biosurfactants have the potential to induce defence against phytophthora infestans in potato
TRANSCRIPT

Biosurfactants Have the Potential to InduceDefence Against Phytophthora infestansin Potato
Therese Bengtsson & Anna Holefors &Erland Liljeroth & Malin Hultberg &
Erik Andreasson
Received: 4 December 2013 /Accepted: 22 January 2015# European Association for Potato Research 2015
Abstract Potato late blight caused by Phytophthora infestans is one of the mostdestructive plant diseases worldwide. Currently, its management mainly relies on thefrequent use of synthetic chemicals, and there is a need to develop more sustainablestrategies. Biosurfactants produced by fluorescent pseudomonads have been shownto rapidly, within 1 min, cause zoospore lyses. This study investigated if thebiosurfactant-producing strain Pseudomonas koreensis 2.74 and its biosurfactantshave the potential to induce resistance in potato to late blight. The experiments wereperformed with treatments of whole plants of the susceptible potato cultivar Bintjeand the partially resistant cultivar Ovatio. A significant disease reduction and aninduced secretion of proteins such as pathogenesis-related (PR) protein 1, within theleaf apoplast, were observed in the potato cultivar Ovatio after treatment with thebiosurfactant at a concentration of 1 mg ml−1. No significant effects on diseasedevelopment were observed after treatment with the bacterial suspension ofPseudomonas koreensis 2.74.
Keywords Apoplast . Induced resistance . Late blight . PR-1 . Secretome .
Solanum tuberosum
Potato ResearchDOI 10.1007/s11540-015-9286-4
T. Bengtsson (*) : E. Liljeroth : E. AndreassonDepartment of Plant Protection Biology, Swedish University of Agricultural Sciences, Alnarp, Swedene-mail: [email protected]
A. HoleforsIn Vitro Plant-Tech AB, Limhamn, Sweden
M. HultbergDepartment of Biosystems and Technology, Swedish University of Agricultural Sciences, Alnarp,Sweden

Introduction
Potato late blight disease caused by the oomycete Phytophthora infestans (Mont.) deBary is one of the most destructive plant diseases worldwide. Plant disease control inagricultural systems today relies heavily on pesticides, and the main managementoption to control late blight in susceptible and moderately resistant cultivars is thefrequent application of fungicides. The number of resistant potato cultivars is limited sofar, and P. infestans has a remarkable capacity to overcome plant resistance (Fry 2008).Furthermore, as regards climate change, P. infestans is expected to be favoured in anyfuture milder and more humid climate (Roos et al. 2011). It is therefore of greatimportance to find new and environmentally sound alternatives to control late blight.
During recent years, there has been increasing interest in the use of antagonisticmicroorganisms as biocontrol agents to provide a sustainable alternative to pesticides.A well-known group of microorganisms used for biocontrol is the fluorescent pseudo-monads. They are indigenous in the plant environment and produce and excretesecondary metabolites that are inhibitory to plant pathogens. Important groups ofsecondary metabolites produced by fluorescent pseudomonads include antibiotics andbiosurfactants (Haas and Defago 2005). Fluorescent pseudomonads capable of produc-ing biosurfactants have recently received increased attention for biocontrol of zoospore-producing pathogens (D’Aes et al. 2010; Tran Thi Thu 2007). Several studies haveshown that the zoospore lyses within a short time, less than 1 min, after exposure tobiosurfactants (De Souza et al. 2003; Stanghellini and Miller 1997). In addition to thedirect lytic effect of the biosurfactant on the zoospores, it has also been reported thatbiosurfactants have the capacity to induce resistance in the plant. Tran et al. (2007)found that treatment with Pseudomonas fluorescens SS101 or its biosurfactant inducedresistance against P. infestans in tomato, both systemically and locally. Also, thebiosurfactant used in a grapevine study by Varnier et al. (2009) showed the ability toinduce resistance and to prime the defence prior to a second challenge.
Considering the importance of zoospores in the spread of late blight and the potentialto induce resistance in the potato plant, biosurfactants and biosurfactant-producingstrains are of interest for biocontrol of late blight. In a previous study based on detachedleaf assays, we observed suppression of late blight when the biosurfactant-producingstrain Pseudomonas koreensis 2.74 or its biosurfactant were applied to potato leaflets(Hultberg et al. 2010).
The aim of the present study was to further investigate the potential for suppressionof late blight, using intact plants in order to mimic natural conditions more accurately,and to examine whether induced defence responses could be observed in the plant aftertreatment.
Material and Methods
The late blight-susceptible potato cultivar Bintje and the partially resistant cultivarOvatio (Liljeroth et al. 2010), both obtained from Agrico Nordic AB (Jönköping,Sweden), were grown in a biotron under controlled conditions with a 16-h/8-h day/night regime and light intensity of 300 μmol m−2 s−1. The temperature was set to 20 °Cand relative humidity to 65%. Bacterial suspension and biosurfactant solution were
Potato Research

prepared from P. koreensis 2.74 (CBS 125413), as described by Hultberg et al. (2010).In both experiments, a biosurfactant concentration of 1 mg ml−1 and a bacterialsuspension with an optical density of 1.0 at 620 nm were used. When the plantsreached an age of 4 weeks, they were treated with the bacterial suspension (Bact) orthe biosurfactant solution (Ten). Plants treated with water (Contr) were used as controls.The treated plants were divided into two experimental units. One experimental unit waskept under the conditions described above and was not infected with P. infestans. Thisis in order to examine any potential phytotoxic effects of the treatments, and if induced,defence responses could be observed in uninfected plants after treatment with thebiosurfactant or bacterial suspension. The other experimental unit was moved to aninfection chamber with 100% relative humidity. After 24 h in the infection chamber,each pot was sprayed with 5 ml of the inoculum consisting of 2.0×104 sporangia ml−1
of P. infestans (Mont.) de Bary isolate SE03058 (mating type A1, virulence 1, 3, 4, 7,10, 11) and kept in the dark for 15 h (temperature 17 °C, relative humidity 100%). Forthe rest of the experiment, the temperature in the infection chamber was kept at 19 °Cduring the day and 17 °C at night, with a light regime of 10 h/14 h, respectively, and arelative humidity of 70%. The light intensity was set to 70 μmol m−2 s−1. Theexperiment was performed with three replicate samples and repeated once. The datawere analysed by two-way ANOVAwith factors cultivar and treatment. The differenceswithin cultivars with respect to effect of the treatments were analysed by one-wayANOVA followed by Tukey’s multiple comparison test, with differences consideredsignificant at p<0.05 (Minitab, version 15).
For quantification of P. infestans colonisation 6 days after infection, quantitative real-time PCR was performed using the Bio-Rad CFX96™ Real-Time PCR system (Bio-Rad Laboratories, Inc.) as previously described by Bengtsson et al. (2013). In parallel,an assessment of the necrotic lesion sizes caused byP. infestanswas carried out using thesoftware Assess 2.0 (APS Press, St. Paul, MN). Ten leaflets, randomly sampled withineach plant, were scanned in a flatbed scanner (Epson Perfection V750), and the fileswere imported to Assess for quantification of the percentage of affected leaflet area.
Four days after infection, four leaves from the fully expanded composite leaves inthe central compartment of the shoots on cv. Ovatio were chosen for analysis ofapoplastic proteins. The leaves were washed in 1% Tween 20 for 30 s to decreasesurface tension as described in Ali et al. (2012). They were then carefully dried, placedin a Petri dish containing infiltration buffer (50 mmol l−1 Na2HPO4, pH 7.0 and150 mmol l−1 NaCl) and vacuum-infiltrated for 3×5 min. To collect the apoplast, theleaves were centrifuged at 2100g for 3 min at 4 °C in a 15-ml Falcon tube containing3 μl of a protease inhibitor cocktail (Sigma, P9599) and a metal ring for separating theleaves from the supernatant.
To analyse the secreted proteins within the apoplast, a 14% SDS polyacrylamide gelwas run. Six μl of a 6× SDS sample buffer containing DTT was added to 30 μl ofsample prior to denaturation at 95 °C for 5 min. The gel was run for 20 min at 80 V,followed by 1.5 h at 100 V. For visualisation of the bands, the gel was stained overnightin Coomassie Brilliant Blue (10% (v/v) acetic acid, 45% (v/v) methanol, 0.25% (w/v)Coomassie R-250) and then briefly washed in dH2O before 3 h in destaining solution(10% (v/v) acetic acid and 40% (v/v) methanol).
Pathogenesis-related (PR) protein-1 bands (14 kDa) were quantified by the ImageJprogram and identified with mass spectrometry by cutting out the band and subsequent
Potato Research

subjection to in-gel tryptic digestion. In brief, the gel pieces were destained and washedand after dithiothreitol reduction and iodoacetamide alkylation, the proteins weredigested with trypsin (modified sequencing grade; Promega) overnight at 37 °C. Thegel pieces were shaken vigorously at room temperature for 15 min, and 0.5 μl of eachsample was spotted directly onto a stainless steel MALDI target and was left to dry. Avolume of 0.5 μl of a matrix solution containing 5 mg/ml α-cyano-4-hydroxy cinnamicacid, 50% acetonitrile, 0.1% TFA and 50 mM citric acid was added and allowed to dry.MALDI-TOF MS and MS/MS spectra were recorded automatically using a 4700Proteomics Analyzer (Applied Biosystems, Framingham, CA, USA). Protein identifi-cation was performed using the GPS Explorer software, with an in-house Mascotsearch engine (Matrix Science, London, UK). The following parameters were appliedfor database-search: database (The Universal Protein Resource (UniProt)), taxonomy(Solanum tuberosum), max missed cleavage (1). A protein was considered as identifiedwhen the level of confidence for the total ion score exceeded 99.95%.
Results
For the infected plants, the amount of P. infestans biomass and the percentage leafletarea occupied by lesions were significantly higher in cv. Bintje than in cv. Ovatio(biomass, p<0.001; lesion, p<0.0001). The two-way ANOVA analysis also showed asignificant reduction in the amount of biomass of P. infestans by the biosurfactanttreatment (p=0.01). However, no significant effect of the biosurfactant treatment on thepercentage area of necrosis could be found (p=0.06). From the qPCR, we could notshow significant effects of the treatments in infected leaves of cv. Bintje (p=0.09), evenwith two independent experimental set-up with three samples in each group. However,the average values for pathogen colonisation of the plants treated with biosurfactant orbacterial suspension were lower compared with the water control (Fig. 1). For the cv.Ovatio, a significant reduction in pathogen colonisation was observed after treatmentwith the biosurfactant (p=0.04) (Fig. 1).
Assessment of the percentage of infected leaflet area of the cv. Bintje showed thatapproximately one third of the total leaflet area was infected on the water-treated plants(Fig. 2). No significant effects of treatment with the biosurfactant or bacterial suspen-sion could be observed on this cultivar (p=0.3). For cv. Ovatio, treatment with the
Fig. 1 Biomass of Phytophthora infestans in potato leaves in the different treatments, measured by qPCR.Bars for a particular cultivar followed by different letters are significantly different (p<0.05, Tukey’s test).Mean and standard error are shown, n=3. Contr control, Ten biosurfactant, Bact Pseudomonas koreensis 2.74
Potato Research

biosurfactant significantly reduced percentage infected area compared with the watertreatment (p=0.02) (Fig. 2).
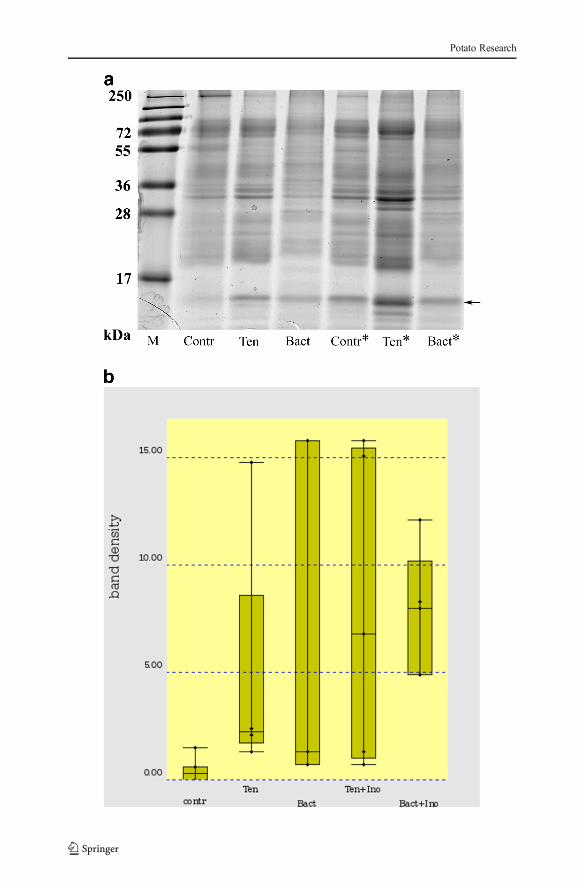
In order to observe changes in the apoplastic PR-1 level after treatment with thebacteria suspension or the biosurfactant solution, the secretome from the cv. Ovatio wasextracted 4 days post inoculation as described in Moushib et al. (2013). The results arepresented in Fig. 3 and in the 14-KDa band referred to as PR-1; we identified the peptideAQVGVGPMSWDAGLASR that was found in two different PR-1 forms in potato(Ali et al. 2012). As a measure of contamination, the rubisco protein at approximately50 kDa was monitored, and it was barely possible to discern a band of that size. For thenon-infected plants kept in the original chamber, a low basal level of the defence markerprotein PR-1 was observed in the apoplast (Fig. 3b). The PR-1 level increased in thebiosurfactant-treated and bacteria-treated plants both before and after infection (Fig. 3b).Thus, a direct effect of defence response was seen after both treatments.
Discussion
This study shows that it is possible to obtain significant reduction in development inlate blight after plant treatment with a biosurfactant. Biosurfactants are generally knownto be less toxic compared with synthetic surfactants and rapidly degraded in theenvironment (Lima et al. 2011; Mulligan 2005). However, phytotoxicity due to leafapplications of certain surfactants has been reported (Bukovac 2005). The biosurfactantused in the present study was tested on uninfected plants, and no signs of phytotoxicitycould be observed with either of the cultivars. Thus, biosurfactants have potential to bedeveloped as a sustainable measure for future management of late blight.
A main difference between the cultivars included in the study is their susceptibility tolate blight as demonstrated in an earlier study (Liljeroth et al. 2010), with cv. Ovatiobeing a partially resistant cultivar and cv. Bintje, a susceptible cultivar (Liljeroth et al.2010). A difference was also observed in the present as well as in our previous study(Hultberg et al. 2010), where a significantly higher degree of late blight was observed oncv. Binje compared with cv. Ovatio. The aggressive disease development on cv. Bintjecould be an explanation for the lack of a significant effect of the biosurfactant treatment.However, in our previous study based on a detached leaf assay, a significant effect of thetreatments was evident in cv. Bintje as well as in cv. Ovatio. In this study, a tendency forreduction of late blight although not significant could be seen in cv. Bintje, and it cannot
Fig. 2 Late blight lesions expressed as percentage of leaflet area for the different treatments. Bars for aparticular cultivar followed by different letters are significantly different (p<0.05, Tukey’s test). Mean andstandard error are shown, n=3. Contr control, Ten biosurfactant, Bact Pseudomonas koreensis 2.74
Potato Research
Potato Research

be ruled out that this would be significant if more experiments were carried out. Potatolate blight is a devastating disease and its management should be based on severalstrategies as discussed by Mizubuti et al. (2007). It is evident that the use ofbiosurfactants cannot be considered a stand-alone method for its control. Instead,biosurfactant-based products should be incorporated into an integrated programmebased on several measures, including resistant or partly resistant cultivars.
The results show that treatment with the purified biosurfactant had a greaterprotective effect against late blight than treatment with the biosurfactant-producingstrain. This is in contrast to findings in our previous study based on detached leaf assay,where a significant decrease of leaf lesion was also observed after treatment with thebacterial suspension (Hultberg et al. 2010). A major difference between the two studiesis that in the present study, intact plants with more intact physiology were infected. Alow production of biosurfactants by P. koreensis 2.74 when introduced into thephyllosphere could be responsible for the lower disease repression compared with apure biosurfactant. The phyllosphere is not a native environment for this strain since itwas isolated from the rhizosphere (Hultberg et al. 2008). In a previous large-scalebiocontrol study, performed on hydroponically cultivated lettuce, where a bacterialsuspension of P. koreensis 2.74 was introduced into the rhizosphere, the protectiveeffect of addition of a bacterial suspension equalled the effect of the purifiedbiosurfactant (Hultberg et al. 2011). It should be pointed out that a biocontrol systembased on living, biosurfactant-producing microorganisms is a less costly and also moresustainable approach compared with the use of the extracted pure compound. This is ofimportance especially taking the fast biodegradation of the biosurfactants (Lima et al.2011) into account and, thereby, the need for frequent additions.
Biosurfactants, applied either as pure compounds or further developed and applied byusing producing strains, have promise for use in dampening the rapid spread of diseasein infected fields due to direct surfactant effect on the zoospores. Less effect can beexpected on plant infection through direct germination of the sporangia since lowertoxicity of biosurfactants on structures others than zoospores have been reported (van deMortel et al. 2009). In this study, increased levels of PR-1 were observed in thebiosurfactant-treated and bacteria-treated plants both before and after infection. Thisfurther strengthens the observations reported in a review by Raaijmakers et al. (2010).There, they report that cyclic lipopeptides produced by Bacillus and Pseudomonas spp.not only can exert disease suppressive effects by direct action on the zoospores but alsohave the capability to induce classical defence responses in the plant.
Acknowledgements This study was supported by grants from the Swedish Research Council for Environ-ment, Agricultural Sciences and Spatial Planning (FORMAS), Swedish Foundation for Strategic Research,Partnership Alnarp, Swedish Potato Research Alnarp and the foundation Stina Werner Fond, which aregratefully acknowledged. We are also grateful to Göran Nilsson, Biotron, Alnarp, for help with plantcultivation.
�Fig. 3 a SDS-PAGE of proteins secreted in the leaf apoplast of cv. Ovatio. Starting from the left, lane 1, PageRuler SM1811 (Fermentas); lanes 2–4, uninfected samples; lanes 5–7, infected samples (*). PR-1 wasidentified in the 14-kDa band (arrow) by MALDI-TOF mass spectrometry. M marker, Contr control, Tenbiosurfactant, Bact Pseudomonas koreensis 2.74. b Box-plot of ImageJ processed PR-1 bands, n=4–7 for eachsample group. contr control, Ten biosurfactant, Bact Pseudomonas koreensis 2.74, Ino inoculation withPhytophthora infestans
Potato Research

References
Ali A et al (2012) Paranoid potato: phytophthora-resistant genotype shows constitutively activated defense.Plant Signal Behav 7:400–408
Bengtsson T, Holefors A, Witzell J, Andreasson E, Liljeroth E (2013) Activation of defence responses toPhytophthora infestans in potato by BABA. Plant Pathol. doi:10.1111/ppa.12069
Bukovac MJ (2005) Maximizing performance of plant growth regulators by improving spray application.Horttechnology 15:222–231
D'Aes J, De Maeyer K, Pauwelyn E, Höfte M (2010) Biosurfactants in plant—Pseudomonas interactions andtheir importance to biocontrol. EnvironMicrobiol Rep 2:359–372. doi:10.1111/j.1758-2229.2009.00104.x
De Souza JT, De Boer M, De Waard P, Van Beek TA, Raaijmakers JM (2003) Biochemical, genetic, andzoosporicidal properties of cyclic lipopeptide surfactants produced by Pseudomonas fluorescens. ApplEnviron Microbiol 69:7161–7172
Fry W (2008) Phytophthora infestans: the plant (and R gene) destroyer. Mol Plant Pathol 9:385–402. doi:10.1111/j.1364-3703.2007.00465.x
Haas D, Defago G (2005) Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat RevMicrobiol 3:307–319
Hultberg M, Bergstrand KJ, Khalil S, Alsanius B (2008) Characterization of biosurfactant-producing strains offluorescent pseudomonads in a soilless cultivation system. Anton Leeuw Int J Gen Mol Microbiol 94:329–334. doi:10.1007/s10482-008-9250-2
Hultberg M, Bengtsson T, Liljeroth E (2010) Late blight on potato is suppressed by the biosurfactant-producing strain Pseudomonas koreensis 2.74 and its biosurfactant. BioControl (Dordrecht) 55:543–550. doi:10.1007/s10526-010-9289-7
Hultberg M, Holmkvist A, Alsanius B (2011) Strategies for administration of biosurfactant-producingpseudomonads for biocontrol in closed hydroponic systems. Crop Prot 30:995–999. doi:10.1016/j.cropro.2011.04.012
Liljeroth E, Bengtsson T, Wiik L, Andreasson E (2010) Induced resistance in potato to Phytophthorainfestans—effects of BABA in greenhouse and field tests with different potato varieties. Eur J PlantPathol 127:171–183. doi:10.1007/s10658-010-9582-4
Lima TM, Procopio LC, Brandao FD, Carvalho AM, Totola MR, Borges AC (2011) Biodegradability ofbacterial surfactants. Biodegradation 22:585–592. doi:10.1007/s10532-010-9431-3
Mizubuti ESG, Lourenco VJ, Forbes GA (2007) Management of late blight with alternate products. PestTechnol 1:106–116
Moushib L, Witzell J, Lenman M, Liljeroth E, Andreasson E (2013) Sugar beet extract induces defenceagainst Phytophthora infestans in potato plants. Eur J Plant Pathol 136:261–271. doi:10.1007/s10658-012-0160-9
Mulligan CN (2005) Environmental applications for biosurfactants. Environ Pollut 133:183–198. doi:10.1016/j.envpol.2004.06.009
Raaijmakers JM, De Bruijn I, Nybroe O, Ongena M (2010) Natural functions of lipopeptides from Bacillusand Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev 34:1037–1062. doi:10.1111/j.1574-6976.2010.00221.x
Roos J, Hopkins R, Kvarnheden A, Dixelius C (2011) The impact of global warming on plant diseases andinsect vectors in Sweden. Eur J Plant Pathol 129:9–19. doi:10.1007/s10658-010-9692-z
Stanghellini ME, Miller RM (1997) Their identity and potential efficacy in the biological control of zoosporicplant pathogens. Plant Dis 81:4–12
Tran Thi Thu H (2007) Interactions between biosurfactant-producing Pseudomonas and Phytophthora speciesTran H, Ficke A, Asiimwe T, Hoefte M, Raaijmakers JM (2007) Role of the cyclic lipopeptide massetolide A
in biological control of Phytophthora infestans and in colonization of tomato plants by Pseudomonasfluorescens. New Phytol 175:731–742. doi:10.1111/j.1469-8137.2007.02138.x
van de Mortel JE, Tran H, Govers F, Raaijmakers JM (2009) Cellular responses of the late blight pathogenPhytophthora infestans to cyclic lipopeptide surfactants and their dependence on G proteins. ApplEnviron Microbiol 75:4950–4957
Varnier A-L et al (2009) Bacterial rhamnolipids are novel MAMPs conferring resistance to Botrytis cinerea ingrapevine. Plant Cell Environ 32:178–193
Potato Research