bone marrow transplantation from alpha1,3-galactosyltransferase gene-knockout pigs in baboons
TRANSCRIPT

Bone marrow transplantation froma1,3-galactosyltransferase gene-knockoutpigs in baboons
Introduction
The number of patients awaiting organ transplan-tation (Tx) each year continues to increase. There isa clear need for an alternative source of organs andtissues. Pigs offer the possibility of an unlimited
source of organs for Tx [1]. However, innate andadaptive immune responses need to be overcome [2].Bone marrow (BM) Tx has been demonstrated
to induce donor-specific tolerance in rodent [3,4],porcine [5], nonhuman primate [6], and humanclinical allogeneic models [7], and has also been
Tseng Y-L, Dor FJMF, Kuwaki K, Ryan D, Wood J, Denaro M,Giovino M, Yamada K, Hawley R, Patience C, Schuurman H-J, AwwadM, Sachs DH, Cooper DKC. Bone marrow transplantation from a1,3-galactosyltransferase gene-knockout pigs in baboons.Xenotransplantation 2004; 11: 361–370. � Blackwell Munksgaard, 2004
Abstract: Background: Successful hematopoietic cell allotransplantationresults in donor-specific tolerance, but this approach has been unsuc-cessful in the wild-type pig-to-baboon xenotransplantation model, as pigcells were lost from the circulation within 5 days. However, after ces-sation of immunosuppressive therapy on day 28, all baboons demon-strated non-specific unresponsiveness on mixed leukocyte reaction(MLR) for at least 30 days. We have now investigated the transplan-tation of bone marrow (BM) cells from miniature swine homozygous fora1,3-galactosyltransferase gene-knockout (GalT-KO).Methods: Baboons (n ¼ 3) were pre-treated with whole body and thy-mic irradiation, anti-thymocyte globulin, and splenectomy, and receivedimmunosuppressive and supportive therapy for 28 days. BM was har-vested from GalT-KO swine (n ¼ 3). The baboons were monitored forthe presence of pig cells by flow cytometry and colony-forming units(CFUs), and for cellular reactivity by MLR.Results: A mean of 11 · 108 BM cells/kg was infused into each baboon.The mean absolute numbers and percentages of pig cells detected in theblood at 2 h and on days 1, 2 and 4, respectively, were 641/ll (9.5%),132/ll (3.4%), 242/ll (3.9%), and 156/ll (2.9%). One baboon died(from accidental hemorrhage) on day 6, at which time chimerism waspresent in the blood (2.0%) and BM (6.4%); pig cell engraftment in theBM was confirmed by polymerase chain reaction (PCR) of CFUs. In thetwo other baboons, blood chimerism was lost after day 5 but returned atlow levels (<1%) between days 9 to 16 and 7 to 17, respectively, in-dicating transient BM engraftment. Both surviving baboons showednon-specific unresponsiveness on MLR until they were euthanized ondays 85 and 110, respectively.Conclusions: By using BM cells from GalT-KO pigs, chimerism wasdetected at levels comparable with previous studies when 30-fold moregrowth factor-mobilized peripheral blood progenitor cells had beentransplanted. In addition, cellular hyporesponsiveness was prolonged.However, long-term engraftment and chimerism were not achieved.
Y.-L. Tseng,1 F.J.M.F Dor,1
K. Kuwaki,1 D. Ryan,2 J. Wood,2
M. Denaro,2 M. Giovino,2
K. Yamada,1 R. Hawley,2
C. Patience,2 H.-J. Schuurman,2
M. Awwad,2 D.H. Sachs1 andD.K.C. Cooper11Transplantation Biology Research Center,Massachusetts General Hospital/Harvard MedicalSchool, Boston, MA, 2Immerge BioTherapeutics, Inc.,Cambridge, MA, USA
Key words: a1,3-galactosyltransferase – bonemarrow transplantation – chimerism –hematopoietic cells – xenotransplantation
Abbreviations: Ab: antibody; BM: bone marrow;CFU: colony-forming unit; Gal: Gala1,3Gal; GalT-KO:a1,3-galactosyltransferase gene-knockout; MLR:mixed leukocyte reaction; PBMC: peripheral bloodmononuclear cells; PBPC: growth factor-mobilizedperipheral blood progenitor cells; PCR: polymerasechain reaction; Tx: transplantation.
Address reprint requests to D.K.C. Cooper MD PhDFRCS, Transplantation Biology Research Center, M-assachusetts General Hospital, MGH East,Building 149-9019, 13th Street, Boston, MA 02129,USA (E-mail: [email protected])
Received 20 April 2004;Accepted 25 April 2004
Xenotransplantation 2004: 11: 361–370Printed in UK. All rights reserveddoi: 10.1111/j.1399-3089.2004.00151.x
Copyright � Blackwell Munksgaard 2004
XENOTRANSPLANTATION
361

successful in concordant rodent [8], nonhumanprimate [9] and pig-to-NOD/SCID mouse [10]xenogeneic models. However, it has not beenachieved in a pig-to-nonhuman primate model.Sablinski [11] and Kozlowski [12,13] and theirrespective colleagues failed to detect macrochimer-ism (i.e. chimerism detectable by flow cytometry, asopposed to microchimerism, that is detectable onlyby polymerase chain reaction) following pig BMTx (2 to 30 · 108 cells/kg) in monkeys andbaboons. Buhler et al. [14–16] transplanted largenumbers of pig growth factor-mobilized peripheralblood progenitor cells (PBPC; 3 to 4.6 · 1010 cells/kg) into baboons, and detected macrochimerismfor up to 5 days; two of the 15 baboons showedreappearance of macrochimerism, suggesting tran-sient engraftment in the baboon BM. The potentialfor natural antibody (Ab) reacting against galac-tosea1,3galactose (Gal) expressed on the pig cellswas considered a major complicating factor in thismodel. Although anti-Gal Ab was immunoad-sorbed before BM or PBPC Tx in all studies[11–16], this Ab began to reappear within hours.The recent availability of pigs homozygous for
a1,3-galactosyltransferase gene-knockout (GalT-KO) [17,18], bred through nuclear transfer/embryotransfer, has enabled us to investigate the Tx ofBM cells that do not express the Gal antigen.
Methods
Animals
Baboons (Papio hamadryas, n ¼ 3), of knownABO blood group with a body weight 8 to 11 kg(Manheimer Foundation, Homestead, FL, USA)were recipients of BM Tx. GalT-KO miniatureswine (n ¼ 3) (bred by nuclear transfer/embryotransfer by Immerge Biotherapeutics, Cambridge,MA, USA), of blood group O, swine leukocyteantigen haplotype dd, and weighing 14 to 20 kg,served as BM donors. All animal care procedureswere in accordance with the Principles of Laborat-ory Animal Care formulated by the NationalSociety for Medical Research and the Guide forthe Care and Use of Laboratory Animals preparedby the Institute of Laboratory Animal Resourcesand published by the National Institutes of Health(NIH publication No. 86-23, revised 1996). TheMassachusetts General Hospital Subcommittee ofResearch Animal Care approved the protocol.
Surgical procedures
All surgical procedures (e.g. splenectomy, intra-venous or intra-arterial line insertion) were
performed during inhalation anesthesia; detailshave been described previously [19], as have themethods of porcine BM processing [20].
Non-myeloablative conditioning regimen
The non-myeloablative conditioning regimen(Fig. 1) consisted of induction therapy with splen-ectomy (performed on day )8), whole body irra-diation (150 cGy ·2 on days )6 and )5), thymicirradiation (700 cGy on day )1), and anti-thymo-cyte globulin (50 mg/kg on days )3, )2 and )1;ATGAM, Upjohn, Kalamazoo, MI, USA). Ifnecessary, additional anti-CD2 monoclonal anti-body (mAb) (LoCD2b 1 to 4 mg/kg; ImmergeBiotherapeutics) was given to maintain the T-cellcount <100 cells/ll for the first week. Mainten-ance therapy consisted of eight doses of a humananti-human CD154 mAb (25 mg/kg; NovartisPharma, Basel, Switzerland) on days 0, 1, 4, 7,10, 14, 19 and 24 (trough level >400 lg/ml);mycophenolate mofetil (110 mg/kg/day by con-tinuous i.v. infusion, from day )6 to day 28 tomaintain a level of 3 to 6 lg/ml), and cobra venomfactor (Advanced Research Technologies, SanDiego, CA, USA) to maintain the CH50 at 0%for the first 2 weeks. Growth factors, pig interleu-kin-3 (100 lg/kg/day for 3 weeks; Immerge Bio-therapeutics) and stem cell factor (50 lg/kg/day for2 weeks; Immerge Biotherapeutics, which is notpig-specific but functions across species barriers)were administered to promote pig BM cell survivaland engraftment. Prostacyclin (PG12; 20 ng/kg/min by continuous i.v. infusion), heparin (10 U/kg/hi.v. infusion), and methylprednisolone (2 mg/kg ·2i.v. for 7 days followed by tapering and discon-tinuation over the next 28 days) were given to
SplenectomyBone marrow aspirationLine insertion
Whole body irradiation150 cGy/each
ATG
Thymic irradiation700 cGy
CVF
MMF (110 mg/kg)
Pig SCF (50 µg/kg)
Prostacyclin (20 ng/kg/min)
Pig IL3 (100 µg/kg)
Heparin
Methylprednisolone
Day -7 -6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
BMTx
Anti-CD2 was given if the T-cell count > 100 /µL from D-1 to D+7
Fig. 1. Conditioning and immunosuppressive regimen for pig-to-baboon BM Tx. All immunosuppressive therapy was stop-ped on day 28. During the first week after BM Tx, an anti-CD2mAb was given to maintain the T-cell count <100 cells/ll. Ahuman anti-human CD154 mAb was administered on dayshighlighted in bold type, and then at 5-day intervals untilday 24.
Tseng et al.
362

prevent baboon platelet aggregation around thepig cells and thrombotic microangiopathy [21].
Monitoring and supportive therapy
Blood cell counts, chemistry, coagulation para-meters, and the blood levels of immunosuppressiveagents were determined by routine methods. Bloodcultures were performed twice weekly. The baboonreceived irradiated (3000 cGy) red blood cells if thehematocrit fell <20%, and a blood type-compa-tible platelet transfusion if the platelet count fell<20 000/ll. Levofloxacin (10 mg/kg/day), ganci-clovir (5 mg/kg/day), and cimetidine (5 mg/kg ·2daily) were administered during the period whenthe baboon had indwelling intravascular catheters.
Flow cytometry for detection of pig cell chimerism
Phenotyping of baboon blood and BM chimerismwas performed with the following mAbs: W6/32(mouse anti-human major histocompatibility com-plex class I IgG; Serotec, Oxford, UK), 1030H-1-19or pan-pig (mouse anti-pig leukocyte IgM), 76-7-4(mouse anti-pig CD1 IgG), 898H2-6-15 (mouseanti-pig CD3 IgG), G7 (mouse anti-pig CD16 IgG),74-22-15 or SWC3a (mouse anti-pig granulocyte-monocytes), 74-12-4 (mouse anti-pig CD4 IgG,),76-2-11 (mouse anti-pig CD8a IgG), 1038H-4-6(mouse anti-pig CD9,). Baboon blood or BM cellswere washed, after which, 1 · 106 cells (100 ll)were stained in FACS media [Hank’s balanced saltsolution (HBSS) with Ca++ and Mg++/0.1%bovine serum albumin (BSA) 0.1% NaN3; Gibco,Grand Island, NY, USA] according to the follow-ing steps for two color analysis; incubation with adirectly-labeled fluorescein isothiocyanate (FITC)-conjugated first mAb (W6/32) (10 ll of predeter-mined optimal concentration), incubation with thebiotinylated second mAb (1030H-1-19, 76-7-4,898H-2-6-15, G7, 74-22-15, 74-12-4, 1038H-4-6)(10 ll), followed by adding 10 ll of phycoerythrin-conjugated streptavidin (Becton Dickinson, Sun-nyvale, CA, USA). Propidium iodide was addedimmediately before acquisition, and dead cells wereexcluded from analysis based on propidium iodidestaining, if necessary. Data were acquired using aFACS fluorescence cytometer (Becton Dickinson)and analyzed using Winlist mode analysis software(Verity Software House, Topsham, ME, USA).
Flow cytometry for determination of total baboon CD3+ cells
The T-cell numbers in the blood were measured byflow cytometry on day )7, day )1, daily to day 20,and then twice a week, by staining with a FITC-
conjugated mouse anti-monkey CD3 Ab (Bio-Source, Camarillo, CA, USA). The total T-cellcount was calculated from the white blood cellcount and flow cytometry data.
Pig colony-forming unit assay in baboon bone marrow
Aspirated baboon BM was set up in colony-forming unit (CFU) assay using MethocultH4230 and H4435 (Stem Cell Technologies, Van-couver, BC, Canada). The BM sample was filteredthrough a strain filter after dilution with 8 mlHBSS. The diluted sample was slowly added andoverlaid onto the lymphocyte separation medium(density 1.077 to 1.08 g/ml; ICN Biomedicals,Aurora, OH, USA). After centrifugation, theinter-face cell layer was removed, washed, andcentrifuged. The supernatant was removed and thepellet was resuspended in mixed leukocyte reaction(MLR) medium [RPMI 1640/without L-glutamine(Fisher Scientific, Agawam, MA, USA) with 0.1%non-essential amino acid, 1% sodium pyruvate,1% hepes buffer, 6% fetal pig serum, 0.1%2-mercaptoethanol, 1% L-glutamine, 200 units/mlpenicillin and streptomycin (Gibco), and0.05 mg/ml gentamicin]. Cells (1 to 2 · 105 cells)were plated into 35 mm dishes (Nunc, Naperville,IL, USA). Methocult H4230 [composed of 1%methylcellulose in Iscove’s modified Dulbecco’smedium containing 30% fetal calf serum (FCS),1%BSA, 10)4 m 2-mercaptoethanol, and 2 mm
L-glutamine] was supplemented with a final con-centration of 100 ng/ml of porcine interleukin-3,and the following recombinant human hemato-poietic growth factors: stem cell factor (50 ng/ml),granulocyte-monocyte colony-stimulating fac-tor (20 ng/ml), interleukin-3 (20 ng/ml), interleu-kin-6 (20 ng/ml), granulocyte colony-stimulatingfactor (20 ng/ml), and erythropoietin (3 U/ml).Methocult H 4435, a methylcellulose-based med-ium, contained the recombinant human cytokinesSCF, GM-CSF, IL-3, IL-6, G-CSF, and erythro-poietin. Total CFUs (or colony-forming cells,CFC) were counted after 11 to 14 days of cultureat 37 �C in 5% CO2. The origin of pooled colonies(i.e. from pig or baboon medium) was determinedper lineage by polymerase chain reaction.
Polymerase chain reaction for detection of pig cell chimerism
A bulk polymerase chain reaction (PCR) assay thatamplifies the porcine cytochrome b gene was usedto detect pig microchimerism in baboon samples.Four 5-fold concentrations of an internal PCRstandard, starting at 10 copies (0.0002%), wereused. This PCR standard was created by adding
GalT-KO bone marrow transplantation
363

the porcine cytochrome b primer sequence (Cyt-F1:5¢ GCC TAT TCA TCC ACG TAG GC3¢, Cyt-R1: 5¢ CAT TCT ACG AGG TCT GTT CCG 3¢)to either side of a fragment of human A-2phospholipase, followed by cloning into a plasmidvector and quantitation using limiting dilutionPCR. Peripheral blood and BM cells were purifiedusing Histopaque 1077 (Sigma, St Louis, MO,USA). The cells were pelleted and frozen at )80 �Cat 2 to 10 · 105 cells until use. Genomic DNA wasextracted using the Qiamp Blood Kit (Qiagen,Valencia, CA, USA), quantified using Hoechst33285 dye and a Hoeffer TKO-100 Fluorometer(Hoeffer Scientific Instruments, San Francisco,CA, USA), and then denatured by heating to100 �C for 10 min. The microchimerism assayconsisted of 250 ng of sample DNA added to atotal PCR reaction volume of 100 ll. The finalreagent mixture consisted of 1 · GeneAmp PCRBuffer II, 2.0 mm MgCl2, 0.4 mm dNTP, and 2.5 Uof Amplitag Gold (Perkin-Elmer, Philadelphia,PA, USA). Eighty picomoles each of Cyt-F1 andCyt-R1 primers were added to the reactions. ThePCR tubes were placed in a Perkin-Elmer Gene-Amp 9600 Thermal Cycler. The initial denaturingstep was 9 min at 95 �C followed by 40 cycles of96 �C for 10 s, 59 �C for 30 s, and 72 �C for 30 s.(Detection of microchimerism by PCR using 40cycles is associated with an approximate 20% falsepositivity, necessitating repeated confirmatory test-ing if the level of positivity is <0.001%). Theprogram concluded with a 5 min incubation at72 �C. The internal standard was added to PCRreactions along with sample DNA to produce a240-bp product that was compared with the 210-bpproduct produced by the porcine cytochromeb gene through ethidium bromide-stained agarosegels. Samples that produced a cytochromeb product that was greater than or equal to theproduct from five copies of the standard sampleswere considered positive for microchimerism.Standard samples of known pig DNA quantitywere run with the experimental samples. Theintensity of the experimental sample band onthe gel was compared visually with the intensityof the standards.
Pig colony-forming unit polymerase chain reaction in baboons
The cellular fractions from the BM aspirate wereisolated, and cells were plated for CFU assays (asabove). Following 10 to 14 days incubation in 5%CO2 at 37 �C, each culture dish was evaluatedfor the presence of CFU-myeloid (CFU-GM),CFU-granulocyte-erythroid-myeloid-megakaryocyte(CFU-GEMM), CFU-erythroid (CFU-E), and
CFU-burst-forming unit-erythroid (BFU-E). Col-onies of each type were then picked using aninverted microscope and pipette, and placed into80 ll phosphate buffered saline (PBS). Thesesamples were then pelleted, the PBS aspirated,and stored at )80 �C until further analysis. DNAwas obtained from pelleted CFUs by incubatingfor 1 h at 55 �C in 50 ll of lysing buffer (10 mm
TrisCl (pH 8.3), 50 mm KCL, 0.45% NP-40,0.45% Tween-20, 100 lg/ml proteinase K; Sigma)followed by an incubation at 100 �C for 10 min.The cytochrome b PCR setup and thermal cyclingwas the same as used for bulk cells (above) exceptthat the reaction was scaled down 2-fold to a 50 lltotal volume. Cell lysate DNA (12.5 ll) was addedto each PCR along with 400 copies of cytochromeb internal standard (about four pig cell-equivalentsof cytochrome b). CFUs that produced a cyto-chrome b product greater than the internal stand-ard product were considered to be of pig origin.
Detection of anti-Gal and anti-nonGal antibodies
Details of these methods have been reportedpreviously [12]. Anti-Gal Ab was measured byenzyme-linked immunosorbent assay. Anti-nonGalIgM and IgG were determined by flow cytometryusing fresh GalT-KO (SLAdd) PBMC as targetcells. GalT-KO pig blood was washed, after which1 · 106 cells (100 ll) were stained in FACS mediaplus 10% normal goat serum with a directly-labeled FITC-conjugated goat anti-human IgM orIgG (Immerge Biotherapeutics). Emergent anti-nonGal Ab, resulting from sensitization to pigcells, was detected by comparing anti-pig reactivityof pre-transplant serum with that of post-trans-plant serum.
Serum cytotoxicity assay
Target cell suspensions (Gal)/) SLAdd PBMC)were diluted to 5 · 106 cells/ml in medium 199(Cellgro, Herndon, VA, USA) supplemented with2% FCS (culture medium). Serum samples wereserially diluted in medium from 1 : 2 to 1 : 2048.Using 96-well U-bottom plates (Costar, Cam-bridge, MA, USA), 25 ll of each dilution werepipetted into each well as designated and 25 ll ofmedium in the control wells for media and com-plement. Twenty-five ll of the appropriate targetcell suspension (Gal)/) SLAdd PBMC) was trans-ferred into the selected wells and rows, includingthe complement and medium control wells. Plateswere incubated in a CO2 incubator for 15 min at37 �C, which then were washed with 125 ll ofmedium. The plate was then centrifuged at
Tseng et al.
364

1200 rpm for 5 min at 4 �C, and supernatants wereflicked out, and the plate was blotted. Twenty-fivell of diluted rabbit complement, at a predeter-mined working dilution in medium, was added toeach well except the medium control wells, inwhich 25 ll of medium was pipetted. Then theplate was incubated in a CO2 incubator at 37 �Cfor 30 min, after which it was centrifuged at1500 rpm for 5 min at 4 �C. The plate was placedon ice and supernatants drawn off from the firstfive wells and discarded. Then 10 ll of trypan bluesolution were pipetted into each of these five wells,and the cells were resuspended. Next, 6 ll of theresuspension were dispensed into one of the fiverings on the microscope slide, on which circularcoverslips were carefully placed. Using the 40·objective, the numbers of dead and live cellswere counted until 100 counts had been made.The number of dead cells was equivalent to thepercentage of dead cells present in each well. Theresult was defined as positive if the percentage ofcells killed was >20% in more than three serialdilutions of serum.
Mixed leukocyte reaction
Details of this assay have been reported previously[11]. All baboons that received BM Tx were testedfor an anti-pig MLR response before and after BMTx at regular intervals until they were killed.Responder baboon cells were stimulated in tripli-cate with pig donor-specific cells (SLAdd), pig non-specific cells (SLAaa, SLAcc), and allogeneicbaboon cells. The results were expressed bystimulation index [Recipient anti-experimental
stimulator response (CPM)/Recipient anti-autolo-gous stimulator response (CPM)].
Results
Two baboons (B213 and B217) survived untilkilled on days 85 and 110, respectively, and thethird (B210) died on day 6 from bleeding from abroken intra-arterial catheter.
Pretransplant anti-pig nonGal antibody status of baboons
Pre-BM Tx, no IgM or IgG binding to GalT-KOpig cells was detected by flow cytometry in theserum of any of the three baboons, and there wasno serum cytotoxicity to GalT-KO PBMC.
Response to conditioning regimen and GalT-KO pig bone marrowinfusion
All three baboons had transient WBC countelevation after BM Tx (not shown). The twosurviving baboons had WBC counts below normalrange (<4000/ll) from days 11 and 12 until days19 and 22, respectively. All three baboons showedgood T-cell depletion before BM Tx (Fig. 2A).T-cell counts were largely maintained <100/llduring the first week, remained low for 2 weeks,and then gradually returned to normal. There wasa trend for the number of baboon monocytes torise during the periods of pig cell chimerism(Fig. 2B,C). The platelet counts fell slowly duringthe first week after BM Tx (Fig. 2D); B213required platelet transfusions only because it dis-connected its indwelling i.v. line on three occasions
–6 –3 0 3 6 9 1215 18 2124 2730 33 360
100200300400500600
B213B210
6501650265036504650
B217
*
Day
CD
3+ T
–cel
ls/µ
l
–6–3 0 3 6 9 1215182124273033360
50100150200250300350400450500
*
Day
k/µl
0 3 6 9 12 15 18 21 24 27 30 33 360
10
20
30
40
50
*
Day
Per
cent
age
0 3 6 9 12 15 18 21 24 27 30 33 360
200400600800
100012001400 *
200030004000500060007000
Day
/µl
A B
C D
Fig. 2. (A) T-cell counts (CD3+ cells/ll)in the three baboons before and during thefirst 35 days after BM Tx. (B and C)Changes in host monocytes in the blood ofthe three baboons after BM Tx. (B) Per-centages, (C) Absolute numbers (per ll).(D) Platelet counts (in k/ll) in the threebaboons before and during the first35 days after BM Tx. (*denotes day ofdeath of B210).
GalT-KO bone marrow transplantation
365

and lost blood. In B213 and B217, the plateletcounts had returned to >100 k/ll on days 16 and18, respectively; B210 had an adequate plateletcount (144 k/ll) on the day of its death (day 6). Nosignificant change was documented in serumlactate dehydrogenase level in any baboon (datanot shown), in contrast to some previous studieswith pig PBPC Tx [16]. Occasional irradiatedpacked red blood cell transfusions were requiredin the 2 surviving baboons to maintain the hemat-ocrit at >20%. No clinical features of a con-sumptive coagulopathy were observed [16].
Pig cell chimerism in baboon blood
The percentages of pig cell chimerism and theabsolute numbers of pig cells in the baboons’ bloodby flow cytometry are shown in Fig. 3(A) and (B),respectively. Chimerism was detected initially for5 days in two (B213, B217), and B210 was stillchimeric when it died on day 6; the mean absolutenumbers of pig cells detected in the blood at 2 hand on days 1, 2 and 4, respectively, were 641, 132,242, and 156/ll, which represented mean chimer-ism of 9.5, 3.4, 3.9, and 2.9%, respectively. The twosurviving baboons (B213, B217) had reappearanceof chimerism from days 9 to 16 and 7 to 17,respectively, with the highest levels (0.77 and0.35%, representing 15 and 27 cells/ll) seen ondays 11 and 12, respectively (Fig. 3A,B). In allthree baboons, the initial chimerism was multilin-eage (Fig. 4A). Continuous chimerism was detec-ted by PCR in B213 and B217 until days 17 and 21,respectively, after which it was lost.
Pig cell chimerism in baboon bone marrow
The BM taken at necropsy on day 6 from B210showed 6.4% chimerism by flow cytometry(Fig. 4B). This engraftment was confirmed byPCR and CFU and was demonstrated to bemultilineage (Table 1). In B213 and B217, BMwas aspirated on day 28, but no chimerism wasdetected by FACS, PCR or CFU assay.
Anti-Gal and anti-nonGal antibody responses
No anti-nonGal Ab was detected by FACS ineither of the two surviving baboons until the day ofeuthanasia (days 85 and 110, respectively), (or inB210 before death on day 6), indicating nosensitization to pig BM cells; serum cytotoxicityto GalT-KO cells remained negative. There wereno changes in anti-Gal Ab levels.
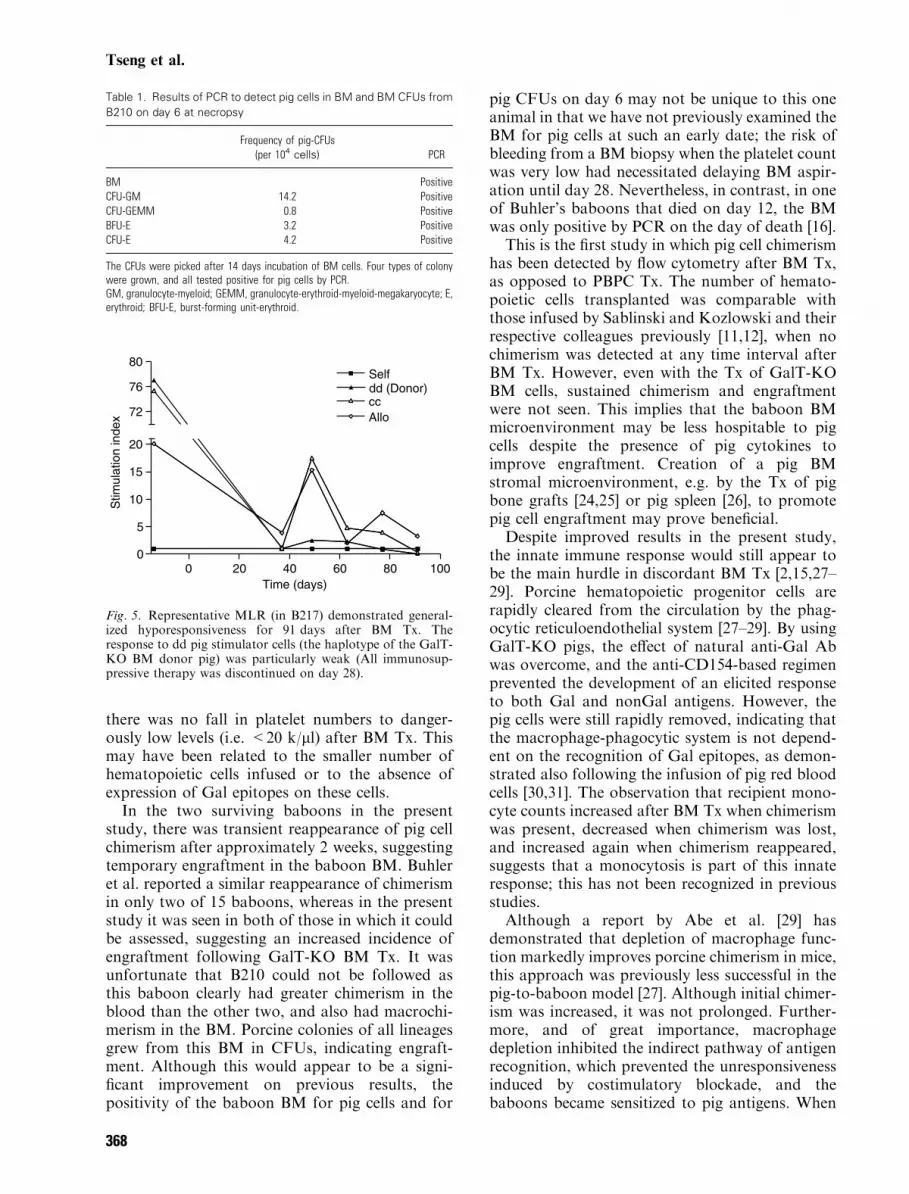
Mixed leukocyte reaction
A representative MLR (from B217) is shown inFig. 5. All immunosuppressive therapy was dis-continued on day 28, but the baboon remainedhyporesponsive to pig and baboon cells for91 days; the response to pig stimulator cells ofthe dd haplotype (that of the GalT-KO donor pig)was especially low. MLRs of B213 also showedgeneralized hyporesponsiveness to pig PBMC untilkilled on day 84.
Discussion
The non-myeloablative conditioning regimen usedin the present study was based on that used byBuhler et al. [14–16]. The major difference wasthat in Buhler’s series the number of PBPCtransplanted was 30-fold greater than the numberof BM cells transplanted in the present study.Eight doses of a murine anti-CD154 mAb wereadministered over 16 days in Buhler’s series,whereas in the present study a human anti-humanCD154 mAb was given at less frequent intervalsover 28 days. The dosages of pig growth factorsadministered to the baboons were considerablyless than used by Buhler (interleukin-3 25 to 50%;stem cell factor 2.5 to 5%). However, even withlower doses of pig growth factors, we obtainedsimilar levels of chimerism following BM Tx withGalT-KO cells although 30-fold less pig cells wereinfused.In Buhler’s reports, platelet aggregation due
to platelet and/or endothelial cell activation by
0 2 4 6 8 10 12 14 16 180
5
10
15
*
Day
Per
cent
age
0 2 4 6 8 10 12 14 16 180
100
B213B210
250
500
750
1000
B217
*
Day
/µl
A B
Fig. 3. Pig cell chimerism detected by flowcytometry in the blood after BM Tx in thethree baboons. (A) Percentages; (B)Absolute numbers of pig cells (per ll).Chimerism reappeared for several days inthe two surviving baboons (B213, B217) ondays 9 and 7, respectively. The baboon thatdied on day 6 (B210) had 243 pig cells/ll inits blood at the time of its death. (*denotesday of death of B210).
Tseng et al.
366

porcine PBPC led to profound thrombocytopenia(<20 k/ll) immediately after infusion of the pigPBPC. In vitro studies indicated that baboon
platelets aggregated around pig PBPC [22,23]. Inthe present study, although there was a gradualreduction in platelet count following irradiation,
3.04%
Pan–pigA
B Pan–pig (6.43%) CD 9 (4.63%)
CD 16 (2.84%) SWC 3 (3.47%)
CD 9 CD 16 SWC 3
101 102 103 104 101 102 103 104 101 102 103 104 101 102 103 104
0 10
2030
4050
6070
80
101 102 103 104 101 102 103 104 101 102 103 104 101 102 103 104
101 102 103 104101 102 103 104101 102 103 104101 102 103 104
101 102 103 104 101 102 103 104
101 102 103 104101 102 103 104
0 10
2030
4050
1020
3040
6050
1020
1020
30
0 10
200
10
010
2030
4050
60
010
2030
4050
60
010
7080
9010
0
1020
3040
1020
3040
50
2040
6080
120
100
140
2040
6080
120
100
140
1020
3040
5060
70
1020
3040
5060
7080
1.57%1.60%2.60%
10.32% 9.22%9.58%8.81%
15.11% 0.87%0.76%14.18%
B213
B210
B217
Fig. 4. (A) Flow cytometry of blood 2 h after BM Tx showed multilineage chimerism, mainly of CD9+, CD16+, or SWC3+ cells, inall three baboons; very small numbers of CD1+ and CD3+ cells were also detected (not shown). (B) Flow cytometry of BM of B210at necropsy on day 6 showed multilineage pig cell chimerism.
GalT-KO bone marrow transplantation
367
there was no fall in platelet numbers to danger-ously low levels (i.e. <20 k/ll) after BM Tx. Thismay have been related to the smaller number ofhematopoietic cells infused or to the absence ofexpression of Gal epitopes on these cells.In the two surviving baboons in the present
study, there was transient reappearance of pig cellchimerism after approximately 2 weeks, suggestingtemporary engraftment in the baboon BM. Buhleret al. reported a similar reappearance of chimerismin only two of 15 baboons, whereas in the presentstudy it was seen in both of those in which it couldbe assessed, suggesting an increased incidence ofengraftment following GalT-KO BM Tx. It wasunfortunate that B210 could not be followed asthis baboon clearly had greater chimerism in theblood than the other two, and also had macrochi-merism in the BM. Porcine colonies of all lineagesgrew from this BM in CFUs, indicating engraft-ment. Although this would appear to be a signi-ficant improvement on previous results, thepositivity of the baboon BM for pig cells and for
pig CFUs on day 6 may not be unique to this oneanimal in that we have not previously examined theBM for pig cells at such an early date; the risk ofbleeding from a BM biopsy when the platelet countwas very low had necessitated delaying BM aspir-ation until day 28. Nevertheless, in contrast, in oneof Buhler’s baboons that died on day 12, the BMwas only positive by PCR on the day of death [16].This is the first study in which pig cell chimerism
has been detected by flow cytometry after BM Tx,as opposed to PBPC Tx. The number of hemato-poietic cells transplanted was comparable withthose infused by Sablinski and Kozlowski and theirrespective colleagues previously [11,12], when nochimerism was detected at any time interval afterBM Tx. However, even with the Tx of GalT-KOBM cells, sustained chimerism and engraftmentwere not seen. This implies that the baboon BMmicroenvironment may be less hospitable to pigcells despite the presence of pig cytokines toimprove engraftment. Creation of a pig BMstromal microenvironment, e.g. by the Tx of pigbone grafts [24,25] or pig spleen [26], to promotepig cell engraftment may prove beneficial.Despite improved results in the present study,
the innate immune response would still appear tobe the main hurdle in discordant BM Tx [2,15,27–29]. Porcine hematopoietic progenitor cells arerapidly cleared from the circulation by the phag-ocytic reticuloendothelial system [27–29]. By usingGalT-KO pigs, the effect of natural anti-Gal Abwas overcome, and the anti-CD154-based regimenprevented the development of an elicited responseto both Gal and nonGal antigens. However, thepig cells were still rapidly removed, indicating thatthe macrophage-phagocytic system is not depend-ent on the recognition of Gal epitopes, as demon-strated also following the infusion of pig red bloodcells [30,31]. The observation that recipient mono-cyte counts increased after BM Tx when chimerismwas present, decreased when chimerism was lost,and increased again when chimerism reappeared,suggests that a monocytosis is part of this innateresponse; this has not been recognized in previousstudies.Although a report by Abe et al. [29] has
demonstrated that depletion of macrophage func-tion markedly improves porcine chimerism in mice,this approach was previously less successful in thepig-to-baboon model [27]. Although initial chimer-ism was increased, it was not prolonged. Further-more, and of great importance, macrophagedepletion inhibited the indirect pathway of antigenrecognition, which prevented the unresponsivenessinduced by costimulatory blockade, and thebaboons became sensitized to pig antigens. When
Table 1. Results of PCR to detect pig cells in BM and BM CFUs from
B210 on day 6 at necropsy
Frequency of pig-CFUs(per 104 cells) PCR
BM PositiveCFU-GM 14.2 PositiveCFU-GEMM 0.8 PositiveBFU-E 3.2 PositiveCFU-E 4.2 Positive
The CFUs were picked after 14 days incubation of BM cells. Four types of colonywere grown, and all tested positive for pig cells by PCR.GM, granulocyte-myeloid; GEMM, granulocyte-erythroid-myeloid-megakaryocyte; E,erythroid; BFU-E, burst-forming unit-erythroid.
0 20 40 60 80 1000
5
10
15
20
ccdd (Donor)
Allo
Self
72
76
80
Time (days)
Stim
ulat
ion
inde
x
Fig. 5. Representative MLR (in B217) demonstrated general-ized hyporesponsiveness for 91 days after BM Tx. Theresponse to dd pig stimulator cells (the haplotype of the GalT-KO BM donor pig) was particularly weak (All immunosup-pressive therapy was discontinued on day 28).
Tseng et al.
368

and how to block the innate immune responsewithout inhibiting the tolerance induced by anti-CD154 mAb-based suppression is a key issuerequiring further exploration.No anti-nonGal Ab developed in the two survi-
ving baboons. A likely explanation for this is thatthe period of pig cell chimerism, i.e. the period oftime in which the baboon was exposed to pigantigens, remained too transient for sensitizationto develop. Costimulatory blockade preventedsensitization during the period of exposure, andthere was no exposure to pig cells once immuno-suppressive therapy was discontinued.The cellular hyporesponsiveness documented in
the present study by MLR to both pig and allo-baboon cells were significantly prolonged in com-parison to previous reports [14–16]. The cellularresponse to donor-specific dd stimulators (thehaplotype of GalT-KO pigs) was especially low inB217. This observation correlates well with therecent report by Kuwaki et al. [32] who demon-strated that a similar regimen based on anti-CD154mAb efficiently suppresses the cellular responseafter pig heart Tx in baboons. However, in thepresent study, the general hyporesponsiveness wassustained for a long period after all drugs had beendiscontinued. The half-life of the human anti-human CD154 mAb used has been estimated to beapproximately 10 to 15 days [33] yet the hypore-sponsiveness to both pig and baboon cells lasted atleast 2 months after discontinuing this agent.In summary, although the presence of Gal on pig
BM cells has been obviated, and the elicited Aband cellular responses can be prevented or minim-ized by a costimulatory blockade-based regimen,several problems in discordant BM Tx still requireresolution. In particular, these include (i) the rapidremoval of the pig cells by the host’s macrophage-phagocytic system, and (ii) modification of thehost’s stromal environment to be more supportiveof pig cell engraftment and proliferation.
Acknowledgments
We thank Leo Buhler MD PhD, and Julian DownPhD for their critical reviews of this manuscript.Y.-L. Tseng is a recipient of a grant from NationalCheng Kung University Hospital, Taiwan.F.J.M.F. Dor is a recipient of grants from theTer Meulen Fund from the Royal NetherlandsAcademy of Arts and Sciences, the Prof. Michael-van Vloten Fund, and the Netherland-AmericaFoundation. Work in our laboratory is supportedin part by NIH Program Project 1PO1 A145897and by a Sponsored Research Agreement betweenthe Massachusetts General Hospital and Immerge
Biotherapeutics. D.H. Sachs and D.K.C. Cooperare consultants to Immerge Biotherapeutics.
References
1. Cooper DKC, Ye Y, Rolf LL, Zuhdi N. The pig aspotential organ donor for man. In: Cooper DKC, Kemp
E, Reemtsma K, White DJG, eds. Xenotransplantation(First edn). Heidelberg: Springer, 1991: 481.
2. Sachs DH, Sykes M, Robson SC, Cooper DKC. Xeno-transplantation. Adv Immunol 2001; 79: 129.
3. Sharabi Y, Sachs DH. Mixed chimerism and permanentspecific transplantation tolerance induced by a nonlethalpreparative regimen. J Exp Med 1989; 169: 493.
4. Lee LA, Sergio JJ, Sykes M. Natural killer cells weaklyresist engraftment of allogeneic long-term multilineage-repopulating hematopoietic stem cells. Transplantation1996; 61: 125.
5. Huang CA, Fuchimoto Y, Scheier-Dolberg R et al.Stable mixed chimerism and tolerance using a nonmyelo-ablative preparative regimen in a large-animal model.J Clin Invest 2000; 105: 173.
6. Kawai T, Cosimi AB, Colvin RB et al. Mixed allogeneicchimerism and renal allograft tolerance in cynomolgusmonkeys. Transplantation 1995; 59: 256.
7. Spitzer TR, Delmonico F, Tolkoff-Rubin N et al.Combined histocompatibility leukocyte antigen-matcheddonor bone marrow and renal transplantation for multiplemyeloma with end stage renal disease: the induction ofallograft tolerance through mixed lymphohematopoieticchimerism. Transplantation 1999; 68: 480.
8. Sharabi Y, Aksentijevich I, Sundt TM III et al. Specifictolerance induction across a xenogeneic barrier: produc-tion of mixed rat/mouse lymphohematopoietic chimerausing a nonlethal preparative regimen. J Exp Med 1990;172: 195.
9. Bartholomew AM, Powelson J, Sachs DH et al. Tol-erance in a concordant nonhuman primate model. Trans-plantation 1999; 68: 1708.
10. Abe M, Yi J, Sykes M et al. Mixed chimerism inducesdonor-specific T-cell tolerance across a highly disparatexenogeneic barrier. Blood 2002; 99: 3823.
11. Sablinski T, Emery DW, Monroy R et al. Long-termdiscordant xenogeneic (porcine-to-primate) bone marrowengraftment in a monkey treated with porcine-specificgrowth factors. Transplantation 1999; 67: 972.
12. Kozlowski T, Monroy R, Xu Y et al. Anti-Gala1-3 Galantibody response to porcine bone marrow in unmodifiedbaboons and baboons conditioned for tolerance induction.Transplantation 1998; 66: 176.
13. Kozlowski T, Monroy R, Giovino M et al. Effect of pig-specific cytokines on mobilization of hematopoietic pro-genitor cells in pigs and on pig bone marrow engraftmentin baboons. Xenotransplantation 1999; 6: 17.
14. Buhler L, Awwad M, Basker M et al. High-dose por-cine hematopoietic cell transplantation combined withCD40 ligand blockade in baboons prevents an inducedantipig humoral response. Transplantation 2000; 69: 2296.
15. Buhler L, Alwayn IPJ, Basker M et al. CD40-CD154pathway blockade requires host macrophage to inducehumoral unresponsiveness to pig hematopoietic cells inbaboons. Transplantation 2001; 72: 1759.
16. Buhler L, Awwad M, Treter S et al. Pig hematopoieticcell chimerism in baboons conditioned with a nonmyelo-ablative regimen and CD154 blockade. Transplantation2002; 73: 12.
GalT-KO bone marrow transplantation
369

17. Phelps CJ, Koike C, Vaught TD et al. Production ofa1,3-galactosyltransferase-deficient pigs. Science 2003;299: 411.
18. Kolber-Simonds D, Lai L, Watt SR et al. a1,3-galact-osyltransferase null pigs via nuclear transfer with fibro-blasts bearing loss of heterozygosity mutations. Proc NatlAcad Sci USA 2004; 19: 7335.
19. Sablinski T, Latinne D, Gianello P et al. Xenotrans-plantation of pig kidneys to nonhuman primates. I.Development of the model. Xenotransplantation 1995; 2:264.
20. Pennington LR, Sakamoto K, Popitz-Bergez FA et al.Bone marrow transplantation in miniature swine. I.Development of the model. Transplantation 1988; 45: 21.
21. Buhler L, Goepfert C, Kitamura H et al. Porcine he-matopoietic cell xenotransplantation in nonhuman pri-mates is complicated by thrombotic microangiopathy.Bone Marrow Transplant 2001; 27: 1227.
22. Alwayn IPJ, Buhler L, Appel JZ III et al. Mechanism ofthrombotic microangiopathy following xenogeneic hema-topoietic progenitor cell transplantation. Transplantation2001; 71: 1601.
23. Appel JZ Jr, Alwayn IPJ, Buhler L et al. Modulation ofplatelet aggregation in baboons: implications for mixedchimerism in xenotransplantation. 1. The roles of indi-vidual components of a transplantation conditioningregimen and of pig peripheral blood progenitor cells.Transplantation 2001; 72: 1299.
24. Varas F, Grande T, Ramirez A, Bueren JA. Implan-tation of bone marrow beneath the kidney capsule resultsin transfer not only of functional stroma but also ofhematopoietic repopulating cells. Blood 2000; 96: 2307.
25. Bingaman AW, Waitze S-G, Alexander DZ. Trans-plantation of the bone marrow microenvironment leads to
hematopoietic chimerism without cytoreductive condi-tioning. Transplantation 2000; 69: 2491.
26. Dor FJMF, Tseng Y-L, Kuwaki K, Ko DSC, Cooper
DKC. Pig spleen transplantation induces transient hema-topoietic cell chimerism in baboons (Letter). Xenotrans-plantation 2004; 11: 298.
27. Basker M, Alwayn IPJ, Buhler L et al. Clearance ofmobilized porcine peripheral blood progenitor cells isdelayed by depletion of the phagocytic reticuloendothelialsystem in baboons. Transplantation 2001; 72: 1278.
28. Fraser CC, Chen BP, Webb S, Van RN, Kraal G.Circulation of human hematopoietic cells in severe com-bined immunodeficient mice after C12MDP-liposome-mediated macrophage depletion. Blood 1995; 86: 183.
29. Abe M, Cheng J, Qi J et al. Elimination of porcine he-matopoietic cells by macrophage in mice. J Immunol 2002;168: 621.
30. Dor FJMF, Rouhani FJ, Cooper DKC. Transfusion ofpig red blood cells into baboons (Letter). Xenotransplan-tation 2004; 11: 295.
31. Rouhani FJ, Dor FJMF, Cooper DKC. Investigation oferythrocytes from a1,3-galactosyltransferase gene-knock-out pigs for human blood transfusion. Transfusion Inpress.
32. Kuwaki K, Knosalla C, Dor FJMF et al. Suppressionof natural and elicited antibodies in pig-to-baboon hearttransplantation using a human anti-CD154 monoclonalantibody-based regimen. Am J Transplant 2004; 4: 363.
33. Knosalla C, Ryan D, Moran K et al. Initial experiencewith the human anti-CD154 monoclonal antibody,ABI793, in pig-to-baboon xenotransplantation. Xeno-transplantation In press.
Tseng et al.
370