, Hamscho N, Savaskan NE, Ninnemann O,Nitsch R, Fujisawa H, Puschel AW, Skutella T. 2000. Sema3C andnetrin-1 differentially affect axon growth in the hippocampal for-mation. Mol Cell Neurosci 15:141–155.
Stoppini L, Buchs PA, Muller D. 1991. A simple method for organo-typic cultures of nervous tissue. J Neurosci Methods 37:173–182.
Suto F, Tsuboi M, Kamiya H, Mizuno H, Kiyama Y, Komai S, Shi-mizu M, Sanbo M, Yagi T, Hiromi Y, Chedotal A, Mitchell KJ,Manabe T, Fujisawa H. 2007. Interactions between Plexin-A2,Plexin-A4, and Semaphorin 6A control lamina-restricted projectionof hippocampal mossy fibers. Neuron 53:535–547.
Suzuki SC, Inoue T, Kimura Y, Tanaka T, Takeichi M. 1997a. Neuro-nal circuits are subdivided by differential expression of type-II clas-sic cadherins in postnatal mouse brains. Mol Cell Neurosci 9:433–447.
Suzuki SC, Inoue T, Kimura Y, Tanaka T, Takeichi M. 1997b. Neuro-nal circuits are subdivided by differential expression of type-II clas-sic cadherins in postnatal mouse brains. Mol Cell Neurosci 9:433–447.
Tanabe K, Takahashi Y, Sato Y, Kawakami K, Takeichi M, NakagawaS. 2006. Cadherin is required for dendritic morphogenesis andsynaptic terminal organization of retinal horizontal cells. Develop-ment 133:4085–4096.
Toda M, Asou H, Miura M, Toya S, Uyemura K. 1994. GFAP trans-fected cells produce laminin, leading to neurite outgrowth promo-tion. Neuroreport 5:1969–1972.
Togashi H, Abe K, Mizoguchi A, Takaoka K, Chisaka O, Takeichi M.2002. Cadherin regulates dendritic spine morphogenesis. Neuron35:77–89.
Tomaselli KJ, Neugebauer KM, Bixby JL, Lilien J, Reichardt LF.1988. N-cadherin and integrins: Two receptor systems that mediateneuronal process outgrowth on astrocyte surfaces. Neuron 1:33–43.
Treubert-Zimmermann U, Heyers D, Redies C. 2002. Targeting axonsto specific fiber tracts in vivo by altering cadherin expression. JNeurosci 22:7617–7626.
Utton MA, Eickholt B, Howell FV, Wallis J, Doherty P. 2001. SolubleN-cadherin stimulates fibroblast growth factor receptor dependentneurite outgrowth and N-cadherin and the fibroblast growth factorreceptor co-cluster in cells. J Neurochem 76:1421–1430.
362 BEKIROV ET AL.
Hippocampus

Voigt T, De Lima AD, Beckmann M. 1993. Synaptophysin immuno-histochemistry reveals inside-out pattern of early synaptogenesis inferret cerebral cortex. J Comp Neurol 330:48–64.
Wilby MJ, Muir EM, Fok-Seang J, Gour BJ, Blaschuk OW, FawcettJW. 1999. N-Cadherin inhibits Schwann cell migration on astro-cytes. Mol Cell Neurosci 14:66–84.
Williams E, Williams G, Gour BJ, Blaschuk OW, Doherty P. 2000. Anovel family of cyclic peptide antagonists suggests that N-cadherinspecificity is determined by amino acids that flank the HAV motif.J Biol Chem 275:4007–4012.
Wilson MT, Snow DM. 2000. Chondroitin sulfate proteoglycanexpression pattern in hippocampal development: Potential regula-tion of axon tract formation. J Comp Neurol 424:532–546.
Wohrn JC, Nakagawa S, Ast M, Takeichi M, Redies C. 1999. Combi-natorial expression of cadherins in the tectum and the sorting ofneurites in the tectofugal pathways of the chicken embryo. Neuro-science 90:985–1000.
Yamagata M, Herman JP, Sanes JR. 1995. Lamina-specific expressionof adhesion molecules in developing chick optic tectum. J Neurosci15:4556–4571.
Yap AS, Brieher WM, Pruschy M, Gumbiner BM. 1997. Lateral clus-tering of the adhesive ectodomain: A fundamental determinant ofcadherin function. Curr Biol 7:308–315.
Yap AS, Niessen CM, Gumbiner BM. 1998. The juxtamembraneregion of the cadherin cytoplasmic tail supports lateral clustering,adhesive strengthening, and interaction with p120ctn. J Cell Biol141:779–789.
Zhu H, Luo L. 2004. Diverse functions of N-cadherin in dendriticand axonal terminal arborization of olfactory projection neurons.Neuron 42:63–75.
Zimmer J, Gahwiler BH. 1984. Cellular and connective organizationof slice cultures of the rat hippocampus and fascia dentata. JComp Neurol 228:432–446.
CAD-8 AND N-CAD IN MOSSY FIBER DEVELOPMENT 363
Hippocampus