buoyancy of some palaeozoic ammonoids and their hydrostatic properties based on empirical 3d-models
TRANSCRIPT

Buoyancy of some Palaeozoic ammonoids and theirhydrostatic properties based on empirical 3D-models
CAROLE NAGLIK, FARHAD RIKHTEGAR AND CHRISTIAN KLUG
Naglik, C., Rikhtegar, F. & Klug, C. 2015: Buoyancy of some Palaeozoic ammonoidsand their hydrostatic properties based on empirical 3D-models. Lethaia, DOI:10.1111/let.12125.
The interpretation of the function of the ammonoid phragmocone as a buoyancydevice is now widely accepted among ammonoid researchers. During the 20th century,several theoretical models were proposed for the role of the chambered shell(phragmocone); accordingly, the phragmocone had hydrostatic properties, whichenabled it to attain neutral buoyancy, presuming it was partially filled with gas. Withnew three-dimensional reconstructions of ammonoid shells, we are now able to testthese hypothetical models using empirical volume data of actual ammonoid shells. Weinvestigated three Palaeozoic ammonoids (Devonian and Carboniferous), namely Fid-elites clariondi, Diallagites lenticulifer and Goniatites multiliratus, to reconstruct theirhydrostatic properties, their syn vivo shell orientation and their buoyancy. Accordingto our models, measurements and calculations, these specimens had aperture orienta-tions of 19°, 64° and 125° during their lives. Although none of our results coincidewith the aperture orientation of the living Nautilus, they do verify the predictions forshell orientations based on published theoretical models. Our calculations also showthat the shorter the body chamber, the poorer was the hydrodynamic stability of theanimal. This finding corroborates the results of theoretical models from the 1990s.With these results, which are based on actual specimens, we favour the rejection ofhypotheses suggesting a purely benthonic mode of life of ammonoids. Additionally, itis now possible to assess hydrodynamic properties of the shells through ontogeny andphylogeny, leading to insights to validate theoretical modes of life and habitat throughthe animal’s life. □ Ammonoidea, buoyancy, grinding tomography, palaeozoic, shell ori-entation, virtual palaeontology.
Carole Naglik [[email protected]], and Christian Klug [[email protected]],Pal€aontologisches Institut und Museum, Universit€at Z€urich, Karl-Schmid-Strasse 4, Z€urich8006, Switzerland; Farhad Rikhtegar [[email protected]], Laboratory of Thermodynamicsin Emerging Technologies, Department of Mechanical and Process Engineering, ETHZ€urich, Z€urich, Switzerland; manuscript received on 27/05/2014; manuscript accepted on30/10/2014.
The function of ammonoid shells as buoyancydevices has been an intriguing topic for naturalistsfor several centuries (e.g. Derham 1726; Owen 1832;Vrolik 1843; Meigen 1870; Willey 1902). In the pastseven decades, the buoyancy of ammonoids has beentheoretically and experimentally studied and docu-mented, starting with the notable pioneer work ofTrueman (1940). Subsequently, numerous authorshave dealt with this topic (e.g. Raup & Chamberlain1967; Saunders & Shapiro 1986; Shigeta 1993;Kr€oger 2002; Klug & Korn 2004; Longridge et al.2009; Tajika et al. 2014). Especially the importantresearch on buoyancy regulation in Recent Nautilida(e.g. Denton 1962, 1974; Denton & Gilpin-Brown1966; Ward & Martin 1978; Ward 1979, 1980, 1982,1986, 1987; Collins et al. 1980; Greenwald et al.1980; Kanie et al. 1980; Ward et al. 1981; Chamber-lain & Moore 1982; Greenwald & Ward 1982; Man-gum & Towle 1982; Chamberlain 1987; Greenwald& Ward 1987; Shapiro & Saunders 1987; Crick 1988;Dunstan et al. 2011; Tsujino & Shigeta 2012) has
promoted an understanding of the mode of life bothof nautilids and ammonoids.
As far as ammonoids are concerned, however,most of the investigations have been carried out onMesozoic material (predominantly Jurassic and Cre-taceous; Trueman 1940; Raup & Chamberlain 1967;Kr€oger 2002; Longridge et al. 2009; Tajika et al.2014; Hoffmann et al. 2015). Few studies haveassessed the buoyancy of Palaeozoic ammonoids,those that have commonly focusing on Carbonifer-ous material (Saunders & Shapiro 1986; Swan &Saunders 1987; Saunders & Work 1996). Even fewerauthors tried to reconstruct buoyancy and locomo-tion for Devonian forms on theoretical grounds(Klug 2001; Klug & Korn 2004; Saunders et al.2004). Globally, the palaeobiology of early ammo-noids has been much less studied compared to youn-ger representatives of this clade. One of the possiblereasons for this difference is the often poorer preser-vation (e.g. no nacre preservation in the Devonian)and the lesser abundance of Palaeozoic outcrops and
DOI 10.1111/let.12125 © 2015 Lethaia Foundation. Published by John Wiley & Sons Ltd

thus the lower availability of suitable material com-pared with the Mesozoic.
Nowadays, rapid technological progresses in phys-ics and computer sciences have provided benefit toan increasing number of palaeontologists, and sincethe last decade, methods of ‘virtual palaeontology’have been further developed and widely used (e.g.Sutton et al. 2001; Garwood et al. 2010; Sutton et al.2014; Longridge et al. 2009). So far, only a few casesof the application of virtual palaeontological meth-ods to ammonoids have been published (e.g. Long-ridge et al. 2009; Kruta et al. 2011; Lukeneder 2012;Hoffmann & Zachow 2011; Hoffmann et al. 2014;Hoffmann et al. 2015; Tajika et al. 2014; Nagliket al. 2015a). Hoffmann et al. (2014) published adiscussion paper on the non-invasive methodsapplied in cephalopod research, where they reviewedthe wealth of research possibilities these new meth-ods are able to offer.
Our present study focuses on the buoyancy ofthree empirical three-dimensional models of threePalaeozoic ammonoid species. Because of the com-mon lack of density contrast between ammonoidshell material and surrounding sediments, our speci-mens could not be investigated with a non-invasiveapproach (Garwood et al. 2010; Tajika et al. 2014;Lukeneder et al. 2014; Naglik et al. 2015a). Thisproblem was resolved by employing the serial sec-tioning/grinding tomography method (Sollas 1904;Garwood et al. 2010), which is indeed a destructivemethod, but which on the other hand usually deliv-ers sufficiently highly resolved image stacks contain-ing colour information. Such image data can beevaluated in order to acquire volumetric data. Thisgrinding tomography method has been applied tothree specimens, two of Devonian and one of Car-boniferous age.
The aims of this study were: (1) to determine themasses of the various shell parts and the soft tissuesas well as other masses affecting the buoyancy ofsome Palaeozoic ammonoids; (2) to determine thebuoyancy of the shells of these ammonoids andthereby answer the question whether they could pos-sibly achieve neutral buoyancy by means of theirphragmocones; (3) to reconstruct their syn vivo shellorientation; and (4) to use this information to dis-cuss their swimming capabilities.
Material and methods
The empirical datasets used here are the same asthose recently published by Naglik et al. (2015a).Three specimens of Palaeozoic ammonoids wereserially ground and virtually reconstructed. These
three specimens were the Middle Devonian(Eifelian) agoniatitid Fidelites clariondi Petter 1959and the anarcestid Diallagites lenticulifer Klug 2002(both belonging to the order Agoniatitida), and theEarly Carboniferous (Visean or Serpukhovian) goni-atitid Goniatites multiliratus Gordon 1962. Both a-goniatitids come from the Anti-Atlas region ofMorocco, and the goniatitid comes from Oklahomain the USA (details of the bearing lithologies in Nag-lik et al. 2015a).
All specimens have been automatically groundinto slices of a thickness of ~0.05 mm per increment.Each ground surface (slice) has been scanned andeach fourth scan of the obtained image stack hasbeen virtually retraced using Adobe Illustrator andPhotoshop before being computed in VGstudio-max2.1 (Volume Graphics GmbH, Heidelberg, Ger-many). Technical details are available in Naglik et al.(2015a). Due to the destructive quality of themethod, no repository numbers of the specimens aregiven; pictures taken of the specimens prior togrinding are provided in Naglik et al. (2015a) andthe original stacks are available upon request to C.Naglik or C. Klug.
Labelling of shell parts
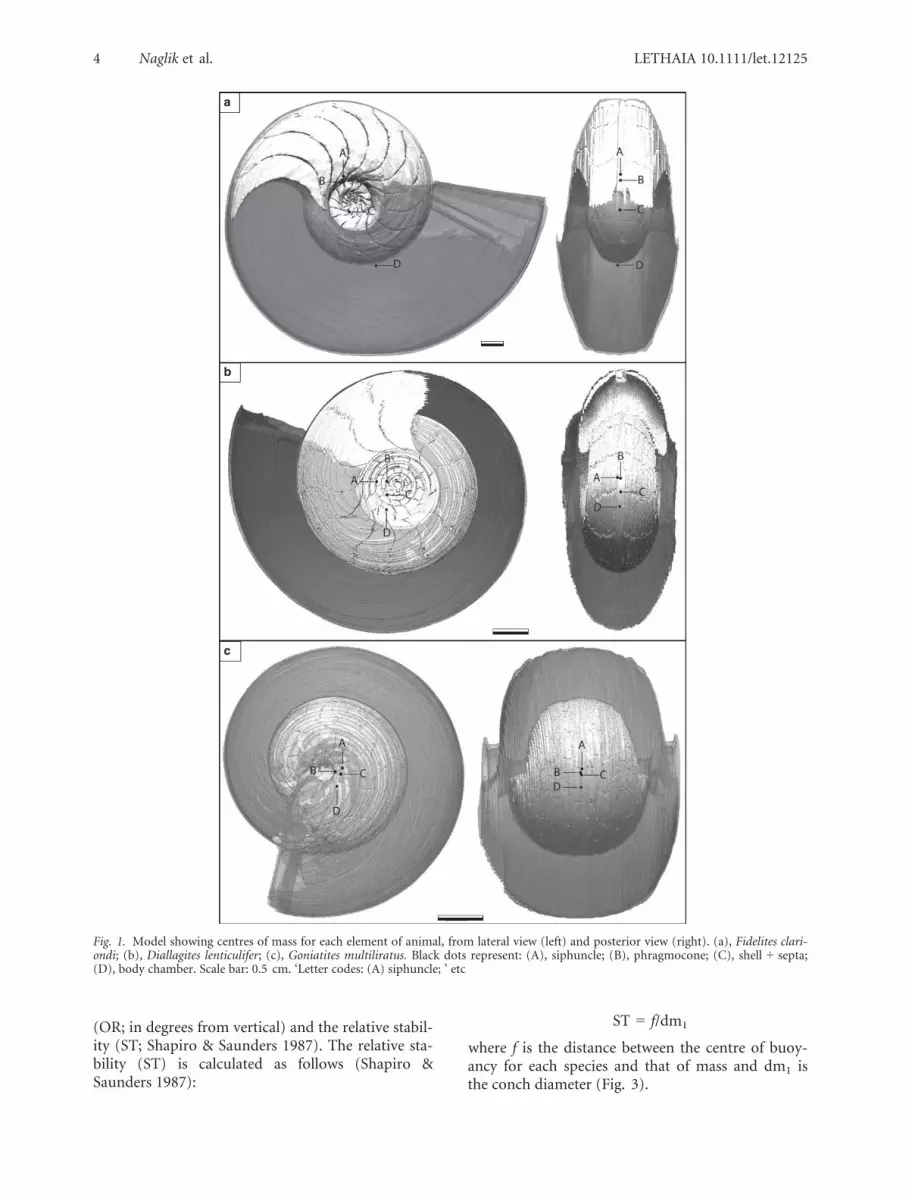
In each fourth slice, the various parts of the shellwith a specific density, i.e. the siphuncle, the cham-bers and the body chamber (segments), were labelledwith a different colour. A STL file (STereoLithogra-phy ASCII) consisting of a mesh of triangles hasbeen created of each segment of the models usingVGstudiomax 2.1. As the polygon-rendering stepgenerated some artifacts in the mesh such as smallholes or overlapping triangles, the meshes have thenbeen processed in Geomagic Studio (Geomagic, Inc.,Morrisville, NC, USA) in order to smooth and repairthem (using built-in functions in the software).Once this step was completed, the obtained volumes(Table 1) were measured directly in the software,and the centres of mass of each segment could bedirectly computed (see Fig. 1 for the centre of massof each segment, for each specimen).
Approximations for calculations
Knowing the volumes of all segments, their massescould be determined using some density approxima-tions from the known densities of equivalent materi-als, which occur in Recent Nautilus. Today’snautilids are the most commonly used actualisticanalogues for studying buoyancy of ammonoids(e.g. Trueman 1940; Saunders & Shapiro 1986;Jacobs & Chamberlain 1996; Westermann 1998,
2 Naglik et al. LETHAIA 10.1111/let.12125

2013; Klug 2001; Kr€oger 2002; Kruta et al. 2014; Taj-ika et al. 2014). Accordingly, we attributed a densityof 1.055 g/cm3 to the soft body (following Saunders& Shapiro 1986). Concerning the soft body, we fol-low the hypothesis assuming that the soft body vol-ume corresponds to the volume of the bodychamber minus the volume of the mantle cavity,which was filled with water (e.g. Shapiro & Saunders1987; Kr€oger 2002).
In the cases of Diallagites and Goniatites, the bodychambers were incompletely preserved. Therefore,we examined more complete specimens from thesame localities (e.g. Klug 2002) and used literaturedata in order to reconstruct their body chamberlength (Klug 2001; Saunders et al. 2004). Dependingon the discrepancy between the preserved bodychamber length and those either reported in the lit-erature and/or known from other specimens at ourdisposal, we removed the last five septa of Diallagiteslenticulifer and the last 16 septa of Goniatites multili-ratus in order to obtain a presumed complete recon-struction of their body chambers.
All three specimens were more or less mature(Naglik et al. 2015a). Using a value for the bodychamber length that is as accurate as possible is ofutmost importance, since one of the most importantfactors involved in the control of the stability in Nau-tilus (and, by analogy, in ammonoids) is the bodychamber length (Shapiro & Saunders 1987). Follow-ing Longridge et al. (2009), we assumed a density of1.055 g/cm3 for the siphuncle. We could not applythe method suggested in Tajika et al. (2014), becausethe connecting rings of the siphuncle could not bereconstructed in all our specimens. Concerning theshell and septa, there are several different published
density values ranging from 2.61 to 2.69 g/cm3 (e.g.Reyment 1958; Kr€oger 2002; Longridge et al. 2009;Hoffmann et al. 2015). Reyment (1958) explainedthat, although the Nautilus shell has a density ofaround 2.61–2.62 g/cm3, its septa contain less organicmaterial and thus have densities of 2.93–2.94 g/cm3,corresponding to the specific density of aragonite inits pure state. Therefore, we compared the centre ofmass positions using 2.62 g/cm3 and using mean val-ues obtained for each single model (calculated mean‘shell + septa’ densities between 2.64 and 2.66 g/cm3).This difference turned out to only insignificantly alterour results; therefore, we decided to keep the com-monly used 2.62 g/cm3 value (Reyment 1958; Saun-ders & Shapiro 1986; Shapiro & Saunders 1987;Kr€oger 2002; Tajika et al. 2014).
Centres of mass and buoyancy
The centre of mass has been calculated using a sim-ple barycentre equation, determining the respectivemass for each coordinate of every segment. Thecoordinates of the centre of mass (X;Y;Z) follows thisbarycentre equation:
X = (mshell+septa * Xshell+septa + msoft part *Xsoft part + mphragmocone * Xphragmocone
+ msiphuncle * Xsiphuncle)/mshell+septa + msoft part
+ mphragmocone + msiphuncle;Y = (mshell+septa * Yshell+septa + msoft part *
Ysoft part + mphragmocone * Yphragmocone
+ msiphuncle * Ysiphuncle)/mshell+septa
+ msoft part + mphragmocone + msiphuncle;Z = (mshell+septa * Zshell+septa+ msoft part * Zsoft part
+ mphragmocone * Zphragmocone + msiphuncle *Zsiphuncle)/mshell+septa+ msoft part+ mphragmocone
+ msiphuncle;
where m(a) is the mass of each segment a, and Xa; Ya;Za, the coordinates of the centre of each segment.
Then, the resulting coordinates have been com-puted in the models using Geomagic Studio andXOS (Fig. 2). The centre of buoyancy was directlycomputed in Geomagic XOS (Fig. 2). The centre ofbuoyancy is corresponding to the centre of mass ofthe sea water volume displaced by the animal (Tru-eman 1940; Raup & Chamberlain 1967; Saunders &Shapiro 1986; Kr€oger 2002). We take the sameassumption as in Tajika et al. (2014) for these calcu-lations, namely that the phragmocone chamberswere filled with gas.
Shell parameters and ratios
Additionally, we measured the body chamber length(BCL; in degrees), the orientation of the aperture
Table 1. Volume, density and mass of each segment constitutingeach specimen.
Segment Volume (cm3) Density (g/cm3) Mass (g)
Fidelites clariondiShell + septa 4.548 2.62 11.916Living chamber 33.712 1.055 35.57Siphuncle 0.124 1.055 0.131Phragmocone 12.821 0 0Total 51.205 – 47.617
Diallagites lenticuliferShell + septa 1.309 2.62 3.429Living chamber 6.582 1.055 6.944Siphuncle 0.081 1.055 0.086Phragmocone 3.421 0 0Total 11.393 – 10.459
Goniatites multiliratusShell + septa 0.620 2.62 1.624Living chamber 6.053 1.055 6.386Siphuncle 0.019 1.055 0.020Phragmocone 1.643 0 0Total 8.335 – 8.030
LETHAIA 10.1111/let.12125 Palaeozoic ammonoid buoyancy 3
(OR; in degrees from vertical) and the relative stabil-ity (ST; Shapiro & Saunders 1987). The relative sta-bility (ST) is calculated as follows (Shapiro &Saunders 1987):
ST = f/dm1
where f is the distance between the centre of buoy-ancy for each species and that of mass and dm1 isthe conch diameter (Fig. 3).
a
b
c
Fig. 1. Model showing centres of mass for each element of animal, from lateral view (left) and posterior view (right). (a), Fidelites clari-ondi; (b), Diallagites lenticulifer; (c), Goniatites multiliratus. Black dots represent: (A), siphuncle; (B), phragmocone; (C), shell + septa;(D), body chamber. Scale bar: 0.5 cm. ‘Letter codes: (A) siphuncle; ’ etc
4 Naglik et al. LETHAIA 10.1111/let.12125
Results
Shell hydrostatics
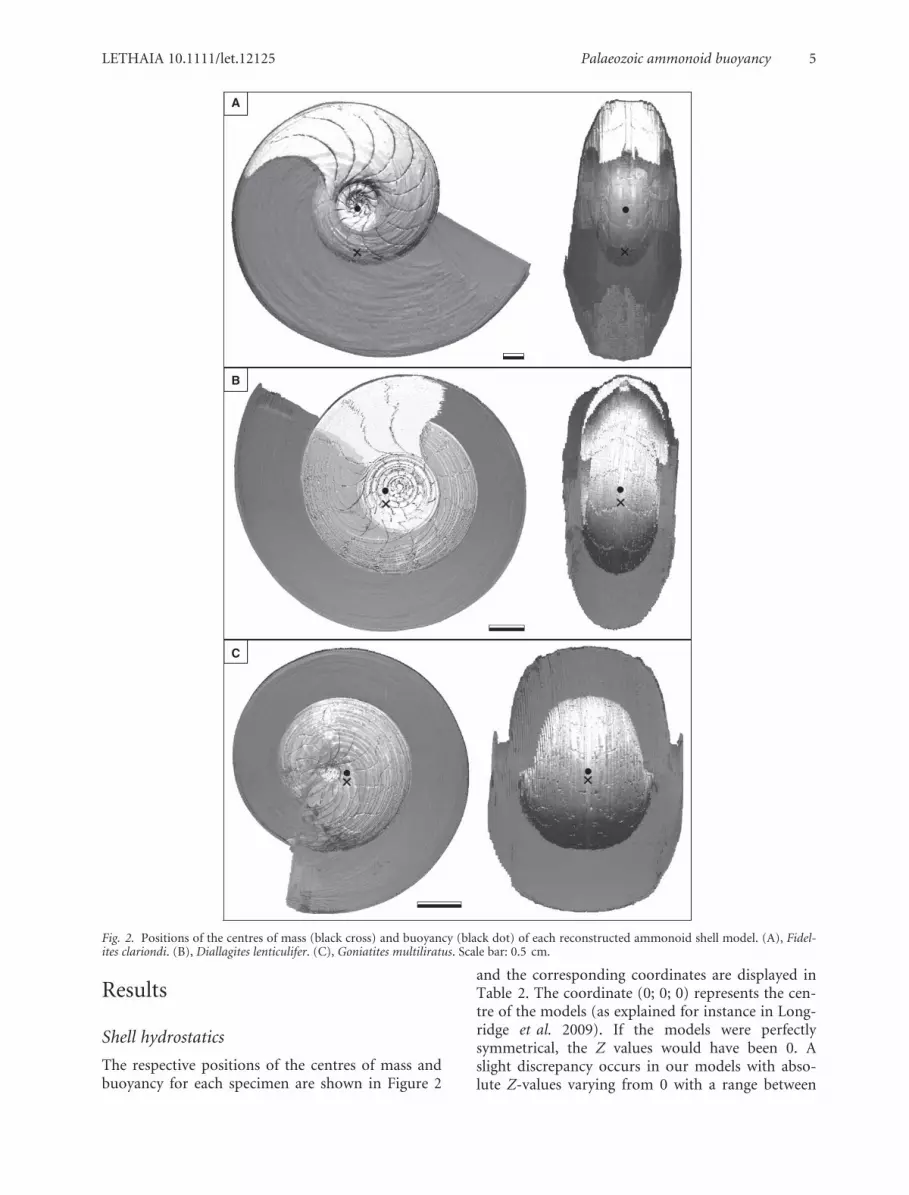
The respective positions of the centres of mass andbuoyancy for each specimen are shown in Figure 2
and the corresponding coordinates are displayed inTable 2. The coordinate (0; 0; 0) represents the cen-tre of the models (as explained for instance in Long-ridge et al. 2009). If the models were perfectlysymmetrical, the Z values would have been 0. Aslight discrepancy occurs in our models with abso-lute Z-values varying from 0 with a range between
A
B
C
Fig. 2. Positions of the centres of mass (black cross) and buoyancy (black dot) of each reconstructed ammonoid shell model. (A), Fidel-ites clariondi. (B), Diallagites lenticulifer. (C), Goniatites multiliratus. Scale bar: 0.5 cm.
LETHAIA 10.1111/let.12125 Palaeozoic ammonoid buoyancy 5

0.014 mm and 0.136 mm. This deviation can haveseveral origins, such as an imperfectly symmetricalshell, a slight asymmetry of the siphuncle, but it canalso result from minor errors during tracing. What-ever the origin of this difference, even if it resultsfrom a small error, this does not compromise therelative positions of the centres of mass and buoy-ancy, and the reconstruction of the syn vivo shell ori-entation in water.
The orientations of the shells shown in Figure 2represent the resting position, with the living orien-tation (i.e., the static orientation of the shell inwater, assuming buoyancy as the main function ofthe shell following Shapiro & Saunders 1987). Thevalues of body chamber lengths (BCL), the orienta-tion of the apertures (OR), the distance between thecentres of mass and buoyancy (f) as well as the rela-tive stability (ST) of each specimen are also given inTable 2. The body chamber length varies from 201°(Fidelites clariondi) to 354° (Goniatites multiliratus)with a length of 282° for Diallagites lenticulifer.Using these values, we reconstructed shell orienta-tions, where the aperture is at 69° from the verticaldirection in F. clariondi, at 125° in D. lenticulifer and19° in G. multiliratus. The former two values appearreasonable and roughly coincide with those mod-elled by Saunders & Shapiro (1986) for similar formsand those modelled by Klug (2001) as well as Klug &Korn (2004) for Devonian taxa.
As far as the hydrodynamic stability is concerned,the distance between the centres of mass and buoy-ancy (f) varies from 10.193 mm to 0.962 mm in thethree ammonoids (Table 2). The direct consequenceof this proximity of the two points is a rather lowrelative hydrodynamic stability. F. clariondi, whichhas a shorter body chamber, possessed a betterhydrodynamic stability with its higher distance f(10.193 mm) and its higher relative stability value of0.13, although this value is still low. D. lenticuliferhas a slightly longer body chamber length and aneven lower stability value of 0.04 with a distance f of1.776. With a hydrodynamic stability value of 0.03,G. multiliratus has both the longest body chamberand the lowest stability value. As suggested bySaunders & Shapiro (1986), a correlation betweenbody chamber length and hydrodynamic stability isapparent, although more data is needed to supportthis (F. clariondi: BCL = 201°, ST = 0.13, D. lentic-ulifer: BCL = 282°, ST = 0.04; G. multiliratus:BCL = 354°, ST = 0.03; Tajika et al. 2014; Norman-nites mitis: BCL = 269°, ST = 0.05). In other words,the longer the body chamber, the poorer the stabil-ity. Nevertheless, this variation in hydrodynamic sta-bility of the shells probably reflects differences in therespective modes of life (Saunders & Shapiro 1986)including more actively horizontally swimmingforms (moderately long body chamber, relativelyhigh ST-value) and other forms, which were mostlikely vertical migrants (very long or very short bodychambers and very low ST-values; Trueman 1940;Westermann 1996).
Buoyancy reconstruction
The virtual reconstructions of these three specimensallowed us to calculate their masses and buoyancies.The volumes, densities and masses of each segmentof each specimen are listed in Table 1. With thesedata, we calculated the total mass of the animalincluding all segments and compare this mass withthe total mass of the displaced seawater. We assumea seawater density of 1.026 g/cm3 (Trueman 1940;Raup & Chamberlain 1967; Saunders & Shapiro1986; Kr€oger 2002; Tajika et al. 2014).
Fig. 3. Parameters used to establish variables and the relativehydrodynamic stability (ST). BCL, body chamber length (indegrees); OR, orientation (in degrees from vertical); f, distancebetween the mass (cross) and the buoyancy (dot) centres; dm1,conch diameter; dm2, conch diameter without the apertureheight (modified from Shapiro & Saunders 1987; with classicalparameter labels from Korn 2010).
Table 2. Mass and buoyancy coordinates of each specimen, as well as their corresponding measured body chamber length (BCL), orien-tation (OR) and their calculated distance between the two centres (f) and their relative stability (ST).
Mass coordinates (mm) Buoyancy coordinates (mm)
BCL (°) OR (°) f (mm) STX Y Z X Y Z
Fidelites clariondi �2.879 �4.981 0.014 �6.971 4.355 0.039 201 69 10.193 0.13Diallagites lenticulifer 0.534 �2.169 �0.326 0.839 �0.428 �0.149 282 125 1.776 0.04Goniatites multiliratus �1.156 �0.116 �0.035 �0.864 0.800 �0.053 354 19 0.962 0.03
6 Naglik et al. LETHAIA 10.1111/let.12125

The total mass of Fidelites clariondi is 47.613 gthen lower than the 52.536 g of displaced water. Thedifference of these two masses is 4.919 g. Similarly,the total mass of Diallagites lenticulifer is 10.459 gand the corresponding mass of displaced wateramounts to 11.689 g, creating a mass difference of1.23 g. Finally, the results for Goniatites multiliratusare congruent with the previous ones, since the totalmass of the animal is 8.030 g compared to 8.552 gfor the corresponding displaced water mass. In thelast case, the mass difference is very low with only0.522 g.
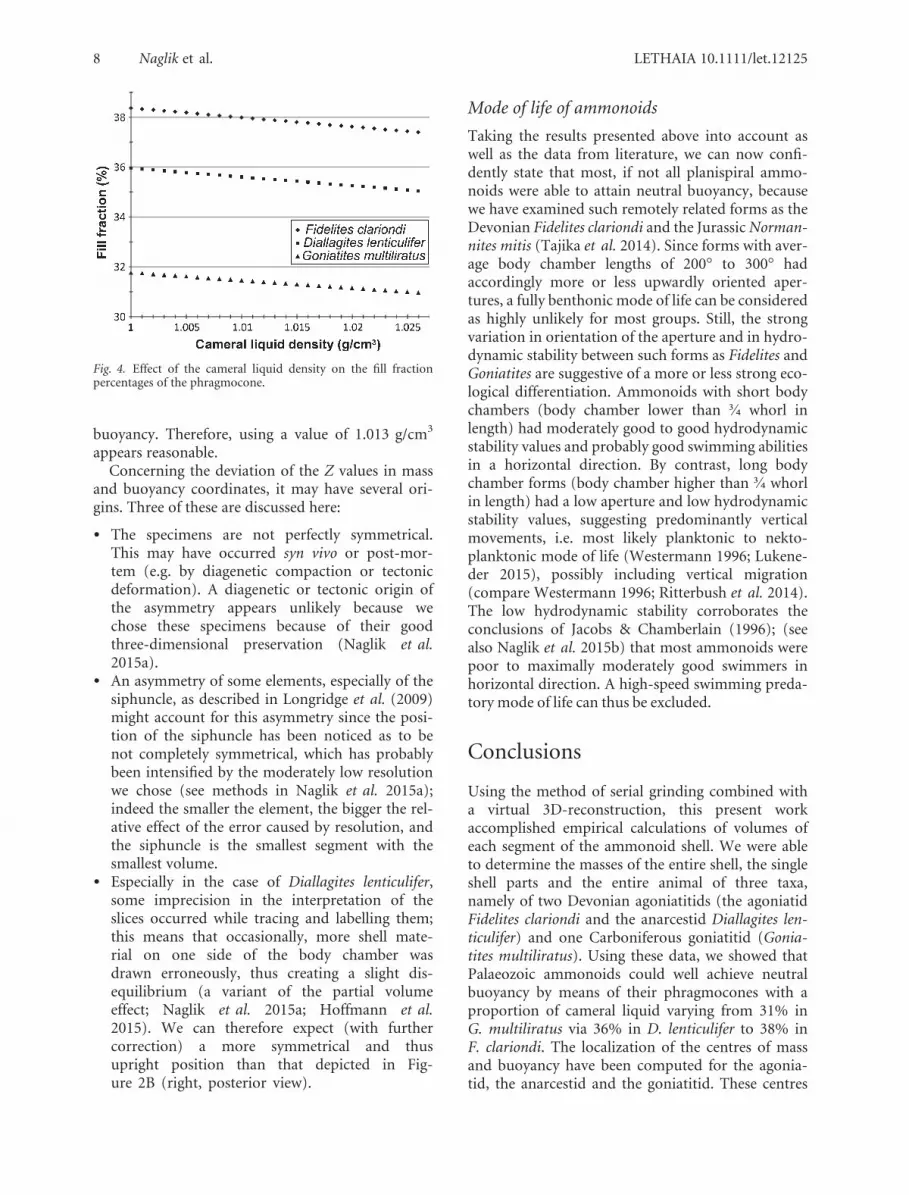
These results show that, under the assumptionthat the phragmocones were filled with only gas, thethree specimens would have been strongly positivelybuoyant, thus causing the animal to stick to thewater surface, which is unlikely. These mass differ-ences represent the maximal theoretical buoyancy ofthese ammonoids (Kr€oger 2002). Hypothesizing thatammonoids were nearly neutrally buoyant, the dif-ference between calculated and neutral buoyancyrepresents the fill fraction (sensu Kr€oger 2002) andprobably equals the mass of chamber liquid (e.g.Kr€oger 2002; Tajika et al. 2014). Assuming a densityof 1.013 g/cm3 for the cameral liquid (Tajika et al.2014), we determined the fill fraction of the cameralliquid needed by the animals to reach neutral buoy-ancy. Accordingly, F. clariondi would have had 38%of the phragmocone filled by cameral liquid, 36% inD. lenticulifer and 31% in G. multiliratus. Theseresults have to be viewed with some reservation sincewe do not know the precise composition of cameralliquid and thus its density, especially if this liquiddiffered in composition from the ambient sea water.Sea water salinity varies with factors such as depth,temperature, distance from fresh water input (brack-ish waters, etc.) and we do not know in detail theprecise environmental conditions of habitats of theexamined species. Nevertheless, these values around30% of cameral liquid are in agreement with whathas been predicted by Heptonstall (1970), Mutvei &Reyment (1973) as well as Reyment (1973) and withwhat has been found by Tajika et al. (2014) for theJurassic ammonite Normannites mitis.
Errors and biases
Hydrostatic properties and buoyancy have beenreconstructed using virtual models of three Palaeo-zoic ammonoid species. Some aspects concerningthe accuracy of such models need to be discussed.The study of Tajika et al. (2014) had the advantagethat Normannites mitis has already been used for abuoyancy study earlier, although Trueman (1940)used evident simplifications in his calculations. The
results are congruent with what has been predictedbased on theoretical models before, taking intoaccount some earlier mistakes such as the shell den-sity value, which was wrong in Trueman (1940), forinstance. On the one hand, this supports the validityof the method used to reconstruct these models, andon the other hand, it shows the minor to negligibleeffect of the resolution limit on the results (Nagliket al. 2015a).
Moreover, some simplifications, generalizationsand approximations have to be discussed here sinceall volumetric models of ammonoids will includesimplifications. This implies that it is not possibleto obtain absolutely accurate volume data, weightvalues and thus buoyancy models of ammonoids.All the hydrostatic and buoyancy calculations inthe literature referring to Raup & Chamberlain’s(1967) equations are based on assumptions, whichwere known to be slightly wrong or not perfectlyappropriate for Nautilus (Shapiro & Saunders1987), such as the circular generating curve and thenon-inclusion of the weight of the shell in the cal-culation of the centre of mass. Still, empiricalexperiments have shown results corresponding wellwith the ones of Raup & Chamberlain (1967).Therefore, we maintain the hypothesis according towhich the soft-tissues are completely included inthe body chamber, as well as the commonly useddensity of 1.055 g/cm3 for the soft body and2.62 g/cm3 for the shell (we tested our models withdensities up to 2.66 g/cm3 without noticeable dif-ferences in the results).
The estimate of the cameral liquid density is morecritical, since we cannot determine its compositionand thus density with certainty; moreover the com-position probably could have differed betweenammonoid taxa, depending on their respective habi-tats, physiology and the corresponding physical andchemical environmental conditions. We hence eval-uated the effect of the cameral liquid density on thefill fraction in percentage of the phragmocone vol-ume (Fig. 4). This figure shows that the change ofthe fill fraction is minor to negligible regarding thefact that we assume density values for the cameralliquid with the same as seawater (1.026 g/cm3). Inall cases, a difference of <1% is observed. We testedsome other densities and the theoretical minimumdensity of cameral liquid to reach neutral buoyancywith more than 90% of phragmocone filling in eachcase, would be a cameral liquid density of 0.4 g/cm3,which appears unlikely. On the other hand, with ahigher -similarly unlikely- density of 2 g/cm3, aliquid filling between only 16% (Goniatites multilira-tus) and 19% (Fidelites clariondi) would be necessaryfor the ammonoid animals to reach neutral
LETHAIA 10.1111/let.12125 Palaeozoic ammonoid buoyancy 7
buoyancy. Therefore, using a value of 1.013 g/cm3
appears reasonable.Concerning the deviation of the Z values in mass
and buoyancy coordinates, it may have several ori-gins. Three of these are discussed here:
• The specimens are not perfectly symmetrical.This may have occurred syn vivo or post-mor-tem (e.g. by diagenetic compaction or tectonicdeformation). A diagenetic or tectonic origin ofthe asymmetry appears unlikely because wechose these specimens because of their goodthree-dimensional preservation (Naglik et al.2015a).
• An asymmetry of some elements, especially of thesiphuncle, as described in Longridge et al. (2009)might account for this asymmetry since the posi-tion of the siphuncle has been noticed as to benot completely symmetrical, which has probablybeen intensified by the moderately low resolutionwe chose (see methods in Naglik et al. 2015a);indeed the smaller the element, the bigger the rel-ative effect of the error caused by resolution, andthe siphuncle is the smallest segment with thesmallest volume.
• Especially in the case of Diallagites lenticulifer,some imprecision in the interpretation of theslices occurred while tracing and labelling them;this means that occasionally, more shell mate-rial on one side of the body chamber wasdrawn erroneously, thus creating a slight dis-equilibrium (a variant of the partial volumeeffect; Naglik et al. 2015a; Hoffmann et al.2015). We can therefore expect (with furthercorrection) a more symmetrical and thusupright position than that depicted in Fig-ure 2B (right, posterior view).
Mode of life of ammonoids
Taking the results presented above into account aswell as the data from literature, we can now confi-dently state that most, if not all planispiral ammo-noids were able to attain neutral buoyancy, becausewe have examined such remotely related forms as theDevonian Fidelites clariondi and the JurassicNorman-nites mitis (Tajika et al. 2014). Since forms with aver-age body chamber lengths of 200° to 300° hadaccordingly more or less upwardly oriented aper-tures, a fully benthonic mode of life can be consideredas highly unlikely for most groups. Still, the strongvariation in orientation of the aperture and in hydro-dynamic stability between such forms as Fidelites andGoniatites are suggestive of a more or less strong eco-logical differentiation. Ammonoids with short bodychambers (body chamber lower than ¾ whorl inlength) had moderately good to good hydrodynamicstability values and probably good swimming abilitiesin a horizontal direction. By contrast, long bodychamber forms (body chamber higher than ¾ whorlin length) had a low aperture and low hydrodynamicstability values, suggesting predominantly verticalmovements, i.e. most likely planktonic to nekto-planktonic mode of life (Westermann 1996; Lukene-der 2015), possibly including vertical migration(compare Westermann 1996; Ritterbush et al. 2014).The low hydrodynamic stability corroborates theconclusions of Jacobs & Chamberlain (1996); (seealso Naglik et al. 2015b) that most ammonoids werepoor to maximally moderately good swimmers inhorizontal direction. A high-speed swimming preda-tory mode of life can thus be excluded.
Conclusions
Using the method of serial grinding combined witha virtual 3D-reconstruction, this present workaccomplished empirical calculations of volumes ofeach segment of the ammonoid shell. We were ableto determine the masses of the entire shell, the singleshell parts and the entire animal of three taxa,namely of two Devonian agoniatitids (the agoniatidFidelites clariondi and the anarcestid Diallagites len-ticulifer) and one Carboniferous goniatitid (Gonia-tites multiliratus). Using these data, we showed thatPalaeozoic ammonoids could well achieve neutralbuoyancy by means of their phragmocones with aproportion of cameral liquid varying from 31% inG. multiliratus via 36% in D. lenticulifer to 38% inF. clariondi. The localization of the centres of massand buoyancy have been computed for the agonia-tid, the anarcestid and the goniatitid. These centres
Fig. 4. Effect of the cameral liquid density on the fill fractionpercentages of the phragmocone.
8 Naglik et al. LETHAIA 10.1111/let.12125

allowed the reconstruction of the syn vivo shell ori-entations with aperture angles (OR) varying fromthe probably erroneously low value of 19° in G. mul-tiliratus via 69° for F. clariondi to 125° in D. lenticu-lifer. These values differ from Nautilus (~42°,Shapiro & Saunders 1987), Normannites mitis (90–100°, Tajika et al. 2014), and from other Palaeozoicammonoid morphologies investigated by Saunders& Shapiro (1986), although they principally coincidewith the discoveries of the latter authors, which werebased on theoretical models. Nevertheless, the orien-tations of the shells of D. lenticulifer and F. clariondiare roughly concordant with the results of Klug(2001). The low angle of G. multiliratus is still rea-sonably congruent with the low angle of 20° to 30°found for the goniatitid morphs in Saunders &Shapiro (1986).
As far as body chamber lengths compared to rela-tive hydrodynamic stabilities of ammonoids are con-cerned, it appears as if specimens with shorter bodychambers had a higher stability value (F. clariondi)than the ones with longer body chambers (D. lentic-ulifer, G. multiliratus). Our results thus confirm thesuggestion of a negative correlation between thebody chamber length and stability (Saunders &Shapiro 1986). In other words, the longer the bodychamber, the poorer the hydrodynamic stability.This variation in shell orientation and hydrody-namic stability probably reflects differences in themode of life. Long body chamber forms with lowapertures and low stability values might have beenrather vertical migrants while short body chamberforms with high stability values and horizontal aper-tures were the better swimmers, although high speedswimming can be ruled out for planispiral ammo-noids (albeit this statement can probably be inferredto heteromorphs as well).
Acknowledgements. – This work was financially supported by theSwiss National Science Foundation SNF (project numbers200021-113956/1, 200020-25029, and 200020-132870). Thegrinding tomography was carried by Stefan G€otz (in memoriam),Enric Pascual-Cebrian and Dominik Hennh€ofer (Heidelberg).We thank the Laboratory of Thermodynamics in Emerging Tech-nologies of ETH Z€urich for giving the initiation of the use of Ge-omagic STUDIO software for mesh repairs. We especially wantto thank Alik Huseynov from the Anthropological Institute andMuseum of the University of Z€urich who kindly let us use thefacilities and allowed us to achieve our work with the softwareGeomagic XOS for measurements. We thank Alexander Lukene-der as well as one anonymous reviewer who helped to improvethe quality of this article.
References
Chamberlain, J.A. Jr 1987: Locomotion of Nautilus. In Saunders,W.B. & Landman, N.H. (eds): Nautilus – The Biology andPaleobiology of a Living Fossil, 489–525. Springer, Dordrecht.
Chamberlain, J.A. Jr & Moore, W.A. Jr 1982: Rupture strength andflow rate of Nautilus siphuncular tube. Paleobiology 8, 408–425.
Collins, D.H., Ward, P.D. & Westermann, G.E.G. 1980: Functionof cameral water in Nautilus. Paleobiology 6, 168–172.
Crick, R.E. 1988: Buoyancy regulation and macroevolution innautiloid cephalopods. Senckenbergiana Lethaea 69, 13–42.
Denton, E.J. 1962: Some recently discovered buoyancy mecha-nisms in marine animals. Proceedings of the Royal Society ofLondon B 265, 366–370.
Denton, E.J. 1974: On buoyancy and the lives of modern and fos-sil cephalopods. Proceedings of the Royal Society of London B185, 273–299.
Denton, E.J. & Gilpin-Brown, J.B. 1966: On the buoyancy of thepearly Nautilus. Journal of the Marine Biological AssociationUK 46, 723–759.
Derham, W. 1726: Philosophical Experiments and Observations ofthe late Eminent Dr. Robert Hooke, Derham, London.
Dunstan, A.J., Ward, P.D. & Marshall, N.J. 2011: Vertical distri-bution and migration patterns of Nautilus pompilius. PLoSONE 6, e16311 1–10.
Garwood, R.J., Rahman, I.A. & Sutton, M.D. 2010: From clergy-men to computers–the advent of virtual palaeontology. Geol-ogy Today 26, 96–100.
Gordon, M. 1962: Species of Goniatites in the Caney Shale ofOklahoma. Journal of Paleontology 36, 355–357.
Greenwald, L. & Ward, P.D. 1982: On the source of cameralliquid in the chambered Nautilus. Veliger 25, 169–170.
Greenwald, L. & Ward, P.D. 1987: Buoyancy in Nautilus. InSaunders, B.W. & Landman, N.H. (eds): Nautilus – The Biol-ogy and Paleobiology of a Living Fossil, 547–560. Springer Ver-lag, Dordrecht.
Greenwald, L., Ward, P.D. & Greenwald, O.E. 1980: Cameralliquid transport and buoyancy control in the chambered nau-tilus (Nautilus macromphalus). Nature 286, 55–56.
Heptonstall, W.B. 1970: Buoyancy control in ammonoids. Le-thaia 3, 317–328.
Hoffmann, R., Schultz, J.A., Schellhorn, R., Rybacki, E., Keupp,H., Gerden, S.R., Lemanis, R. & Zachow, S. 2014: Non-inva-sive imaging methods applied to neo- and paleontologicalcephalopod research. Biogeosciences 11, 2721–2739.
Hoffmann, R. & Zachow, S. 2011: Non-invasive approach toshed new light on the buoyancy business of chamberedcephalopods (Mollusca). IAMG, 506–516. doi: 10.5242/iamg.2011.0163
Hoffmann, R., Lemanis, R., Naglik, C. & Klug, C. 2015: Ammo-noid buoyancy. In Klug, C., Korn, D., De Baets, K., Kruta, I. &Mapes, R.H. (eds): Ammonoid Paleobiology: Volume 1, FromAnatomy to Ecology. Topics in Geobiology 43.
Jacobs, D.K. & Chamberlain, J.A. 1996: Buoyancy and hydrody-namics in ammonoids. In Landman, N., Tanabe, K. & Davis,R.A. (eds): Ammonoid Paleobiology. Topics in Geobiology, 13,169–223. Plenum Press, New York.
Kanie, Y., Fukuda, Y., Nakahara, K., Seki, K. & Hattori, H. 1980:Implosion of living Nautilus under increased pressure. Paleobi-ology 6, 44–47.
Klug, C. 2001: Life-cycles of Emsian and Eifelian ammonoids(Devonian). Lethaia 34, 215–233.
Klug, C. 2002: Quantitative stratigraphy and taxonomy of lateEmsian and Eifelian ammonoids of the Anti-Atlas (Morocco).Courier Forschungsinstitut Senckenberg 238, 1–109.
Klug, C. & Korn, D. 2004: The origin of ammonoid locomotion.Acta Palaeontologica Polonica 49, 235–242.
Korn, D. 2010: A key for the description of Palaeozoic ammo-noids. Fossil Record 13, 5–12.
Kr€oger, B. 2002: On the efficiency of the buoyancy apparatus inammonoids: evidences from sublethal shell injuries. Lethaia35, 61–70.
Kruta, I., Landman, N.H., Rouget, I., Cecca, F. & Tafforeau, P.2011: The role of ammonites in the Mesozoic marine food webrevealed by jaw preservation. Science 331, 70–72.
Kruta, I., Landman, N.H. & Cochran, J.K. 2014: A new approachfor the determination of ammonite and nautilid habitats. PLoSONE 9, e87479 1–7.
LETHAIA 10.1111/let.12125 Palaeozoic ammonoid buoyancy 9

Longridge, L.M., Smith, P.L., Rawlings, G. & Klaptocz, V. 2009:The impact of asymmetries in the elements of the phragmo-cone of Early Jurassic ammonites. Palaeontologia Electronica12, 1–15.
Lukeneder, A. 2012: Computed 3D visualisation of an extinctcephalopod using computer tomographs. Computers and Geo-sciences 45, 68–74.
Lukeneder, A. 2015: Ammonoid habitats and life history. InKlug, C., Korn, D., De Baets, K., Kruta, I. & Mapes, R.H.(eds): Ammonoid Paleobiology, volume 1, From Anatomy toEcology. Springer, Dordrecht.
Lukeneder, S., Lukeneder, A. & Weber, G.W. 2014: Computedreconstruction of spatial ammonoid-shell orientation capturedfrom digitized grinding and landmark data. Computers andGeosciences 64, 104–114.
Mangum, C.P. & Towle, D.W. 1982: The Nautilus siphuncle asan ion pump. Pacific. Science. 36, 273–282.
Meigen, W. 1870: €Uber den hydrostatischen Apparat bei Nautiluspompilius. Archiv f€ur Naturgeschichte 36, 1–36.
Mutvei, H. & Reyment, R. 1973: Buoyancy control and siphunclefunction in ammonites. Palaeontology 6, 623–636.
Naglik, C., Monnet, C., G€otz, S., Kolb, C., De Baets, K. & Klug, C.2015a: Growth trajectories in chamber and septum volumes inmajor subclades of Palaeozoic ammonoids. Lethaia, 48, 29–46.
Naglik, C., Tajika, A., Chamberlain, J. & Klug, C. 2015b: Ammo-noid locomotion. In Klug, C., Korn, D., De Baets, K., Kruta, I.& Mapes, R.H. (eds): Ammonoid Paleobiology, volume 1, FromAnatomy to Ecology. Springer, Dordrecht.
Owen, R. 1832: Memoir of the Pearly Nautilus (Nautilus Pompi-lius, Linn.). 1–68, Royal College of Surgeons, London.
Petter, G. 1959: Goniatites d�evoniennes du Sahara. Publicationsdu Service de la Carte G�eologique de l’Alg�erie, Nouvelle S�erie,Pal�eontologie 2, 1–313.
Raup, D.M. & Chamberlain, J.A. 1967: Equations for volume andcentre of gravity in ammonoid shells. Journal of Paleontology41, 566–574.
Reyment, R.A. 1958: Some factors in the distribution of fossilCephalopods. Acta Universitatis Stockholmiensis — Stock-holm Contribitions in. Geology 1, 97–184.
Reyment, R.A. 1973: Factors in the distribution of fossil cephalo-pods. Part 3: experiments with exact models of certain shelltypes. Bulletin of the Geological Institution of the University ofUppsala New Series. 4, 7–41.
Ritterbush, K., Hoffmann, R., Lukeneder, A. & De Baets, K.2014: Pelagic palaeoecology: the importance of recent con-straints on ammonoid palaeobiology and life history. Journalof Zoology 292, 229–241.
Saunders, W.B. & Shapiro, E.A. 1986: Calculation and simulationof ammonoid hydrostatics. Paleobiology 12, 64–79.
Saunders, W.B. & Work, D.M. 1996: Shell morphology andsuture complexity in upper carboniferous ammonoids. Paleo-biology 22, 189–218.
Saunders, W.B., Work, D.M. & Nikolaeva, S.V. 2004: The evolu-tionary history of shell geometry in Paleozoic ammonoids.Paleobiology 30, 19–43.
Shapiro, E.A. & Saunders, W.B. 1987: Nautilus Shell Hydrostatics.In Saunders, W.B. & Landman, N.H. (eds): Nautilus: The Biol-ogy and Paleobiology of a Living Fossil, 33, 527–545. PlenumPress, New York.
Shigeta, Y. 1993: Post-hatching early life history of CretaceousAmmonoidea. Lethaia 26, 133–146.
Sollas, W.J. 1904: A method for the investigation of fossils byserial section. Philosophical Transactions of the Royal Society ofLondon 196, 259–265.
Sutton, M.D., Briggs, D.E.G., Siveter, D.J. & Siveter, D.J. 2001:Methodologies for the visualization and reconstruction ofthree-dimensional fossils from the Silurian Herefordshire Lag-erst€atte. Palaeontologia Electronica 4, 1–17.
Sutton, M.D., Rahman, I.A. & Garwood, R.J. 2014: Techniquesfor Virtual Palaeontology. Wiley, New York. doi:10.1002/9781118591192.
Swan, A.R.H. & Saunders, W.B. 1987: Function and shape in latePaleozoic (mid-Carboniferous) ammonoids. Paleobiology 13,297–311.
Tajika, A., Naglik, C., Morimoto, N., Pascual-Cebrian, E.,Hennh€ofer, D. & Klug, C. 2014: Empirical 3D-model of theconch of the Middle Jurassic ammonite microconch Nor-mannites: its buoyancy, the physical effects of its maturemodifications and speculations on their function. HistoricalBiology: An International Journal of Paleobiology, 1–11, doi:10.1080/08912963.2013.872097
Trueman, A.E. 1940: The ammonite body chamber, with specialreference to the buoyancy and mode of life of the livingammonite. Quarterly Journal of the Geological Society, London96, 339–383.
Tsujino, Y. & Shigeta, Y. 2012: Biological response to experimen-tal damage of the phragmocone and siphuncle in Nautiluspompilius Linnaeus. Lethaia 45, 443–449.
Vrolik, W. 1843: On the anatomy of the pearly Nautilus. Annalsand Magazine of Natural History 12, 173–175.
Ward, P.D. 1979: Cameral liquid in Nautilus and ammonites.Paleobiology 5, 40–49.
Ward, P.D. 1980: Restructuring the chambered Nautilus. Paleobi-ology 6, 247–249.
Ward, P.D. 1982: The relationship of siphuncle size to emptyingrates in chambered cephalopods: implications for cephalopodpaleobiology. Paleobiology 8, 426–433.
Ward, P.D. 1986: Rates and processes of compensatorybuoyancy change in Nautilus macromphalus. Veliger 28,356–368.
Ward, P.D. 1987: The Natural History of Nautilus. Allen &Unwin, Boston.
Ward, P.D. & Martin, A.W. 1978: On the buoyancy of the PearlyNautilus. Journal of Experimental Zoology 205, 5–12.
Ward, P.D., Greenwald, L. & Magnier, Y. 1981: The chamber for-mation cycle in Nautilus macromphalus. Paleobiology 7, 481–493.
Westermann, G.E.G. 1996: Ammonoid life and habitat, In Land-man, N.H., Tanabe, K. & Davis, R.A. (eds): Ammonoid Paleo-biology. Topics in Geobiology, 13, 607–707. Plenum Press, NewYork.
Westermann, G.E.G. 1998: Life habits of ammonoids. In Savazzi,E. (ed.): Functional Morphology of the Invertebrate Skeleton,263–298. John Wiley & Sons, Chichester.
Westermann, G.E.G. 2013: Hydrostatics, propulsion and life-habits of the Cretaceous ammonoid Baculites. Revue dePal�eobiologie 32, 249–265.
Willey, A. 1902: Contributions to the natural history of thepearly Nautilus : Zoological results based on material from NewBritain, New Guinea, Loyalty Islands and elsewhere, collectedduring the years 1895, 1896 and 1897, volume 6, UniversityPress, Cambridge, England. 691–830.
10 Naglik et al. LETHAIA 10.1111/let.12125