a novel postsynaptic density protein: the monocarboxylate transporter mct2 is co-localized with...
TRANSCRIPT

Abstract Confocal immunofluorescence microscopyshowed strong monocarboxylate transporter 2 (MCT2)labeling of Purkinje cell bodies and punctate labeling inthe molecular layer. By immunogold cytochemistry, itcould be demonstrated that the MCT2 immunosignalwas concentrated at postsynaptic densities of parallel fiber–Purkinje cell synapses. The distribution of MCT2transporters within the individual postsynaptic densitiesmimicked that of the δ2 glutamate receptor, as shown byuse of two different gold-particle sizes. The MCT2 dis-tribution was also compared with the distributions ofother monocarboxylate transporters (MCT1 and MCT4).The MCT1 immunolabeling was localized in the endo-thelial cells, while MCT4 immunogold particles were as-sociated with glial profiles, including those abutting thesynaptic cleft of the parallel fiber-spine synapses. Thepostsynaptic density (PSD) molecules identified so farcan be divided into five classes: receptors, their anchor-ing molecules, molecules involved in signal transduc-tion, ion channels, and attachment proteins. Here, weprovide evidence that this list of molecules must now beextended to comprise an organic molecule transporter:the monocarboxylate transporter MCT2. The present da-ta suggest that MCT2 has specific transport functions re-
lated to the synaptic cleft and that this transporter mayallow an influx of lactate derived from perisynaptic glialprocesses. The expression of MCT2 in synaptic mem-branes may allow energy supply to be tuned to the exci-tatory drive.
Keywords Monocarboxylate transporters · Cerebellum ·Parallel fiber-spine synapses · Immunocytochemistry ·Glutamate receptors
Introduction
The monocarboxylate transporters (MCTs) are a familyof proton-linked transporters with a number of sub-strates, including branched-chain oxo acids derived fromleucine, valine, and isoleucine; the keton bodies acetoac-etate, β-hydroxybutyrate, and acetate; as well as pyru-vate and lactate (Halestrap and Price 1999). Lactate (co-transported with protons) is the most important substrate.Several isoforms of MCT, all with 12 predicted trans-membrane regions (Halestrap and Price 1999), have beencharacterized with respect to function and tissue distribu-tion. The picture that has emerged is that the kineticproperties and substrate specificity of a given isoformare related to the unique metabolic requirements of thetissue in which it is localized (Halestrap and Price 1993,1999) MCT1, -2, and -4 are the monocarboxylate trans-porters that have been studied most extensively. The firstof these was cloned from hamster ovary cells by Garciaet al. (1994) and found to be widely expressed, with par-ticularly high concentrations in the heart and red muscle.The expression level in muscle was upregulated in re-sponse to work, suggesting a special role coupled to lac-tic-acid oxidation. The second isoform (MCT2) was iso-lated from a Syrian hamster liver library and was shownto have a ~60% amino acid sequence identity withMCT1 (Garcia et al. 1995). Later Jackson et al. (1997)cloned a rat MCT2 homologue. MCT2 has a ten-fold
L. Bergersen · O. Wærhaug · J. Helm · M. Thomas O.P. Ottersen (✉ )Department of Anatomy, Institute of Basic Medical Sciences, University of Oslo, POB 1105 Blindern, 0317 Oslo, Norwaye-mail: [email protected].: +47-22851270, Fax: +47-22851299
A.J. Davies · M.C. Wilson · A.P. HalestrapDepartment of Biochemistry, School of Medical Sciences, University of Bristol, Bristol BS8 1TD, UK
P. LaakeSection of Medical Statistics, University of Oslo, POB 1122 Blindern, 0317 Oslo, Norway
L. Bergersen · O. WærhaugThe Norwegian University of Sport and Physical Education, POB 4014 Ullevål Hageby, 0806 Oslo, Norway
Exp Brain Res (2001) 136:523–534DOI 10.1007/s002210000600
R E S E A R C H A RT I C L E
Linda Bergersen · Ola Wærhaug · Johannes Helm Marion Thomas · Petter Laake · Andrew J. Davies Mariangela C. Wilson · Andrew P. Halestrap Ole P. Ottersen
A novel postsynaptic density protein: the monocarboxylate transporterMCT2 is co-localized with δ-glutamate receptors in postsynaptic densities of parallel fiber–Purkinje cell synapsesReceived: 4 July 2000 / Accepted: 5 October 2000 / Published online: 15 December 2000© Springer-Verlag 2000

higher affinity for substrates than MCT1 and MCT4 andis found in cells where rapid uptake at low substrate con-centrations is required, including proximal kidney-tubulecells and sperm tails. By contrast, MCT4 is most evidentin white muscle and other cells with a high glycolyticrate, such as tumor cells and white blood cells, suggest-ing that it is expressed at sites with a predominant lactic-acid efflux (Halestrap and Price 1999).
Brain tissue is heterogeneous in terms of energy me-tabolism. Several lines of evidence indicate that there isan exchange of energy substrates and metabolites be-tween neurons and glia and that this cellular interactionplays a critical role in the brain’s energy homeostasis(Poitry-Yamate et al. 1995; Pellerin et al. 1998a; Magistretti et al. 1999). However, the nature of this in-teraction has not been worked out in any detail. The aimof the present study was to assess the distribution ofthree major MCTs (MCT1, 2, and 4) in brain neuropil,using the cerebellum as a model. Extrapolating from thesituation in peripheral tissues, we assumed that cellularand subcellular heterogeneities in energy metabolismwould be reflected by differences in the complement andexpression levels of monocarboxylate transporters. Weshow that MCT4 is mainly expressed by astrocytes,while MCT2 is selectively expressed in Purkinje cellspines, with an enrichment corresponding to the postsyn-aptic density (PSD). MCT1 was located in endothelialcells, as described previously (Gerhart et al. 1997). Theaccumulation of MCT2 at the PSD suggests that thepostsynaptic membrane is not only involved in mediat-ing synaptic transmission, but that it also contributes totransport of energy substrates and to the regulation oflactate and proton concentrations in the synaptic cleft.
Materials and methods
Animals and preparation of tissues
Adult male Wistar rats (250–300 g; n=5, from Møllegaard, Ejby,Denmark) were used. They had been allowed free access to foodand drinking water. All experiments were performed in accor-dance with the guidelines of the Norwegian Committee on AnimalExperimentation. The animals were killed with an intraperitonealinjection of Equithesin (0.4 ml per 100 g b.w.) and subjected totranscardiac perfusion with 2% dextran (MW 70,000) in 0.1 M so-dium-phosphate buffer (PB; pH 7.4, 4°C, 15 s), followed by amixture of glutaraldehyde (0.1%) and formaldehyde (4%; freshlydepolymerized from paraformaldehyde) in the same buffer (roomtemperature, 50 ml/min for 20 min). The brain was left in situovernight (4°C). Specimens from the cerebellum (lobule VI) wereisolated, cryoprotected in graded concentrations of phosphate-buffered glycerol, and rapidly frozen in liquid propane (–170°C)in a cryofixation unit (Reichert KF80, Vienna, Austria). The speci-mens were transferred to 0.5% uranyl acetate dissolved in an-hydrous methanol (–90°C) in a cryosubstitution unit (AFS; Reichert). The temperature was raised stepwise to –45°C. Thesamples were infiltrated with Lowicryl HM20 resin (Lowi, Wald-kraiburg, Germany), and polymerization was induced by UV lightfor 48 h. A detailed description of the procedure has been pub-lished (van Lookeren Campagne et al. 1991; Hjelle et al. 1994).
Expression of MCT2 in COS-cells
Expression of MCT2 in COS-cells was performed as previouslydescribed (Kirk et al. 2000). In brief, the coding region of MCT2was subcloned into a mammalian expression vector (Promega),and the response plasmid used in the Tet-Off gene-expressionsystem (Clontech). The resulting constructs were expressed inCOS cells by a liposome-mediated transfection procedure. Aftertransfection, the cells were fixed and stained with the MCT2 anti-bodies for confocal microscopy. Western blots of membrane prep-arations were derived from transfected and nontransfected cells.
Immunoblots of total brain extracts
Proteins were isolated as previously described (Han et al. 1995;McCullagh et al. 1996). Protein samples of cerebellum and fore-brain were separated on 8% SDS-polyacrylamide gels (130 mA,9 h). Proteins were then transferred to Immobilon polyvinylidenedifluoride membranes (100 V, 60 min). The membranes were incu-bated with the MCT2 antibody (0.1 µg/ml overnight), followed bydonkey anti-rabbit IgG conjugated to horseradish peroxidase(Amersham, 0.1 µg/ml, 1 h). MCT2 was detected using the en-hanced chemiluminescence method (Hyperfilm-ECL, Amersham).Molecular-weight markers were included.
Immunofluorescence
Cerebellum was dissected out after fixation, as described above.After fixation, the cerebellum was rinsed in PB and incubated se-quentially in 10% (3 h), 20% (3 h), and 30% (overnight) sucrosein PB for cryoprotection. The cerebellum was sectioned at12–15 µm thickness on a cryostat. Sections were collected on gel-atine-coated slides and stored at –20°C before use in indirect im-munofluorescence experiments, which were carried out as de-scribed (Veruki and Wässle 1996). The antibodies were diluted1:5000 (0.4 µg/ml; MCT1), 1:2000 (0.1 µg/ml; MCT2), 1:5000(0.4 µg/ml; MCT4), and 1:2000 (1 µg/ml; δ2 glutamate receptor)in 0.1 M PB with 3% normal goat serum, 1% bovine serum albu-min, 0.5% Triton-X-100, and 0.05% sodium azide, pH 7.4. Theprimary antibodies were revealed by a carboxymethylindocyanine(Cy3)-coupled secondary antibody (1:1000, Jackson Immuno Re-search Laboratories, West Grove, Penn., USA). Secondary anti-bodies were diluted in the same solution as the primary antibodies.Cerebellar sections were viewed with a confocal microscope (Leica TCS SP, Tektronix Phaser 440, dye sublimation).
Postembedding immunogold cytochemistry
Ultrathin sections were mounted on nickel grids and processed forimmunogold cytochemistry as described by Matsubara et al.(1996). Briefly, the sections were treated with saturated solutionsof NaOH in absolute ethanol (2–3 s), rinsed, and incubated se-quentially in; (1) 0.1% sodium borohydride and 50 mM glycine inTris buffer containing 0.05 M NaCl and 0.1% Triton X-100(TBNT); (2) 2% human serum albumin (HSA) in TBNT; (3) anti-bodies to MCT1 (dilution: 2 µg/ml), MCT2 (dilution: 1 µg/ml),MCT4 (dilution: 2 µg/ml), δ2 glutamate receptor (dilution:1 µg/ml), GluR2/3 (dilution: 2 µg/ml), in TBNT and 2% HSA; (4)2% HSA in TBNT; and (5) goat anti-rabbit immunoglobulins cou-pled to 10-nm gold particles (Amersham, Arlington Heights, Ill.,USA) and diluted 1:20 in TBNT with 2% HSA and 5 mg/ml poly-ethyleneglycol. In double-labeling experiments (Ottersen et al.1992), the sections were first treated with antibodies againstMCT2 (dilution: 1 µg/ml) and, then, with antibodies against δ2glutamate receptors (dilution: 1 µg/ml). Formaldehyde vapor(80°C, 1 h) was used between the sequential incubations to pre-vent interference (Wang and Larsson 1985). The MCT2 and δ2-glutamate receptors were distinguished by means of different goldparticle sizes (10 nm for MCT2 and 15 nm for the δ2 receptor).
524

The sections were examined in a Philips CM10 transmission elec-tron microscope.
Antibodies
Antibodies against rat MCT1, MCT2, and MCT4 were raised inNew-Zealand white rabbits against C-terminal peptides, conjugat-ed to keyhole limpet haemocyanin (KLH), and subjected to affini-ty purification as described previously (Poole et al. 1996; Jacksonet al. 1997; Wilson et al. 1998). The peptide sequence used for im-munization was CPQQNSSGDPAEEESPV for MCT1, NTH-NPPSDRDKESSI for MCT2, and CEPEKNGEVVHPPETSV forMCT4. A search in the Swiss-Prot sequence database revealed noproteins with significant homologies to the peptide sequence usedfor immunization (Fig. 1). The δ2-glutamate-receptor antibodywas raised against a synthetic peptide QPTPTLGLNLGND-PDRGTSI, corresponding to the C-terminus of the rat δ2-receptorsubunit (Mayat et al. 1995). This antibody also recognizes the δ1-receptor subunit, but does not label other glutamate receptors, in-cluding α-amino-3-hydoxy-5-methyl-4-isoxazole propionic acid(AMPA) and N-methyl-D-aspartate (NMDA) receptors (Mayat et al. 1995). The AMPA receptor antibody (Ab25; Wenthold et al.1992) used for control reacts with GluR2 as well as GluR3.
Quantitative analysis
Ultrathin sections were either singly labeled with 10-nm gold par-ticles to reveal the localization of MCT2, or double labeled with10-nm gold particles and 15-nm gold particles for simultaneouslocalization of MCT2 and δ2 receptors, respectively. Synapsesthroughout the different layers of the cerebellum were identified inthe electron microscope according to morphological criteria (Palayand Chan-Palay 1974). Transversely cut synaptic profiles withclearly visible postsynaptic membrane and postsynaptic densitywere selected for photography, and quantitative analysis was per-formed on photomicrographs magnified ×63 000, ×86 250, or×97 500. At synapses between parallel fibers and Purkinje cellspines, the positions of gold particles were determined along twoaxes; an axis perpendicular to the postsynaptic membrane and anaxis tangential to this. All particles lying within a field limited by
at least 200 nm in pre- and postsynaptic directions from the middle of the postsynaptic membrane were recorded. The lengthof the synaptic profiles, the thickness of the postsynaptic densities,and the width of the synaptic clefts were measured
Statistics
The data were entered in Statview v.4.5 (Abacus Concept) andSystat v.9.0 (SPSS) for statistical treatment. The analysis was per-formed only on synaptic profiles longer than 150 nm (Landsend etal. 1997) To ensure that all immunoreactivity possibly related tothe synapse was detected, particles occurring within 50 nm of themiddle of the postsynaptic membrane and within 50 nm of the pe-ripheral edge of the PSD were included (Matsubara et al. 1996). Inaddition to routine descriptive statistics, two-dimensional q-q plotsand two-sample Kolmogorov-Smirnov nonparametric tests wereused to compare the distribution of gold particles and to examinethe significance of difference between the distributions. The levelof significance was determined to be 0.05.
A q-q plot is a graphical display where the quantiles of the dis-tributions of the two variables are plotted in two dimensions. If thetwo distributions are equal, the two quantiles will lie on a straightline. Discrepancies from a straight line will point to specific dif-ferences in the distributions. The two-sample Kolmogorov-Smirnov test gives an overall comparison of the two distributions.The comparison is based on the maximum of the vertical distancesbetween the two distributions and gives no specific indication ofwhere the differences between the distributions are.
Control experiments
In addition to the transfection experiments (see above), control ex-periments included replacement of the antibodies with non-immune IgG or pre-adsorption with the peptide used for immuni-zation. Such controls were performed for Western blot, immuno-fluorescence, and electron-microscopic experiments.
Results
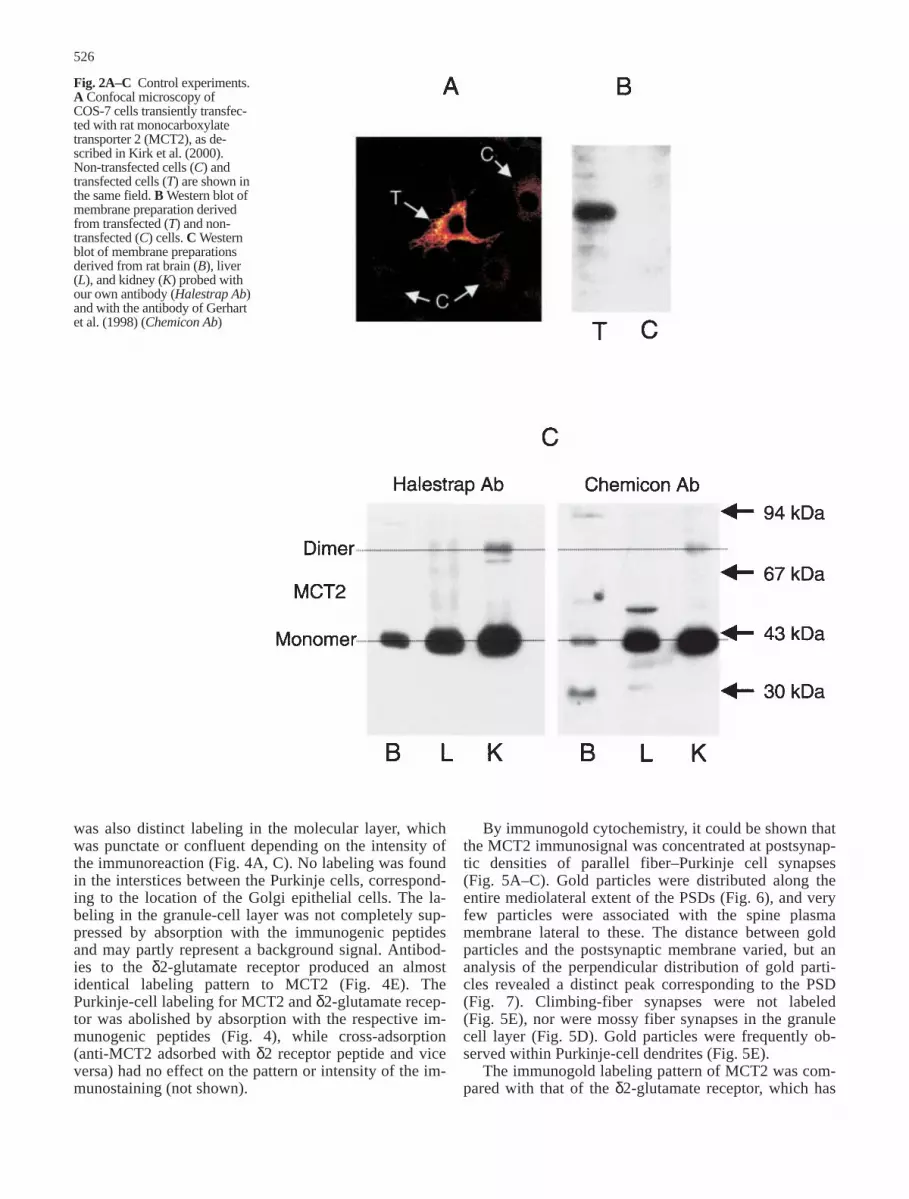
Cells that had been transfected with MCT2 showedstrong immunolabeling with the MCT2 antibody, where-as control cells were unlabelled (Fig. 2A). Immunoblotsof membrane preparations from the transfected cellsshowed a single band (Fig. 2B).
The blots that had been prepared from membranepreparations of rat brain, liver, and kidney (Fig. 2C) alsoshowed single bands, except for a weak additional bandin the kidney fraction. The latter band had a molecularweight of about 80 kDa, suggesting that it was due to di-merization. The results obtained with our own antibodywere compared with those obtained with the antibody ofGerhart et al. (1998). The latter antibody produced someextra bands, notably in the brain and liver (Fig. 2C).
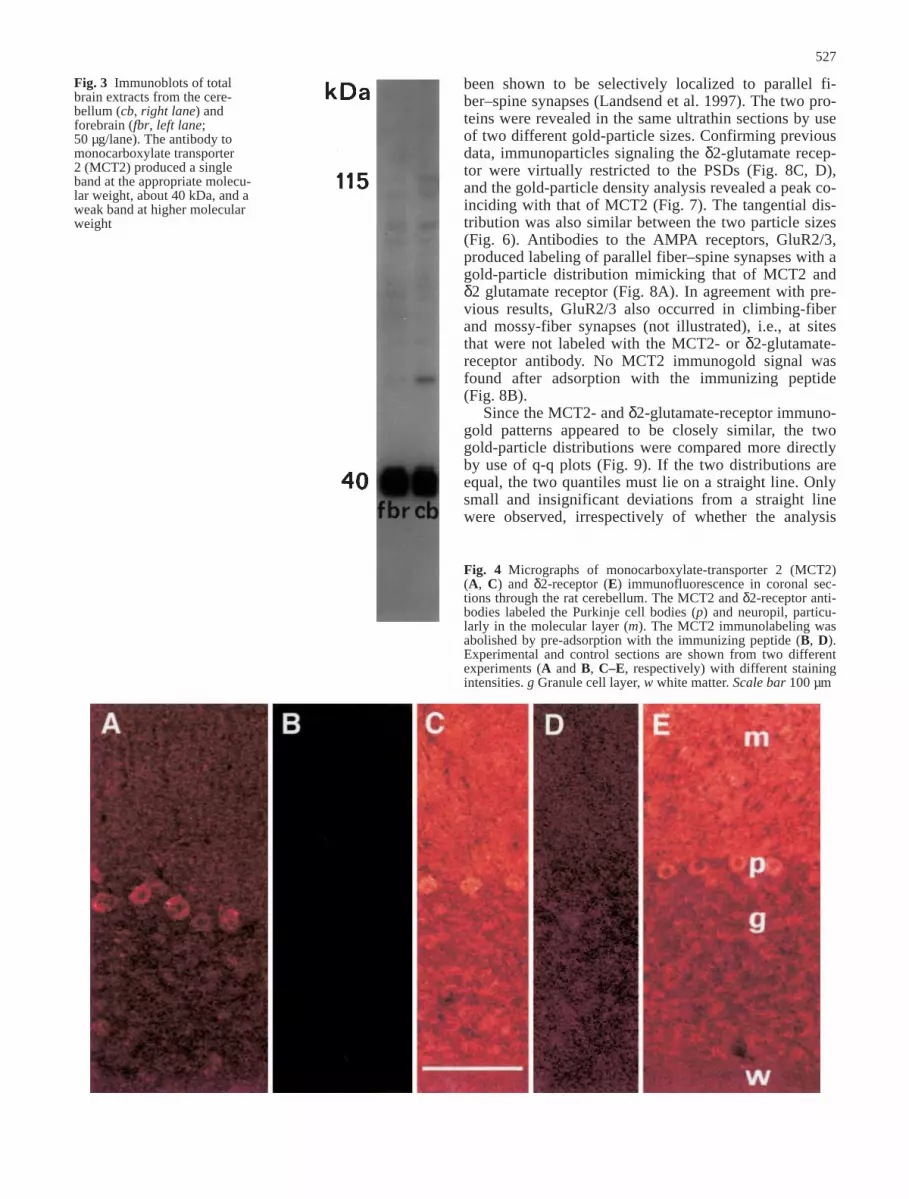
Immunoblots of total brain extracts (Fig. 3) revealed asingle, strong MCT2-immunoreactive band with an ap-proximate MW of about 40 kDa in both the cerebellumand forebrain. This corresponds to the estimated size ofthe cloned MCT2 transporter (Jackson et al. 1997). Aweak band was observed at a higher molecular weight(Fig. 3). The peptide block of the MCT2 antibody result-ed in no significant labeling (data not shown)
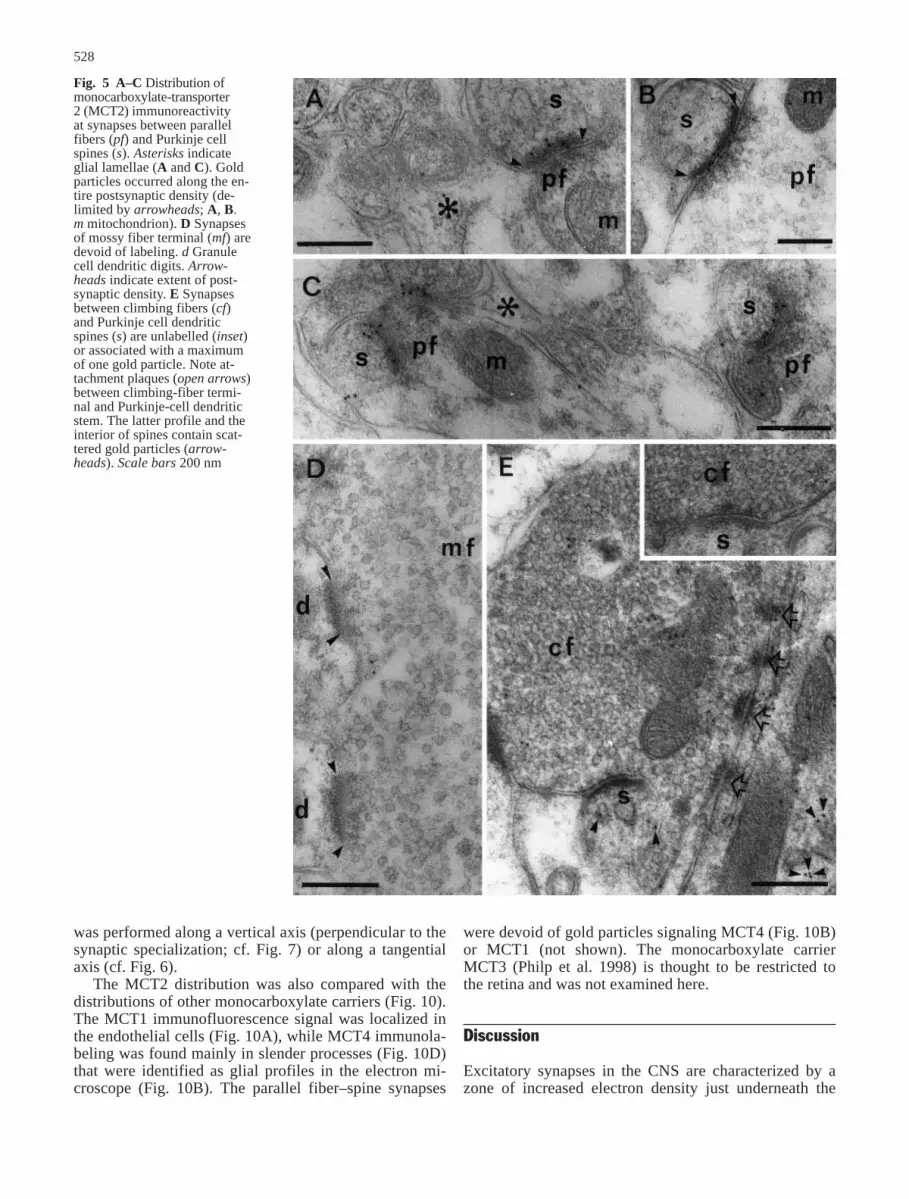
Confocal immunofluorescence microscopy showedMCT2 labeling of Purkinje cell bodies (Fig. 4). There
525
Fig. 1 Upper part: Sequence alignment of the C-termini of differ-ent monocarboxylate transporter (MCT) isoforms and proteinsknown to be associated with postsynaptic specializations of paral-lel fiber–Purkinje cell synapses. [Kir2.1 is also included; this K+
channel has been demonstrated in Purkinje cells, but its synapticlocalization remains to be determined (Miyashita and Kubo1997)]. Lower part: MCT4 and MCT1 (bottom part of figure)have extrasynaptic localizations in the molecular layer of the cere-bellum. The terminal TXV/SXV sequence of the latter MCT iso-forms is known to interact with PSD 95 in other systems. PSD 95is not expressed by Purkinje cells
was also distinct labeling in the molecular layer, whichwas punctate or confluent depending on the intensity ofthe immunoreaction (Fig. 4A, C). No labeling was foundin the interstices between the Purkinje cells, correspond-ing to the location of the Golgi epithelial cells. The la-beling in the granule-cell layer was not completely sup-pressed by absorption with the immunogenic peptidesand may partly represent a background signal. Antibod-ies to the δ2-glutamate receptor produced an almostidentical labeling pattern to MCT2 (Fig. 4E). The Purkinje-cell labeling for MCT2 and δ2-glutamate recep-tor was abolished by absorption with the respective im-munogenic peptides (Fig. 4), while cross-adsorption (anti-MCT2 adsorbed with δ2 receptor peptide and viceversa) had no effect on the pattern or intensity of the im-munostaining (not shown).
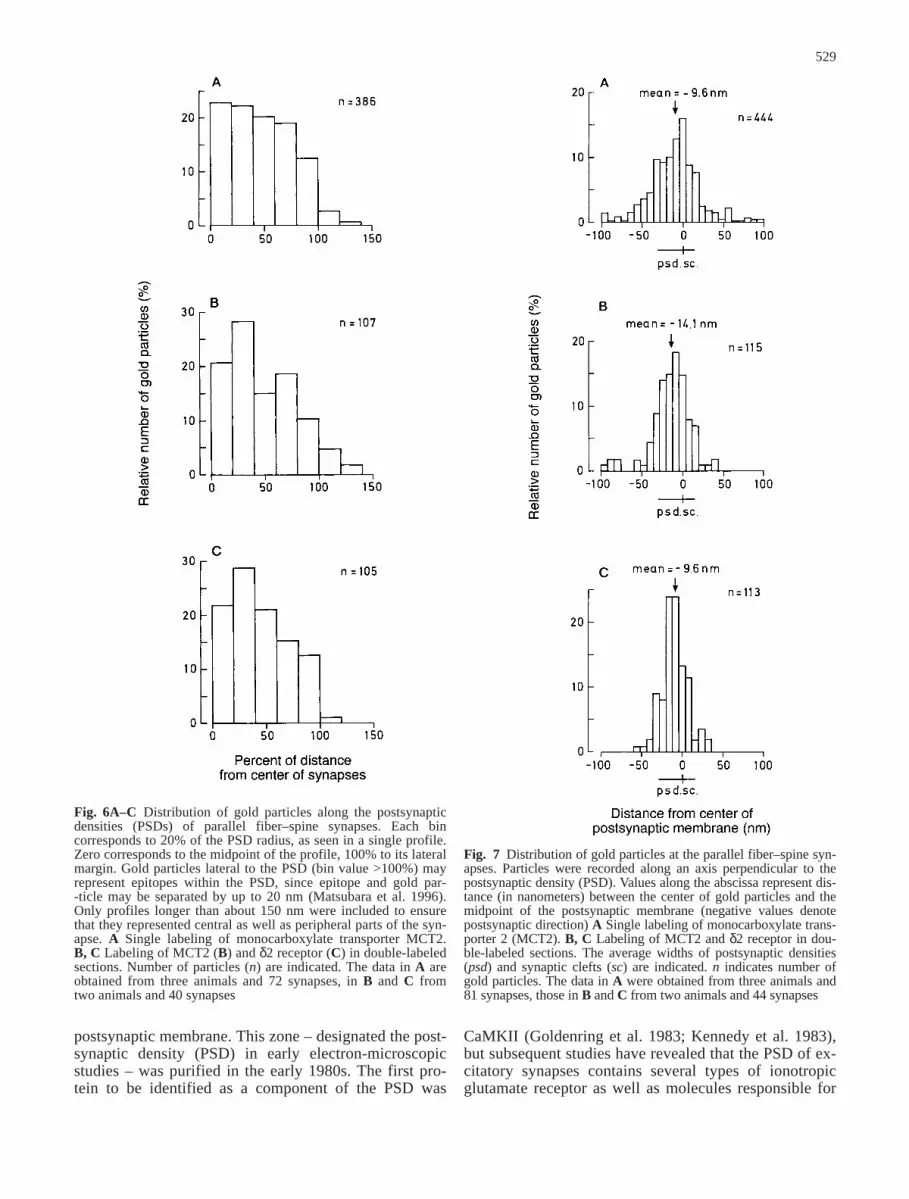
By immunogold cytochemistry, it could be shown thatthe MCT2 immunosignal was concentrated at postsynap-tic densities of parallel fiber–Purkinje cell synapses(Fig. 5A–C). Gold particles were distributed along theentire mediolateral extent of the PSDs (Fig. 6), and veryfew particles were associated with the spine plasmamembrane lateral to these. The distance between goldparticles and the postsynaptic membrane varied, but ananalysis of the perpendicular distribution of gold parti-cles revealed a distinct peak corresponding to the PSD(Fig. 7). Climbing-fiber synapses were not labeled(Fig. 5E), nor were mossy fiber synapses in the granulecell layer (Fig. 5D). Gold particles were frequently ob-served within Purkinje-cell dendrites (Fig. 5E).
The immunogold labeling pattern of MCT2 was com-pared with that of the δ2-glutamate receptor, which has
526
Fig. 2A–C Control experiments.A Confocal microscopy of COS-7 cells transiently transfec-ted with rat monocarboxylatetransporter 2 (MCT2), as de-scribed in Kirk et al. (2000).Non-transfected cells (C) andtransfected cells (T) are shown inthe same field. B Western blot ofmembrane preparation derivedfrom transfected (T) and non-transfected (C) cells. C Westernblot of membrane preparationsderived from rat brain (B), liver(L), and kidney (K) probed withour own antibody (Halestrap Ab)and with the antibody of Gerhartet al. (1998) (Chemicon Ab)
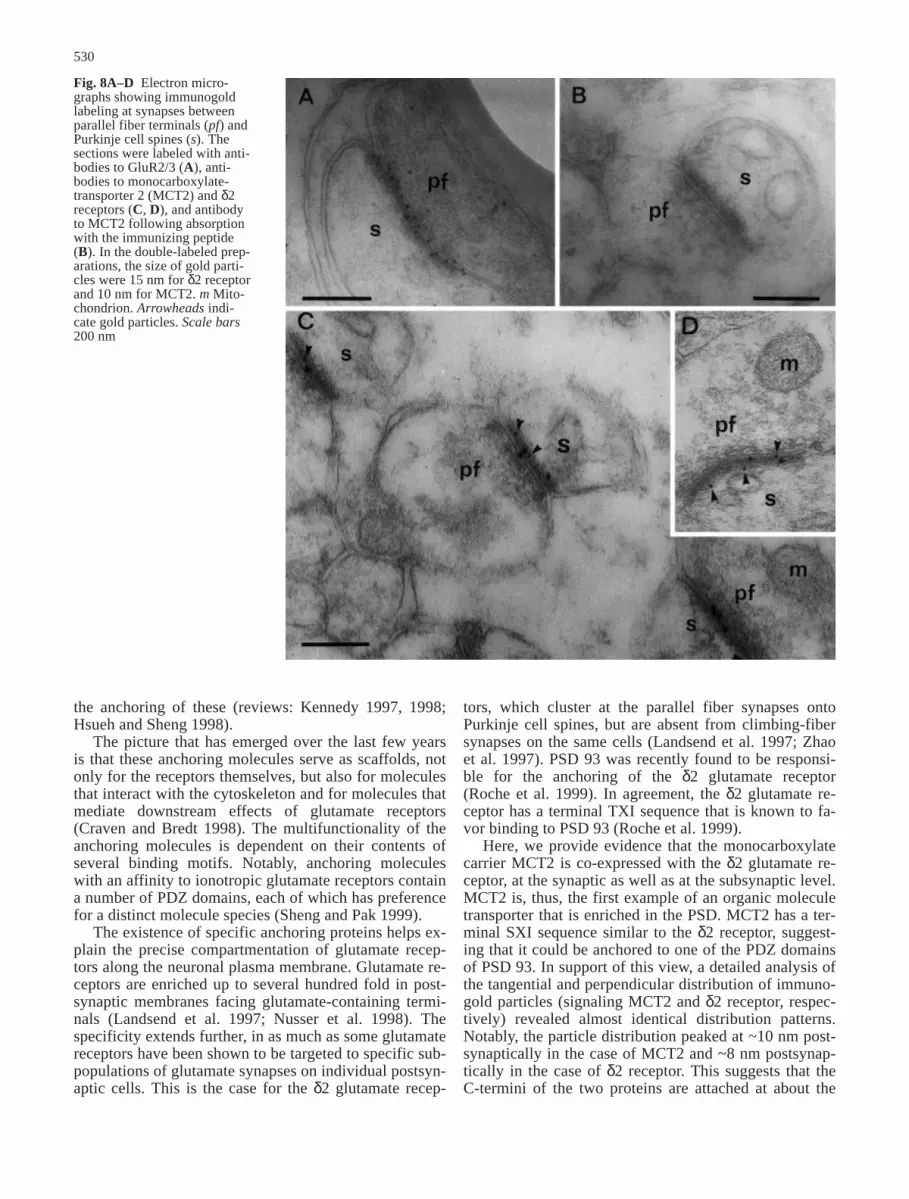
been shown to be selectively localized to parallel fi-ber–spine synapses (Landsend et al. 1997). The two pro-teins were revealed in the same ultrathin sections by useof two different gold-particle sizes. Confirming previousdata, immunoparticles signaling the δ2-glutamate recep-tor were virtually restricted to the PSDs (Fig. 8C, D),and the gold-particle density analysis revealed a peak co-inciding with that of MCT2 (Fig. 7). The tangential dis-tribution was also similar between the two particle sizes(Fig. 6). Antibodies to the AMPA receptors, GluR2/3,produced labeling of parallel fiber–spine synapses with agold-particle distribution mimicking that of MCT2 andδ2 glutamate receptor (Fig. 8A). In agreement with pre-vious results, GluR2/3 also occurred in climbing-fiberand mossy-fiber synapses (not illustrated), i.e., at sitesthat were not labeled with the MCT2- or δ2-glutamate-receptor antibody. No MCT2 immunogold signal wasfound after adsorption with the immunizing peptide(Fig. 8B).
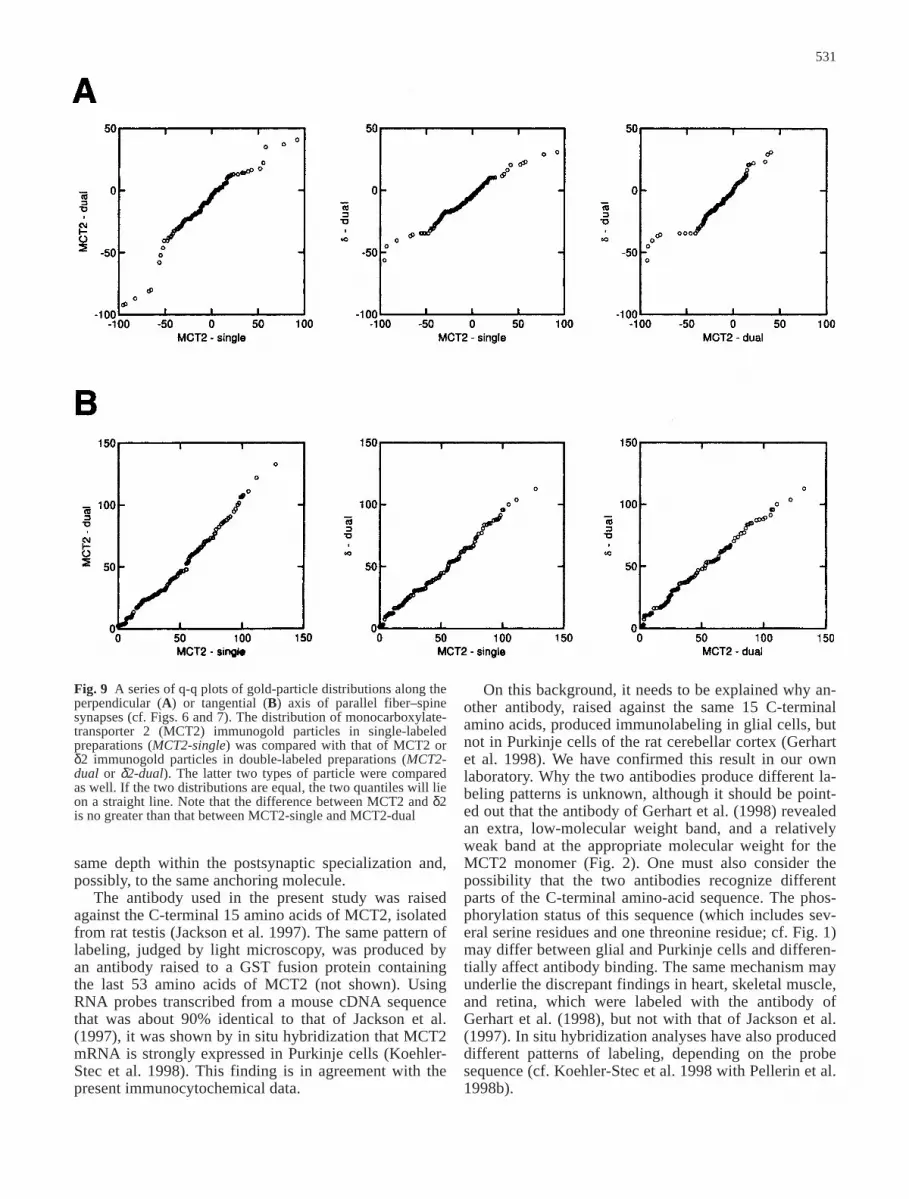
Since the MCT2- and δ2-glutamate-receptor immuno-gold patterns appeared to be closely similar, the twogold-particle distributions were compared more directlyby use of q-q plots (Fig. 9). If the two distributions areequal, the two quantiles must lie on a straight line. Onlysmall and insignificant deviations from a straight linewere observed, irrespectively of whether the analysis
527
Fig. 3 Immunoblots of totalbrain extracts from the cere-bellum (cb, right lane) andforebrain (fbr, left lane;50 µg/lane). The antibody tomonocarboxylate transporter 2 (MCT2) produced a singleband at the appropriate molecu-lar weight, about 40 kDa, and aweak band at higher molecularweight
Fig. 4 Micrographs of monocarboxylate-transporter 2 (MCT2)(A, C) and δ2-receptor (E) immunofluorescence in coronal sec-tions through the rat cerebellum. The MCT2 and δ2-receptor anti-bodies labeled the Purkinje cell bodies (p) and neuropil, particu-larly in the molecular layer (m). The MCT2 immunolabeling wasabolished by pre-adsorption with the immunizing peptide (B, D).Experimental and control sections are shown from two differentexperiments (A and B, C–E, respectively) with different stainingintensities. g Granule cell layer, w white matter. Scale bar 100 µm
was performed along a vertical axis (perpendicular to thesynaptic specialization; cf. Fig. 7) or along a tangentialaxis (cf. Fig. 6).
The MCT2 distribution was also compared with thedistributions of other monocarboxylate carriers (Fig. 10).The MCT1 immunofluorescence signal was localized inthe endothelial cells (Fig. 10A), while MCT4 immunola-beling was found mainly in slender processes (Fig. 10D)that were identified as glial profiles in the electron mi-croscope (Fig. 10B). The parallel fiber–spine synapses
were devoid of gold particles signaling MCT4 (Fig. 10B)or MCT1 (not shown). The monocarboxylate carrierMCT3 (Philp et al. 1998) is thought to be restricted tothe retina and was not examined here.
Discussion
Excitatory synapses in the CNS are characterized by azone of increased electron density just underneath the
528
Fig. 5 A–C Distribution ofmonocarboxylate-transporter 2 (MCT2) immunoreactivity at synapses between parallel fibers (pf) and Purkinje cellspines (s). Asterisks indicateglial lamellae (A and C). Goldparticles occurred along the en-tire postsynaptic density (de-limited by arrowheads; A, B.m mitochondrion). D Synapsesof mossy fiber terminal (mf) aredevoid of labeling. d Granulecell dendritic digits. Arrow-heads indicate extent of post-synaptic density. E Synapsesbetween climbing fibers (cf)and Purkinje cell dendriticspines (s) are unlabelled (inset)or associated with a maximumof one gold particle. Note at-tachment plaques (open arrows)between climbing-fiber termi-nal and Purkinje-cell dendriticstem. The latter profile and theinterior of spines contain scat-tered gold particles (arrow-heads). Scale bars 200 nm
postsynaptic membrane. This zone – designated the post-synaptic density (PSD) in early electron-microscopicstudies – was purified in the early 1980s. The first pro-tein to be identified as a component of the PSD was
CaMKII (Goldenring et al. 1983; Kennedy et al. 1983),but subsequent studies have revealed that the PSD of ex-citatory synapses contains several types of ionotropicglutamate receptor as well as molecules responsible for
529
Fig. 6A–C Distribution of gold particles along the postsynapticdensities (PSDs) of parallel fiber–spine synapses. Each bin corresponds to 20% of the PSD radius, as seen in a single profile.Zero corresponds to the midpoint of the profile, 100% to its lateralmargin. Gold particles lateral to the PSD (bin value >100%) mayrepresent epitopes within the PSD, since epitope and gold par--ticle may be separated by up to 20 nm (Matsubara et al. 1996).Only profiles longer than about 150 nm were included to ensurethat they represented central as well as peripheral parts of the syn-apse. A Single labeling of monocarboxylate transporter MCT2.B, C Labeling of MCT2 (B) and δ2 receptor (C) in double-labeledsections. Number of particles (n) are indicated. The data in A areobtained from three animals and 72 synapses, in B and C fromtwo animals and 40 synapses
Fig. 7 Distribution of gold particles at the parallel fiber–spine syn-apses. Particles were recorded along an axis perpendicular to thepostsynaptic density (PSD). Values along the abscissa represent dis-tance (in nanometers) between the center of gold particles and themidpoint of the postsynaptic membrane (negative values denotepostsynaptic direction) A Single labeling of monocarboxylate trans-porter 2 (MCT2). B, C Labeling of MCT2 and δ2 receptor in dou-ble-labeled sections. The average widths of postsynaptic densities(psd) and synaptic clefts (sc) are indicated. n indicates number ofgold particles. The data in A were obtained from three animals and81 synapses, those in B and C from two animals and 44 synapses
the anchoring of these (reviews: Kennedy 1997, 1998;Hsueh and Sheng 1998).
The picture that has emerged over the last few yearsis that these anchoring molecules serve as scaffolds, notonly for the receptors themselves, but also for moleculesthat interact with the cytoskeleton and for molecules thatmediate downstream effects of glutamate receptors (Craven and Bredt 1998). The multifunctionality of theanchoring molecules is dependent on their contents ofseveral binding motifs. Notably, anchoring moleculeswith an affinity to ionotropic glutamate receptors containa number of PDZ domains, each of which has preferencefor a distinct molecule species (Sheng and Pak 1999).
The existence of specific anchoring proteins helps ex-plain the precise compartmentation of glutamate recep-tors along the neuronal plasma membrane. Glutamate re-ceptors are enriched up to several hundred fold in post-synaptic membranes facing glutamate-containing termi-nals (Landsend et al. 1997; Nusser et al. 1998). Thespecificity extends further, in as much as some glutamatereceptors have been shown to be targeted to specific sub-populations of glutamate synapses on individual postsyn-aptic cells. This is the case for the δ2 glutamate recep-
tors, which cluster at the parallel fiber synapses ontoPurkinje cell spines, but are absent from climbing-fibersynapses on the same cells (Landsend et al. 1997; Zhaoet al. 1997). PSD 93 was recently found to be responsi-ble for the anchoring of the δ2 glutamate receptor(Roche et al. 1999). In agreement, the δ2 glutamate re-ceptor has a terminal TXI sequence that is known to fa-vor binding to PSD 93 (Roche et al. 1999).
Here, we provide evidence that the monocarboxylatecarrier MCT2 is co-expressed with the δ2 glutamate re-ceptor, at the synaptic as well as at the subsynaptic level.MCT2 is, thus, the first example of an organic moleculetransporter that is enriched in the PSD. MCT2 has a ter-minal SXI sequence similar to the δ2 receptor, suggest-ing that it could be anchored to one of the PDZ domainsof PSD 93. In support of this view, a detailed analysis ofthe tangential and perpendicular distribution of immuno-gold particles (signaling MCT2 and δ2 receptor, respec-tively) revealed almost identical distribution patterns.Notably, the particle distribution peaked at ~10 nm post-synaptically in the case of MCT2 and ~8 nm postsynap-tically in the case of δ2 receptor. This suggests that theC-termini of the two proteins are attached at about the
530
Fig. 8A–D Electron micro-graphs showing immunogoldlabeling at synapses betweenparallel fiber terminals (pf) andPurkinje cell spines (s). Thesections were labeled with anti-bodies to GluR2/3 (A), anti-bodies to monocarboxylate-transporter 2 (MCT2) and δ2receptors (C, D), and antibodyto MCT2 following absorptionwith the immunizing peptide(B). In the double-labeled prep-arations, the size of gold parti-cles were 15 nm for δ2 receptorand 10 nm for MCT2. m Mito-chondrion. Arrowheads indi-cate gold particles. Scale bars200 nm
same depth within the postsynaptic specialization and,possibly, to the same anchoring molecule.
The antibody used in the present study was raisedagainst the C-terminal 15 amino acids of MCT2, isolatedfrom rat testis (Jackson et al. 1997). The same pattern oflabeling, judged by light microscopy, was produced byan antibody raised to a GST fusion protein containingthe last 53 amino acids of MCT2 (not shown). UsingRNA probes transcribed from a mouse cDNA sequencethat was about 90% identical to that of Jackson et al.(1997), it was shown by in situ hybridization that MCT2mRNA is strongly expressed in Purkinje cells (Koehler-Stec et al. 1998). This finding is in agreement with thepresent immunocytochemical data.
On this background, it needs to be explained why an-other antibody, raised against the same 15 C-terminalamino acids, produced immunolabeling in glial cells, butnot in Purkinje cells of the rat cerebellar cortex (Gerhartet al. 1998). We have confirmed this result in our ownlaboratory. Why the two antibodies produce different la-beling patterns is unknown, although it should be point-ed out that the antibody of Gerhart et al. (1998) revealedan extra, low-molecular weight band, and a relativelyweak band at the appropriate molecular weight for theMCT2 monomer (Fig. 2). One must also consider thepossibility that the two antibodies recognize differentparts of the C-terminal amino-acid sequence. The phos-phorylation status of this sequence (which includes sev-eral serine residues and one threonine residue; cf. Fig. 1)may differ between glial and Purkinje cells and differen-tially affect antibody binding. The same mechanism mayunderlie the discrepant findings in heart, skeletal muscle,and retina, which were labeled with the antibody of Gerhart et al. (1998), but not with that of Jackson et al.(1997). In situ hybridization analyses have also produceddifferent patterns of labeling, depending on the probe sequence (cf. Koehler-Stec et al. 1998 with Pellerin et al.1998b).
531
Fig. 9 A series of q-q plots of gold-particle distributions along theperpendicular (A) or tangential (B) axis of parallel fiber–spinesynapses (cf. Figs. 6 and 7). The distribution of monocarboxylate-transporter 2 (MCT2) immunogold particles in single-labeledpreparations (MCT2-single) was compared with that of MCT2 orδ2 immunogold particles in double-labeled preparations (MCT2-dual or δ2-dual). The latter two types of particle were comparedas well. If the two distributions are equal, the two quantiles will lieon a straight line. Note that the difference between MCT2 and δ2is no greater than that between MCT2-single and MCT2-dual

On the basis of the above data, one cannot excludethe possibility that the glial cells express a pool ofMCT2 that is not visualized by the Jackson antibodyused here. However, our findings suggest that MCT4 is amajor monocarboxylate transporter in cerebellar astro-cytes, while MCT2 is the predominant monocarboxylatetransporter in Purkinje cells. The possibility remains thatboth cell types contain additional transporters, whichwere not included among those that were investigated inthe present study.
MCT4 is known to be responsible for the export oflactate from glycolytic muscle fibers (Juel and Halestrap1999) and may have an analogous function in the brain.There is ample evidence that there is a flux of lactatefrom glial cells to neurons (Pellerin et al. 1998a). MCT4and MCT2 could act in sequence to facilitate lactatetransport between the Bergmann glial processes and thePurkinje cells that they ensheath. The Purkinje cell den-drites have an extraordinary high density of mitochon-dria (Palay and Chan-Palay 1974), pointing to an activeoxidative metabolism, and the high affinity of MCT2 forlactate (Broer et al. 1999) make it ideally suited for up-take of this metabolic fuel. The surrounding glial pro-
cesses, in contrast, contain few mitochondria and maydepend largely on glycolytic pathways.
Glutamate uptake is coupled to a stoichiometric up-take of H+ (Danbolt 1994). This will rapidly create a def-icit of H+ in the synaptic cleft, which has to be compen-sated for by outward movement of H+ from surroundingcellular compartments. In the case of glia, the extracellu-lar deficit of H+ could energize outward lactate/H+ trans-port, through MCT4, essentially leading to an exchangeof glutamate with lactate across the plasma membrane. Ithas also been proposed that part of the glutamate couldbe metabolically converted to lactate after uptake intoastrocytes (Hassel and Sonnewald 1995) and that glialexport of lactate would be favored by the increased gly-colytic rate imposed by glutamate uptake (Pellerin andMagistretti 1994, 1997). Lactate derived from glia couldbe an important metabolic substrate for neurons (re-viewed by Magistretti et al. 1999).
In the case of the Purkinje cells, the extracellular H+
deficit could drive the Na+/H+ exchange. Neurons areknown to express several subtypes of Na+/H+ antiporters(Attaphitaya et al. 1999), and one of these (NHE3) is al-most exclusively localized to Purkinje cells (Ma andHaddad 1997). Exchange of H+ for Na+ could allowMCT2 to operate in the inward direction (i.e., along thesupposed concentration gradient for lactate), allowingthe glial-derived lactate to be accumulated in the den-dritic spines and stems.
If MCT2 serves as a point of entry of an energy sub-strate, why is it concentrated at the synapse? The spatial
532
Fig. 10 Comparison between the immunofluorescence patternsfor monocarboxylate transporter 1 (MCT1) (A), MCT2 (C), andMCT4 (D). MCT4 occurred in slender processes, which wereidentified as astrocyte profiles (asterisks) by immunogold electronmicroscopy (B). MCT1 was concentrated in the endothelial cells.Abbreviations as in Fig. 4. Scale bar 50 µm in A, C, D and200 nm in B

coupling between glutamate exocytosis, glutamate up-take, and lactate transport provides a unique way of ad-justing energy supply to excitatory drive. Uptake of glu-tamate by the glutamate transporter EAAT1, which is en-riched at those astrocyte-membrane domains that facethe parallel fiber–spine synapse (Chaudhry et al. 1995),will directly and indirectly promote lactate releasethrough MCT4, which is enriched in the same membranedomains (cf. Fig. 10B). Released lactate will then betransported into the Purkinje-cell spines by the synapticMCT2 molecules. Thus, the more glutamate that is re-leased from the parallel fibers, the more lactate will bereleased from the glial processes and be available forpostsynaptic uptake. Glutamate – itself an energy sub-strate – may also be taken up directly into the postsynap-tic Purkinje cells, through EAAT4 (which is concentratedin the spine membrane lateral to the PSD; Dehnes et al.1998) and through EAAT3 (demonstrated in Purkinjecells by enzyme-based immunocytochemistry; Rothsteinet al. 1995). We are thus beginning to realize that thesynapse not only serves as a site of interneuronal signaltransfer – it may also act as an important site of meta-bolic coupling.
Acknowledgements We thank Bjørg Riber, Karen M. Gujord,Gunnar Lothe, Carina Knudsen, and Kari Ruud for technical assis-tance, and Dr. M. Krestchatisky for helpful discussions. This workwas supported by the Norwegian Research Council and ProfessorLetten F. Saugstad’s Fund.
References
Attaphitaya S, Park K, Melvin JE (1999) Molecular cloning andfunctional expression of a rat Na+/H+ exchanger (NHE5) high-ly expressed in brain. J Biol Chem 274:4383–4388
Broer S, Broer A, Schneider HP, Stegen C, Halestrap AP, DeitmerJW (1999) Characterization of the high-affinity monocarbox-ylate transporter MCT2 in Xenopus laevis oocytes. Biochem J 341:529–535
Chaudhry FA, Lehre KP, Van Lookeren Campagne M, OttersenOP, Danbolt NC, Storm-Mathisen J (1995) Glutamate trans-porters in glial plasma membranes: highly differentiated local-izations revealed by quantitative ultrastructural immunocyto-chemistry. Neuron 15:711–720
Craven SE, Bredt DS (1998) PDZ proteins organize synaptic sig-naling pathways. Cell 93:495–498
Danbolt NC (1994) The high affinity uptake system for excitatoryamino acids in the brain. Prog Neurobiol 44:377–396
Dehnes Y, Chaudhry FA, Ullensvang K, Lehre KP, Storm-Mathisen J, Danbolt NC (1998) The glutamate transporterEAAT4 in rat cerebellar Purkinje cells: a glutamate-gatedchloride channel concentrated near the synapse in parts of thedendritic membrane facing astroglia. J Neurosci 18:3606–3619
Garcia CK, Li X, Luna J, Francke U (1994) cDNA cloning of thehuman monocarboxylate transporter 1 and chromosomal local-ization of the SLC16A1 locus to 1p13.2-p12. Genomics 23:500–503
Garcia CK, Brown MS, Pathak RK, Goldstein JL (1995) cDNAcloning of MCT2, a second monocarboxylate transporter ex-pressed in different cells than MCT1. J Biol Chem 270:1843–1849
Gerhart DZ, Enerson BE, Zhdankina OY, Leino RL, Drewes LR(1997) Expression of monocarboxylate transporter MCT1 bybrain endothelium and glia in adult and suckling rats. Am J Physiol 273:207–213
Gerhart DZ, Enerson BE, Zhdankina OY, Leino RL, Drewes LR(1998) Expression of the monocarboxylate transporter MCT2by rat brain glia. Glia 22:272–281
Goldenring JR, Gonzalez B, McGuire JS Jr., DeLorenzo RJ (1983)Purification and characterization of a calmodulin-dependentkinase from rat brain cytosol able to phosphorylate tubulin andmicrotubule-associated proteins. J Biol Chem 258:12632–12640
Halestrap AP, Price NT (1993) Transport of lactate and othermonocarboxylates across mammalian plasma membranes. AmJ Physiol 264:761–782
Halestrap AP, Price NT (1999) The proton-linked monocarboxyl-ate transporter (MCT) family: structure, function and regula-tion. Biochem J 343:281–299
Han XX, Handberg A, Petersen LN, Ploug T, Galbo H (1995) Sta-bility of GLUT-1 and GLUT-4 expression in perfused rat mus-cle stimulated by insulin and exercise. J Appl Physiol 78:46–52
Hassel B, Sonnewald U (1995) Glial formation of pyruvate andlactate from TCA cycle intermediates: implications for the in-activation of transmitter amino acids? J Neurochem 65:2227–2234
Hjelle OP, Chaudhry FA, Ottersen OP (1994) Antisera to glutathi-one: characterization and immunocytochemical application tothe rat cerebellum. Eur J Neurosci 6:793–804
Hsueh YP, Sheng M (1998) Anchoring of glutamate receptors atthe synapse. Prog Brain Res 116:123–131
Jackson VN, Price NT, Carpenter L, Halestrap AP (1997) Cloningof the monocarboxylate transporter isoform MCT2 from rattestis provides evidence that expression in tissues is species-specific and may involve post-transcriptional regulation. Bio-chem J 324:447–453
Juel C, Halestrap AP (1999) Lactate transport in skeletal muscle –role and regulation of the monocarboxylate transporter. J Physiol 517:633–642
Kennedy MB (1997) The postsynaptic density at glutamatergicsynapses. Trends Neurosci 20:264–268
Kennedy MB (1998) Signal transduction molecules at the gluta-matergic postsynaptic membrane. Brain Res Brain Res Rev26:243–257
Kennedy MB, Bennett MK, Erondu NE (1983) Biochemical andimmunochemical evidence that the “major postsynaptic densi-ty protein” is a subunit of a calmodulin-dependent protein ki-nase. Proc Natl Acad Sci USA 80:7357–7361
Kirk P, Wilson MC, Heddle C, Brown MH, Barclay AN, HalestrapAP (2000) CD147 is tightly associated with lactate transport-ers MCT1 and MCT4 and facilitates their cell surface expres-sion. EMBO J 19:3896–3904
Koehler-Stec EM, Simpson IA, Vannucci SJ, Landschulz KT,Landschulz WH (1998) Monocarboxylate transporter expres-sion in mouse brain. Am J Physiol 275:516–524
Landsend AS, Amiry-Moghaddam M, Matsubara A, Bergersen L,Usami S, Wenthold RJ, Ottersen OP (1997) Differential local-ization of delta glutamate receptors in the rat cerebellum: co-expression with AMPA receptors in parallel fiber–spine synap-ses and absence from climbing fiber–spine synapses. J Neuro-sci 17:834–842
Ma E, Haddad GG (1997) Expression and localization of Na+/H+
exchangers in rat central nervous system. Neuroscience 79:591–603
Magistretti PJ, Pellerin L, Rothman DL, Shulman RG (1999) Energy on demand. Science 283:496–497
Matsubara A, Laake JH, Davanger S, Usami S, Ottersen OP(1996) Organization of AMPA receptor subunits at a glutamatesynapse: a quantitative immunogold analysis of hair cell syn-apses in the rat organ of Corti. J Neurosci 16:4457–4467
Mayat E, Petralia RS, Wang YX, Wenthold RJ (1995) Immunopre-cipitation, immunoblotting, and immunocytochemistry studiessuggest that glutamate receptor delta subunits form novel post-synaptic receptor complexes. J Neurosci 15:2533–2546
McCullagh KJ, Poole RC, Halestrap AP, O’Brien M, Bonen A(1996) Role of the lactate transporter (MCT1) in skeletal muscles. Am J Physiol 271:143–150
533

Miyashita T, Kubo Y (1997) Localization and developmentalchanges of the expression of two inwardrectifying K+-channelproteins in the rat brain. Brain Res 750:251–263
Nusser Z, Sieghart W, Somogyi P (1998) Segregation of differentGABAA receptors to synaptic and extrasynaptic membranesof cerebellar granule cells. J Neurosci 18:1693–1703
Ottersen OP, Zhang N, Walberg F (1992) Metabolic compartmen-tation of glutamate and glutamine: morphological evidenceobtained by quantitative immunocyto-chemistry in rat cerebel-lum. Neuroscience 46:519–534
Palay SL, Chan-Palay V (1974) Cerebellar cortex: cytology andorganization. Springer, Berlin Heidelberg New York
Pellerin L, Magistretti PJ (1994) Glutamate uptake into astrocytesstimulates aerobic glycolysis: a mechanism coupling neuronalactivity to glucose utilization. Proc Natl Acad Sci USA 91:10625–10629
Pellerin L, Magistretti PJ (1997) Glutamate uptake stimulatesNa+,K+-ATPase activity in astrocytes via activation of a dis-tinct subunit highly sensitive to ouabain. J Neurochem 69:2132–2137
Pellerin L, Pellegri G, Bittar PG, Charnay Y, Bouras C, Martin JL,Stella N, Magistretti PJ (1998a) Evidence supporting the exis-tence of an activity-dependent astrocyte-neuron lactate shuttle.Dev Neurosci 20:291–299
Pellerin L, Pellegri G, Martin JL, Magistretti PJ (1998b) Expres-sion of mono-carboxylate transporter mRNAs in mouse brain:support for a distinct role of lactate as an energy substrate forthe neonatal vs. adult brain. Proc Natl Acad Sci USA 95:3990–3995
Philp NJ, Yoon H, Grollman EF (1998) Monocarboxylate trans-porter MCT1 is located in the apical membrane and MCT3 inthe basal membrane of rat RPE. Am J Physiol 274:1824–1828
Poitry-Yamate CL, Poitry S, Tsacopoulos M (1995) Lactate re-leased by Muller glial cells is metabolized by photoreceptorsfrom mammalian retina. J Neurosci 15:5179–5191
Poole RC, Sansom CE, Halestrap AP (1996) Studies of the mem-brane topology of the rat erythrocyte H+/lactate cotransporter(MCT1). Biochem J 320:817–824
Roche KW, Ly CD, Petralia RS, Wang YX, McGee AW, Bredt DS,Wenthold RJ (1999) Postsynaptic density-93 interacts with thedelta2 glutamate receptor subunit at parallel fiber synapses. JNeurosci 19:3926–3934
Rothstein JD, Van Kammen M, Levey AI, Martin LJ, Kuncl RW(1995) Selective loss of glial glutamate transporter GLT-1 inamyotrophic lateral sclerosis. Ann Neurol 38:73–84
Sheng M, Pak DT (1999) Glutamate receptor anchoring proteinsand the molecular organization of excitatory synapses. Ann N Y Acad Sci 868:483–493
Van Lookeren Campagne M, Oestreicher AB, Krift TP van der,Gispen WH, Verkleij AJ (1991) Freeze-substitution and Lowicryl HM20 embedding of fixed rat brain: suitability forimmunogold ultrastructural localization of neural antigens. JHistochem Cytochem 39:1267–1279
Veruki ML, Wässle H (1996) Immunohistochemistry localizationof dopamine D1 receptors in rat retina. Eur J Neurosci 8:2286–2297
Wang BL, Larsson LI (1985) Simultaneous demonstration of mul-tiple antigens by indirect immunofluorescence or immunogoldstaining. Novel light and electron microscopical double andtriple staining method employing primary antibodies from thesame species. Histochemistry 83:47–56
Wenthold RJ, Yokotani N, Doi K, Wada K (1992) Immunochemi-cal characterization of the non-NMDA glutamate receptor using subunit-specific antibodies. Evidence for a hetero-oligomeric structure in rat brain. J Biol Chem 267:501–507
Wilson MC, Jackson VN, Heddle C, Price NT, Pilegaard H, JuelC., Bonen A, Hutter OF, Halestrap AP (1998) Lactic acid ef-flux from white skeletal muscle is catalyzed by the monocar-boxylate transporter isoform MCT3. J Biol Chem 273:15920–15926
Zhao HM, Wenthold RJ, Wang YX, Petralia RS (1997) Delta-glutamate receptors are differentially distributed at paralleland climbing fiber synapses on Purkinje cells. J Neurochem68:1041–1052
534