a review of the early palaeozoic biogeography of bryozoans
TRANSCRIPT

Chapter 12
A review of the Early Palaeozoic biogeography of bryozoans
CAROLINE J. BUTTLER1*, PATRICK N. WYSE JACKSON2, ANDREJ ERNST3 & FRANK K. MCKINNEY4†
1Department of Geology, National Museum of Wales, Cathays Park, Cardiff CF10 3NP, Wales, UK2Department of Geology, Trinity College, Dublin 2, Ireland
3Institut fur Geowissenschaften der Christian-Albrechts-Universitat zu Kiel, Ludewig-Meyn-Str. 10, D-24118 Kiel, Germany4Formerly at Department of Geology, Appalachian State University, Boone, NC 28608, USA
†Deceased
*Corresponding author (e-mail: [email protected])
Abstract: The palaeogeographical distributions of Early Palaeozoic bryozoan faunas are reviewed. Previous studies are examined andnew databases have been assembled of the stratigraphical and geographical distribution of Ordovician and Silurian taxa. Analysis wascarried out using cluster analysis based on Jaccard’s coefficient and paired group method, as well as principal coordinate analysis basedon Jaccard’s coefficient, to examine the relationships between different localities. Bryozoan faunas increased in diversity throughout theOrdovician peaking with 133 genera during the Katian. In the earliest Ordovician provincialism is difficult to determine, but by the Dar-riwilian five distinct provinces developed, decreasing to four in the Sandbian. There was a decrease in provinciality throughout the Katianas faunas became less endemic, caused by the reduction of geographical barriers. Following the extinction of many genera at the end ofthe Ordovician, early Silurian faunas contain remnant taxa. Subsequently fenestrates began to dominate faunas. During the Llandoverybryozoans began to show distinct provincialism, but this declined during the Wenlock, only to re-emerge during the Ludlow. Late Silur-ian (Pridoli) faunas are sparse but nevertheless show possible division into two provinces.
There have been a number of studies examining the biogeographi-cal distribution of Early Palaeozoic bryozoans. Some have inves-tigated particular areas, for example, the Ordovician RussianArctic (Nekhorosheva 1976), North America (Anstey 1974,1986) and Mongolia (Kopaevich 1984; Ariunchimeg 2009a, b),and others have examined the worldwide generic distribution ofbryozoan faunas throughout the Ordovician (Ross 1985; Xiaet al. 2010) or for part of this period (Anstey 1986; Jimenez-Sanchez & Villas 2009). Tuckey (1990a, b) has described indetail the distributions of Ordovician and Silurian faunas andrecently McCoy & Anstey (2010) examined the biogeographicalassociations of Silurian genera in North America, Baltica andSiberia. Gorjunova et al. (2004) provided a synthesis of biogra-phical distributions and patterns for the whole of the Palaeozoic.
Our knowledge of Early Palaeozoic bryozoans is patchy; untilrelatively recently the majority of research had been undertakenby North American and Russian workers. There are still areas ofthe globe where bryozoans of this age are abundant, but our knowl-edge is incomplete. Palaeobiogeographical models are only asgood as the available data and will be refined as new faunasare described.
This paper is a review of Early Palaeozoic bryozoan faunas.New databases have also been assembled of the stratigraphical andgeographical distributions of Ordovician and Silurian bryozoangenera. The compilation of the databases and the analysis of thepalaeobiogeographical relations of the bryozoan faunas beloware based on regional taxonomical papers. Full citation of thesepublications (more than 200) is beyond of the scope of the presentpaper. The compiled sources are assigned in general to the follow-ing areas: North Europe (Estonia, NW Russia, Sweden, Norway,Great Britain, Greenland), southern Europe (France, Italy, Spain,Portugal), Belarus, North Africa, North America (USA, Canada),Siberia (Arctic Urals, Novaya Zemlya, Taymyr, Vaigach), Mongo-lia, China, Australia, Kazakhstan and Argentina.
The databases were converted to presence/absence data matri-ces, which were used for cluster and principal coordinate analy-sis, in order to show similarities between localities. The cluster
analysis has been performed measuring the Jaccard similaritycoefficient, using the Unweighted Pair Group Algorithm withArithmetic Mean (PAST version 1.81, Hammer et al. 2001). Theprincipal coordinate analysis was performed using Jaccard’s coef-ficient for measurement of similarities. Jaccard’s coefficient is asimilarity index for binary data, measuring similarity betweensample sets (Jaccard 1901a, b). This method has proven to be arobust and quite reliable statistic procedure for calculating faunalresemblance between regions (e.g. Legendre & Legendre 1998;Glasby & Alvarez 1999; Murguıa & Villasenor 2003; Schmach-tenberg 2008).
The stratigraphical framework used herein is the InternationalStratigraphic Chart (version of September 2010) published bythe International Commission on Stratigraphy (2010). Older pub-lished distributions (Tuckey 1990a; Tuckey & Anstey 1992) andrecords derived from taxonomic works have been examined andolder stage assignations for the Ordovician correlated using datain Ogg et al. (2008). In contrast to Tuckey (1990a, b), who didnot include in his analyses localities with small datasets (wherefewer than five taxa were recorded), we do include these data here.
Ordovician
A supposed cryptostome bryozoan has been recorded from theUpper Cambrian of southern Mexico (Landing et al. 2010) butthis has now been re-identified as an octocoral (Taylor et al. inpress). The oldest known unequivocal bryozoans are of Late Tre-madoc age from China (Hu & Spjeldnæs 1991; Xia et al. 2007;Zhang et al. 2009; Cuffey et al. 2012) and Severnaya andNovaya Zemlya (Astrova 1965; Ernst et al. 2013). They are notknown from any other location at this time. Bryozoans wererecorded throughout the Floian-Dapingian; the oldest is a singletrepostome specimen from the Floian of Wales (Taylor & Cope1987). Early bryozoan faunas have also been described fromRussia, Estonia, Poland, North America and Ireland. All fiveorders of bryozoans are recognized (Taylor & Ernst 2004).
From: Harper, D. A. T. & Servais, T. (eds) 2013. Early Palaeozoic Biogeography and Palaeogeography.
Geological Society, London, Memoirs, 38, 145–155. http://dx.doi.org/10.1144/M38.12
# The Geological Society of London 2013. Publishing disclaimer: www.geolsoc.org.uk/pub_ethics

Tuckey (1990a) examined the worldwide patterns of Ordovi-cian bryozoans using multivariate statistical techniques to deter-mine biogeographical associations. In the Floian–Dapingian herecognized a North American and a Baltic Province and consideredthat China had similarities to North America.
During the Darriwilian an increase in diversity and provincialitywas recognized by Tuckey (1990a). He described three provinces:the Baltic Province, subdivided into the Baltic Platform and theUralian Geosyncline biomes, the North American Province,which included the Chazyan reef biome and Appalachian–Oua-chita Geosyncline, and the Siberian Province. Tuckey (1990a)recognized the same provincial patterns in the late Darriwilian,with a blurring of provincial boundaries and increasing differ-entiation of North American faunas. He also identified a fourthMediterranean province represented by Moroccan faunas. Spjeld-næs (1981) suggested a similarity to faunas from southern Europe,and that there was a climatic rather than a wide oceanic barrierbetween the Mediterranean and Baltic Provinces. Spjeldnæs(1981) also identified this time as when the first major faunalexchange occurred across the Iapetus Ocean.
Tuckey (1990a) identified the same four provinces in the Sand-bian. The Mediterranean Province at this time included faunasfrom Bohemia, Sardinia and the Carnic Alps. He included faunasfrom Burma within the Baltic Province; this was consistent withthe findings of Williams (1973), who included it on the basis ofthe brachiopod faunas. In the upper Katian Tuckey recognizedthat faunas were becoming more cosmopolitan and provincialitydecreasing with only two identifiable provinces: the NorthAmerican–Siberian Province and the Baltic–MediterraneanProvince.
Taylor & Ernst (2004) examined the distribution of the eightmajor stenolaemate clades within the four Ordovician provinces(North American, Baltic, Siberian and Mediterranean) describedby Tuckey (1990a). They recognized that halloporine trepostomeswere the most species-rich group in all provinces with the excep-tion of Siberia, where ptilodictyine cryptostomes dominated.Esthonioporines were common in the Baltic Province but less soelsewhere, especially in North America. This reflected the agesof the bryozoan faunas present, the Baltic Province includingolder bryozoan faunas in which the more primitive esthonioporinestrepostomes are abundant.
Jimenez-Sanchez & Villas (2009) performed a quantitativestudy of the distribution of the late Katian bryozoans, with afocus primarily on Mediterranean bryozoans, comparing their dis-tribution with those from other Gondwanan terranes and the restof the Late Ordovician palaeocontinents. This study showed thatmigrations of bryozoans were strongly influenced by climaticfactors and transgression/regression events. The decrease in ende-mism was a result of the global warming event (Boda event), whichcaused an intermixture of tropical and temperate faunas.
In the Late Ordovician three distinct extinction events have beenidentified (Tuckey & Anstey 1992): (1) early Katian (Onnian),which was concentrated on the continents of Baltica and Siberia;(2) late Katian (Rawtheyan), which was most effective on theNorth American Plate; and (3) Hirnantian, which was most pro-nounced in Baltica.
Tremadocian
The earliest unquestionable bryozoans are known from the Trema-docian of China and include Nekhorosheviella, Orbiramusand Prophyllodictya (Xia et al. 2007; Zhang et al. 2009; Adachiet al. 2012). Hu & Spjeldnæs (1991) mentioned two trepostomegenera, Hubeipora and Yichangpora from the Tremadocian ofChina. However, these genera were never formally described.The cystoporate Profistulipora was recorded from Siberia(Astrova 1965).
Floian
The bryozoan record from the Floian is quite sparse. Five generaare known from NW Russia and Estonia, all belonging to eitherthe order Trepostomata (Hemiphragma, Phragmopora, Estonio-pora and Dittopora; Pushkin & Popov 1999) or the order Cystopor-ata (Revalotrypa and Lynnopora; Gorjunova & Koromyslova2008). Elsewhere the trepostome Orbipora was recorded fromWales (Taylor & Cope 1987), and the cryptostome Profistuliporafrom Siberia (Astrova 1965), while the trepostome Nicholsonellawas mentioned by Carrera (1995) and Carrera & Ernst (2010)from the Floian part of the San Juan Formation of Argentina.
Dapingian
The Dapingian saw an increase in the diversity and distribution ofbryozoan taxa. In Baltica the first known fenestrate Alwynopora(Taylor & Curry 1985) and cyclostome Goryunovia (Taylor &Rozhnov 1996) appeared, as well as the cryptostome Prophyllodic-tya, cystoporate Revalotrypa and six trepostome genera: Ana-phragma, Cyphotrypa, Dianulites, Dittopora, Estoniopora andOrbipora. Five genera are known from the Kanosh Formation ofUtah, USA (Ernst et al. 2007): Orbipora, Kanoshipora, Ibexella,Eridotrypa and ?Diplotrypa. The trepostome Amplexopora isknown from Australia (Ross 1961), and one cryptostome fromSiberia: Arthrophragma (Astrova 1965).
Darriwilian
During the Darriwilian 36 bryozoan genera are identified, dis-tributed in five provinces. Twenty-one bryozoan genera havebeen recorded from Baltica, seven genera are known from NorthAmerica (USA and Canada), 19 from Belarus, eight fromSiberia, and a single genus (Revalotrypa) from China (Yang 1957).
According to the cluster analysis of the bryozoan generic distri-bution (Fig. 12.1), Belarus and Baltica show the closest relation-ship, sharing 10 genera. Siberia is close to the cluster Baltica–Belarus. The North American province is separate from bothBaltica–Belarus and Siberia, sharing three genera with Balticaand only one genus with Belarus and Siberia. China shares thegenus Revalotrypa with Baltica and Siberia.
Ordovician bryozoans of Argentina are quite diverse and abun-dant, but have only recently been studied. The trepostome bryo-zoans Lamottopora and Aostipora as well as a problematicphylloporinid genus were described from the Dariwillian part ofthe San Juan Formation of Argentina (Carrera & Ernst 2010).These findings suggest a close relationship of the Argentineanbryozoan faunas to those of North America and Baltica.
Sandbian
In the Sandbian four provinces can be recognized in which 105bryozoan genera are known. The Baltic Province contains 62genera, North America 55, Belarus 33 and Siberia 23.
The closest relationship existed between Baltica and NorthAmerican provinces (Fig. 12.2). These provinces share 28bryozoan genera. Belarus has 12 genera in common withBaltica/North America. Siberia has a different generic compo-sition; about half of Siberian genera are endemic. However, itshares 11 genera with North America, eight with Baltica, andfive with Belarus. Carrera (2003) reported the trepostome Praso-pora from the Sandbian of Argentina, and it is also known fromBaltica, Belarus and North America.
C. J. BUTTLER ET AL.146

Katian
The Katian shows the highest bryozoan diversity of the LowerPalaeozoic, with 133 genera known from this stage world-wide. Bryozoans are reported from a greater number of areasand the relationships between these are very complex. For thepurposes of cluster analysis 11 geographical regions are defined(Fig. 12.3). North America contains the greatest number ofgenera (79), followed by Baltica (68). From Belarus 29 bryozoangenera are recorded. France, Italy and Spain, in the Mediterraneanregion, contain 38, 13 and 18 genera, respectively. Thirty-sixgenera are known from Siberia, 20 from India, 16 from Mongolia,five from China and six from Australia. Bryozoan faunas fromNorth Africa (Morocco, Libya and Algeria) seem to be very rich,but only eight genera have been reported from this region(Buttler & Massa 1996; Becq-Giraudon & Massa 1997; Buttleret al. 2007).
Baltica and North America show the closest similarity, sharing40 bryozoan genera. Belarus has 21 genera in common withBaltica and 17 with North America. These three localities share12 genera. This cluster is close to the Mediterranean cluster verydistinctly produced by France and Italy. Bryozoan faunas fromFrance and Italy share 12 genera and they cluster together withBelarus, with which they share seven genera.
Spain produces a distinct cluster with India, with which it sharessix genera. It shares five genera with the France–Italy cluster and
four genera with Belarus, and in total three genera with theFrance–Italy–Belarus cluster.
France, Italy, Spain, India and North America share four genera:Kukersella, Ceramopora, Eridotrypa and Heterotrypa. The twolatter genera occur also in Belarus.
Mongolia shows diverse relationships; it shares genera withSiberia (eight) as well as with North America (11 genera),Baltica (seven genera) and Belarus (six genera). The relationshipswith Mediterranean faunas are quite weak. Mongolia shares twogenera with France and Italy, and three genera with Spain. Further-more, Mongolia shares two genera with India and four genera withAustralia. Only one genus (Homotrypa) is common to Mongoliaand China.
Australian bryozoan faunas show relationships with Baltica(four genera), Mongolia (four genera) and North America (threegenera) as well as Siberia and Belarus (both two genera). Siberiashares 21 genera with North America, suggesting a close relation-ship. Despite the similarity between Baltica and North America,Siberia contains only seven genera, which occur in both ofthese provinces.
Four genera from China are also known from North America andSiberia. The trepostome genus Prasopora occurs in China, NorthAmerica, Siberia and Baltica. Two species of Prasopora werealso described from the Katian of Kazakhstan (Pushkin & Popov1991). In general, information on Ordovician bryozoans fromKazakhstan is very restricted. Five species, belonging to four
Baltica
N. America
Siberia
Belarus
-3 -2.4 -1.8 -1.2 -0.6 0 0.6 1.2 1.8 2.4Coordinate 1
-3
-2.4
-1.8
-1.2
-0.6
0
0.6
1.2
1.8
2.4
Coo
rdin
ate
2
N. America
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Similarity
Belarus
Baltica
Siberia
Fig. 12.2. Cluster diagram and principal coordinate analysis showing
similarities between bryozoan faunas of different areas during the Sandbian (see
text for explanations).
Baltica N. America
Siberia
Belarus
-3 -2.4 -1.8 -1.2 -0.6 0 0.6 1.2 1.8 2.4Coordinate 1
-3
-2.4
-1.8
-1.2
-0.6
0
0.6
1.2
1.8
2.4
Coo
rdin
ate
2
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Similarity
Siberia
Belarus
Baltica
N. America
Fig. 12.1. Cluster diagram and principal coordinate analysis showing
similarities between bryozoan faunas of different areas during the Dariwillian
(see text for explanations).
EARLY PALAEOZOIC BIOGEOGRAPHY OF BRYOZOANS 147

genera (cystoporate Revalopora and three trepostomes, Nicholso-nella, Batostoma and Prasopora) are formally described (Pushkin& Popov 1991).The record of the species Nicholsonella cf. poly-morpha (Kopaevich 1984) suggests a relationship of Kazakhstanwith Mongolia, whereas the cystoporate Revalopora shows alink with the Baltic Province. Batostoma is, like Prasopora,widely distributed during the Katian, occurring in Baltica, theMediterranean (France and Italy), North America, Siberia andAustralia.
The bryozoan faunas from northern Africa are rich but poorlyinvestigated. Eight genera are recorded from this region, six ofwhich also occur in France, Italy, the Baltic and North America.One genus (Jifarahpora) is endemic (to Libya), and another (Poly-teicus) is also known from Bohemia.
Ross et al. (2000) described a bryozoan fauna from Iran andidentified six taxa from which two genera (Hallopora and Erido-trypa) are distinct and show a cosmopolitan distribution duringthe Katian. In general, bryozoans of this region are poorly known.
China, India, North America as well as the Mediterraneancluster (France and Italy) share the trepostome genus Dekayia.This genus was not reported from the Katian of Baltica untilnow, but it is certainly present there (A. Ernst, pers. obs. 2003).
Ernst & Carrera (2008) recorded Moyerella and Phylloporinafrom the Katian Sassito Formation of Argentina. Moyerella indi-cates connections to Siberia and Spain.
Our data indicate that, in the Sandbian and at the beginning ofthe Katian, provincialism was high and this resulted in the highlevels of bryozoan diversity that are recognized. However, there
0 0.12 0.24 0.36 0.48 0.6 0.72 0.84 0.96
Similarity
N. Africa
China
Mongolia
India
Spain
Siberia
N. America
Baltica
Belarus
France
Australia
Italy
Baltica
FranceItaly N. America
SiberiaBelarus
China
Mongolia
India
Australia
Spain
N. Africa
-0.8 -0.64 -0.48 -0.32 -0.16 0 0.16 0.32 0.48 0.64
Coordinate 1
-0.8
-0.64
-0.48
-0.32
-0.16
0
0.16
0.32
0.48
0.64
Coo
rdin
ate
2
Fig. 12.3. Cluster diagram and principal
coordinate analysis showing similarities
between bryozoan faunas of different areas
during the Katian (see text for explanations).
C. J. BUTTLER ET AL.148

was a decrease in provincialism along with diversity during theKatian owing to the reduction of geographical barriers. The dis-tributional patterns of bryozoans at this time are complex andthe designation of individual provinces is open to differinginterpretations. One might argue that, as the Baltica and NorthAmerica provinces share 40 genera, this represents a cosmopolitanstate. Alternatively these provinces could be interpreted as beingdistinctive given the large number of endemic genera confined toone province and not occurring in the other.
Hirnantian
The Hirnantian shows a greatly reduced diversity, only 37 generabeing known from rocks of this age. The bryozoan record allowsrecognition of four provinces (Fig. 12.4). The most diversebryozoan fauna occurred in Baltica (25 genera), followed by theNorth American Province (18). Six genera are known fromBelarus, four genera from Mongolia and three from Siberia.
Baltica and North America cluster together, sharing nine genera.Belarus is close to this cluster, sharing with them four genera, andis closer to North America, with which it shares one more genusthan with Baltica. Siberia shares no taxa with these three pro-vinces. In contrast Mongolia shares Metadictya and Phaenoporawith the cluster Baltica–North America.
Silurian
When compared with the volume of information published onOrdovician bryozoans, that available on the distribution anddiversity of Silurian bryozoans is considerably poorer. There arefewer papers published on Silurian faunas, most likely owing tothe rarity of bryozoans in Silurian successions. Brood (1989) attri-butes this to the fact that many early Silurian successions inWestern Europe and the Baltic regions are dominated by sand-stones and shales which are not conducive to the preservationof bryozoans.
BalticaN. America
SiberiaBelarus
Mongolia
-2 -1.6 -1.2 -0.8 -0.4 0 0.4 0.8 1.2 1.6
Coordinate 1
-2
-1.6
-1.2
-0.8
-0.4
0
0.4
0.8
1.2
1.6
Coo
rdin
ate
2
Siberia
Belarus
Baltica
N. America
Similarity
Mongolia
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Fig. 12.4. Cluster diagram and principal
coordinate analysis showing similarities
between bryozoan faunas of different areas
during the Hirnantian (see text for
explanations).
EARLY PALAEOZOIC BIOGEOGRAPHY OF BRYOZOANS 149

At the end of the Ordovician many genera disappeared butnevertheless the bryozoan faunas and taxa that remained weresimilar to those of the late Ordovician, and as Brood (1989,p. 231) has noted, represented ‘an impoverished upper Ordovicianassemblage’. The fenestrate bryozoans were poorly represented inthe Ordovician but rapidly diversified during the early Silurianwhen representatives of the Family Fenestellidae began to domi-nate, and the Phylloporinidae declined.
Llandovery
The classic papers documenting Llandovery bryozoans are thosewhich have reported on faunas from Estonia (Kopaevich 1975),
Gotland (Hennig 1905) and Anticosti Island (Bassler 1928).However, faunas of this age are poorly known and requirereassessment.
The Silurian saw the closing of the Iapetus Ocean, which ceasedto be a barrier to brachiopod and trilobite larvae (Cocks & Fortey1982). Cocks (2001) identified the early Silurian as a period ofexceptional cosmopolitanism for benthic larvae such as brachio-pods and trilobites.
Tuckey (1990b), using the multivariate statistical techniquesof gradient and cluster analysis, distinguished three differentbryozoan faunal provinces in the Llandovery: Baltica, NorthAmerican–Siberian and Mongolian provinces, which was alsopresent in Siberia. McCoy & Anstey (2010) re-examined the
Similarity
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Kazakhstan
Mongolia
Siberia
China
N. America
Baltica
Baltica
N. America
Siberia
China
Mongolia
Kazakhstan
-2 -1.6 -1.2 -0.8 -0.4 0 0.4 0.8 1.2 1.6
Coordinate 1
-2
-1.6
-1.2
-0.8
-0.4
0
0.4
0.8
1.2
1.6
Coo
rdin
ate
2
Fig. 12.5. Cluster diagram and principal
coordinate analysis showing similarities
between bryozoan faunas of different areas
during the Llandovery (see text for
explanations).
C. J. BUTTLER ET AL.150

palaeogeography of Silurian bryozoans by incorporating addi-tional qualitative techniques including parsimony analysis ofendemicity. They also identified three palaeogeographicalregions in the Llandovery: Mongolian Province present inSiberia, Brassfield Subprovince in North America with one localarea in Siberia, and the Visby Subprovince, predominantly inBaltica.
In contrast to previous interpretations, our analysis (Fig. 12.5)shows the Mongolian fauna as a separate entity to those ofSiberia, which plots closer to those of China. North America andBaltica form a distinct province.
Wenlock
Wenlock faunas are well documented for certain regions:England (Snell 2004), Gotland (Hennig 1905, 1906), Tuva(Astrova 1957) and the USA (Hall 1883; Bassler 1906). In theWenlock Tuckey (1990b) considered that the Baltic and NorthAmerican–Siberian Province had merged, and included thefauna of Kazakhstan. He also identified the Mongolian Province,which comprised faunas from northwestern and eastern Mongo-lia, Tuva and central China. However, McCoy & Anstey (2010)recognized four subprovinces in the Wenlock: the Mongolian
0 0.12 0.24 0.36 0.48 0.6 0.72 0.84 0.96Similarity
Siberia
Australia
Mongolia
China
N. America
Baltica
Baltica
N. America
Siberia
China
Mongolia
Australia
-2 -1.6 -1.2 -0.8 -0.4 0 0.4 0.8 1.2 1.6
Coordinate 1
-2
-1.6
-1.2
-0.8
-0.4
0
0.4
0.8
1.2
1.6
Coo
rdin
ate
2
Fig. 12.6. Cluster diagram and principal
coordinate analysis showing similarities
between bryozoan faunas of different areas
during the Wenlock (see text for
explanations).
EARLY PALAEOZOIC BIOGEOGRAPHY OF BRYOZOANS 151

Province in Siberia, the Visby Subprovince (dominant in Baltica),the Wenlock Subprovince in SW Baltica and the Rochester Sub-province in North America.
From our analysis (Fig. 12.6) we note the similarities ofthe Siberian and Australian faunas, and the distinctive NorthAmerican and Baltic assemblages which constrain the two pro-vinces. The Mongolian and Chinese faunas appear somewhatendemic.
Ludlow
Ludlovian bryozoans are well documented from the Czech Repub-lic (Pocta 1894), and Estonia (Kopaevich 1975) and have beenreported from China, Russia, Kazakhstan and Wales. By the Lud-low the northern Iapetus had completely closed (Cocks & McKer-row 1973). Tuckey (1990b) described the Ludlow as a time ofcosmopolitanism for bryozoans. The Mongolian Province had
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
Similarity
Australia
Kazakhstan
Mongolia
N. America
Siberia
China
Baltica
Baltica
N. America
Siberia
China
Mongolia
Australia
Kazakhstan
-2 -1.6 -1.2 -0.8 -0.4 0 0.4 0.8 1.2 1.6Coordinate 1
-2
-1.6
-1.2
-0.8
-0.4
0
0.4
0.8
1.2
1.6
Coo
rdin
ate
2
Fig. 12.7. Cluster diagram and principal
coordinate analysis showing similarities
between bryozoan faunas of different areas
during the Lludlovian (see text for
explanations).
C. J. BUTTLER ET AL.152
disappeared and the faunas from Mongolia and Tuva showedsimilarities to European and American faunas. He identified dis-tinctive faunas at single localities rather than provinciality. Thiscosmopolitanism continued to the Pridoli and little biogeo-graphical differentiation in a Baltic–North American–SiberianProvince could be identified. As in the Wenlock, some distinc-tive faunas could be identified, which Tuckey identified as commu-nities within the large cosmopolitan province. These were theHamra Formation from Gotland and the Taugantelyski Formationof Tuva, which has a low diversity and genera that are morecommon in the Ordovician.
McCoy & Anstey (2010) examined the Ludlow and Pridolitogether and interpreted the data to represent four subprovincesrather than one large merged province. They indentified three pro-vinces that remained similar to those of the Wenlock: the MongoliaProvince in Siberia, the Wenlock Subprovince in Avalonia/Balticaand the Rochester Subprovince in North America. The KeyserSubprovince was also identified which replaced the Visby Subpro-vince in Baltica and the Rochester Subprovince in southeasternNorth America.
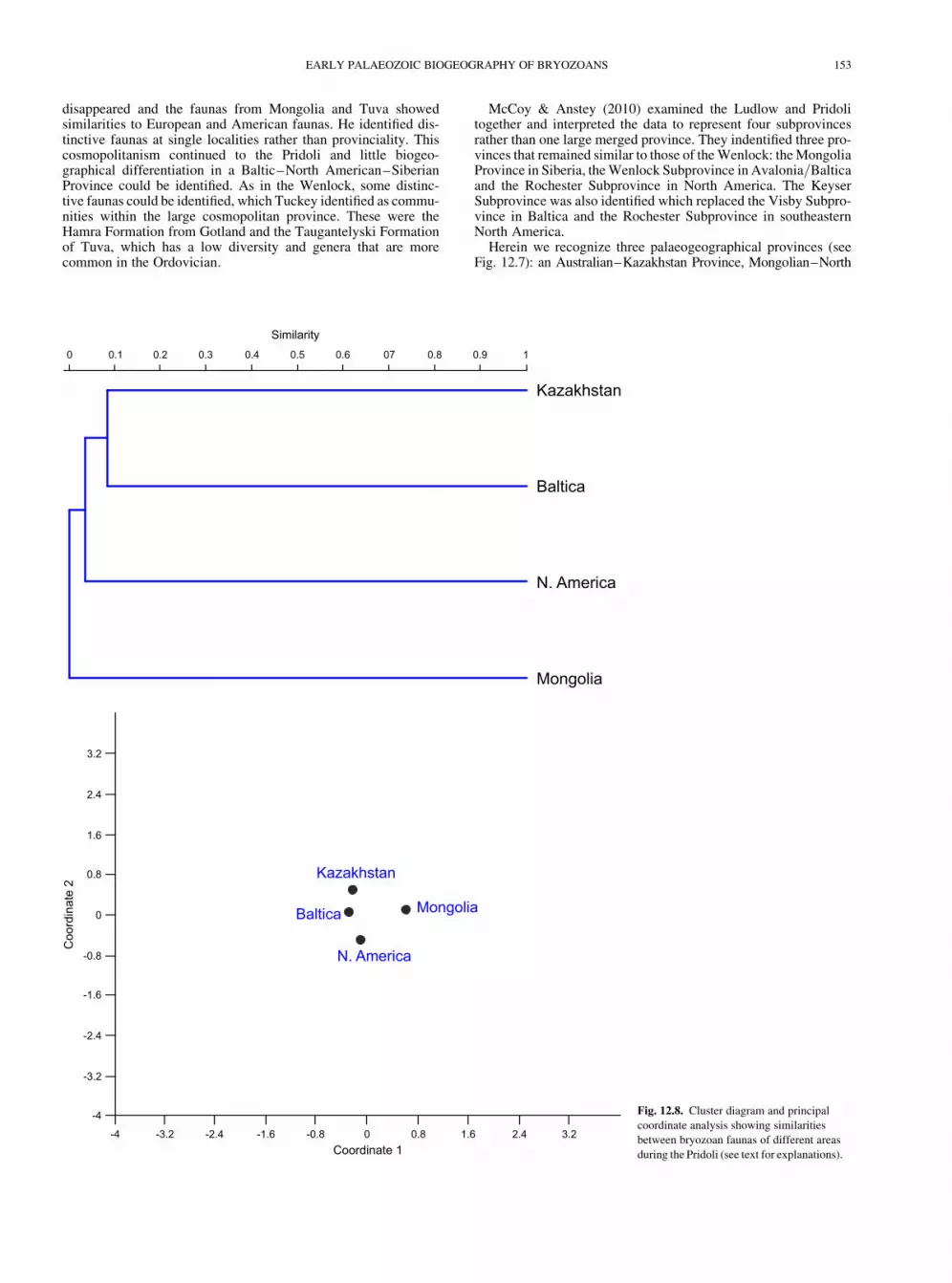
Herein we recognize three palaeogeographical provinces (seeFig. 12.7): an Australian–Kazakhstan Province, Mongolian–North
0 0.1 0.2 0.3 0.4 0.5 0.6 07 0.8 0.9 1
Similarity
Kazakhstan
Baltica
N. America
Mongolia
Baltica
N. America
Mongolia
Kazakhstan
-4 -3.2 -2.4 -1.6 -0.8 0 0.8 1.6 2.4 3.2Coordinate 1
-4
-3.2
-2.4
-1.6
-0.8
0
0.8
1.6
2.4
3.2
Coo
rdin
ate
2
Fig. 12.8. Cluster diagram and principal
coordinate analysis showing similarities
between bryozoan faunas of different areas
during the Pridoli (see text for explanations).
EARLY PALAEOZOIC BIOGEOGRAPHY OF BRYOZOANS 153

American Province and a Baltic–Siberian–China Province. Thedistribution of bryozoans is probably strongly controlled by latitu-dinal factors.
Pridoli
Pridoli faunas are poorly represented with records being confinedto a small number of locations, including the Czech Republic(Pocta 1894), Podolia (Astrova 1970), Turkestan (Pushkin et al.1990), and Belarus and Latvia (Pushkin 1975). Tuckey (1990b)noted that the bryozoan faunas remained cosmopolitan into thePridoli. Our analysis, however (Fig. 12.8), suggests moderatelyclose links between the Baltic and Kazakhstan faunas and moredistant similarities with the North American faunas.
Conclusions
Analysis of bryozoan faunas from throughout the Lower Palaeo-zoic show distinct changes in provincialism and diversity.Generic diversity increases throughout the Ordovician and isgreatly reduced after the Ordovician extinction events. Provincial-ism is shown to decrease in the Upper Ordovician (Katian) andfaunas become less endemic. Following the Late Ordovicianextinctions bryozoan diversity is greatly reduced, a patternwhich is continued in the Silurian. Separate studies have produceddiffering interpretations as to the exact relationship of LowerPalaeozoic bryozoan geographical distributions. There are stillregions known to contain rich bryozoan faunas that have beenpoorly investigated, for example North Africa, Kazakhstan, Aus-tralia and Argentina. With greater knowledge of these areaspalaeogeographical analysis will become more refined.
We would like to thank P. Taylor (Natural History Museum, London, UK) and
H. A. Nakram (University of Oslo, Natural History Museum, Norway) for their
helpful reviews of this paper. A. E. thanks the German Scientific Foundation
(DFG) for funding the research project ER 278/2-1. u. 2-2 ‘Radiation of Ordovi-
cian bryozoans’ (2000–2002).
References
Adachi, N., Ezaki, Y. & Liu, J. 2012. The oldest bryozoan reefs: a uniqueEarly Ordovician skeletal framework construction. Lethaia, 45,14–23, http://dx.doi.org/10.1111/j.1502-3931.2011.00268.x
Anstey, R. L. 1974. Geographic diversity of Late Ordovician corals andbryozoans in North America. Journal of Paleontology, 48,1141–1148.
Anstey, R. L. 1986. Bryozoan provinces and patterns of generic evolutionand extinction in the Late Ordovician of North America. Lethaia, 19,33–51.
Ariunchimeg, Y. 2009a. New Late Ordovician bryozoans from theTsagaan–Del Section of Central Mongolia. PaleontologicalJournal, 43, 401–407.
Ariunchimeg, Y. 2009b. Stratigraphy and paleogeographic distributionof the Upper Ordovician bryozoans of Mongolia. PaleontologicalJournal, 43, 1432–1438.
Astrova, G. G. 1957. Some new bryozoan species from the Silurianof Tuva. Materiali k osnovam paleontologhii Akad Nauk SSSR, 1,5–14 [in Russian].
Astrova, G. G. 1965. Morphology, history of development, and systema-tics of Ordovician and Silurian Bryozoa. Trudy Paleontologiches-kogo Instituta Akademii Nauk SSSR, 106, 1–432 [in Russian].
Astrova, G. G. 1970. New Silurian and Early Devonian Bryozoa, Cysto-porata and Trepostomata in Estonia and Podolia. In: Astrova, G. G.& Chudinova, I. I. (eds) New Species of Paleozoic Bryozoa andCorals. Nauk, Moscow, 7–22 [in Russian].
Bassler, R. S. 1906. The bryozoan fauna of the Rochester Shale. UnitedStates Geological Survey Bulletin, 292, 1–65.
Bassler, R. S. 1928. Bryozoa. In: Twenhofel, W. H. (ed.) Geology ofAnticosti Island. Geological Survey of Canada, Ottawa, Memoirs,154, 143–168.
Becq-Giraudon, J. F. & Massa, D. 1997. Dreyfussina libyca nov. sp.(Trilobita, Phacopacea) de la formation Melez-Chograne, Caradocsuperieur de Libye. Geobios, 30 (suppl. 1), 49–54.
Brood, K. 1989. Bryozoans. In: Holland, C. H. & Basset, M. G. (eds) AGlobal Standard for the Silurian System. National Museum of Wales,Cardiff, Geological Series, 9, 231–232.
Buttler, C. & Massa, D. 1996. Late Ordovician bryozoans from carbon-ate buildups, Tripolitania, Libya. In: Gordon, D. P., Smith, A. M. &Grant-Mackie, J. A. (eds) Bryozoans in Space and Time. NIWA,Wellington, 63–68.
Buttler, C., Cherns, L. & Massa, D. 2007. Bryozoan mud-moundsfrom the Upper Ordovician Jifarah (Djeffara) Formation of Tripolita-nia, north-west Libya. Palaeontology, 50, 479–494.
Carrera, M. G. 1995. El genero Nicholsonella Ulrich (Bryozoa) en elOrdovicio de la Precordillera Argentina. Su significando paleoecolo-gico y paleogeografico. Ameghiniana, 32, 181–190.
Carrera, M. G. 2003. The genus Prasopora (Bryozoa) from the MiddleOrdovician of the Argentine Precordillera. Ameghiniana, 40,197–203.
Carrera, M. & Ernst, A. 2010. Darriwilian bryozoans from the San JuanFormation (Ordovician), Argentine Precordillera. Ameghiniana, 47,343–354.
Cocks, L. R. M. 2001. Ordovician and Silurian global geography. Journalof the Geological Society, 158, 197–210.
Cocks, L. R. M. & Fortey, R. A. 1982. Faunal evidence for oceanic sep-arations in the Palaeozoic of Britain. Journal of the GeologicalSociety, 139, 465–478.
Cocks, L. R. M. & McKerrow, W. S. 1973. Brachiopod distributions andfaunal provinces in the Silurian and Lower Devonian. In: Hughes,N. R. (ed.) Organisms and Continents through Time. PalaeontologicalAssociation, London, Special Papers in Palaeontology, 12, 291–304.
Cuffey, R. J., Xiao, C., Zhu, Z., Spjeldnaes, N. & Hu, Z.-X. 2012.The world’s oldest-known Bryozoan reefs: late Tremadocian, mid-Early Ordovician; Yichang, central China. In: Ernst, A., Schafer,P. & Scholz, J. (eds) Bryozoan Studies 2010. Springer, Heidelberg,13–27.
Ernst, A. & Carrera, M. 2008. Cryptostomid bryozoans from the SassitoFormation, Upper Ordovician cool-water carbonates of the Argenti-nean Precordillera. Palaeontology, 51, 1117–1127.
Ernst, A., Taylor, P. D. & Wilson, M. A. 2007. Ordovician bryozoansfrom the Kanosh Formation (Whiterockian) of Utah, USA. Journal ofPalaeontology, 81, 998–1008.
Ernst, A., Bogolepova, O. K., Hubmann, B., Golubkova, E. Y. &Gubanov, A. P. 2013. Dianulites (Trepostomata, Bryozoa) fromthe Early Ordovician of Severnaya Zemlya, Arctic Russia. Geologi-cal Magazine, FirstView Article, 1–11.
Glasby, C. J. & Alvarez, B. 1999. Distribution patterns and biogeo-graphic analysis of Austral Polychaeta (Annelida). Journal of Bio-geography, 26, 507–533.
Gorjunova, R. V. & Koromyslova, A. V. 2008. A new genus of the Are-nigian bryozoans, Lynnopora, and its systematic position in theFamily Revalotrypidae Gorjunova, 1988. Paleontological Journal,42, 491–499.
Gorjunova, R. V., Markov, A. V. & Naimark, E. B. 2004. Evolutionand Biogeography of Paleozoic Bryozoans: The Results of Quantitat-ive Analysis. GEOS, Moscow [in Russian].
Hall, J. 1883. Bryozoans of the Upper Helderberg and Hamilton Groups.Transactions of the Albany Institute, 10, 145–197.
Hammer, Ø., Harper, D. A. T. & Ryan, P. D. 2001. PAST: paleontologi-cal statistics software package for education and data analysis.Palaeontologia Electronica, 4, 1–9, http://palaeo-electronica.org/2001_1/past/issue1_01.htm.
Hennig, A. H. 1905. Gotlands Silur-Bryozoer, 1. Arkiv for zoology, 2,1–37.
Hennig, A. H. 1906. Gotlands Silur-Bryozoer, 2. Arkiv for zoologi, 3,1–62.
Hu, Z.-X. & Spjeldnæs, N. 1991. Early Ordovician bryozoans fromChina. In: Bigey, F. P. & d’Hondt, J.-L. (eds) Bryozoaires Actuelset Fossiles: Bryozoa Living and Fossil. Societe des Sciences
C. J. BUTTLER ET AL.154

Naturelles de l’Ouest de la France Memoire HS, Bulletins, Nantes,1, 179–185.
INTERNATIONAL COMMISSION ON STRATIGRAPHY 2010. Inter-national Stratigraphic Chart (September 2010 version), http://www.stratigraphy.org/index.php/ics-chart-timescale
Jaccard, P. 1901a. Distribution de la flore alpine dans le bassin desDranses et dans quelques regions voisines. Bulletin de la Societe Vau-doise des Sciences Naturelles, 37, 241–272.
Jaccard, P. 1901b. Etude comparative de la distribution florale dans uneportion des Alpes et des Jura. Bulletin de la Societe Vaudoise desSciences Naturelles, 37, 547–579.
Jimenez-Sanchez, A. & Villas, E. 2009. The bryozoan dispersion intothe Mediterranean margin of Gondwana during the pre-glacial LateOrdovician. Palaeogeography, Palaeoclimatology, Palaeoecology,294, 220–231.
Kopaevich, G. V. 1975. Silurian bryozoans from Estonia and Podolia.Trudy Paleontologischeskogo Instituta Akademiya Nauk SSSR, 151,5–153 [in Russian].
Kopaevich, G. V. 1984. Atlas of Bryozoa from the Ordovician, Silurianand Devonian of Mongolia. Sovmestnaya Sovetsko-MongolskayaPaleontologischeskaya Ekspeditsiya Trudy, 22, 1–164 [in Russian].
Landing, E., English, A. & Keppie, J. D. 2010. Cambrian origin of allskeletalized metazoan phyla – discovery of Earth’s oldest bryozoans(Upper Cambrian, southern Mexico). Geology, 38, 547–550.
Legendre, P. & Legendre, L. 1998. Numerical Ecology. Developmentsin Environmental Modelling. Elsevier, Amsterdam.
McCoy, V. E. & Anstey, R. L. 2010. Biogeographic associations of Silur-ian bryozoan genera in North America, Baltica and Siberia. Palaeo-geography, Palaeoclimatology, Palaeoecology, 297, 420–427.
Murguıa, M. & Villasenor, J. L. 2003. Estimating the effect of simi-larity coefficient and the cluster algorithm in biogeographic classifi-cations. Annales Botanici Fennici, 40, 415–421.
Nekhorosheva, L. V. 1976. Ordovician Bryozoa of the Soviet Arctic. In:Bassett, M. G. (ed.) The Ordovician System. University of WalesPress and National Museum of Wales, Cardiff, 575–582.
Ogg, J. G., Ogg, G. & Gradstein, F. M. 2008. The Concise GeologicTime Scale. Cambridge University Press, Cambridge.
Pocta, P. 1894. 1. Bryozoaires. In: Barrande, J. (ed.) Systeme Silurien duCentre de la Boheme. Charles Bellmann, Prague, 8, 3–132.
Pushkin, V. I. 1975. About the genus Mediapora and its new represen-tatives from the Upper Silurian in Belarus and Latvia. Transactionsof the Geological Museum of Moscow University, 11, 113–120[in Russian].
Pushkin, V. I. & Popov, L. E. 1991. The first information about LateOrdovician bryozoans from mountain range Tarbagataj (Kazakh-stan). Ezhegodnik Vsesoyuznogo Paleontologicheskoga obstchestva,34, 19–34 [in Russian].
Pushkin, V. I. & Popov, L. E. 1999. Early Ordovician bryozoans from theNorth-western Russia. Palaeontology, 42, 171–189.
Pushkin, V. I., Nekhorosheva, L. V., Kopaevich, G. V. & Yaroshins-
kaya, A. M. 1990. Pridoli Bryozoa of the USSR. Nauk, Moscow [inRussian].
Ross, J. R. P. 1961. Ordovician, Silurian and Devonian Bryozoa of Austra-lia. Bureau of Mineral Resources, Bulletin, 50, 1–172.
Ross, J. R. P. 1985. Biogeography of Ordovician ectoproct (bryozoan)faunas. In: Nielsen, C. & Larwood, G. P. (eds) Bryozoa: Ordovi-cian to Recent. Olsen & Olsen, Fredensborg, 265–271.
Ross, J. R. P., Hamedi, M. A. & Wright, A. J. 2000. Late Ordovician(Caradoc) bryozoans from the Kerman district, east-central Iran. In:Herrera, C. A. & Jackson, J. B. C. (eds) Proceedings of the 11thInternational Bryozoology Association Conference. SmithsonianTropical Research Institute, Balboa, R.P., 329–335.
Schmachtenberg, W. 2008. Resolution and limitations of faunal simi-larity indices of biogeographic data for testing predicted paleogeo-graphic reconstructions and estimating intercontinental distances: atest case of modern and Cretaceous bivalves. Palaeogeography,Palaeoclimatology, Palaeoecology, 265, 255–261.
Snell, J. F. 2004. Bryozoa from the Much Wenlock Limestone(Silurian) Formation of the West Midlands and Welsh Borderland.Monographs of the Palaeontographical Society (London), 157,1–136.
Spjeldnæs, N. 1981. Lower Palaeozoic Palaeoclimatology. In: Holland,C. H. (ed.) Lower Palaeozoic of the Middle East, Eastern andSouthern Africa, and Antarctica. John Wiley & Sons, Chichester,199–256.
Taylor, P. D. & Cope, J. C. W. 1987. A trepostome bryozoan from theLower Arenig of South Wales: implications of the oldest describedbryozoan. Geological Magazine, 124, 367–371.
Taylor, P. D. & Curry, G. B. 1985. The earliest known fenestratebryozoan, with short review of Lower Ordovician Bryozoa. Palaeon-tology, 28, 147–158.
Taylor, P. D. & Ernst, A. 2004. Bryozoans. In: Webby, B. D., Paris, F.,Droser, M. L. & Percival, I. G. (eds) The Great OrdovicianBiodiversification Event. Columbia University Press, New York,147–156.
Taylor, P. D. & Rozhnov, S. 1996. A new early cyclostome bryozoanfrom the Lower Ordovician (Volkhov Stage) of Russia. Palaontolo-gische Zeitschrift, 70, 171–180.
Taylor, P. D., Berning, B. & Wilson, M. A. Reinterpretation of theCambrian ‘bryozoan’ Pywackia as an octocoral. Journal of Paleon-tology, in press.
Tuckey, M. E. 1990a. Biogeography of Ordovician bryozoans. Palaeo-geography, Palaeoclimatology, Palaeoecology, 77, 91–126.
Tuckey, M. E. 1990b. Distributions and extinctions of Silurian Bryozoa.In: McKerrow, W. S. & Scotese, C. R. (eds) Palaeozoic Palaeogeo-graphy and Biogeography. Geological Society, London, Memoirs,12, 197–206.
Tuckey, M. E. & Anstey, R. L. 1992. Late Ordovician extinctions ofbryozoans. Lethaia, 25, 111–117.
Williams, A. 1973. Distribution of brachiopod assemblages in relationto Ordovician palaeogeography. In: Hughes, N. R. (ed.) Organismsand Continents through Time. Palaeontological Association,London, Special Papers in Palaeontology, 12, 241–269.
Xia, F., Zhang, S. & Wang, Z. 2007. The oldest bryozoans: new evi-dence from the Late Tremadocian (Early Ordovician) of EastYangtze Gorges in China. Journal of Paleontology, 81, 1308–1326.
Xia, F., Ma, J. & Hao, J. 2010. Ordovician bryozoan diversity changesand bearing on the origin of Bryozoa. Acta PalaeontologicalSinica, 49, 139–163 [in Chinese].
Yang, K. 1957. Some Bryozoa from the upper part of the Lower Ordovi-cian of Liangshan, Southern Shensi (including a new genus). ActaPalaeontologica Sinica, 5, 1–10 [in Chinese].
Zhang, S., Xia, F., Yan, H. & Wang, Z. 2009. Horizon of the oldestknown bryozoans (Ordovician). Palaeoworld, 18, 67–73.
EARLY PALAEOZOIC BIOGEOGRAPHY OF BRYOZOANS 155