bursting in substantia nigra pars reticulata neurons in vitro: possible relevance for parkinson...
TRANSCRIPT

Bursting in Substantia Nigra Pars Reticulata Neurons In Vitro: PossibleRelevance for Parkinson Disease
Osvaldo Ibanez-Sandoval,1 Luis Carrillo-Reid,1 Elvira Galarraga,1 Dagoberto Tapia,1 Ernesto Mendoza,1
Juan C. Gomora,1 Jorge Aceves,2 and Jose Bargas1
1Departamento de Biofısica, Instituto de Fisiologıa Celular, Universidad Nacional Autonoma de Mexico; and 2Departamento deFisiologıa, Biofısica y Neurociencias, Centro de Investigacion y Estudios Avanzados, Mexico City, Mexico
Submitted 5 June 2007; accepted in final form 16 August 2007
Ibanez-Sandoval O, Carrillo-Reid L, Galarraga E, Tapia D, Men-doza E, Gomora JC, Aceves J, Bargas J. Bursting in substantianigra pars reticulata neurons in vitro: possible relevance for Parkinsondisease. J Neurophysiol 98: 2311–2323, 2007. First published August22, 2007; doi:10.1152/jn.00620.2007. Projection neurons of the sub-stantia nigra reticulata (SNr) convey basal ganglia (BG) processing tothalamocortical and brain stem circuits responsible for movement.Two models try to explain pathological BG performance duringParkinson disease (PD): the rate model, which posits an overexcita-tion of SNr neurons due to hyperactivity in the indirect pathway andhypoactivity of the direct pathway, and the oscillatory model, whichexplains PD as the product of pathological pattern generators dis-closed by dopamine reduction. These models are, apparently, incom-patible. We tested the predictions of the rate model by increasing theexcitatory drive and reducing the inhibition on SNr neurons in vitro.This was done pharmacologically with bath application of glutamateagonist N-methyl-D-aspartate and GABAA receptor blockers, respec-tively. Both maneuvers induced bursting behavior in SNr neurons.Therefore synaptic changes forecasted by the rate model induce theelectrical behavior predicted by the oscillatory model. In addition, wefound evidence that CaV3.2 Ca2� channels are a critical step ingenerating the bursting firing pattern in SNr neurons. Other ionchannels involved are: hyperpolarization-activated cation channels,high-voltage-activated Ca2� channels, and Ca2�-activated K� chan-nels. However, although these channels shape the temporal structureof bursting, only CaV3.2 Ca2� channels are indispensable for theinitiation of the bursting pattern.
I N T R O D U C T I O N
Together with the internal globus pallidus, neurons of thesubtantia nigra reticulata (SNr) constitute the “output” of thebasal ganglia (Albin et al. 1989; Graybiel 2004; Grillner et al.2005a; Smith et al. 1998; Wichmann and DeLong 2003). SNrneurons exhibit a tonic firing rate of activity (Atherton andBevan 2005; Richards et al. 1997) regulated by inhibitioncoming in part from the direct pathway of the basal ganglia(BG) conveyed by strionigral GABAergic afferents (Albinet al. 1989; Graybiel 2004; Smith and Bolam 1991) and also byexcitation coming from the subthalamic nucleus (STN) (Iribeet al. 1999; Nakanishi et al. 1987; Robledo and Feger 1990),the last relay of the indirect pathway. Balance between directand indirect pathways selects and unselects, respectively, mo-tor programs, by regulating the firing rate of SNr outputneurons (Graybiel 2004, 2005; Grillner et al. 2005a; Obesoet al. 2002).
The rate model of Parkinson’s disease (PD) posits that theabsence of dopamine induces, among other changes, an over-activation of the indirect pathway ending at glutamatergicsubthalamo-nigral afferents, thus leading to overexcitation ofSNr neurons. This model also proposes that PD patients havean underactivation of the GABAergic direct pathway, leadingto a decrease in inhibition from strionigral afferents (e.g., Albinet al. 1989; Day et al. 2006; Mallet et al. 2006; Murer et al.2002; Walters et al. 2007; Windels et al. 2005). The end resultof these synaptic changes is proposed to be a generalizedincrease in firing rate of SNr neurons (Albin et al. 1989; Obesoet al. 2002; Smith et al. 1998; Takakusaki et al. 2004),producing overinhibition of the nuclei in charge of movement.This would explain hypokinesia, a hallmark of PD.
In the present work, we tested this prediction of the ratemodel in SNr neurons from normal animals. The excitatorydrive on SNr neurons was increased by the application of theglutamatergic agonist: N-methyl-D-aspartate (NMDA), and adecrease in inhibition of SNr neurons was produced by block-ing their GABAA receptors.
The result of these manipulations was not just an increase infiring rate. Instead, SNr neurons entered into a bursting firingpattern (e.g., Hutchinson et al. 2004; Walters et al. 2007;Wichmann et al. 1999). However, the palette of ionic channelsthat bestow SNr neurons with the ability to generate rhythmicbursting has not been described. Therefore a preliminarysearch into the ionic mechanisms of bursting in SNr is pre-sented.
The oscillatory model of PD disease posits that dopamineabsence discloses pathologic central pattern generators (CPGs)that display unusual numbers of neurons engaged in rhythmicbursting in different BG nuclei (Hutchison et al. 2004). In fact,bursting patterns are exhibited by active CPGs (Bevan et al.2002; Grillner et al. 2005a,b; Plenz and Kitai 1999). NMDA iscapable of activating CPG circuits in vitro (e.g., Alford et al.2003; Enomoto et al. 2002; Gordon and Whelan 2006; Grillneret al. 1981; Guertin and Hounsgaard 1998; Hochman et al.1994; Hsiao et al. 1998; Kiehn 2006; Takakusaki et al. 2004).The rate and oscillatory models of PD are apparently incom-patible. Nevertheless, this work shows that simulating thesynaptic changes proposed by the rate model results in firingbehaviors similar to those proposed by the oscillatory model. Italso demonstrates that intrinsic ion channels possessed by SNrneurons can explain the bursting firing pattern. A report of
Address for reprint requests and other correspondence: J. Bargas. Institutode Fisiologıa Celular UNAM, PO Box: 70-253, Mexico City, DF 04510Mexico (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked “advertisement”in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol 98: 2311–2323, 2007.First published August 22, 2007; doi:10.1152/jn.00620.2007.
23110022-3077/07 $8.00 Copyright © 2007 The American Physiological Societywww.jn.org

these results has been presented in abstract form (Ibanez-Sandoval et al. 2005).
M E T H O D S
Preparation of slices
All procedures were carried out in accordance with the NationalInstitutes of Health Guide for Care and Use of Laboratory Animalsand were approved by the Institutional Animal Care Committees ofthe UNAM. The experiments were performed on brain slices obtainedfrom Wistar juvenile rats (PD 14–21). The rats were anesthetized anddecapitated. The brain was rapidly obtained an immersed for one minin cold oxygenated saline (ca 4°C; 95% O2-5% CO2) of the followingcomposition (in mM): 124 NaCl, 2.5 KCl, 1.3 MgCl2, 26 NaHCO3,1.2 NaH2PO4, 2.4 CaCl2, and 10 glucose. The same saline but withcholine chloride (124 mM) instead of NaCl was used during theslicing procedure. Parasagittal slices (300 �m) containing the SNrwere cut on a vibratome (Pelco 101 Series 1000; Pelco, St. Louis,MO) and transferred to the saline with NaCl (preceding text). Theslices were left for equilibration for �1 h in oxygenated saline at roomtemperature (ca 25°C). After equilibration, a single slice was trans-ferred to a recording chamber placed on the stage of an uprightmicroscope and was continuously superfused (2–3 ml/min) withoxygenated saline at room temperature.
Whole cell recordings
Recordings were made at room temperature (�25°C) from neuronslocated inside the SNr boundaries as seen in the parasagittal slice. Thisregion mainly contains GABAergic neurons (see following text).Neurons were visualized using infrared differential interferencevideomicrosocopy with an �40 water-immersion objective. Micropi-pettes for whole cell recordings were pulled (Sutter Instrument,Novato, CA) from borosilicate glass tubes (1.5 mm OD, WPI, Sara-sota, FL) for a final resistance of 2–5 M� when filled with internalsaline of the following composition (in mM): 120 KSO3CH4, 10NaCl, 10 K2EGTA, 10 HEPES, 1 CaCl2, 2 MgCl2, 2 ATP-Mg, 0.3GTP-Na (pH 7.3, 290 mosM/l). Voltage-clamp recordings were madewith an Axopatch 200B amplifier (Axon Instruments, Foster City,CA). Liquid junction potentials (�8 mV) were not corrected. Current-voltage relationships made in voltage-clamp mode with 60–80%compensated series resistance, were performed by stepping the mem-brane potential to hyperpolarized and depolarized values with near 1-svoltage commands. Current values in response to these commandswere measured at the end of the step and plotted as a function ofvoltage so that I-V plots represent activity of currents underlying thespikes. Action currents were unclamped. Firing was readily obtainedafter switching to current-clamp mode. Coincidence between inputresistance measured in voltage- and current-clamp suggested thatneither bridge balance nor series resistance (�15 M�) represent aproblem (see Ibanez-Sandoval et al. 2006).
Immunocytochemical procedures
To identify the recorded cells, 1% byocitin was included in thepipette solution. For immunocytochemistry, the slices were incubatedin streptavidin conjugated with Cy3 (1:200 dissolved in PBS, ZymedLaboratories, San Francisco CA). This allowed us to visualize therecorded neuron (1 per slice). Thereafter slices were incubated 30 minwith 1% bovine albumin to block nonspecific binding sites and for36 h with a mouse monoclonal antibody against parvalbumin (anti-PV; 1:2000, Sigma-Aldrich dissolved in PBS containing 0.25%Triton-X). The slices were then rinsed thrice with PBS and incubatedwith a goat versus mouse secondary antibody during 1 h. Thisantibody was conjugated with Cy5 (Jackson Inmuno Res Lab, WestGrove, PA). To show the presence of voltage-gated calcium channel
of the CaV3.2 type (T-channels), the slices were again incubated 30min with 1% bovine albumin to block nonspecific binding sites. Thenincubation for 36 h with a goat policlonal antibody against CaV3.2channels followed (anti-CaV3.2; 1:100, dissolved in PBS containing0.25% Triton-X). The slices were then rinsed thrice with PBS andincubated with goat secondary antibody during 1 h. This antibody wasconjugated with fluorescein (Vector Laboratories, Burlingame, CA).Next, slices were mounted on covered slides and observed withdifferent fluorescent filters (Vectashield, Vector Laboratories) in antransillumination or in a confocal microscope (Bio-Rad Microscience,London, UK). Omission of primary anti-sera resulted in no detectablesignal (data not shown).
Drugs
Drugs were stored as dry aliquots and stock solutions were preparedjust prior to each experiment and added to the perfusion solution in thefinal concentration indicated. NMDA, biocytin, TEA-Cl, ATP-Mg,ATP-Na, picrotoxin, and bicuculline methiodide or hydrochloridewere obtained from Sigma (St. Louis, MO). ZD 7288 was obtainedfrom Tocris (Ellisville, MO). Tetrodotoxin (TTX), calcicludine,iberiotoxin and apamin were obtained from Alomone Labs (Jerusa-lem, Israel).
Data analysis
Digitized data were imported for analysis and graphing into com-mercial software (Origin v.6. Microcal, Northampton MA). All dataare given as means � SE plus median and range, unless statedotherwise. Free-distribution statistical tests were used to assess statis-tical significance: Mann-Whitney’s U test or Wilcoxon’s t-test (de-pending on nonpaired or paired samples; Systat, Richmond CA) andFriedman�s one-way ANOVA with post hoc Dunn�s test or post hocStudent-Newman-Keuls test were also employed to compare morethan two samples. Statistical differences of fitted functions wereassessed by comparing the obtained parameters and their estimationerrors with Student�s t-test.
R E S U L T S
SNr projection neurons may burst
Ninety percent (n � 63/70) of SNr neurons recorded in vitro incurrent-clamp mode exhibited spontaneous activity (Fig. 1A1). Aspreviously described (Atherton and Bevan 2005; Nakanishiet al. 1987; Richards et al. 1997), spontaneous activity ischaracterized by regular single-spike firing at a frequency of10 � 2 Hz (n � 20; range: 4–29 Hz) with action potentialthreshold at 46 � 0.5 mV. Action potential duration athalf-amplitude is 0.87 � 0.02 ms (n � 35). Regularity ofspontaneous firing was assessed by several means. First, aphase-portrait of membrane potential versus its time derivativeexhibited superimposed regular orbits with clear and fixedlandmarks (Fig. 1A2). Second, distribution of interspike inter-vals had a single mode (Fig. 1B1) lasting 77 � 0.3 ms with acoefficient of variation of 0.11 � 0.01 (n � 20). Third, adiagram of interspike intervals (ISIs) against subsequent onesshowed a single cluster (Fig. 1B2). Sixty-five percent of theserecorded neurons (Fig. 1, C, 1–3) were inmunoreactive againstparvalbumin (PV). PV� neurons have been described beforeto be GABAergic projection neurons (Gerfen et al. 1985;Gonzalez-Hernandez and Rodrıguez 2000; Ibanez-Sandovalet al. 2006).
In our hands, 11% (n � 7/63) of tonically firing neuronsexhibited spontaneous transitions in their firing pattern from
2312 IBANEZ-SANDOVAL ET AL.
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org
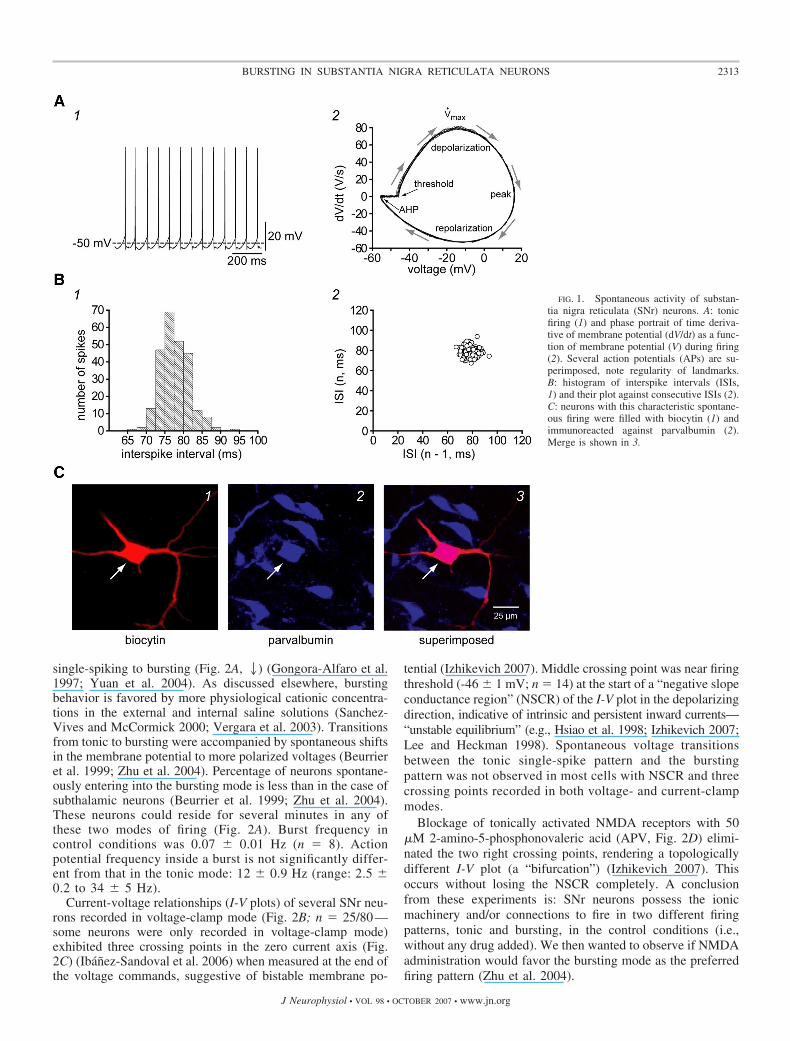
single-spiking to bursting (Fig. 2A, 2) (Gongora-Alfaro et al.1997; Yuan et al. 2004). As discussed elsewhere, burstingbehavior is favored by more physiological cationic concentra-tions in the external and internal saline solutions (Sanchez-Vives and McCormick 2000; Vergara et al. 2003). Transitionsfrom tonic to bursting were accompanied by spontaneous shiftsin the membrane potential to more polarized voltages (Beurrieret al. 1999; Zhu et al. 2004). Percentage of neurons spontane-ously entering into the bursting mode is less than in the case ofsubthalamic neurons (Beurrier et al. 1999; Zhu et al. 2004).These neurons could reside for several minutes in any ofthese two modes of firing (Fig. 2A). Burst frequency incontrol conditions was 0.07 � 0.01 Hz (n � 8). Actionpotential frequency inside a burst is not significantly differ-ent from that in the tonic mode: 12 � 0.9 Hz (range: 2.5 �0.2 to 34 � 5 Hz).
Current-voltage relationships (I-V plots) of several SNr neu-rons recorded in voltage-clamp mode (Fig. 2B; n � 25/80—some neurons were only recorded in voltage-clamp mode)exhibited three crossing points in the zero current axis (Fig.2C) (Ibanez-Sandoval et al. 2006) when measured at the end ofthe voltage commands, suggestive of bistable membrane po-
tential (Izhikevich 2007). Middle crossing point was near firingthreshold (-46 � 1 mV; n � 14) at the start of a “negative slopeconductance region” (NSCR) of the I-V plot in the depolarizingdirection, indicative of intrinsic and persistent inward currents—“unstable equilibrium” (e.g., Hsiao et al. 1998; Izhikevich 2007;Lee and Heckman 1998). Spontaneous voltage transitionsbetween the tonic single-spike pattern and the burstingpattern was not observed in most cells with NSCR and threecrossing points recorded in both voltage- and current-clampmodes.
Blockage of tonically activated NMDA receptors with 50�M 2-amino-5-phosphonovaleric acid (APV, Fig. 2D) elimi-nated the two right crossing points, rendering a topologicallydifferent I-V plot (a “bifurcation”) (Izhikevich 2007). Thisoccurs without losing the NSCR completely. A conclusionfrom these experiments is: SNr neurons possess the ionicmachinery and/or connections to fire in two different firingpatterns, tonic and bursting, in the control conditions (i.e.,without any drug added). We then wanted to observe if NMDAadministration would favor the bursting mode as the preferredfiring pattern (Zhu et al. 2004).
FIG. 1. Spontaneous activity of substan-tia nigra reticulata (SNr) neurons. A: tonicfiring (1) and phase portrait of time deriva-tive of membrane potential (dV/dt) as a func-tion of membrane potential (V) during firing(2). Several action potentials (APs) are su-perimposed, note regularity of landmarks.B: histogram of interspike intervals (ISIs,1) and their plot against consecutive ISIs (2).C: neurons with this characteristic spontane-ous firing were filled with biocytin (1) andimmunoreacted against parvalbumin (2).Merge is shown in 3.
2313BURSTING IN SUBSTANTIA NIGRA RETICULATA NEURONS
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org
Enhancing the excitatory drive on SNr neurons
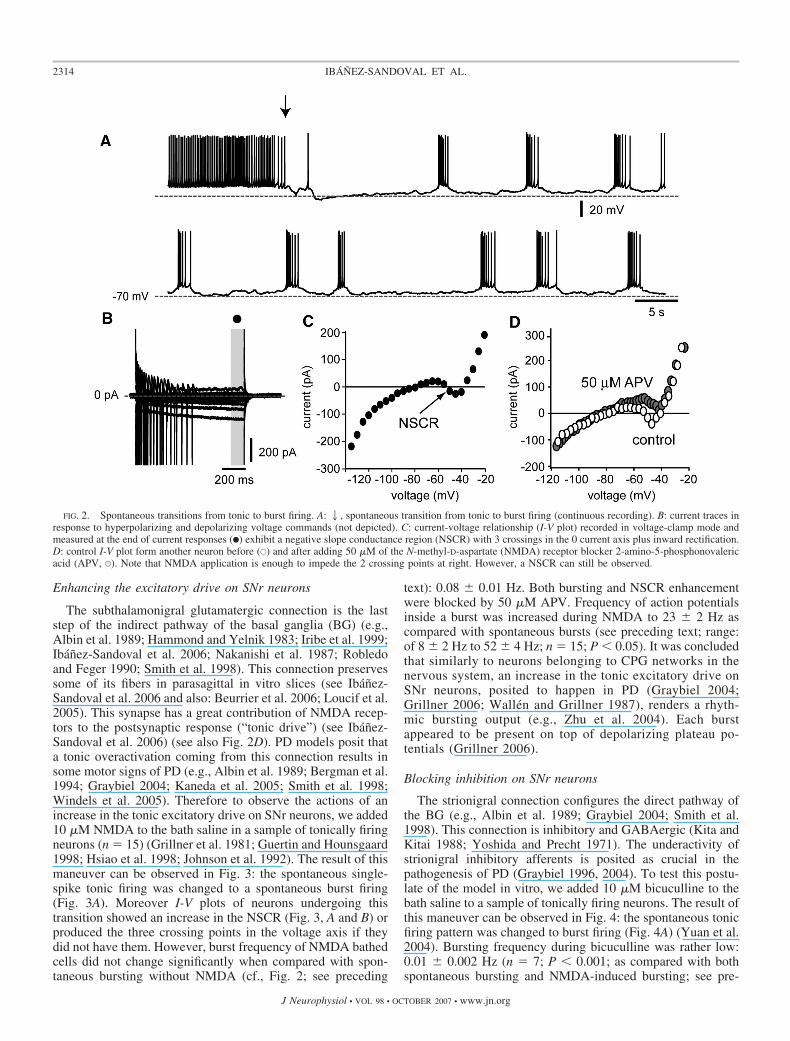
The subthalamonigral glutamatergic connection is the laststep of the indirect pathway of the basal ganglia (BG) (e.g.,Albin et al. 1989; Hammond and Yelnik 1983; Iribe et al. 1999;Ibanez-Sandoval et al. 2006; Nakanishi et al. 1987; Robledoand Feger 1990; Smith et al. 1998). This connection preservessome of its fibers in parasagittal in vitro slices (see Ibanez-Sandoval et al. 2006 and also: Beurrier et al. 2006; Loucif et al.2005). This synapse has a great contribution of NMDA recep-tors to the postsynaptic response (“tonic drive”) (see Ibanez-Sandoval et al. 2006) (see also Fig. 2D). PD models posit thata tonic overactivation coming from this connection results insome motor signs of PD (e.g., Albin et al. 1989; Bergman et al.1994; Graybiel 2004; Kaneda et al. 2005; Smith et al. 1998;Windels et al. 2005). Therefore to observe the actions of anincrease in the tonic excitatory drive on SNr neurons, we added10 �M NMDA to the bath saline in a sample of tonically firingneurons (n � 15) (Grillner et al. 1981; Guertin and Hounsgaard1998; Hsiao et al. 1998; Johnson et al. 1992). The result of thismaneuver can be observed in Fig. 3: the spontaneous single-spike tonic firing was changed to a spontaneous burst firing(Fig. 3A). Moreover I-V plots of neurons undergoing thistransition showed an increase in the NSCR (Fig. 3, A and B) orproduced the three crossing points in the voltage axis if theydid not have them. However, burst frequency of NMDA bathedcells did not change significantly when compared with spon-taneous bursting without NMDA (cf., Fig. 2; see preceding
text): 0.08 � 0.01 Hz. Both bursting and NSCR enhancementwere blocked by 50 �M APV. Frequency of action potentialsinside a burst was increased during NMDA to 23 � 2 Hz ascompared with spontaneous bursts (see preceding text; range:of 8 � 2 Hz to 52 � 4 Hz; n � 15; P � 0.05). It was concludedthat similarly to neurons belonging to CPG networks in thenervous system, an increase in the tonic excitatory drive onSNr neurons, posited to happen in PD (Graybiel 2004;Grillner 2006; Wallen and Grillner 1987), renders a rhyth-mic bursting output (e.g., Zhu et al. 2004). Each burstappeared to be present on top of depolarizing plateau po-tentials (Grillner 2006).
Blocking inhibition on SNr neurons
The strionigral connection configures the direct pathway ofthe BG (e.g., Albin et al. 1989; Graybiel 2004; Smith et al.1998). This connection is inhibitory and GABAergic (Kita andKitai 1988; Yoshida and Precht 1971). The underactivity ofstrionigral inhibitory afferents is posited as crucial in thepathogenesis of PD (Graybiel 1996, 2004). To test this postu-late of the model in vitro, we added 10 �M bicuculline to thebath saline to a sample of tonically firing neurons. The result ofthis maneuver can be observed in Fig. 4: the spontaneous tonicfiring pattern was changed to burst firing (Fig. 4A) (Yuan et al.2004). Bursting frequency during bicuculline was rather low:0.01 � 0.002 Hz (n � 7; P � 0.001; as compared with bothspontaneous bursting and NMDA-induced bursting; see pre-
FIG. 2. Spontaneous transitions from tonic to burst firing. A:2, spontaneous transition from tonic to burst firing (continuous recording). B: current traces inresponse to hyperpolarizing and depolarizing voltage commands (not depicted). C: current-voltage relationship (I-V plot) recorded in voltage-clamp mode andmeasured at the end of current responses (F) exhibit a negative slope conductance region (NSCR) with 3 crossings in the 0 current axis plus inward rectification.D: control I-V plot form another neuron before (E) and after adding 50 �M of the N-methyl-D-aspartate (NMDA) receptor blocker 2-amino-5-phosphonovalericacid (APV, U). Note that NMDA application is enough to impede the 2 crossing points at right. However, a NSCR can still be observed.
2314 IBANEZ-SANDOVAL ET AL.
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org

ceding text). I-V plots of all neurons bathed with bicucullineshowed an increase in the NSCR and displayed bursting (Fig.4, A and B), indicating that tonic inhibition is capable ofrestraining intrinsic persistent inward currents. The same resultwas obtained in two experiments after the application ofpicrotoxin (50 �M). Frequency of action potentials inside aburst was increased: 20 � 1 Hz (range: 3–47 Hz) with respectto spontaneous bursting (see preceding text; n � 9; P � 0.05)after GABAA receptor blockage.
Reduction of inhibition is also known to induce rhythmicpatterns in CPG networks by unbalancing a tonic excitatorydrive (Grillner 2006; Schmitt et al. 2004; Takakusaki et al.2004), including the SNr (Yuan et al. 2004). Therefore theblockade of an inhibitory tone discloses and confirms thepresence of a tonic excitatory tone. Previous experiments usingextracellular recordings could not observe the subthresholdbehavior during bursting (Gongora-Alfaro et al. 1997; Yuanet al. 2004).
Figure 4 also shows that addition of 10 �M NMDA in thecontinuous presence of bicuculline made the oscillations morerobust and frequent, suggesting that both maneuvers haveadditive actions and that the system is far from saturation withany of the maneuvers. Burst duration increased from 3.5 � 0.1to 6.2 � 0.2 s, and action potential frequency inside burstsbecame 27 � 0.8 Hz (n � 5; range: 13–61 Hz). Because theI-V plot was clearly changed after agonists and antagonists ofboth transmitter receptors: glutamate and GABA, it is sug-gested that SNr neurons receive important excitatory andinhibitory tonic synaptic drives in such a way as to restrain the
intrinsic conductances that generate bursting, thus limiting thefiring pattern to tonic single-spike firing.
Ionic mechanisms of bursting in SNr neurons
Having demonstrated that pharmacological procedures thatdisclose pattern generation in other parts of the nervous sys-tem, disclose bursting firing pattern in SNr neurons, we begansearching into the ionic mechanisms underlying this bursting.Ionic substitution experiments were then performed in thepresence of 10 �M NMDA.
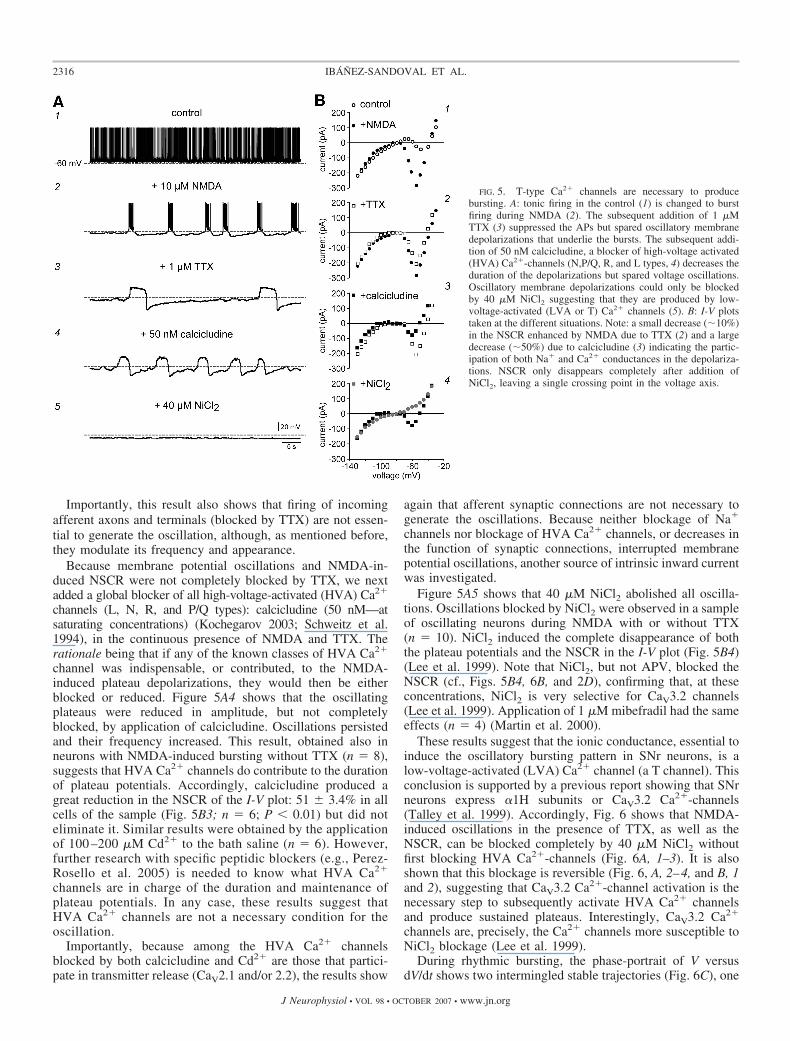
In a sample of neurons (n � 6) exhibiting oscillatorybursting induced by 10 �M NMDA (cf., Figs. 5A1 and 2),addition of 1 �M TTX, abolished the firing of action potentials(Fig. 5A3), and disclosed the slow and repetitive membranepotential plateaus, that appear to underlie burst firing. Thus theoscillation persisted after TTX in all neurons tested in this way(e.g., Beurrier et al. 1999; Guertin and Hounsgaard 1998;Hsiao et al. 1998; Zhu et al. 2004). Plateaus had longerduration than control NMDA-induced bursts but exhibited alower frequency (Fig. 5A3). However, the NSCR in the I-V plotwas decreased by TTX in all neurons of this sample by 11 �1% (n � 6; cf., Figs. 5B1 and 2) as though persistent Na�
currents contributed, in part, to generate the slow depolariza-tions underlying the bursts (Grillner 2006; Hsiao et al.1998;Zhu et al. 2004).
FIG. 4. Bursting induced by bicuculline. A: spontaneous tonic firing in thecontrol (top) is changed to burst firing after addition of 10 �M bicuculline(middle). Burst frequency is further enhanced by the subsequent addition of 10�M NMDA (bottom). - - -, 60 mV. B: current traces in response to hyper-polarizing and depolarizing voltage commands (1) in all 3 conditions areplotted in 2. I-V plots show that bicuculline is enough to augment the NSCR.NMDA has additive effects on those produced by bicuculline.
FIG. 3. Bursting induced by NMDA. A: spontaneous tonic firing in thecontrol (top) is changed to burst firing after addition of 10 �M NMDA(bottom). - - -, 60 mV. B: current traces in response to hyperpolarizing anddepolarizing voltage commands before (top) and during NMDA administration(bottom, 1) were used to build I-V plots in both conditions (2). Note enhance-ment of NSCR during NMDA (F).
2315BURSTING IN SUBSTANTIA NIGRA RETICULATA NEURONS
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org
Importantly, this result also shows that firing of incomingafferent axons and terminals (blocked by TTX) are not essen-tial to generate the oscillation, although, as mentioned before,they modulate its frequency and appearance.
Because membrane potential oscillations and NMDA-in-duced NSCR were not completely blocked by TTX, we nextadded a global blocker of all high-voltage-activated (HVA) Ca2�
channels (L, N, R, and P/Q types): calcicludine (50 nM—atsaturating concentrations) (Kochegarov 2003; Schweitz et al.1994), in the continuous presence of NMDA and TTX. Therationale being that if any of the known classes of HVA Ca2�
channel was indispensable, or contributed, to the NMDA-induced plateau depolarizations, they would then be eitherblocked or reduced. Figure 5A4 shows that the oscillatingplateaus were reduced in amplitude, but not completelyblocked, by application of calcicludine. Oscillations persistedand their frequency increased. This result, obtained also inneurons with NMDA-induced bursting without TTX (n � 8),suggests that HVA Ca2� channels do contribute to the durationof plateau potentials. Accordingly, calcicludine produced agreat reduction in the NSCR of the I-V plot: 51 � 3.4% in allcells of the sample (Fig. 5B3; n � 6; P � 0.01) but did noteliminate it. Similar results were obtained by the applicationof 100 –200 �M Cd2� to the bath saline (n � 6). However,further research with specific peptidic blockers (e.g., Perez-Rosello et al. 2005) is needed to know what HVA Ca2�
channels are in charge of the duration and maintenance ofplateau potentials. In any case, these results suggest thatHVA Ca2� channels are not a necessary condition for theoscillation.
Importantly, because among the HVA Ca2� channelsblocked by both calcicludine and Cd2� are those that partici-pate in transmitter release (CaV2.1 and/or 2.2), the results show
again that afferent synaptic connections are not necessary togenerate the oscillations. Because neither blockage of Na�
channels nor blockage of HVA Ca2� channels, or decreases inthe function of synaptic connections, interrupted membranepotential oscillations, another source of intrinsic inward currentwas investigated.
Figure 5A5 shows that 40 �M NiCl2 abolished all oscilla-tions. Oscillations blocked by NiCl2 were observed in a sampleof oscillating neurons during NMDA with or without TTX(n � 10). NiCl2 induced the complete disappearance of boththe plateau potentials and the NSCR in the I-V plot (Fig. 5B4)(Lee et al. 1999). Note that NiCl2, but not APV, blocked theNSCR (cf., Figs. 5B4, 6B, and 2D), confirming that, at theseconcentrations, NiCl2 is very selective for CaV3.2 channels(Lee et al. 1999). Application of 1 �M mibefradil had the sameeffects (n � 4) (Martin et al. 2000).
These results suggest that the ionic conductance, essential toinduce the oscillatory bursting pattern in SNr neurons, is alow-voltage-activated (LVA) Ca2� channel (a T channel). Thisconclusion is supported by a previous report showing that SNrneurons express �1H subunits or CaV3.2 Ca2�-channels(Talley et al. 1999). Accordingly, Fig. 6 shows that NMDA-induced oscillations in the presence of TTX, as well as theNSCR, can be blocked completely by 40 �M NiCl2 withoutfirst blocking HVA Ca2�-channels (Fig. 6A, 1–3). It is alsoshown that this blockage is reversible (Fig. 6, A, 2–4, and B, 1and 2), suggesting that CaV3.2 Ca2�-channel activation is thenecessary step to subsequently activate HVA Ca2� channelsand produce sustained plateaus. Interestingly, CaV3.2 Ca2�
channels are, precisely, the Ca2� channels more susceptible toNiCl2 blockage (Lee et al. 1999).
During rhythmic bursting, the phase-portrait of V versusdV/dt shows two intermingled stable trajectories (Fig. 6C), one
FIG. 5. T-type Ca2� channels are necessary to producebursting. A: tonic firing in the control (1) is changed to burstfiring during NMDA (2). The subsequent addition of 1 �MTTX (3) suppressed the APs but spared oscillatory membranedepolarizations that underlie the bursts. The subsequent addi-tion of 50 nM calcicludine, a blocker of high-voltage activated(HVA) Ca2�-channels (N,P/Q, R, and L types, 4) decreases theduration of the depolarizations but spared voltage oscillations.Oscillatory membrane depolarizations could only be blockedby 40 �M NiCl2 suggesting that they are produced by low-voltage-activated (LVA or T) Ca2� channels (5). B: I-V plotstaken at the different situations. Note: a small decrease (10%)in the NSCR enhanced by NMDA due to TTX (2) and a largedecrease (50%) due to calcicludine (3) indicating the partic-ipation of both Na� and Ca2� conductances in the depolariza-tions. NSCR only disappears completely after addition ofNiCl2, leaving a single crossing point in the voltage axis.
2316 IBANEZ-SANDOVAL ET AL.
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org
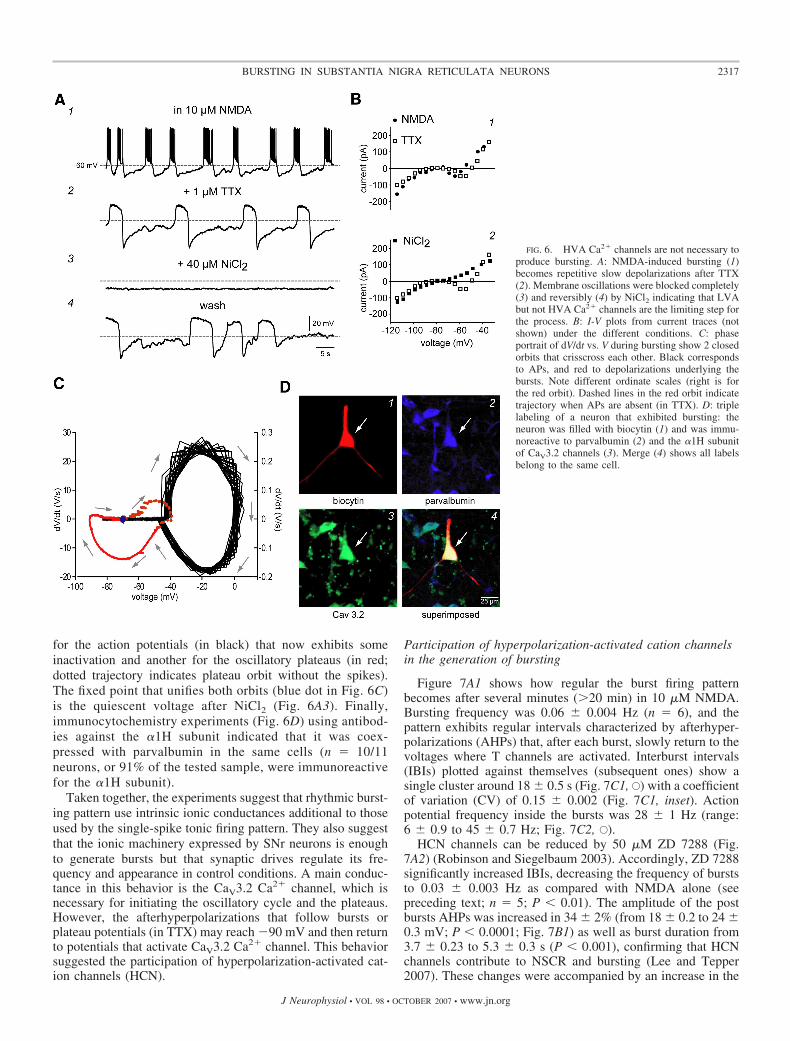
for the action potentials (in black) that now exhibits someinactivation and another for the oscillatory plateaus (in red;dotted trajectory indicates plateau orbit without the spikes).The fixed point that unifies both orbits (blue dot in Fig. 6C)is the quiescent voltage after NiCl2 (Fig. 6A3). Finally,immunocytochemistry experiments (Fig. 6D) using antibod-ies against the �1H subunit indicated that it was coex-pressed with parvalbumin in the same cells (n � 10/11neurons, or 91% of the tested sample, were immunoreactivefor the �1H subunit).
Taken together, the experiments suggest that rhythmic burst-ing pattern use intrinsic ionic conductances additional to thoseused by the single-spike tonic firing pattern. They also suggestthat the ionic machinery expressed by SNr neurons is enoughto generate bursts but that synaptic drives regulate its fre-quency and appearance in control conditions. A main conduc-tance in this behavior is the CaV3.2 Ca2� channel, which isnecessary for initiating the oscillatory cycle and the plateaus.However, the afterhyperpolarizations that follow bursts orplateau potentials (in TTX) may reach 90 mV and then returnto potentials that activate CaV3.2 Ca2� channel. This behaviorsuggested the participation of hyperpolarization-activated cat-ion channels (HCN).
Participation of hyperpolarization-activated cation channelsin the generation of bursting
Figure 7A1 shows how regular the burst firing patternbecomes after several minutes (�20 min) in 10 �M NMDA.Bursting frequency was 0.06 � 0.004 Hz (n � 6), and thepattern exhibits regular intervals characterized by afterhyper-polarizations (AHPs) that, after each burst, slowly return to thevoltages where T channels are activated. Interburst intervals(IBIs) plotted against themselves (subsequent ones) show asingle cluster around 18 � 0.5 s (Fig. 7C1, E) with a coefficientof variation (CV) of 0.15 � 0.002 (Fig. 7C1, inset). Actionpotential frequency inside the bursts was 28 � 1 Hz (range:6 � 0.9 to 45 � 0.7 Hz; Fig. 7C2, E).
HCN channels can be reduced by 50 �M ZD 7288 (Fig.7A2) (Robinson and Siegelbaum 2003). Accordingly, ZD 7288significantly increased IBIs, decreasing the frequency of burststo 0.03 � 0.003 Hz as compared with NMDA alone (seepreceding text; n � 5; P � 0.01). The amplitude of the postbursts AHPs was increased in 34 � 2% (from 18 � 0.2 to 24 �0.3 mV; P � 0.0001; Fig. 7B1) as well as burst duration from3.7 � 0.23 to 5.3 � 0.3 s (P � 0.001), confirming that HCNchannels contribute to NSCR and bursting (Lee and Tepper2007). These changes were accompanied by an increase in the
FIG. 6. HVA Ca2� channels are not necessary toproduce bursting. A: NMDA-induced bursting (1)becomes repetitive slow depolarizations after TTX(2). Membrane oscillations were blocked completely(3) and reversibly (4) by NiCl2 indicating that LVAbut not HVA Ca2� channels are the limiting step forthe process. B: I-V plots from current traces (notshown) under the different conditions. C: phaseportrait of dV/dt vs. V during bursting show 2 closedorbits that crisscross each other. Black correspondsto APs, and red to depolarizations underlying thebursts. Note different ordinate scales (right is forthe red orbit). Dashed lines in the red orbit indicatetrajectory when APs are absent (in TTX). D: triplelabeling of a neuron that exhibited bursting: theneuron was filled with biocytin (1) and was immu-noreactive to parvalbumin (2) and the �1H subunitof CaV3.2 channels (3). Merge (4) shows all labelsbelong to the same cell.
2317BURSTING IN SUBSTANTIA NIGRA RETICULATA NEURONS
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org
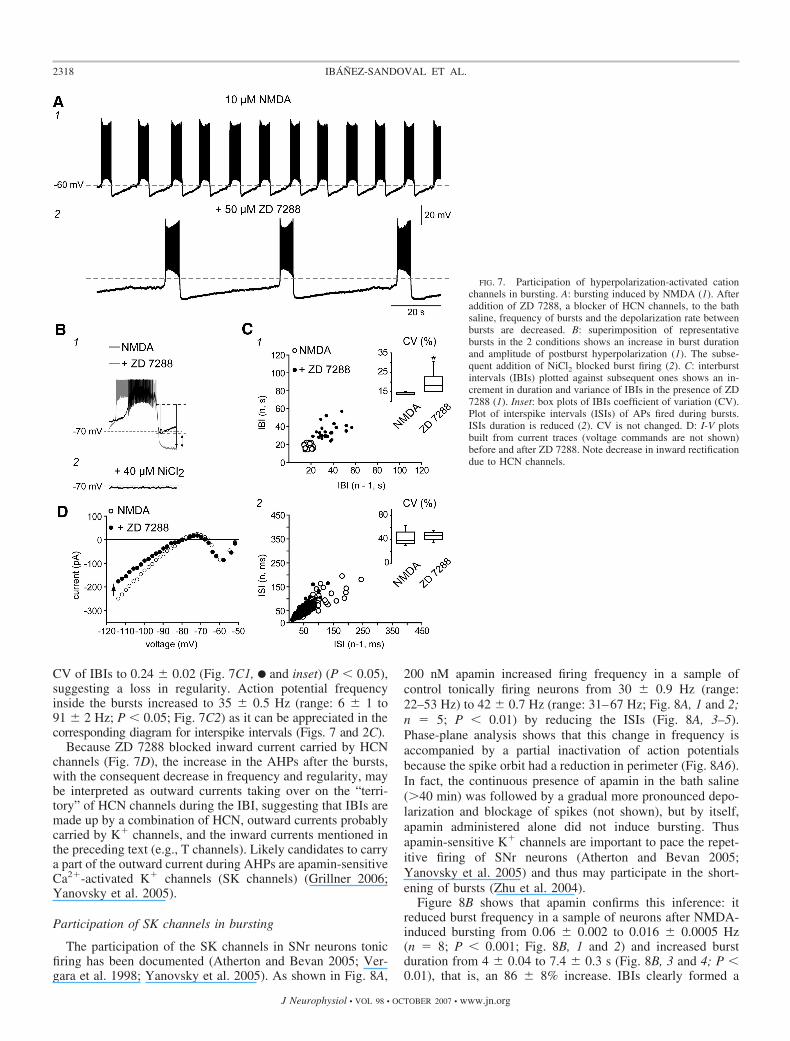
CV of IBIs to 0.24 � 0.02 (Fig. 7C1, F and inset) (P � 0.05),suggesting a loss in regularity. Action potential frequencyinside the bursts increased to 35 � 0.5 Hz (range: 6 � 1 to91 � 2 Hz; P � 0.05; Fig. 7C2) as it can be appreciated in thecorresponding diagram for interspike intervals (Figs. 7 and 2C).
Because ZD 7288 blocked inward current carried by HCNchannels (Fig. 7D), the increase in the AHPs after the bursts,with the consequent decrease in frequency and regularity, maybe interpreted as outward currents taking over on the “terri-tory” of HCN channels during the IBI, suggesting that IBIs aremade up by a combination of HCN, outward currents probablycarried by K� channels, and the inward currents mentioned inthe preceding text (e.g., T channels). Likely candidates to carrya part of the outward current during AHPs are apamin-sensitiveCa2�-activated K� channels (SK channels) (Grillner 2006;Yanovsky et al. 2005).
Participation of SK channels in bursting
The participation of the SK channels in SNr neurons tonicfiring has been documented (Atherton and Bevan 2005; Ver-gara et al. 1998; Yanovsky et al. 2005). As shown in Fig. 8A,
200 nM apamin increased firing frequency in a sample ofcontrol tonically firing neurons from 30 � 0.9 Hz (range:22–53 Hz) to 42 � 0.7 Hz (range: 31–67 Hz; Fig. 8A, 1 and 2;n � 5; P � 0.01) by reducing the ISIs (Fig. 8A, 3–5).Phase-plane analysis shows that this change in frequency isaccompanied by a partial inactivation of action potentialsbecause the spike orbit had a reduction in perimeter (Fig. 8A6).In fact, the continuous presence of apamin in the bath saline(�40 min) was followed by a gradual more pronounced depo-larization and blockage of spikes (not shown), but by itself,apamin administered alone did not induce bursting. Thusapamin-sensitive K� channels are important to pace the repet-itive firing of SNr neurons (Atherton and Bevan 2005;Yanovsky et al. 2005) and thus may participate in the short-ening of bursts (Zhu et al. 2004).
Figure 8B shows that apamin confirms this inference: itreduced burst frequency in a sample of neurons after NMDA-induced bursting from 0.06 � 0.002 to 0.016 � 0.0005 Hz(n � 8; P � 0.001; Fig. 8B, 1 and 2) and increased burstduration from 4 � 0.04 to 7.4 � 0.3 s (Fig. 8B, 3 and 4; P �0.01), that is, an 86 � 8% increase. IBIs clearly formed a
FIG. 7. Participation of hyperpolarization-activated cationchannels in bursting. A: bursting induced by NMDA (1). Afteraddition of ZD 7288, a blocker of HCN channels, to the bathsaline, frequency of bursts and the depolarization rate betweenbursts are decreased. B: superimposition of representativebursts in the 2 conditions shows an increase in burst durationand amplitude of postburst hyperpolarization (1). The subse-quent addition of NiCl2 blocked burst firing (2). C: interburstintervals (IBIs) plotted against subsequent ones shows an in-crement in duration and variance of IBIs in the presence of ZD7288 (1). Inset: box plots of IBIs coefficient of variation (CV).Plot of interspike intervals (ISIs) of APs fired during bursts.ISIs duration is reduced (2). CV is not changed. D: I-V plotsbuilt from current traces (voltage commands are not shown)before and after ZD 7288. Note decrease in inward rectificationdue to HCN channels.
2318 IBANEZ-SANDOVAL ET AL.
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org

different cluster after apamin (Fig. 8B6; P � 0.05). As ex-pected, firing frequency inside the bursts was increased from33 � 0.5 Hz (range: 16 � 0.5 to 52 � 2 Hz) to 45 � 1.1 Hzin this sample (range: 16 � 1.2 to 98 � 3 Hz; P � 0.05).
The diagram of ISIs was simply spread toward minor inter-vals in both tonic and burst firing allowing the firing of morespikes (Fig. 8, A5 and B5). In contrast, IBIs clearly separatedinto different clusters (Fig. 8B6), suggesting an inverse rela-tionship between ISIs and IBIs duration. This suggests thatSK-channel reduction may slow down burst repolarization witha consequent increase in burst duration, allowing more Ca2�
influx to generate more prolonged AHPs and IBIs. This sug-gests that other K� channels, perhaps BK channels, shape thedischarge during the burst.
Participation of BK channels
Submillimolar concentrations of tetraethylammonium(TEA) are known to be selective in blocking BK channels(Bargas et al. 1999; Pineda et al. 1992; Vergara et al. 1998).Therefore 500 �M TEA were used to infer the participation ofthese channels in tonic and burst firing (Fig. 9). Basically, TEAslowed down the repolarization of the action potential andblocked the initial part of the AHP during control tonic firing(Fig. 9A, 1–3). This prolonged the action potentials from0.83 � 0.02 to 1.16 � 0.02 ms in a sample of neurons recorded
in control conditions (Fig. 9A, 3 and 5; n � 20; P � 0.0001).Phase portrait shows that only the repolarization phase and thepeak of the AHP were affected. In contrast with apaminactions, these effects were not enough to change firing fre-quency: from 9.3 � 1.7 in the control to 9.4 � 1.3 Hz afterTEA (NS). ISIs or their CVs were not changed significantly.
In the same way, TEA had minor effects in either burstduration or IBIs in a sample of neurons during bursting inducedby NMDA (n � 6; Fig. 9B). The most detectable actions wereon spikes. Action potential frequency decreased from 24 � 0.4Hz (range: 10 � 1 to 46 � 2 Hz) to 16 � 0.3 Hz (range: 1 �0.05 to 34 � 3 Hz; P � 0.05), suggesting that at high firingfrequencies, the small change on the fast AHP does have aneffect in frequency. Action potentials during the bursts tendedto inactivate (Fig. 9B, 3 and 4) disabling the neuron formaintaining high frequencies during the bursts. Instead ofaction potentials, high-frequency oscillations on top of theplateau potentials were evident (Fig. 9B, 3 and 4). The durationof bursts suffered a decrease, from 2.6 � 0.06 to 1.6 � 0.07 s(P � 0.01). However, neither IBIs nor ISIs distributionsshowed significant changes (Fig. 9B, 5 and 6). Iberiotoxin, aselective BK-channel blocker had the same effects as micro-molar TEA (n � 5). To conclude, both BK and SK channelsparticipate in the firing of actions potentials during bursts butonly SK channels participate in fixing bursting frequency.
FIG. 8. Participation of Ca2�-activatedK� channels of small conductance in burst-ing. A: apamin (200 nM), a blocker of smallconductance (SK) Ca2�-activated K� chan-nels decreases ISIs and increases firing fre-quency if applied during spontaneous tonicfiring: control (1), in apamin (2), superimpo-sition of APs and ISIs at different time base(3 and 4) shows a decrease in the AHP,which leads to a decrease in the ISI (5).However, CV of ISIs decreased afterapamin. The dV/dt vs. V portrait shows thatthe area of the orbit corresponding to tonicAPs is reduced (6). B: apamin actions duringburst firing. Frequency of NMDA-inducedbursting (1) decreased after apamin (2),whereas the duration of bursts increased(3 and 4). In contrast to tonic firing (A),apamin did not change significantly intra-burst ISIs or their CV. However, apaminsignificantly changed IBIs.
2319BURSTING IN SUBSTANTIA NIGRA RETICULATA NEURONS
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org

Further research is needed to disentangle the complete set ofoutward currents in charge of shaping the bursts and burstingfiring pattern (e.g., Na-activated K channels, slowly inactivat-ing voltage-dependent K channels, and so on) (Grillner 2006),but clearly, HCN, SK, and T channels are important in fixingthe temporal structure of oscillatory bursting in SNr neurons.
D I S C U S S I O N
NMDA bursting
Because the rate model of PD posits that pathologicalchanges are a result of an overexcitation of the indirect path-way and an overinhibition of the direct pathway of the BG(Albin et al.1989; Day et al. 2006; Graybiel 1996; Murer et al.2002; Windels 2005), we decided to test this model by observ-ing what happens to normal SNr neurons after producing anincrease in their excitatory tonic drive and/or a decrease in theirincoming inhibition.
As in many other studies (Grillner et al. 1981; Hochmanet al. 1994; Hsiao et al. 1998; Wallen and Grillner 1987),NMDA increase the excitatory drive on SNr neurons. As incircuits embodying CPGs, the increase in the excitatory drivewas transformed into patterns of bursting activity (Grillner2006; Takakusaki et al. 2004). Coincidently, the oscillatory
model of PD posits that synaptic changes induced by dopaminedepletion disclose pathological or uncontrolled CPGs, mani-fested by bursting and abnormal rhythmic firing (Hutchinson etal. 2004; Plenz and Kitai 1999). Indeed, bursting in SNrneurons is observed during parkinsonism (Bergman et al. 1994;Hutchison et al. 2004; Walters et al. 2007; Wichmann et al.1999).
In a comparable way, a reduction of inhibition on SNrneurons also induced bursting (Grillner 2006; Takakusaki et al.2004; Yuan et al. 2004). Accordingly, it was found that bysimulating the synaptic changes proposed by the rate model ofPD, we obtained pattern generation, a sign proposed as a mainelectrophysiological characteristic of PD by the oscillatorymodel. To our knowledge, neither bursting nor its ionic andinduction mechanisms had been reported previously for SNrneurons, much less in the context of synaptic changes proposedto characterize PD.
Note that according to the rate model of PD, GABAergicinhibitory afferents connecting the globus pallidus (external)and the SNr (Graybiel 2004, 2005; Kita 2001; Parent andHazrati 1995) would also be underactive in PD because thestriatal neurons that inhibit pallidal neurons would be overac-tive (Day et al. 2006; Mallet et al. 2006). However, it has beenproposed that pallidal neurons participate in the connections of
FIG. 9. Participation of Ca2�-activatedK� channels of big conductance in intraburstfiring. A: 500 �M tetraethylammonium(TEA) is a blocker of big conductance (BK)Ca2�-activated K� channels. TEA does notchange the frequency of spontaneous tonicfiring (1 and 2). TEA decreased AP repolar-ization (3) without changing ISIs duration(4). The dV/dt vs. V portrait shows a reduc-tion in the trajectory corresponding to APrepolarization rate (5) without great changesin ISIs CV (6). B: TEA induces APs inacti-vation during the bursts. In contrast toapamin (Fig. 8), frequency of NMDA-in-duced bursting (1) is not changed after TEA(2). However, TEA alters firing during burst-ing (3 and 4): it partially inactivates APsdisclosing a fast frequency oscillation on topof the depolarization that underlies the bursts(4, bottom). Neither ISIs nor IBIs changeafter TEA; except for ISIs CV.
2320 IBANEZ-SANDOVAL ET AL.
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org

the CPG that emerges after dopamine depletion (Hutchinson etal. 2004; Plenz and Kitai 1999). Therefore GABA concentra-tion may in fact be fluctuating (Bevan et al. 2002; Walters et al.2007; Windels et al. 2005) in PD due to CPG activity.
The present experiments also revealed that SNr projectionneurons have an intrinsic pacemaking capability to burst sim-ilar to that found in STN neurons (Beurrier et al. 1999). Neitherthe blockage of all synaptic transmission with Cd2� nor theblockage of action potential firing with TTX abolished thesubthreshold oscillation underlying bursting. It is known thatboth NMDA and bicuculline initiate locomotion or augmentedmuscle tone if injected in locomotor regions controlled by theSNr (Takakusaki et al. 2004).
Ionic conductances necessary for bursting in SNr neurons
An initial investigation into the ionic conductances respon-sible for bursting in SNr neurons showed that several ioniccurrents participate in burst generation, such as: HCN, SK, andHVA Ca2� channels. However, it was found that activation ofCaV3.2 Ca2� channels is the necessary step to induce theoscillatory pattern. In SNr neurons, Na� and HVA Ca2�
conductances participate in fixing the duration and amplitudeof the plateau potential that underlies the bursts, and HCN andSK conductances participate in fixing the temporal structure ofbursting, but it is the CaV3.2 Ca2� channel that links both setsof conductances to produce bursting (Grillner 2006). Theexperiments also show that both excitatory and inhibitory tonicdrives are readily detected on SNr neurons by means ofchanges in the NSCR of the steady-state I-V plot after phar-macological manipulations (NMDA, APV, bicuculline).Therefore the fact that neither TTX nor HVA Ca2�-channelblockade affected the generation of membrane oscillations(pacemaking properties) does not imply that synaptic drives areunimportant. On the contrary, synaptic changes had to bedisposed to disclose the bursting pattern, which means that theabsence of this pattern during tonic firing implies important ongoing and balanced synaptic influences, even in the in vitroslice preparation (Beurrier et al. 2006). In this respect, burstingfrequencies found in the present study are rather low probablydue to incomplete circuitry connections (Loucif et al. 2005),favoring the view that the cortex may be the origin of the mainpacemaker (Magill et al. 2001; Yuste et al. 2005).
Functional implications
Bursting SNr neurons is a common finding in PD patientsand parkinsonian animals (Bergman et al. 1994; Meissner et al.2006; Tseng et al. 2001; Walters et al. 2007; Wichmann et al.1999). The fact that bursting can appear on SNr neurons afterpharmacological manipulations of their synaptic inputs and, inaddition, the fact that all dopaminergic actions on SNr neuronsrely on the activity of the dopamine presynaptic receptorspresent at these synaptic inputs (Ibanez-Sandoval et al. 2006;Misgeld et al. 2007) (no dopamine postsynaptic receptors havebeen described for SNr neurons), gives a cue about whydopamine depletion may lead to impaired transmission andbursting: synaptic inputs are impeding bursting. The derange-ment of their control, by dopamine depletion, allows theappearance of this firing pattern. Therefore presynaptic recep-tors become potential targets for therapeutics.
By the same token, CaV3.2 channels turn to be the essentialintrinsic component of the bursting mechanism disclosed byuncontrolled synaptic entries. Therefore these channels, too,become a potential therapeutic target.
It has been proposed that BG select among a group of CPGsto carry out innate or acquired behavioral routines and motorprograms (Barnes et al. 2005; Graybiel 1995, 1996; Grillneret al. 2005a,b, 2006; Takakusaki et al. 2004). The essentialproperties of these circuitries are maintained throughout ver-tebrate evolution (Graybiel 1995; Grillner 2006). Disinhibitionis the mechanism proposed for the BG to release (select) someCPGs and then elicit a motor behavior or synergy (Graybiel1995, 1997, 2004; Grillner 2005a,b, 2006). Therefore BGcircuitry can be seen as part of a set of CPGs.
Although more research is needed into the mechanisms ofSNr firing in animal models of PD to further substantiate thepresent findings, our results indicate that the rate and oscilla-tory models of PD could be made compatible if one thinks thatthe synaptic changes proposed by the rate model will have asa consequence changes in firing pattern. In this respect, CPGtheory proposes that unbalancing synaptic drives draws forthfiring pattern generation.
A C K N O W L E D G M E N T S
The technical assistance of A. Laville is greatly appreciated. We thank Drs.Nicolas Vautrelle and Victor Mendoza for critically commenting the presentmanuscript.
G R A N T S
This work was supported by grants from a Program Project grantIMPULSA 03 to E. Galarraga and J. Bargas, by Consejo Nacional de Cienciay Tecnologıa (Mexico) Grants G34706 to J. Aceves, 42636 to E. Galarraga,and 49484 to J. Bargas, and by grants from Direccion General de Asuntos delPersonal Academico. Universidad Nacional Autonoma de Mexico: IN201607to J. Bargas and IN201507 to E. Galarraga.
R E F E R E N C E S
Albin RL, Young AB, Penney JB. The functional anatomy of basal gangliadisorders. Trends Neurosci 12: 366–375, 1989.
Alford S, Schwartz E, Di Prisco GV. The pharmacology of vertebrate spinalcentral pattern generators. Neuroscientist 9: 217–228, 2003.
Atherton JF, Bevan MD. Ionic mechanisms underlying autonomous actionpotential generation in the somata and dendrites of GABAergic substantianigra pars reticulata neurons in vitro. J Neurosci 25: 8272–8281, 2005.
Bargas J, Ayala CX, Vilchis C, Pineda JC, Galarraga E. Ca2�-activatedoutward currents in neostriatal neurons. Neurosci 88: 479–488, 1999.
Barnes TD, Kubota Y, Hu D, Jin DZ, Graybiel AM. Activity of striatalneurons reflects dynamic encoding and recoding of procedural memories.Nature 437: 1158–1161, 2005.
Bergman H,Wichmann T, Karmon B, Delong MR. The primate subthalamicnucleus. II. Neuronal activity in the MPTP model of parkinsonism. J Neu-rophysiol 72: 507–520, 1994.
Beurrier C, Ben-Ari Y, Hammond C. Preservation of the direct and indirectpathways in an in vitro preparation of the mouse basal ganglia. Neuroscience140: 77–86, 2006.
Beurrier C, Congar P, Bioulac B, Hammond C. Subthalamic nucleusneurons switch from single-spike activity to burst-firing mode. J Neurosci19: 599–609, 1999.
Bevan MD, Magill PJ, Terman D, Bolam JP, Wilson CJ. Move to therhythm: oscillations in the subthalamic nucleus-external globus pallidusnetwork. Trends Neurosci 25: 525–531, 2002.
Day M, Wang Z, Ding J, An X, Ingham CA, Shering AF, Wokosin D, IlijicE, Sun Z, Sampson AR, Mugnaini E, Deutch AY, Sesack SR, Arbuth-nott GW, Surmeier DJ. Selective elimination of glutamatergic synapses onstriatopallidal neurons in Parkinson disease models. Nat Neurosci 9: 251–259, 2006.
2321BURSTING IN SUBSTANTIA NIGRA RETICULATA NEURONS
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org

Enomoto E, Kogo M, Koizumi K, Ishihama K, Yamanishi T. Localizationof premotoneurons for an NMDA-induced repetitive rhythmical activity toTMNs. Neuroreport 13: 2303–2307, 2002.
Gerfen CR, Baimbridge KG, Miller JJ. The neostriatal mosaic: compart-mental distribution of calcium-binding protein and parvalbumin in the basalganglia of the rat and monkey. Proc Natl Acad Sci USA 82: 8780–8784,1985.
Gongora-Alfaro JL, Hernandez-Lopez S, Flores-Hernandez J, GalarragaE. Firing frequency modulation of substantia nigra reticulata neurons by5-hydroxytryptamine. Neurosci Res 29: 225–231, 1997.
Gonzalez-Hernandez T, Rodrıguez M. Compartmental organization andchemical profile of dopaminergic and GABAergic neurons in the substantianigra of the rat. J Comp Neurol 421: 107–135, 2000.
Gordon IT, Whelan PJ. Deciphering the organization and modulation ofspinal locomotor central pattern generators. J Exp Biol 209: 2007–2014,2006.
Graybiel AM. Building action repertoires: memory and learning functions ofthe basal ganglia. Curr Opin Neurobiol 5: 733–741, 1995.
Graybiel AM. Basal ganglia: new therapeutic approaches to Parkinson�sdisease. Current Biol 6: 368–371, 1996.
Graybiel AM. The basal ganglia and cognitive pattern generators. SchizophrBull 23: 459–469, 1997.
Graybiel AM. Network-level neuroplasticity in cortico-basal ganglia path-ways. Parkinsonism Relat Disord. 10: 293–296, 2004.
Graybiel AM. The basal ganglia: learning new tricks and loving it. Curr OpinNeurobiol 15: 638–644, 2005.
Grillner S. Biological pattern generation: the cellular and computational logicof networks in motion. Neuron 52: 751–766, 2006.
Grillner S, Hellgren J, Menard A, Saitoh K, Wikstrom MA. Mechanismsfor selection of basic motor programs—roles for the striatum and pallidum.Trends Neurosci 28: 364–370, 2005a.
Grillner S, Markram H, De Schutter E, Silberberg G, LeBeau FE.Microcircuits in action–from CPGs to neocortex. Trends Neurosci 28:525–533, 2005b.
Grillner S, McClellan A, Sigvardt K, Wallen P, Wilen M. Activation ofNMDA-receptors elicits “fictive locomotion” in lamprey spinal cord in vitro.Acta Physiol Scand 113: 549–551, 1981.
Guertin PA, Hounsgaard J. NMDA-induced intrinsic voltage oscillationsdepend on L-type calcium channels in spinal motoneurons of adult turtles.J Neurophysiol 80: 3380–3382, 1998.
Hammond C, Yelnik J. Intracellular labelling of rat subthalamic neuroneswith horseradish peroxidase: computer analysis of dendrites and character-ization of axon arborization. Neuroscience 8: 781–790, 1983.
Hochman S, Jordan LM, MacDonald JF. N-Methyl-D-aspartate receptor-mediated voltage oscillations in neurons surrounding the central canal inslices of rat spinal cord. J Neurophysiol 72: 567–577, 1994.
Hsiao C, del Negro CA, Trueblood PR, Chandler SH. Ionic basis forserotonin-induced bistable membrane properties in guinea pig trigeminalmotoneurons. J Neurophysiol 79: 2847–2856, 1998.
Hutchinson WD, Dostrovsky JO, Walters JR, Courtemanche R, BoraudT, Goldberg J, Brown P. Neuronal oscillations in the basal ganglia andmovement disorders: evidence from whole animal and human recordings.J Neurosci 24: 9240–9243, 2004.
Ibanez-Sandoval O, Carrille-Reid L, Galarraga E, Tapia D, Mendoza E,Gomora JC, Aceves J, Bargas J. Firing patterns and membrane potentialoscillations in projection neurons of the substantia nigra pars reticulata.Soc Neurosci Abst 35: 298–301, 2005.
Ibanez-Sandoval O, Hernandez A, Floran B, Galarraga E, Tapia D,Valdiosera R, Erlij D, Aceves J, Bargas J. Control of the subthalamicinnervation of substantia nigra pars reticulata by D1 and D2 dopaminereceptors. J Neurophysiol 95: 1800–1811, 2006.
Iribe Y, Moore K, Pang KC, Tepper JM. Subthalamic stimulation-inducedsynaptic responses in substantia nigra pars compacta dopaminergic neuronsin vitro. J Neurophysiol 82: 925–933, 1999.
Izhikevich EM. Dynamical Systems in Neuroscience. Cambridge, MA: MITPress, 2007.
Johnson SW, Seutin V, North RA. Burst firing in dopamine neurons inducedby N-methyl-D-aspartate: role of electrogenic sodium pump. Science 258:665–667, 1992.
Kaneda K, Tachibana Y, Imanishi M, Kita H, Shigemoto R, Nambu A,Takada M. Down-regulation of metabotropic glutamate receptor 1� inglobus pallidus and substantia nigra of parkinsonian monkeys. Eur J Neu-rosci 22: 3241–3254, 2005.
Kiehn O. Locomotor circuits in the mammalian spinal cord. Annu RevNeurosci 29: 279–306, 2006.
Kita H. Neostriatal and globus pallidus stimulation induced inhibitorypostsynaptic potentials in entopeduncular neurons in rat brain slice prepa-rations. Neuroscience 105: 871–879, 2001.
Kita H, Kitai ST. Glutamate decarboxylase immunoreactive neurons in ratneostriatum: their morphological types and populations. Brain Res 447:346–352, 1988.
Kochegarov AA. Pharmacological modulators of voltage-gated calcium chan-nels and their therapeutical application. Cell Calcium 33: 145–162, 2003.
Lee CR, Tepper JM. A calcium-activated nonselective cation conductanceunderlies the plateau potential in rat substantia nigra GABAergic neurons.J Neurosci 27: 6531–6541, 2007.
Lee JH, Gomora JC, Cribbs LL, Perez-Reyes E. Nickel block of threecloned T-type calcium channels: low concentrations selectively block �1H.Biophys J 77: 3034–3042, 1999.
Lee RH, Heckman CJ. Bistability in spinal motoneurons in vivo: systematicvariations in persistent inward currents. J Neurophysiol 80: 583–593, 1998.
Loucif KC, Wilson CL, Baig R, Lacey MG, Stanford IM. Functionalinterconnectivity between the globus pallidus and the subthalamic nucleus inthe mouse brain slice. J Physiol 567: 977–987, 2005.
Magill PJ, Bolam JP, Bevan MD. Dopamine regulates the impact of thecerebral cortex on the subthalamic nucleus-globus pallidus network. Neu-roscience 106: 313–330, 2001.
Mallet N, Ballion B, Le Moine C, Gonon F. Cortical inputs and GABAinterneurons imbalance projection neurons in the striatum of parkinsonianrats. J Neurosci 26: 3875–3884, 2006.
Martin RL, Lee JH, Cribbs LL, Perez-Reyes E, Hanck DA. Mibefradilblock of cloned T-type calcium channels. J Pharmacol Exp Ther 295:302–308, 2000.
Meissner W, Ravenscroft P, Reese R, Harnack D, Morgenstern R, KupschA, Klitgaard H, Bioulac B, Gross CE, Bezard E, Boraud T. Increasedslow oscillatory activity in substantia nigra pars reticulata triggers abnormalinvoluntary movements in the 6-OHDA-lesioned rat in the presence ofexcessive extracellular striatal dopamine. Neurobiol Dis 22: 586–598, 2006.
Misgeld U, Drew G, Yanovsky Y. Presynaptic modulation of GABA releasein the basal ganglia. Prog Brain Res 160: 245–259, 2007.
Murer MG, Tseng KY, Kasanetz F, Belluscio M, Riquelme LA. Brainoscillations, medium spiny neurons, and dopamine. Cell Mol Neurobiol 22:611–632, 2002.
Nakanishi H, Kita H, Kitai ST. (1987) Intracellular study of rat substantianigra pars reticulata neurons in an vitro slice preparation: electrical mem-brane properties and response characteristics to subtalamic stimulation.Brain Res 437: 45–55, 1987.
Obeso JA, Rodriguez-Oroz MC, Rodriguez M, Arbizu J, Gimenez-AmayaJM. The basal ganglia and disorders of movement: pathophysiologicalmechanisms. News Physiol Sci 17: 51–55, 2002.
Parent A, Hazrati LN. Functional anatomy of the basal ganglia. II. The placeof subthalamic nucleus and external pallidum in basal ganglia circuitry.Brain Res Rev 20: 128–154, 1995.
Perez-Rosello T, Figueroa A, Salgado H, Vilchis C, Tecuapetla F, GuzmanJN, Galarraga E, Bargas J. Cholinergic control of firing pattern andneurotransmission in rat neostriatal projection neurons: role of CaV2.1 andCaV2.2 Ca2� channels. J Neurophysiol 93: 2507–2519, 2005.
Pineda JC, Galarraga E, Bargas J, Cristancho JM, Aceves J. Charybdo-toxin and apamin sensitivity of the calcium-dependent repolarization and theafterhyperpolarization in neostriatal neurons. J Neurophysiol 287–294,1992.
Plenz D, Kitai ST. A basal ganglia pacemaker formed by the subthalamicnucleus and external globus pallidus. Nature 400: 677–682, 1999.
Richards CD, Shiroyama T, Kitai ST. Electrophysiological and immunocy-tochemical characterization of GABA and dopamine neurons in the sub-stantia nigra of the rat. Neuroscience 80: 545–557, 1997.
Robinson RB, Siegelbaum SA. Hyperpolarization-activated cation currents:from molecules to physiological function. Annu Rev Physiol 65: 453–480,2003.
Robledo P, Feger J. Excitatory influence of rat subthalamic nucleus tosubstantia nigra pars reticulata and the pallidal complex: electrophysiolog-ical data. Brain Res 518: 47–54, 1990.
Sanchez-Vives MV, McCormick DA. Cellular and network mechanisms ofrhythmic recurrent activity in neocortex. Nat Neurosci 3: 27–034, 2000.
Schmitt DE, Hill RH, Grillner S. The spinal GABAergic system is a strongmodulator of burst frequency in the lamprey locomotor network. J Neuro-physiol 92: 2357–2367, 2004.
2322 IBANEZ-SANDOVAL ET AL.
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org

Schweitz H, Heurteaux C, Bois P, Moinier D, Romey G, Lazdunski M.Calcicludine, a venom peptide of the Kunitz-type protease inhibitor family,is a potent blocker of high-threshold Ca2� channels with a high affinity forL-type channels in cerebellar granule neurons. Proc Natl Acad Sci USA 91:878–882, 1994.
Smith Y, Bevan MD, Shink E, Bolam JP. Microcircuitry of the direct andindirect pathways of the basal ganglia. Neuroscience 86: 353–387, 1998.
Smith Y, Bolam JP. Convergence of synaptic inputs from the striatum and theglobus pallidus onto identified nigrocollicular cells in the rat: a doubleanterograde labelling study. Neuroscience 144: 45–73, 1991.
Takakusaki K, Saitoh K, Harada H, Kashiwayanagi M. Role of basalganglia-brain stem pathways in the control of motor behaviors. Neurosci Res50: 137–151, 2004.
Talley EM, Cribbs LL, Lee JH, Daud A, Perez-Reyes E, Bayliss DA. Differ-ential distribution of three members of a gene family encoding low voltage-activated (T-type) calcium channels. J Neurosci 19: 1895–1911, 1999.
Tseng KY, Kasanetz F, Kargieman L, Pazo JH, Murer MG, Riquelme LA.Subthalamic nucleus lesions reduce low frequency oscillatory firing ofsubstantia nigra pars reticulata neurons in a rat model of Parkinson’s disease.Brain Res 904: 93–103, 2001.
Vergara C, LaTorre R, Marrion NV, Ademan JP. Calcium-activatedpotassium channels. Curr Opin Neurobiol 8: 321–329, 1998.
Vergara R, Rick C, Hernandez-Lopez S, Laville JA, Guzman JN, Galar-raga E, Surmeier DJ, Bargas J. Spontaneous voltaje oscillations in striatalprojection neurons in a rat corticostriatal slice. J Physiol 553: 169–182,2003.
Wallen P, Grillner S. N-methyl-D-aspartate receptor-induced, inherent oscil-latory activity in neurons active during fictive locomotion in the lamprey.J Neurosci 7: 2745–2755, 1987.
Walters JR, Hu D, Itoga CA, Parr-Brownlie LC, Bergstrom DA. Phaserelationships support a role for coordinated activity in the indirect pathwayin organizing slow oscillations in basal ganglia output after loss of dopa-mine. Neuroscience 144: 762–776, 2007.
Wichmann T, Bergman H, Starr PA, Subramanian T, Watts RL, DeLongMR. Comparison of MPTP-induced changes in spontaneous neuronal dis-charge in the internal pallidal segment and in the substantia nigra parsreticulata in primates. Exp Brain Res 125: 397–409, 1999.
Wichmann T, DeLong MR. Pathophysiology of Parkinson�s disease: theMPTP primate model of the human disorder. Ann NY Acad Sci 991:199–213, 2003.
Windels F, Carcenac C, Poupard A, Savasta M. Pallidal origin of GABArelease within the substantia nigra pars reticulata during high-frequencystimulation of the subthalamic nucleus. J Neurosci 25: 5079–5086, 2005.
Yanovsky Y, Zhang W, Misgeld U. Two pathways for the activation ofsmall-conductance potassium channels in neurons of substantia nigra parsreticulata. Neuroscience 136: 1027–1036, 2005.
Yoshida M, Precht W. Monosynaptic inhibition of neurons of the substantianigra by caudato-nigral fibers. Brain Res 32: 225–228, 1971.
Yuan H, Yamada K, Inagakia N. Multiminute oscillations in mouse sub-stantia nigra pars reticulata neurons in vitro. Neurosci Lett 355: 136–140,2004.
Yuste R, MacLean JN, Smith J, Lansner A. The cortex as a central patterngenerator. Nat Rev Neurosci 6: 477–483, 2005.
Zhu ZT, Munhall A, Shen KZ, Johnson SW. Calcium-dependent subthresh-old oscillations determine bursting activity induced by N-methyl-D-aspartatein rat subthalamic neurons in vitro. Eur J Neurosci 19: 1296–1304,2004.
2323BURSTING IN SUBSTANTIA NIGRA RETICULATA NEURONS
J Neurophysiol • VOL 98 • OCTOBER 2007 • www.jn.org