biologie des populations de tourterelles à queue carrée sur l'île de la barbade apports de...
TRANSCRIPT

UNIVERSITE DE BOURGOGNE
UFR Sciences Vie Terre Environnement
Ecole Doctorale Environnements - Santé - STIC (n°490)
THESE POUR OBTENIR LE GRADE DE
DOCTEUR DE L’UNIVERSITE DE BOURGOGNE
Discipline : Sciences de la Vie
par Karine MONCEAU
le 21 décembre 2009
Biologie des populations de tourterelles à queue carrée
sur l'île de la Barbade
Apports de l'outil moléculaire
Directeur de thèse : Pr. Frank CEZILLY
Co-directeur de thèse : Dr. François-Xavier DECHAUME-MONCHARMONT
JURY
Dominique ALLAINE Rapporteur Professeur, Université Lyon I
Loïc BOLLACHE Examinateur Professeur, Université de Bourgogne
Frank CEZILLY Examinateur Professeur, Université de Bourgogne
François-Xavier
DECHAUME-MONCHARMONT Examinateur Maître de Conférences, Université de Bourgogne
Olivier DEHORTER Examinateur Ingénieur de Recherche CNRS, CRBPO
Philipp HEEB Rapporteur Chargé de Recherche CNRS, HDR, Université Toulouse III

REMERCIEMENTS
En premier lieu, je souhaiterais remercier Frank Cézilly, pour m’avoir fait confiance sur un
sujet qui, je le sais, lui tenait particulièrement à cœur. Je tiens à lui témoigner ici toute ma gratitude
pour son encadrement aussi bien à Dijon qu’à la Barbade (et aussi pour le resto de pierrade où il m’a
amené les jours où le manque de protéines se faisait sentir sur le terrain).
Merci également à François-Xavier Dechaume-Moncharmont (ouf ! ça a tout de même était
compliqué de faire tenir ton nom sur la liste du jury sans que ça dépasse trop !) pour sa disponibilité et
son encadrement. Il m’a également permis de découvrir le monde fabuleux de R dont je ne peux plus
me passer maintenant.
Je remercie tout particulièrement Philipp Heeb et Dominique Allainé pour avoir accepté d’être
rapporteur de mes travaux. Je les remercie d’autant plus qu’ils m’ont tous deux accordés un sursis
réellement nécessaire pour boucler le manuscrit, tout en sachant que de leur côté, cela leur réduisait
leur temps de correction, un temps qui je le sais bien est précieux pour des personnes aussi occupées.
Merci également à Loïc Bollache et Olivier Dehorter d’avoir accepté de participer à mon jury
de thèse et d’examiner mes travaux.
Un grand merci également à Thierry Rigaud, avec qui j’ai beaucoup appris en M2, notamment
à ne pas paniquer lorsque les expériences réalisées ne donnent que des résultats négatifs. Finalement,
cela a été très instructif, même si je sais que le déroulement de mon stage l’avait, à l’époque, quelque
peu stressé. Je ne regrettte certes pas ces satanés microsporidies mais encore moins d’avoir travaillé
sous la direction de quelqu’un qui est pour moi, un grand Monsieur.
Je voudrais également remercier Rémi Wattier (Mr Génétique) pour son soutien, sa
disponibilité et sa bonne humeur.
Merci également à Coraline Caullet, Christine Dubreuil, Maria Gaillard et Julia Geraci pour
leur aide et/ou conseils en génétique et plus particulièrement face au redoutable Licor, qui nous en
aura fait voir de toutes les couleurs.
Je remercie également Morgan David, Maria Gaillard (bis), Estelle Harrang, Sébastien
Motreuil, Jérôme Moreau et George Prato pour leur aide précieuse sur le terrain à la Barbade, avec
pour certains beaucoup d’anicroches techniques durant les trajets !

Merci également à l’équipe canadienne : Laure Cauchard, Kimberley-Anne Côté et Sarah
Overington pour les bons moments passés à la Barbade et à Neeltje Boogert pour les échanges sur les
tourterelles, dommage que nous ne nous soyions jamais croisées là-bas. J’en profite également pour
remercier Louis Lefebvre sans qui Frank n’aurait peut être jamais commencé à travailler sur les
tourterelles à queue carrée.
Je remercie également mes stagiaires, encadrés sur les thématiques liées aux tourterelles :
Aurore Barat, Eloise Colin, Gwladys Doyen et Amélie Slaski ou sur d’autres sujet : Jérémie Cornuau
et Matthias Galipaud (je fais grâce de leur surnom). En tout cas, j’espère qu’ils auront tous gardé un
petit quelque chose que j’aurais pu leur transmettre. En ce qui me concerne cela aura été un véritable
plaisir de travailler avec eux.
Je remercie vivement Emilie Arnoux, Alexandre Bauer, Jean-Sébastien Bolduc, Marie-Jeanne
Perrot-Minnot, Yannick Moret, Jean-Philippe Troussard ainsi que Nathalie Franceschi, Nicolas
Kaldonski et tous les membres précédemment cités de l’équipe Ecologie Evolutive de l’UMR CNRS
5561 Biogéosciences pour leur soutien et leur amitié.
Merci également à tous les étudiants en M1 et M2 que j’ai pu côtoyer ces trois dernières
années et qui contribuent également à la bonne ambiance générale. Désolée de ne pas tous les citer
mais la liste serait trop longue.
Je remercie également les familles Bolduc, Bouvier-Kaldonski, Cézilly, Dechaume-
Moncharmont, Moret, Martinaud, Moreau, Rigaud et Troussard pour tous les bons moments partagés
autour de bonnes victuailles.
Merci également à Vivi, Tauta et Vince car même si on ne se voit pas souvent vous êtes
toujours là.
Je ne remercierais jamais assez mes parents pour m’avoir permis d’aller aussi loin, ainsi que
ma famille et mes proches dont le soutien indéfectible m’a toujours aidé à avancer.
Et la dernière personne que je souhaiterais remercier est Jérôme Moreau qui entre, en fait,
dans plusieurs cases : celle de collègue, d’ami, de famille et surtout de compagnon. Toutes ces
casquettes n’ont sans doute pas dues être facile à porter compte tenu de mon caractère, mais il n’a
jamais raccroché et reste toujours présent. Je lui témoigne ici toute ma gratitude et mon amour.

SOMMAIRE
PREAMBULE 1
CHAPITRE I :
PRESENTATION DU MODELE D ’ETUDE ET DES OUTILS METHODOLOGIQUES 9
I. INTRODUCTION 10
II. METHODE GENERALE : MODELE BIOLOGIQUE ET ACQUISITION DES DONNEES 12
1. La tourterelle à queue carrée, Zenaida aurita 12
2. Capture, acquisition des données morphométriques et prélèvements sanguins 15
3. Préparation des échantillons sanguins pour les analyses moléculaires subséquentes :
l’extraction d’ADN 17
III. SEXAGE ET DIMORPHISME SEXUEL CHEZ LA TOURTERELLE A QUEUE CARREE : LES
PROBLEMES METHODOLOGIQUES 18
1. Sexage moléculaire 18
a. Généralités sur le sexage chez les oiseaux 18
b. Méthodes de sexage moléculaire 19
i. Généralités 19
ii. Mise en place du sexage moléculaire chez la tourterelle à queue
carrée 20
2. Sexage via l’utilisation de données morphométriques : les biais
méthodologiques 26
IV. ETUDE DE LA VARIABILITE INTERINDIVIDUELLE : MISE AU POINT DES MARQUEURS
MICROSATELLITES 29

CHAPITRE II :
POLYMORPHISME DE RESSOURCES CHEZ LA TOURTERELLE A QU EUE
CARREE 32
CHAPITRE III :
HETEROZYGOTIE MULTI -LOCI ET APTITUDE PHENOTYPIQUE 36
I. INTRODUCTION 37
II. MATERIEL & METHODES 41
1. Modèle biologique 41
2. Site d’études, captures, prises de mesures et prélèvements sanguins 42
3. Analyses génétiques 43
a. Généralités 43
b. Analyses préliminaires 43
c. Mesures de l’hétérozygotie 44
4. Relation entre hétérozygotie et aptitude phénotypique 45
a. Hétérozygotie, stratégie d’approvisionnement et âge (adultes et juvéniles) 45
b. Hétérozygotie et asymétrie fluctuante (adultes uniquement) 45
i. Analyses préliminaires 46
α. Erreur de mesure et asymétrie 46
β. Antisymétrie (AS) 47
γ. Asymétrie directionnelle (AD) 49
δ. Indépendance de la différence droite - gauche vis-à-vis de la taille
des individus 49
ii. Test de la relation entre hétérozygotie et asymétrie fluctuante 49
5. Logiciels de traitement des données 49
III. RESULTATS 50
1. Analyses génétiques 50
a. Analyses préliminaires 50
b. Mesure de l’hétérozygotie 52

2. Relation entre hétérozygotie et aptitude phénotypique 53
a. Hétérozygotie, stratégie d’approvisionnement et âge 53
i. Hétérozygotie et stratégie d’approvisionnement 53
ii. Hétérozygotie et âge 53
b. Hétérozygotie et asymétrie fluctuante 55
i. Erreur de mesure et asymétrie 55
ii. Antisymétrie (AS) 55
iii. Asymétrie directionnelle (AD) 58
iv. Indépendance de la différence droite-gauche vis-à-vis de la taille des
individus 58
v. Test de la relation entre hétérozygotie et asymétrie fluctuante 59
α. Chez les mâles 59
β. Chez les femelles 62
IV. DISCUSSION 65
DISCUSSION GENERALE ET PERSPECTIVES D’ETUDES 70
REFERENCES BIBLIOGRAPHIQUES 77
ANNEXES 105

1
PREAMBULE

2
Le milieu insulaire
Etymologiquement proche du terme isolement, l’île est définie comme une étendue de
terre bordée de toutes parts d’eau. Le milieu insulaire, par extension de l’origine latine insula,
excite depuis toujours la curiosité et l’imaginaire des hommes par sa singularité. Dans
l’inconscient collectif, l’île est le symbole de l’isolement ultime, tel qu’il est décrit dans
Robinson Crusoe (Defoe 1719). Cette caractéristique majeure du milieu insulaire a également
été largement exploitée notamment pour des raisons de sécurité dans le cadre de détentions
(Alcatraz, le bagne de Cayenne). Les îles sont également souvent associées au terme
paradisiaque décrivant une végétation luxuriante et une faune riche et extrêmement spécifique
ou endémisme. Cette diversité n’a pas échappé à Darwin lors d’une étape dans l’archipel des
Galápagos durant son voyage sur le Beagle et a contribué à l’élaboration de sa théorie de
l’évolution dont nous célébrons cette année les cent cinquante ans (première édition de On the
Origin of Species Darwin 1859). Cependant, certains éléments manquaient pour asseoir sa
théorie de la sélection naturelle.
Du phénotype au génotype
D’un point de vue strictement mécanistique, la théorie de la sélection naturelle
énoncée par Darwin s’appuie sur des observations phénotypiques et manque de preuves
basales et plus précisément des bases génétiques qui, à l’époque, ne sont pas connues. Alors
que Mendel publie ses travaux sur l’hérédité en 1866, ceux-ci restent inconnus pour Darwin et
ce n’est qu’au début du XXème siècle, dans les années trente que des approches génétiques
sont proposées par Fisher, Haldane et Wright, accompagnées du développement de concepts
génétiques théoriques qui sont progressivement intégrés à la théorie de l’évolution de Darwin
(Powell 1987, Ayala & Fitch 1997). Néanmoins, la nature mathématique de ceux-ci n’est pas
abordable par tous les biologistes ce qui est par la suite pallié par la publication de Genetics
and the Origin of Species de Dobzhansky (1937) permettant de faire le lien entre généticiens
et évolutionnistes (Powell 1987, Ayala & Fitch 1997). Cette nouvelle théorie baptisée, théorie
synthétique de l’évolution, utilise une notion qui jusque là restait floue à savoir l’espèce. Bien
PREAMBULE

Préambule
3
que couramment employée, celle-ci n’est pas réellement délimitée (Mayr 1940). En effet,
depuis Aristote (environ 350 avant J.C.) jusqu’à Buffon (1749-1788) en passant par Linné
(1758), la classification du monde vivant est effectuée sur des critères morphologiques et/ou
anatomiques. Mayr (1942) propose alors une définition basant l’unité de l’espèce sur un
ensemble d’individus pouvant se reproduire entre eux et avoir une descendance féconde
(Borges 2005). Cette définition, couramment admise comme une définition biologique
classique, cache en fait sa dimension génétique (γενητικός du grec "apte à procréer"). Par la
suite, l’essor de la génétique, aussi bien d’un point de vue fondamental avec la découverte de
la structure de l’ADN (Watson & Crick 1953) que technique (toujours d’actualité), offre un
nouveau niveau d’analyse puisque qu’elle permet de mettre en évidence des différences et/ou
des similarités potentiellement silencieuses d’un point de vue morphologique (Dobzhansky
1973). Aujourd’hui, les apports de la génétique sont incontestables quelle que soit la
discipline considérée en biologie.
Le milieu insulaire : un laboratoire de l’évolution
Outre la publication des écrits de Darwin et celle des essais de son contemporain
Wallace (1869, 1876, 1880), peu de travaux impliquant le milieu insulaire ont été réalisés
jusqu’à la seconde moitié du XXème siècle. Ce n’est que depuis la publication de la théorie de
la biogéographie insulaire de MacArthur & Wilson (1967) qu’un regain d’intérêt pour ce
milieu se fait sentir, notamment depuis l’avancée des connaissances en matière de génétique
(Monge-Nájera 2008).
Globalement, les îles présentent une grande diversité d’âge, de taille, de forme, de
localisation et de niveau d’isolement ce qui confère à chacune d’elles une combinaison unique
de caractéristiques en termes écologiques (MacArthur & Wilson 1967, Cronk 1997, Whittaker
& Fernández-Palacios 2007). Considéré comme un système isolé et stable, le milieu insulaire
présente un taux d’endémisme important, voire dans certains cas spectaculaire. En 1973,
Dobzhansky évoquait par exemple l’existence d’environ 2000 espèces de drosophiles, dont un
quart d’entre elles étaient présentes sur l’archipel Hawaïen et parmi elles, 17 étaient
endémiques soit (0,85%) alors que cet archipel représente à peine 0,20% de la surface totale
des terres émergées (pour une vision actualisée voir Magnacca et al. 2008). Globalement,
81% des oiseaux, 91% des angiospermes 99% des insectes et des mollusques hawaïens
seraient endémiques de cet archipel (Sohmers & Gustafson 1993 cités dans Whittaker &

Préambule
4
Fernández-Palacios 2007). Le même cas s’observe aux Galápagos, évoquées précédemment,
où les 13 espèces de pinsons de Darwin (sous-famille des Geospizinae) sont inféodées à cet
archipel (une autre espèce étant présente sur Coco Island au nord de l’archipel) (Grant &
Grant 2008). Ceci ne signifie pas que ces espèces ont colonisé exclusivement ces milieux
mais qu’elles sont issues d’un ensemble de processus ayant pour conséquence une
diversification, voire une spéciation. Dans le cas des pinsons de Darwin, la radiation
adaptative observée a été confirmée génétiquement (marqueurs microsatellites et
mitochondriaux) comme résultant de la colonisation par un ancêtre commun (Petren et al.
1999, Sato et al. 1999, Grant & Grant 2008). Globalement, ce sont les conséquences du
niveau d’isolement et de la plus ou moins grande connectivité des îles qui intéressent les
biologistes évolutionnistes (Losos & Ricklefs 2009).
Dans ce cadre, certains archipels ont été plus étudiés que d’autres et sont considérés
comme des laboratoires de l’évolution en raison de la radiation endémique qu’ils présentent
(Whittaker & Fernández-Palacios 2007, Ricklefs & Bermingham 2008, Losos & Ricklefs
2009). Ceci est le cas par exemple des îles d’Hawaii, archipel le plus isolé des continents.
Situé dans la partie médiane de l’océan Pacifique, cet archipel héberge plusieurs milliers
d’espèces endémiques dont, entre autre, des drosophiles (Magnacca et al. 2008) mais
également des moucherons de la famille des Limoniidae (Nitta & O’Grady 2008). De même,
et peut être avec une valeur historique supplémentaire, les Galápagos ont fait l’objet de
beaucoup d’attention de la part des évolutionnistes et particulièrement pour les pinsons de
Darwin, étudiés depuis plus d’une trentaine d’années par P. et R. Grant et leur équipe de
l’université de Princeton (Grant & Grant 2008 pour une vue d’ensemble). Certains archipels,
à l’inverse de ceux-ci, ont bénéficié de moins d’attention, bien que présentant des
caractéristiques toutes aussi attrayantes.
L’arc Caribéen : un archipel peu étudié
Les Caraïbes forment un archipel de plus d’une centaine d’îles et îlets au large des
côtes ouest du continent américain entre Amérique du Nord et du Sud (figure 1).
Globalement, trois parties peuvent être distinguées (du Nord au Sud) : les Caraïbes du nord
(au large de la Floride incluant les Keys, les Bahamas et îles Turques-et-Caïques), les Grandes
Antilles (les quatre plus grandes îles Cuba, la Jamaïque, Hispaniola et Porto Rico) et les
Petites Antilles (une quarantaine d’îles délimitant l’arc caribéen). D’un point de vue

Préambule
5
historique, sa formation complexe offre une diversité géologique et écologique élevée (Dengo
& Case 1990, Donovan & Jackson 1994, tous deux cités dans Hedges 2001). Au Jurassique (-
170Ma), la division de la Pangée a créé un espace pour l’apparition ultérieure de la plaque
caribéenne au Crétacé où les îles de l’arc proto-antillais issues du volcanisme ont émergé
progressivement sans que ni l’ordre ni le moment exact de leur émergence ne soient connus.
A cette période, et sur environ 30 millions d’années, la plaque proto-antillaise est restée plus
ou moins connectée aux continents nord et sud américains (-100 à -70 Ma). Puis, au début du
Cénozoïque (-60 Ma), elle heurte et fusionne avec la plaque des Bahamas issue du continent
nord-américain et le tout glisse vers l’est. Depuis le Pliocène (environ -5 Ma), l’ensemble se
retrouve donc isolé vis-à-vis de la plaque continentale avoisinante dans une situation
géographique proche de celle que nous connaissons aujourd’hui. Contrairement à la partie
nord de l’archipel (Caraïbes nord et Grandes Antilles) où aucune activité volcanique n’est
connue, certains des volcans situés dans les Petites Antilles sont toujours actifs. Toutefois,
toutes les îles de cette zone géographique ne sont pas de type volcanique comme l’île de
Trinité issue d’un détachement du continent sud-américain ou encore la Barbade provenant
d’un soulèvement tectonique daté d’environ 700 000 ans (Speed 1994 cité dans Lovette et al.
1999).
D’un point de vue biologique, les champignons, protistes, bactéries, invertébrés et
plantes ne sont que peu ou pas étudiés contrairement aux vertébrés plus fortement suivis
notamment pour des aspects phylogéographiques, biogéographiques et de spéciation (Hedges
et al. 1992, Lovette et al. 1999, Hedges 2001, Pramuk et al. 2001, Rodriguez-Durán & Kunz
2001, Hower & Hedges 2003, Dávalos 2004, Ricklefs & Bermingham 2004, Rocca et al.
2004, Glor et al. 2005, Losos et al. 2006, Heinicke et al. 2007, Hedges et al. 2009).
En ce qui nous concerne, nous avons choisi de travailler sur un modèle biologique
vertébré, la tourterelle à queue carrée, Zenaida aurita, colombidé endémique des Caraïbes et
ce pour diverses raisons dont leur proximité à l’homme sur certaines îles. Notre
problématique initiale, consistant en l’étude de la fidélité et des liens du couple chez une
espèce monogame, ne trouve pas son intérêt dans un contexte insulaire en tant que tel mais
plutôt par rapport à la localisation de l’archipel à savoir la zone tropicale car finalement peu
d’études concernant cet axe de recherche s’y sont intéressées (Stutchbury & Morton 2001).
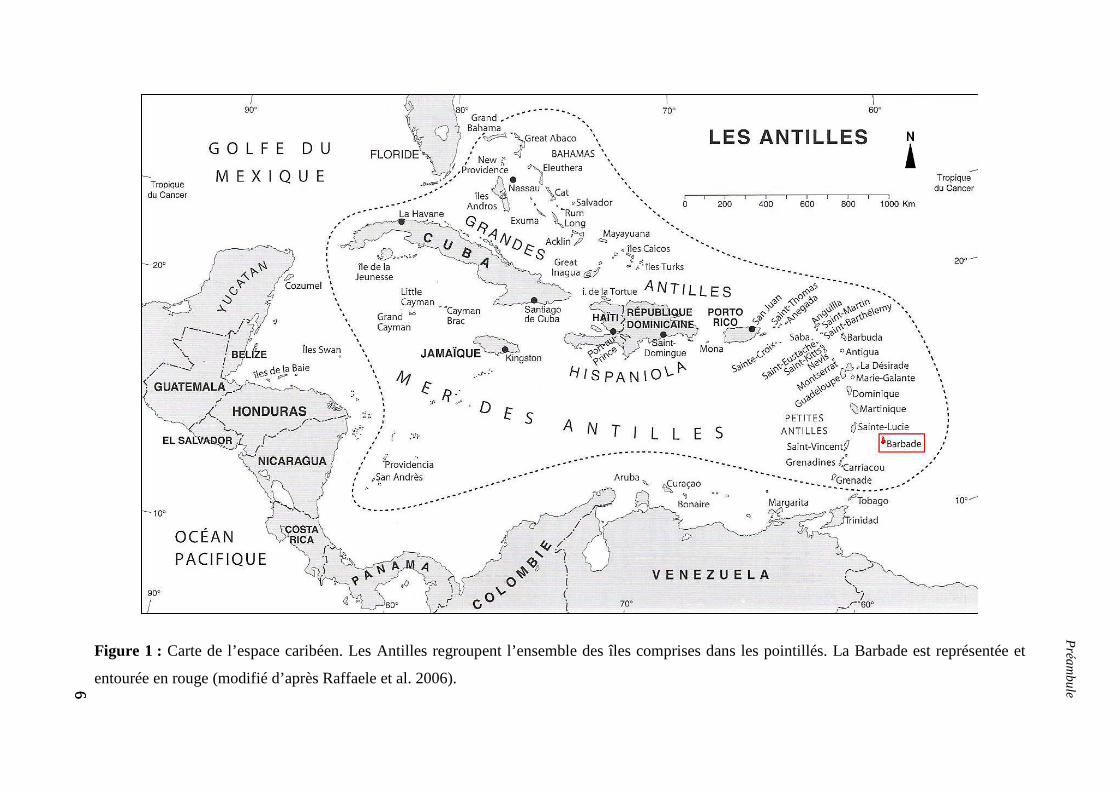
6
Figure 1 : Carte de l’espace caribéen. Les Antilles regroupent l’ensemble des îles comprises dans les pointillés. La Barbade est représentée et
entourée en rouge (modifié d’après Raffaele et al. 2006).
Pré
am
bu
le
6

Préambule
7
De la monogamie à des questions de biologie des populations
En écologie comportementale, un des enjeux majeurs est la compréhension des
systèmes d’appariement et notamment de la monogamie qui est somme toute assez singulière
dans le règne animal tant dans sa distribution taxonomique que dans son maintien évolutif.
Définie comme étant une association d’un mâle avec une femelle pour au moins un épisode
de reproduction, ce régime d’appariement apparait comme un paradoxe car d’un point de vue
adaptatif, les mâles ne devraient pas se restreindre qu’à une seule partenaire (voir Cézilly
2006 pour une revue détaillée de cette problématique). Chez les oiseaux, où la monogamie est
le régime prédominant, la plupart des études ont été réalisées en milieu tempéré où les saisons
rythment la reproduction ce qui ajoute à la difficulté de l’étude de la fidélité, un nouveau
paramètre de séparation des couples, à savoir les coupures potentielles temporaires ou
permanentes des liens entre partenaires. A l’inverse, en milieu tropical, les saisons sont
restreintes. Ainsi, compte tenu de la stabilité du milieu, les couples monogames peuvent
perdurer tout au long de l’année. De cette façon, les causes de séparations éventuelles
("divorces") pourraient être analysées plus aisément sans effet parasite ou du moins pas
imputable à des changements saisonniers. Toutefois, en dépit de ces avantages, peu de travaux
y ont été réalisés (Stutchbury & Morton 2001 pour un aperçu).
L’ambition du projet dans lequel s’inscrivait intialement cette thèse était donc placée
dans l’optique d’effectuer un suivi de couples après les avoir marqués par baguage. Ces
aspects comportementaux devaient constituer la majeure partie de mes travaux de thèse. Pour
cela, nous avons choisi de travailler à la Barbade (figure 1), notamment en raison du soutien
logistique dont la station biologique de Bellairs (Holetown, St James Parish) de l’Université
de McGill (Montréal, Canada) par notre collaboration avec le Pr. Louis Lefebvre (l’Université
de McGill). Cependant, bien que nous ayons effectué un baguage intensif lors de la première
session de capture en 2007 sur l’île de la Barbade (260 oiseaux bagués sur les territoires en
trois mois avec en moyenne deux personnes sur le terrain en permanence), le suivi des
couples s’est révélé plus difficile que ce qui était prévisible à l’origine. En effet, peu de
couples identifiés au cours de la première saison de terrain (six sur un total de 46) ont pu être
retrouvés lors de la session de capture suivante en 2008. De ce fait, le faible taux de
recaptures visuelles compromettait fortement l’obtention de données sur le taux de "divorce".
En parallèle de ce suivi comportemental et des analyses morphométriques sur les couples
bagués, des marqueurs microsatellites ont été développés afin d’analyser les critères

Préambule
8
d’appariement des individus sur des critères génétiques. Compte tenu du fait que nous avons
obtenu un grand nombre de marqueurs (23) avec un niveau minimum de polymorphisme
correct (au moins quatre allèles par locus), une analyse génétique a été associée à la
morphométrie afin d’envisager de nouvelles problématiques. De ce fait, les aspects liés au
régime d’appariement ne seront que peu développés (et même de façon plutôt indirecte au
travers du dimorphisme sexuel) dans le cadre de mes travaux de thèse, réorientée vers une
problématique articulée autour du lien entre génotype et phénotype.
Avant de rentrer dans le vif du sujet, l’étude chez cette espèce a nécessité dans un
premier temps, la mise au point de certaines techniques génétiques (dont les marqueurs
microsatellites et des marqueurs moléculaires d’identification du sexe) mais également une
mise au point technique pour l’exploitation des données morphométriques (chapitre I). Dans
l’ensemble de notre raisonnement, morphologie et génétique sont intimement liées l’une à
l’autre. Ceci est vrai que ce soit en tant qu’outils de diagnostic comme cela est le cas pour
l’utilisation des ressources par les tourterelles au sein de la Barbade (chapitre II) ou au sens
mécanistique pour la relation entre hétérozygotie et aptitude phénotypique (chapitre III).
Finalement, bien que notre fil conducteur ait toujours été la relation phénotype-génotype,
celle-ci a néanmoins été abordée sous des angles très divers. Ainsi, les conclusions et
perspectives retenues à l’issue de chaque chapitre ont été associées afin de présenter une
synthèse critique sur les avancées générées par ces trois années d’études et de présenter un
ensemble de perspectives envisageables à plus ou moins long terme.

9
CHAPITRE I
-
PRESENTATION DU MODELE D’ETUDE
ET DES OUTILS METHODOLOGIQUES

10
CHAPITRE I
-
PRESENTATION DU MODELE D’ETUDE
ET DES OUTILS METHODOLOGIQUES
I. INTRODUCTION
A la publication de l’Origine des Espèces en 1859, Darwin reçut de vives critiques
quant au support de la sélection naturelle. Alors que Mendel publiait ses travaux sur l’hérédité
en 1866, ce n’est qu’une vingtaine d’années plus tard que Weismann, au travers d’une
présentation sur l’hérédité en 1883 à l’Université de Fribourg-en-Brisgau (Allemagne,
Weismann 1891), proposait l’existence d’un élément physique de l’hérédité. Néanmoins, il a
fallu encore attendre 80 ans pour que la structure de l’ADN soit identifiée (Watson & Crick
1953).
L’intime relation entre phénotype et génotype amène les biologistes à repenser leur
vision de l’individu en considérant non seulement ces deux caractéristiques, mais également
l’interaction entre l’environnement et le patrimoine génétique. Les gènes et les produits de
leur expression (protéines) deviennent alors des marqueurs de variations environnementales
(Lambert 1995, Beebee & Rowe 2008). Cependant, une quarantaine d’années a été nécessaire
pour que des techniques initialement réservées aux généticiens et aux biologistes moléculaires
soient introduites en écologie comme outils de diagnostic. Ceci marque l’essor de l’écologie
moléculaire - discipline apparue dans les années 1960 - qui connut une reconnaissance
internationale notamment avec la création en 1992 d’une revue spécifique Molecular Ecology
(Burke et al. 1992, Lambert 1995, Avise 2006).
Les apports des techniques moléculaires ont permis des avancées majeures dans divers
courants tels que la biologie de la conservation qui a, par exemple, permis d’identifier
l’origine de la faible fertilité des guépards africains, Acinonyx jubatus. Par l’utilisation

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
11
d’outils moléculaires, une baisse de la diversité génétique, consécutive à deux réductions
majeures de l’effectif des populations, a pu être mise en évidence (Menotti-Raymond &
O’Brien 1993). Cependant, les apports les plus spectaculaires concernent sans doute la remise
en question d’un principe considéré jusqu’alors comme universel. Chez les oiseaux, plus de
93 % des espèces de passereaux étaient considérées comme monogames (Lack 1968). Une
revue de la littérature recensant les taux de paternités hors couples - déterminées par
l’utilisation de marqueurs microsatellites - a permis de montrer qu’environ 86 % de ces
espèces pratiquaient les copulations hors couples (Griffith et al. 2002). Ainsi, l’utilisation de
marqueurs moléculaires a permis de rectifier la vision "idyllique" de la monogamie aviaire en
apportant une nuance à la définition de la monogamie telle qu’elle était vue par Lack (1968)
en la qualifiant de monogamie sociale.
Dans l’ensemble, la biologie moléculaire a réellement révolutionné l’écologie et la
biologie évolutive (Ellegren 2008). Toutefois, l’avantage de ces techniques qui est de mettre
en évidence la variabilité se révèle être également une difficulté technique puisque cela
signifie que ces outils doivent être adaptés à leur cible. Or, ceci n’est pas toujours possible
(limitation du matériel biologique, défaut de compétences, de temps ou de financement pour
le développement). Dans le cas des microsatellites par exemple, bien qu’à l’heure actuelle leur
développement puisse être relativement aisé (à raison de temps et/ou d’argent),
l’amplification transpécifique de certains marqueurs peut s’avérer être une alternative
intéressante dans le cas où ceux-ci ont pu être développés chez des espèces proches, du même
genre voire parfois de la même famille (Wilson et al. 2004, Selkoe & Toonen 2006). Ainsi,
bon nombre d’articles introduisant de nouveaux marqueurs microsatellites intègrent une
section concernant l’amplification transpécifique (pour des exemples récents : Hoffman 2009,
Ma et al. 2009, Miles et al. 2009). Globalement, et ce quelle que soit la technique utilisée, la
première étape dans toute analyse moléculaire est de rechercher ce qui est disponible chez des
espèces proches.
Dans le cas de la tourterelle à queue carrée, Zenaida aurita, certaines populations
(Barbade et Porto Rico principalement) ont déjà été étudiées principalement sur des aspects
démographiques, comportementaux et morphologiques (Nellis & Dewey 1978, Nellis et al.
1984, Burger et al. 1989, Wiley 1991, Rivera-Milán 1992, 1996, 1997, 1999, 2001, Carlier &
Lefebvre 1996, Dolman et al. 1996, Lefebvre et al. 1996, 1997, Rivera-Milán & Vásquez
2000, Goldberg et al. 2001, Seferta et al. 2001, Rivera-Milán & Schaffner 2002, Griffin et al.
2005, Sol et al. 2005) mais peu en génétique et uniquement via un séquençage de cibles
mitochondriales (Johnson & Clayton 2000a et b). Comme bon nombre de colombidés, cette

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
12
espèce est monomorphe, la question du sexage moléculaire marque donc le début de nos
investigations d’un point de vue génétique chez cette espèce. Par contre, d’autres espèces du
genre Zenaida ont fait l’objet d’analyses génétiques. Cela est notamment le cas de Z.
galapagoensis dont la variabilité génétique dans l’archipel des Galápagos a été étudiée à
l’aide de marqueurs microsatellites initialement développés chez Z. asiatica (Santiago-
Alarcon et al. 2006) De ce fait, ces marqueurs constituaient une piste intéressante pour
débuter, du moins en ce qui concerne l’étude de la variabilité inter-individuelle car en ce qui
concerne le sexage les seuls colombidés sexés génétiquement n’étaient pas aussi proches
phylogénétiquement parlant (Barré et al. 2003, Pike 2005).
La première phase de l’étude entreprise sur la tourterelle à queue carrée a donc été de
mettre en place une ossature méthodologique constituée d’outils aussi bien morphologiques
par l’acquisition de données morphométriques que génétiques via le sexage moléculaire et les
marqueurs microsatellites. Afin d’établir une base générale pour ce chapitre mais également
pour les suivants, nous procédons ici à une brève présentation du modèle biologique et de la
méthode d’acquisition des données ou en d’autres termes du travail de terrain. Par la suite, le
sexage moléculaire ainsi que l’application de cette méthode couplée à des techniques
morphométriques d’un point de vue méthodologique, seront exposés. Enfin, le développement
des marqueurs microsatellites a également été inclus dans ce chapitre. Toutefois, leur
application étant présentée dans les chapitres suivants, aucun exemple concret de leur
utilisation n’a été ajouté ici.
II. METHODE GENERALE : MODELE BIOLOGIQUE ET ACQUISITION
DES DONNEES
1. La tourterelle à queue carrée, Zenaida aurita
La tourterelle à queue carrée est un colombidé d’une taille corporelle (longueur du
corps) d’environ trente centimètres. Elle appartient au genre Zenaida regroupant sept espèces
de tourterelles exclusivement inféodées au Nouveau Monde (tableau 1). Certaines espèces de
ce genre sont largement distribuées à l’échelle du continent américain contrairement à
d’autres qui sont strictement inféodées à des complexes insulaires ce qui est le cas de notre
modèle biologique. Z. aurita est présente dans l’ensemble des Caraïbes (Bahamas, Grandes et
Petites Antilles, Evans 1990, Raffaele et al. 2006).

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
13
Tableau 1 : Espèces appartenant au genre Zenaida et leur aire de répartition.
Nom vernaculaire
Espèce Français Anglais Localisation
Z. macroura Tourterelle triste Mourning dove Amérique du Nord
Z. auriculata Tourterelle oreillarde Eared dove Amérique du Sud
Z. galapagoensis Tourterelle des Galápagos Galápagos dove Galápagos
Z. graysoni Tourterelle de Socorro Socorro dove Socorro*
Z. asiatica Tourterelle à ailes blanches White-winged dove Amérique du Nord et Centrale
Z. meloda Tourterelle mélodieuse Pacific dove Amérique du Sud
Z. aurita Tourterelle à queue carrée Zenaida dove Caraïbes
* population naturelle éteinte, populations captives en zoo uniquement.
Dans la plupart des îles caribéennes, les tourterelles à queue carrée occupent des
habitats principalement de type forestier mais elles sont généralistes dans leurs préférences de
nidification et d’approvisionnement (Nellis et al. 1984, Burger et al. 1989, Wiley 1991,
Rivera-Milán 1997, 1999), excepté à la Barbade où elles sont relativement anthropophiles,
notamment à cause de la destruction de la quasi-totalité de l’espace forestier (seulement une
faible surface de forêt primitive a été épargnée par la culture de la canne à sucre, Welchman
Hall Gully, St.Thomas Parish) et vivent en milieux faiblement urbanisés dans des parcs et
jardins arborés. Cette espèce est territoriale : les couples s’établissent sur des territoires qu’ils
défendent tout au long de l’année contre l’intrusion de congénères avec un comportement
ritualisé consistant en un lever d’aile ("wing raising", Lefebvre et al. 1996, figure 1).

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
14
Figure 1 : Comportement ritualisé de défense du territoire par lever d’aile ("wing raising")
chez la tourterelle à queue carrée, Zenaida aurita.
A l’image de bon nombre de colombidés y compris d’autres espèces du genre Zenaida
telles que Z. asiatica (Swanson & Rappole 1992) et Z. galapagoensis (Santiago-Alarcon &
Parker 2007), le dimorphisme sexuel chez la tourterelle à queue carrée est peu ou pas marqué
même si certains aspects du plumage ont pu être utilisés jusque là comme clés potentielles
d’identification sexuelle (del Hoyo et al. 1997, Gibbs et al. 2001, Sol et al. 2005). Toutefois,
ceci suppose que la variabilité interindividuelle du plumage chez Z. aurita ne puisse pas
biaiser l’assignement du sexe. De plus, chez les juvéniles, le sexage est impossible sur des
critères phénotypiques car leur plumage grisonnant permet, certes, leur discrimination vis-à-
vis des adultes, mais ne présente aucune différence entre sexes. Compte tenu du fait que
l’observation de comportements sexuels n’est pas toujours possible, plusieurs méthodes sont
potentiellement réalisables dont le sexage moléculaire et l’utilisation des mensurations des
individus qui ont été abordés un peu plus loin dans ce chapitre.

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
15
2. Capture, acquisition des données morphométriques et prélèvements
sanguins
Les captures de tourterelles ont été réalisées à l’aide de cages à porte rabattable
("walk-in baited drop traps") appâtées avec du riz cuit mais également avec des clapnets
appâtés avec du pain (figure 2).
Figure 2 : Pièges types utilisés pour la capture des tourterelles à queue carrée, a) cage à porte
rabattable et b) clapnet.
Une fois capturés, les oiseaux ont été bagués avec une combinaison unique de bagues
plastiques (darvic) colorées (A.C. Hughes Ltd., Hampton Hill, United Kingdom) et une bague
aluminium numérotée du Muséum d’Histoire Naturelle de Paris, un programme de baguage
du C.R.B.P.O. (Centre de Recherches par le Baguage des Populations d’Oiseaux) étant en
cours pour cette espèce. Par la suite, l’acquisition des données morphométriques a été
effectuée selon les recommandations du C.R.B.P.O. (Brucy et al. 2007) et ce, en double afin
d’évaluer l’erreur de mesure et la répétabilité de nos mesures (figure 3). La longueur, la
largeur et la hauteur du bec ainsi que la longueur totale de la tête et les tarses (droit et gauche)
ont été réalisées à l’aide d’un pied à coulisse électronique (précision = 0,2 mm). Les mesures
de la longueur des ailes (droite et gauche) et de la queue ont quant à elles été obtenues à l’aide
d’un réglet (précision: ± 1 mm). Une fois les mesures réalisées, deux échantillons de sang
d’environ 40 µl ont été prélevés par ponction de la veine brachiale (entre le radius et l’ulna) à
l’aide d’une aiguille stérile gauge 27 et un capillaire hépariné. Les prélèvements étaient
immédiatement transférés dans 800µl de tampon de conservation adapté aux conditions
tropicales (pas de réfrigération) et constitué de 70% d’éthanol absolu et de 30% de TE (Tris-
EDTA pH 8).

16
Figure 3 : Mesures et prélèvement sanguins effectués sur chaque tourterelle à queue carrée capturée. a) longueur du bec, b) largeur du bec, c)
hauteur du bec, d) longueur total de la tête, e) longueur du tarse, f) longueur de l’aile, g) longueur de la queue, h) prise de sang et i) pesée.
Ch
ap
itre I –
Pré
sen
tatio
n d
u m
od
èle
d’é
tud
e e
t de
s ou
tils mé
tho
do
log
iqu
es

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
17
Une fois le prélèvement terminé, les oiseaux ont été pesés grâce à une balance digitale
de poche PESOLA® MS 500 (précision: ± 0.1 g, Suisse). Une fois l’ensemble du traitement
effectué, les oiseaux étaient relâchés sur leur site de capture.
3. Préparation des échantillons sanguins pour les analyses moléculaires
subséquentes : l’extraction d’ADN
Le sang a été choisi comme support pour les analyses moléculaires pour différentes
raisons. Chez les oiseaux le volume sanguin représente 7 à 10 % de la masse corporelle. Chez
la tourterelle triste, Zenaida macroura, qui est de taille et de masse comparables à Z. aurita,
un prélèvement sanguin d’un volume allant jusqu’à 0,5 ml ne semble pas porter atteinte à
l’oiseau en termes de survie (Bigler et al. 1977). De ce fait, en ne prélevant que 80 µl, la prise
de sang n’est que peu contraignante pour l’individu (excepté le stress de la manipulation). Par
ailleurs, bon nombre d’études se sont penchées sur les conséquences d’une prise de sang sur
des espèces sauvages et globalement, cette technique ne semble pas être dommageable pour
les oiseaux (Hoysak & Wetherhead 1991, Sheldon et al. 2008). Même si de l’ADN peut être
extrait à partir d’autres supports tels que la pulpe des plumes fraichement arrachées, le sang,
même en faible quantité, permet d’obtenir une très grande quantité d’ADN de bonne qualité,
les hématies des oiseaux étant nucléées. Ainsi, le panel d’analyses génétiques potentiellement
réalisables n’est pas limité ce qui n’est pas forcément possible dans le cas des plumes
(Taberlet & Bouvet 1991, Hogan et al. 2007).
Le protocole d’extraction a été modifié d’après le protocole phénol-chloroforme
d’Hillis et al. (1996) afin d’être adapté à nos échantillons, notamment en ce qui concerne le
tampon de stockage. En effet, celui-ci contient de l’éthanol qui est un inhibiteur de la
protéinase K, utilisée pour lyser les parois des hématies. La première étape du protocole était
donc de retirer ce tampon afin de ne garder que le sang. Pour cela, 100 µl de l’échantillon ont
été prélevés et centrifugés à 4000 rpm pendant une minute puis le surnageant contenant
seulement le tampon a été retiré. Au culot, 200 µl de tampon de lyse (Queen’s Lysis Buffer,
Seutin et al. 1991) et 0,1 mg de protéinase K ont été ajoutés et le tout a été laissé à incuber à
55°C pour un minimum de trois heures. Par la suite, un double lavage au phénol-chloroforme
a été réalisé suivi d’un autre uniquement au chloroforme afin d’ôter un maximum de lipides et
de phénol. Entre chaque lavage, le surnageant a été récupéré après centrifugation à 12 000
rpm pendant 8 min à 15°C. A la fin du troisième lavage, 375 µl d’isopropanol glacé (-20°C)

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
18
ont été ajoutés au surnageant afin de précipiter l’ADN. Pour finir, l’ADN a été lavé avec trois
bains successifs d’éthanol 70% glacé (-20°C) couplés à une centrifugation à 15 000 rpm
pendant 5 min à 4°C, puis séché et enfin mis en suspension dans 100 µl un tampon d’élution
(Tris-EDTA ou TE pH8). Les extraits étaient alors utilisables sous un délai d’une demi-
journée, délai minimum nécessaire à leur bonne remise en suspension. Par la suite, leur
conservation s’effectuait à 4°C tant qu’ils étaient utilisés, puis à -20°C pour un stockage sur le
long terme.
III. SEXAGE ET DIMORPHISME SEXUEL CHEZ LA TOURTERELLE A
QUEUE CARREE : LES PROBLEMES METHODOLOGIQUES
1. Sexage moléculaire
a. Généralités sur le sexage chez les oiseaux
Chez les oiseaux, comme nous l’avons vu précédemment, bon nombre d’espèces sont
monogames que ce soit socialement ou génétiquement. Les origines du maintien de ce régime
d’appariement sont multiples. Parmi elles, les soins biparentaux aux jeunes sont souvent
avancés comme une contrainte majeure (celle-ci découlant de l’oviparité) ayant favorisée le
maintien de la monogamie mais la territorialité par exemple, en est une autre (Mock 1985,
Gowaty 1996, Cézilly 2006). Globalement, l’ensemble de ces contraintes tend à réduire la
variabilité du succès reproducteur des mâles monogames comparés aux mâles polygynes et en
conséquences diminuerait l’intensité de la sélection sexuelle et par extension les différences
entre mâles et femelles d’un point de vue phénotypique.
Un faible dimorphisme sexuel et des organes reproducteurs internes font que les
espèces aviaires monomorphe sont difficiles à sexer. Jusque vers la fin du XXème siècle,
diverses méthodes anatomiques par observation des organes sexuels internes (Petrides 1950,
Miller & Wagner 1955, Shamis & Forrester 1977, Bancroft 1984, Malacalaza & Hall 1988,
Swanson & Rappole 1992, Lorentsen & Røv 1994 Small et al. 2005), analyses des
vocalisations (Blakesley et al. 1990, Murie et al. 1991, Bretagnolle & Thibault 1995, Lo
Valvo 2001, Gill & Vonhof 2006, Bourgeois et al. 2007) ou des comportements sexe-
spécifiques (Castoro & Guhl 1958, Chardine & Morris 1989, Desrochers 1990, Hull 1996,

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
19
Martínez-Gómez & Curry 1998, Flux & Innes 2001, Fletcher & Hamer 2003, Alarcos et al.
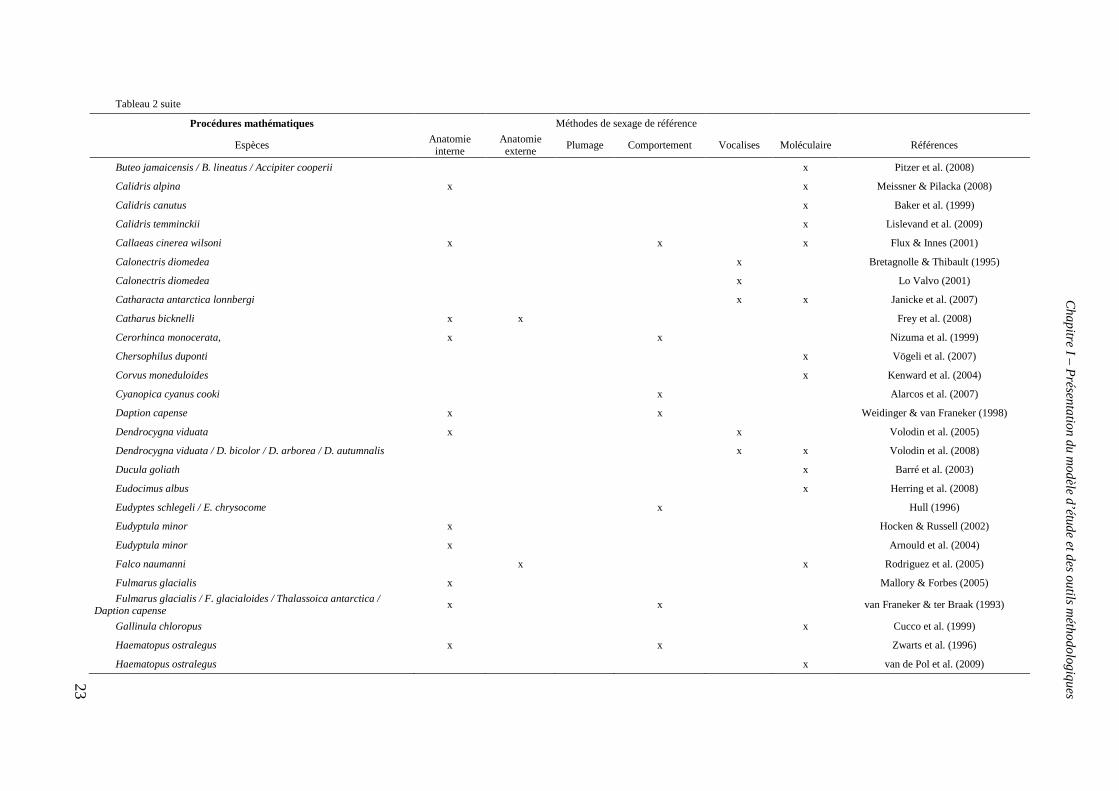
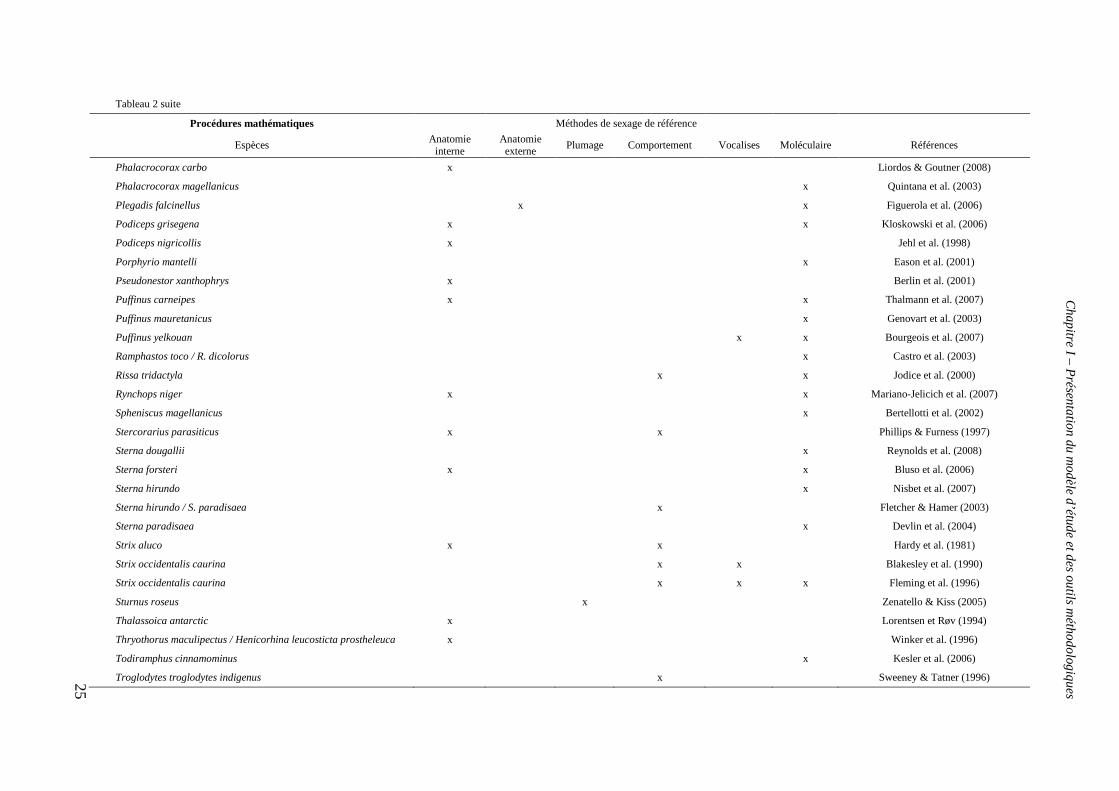
2007) mais également des procédures mathématiques telles que l’analyse discriminante
associées aux techniques citées précédemment (tableau 2). Puis dans les années 1990, les
premières techniques moléculaires dédiées au sexage des oiseaux apparaissent et deviennent
quasiment incontournables dès lors que le déterminisme du sexe est nécessaire (pour une
présentation de ces techniques se référer à Dubiec & Zagalska-Neubauer 2006).
b. Méthodes de sexage moléculaire
i. Généralités
Chez les oiseaux non-ratites, le sexage moléculaire est essentiellement basé sur des
variations de séquences du gène CHD1 (codant pour une protéine, la "Chromo-Helicase-DNA
binding protein", Griffiths & Tiwari 1993, Ellegren 1996, Griffiths et al. 1996, Dubiec &
Zagalska-Neubauer 2006). Ce gène est situé sur les deux chromosomes sexuels, Z et W,
présents chez les oiseaux et les séquences arborent un haut degré de conservation entre elles
(Ellegren 1996, Griffiths et al. 1996, Ellegren & Fridolfsson 1997, Griffiths & Korn 1997,
Fridolfsson et al. 1998). A l’inverse des humains où les hommes sont hétérogamétiques, chez
les oiseaux se sont les femelles qui le sont (ZW) alors que les mâles sont homogamétiques
(ZZ). Ainsi, le gène CHD1 est un marqueur intéressant pour le déterminisme du sexe. Chaque
copie du gène (CHD1-W et CHD1-Z) présent sur l’un ou l’autre des chromosomes sexuels
diffère de l’autre par leurs séquences ou la taille d’un intron. Bon nombre de techniques
moléculaires basées sur l’amplification de différentes sections du gène ont été développées à
ce jour (Dubiec & Zagalska-Neubauer 2006). Toutefois, le choix d’une méthode de
visualisation de ce polymorphisme n’est pas évident car il est dépendant de différentes
contraintes techniques inhérentes à la PCR. Dans un premier temps, la conservation des
séquences des amorces doit être vérifiée ce qui se traduit par une bonne qualité
d’amplification pour les copies Z et W (absence d’allèle nul comme pour les microsatellites,
Pemberton et al. 1995). Un autre problème émane du fait que les techniques de PCR
couramment employées sont basées sur la détection d’un polymorphisme de taille des allèles.
Dans certains cas, seul le plus petit allèle est amplifié ("short allele dominance", voir Wattier
et al. 1998). Ceci peut potentiellement poser problème lorsque que l’allèle diagnostique
(spécifique aux femelles) à savoir CHD1-W n’est pas le plus petit. En effet, si CHD1-Z est
préférentiellement amplifié à défaut de CHD1-W, certaines femelles seront identifiées comme

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
20
mâles. Enfin, le dernier point important est la quantité d’ADN disponible. Si celle-ci n’est pas
suffisante, seulement un des deux allèles peut être aléatoirement amplifié, ce qui, comme
précédemment, peut induire une assignation au mauvais sexe ("allelic drop out", voir Taberlet
et al. 1996). Par la suite, les résultats obtenus via les analyses moléculaires doivent être
confrontés à ceux obtenus par une autre méthode (observations comportementales, anatomie,
vocalises) ce qui ne peut être souvent réalisé que sur un faible nombre d’individus (Robertson
& Gemmell 2006).
ii. Mise en place du sexage moléculaire chez la tourterelle à queue
carrée
Afin de choisir la méthode moléculaire la plus appropriée au sexage des tourterelles à
queue carrée, deux couples d’amorces ont été testés. Le premier couple, P2 (5′-
TCTGCATCGCTAAATCCTTT-3’) et P8 (5′-CTCCCAAGGATGAGRAAYTG-3′), a été
défini par Griffiths et al. (1998) et le second, 2550F (5′-GTTACTGATTCGTCTACGAGA-
3′) et 2718R (5′-ATTGAAATGATCCAGTGCTTG-3′), a été défini par Fridolfsson &
Ellegren (1999). Le choix pour ces deux paires d’amorces ne s’est pas fait au hasard. Non
seulement ils amplifient le gène CHD1 sur des portions différentes (incluant chacune un
intron), ce qui offre une base méthodologique pour comparer la qualité des deux techniques
mais ce sont également les plus employés (Griffiths et al. 1998 ont été cités 773 fois contre
335 citations pour Fridolfsson & Ellegren 1999 ; source : Web of Knowledge, octobre 2009).
De plus, le couple 2550F / 2718R a déjà été utilisé pour le sexage chez des colombidés
comme le pigeon domestique, Columba livia domestica (Pike 2005) ou chez le notou, Ducula
goliath (Barré et al. 2003).
Dans un premier temps, trois couples de tourterelles (observations comportementales)
ont permis de tester le couple P2 / P8 puis un groupe de 88 individus (sélectionnés au hasard)
ont été sexés par P2 / P8 et 2550F / 2718R. Les PCR étaient effectuées dans un volume total
de 10 µl composés de 100 ng d’ADN (pas d’"allelic drop-out"), 200 µM de chaque dNTP,
200 nM de chaque amorce, 1X de tampon de réaction et 0,45 U de Taqpolymérase HotMaster
(5-PRIME). L’amplification de l’ADN était effectuée dans un thermocycler T3 (Biometra) et
commençait par une dénaturation initiale à 94°C pour 1 min. 30 suivi de 35 cycles de 20 s. à
94°C, 30 s. à 46°C et 30 s. à 65°C pour terminer avec une élongation finale à 65°C pendant 3
min. Etant donné que les produits issus de l’amplification avec P2 / P8 ne différaient pas en

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
21
taille, une étape supplémentaire de restriction a été appliquée sur ce couple avant
l’électrophorèse. A cette fin, les produits de PCR issus de P2 / P8 ont été digérés avec une
unité d’enzyme de restriction DdeI pendant 5 h à 37°C, coupant spécifiquement l’allèle
CHD1-W (Griffiths et al. 1996, Bermúdez-Humarán et al. 2002). Les variations de taille des
introns ont été par la suite visualisées après une électrophorèse de 30 min (100 V) sur gel
d’agarose à 3 % dans un tampon TBE 1 X (Tris-borate-EDTA) et une révélation sous lampe
UV après immersion dans un bain de bromure de BET (bromure d’éthidium). Les tailles
obtenues différaient en fonction du couple d’amorces utilisées avec CHD1-W = 290 + 90 pb
et CHD1-Z = 380 pb pour P2 / P8 (après restriction) et CHD1-W = 450 pb et CHD1-Z = 750
pb pour 2550F / 2718R. Sur les 88 individus sexés, les deux méthodes apportent les mêmes
résultats. La différence majeure entre ces deux techniques, outre le temps de manipulation,
était liée au biais potentiellement introduit par le recours à l’enzyme de restriction. En effet, si
l’enzyme ne coupait pas (mauvaise manipulation, inactivation de l’enzyme), les femelles
seraient identifiées comme étant des mâles ce qui implique de réaliser la manipulation deux
fois, du moins sur les individus identifiés comme mâles. De ce fait, le couple d’amorces
retenu pour le sexage chez la tourterelle à queue carrée a été 2550F / 2718 R. De plus, ce
couple d’amorce produit des fragments dont la différence de taille (300bp) est très aisément
détectable sur gel d’agarose à 2%.
Depuis cette étude préliminaire, des travaux impliquant un sexage moléculaire chez
d’autres colombidés réalisé avec ce même couple d’amorces ont été publiés notamment chez
la tourterelle turque, Streptopelia decaocto (Eraud et al. 2008) mais également chez une
espèce appartenant au genre Zenaida, Z. galapagoensis (Santiago-Alarcon & Parker 2007).

Ch
ap
itre I –
Pré
sen
tatio
n d
u m
od
èle
d’é
tud
e e
t de
s ou
tils mé
tho
do
log
iqu
es
22
Tableau 2: Exemples d’études incluant un sexage via une procédure mathématique (analyse en composante principale, modèle linéaire
généralisé, régression linéaire et analyse discriminante) et une ou plusieurs méthodes de référence.
Procédures mathématiques Méthodes de sexage de référence
Espèces Anatomie
interne Anatomie externe
Plumage Comportement Vocalises Moléculaire Références
Analyse en composante principale
Biziura lobata McCracken et al. (2000)
Bubo bubo x del mar Delgado & Penteriani (2004)
Chlidonias niger x Shealer & Cleary (2007)
Limosa limosa limosa x Schroeder et al. (2008)
Mycteria leucocephala x Urfi & Kalam (2006)
Phalaropus lobatus x Rubega (1996)
Tringa glareola x Remisiewicz & Wenerberg (2006)
Modèle linéaire généralisé
Calidris maritima x Hallgrimsson et al. (2008)
Régression linéaire
Caracara plancus x Morrison & Maltbie (1999)
Gallinago hardwickii x Ura et al. (2005)
Loxioides bailleui x Jeffrey et al. (1993)
Porphyrio porphyrio melanotus x Millar et al. (1996)
Thryothorus leucotis x x Gill & Vonhof (2006)
Analyse discriminante et régression linéaire
Charadrius montanus x Iko et al. (2004)
Analyse discriminante
Aegolius funereus x Hipkiss (2007)
Alca torda x Grecian et al. (2003)
Alle alle x Jakubas & Wojczulanis (2007)
Anous stolidus x Chardine & Morris (1989)
Buteo jamaicensis x Donohue & Dufty (2006)
Ch
ap
itre I –
Pré
sen
tatio
n d
u m
od
èle
d’é
tud
e e
t de
s ou
tils mé
tho
do
log
iqu
es
23
Tableau 2 suite
Procédures mathématiques Méthodes de sexage de référence
Espèces Anatomie
interne Anatomie externe
Plumage Comportement Vocalises Moléculaire Références
Buteo jamaicensis / B. lineatus / Accipiter cooperii x Pitzer et al. (2008)
Calidris alpina x x Meissner & Pilacka (2008)
Calidris canutus x Baker et al. (1999)
Calidris temminckii x Lislevand et al. (2009)
Callaeas cinerea wilsoni x x x Flux & Innes (2001)
Calonectris diomedea x Bretagnolle & Thibault (1995)
Calonectris diomedea x Lo Valvo (2001)
Catharacta antarctica lonnbergi x x Janicke et al. (2007)
Catharus bicknelli x x Frey et al. (2008)
Cerorhinca monocerata, x x Nizuma et al. (1999)
Chersophilus duponti x Vögeli et al. (2007)
Corvus moneduloides x Kenward et al. (2004)
Cyanopica cyanus cooki x Alarcos et al. (2007)
Daption capense x x Weidinger & van Franeker (1998)
Dendrocygna viduata x x Volodin et al. (2005)
Dendrocygna viduata / D. bicolor / D. arborea / D. autumnalis x x Volodin et al. (2008)
Ducula goliath x Barré et al. (2003)
Eudocimus albus x Herring et al. (2008)
Eudyptes schlegeli / E. chrysocome x Hull (1996)
Eudyptula minor x Hocken & Russell (2002)
Eudyptula minor x Arnould et al. (2004)
Falco naumanni x x Rodriguez et al. (2005)
Fulmarus glacialis x Mallory & Forbes (2005)
Fulmarus glacialis / F. glacialoides / Thalassoica antarctica / Daption capense
x x van Franeker & ter Braak (1993)
Gallinula chloropus x Cucco et al. (1999)
Haematopus ostralegus x x Zwarts et al. (1996)
Haematopus ostralegus x van de Pol et al. (2009)

Ch
ap
itre I –
Pré
sen
tatio
n d
u m
od
èle
d’é
tud
e e
t de
s ou
tils mé
tho
do
log
iqu
es
24
Tableau 2 suite
Procédures mathématiques Méthodes de sexage de référence
Espèces Anatomie
interne Anatomie externe
Plumage Comportement Vocalises Moléculaire Références
Haliaeetus albicilla x Helander et al. (2007)
Hieraaetus fasciatus x Palma et al. (2001)
Hirundo rustica x Hermosell et al. (2007)
Hylophilus ochraceiceps ochraceiceps / H. decurtatus decurtatus / Xenops minutus mexicanus / Automolus ochrolaemus cervinigularis
x Winker et al. (1994)
Lagopus lagopus x x x Gruys & Hannon (1993)
Larus argentatus x Evans et al. (1995)
Larus atricilla x Hanners & Patton (1985)
Larus cachinnans x Bosch (1996)
Larus californicus x Schnell et al. (1985)
Larus californicus x Rodriguez et al. (1996)
Larus crassirostris x Chochi et al. (2002)
Larus michahellis lusitanius x Arizaga et al. (2008)
Larus michahellis lusitanius x Galarza et al. (2008)
Larus novaehollandiae scopulinus x Mills (1971)
Lonchura striata phaethontoptila / Lonchura striata var. domestica x x Mizuta et al. (2003)
Lymnocryptes minimus x Sikora & Dubiec (2007)
Macronectes giganteus x Copello et al. (2006)
Megadyptes antipodes x Setiawan et al. (2004)
Miliaria calandra x Campos et al. (2005)
Mimodes graysoni x Martinez-Gomez & Cury (1998)
Nestor notabilis x Bond et al. (1991)
Parus atricapillus x Desrochers (1990)
Passerculus sandwichensis x x Wheelwright et al. (1994)
Pelecanus erythrorhynchos x Dorr et al. (2005)
Phalacrocorax albiventer x Malacalaza & Hall (1988)
Phalacrocorax atriceps x x x Svagelj & Quintana (2007)
Phalacrocorax auritus auritus x Glahn & McCoy (1995)
Ch
ap
itre I –
Pré
sen
tatio
n d
u m
od
èle
d’é
tud
e e
t de
s ou
tils mé
tho
do
log
iqu
es
25
Tableau 2 suite
Procédures mathématiques Méthodes de sexage de référence
Espèces Anatomie
interne Anatomie externe
Plumage Comportement Vocalises Moléculaire Références
Phalacrocorax carbo x Liordos & Goutner (2008)
Phalacrocorax magellanicus x Quintana et al. (2003)
Plegadis falcinellus x x Figuerola et al. (2006)
Podiceps grisegena x x Kloskowski et al. (2006)
Podiceps nigricollis x Jehl et al. (1998)
Porphyrio mantelli x Eason et al. (2001)
Pseudonestor xanthophrys x Berlin et al. (2001)
Puffinus carneipes x x Thalmann et al. (2007)
Puffinus mauretanicus x Genovart et al. (2003)
Puffinus yelkouan x x Bourgeois et al. (2007)
Ramphastos toco / R. dicolorus x Castro et al. (2003)
Rissa tridactyla x x Jodice et al. (2000)
Rynchops niger x x Mariano-Jelicich et al. (2007)
Spheniscus magellanicus x Bertellotti et al. (2002)
Stercorarius parasiticus x x Phillips & Furness (1997)
Sterna dougallii x Reynolds et al. (2008)
Sterna forsteri x x Bluso et al. (2006)
Sterna hirundo x Nisbet et al. (2007)
Sterna hirundo / S. paradisaea x Fletcher & Hamer (2003)
Sterna paradisaea x Devlin et al. (2004)
Strix aluco x x Hardy et al. (1981)
Strix occidentalis caurina x x Blakesley et al. (1990)
Strix occidentalis caurina x x x Fleming et al. (1996)
Sturnus roseus x Zenatello & Kiss (2005)
Thalassoica antarctic x Lorentsen et Røv (1994)
Thryothorus maculipectus / Henicorhina leucosticta prostheleuca x Winker et al. (1996)
Todiramphus cinnamominus x Kesler et al. (2006)
Troglodytes troglodytes indigenus x Sweeney & Tatner (1996)

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
26
2. Sexage via l’utilisation de données morphométriques : les biais
méthodologiques
ANNEXE 1
Monceau, K., Cézilly, F. & Dechaume-Moncharmont, F.-X. Sexing birds using discriminant
function analysis based on morphometric measurements: A critical appraisal.
Manuscrit en préparation
Comme présenté en introduction de ce chapitre, plusieurs méthodes de sexage ont été
employées lors d’études sur des espèces aviaires présentant un faible dimorphisme sexuel.
Parmi les méthodes couramment retrouvées dans la littérature, l’analyse discriminante (AD),
procédure statistique de classement des individus, apparait comme étant une méthode rapide
et peu invasive au moment du baguage. Le principe de l’AD est simple dans sa philosophie
mais sa mise en œuvre pratique peut s’avérer dans certains cas un peu technique. Il s’agit de
trouver la combinaison (linéaire ou non) de variables, discriminant au mieux les individus
mesurés. Dans notre cas, l’analyse est effectuée sur un groupe d’oiseaux de sexe identifié
(avec certitude) à l’avance par une méthode alternative (sexage moléculaire) et sur lesquels
une série de mesures morphométriques a pu être effectuée. L’AD permet d’établir une
équation prédisant au mieux le sexe des individus. Grâce à celle-ci, chaque oiseau obtient un
score l’assignant à un groupe, en l’occurrence ici soit aux femelles soit aux mâles. Une fois
cette équation déterminée, le pouvoir discriminant global (ou score de discrimination) peut
être évalué par une méthode de validation.
D’une manière générale et comme pour toute procédure statistique, il convient de
porter une attention particulière aux nombreux biais potentiels pouvant influencer la
pertinence de l’analyse. Dans le cas particulier de l’AD, un examen de la littérature révèle que
les premiers auteurs à utiliser cette technique y étaient très sensibles. Avec la popularisation
de cette méthode et l’augmentation du nombre de logiciels d’analyses statistiques proposant
des algorithmes d’AD (plus ou moins automatisés), certains auteurs semblent avoir négligé de

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
27
nombreuses recommandations pourtant clairement évoquées par ailleurs dans la littérature
statistique. Celles-ci sont de divers ordres. Tout d’abord, la taille de l’échantillon est un
paramètre incontournable en statistique limitant souvent la puissance des tests. Ainsi,
intuitivement nous nous attendons à ce que le niveau de discrimination augmente avec une
taille d’échantillon croissante. Le second point concerne les erreurs de mesures. Celui-ci est
surtout inhérent à la prise de données morphométriques. Dans notre cas, l’enjeu est de réussir,
à identifier mâles et femelles chez des espèces à faible dimorphisme sexuel et donc chez
lesquelles les différences mesurées sont faibles. De ce fait, l’erreur de mesure pourrait
affaiblir le niveau de discrimination de l’AD en réduisant artificiellement ces différences. Le
troisième point concerne la méthode de réduction du nombre de variables incluses dans la
fonction discriminante finale consistant en une procédure automatique ou non d’élimination
ou insertion "pas à pas" ("stepwise") de variables. En effet, l’utilisation de routines
automatiques, implémentées dans bon nombre de logiciels statistiques classiques, est
fortement remise en cause non seulement dans des revues de statistiques mais également dans
des domaines moins spécialisés comme dans des revues d’écologie (Williams & Titus 1988)
ou de psychologie (Thompson 1995). Enfin le dernier point concerne la méthode de validation
à l’issue de l’AD à proprement parler. En fait, trois grandes familles de méthodes de
validation existent (Eisenbeis 1977). Tout d’abord, la technique de re-substitution consiste à
utiliser directement la population ayant servi à calibrer l’équation discriminante. La raison de
sa popularité réside dans sa grande simplicité. Toutefois, elle présente de nombreux biais
évidents du fait de la circularité gênante consistant à évaluer une fonction avec les données
mêmes ayant servi à sa calibration. De ce fait, le score discriminant obtenu est toujours
surestimé (Eisenbeis 1977, Ryder 1978). La seconde technique envisageable consiste à
effectuer des validations croisées ("leave-one-out procedure" également connu sous le nom de
"jackknife") : un individu de la population des n individus que contient le jeu de données
initial est exclu puis l’équation discriminante est ajustée sur les n-1 individus restants. Enfin,
le score de l’individu initialement retiré de l’analyse est alors estimé. Cette procédure est
répétée pour les n individus de la population. Elle conduit à un score de discrimination non
biaisé et qui devrait donc être moins surévalué que celui obtenu par re-substitution. Cette
méthode de jackknife est assez gourmande en calcul mais elle est maintenant implémentée en
routine sur la plupart des grands logiciels de statistiques (R, SAS, SPSS). La troisième grande
classe de méthodes consiste à diviser le jeu de données en deux sous-échantillons ("sample
splitting"), l’un servant à ajuster l’équation discriminante et l’autre en tester le pouvoir
discriminant. Cette méthode est par définition, elle aussi, exempte de biais et présente

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
28
l’avantage d’être très intuitive et facile à mettre en œuvre. Pourtant, plusieurs problèmes
peuvent être notés. L’équation et le score discriminant associé sont intimement dépendants du
sous-échantillonnage effectué aléatoirement. En d’autres termes, avec un autre
échantillonnage, l’équation et le score obtenus sont potentiellement différents.
Globalement, une revue de la littérature (85 articles) nous a permis de constater que la
plus grande confusion régnait quant à l’utilisation de routines automatiques de réduction de
variables et à la méthode employée pour l’évaluation du score de discrimination. Dans le
premier cas, une fraction très importante des travaux que nous avons évalués (60%) ont eu
recours aux méthodes automatiques. De même pour les techniques de validation où 43,5% des
travaux rapportaient clairement aucune méthode d’évaluation ou avaient utilisé la technique
de re-substitution, 35,3% avaient utilisé le jackknife, 21,2% le sous-échantillonnage. Cette
diversité des méthodes est d’autant plus problématique, que de nombreux auteurs comparaient
les scores de discrimination de plusieurs études aux méthodes différentes. De plus, seulement
cinq études sur les 85 étudiées évoquaient l’existence de l’erreur de mesure (Jodice et al.
2000, Flux & Innes 2001, Devlin et al. 2004, Kenward et al. 2004, Mallory & Forbes 2005).
A partir de simulations réalisées sur un jeu de données important récoltées sur 525
tourterelles à queue carrée capturées en 2007, les effets de ces quatre paramètres (taille de
l’échantillon, erreur de mesure, réduction pas à pas du nombre de variables et effet de la
méthode d’évaluation du score de discrimination) ont été testés. Cette espèce est d’autant plus
un bon modèle pour ce type d’analyse que le niveau de dimorphisme sexuel qu’elle présente
est faible.
A l’issue des simulations, les effets de la taille d’échantillon sont contradictoires en
fonction de la méthode de validation employée, l’une augmentant le taux de discrimination de
pair avec un effectif croissant, alors que l’autre le diminue. Ainsi, en utilisant uniquement le
jackknife (procédure la moins biaisée), l’erreur de mesure semble fortement diminuer le
niveau de discrimination de l’AD.
En fin de compte, un problème méthodologique récurrent au sein de la littérature
scientifique semble émerger. Il concerne le niveau d’imprécision de certains articles publiés
dont les auteurs omettent consciemment ou non, de préciser la méthode de validation
employée. Derrière cela se cache également un problème lié aux logiciels statistiques
couramment employés tels que Jmp ou Statistica qui ne précisent pas quel type de méthode
est incrémenté dans leur fonction. De ce fait, le taux de discrimination de certaines analyses
pourrait être involontairement surévalué.

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
29
IV. ETUDE DE LA VARIABILITE INTERINDIVIDUELLE : MISE AU POINT DES MARQUEURS MICROSATELLITES
ANNEXE 2
Monceau, K., Gaillard, M., Harrang, E., Santiago-Alarcon, D., Parker, P., Cézilly, F. &
Wattier, R. 2009. Twenty-three polymorphic microsatellite markers for the Caribbean
endemic Zenaida dove, Zenaida aurita, and its conservation in related Zenaida species.
Conservation Genetics, 10, 1577-1581.
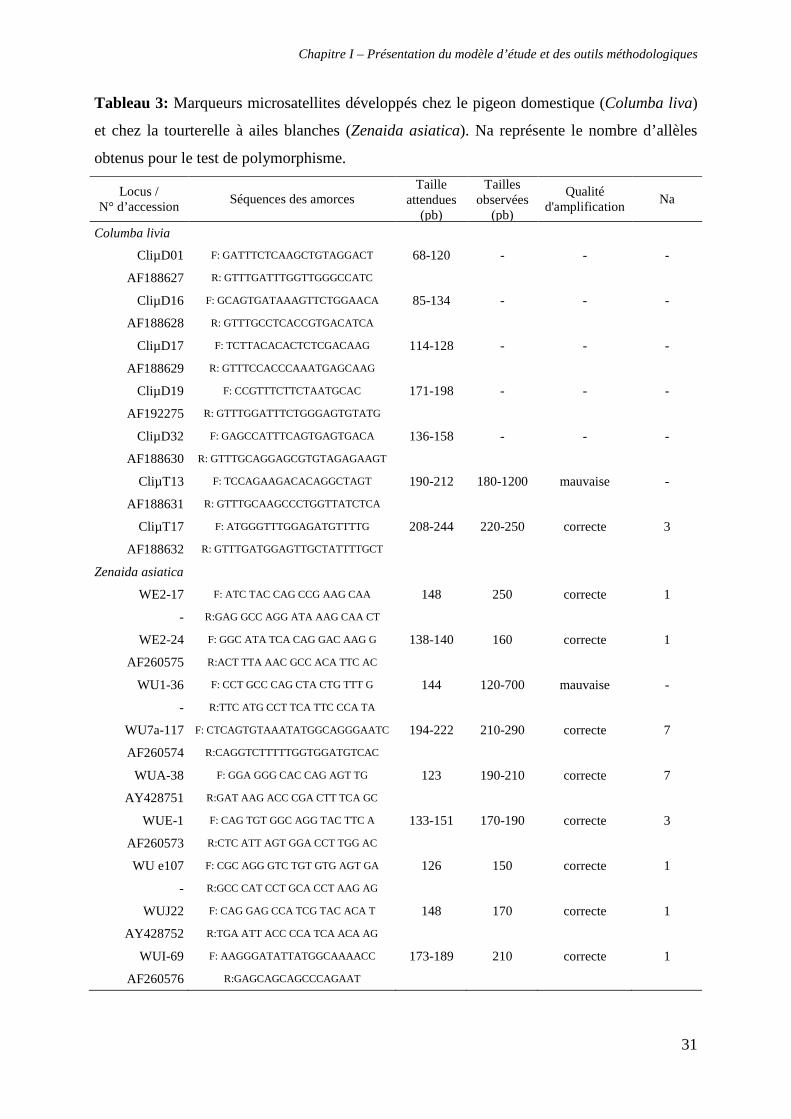
Compte tenu de l’existence de marqueurs microsatellites développés pour Z. asiatica,
polymorphes en transpécifique chez Z. galapogoensis (Santiago-Alarcon et al. 2006), des tests
d’amplification chez Z. aurita ont été réalisés. En parallèle, des marqueurs développés chez le
pigeon domestique, Columba livia (Traxler et al. 2000), dont un était polymorphe chez la
tourterelle des Galápagos (CliµT17, Santiago-Alarcon et al. 2006), ont été ajoutés au groupe
de loci à tester. Ainsi, la qualité d’amplification de 16 microsatellites a été évaluée (tableau
3). Compte tenu du faible taux de réussite (quatre loci exploitables), des marqueurs
spécifiquement développés pour Z. aurita étaient indispensables pour pouvoir bénéficier d’un
plus grand nombre de loci. Les principales étapes du développement de ces marqueurs
(création de la banque, enrichissement pour des motifs microsatellites, séquençage des clones
positifs et définition des amorces) ont été effectuées par la société californienne Genetic
Identification Services (G.I.S., www.genetic-id-services.com). Par la suite, huit individus
issus de la même population ont été typés pour les 33 loci potentiels développés par le G.I.S.
pour tester leur qualité d’amplification et leur niveau de polymorphisme. Sur l’ensemble, dix
n’étaient pas polymorphes ou ne présentaient pas de motifs lisibles. Les 23 restants ont été
testés sur un nombre plus important d’individus afin d’affiner leur niveau de polymorphisme,
de tester la présence d’un déséquilibre de liaison et la conformité de la stucture génotypique
observée avec celle attendue sous l’hypothèse d’un équilibre de Hardy-Weinberg. De même,
des tests d’amplification transpécifique ont été réalisés chez quatre autres espèces du genre
Zenaida : Z. galapagoensis, Z. auriculata, Z. macroura et Z. meloda ainsi que chez une autre
espèce de tourterelle fréquemment rencontrée sous les mêmes latitudes, la colombe à queue

Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
30
noire, Columbina passerina. Pour toutes ces espèces, les échantillons d’ADN nous ont été
procurés par D. Santiago-Alarcon et P. Parker (Université du Missouri, St Louis, Etats Unis
d’Amérique). Les détails techniques d’amplification et de visualisation du polymorphisme
sont présentés en détail dans la note technique (Annexe 2 : Monceau et al. 2009).
Au final, 23 loci microsatellites sont disponibles pour l’étude de la variabilité
génétique chez la tourterelle à queue carrée avec un niveau de polymorphisme allant de deux
(ZaD127) à 11 allèles (Za5). Trois multiplex combinant gammes de variations alléliques non
chevauchantes, niveau de polymorphisme élevé (mimum six alléles), qualité d’amplification,
lisibilité et absence d’allèles nuls ont été constitués. Ainsi, un ensemble de sept à dix
marqueurs a été utilisé par la suite pour nos analyses (voir chapitres II, III).
Chapitre I – Présentation du modèle d’étude et des outils méthodologiques
31
Tableau 3: Marqueurs microsatellites développés chez le pigeon domestique (Columba liva)
et chez la tourterelle à ailes blanches (Zenaida asiatica). Na représente le nombre d’allèles
obtenus pour le test de polymorphisme.
Locus / N° d’accession
Séquences des amorces Taille
attendues (pb)
Tailles observées
(pb)
Qualité d'amplification
Na
Columba livia
CliµD01 F: GATTTCTCAAGCTGTAGGACT 68-120 - - -
AF188627 R: GTTTGATTTGGTTGGGCCATC
CliµD16 F: GCAGTGATAAAGTTCTGGAACA 85-134 - - -
AF188628 R: GTTTGCCTCACCGTGACATCA
CliµD17 F: TCTTACACACTCTCGACAAG 114-128 - - -
AF188629 R: GTTTCCACCCAAATGAGCAAG
CliµD19 F: CCGTTTCTTCTAATGCAC 171-198 - - -
AF192275 R: GTTTGGATTTCTGGGAGTGTATG
CliµD32 F: GAGCCATTTCAGTGAGTGACA 136-158 - - -
AF188630 R: GTTTGCAGGAGCGTGTAGAGAAGT
CliµT13 F: TCCAGAAGACACAGGCTAGT 190-212 180-1200 mauvaise -
AF188631 R: GTTTGCAAGCCCTGGTTATCTCA
CliµT17 F: ATGGGTTTGGAGATGTTTTG 208-244 220-250 correcte 3
AF188632 R: GTTTGATGGAGTTGCTATTTTGCT
Zenaida asiatica
WE2-17 F: ATC TAC CAG CCG AAG CAA 148 250 correcte 1
- R:GAG GCC AGG ATA AAG CAA CT
WE2-24 F: GGC ATA TCA CAG GAC AAG G 138-140 160 correcte 1
AF260575 R:ACT TTA AAC GCC ACA TTC AC
WU1-36 F: CCT GCC CAG CTA CTG TTT G 144 120-700 mauvaise -
- R:TTC ATG CCT TCA TTC CCA TA
WU7a-117 F: CTCAGTGTAAATATGGCAGGGAATC 194-222 210-290 correcte 7
AF260574 R:CAGGTCTTTTTGGTGGATGTCAC
WUA-38 F: GGA GGG CAC CAG AGT TG 123 190-210 correcte 7
AY428751 R:GAT AAG ACC CGA CTT TCA GC
WUE-1 F: CAG TGT GGC AGG TAC TTC A 133-151 170-190 correcte 3
AF260573 R:CTC ATT AGT GGA CCT TGG AC
WU e107 F: CGC AGG GTC TGT GTG AGT GA 126 150 correcte 1
- R:GCC CAT CCT GCA CCT AAG AG
WUJ22 F: CAG GAG CCA TCG TAC ACA T 148 170 correcte 1
AY428752 R:TGA ATT ACC CCA TCA ACA AG
WUI-69 F: AAGGGATATTATGGCAAAACC 173-189 210 correcte 1
AF260576 R:GAGCAGCAGCCCAGAAT

32
CHAPITRE II
-
POLYMORPHISME DE RESSOURCES CHEZ
LA TOURTERELLE A QUEUE CARREE

33
CHAPITRE II
-
POLYMORPHISME DE RESSOURCES
CHEZ LA TOURTERELLE A QUEUE CARREE
Quel que soit le champ disciplinaire invoqué, la validité d’une expérience scientifique,
d’un point de vue méthodologique, est basée sur trois principes : l’établissement d’un groupe
contrôle, le traitement aléatoire (randomisation, de l’anglais "random" au hasard) ainsi que la
réplication (Johnson 2002, 2006). Bien que l’utilisation d’un groupe témoin soit dans certains
cas discutable, la randomisation ne peut être effectuée que si le traitement est dupliqué
(Johnson 2006). Outre ce paramètre aléatoire, la réplication permet de limiter la surestimation
d’un effet en limitant les erreurs potentielles. Ceci est vrai, d’une part parce qu’une erreur
unique surpasse une moyenne d’erreurs indépendantes, notamment à cause du fait que les
erreurs sont distribuées aléatoirement grâce à la randomisation, mais également car la
variabilité observée sur l’effet mesuré estime l’erreur (Johnson 2006).
Cette définition de la réplication s’applique dans le cadre d’une expérimentation mais un
niveau supérieur de résolution peut être et doit être envisagé à savoir la métaréplication
(Johnson 2002, 2006, McCaffery & Ruthrauff 2004, Tallmon & Mills 2004, Tigreros et al.
ANNEXE 3
Monceau, K., Dechaume-Moncharmont, F.-X., M., Wattier, R. A., Motreuil, S. & Cézilly, F.
Territoriality vs. flocking in the Zenaida dove, Zenaida aurita: Resource polymorphism
revisited using morphological and genetic analyses.
Manuscrit à soumettre à Ecology.

Chapitre II – Polymorphisme de ressources chez la tourterelle à queue carrée
34
2009). Encore appelée réplication conceptuelle (Kelly 2006), elle désigne la réitération d’une
étude complète mais en différents lieux et temps, avec de nouveaux expérimentateurs et
surtout une autre méthode, ceci dans le seul but de renforcer la véracité de l’effet observé en
éliminant un artefact dû à des circonstances particulières. Bien que la métaréplication soit
encore peu pratiquée en biologie évolutive et en écologie (Kelly 2006), quelques études ont
d’ores et déjà été publiées (McCaffery & Ruthrauff 2004, Tigreros et al. 2009). Toutefois,
certaines expériences empiriques nécessiteraient d’être systématiquement dupliquées afin
d’éviter que leur influence ne devienne trop importante au regard de leur légitimité comme
cela est le cas pour l’article de Bateman (1948), cité 1080 fois (source Web of Knowledge), et
récemment remis en cause par Snyder & Gowaty (2007). Néanmoins, comme le précisent
dans ce cas là les auteurs, le but de ce type d’article n’est pas de faire un procès à leur
prédécesseur, mais avant tout d’informer et de faire progresser la recherche sur le thème
considéré (Snyder & Gowaty 2007).
C’est justement dans cet esprit que nous avons décidé d’appréhender sous un autre
angle l’étude réalisée par Sol et al. (2005) sur la ségrégation des ressources observée chez les
tourterelles à queue carrée de la Barbade. Bien que les individus de cette espèce soient
territoriaux et défendent un site tout au long de l’année contre l’intrusion de congénères, une
partie d’entre eux se nourrit en groupe sans qu’il y ait de comportement agonistique (Carlier
& Lefebvre 1996, Dolman et al. 1996, figure 1). En se basant sur des comparaisons
morphologiques, comportementales et écologiques (par l’étude de leurs préférences
alimentaires), Sol et al. (2005) ont déduit qu’il s’agissait d’un cas de polymorphisme de
ressources (Skúlason & Smith 1995, Smith & Skúlason 1996). Toutefois, certains points
méthodologiques sont susceptibles d’être améliorés. Le premier concerne l’absence de
réplication pour ce qui est des stratégies d’approvisionnement car un seul site territorial et un
seul site de nourrissage en groupe ont été échantillonnés et ce sur une seule et même année.
Le second point concerne la méthode de sexage, basée sur la couleur du plumage (del Hoyo et
al. 1997, Gibbs et al. 2001). Chez cette espèce, le dimorphisme sexuel est faible (chapitre I) et
la variation de coloration des plumes peut notamment être imputable à une simple variabilité
interindividuelle. De plus, les juvéniles ne peuvent pas être sexés selon cette méthode et ils
ont donc été exclus de leur analyse morphologique.
Afin, d’améliorer d’un point de vue méthodologique l’étude menée par Sol et al. (2005), un
réplicat pour chaque site a été ajouté, et ce pour deux années consécutives. De plus, nous
avons réalisé un sexage moléculaire de l’intégralité des individus adultes et juvéniles
capturés. Enfin, nous avons également réalisé une analyse complémentaire de différenciation

Chapitre II – Polymorphisme de ressources chez la tourterelle à queue carrée
35
génétique afin de déterminer l’existence potentielle d’un isolement entre sites puisque nous
disposions de marqueurs microsatellites développés chez cette espèce (Monceau et al. 2009,
annexe 2). D’un point de vue général, notre étude correspond à la définition d’une
métaréplication au sens de Johnson (2002), à savoir une réitération d’une étude
précédemment entreprise mais avec une méthodologie propre (au sens personnel du terme).
Les résultats que nous obtenons par notre méthode ne concordent pas avec l’hypothèse
du polymorphisme de ressources défendue par Sol et al. (2005). De plus, les différences
observées d’un point de vue morphologique entre les deux articles pourraient en partie être
imputables à un mauvais sexage lors de la première étude. Globalement, nos travaux montrent
non seulement l’intérêt de la métaréplication qui devrait être plus souvent rencontrée dans la
littérature mais également, et encore une fois, l’intérêt du sexage moléculaire dont nous avons
déjà affirmé l’importance pour les études concernant des espèces aviaires ayant un faible
dimorphisme sexuel.
Figure 1 : Exemple de session de capture sur un site de nourrissage en groupe (Harbour).

36
CHAPITRE III
-
HETEROZYGOTIE MULTI-LOCI
ET
APTITUDE PHENOTYPIQUE

37
CHAPITRE III
-
HETEROZYGOTIE MULTI-LOCI ET
APTITUDE PHENOTYPIQUE
REMARQUE
Ce chapitre a été rédigé comme le serait un manuscrit en construction, les données
présentées ici devant être complétées par des analyses supplémentaires (augmentation du
nombre d’individus et de loci) avant de constituer un manuscrit à soumettre à X.
I. INTRODUCTION
Depuis le début du siècle dernier, une avancée notable des connaissances dans le
domaine des relations génotype-phénotype a été réalisée avec les travaux de Shull décrivant
notamment les qualités de rendements de variétés de maïs "hybride" (Shull 1946, 1948, Crow
1998). Contrairement à son sens premier, désignant le résultat de l’union de deux espèces
proches comme par exemple le mulet (croisement entre une jument et un âne), le mot
"hybride" a tout d’abord été employé en lieu et place du terme hétérozygote (Shull 1946).
L’hétérosis (vigueur hybride), introduit par Shull en 1914 (Shull 1948), désignait et désigne
l’avantage en termes d’aptitude phénotypique d’un individu hétérozygote par rapport à un
homozygote. Toutefois, ceci est défini pour la globalité du génome du porteur. Ainsi, cette
notion est le plus souvent testée via un ensemble de croisements consanguins visant à obtenir
des individus homozygotes afin de les comparer à des individus hétérozygotes, ce qui a,
majoritairement, été développé chez les plantes (Hochholdinger & Hoecker 2007,

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
38
Krystkowiak et al. 2009). Néanmoins, cela implique de pouvoir effectuer des croisements
contrôlés afin de maîtriser le pedigree des individus ce qui est, a priori, plus difficile sur des
populations naturelles (voir toutefois Marr et al. 2002). Cependant, depuis l’avènement de
l’écologie moléculaire à la fin du XXème siècle et le développement de techniques de
visualisation de la variabilité génétique, la question du maintien du niveau de polymorphisme
au sein des populations a pu être investie sous un nouvel angle (Mitton 1994). En effet, de
nombreuses études ont tenté de corréler un niveau d’hétérozygotie pour des marqueurs
génétiques (enzymes et microsatellites généralement) avec un ou plusieurs paramètres
reflétant l’aptitude phénotypique des individus (pour une revue se référer à Mitton 1994,
David 1998, Chapman et al. 2009). Les indices représentant l’aptitude phénotypique peuvent
être de différentes natures. Chez les plantes telles que Clematis acerifolia, la surface et le
nombre de feuilles ont été utilisés, compte tenu de leur implication dans la photosynthèse et,
par extension, pour la survie (López-Pujol et al. 2008). Chez les animaux, les paramètres liés
à la survie (âge, durée de vie) ont également été employés mais aussi des traits impliqués dans
la reproduction, le comportement territorial, l’immunité, les traits sexuels (ornementations,
chants), la taille corporelle et les asymétries de développement (Coulson et al. 1998, 1999,
Merilä et al. 2003, Neff 2004a, Seddon et al. 2004, Lieutenant-Gosselin & Bernatchez 2006,
Kempenaers 2007, Da Silva et al. 2008, Mainguy et al. 2009).
Dans le cadre de notre étude sur la tourterelle à queue carrée, la corrélation entre
hétérozygotie et aptitude phénotypique pouvait être testée sous différentes formes. Tout
d’abord, l’hétérozygotie pouvait être comparée entre les deux stratégies d’approvisionnement
pratiquées par les tourterelles à queue carrée à la Barbade. En effet, en milieu tropical, bon
nombre d’espèces défendent leur territoire tout au long de l’année contre des intrusions de
congénères (Stutchbury & Morton 2001), ce qui est notamment le cas des tourterelles à queue
carrée à la Barbade (chapitre II). En conséquence d’un taux de survie adulte supposé être
particulièrement élevé dans ce type de milieu, le remplacement des individus territoriaux
apparait relativement faible, conduisant ainsi à une forte compétition pour l’acquisition d’un
territoire (Greenberg & Gradwohl 1997, Morton et al. 2000, Stutchbury & Morton 2008). De
ce fait, une partie des individus de la population ne possède pas de territoire leur permettant
entre autres de se nourrir. Ceux-ci sont qualifiés de "flotteurs" notamment à cause de leur
prospection sur différents sites territoriaux d’où ils se font activement chassés (Brown 1969,
Winker 1998, Sol et al. 2005). Ces individus peuvent potentiellement s’agréger sur des sites
de nourrissage collectif, tout comme cela a pu être décrit chez Z. aurita (chapitre II). Ainsi,

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
39
flotteurs et individus territoriaux peuvent différer sur des critères liés à leur capacité à
acquérir et défendre un site, témoins de leur capacité compétitrice (Smith & Arcese 1989,
Stutchbury 1992, Lozano 1994). De ce fait, la capacité à acquérir et défendre un territoire peut
être considérée comme étant le reflet de l’aptitude phénotypique de l’individu, ce qui pourrait
être corrélé à son degré d’hétérozygotie, qui serait donc supérieur chez les territoriaux par
rapport aux flotteurs. Ce type de relation a notamment été démontré chez les tétras lyre,
Tetrao tetrix, chez lesquels les mâles les plus hétérozygotes avaient de meilleures chances
d’obtenir un territoire (Höglund et al. 2002). Toutefois, ceci ne pouvait concerner que les
adultes puisque que nous nous plaçons implicitement dans un contexte de sélection sexuelle,
l’acquisition d’un territoire étant liée à l’initiation de la reproduction. Pour aller plus loin sur
notre modèle d’étude, certains traits tels que la survie sont plus variables dans les premiers
stades de développement notamment à cause du fait que les phénotypes peu aptes à survivre
sont éliminés plus précocement (Koehn & Gaffney 1984, David 1998, Cohas et al. 2009).
Compte tenu de l’avantage que leur confère leur variabilité génétique, les individus
hétérozygotes pourraient avoir une probabilité de survie supérieure à celle des individus plus
homozygotes. Ces derniers disparaissant, la variabilité du niveau d’hétérozygotie dans la
population devrait décroître avec l’âge, entraînant également une augmentation du niveau
moyen comme cela a récemment été montré chez la marmotte des Alpes, Marmota marmota
(Cohas et al. 2009). Dans notre cas, étant donné que nous ne sommes pas en mesure
d’attribuer un âge plus précis que la catégorisation en adultes et juvéniles, cette hypothèse
devrait se traduire par une variabilité du niveau d’hétérozygotie chez les juvéniles supérieure
à celle observée chez les adultes, ceux-ci ayant un niveau moyen supérieur aux jeunes.
Comme cela a été évoqué précédemment, des caractères tels que la survie ou les traits liés à la
reproduction ont souvent été employés pour ce type d’approche. Toutefois, ces paramètres ne
seraient pas pertinents à ce stade de l’étude, étant donné que nous ne disposons pas, à l’heure
actuelle, de suffisamment de données. Néanmoins, ces traits classiques sembleraient pouvoir
être estimés avec une bonne sensibilité par l’analyse de l’asymétrie fluctuante (Lens et al.
2002a). Celle-ci est définie comme étant un écart à la symétrie bilatérale pour les caractères
pairs (Palmer & Strobeck 1986). L’amplitude de ces déviations reflèterait la capacité de
l’individu à produire un phénotype le plus proche de la symétrie bilatérale idéale, quel que
soit le niveau de stress (parasitisme par exemple) rencontré au cours du développement
(Palmer & Strobeck 1992, Clarke 1993, Palmer 1994, van Dongen & Lens 2000, Debat &
David 2001). Par un effet tampon, dû à la plus grande diversité allélique, les individus
hétérozygotes auraient potentiellement un développement plus stable que les homozygotes et

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
40
en conséquence leur symétrie bilatérale pour les tarses et les ailes serait meilleure (Clarke
1993, Leamy & Klingenberg 2005). Toutefois, cette analyse ne pouvait concerner, encore une
fois, que les adultes car les juvéniles sont, par définition, en cours de développement et donc,
leur morphologie est encore soumise à modification.
Afin de tester nos différentes hypothèses quant à la relation entre génotype et aptitude
phénotypique, un ensemble de marqueurs microsatellites polymorphes ainsi que divers
estimateurs de l’hétérozygotie ont été utilisés. Toutefois, le choix d’un tel indice n’est pas aisé
car ils sont non seulement nombreux, mais également déclinés sous diverses formes et
l’utilisation de chacun peut être soumise à controverse. Par exemple, une mesure basique de la
diversité génétique est la proportion de loci pour lesquels l’individu est hétérozygote (noté par
la suite H, Coltman et al. 1998, Coulson et al. 1998, 1999, Marshall & Spalton 2000, Slate et
al. 2000, Hansson et al. 2001, Rossiter et al. 2001, Höglund et al. 2002, Slate & Pemberton
2002, Foerster et al. 2003, Richardson et al. 2004, Kleven & Lifjeld 2005, MacDougall-
Shackleton et al. 2005, Dowling & Mulder 2006, Freeman-Gallant et al. 2006, Jouventin et al.
2007, Beltran et al. 2008, Da Silva et al. 2008, Ortego et al. 2009, Ruiz-López et al. 2009). Le
problème est que cet indice peut être biaisé lorsque les individus ne sont pas tous typés avec
les mêmes marqueurs, que le nombre de loci est faible et/ou que ceux-ci diffèrent dans leur
niveau de polymorphisme (Aparicio et al. 2006). Pour éviter ces problèmes, Coltman et al.
(1999) ont proposé un indice d’hétérozygotie standardisé par l’hétérozygotie moyenne aux
loci pour lesquels l’individu a été typé (noté par la suite SH) qui a été tout autant utilisé que le
premier (Slate et al. 2000, Amos et al. 2001, Slate & Pemberton 2002, Master et al. 2003,
Kleven & Lifjeld 2005, Tarvin et al. 2005, Cohas et al. 2006, 2007, 2009, Bishop et al. 2007,
Rubenstein 2007, Fossøy et al. 2008, Lyons et al. 2009, Ruiz-López et al. 2009). H ou SH
(souvent regroupés sous la désignation MLH pour "multilocus heterozygosity") ont souvent
été associés et comparés avec un autre paramètre, mean d², introduit par Coulson et al. 1998
(noté par la suite MD, voir Coltman & Slate 2003). Celui-ci, valable uniquement pour les
marqueurs microsatellites, est dérivé d’un indice utilisé au niveau populationnel développé
par Goldstein et al. (1995) et basé sur la relation entre la distance - en unités de répétition -
entre deux allèles et le temps écoulé depuis leur coalescence. Au niveau individuel, MD
représente le niveau de parenté entre les haplotypes des parents de l’individu. Ce dernier a
également été décliné sous différentes formes, par exemple en le standardisant par la variance
de la différence au carré par locus afin de limiter la trop forte influence des loci hautement
polymorphes (Hedrick et al. 2001, Slate & Pemberton 2002). D’un point de vue de la

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
41
résolution de l’analyse, H, SH et MD sembleraient être tous trois surpassés par l’indice de
parenté interne (noté par la suite IR pour "internal relatedness") développé par Amos et al.
(2001). Cet indice, bien que moins répandu, offre l’avantage d’être également pondéré par les
fréquences alléliques en donnant plus de poids aux allèles rares (Masters et al. 2003, Tarvin et
al. 2005, Oh & Badyaev 2006, Lindstedt et al. 2007, Ortego et al. 2007a, Rubenstein 2007,
Rijks et al. 2008, Acevedo-Whitehouse et al. 2009, Ruiz-López et al. 2009). Enfin, plus
récemment, une nouvelle méthode a été proposée par Aparicio et al. (2006) pour estimer à
l’origine l’homozygotie (noté par la suite HL pour "homozygosity by loci") et, par extension,
l’hétérozygotie prenant en compte la variabilité allélique de chaque locus (Ortego et al. 2007a,
2007b, 2009, Rijks et al. 2008, Acevedo-Whitehouse et al. 2009, Haag-Liautard et al. 2009,
Ruiz-López et al. 2009). Un autre avantage non négligeable de HL est que le niveau de
significativité des corrélations potentiellement observées avec IR serait atteint avec un effectif
moitié moins important (Aparicio et al. 2006).
Compte tenu du fait que le débat reste toujours ouvert quant à l’efficacité de chaque
index d’hétérozygotie (Chapman et al. 2009), le choix d’un indice plutôt qu’un autre est
complexe, notamment compte tenu des qualités et défauts de chacun. De ce fait, nous avons
fait le choix, dans le cadre de notre étude visant à vérifier l’existence d’une relation entre
génétique et aptitude phénotypique, de ne retenir que quatre des cinq indices précédemment
introduits à savoir, SH, MD, IR et HL. H a été délibérément exclu de notre étude dû à son
manque de fiabilité lorsque tous les individus n’étaient pas typés au même loci, ce qui est le
cas dans notre étude.
II. MATERIEL & METHODES
1. Modèle biologique
La tourterelle à queue carrée, Zenaida aurita, est un colombidé endémique des
Caraïbes. Cette espèce est monogame et les couples s’établissent sur des territoires qu’ils
défendent tout au long de l’année contre l’intrusion de congénères (Lefebvre et al. 1996).
Néanmoins, sur certains sites de production et stockage agraire, elles adoptent une stratégie
alternative d’approvisionnement en groupe où peu de comportements agonistiques sont
observés entre congénères (Carlier & Lefebvre 1996, Dolman et al. 1996, Sol et al. 2005,

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
42
chapitre II). Le dimorphisme sexuel étant peu marqué un sexage moléculaire a été réalisé pour
tous les individus (chapitre I). Pour ce qui est de l’âge, les juvéniles sont facilement
discernables des adultes par leur plumage grisâtre et l’absence de taches iridescentes autour
du cou. Pour plus de détails sur l’espèce et les méthodes de sexage moléculaire, se référer au
chapitre I.
2. Sites d’études, captures, prises de mesures et prélèvements
sanguins
Tout comme les travaux précédents (chapitre I et II), les tourterelles ont été capturées
sur quatre sites distincts sur l’île de la Barbade : deux sites territoriaux, Bellairs et Sunset
Crest et deux sites de nourrissage en groupe, Roberts et Harbour. Etant donné qu’aucune
différence d’ordre génétique ou morphologique n’a pu être mise en évidence précédemment
entre les deux sites territoriaux d’une part et les deux sites de nourrissage en groupe d’autre
part (chapitre II), ceux-ci ont été regroupés afin de n’avoir que deux groupes tests.
La méthode d’acquisition des données morphométriques et des prélèvements sanguins
nécessaires aux analyses moléculaires suivaient le même protocole que celui présenté au
chapitre I. Un point important pour une partie des analyses suivantes est à souligner ici. Les
mesures des tarses et des ailes ont toujours été réalisées en double afin de pouvoir évaluer
l’erreur de mesure ainsi que la répétabilité de celles-ci, toutes les mesures n’ayant pas été
réalisées par le même expérimentateur.
Au total, sur une période de trois ans, 359 tourterelles ont été utilisées pour cette étude
dont 276 adultes et 83 juvéniles. Le détail des effectifs selon les stratégies
d’approvisionnement et les classes d’âge est présenté dans le tableau 1.
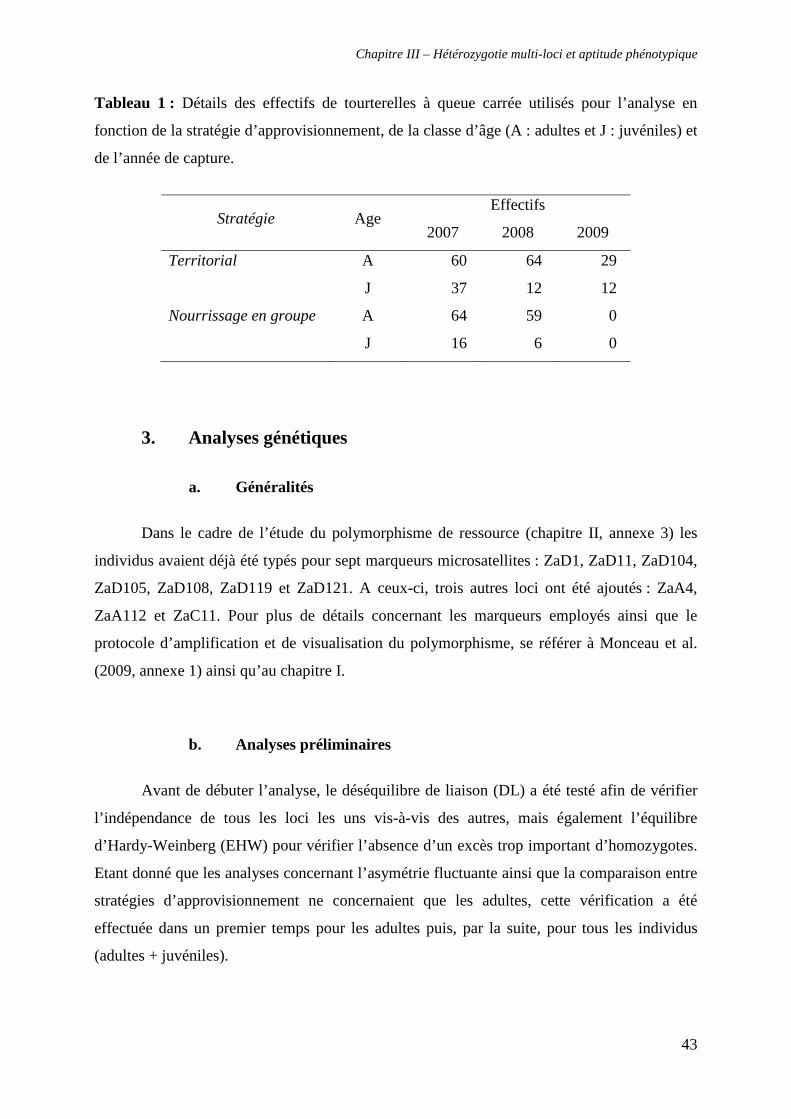
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
43
Tableau 1 : Détails des effectifs de tourterelles à queue carrée utilisés pour l’analyse en
fonction de la stratégie d’approvisionnement, de la classe d’âge (A : adultes et J : juvéniles) et
de l’année de capture.
Stratégie Age Effectifs
2007 2008 2009
Territorial A 60 64 29
J 37 12 12
Nourrissage en groupe A 64 59 0
J 16 6 0
3. Analyses génétiques
a. Généralités
Dans le cadre de l’étude du polymorphisme de ressource (chapitre II, annexe 3) les
individus avaient déjà été typés pour sept marqueurs microsatellites : ZaD1, ZaD11, ZaD104,
ZaD105, ZaD108, ZaD119 et ZaD121. A ceux-ci, trois autres loci ont été ajoutés : ZaA4,
ZaA112 et ZaC11. Pour plus de détails concernant les marqueurs employés ainsi que le
protocole d’amplification et de visualisation du polymorphisme, se référer à Monceau et al.
(2009, annexe 1) ainsi qu’au chapitre I.
b. Analyses préliminaires
Avant de débuter l’analyse, le déséquilibre de liaison (DL) a été testé afin de vérifier
l’indépendance de tous les loci les uns vis-à-vis des autres, mais également l’équilibre
d’Hardy-Weinberg (EHW) pour vérifier l’absence d’un excès trop important d’homozygotes.
Etant donné que les analyses concernant l’asymétrie fluctuante ainsi que la comparaison entre
stratégies d’approvisionnement ne concernaient que les adultes, cette vérification a été
effectuée dans un premier temps pour les adultes puis, par la suite, pour tous les individus
(adultes + juvéniles).

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
44
c. Mesures de l’hétérozygotie
Les quatre indices suivant l’hétérozygotie ont été calculés pour chaque individu :
• SH : Hétérozygotie standardisée par l’hétérozygotie moyenne des loci typés (Coltman et
al. 1999)
H t / moyenne Ht
avec :
Ht proportion de loci typés hétérozygotes
• IR : Indice de parenté (Amos et al. 2001)
(2H - ∑f i) / (2N - ∑f i)
avec :
H le nombre de loci homozygotes
N le nombre de loci
f i la fréquence de l’allèle i
• MD : Mean d² standardisé par la valeur maximum individuelle observée à chaque locus
(Coulson et al. 1998, Hedrick et al. 2001)
(1 / N) ∑ [(ni1 - ni2)² / (max (ni1 - ni2))²]
avec :
N le nombre de loci
ni1 et ni2 le nombre d’unité de répétitions des allèles 1 et 2
• HL : Homozygotie pondérée par locus (Aparicio et al. 2006)
∑Eh / (∑Eh + ∑Ej)
avec :
Eh l’hétérozygotie attendue aux loci où l’individu est homozygote
Ej l’hétérozygotie attendue aux loci où l’individu est hétérozygote

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
45
Les corrélations entre paires d’indices ont été effectuées, et ce sur l’ensemble de
l’échantillon (adultes et juvéniles) grâce à un test de Spearman (les variables ne suivant pas
une loi normale) afin de préciser leur relation.
4. Relation entre hétérozygotie et aptitude phénotypique
a. Hétérozygotie, stratégie d’approvisionnement et âge (adultes et
juvéniles)
Après avoir vérifié la normalité des indices, ceux-ci ont été comparés entre stratégies
d’approvisionnement (territoriaux vs nourrissage en groupe), mais également par âge (adultes
vs juvéniles). Dans ce dernier cas, compte tenu du fait que nous nous attendons à ce qu’il y ait
une augmentation du niveau d’hétérozygotie global au stade adulte, un test unilatéral a été
réalisé à la place du test bilatéral.
Etant donné que nous ne disposions que de deux groupes pour tester les différences en
termes de variance entre adultes et juvéniles, un simple test d’homogénéité des variances de
Brown-Forsythe (dérivé non paramétrique du test classique de Levene, Brown & Forsythe
1974 cités dans Hui et al. 2008) a été réalisé.
b. Hétérozygotie et asymétrie fluctuante (adultes uniquement)
Avant de s’intéresser à l’asymétrie fluctuante (AF), plusieurs vérifications ont été
effectuées. Celles-ci comportaient quatre étapes. Tout d’abord, l’effet de l’erreur de mesure
par rapport à l’asymétrie globale a dû être évalué afin de s’assurer que celle-ci n’était pas
supérieure à l’asymétrie observable. Puis, la présence d’autres formes d’asymétrie, à savoir
l’antisymétrie (AS) et l’asymétrie directionnelle (AD) ont dû être écartées (Palmer &
Strobeck 1986, 1992, 2003, Palmer 1994, figure 1). Enfin, l’indépendance des indices d’AF
vis-à-vis de la taille du caractère considéré devait être testée. Ces précautions étaient
nécessaires car l’analyse de l’AF peut être faussée dans le cas où l’erreur de mesure serait trop
importante ou que les autres formes d’asymétrie se mélangeraient et se surimposeraient à
l’AF.
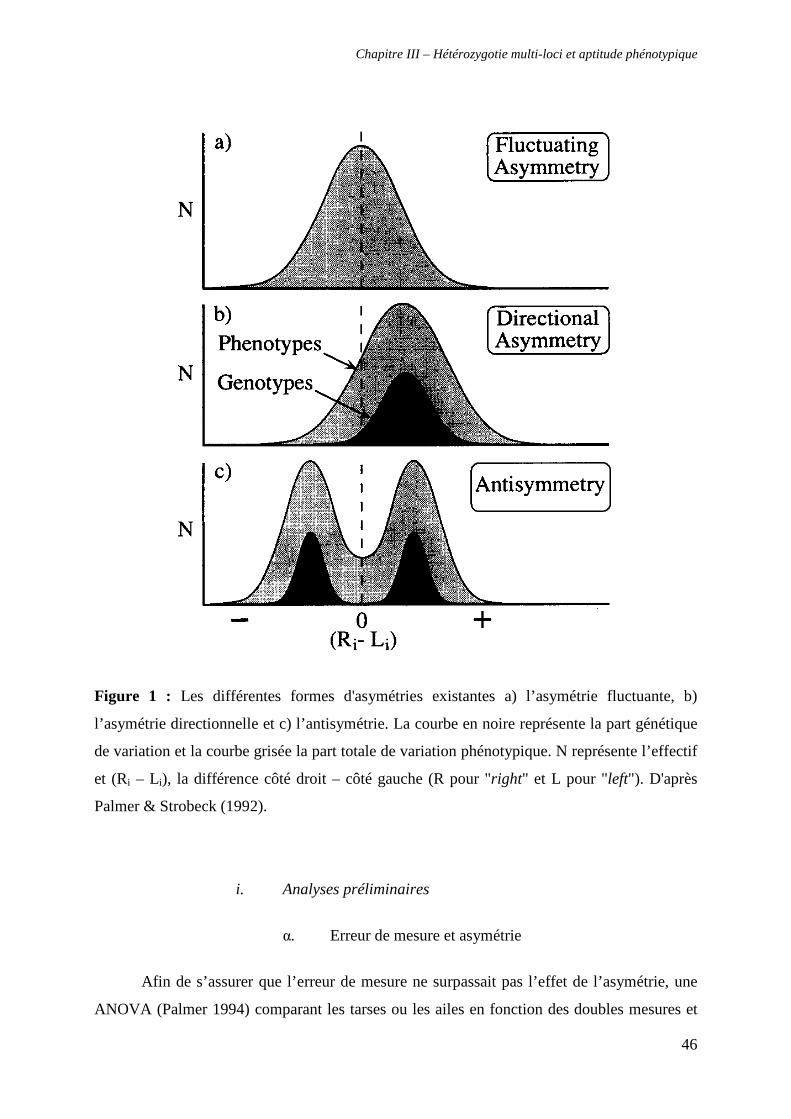
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
46
Figure 1 : Les différentes formes d'asymétries existantes a) l’asymétrie fluctuante, b)
l’asymétrie directionnelle et c) l’antisymétrie. La courbe en noire représente la part génétique
de variation et la courbe grisée la part totale de variation phénotypique. N représente l’effectif
et (Ri – Li), la différence côté droit – côté gauche (R pour "right" et L pour "left"). D'après
Palmer & Strobeck (1992).
i. Analyses préliminaires
α. Erreur de mesure et asymétrie
Afin de s’assurer que l’erreur de mesure ne surpassait pas l’effet de l’asymétrie, une
ANOVA (Palmer 1994) comparant les tarses ou les ailes en fonction des doubles mesures et

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
47
des côtés a été effectuée afin de vérifier que l’asymétrie potentiellement existante était
supérieure à l’effet de l’erreur de mesure. Ceci a été réalisé après vérification de la conformité
des assomptions de l’ANOVA (homogénéité des variances par un test de Levene et normalité
par un test de Shapiro-Wilk). De plus, la répétabilité des mesures a été estimée afin de
simplifier les analyses en utilisant les moyennes des doubles mesures (Lessells & Boag 1987).
β. Antisymétrie (AS)
L’antisymétrie (AS) est définie comme un écart à la symétrie bilatérale dû au
développement prononcé d’un caractère, et ce aussi bien à gauche qu’à droite (Palmer &
Strobeck 1986, 1992, 2003, Palmer 1994, figure 1c). L’AS se manifeste par un écart à la
distribution normale des différences droite-gauche. Cet écart peut se traduire de différentes
façons : par une asymétrie de la distribution et/ou par l’aplatissement de celle-ci (figure 2).
Afin de tester la présence d’AS, la normalité de la distribution du caractère a tout
d’abord été testée (test de Shapiro-Wilk). Dans le cas où celle-ci ne suivait pas une loi
normale, la forme générale de la distribution a été testée avec le coefficient d’aplatissement ou
kurtosis (K) puis comparée à la valeur théorique du t de Student pour un degré de liberté
infini (t0,05 ; ∞= 1,96) avec :
tk = K / sK
où K est la valeur calculée de l’aplatissement
sK = √ [(24n(n-1)² / (n-3)(n-2)(n+3)(n+5)] est l’erreur standard de K et n la taille de
l’échantillon.
Si K est négatif, la distribution est platykurtique (distribution large ou bimodale, pic avec
queues courtes) et si K est positif, elle est leptokurtique (aplatie avec un faible pic et de
longues queues) (figure 2a et b).
De même, la symétrie de la distribution a été estimée par le coefficient de symétrie ou
skewness (S) puis comparée à la valeur théorique du t de Student pour un degré de liberté
infini (t0,05 ; ∞= 1,96) avec :
ts = S / sS
où S est la valeur calculée de la symétrie,
sS = √[(6n(n-1) / (n-2)(n+1)(n+3)] est l’erreur standard de S et n la taille de l’échantillon.
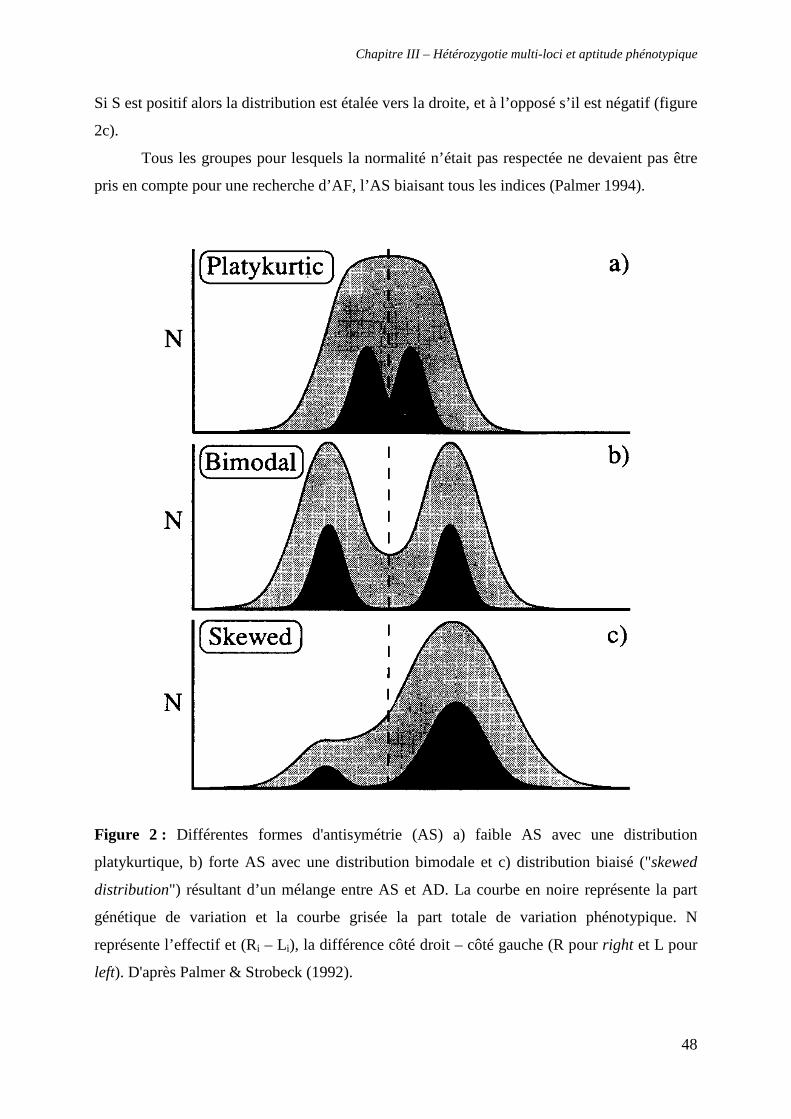
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
48
Si S est positif alors la distribution est étalée vers la droite, et à l’opposé s’il est négatif (figure
2c).
Tous les groupes pour lesquels la normalité n’était pas respectée ne devaient pas être
pris en compte pour une recherche d’AF, l’AS biaisant tous les indices (Palmer 1994).
Figure 2 : Différentes formes d'antisymétrie (AS) a) faible AS avec une distribution
platykurtique, b) forte AS avec une distribution bimodale et c) distribution biaisé ("skewed
distribution") résultant d’un mélange entre AS et AD. La courbe en noire représente la part
génétique de variation et la courbe grisée la part totale de variation phénotypique. N
représente l’effectif et (Ri – Li), la différence côté droit – côté gauche (R pour right et L pour
left). D'après Palmer & Strobeck (1992).

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
49
γ. Asymétrie directionnelle (AD)
Une asymétrie directionnelle est définie comme une différence de symétrie
systématiquement exprimée d’un même côté (Palmer & Strobeck 1986, 1992, 2003, Palmer
1994, figure 1b). Afin de vérifier son existence, les moyennes des caractères droits et gauches
ont été comparées par un test t.
δ. Indépendance de la différence droite - gauche vis-à-vis
de la taille des individus
Afin d’éviter qu’un effet de la taille du caractère puisse biaiser les résultats,
l’indépendance de la taille a dû être testée. Pour cela, une régression linéaire de la différence
absolue droite - gauche sur la moyenne du caractère a été réalisée après vérification de la
normalité des distributions (test de Shapiro-Wilk).
ii. Test de la relation entre hétérozygotie et asymétrie fluctuante
Tout comme pour l’hétérozygotie, différents indices étaient disponibles. Les plus
couramment employés, du fait de leur simplicité de manipulation et d’interprétation, étaient la
différence droite - gauche (FA4) ainsi que la différence absolue (FA1) (Palmer 1994).
Après avoir vérifié la normalité des indices d’AF (FA1 et FA4) et d’hétérozygotie (test
de Shapiro-Wilk), les régressions linéaires entre ces deux types de variables ont été
effectuées. Afin d’estimer la qualité de celles-ci, la distribution des résidus a été vérifiée afin
de savoir si elle s’écartait de la normalité (test de Shapiro-Wilk).
5. Logiciels de traitement des données
Le traitement des données de génétique des populations, à savoir le déséquilibre de
liaison et l’équilibre d’Hardy-Weinberg, a été réalisé grâce au logiciel GENEPOP (Raymond
& Rousset 1995). L’estimation de la fréquence des allèles nuls (selon la méthode décrite dans
Allen et al. 1995, W. Amos com. pers.), ainsi que le calcul des différents indices
d’hétérozygotie ont été obtenus grâce à la macro Excel "IRmacroN4"

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
50
(http://www.zoo.cam.ac.uk/zoostaff/amos) développée par W. Amos (Department of
Zoology, Cambridge University, UK).
L’ensemble du traitement statistique des données a été réalisé avec le logiciel R 2.9.1
(R Development Core Team 2008) implémenté des packages "car" et "lawstat" pour les
analyses de variances, "nlme" pour l’erreur de mesure et la répétabilité ainsi que "Hmisc"
pour les outils graphiques.
III. RESULTATS
1. Analyses génétiques
a. Analyses préliminaires
Pour chacun des deux groupes (adultes seuls et adultes + juvéniles), aucun
déséquilibre de liaison entre paires de loci n’a été détecté. Pour l’ensemble des loci, la
diversité allélique était en moyenne de 9,1 allèles par locus (6 à 14 allèles). En ce qui
concerne l’équilibre d’Hardy Weinberg (EHW), tous les loci étaient à l’équilibre, excepté
ZaA112 et ZaC11 dans les deux cas (tableau 2). Pour ZaC11, l’excès d’homozygotes constaté
étaient potentiellement imputable à la présence d’allèles nuls dont la fréquence était
supérieure à 5%, alors que pour ZaA112, l’écart à l’équilibre d’Hardy-Weinberg été causé par
un excès d’hétérozygotes (tableau 2). Pour éviter tous biais dans les analyses, ces deux loci
ont été exclus.
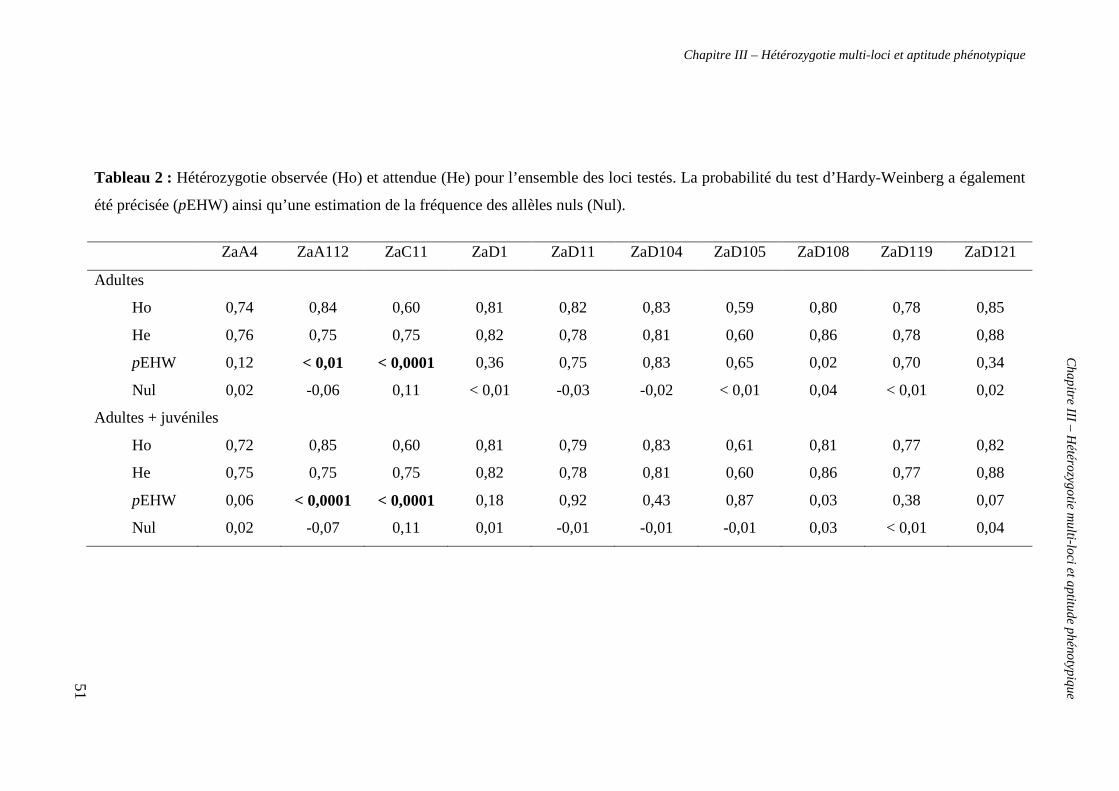
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
Ch
ap
itre III –
Hé
téro
zygo
tie m
ulti-lo
ci et a
ptitu
de
ph
éno
typiq
ue
51
Tableau 2 : Hétérozygotie observée (Ho) et attendue (He) pour l’ensemble des loci testés. La probabilité du test d’Hardy-Weinberg a également
été précisée (pEHW) ainsi qu’une estimation de la fréquence des allèles nuls (Nul).
ZaA4 ZaA112 ZaC11 ZaD1 ZaD11 ZaD104 ZaD105 ZaD108 ZaD119 ZaD121
Adultes
Ho 0,74 0,84 0,60 0,81 0,82 0,83 0,59 0,80 0,78 0,85
He 0,76 0,75 0,75 0,82 0,78 0,81 0,60 0,86 0,78 0,88
pEHW 0,12 < 0,01 < 0,0001 0,36 0,75 0,83 0,65 0,02 0,70 0,34
Nul 0,02 -0,06 0,11 < 0,01 -0,03 -0,02 < 0,01 0,04 < 0,01 0,02
Adultes + juvéniles
Ho 0,72 0,85 0,60 0,81 0,79 0,83 0,61 0,81 0,77 0,82
He 0,75 0,75 0,75 0,82 0,78 0,81 0,60 0,86 0,77 0,88
pEHW 0,06 < 0,0001 < 0,0001 0,18 0,92 0,43 0,87 0,03 0,38 0,07
Nul 0,02 -0,07 0,11 0,01 -0,01 -0,01 -0,01 0,03 < 0,01 0,04
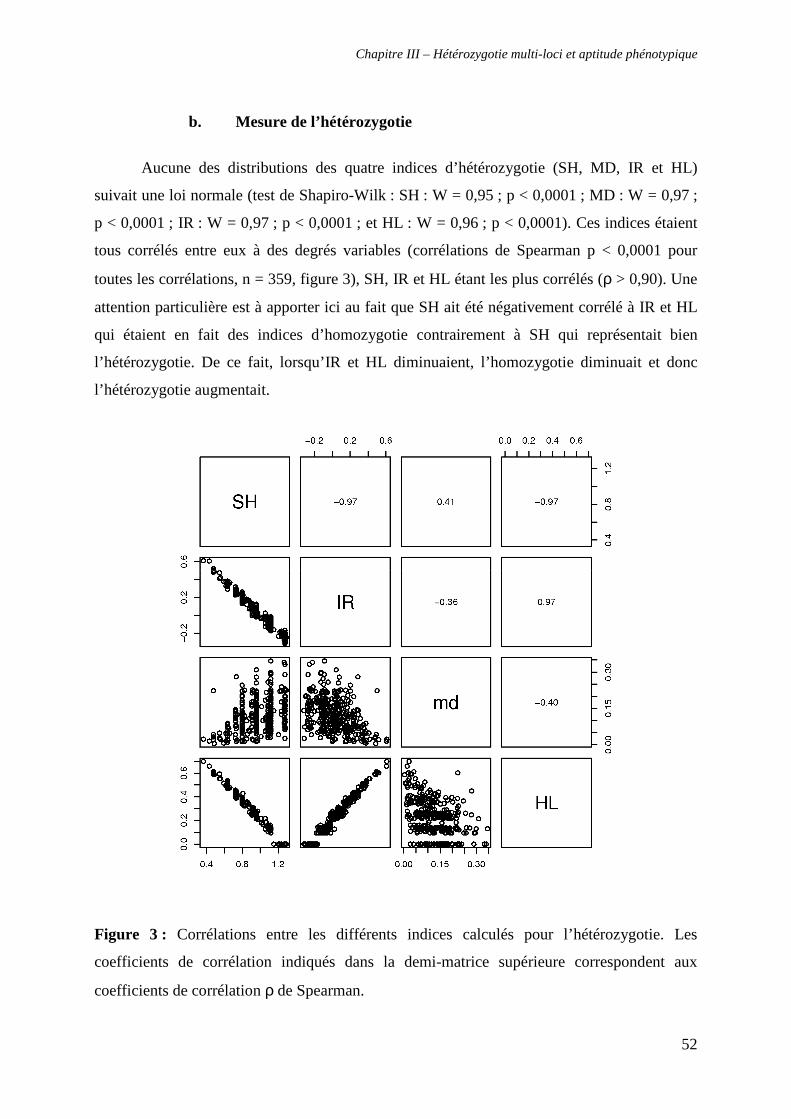
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
52
b. Mesure de l’hétérozygotie
Aucune des distributions des quatre indices d’hétérozygotie (SH, MD, IR et HL)
suivait une loi normale (test de Shapiro-Wilk : SH : W = 0,95 ; p < 0,0001 ; MD : W = 0,97 ;
p < 0,0001 ; IR : W = 0,97 ; p < 0,0001 ; et HL : W = 0,96 ; p < 0,0001). Ces indices étaient
tous corrélés entre eux à des degrés variables (corrélations de Spearman p < 0,0001 pour
toutes les corrélations, n = 359, figure 3), SH, IR et HL étant les plus corrélés (ρ > 0,90). Une
attention particulière est à apporter ici au fait que SH ait été négativement corrélé à IR et HL
qui étaient en fait des indices d’homozygotie contrairement à SH qui représentait bien
l’hétérozygotie. De ce fait, lorsqu’IR et HL diminuaient, l’homozygotie diminuait et donc
l’hétérozygotie augmentait.
Figure 3 : Corrélations entre les différents indices calculés pour l’hétérozygotie. Les
coefficients de corrélation indiqués dans la demi-matrice supérieure correspondent aux
coefficients de corrélation ρ de Spearman.

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
53
2. Relation entre hétérozygotie et aptitude phénotypique
a. Hétérozygotie, stratégie d’approvisionnement et âge
i. Hétérozygotie et stratégie d’approvisionnement
Aucune différence, en termes de niveau d’hétérozygotie mesurée par SH, MD, IR et
HL n’a été décelée entre stratégies d’approvisionnement (test de Wilcoxon, SH : W = 9828,5 ;
p = 0,52 ; MD : W = 9589 ; p = 0,79 ; IR : W = 8563 ; p = 0,20 et HL : W = 8722 ; p = 0,30).
ii. Hétérozygotie et âge
Les variances de SH, MD, IR et HL ne différaient pas entre juvéniles et adultes même
si une tendance à la réduction du stade juvénile au stade adulte était observée pour IR et HL
(tableau 3).
Les médianes étaient différentes selon SH, IR et HL (test unilatéral de Wilcoxon : SH :
W = 9912,5 ; p = 0,03 ; IR : W = 12960 ; p = 0,03 et HL : W = 13184 ; p = 0,02 ; figure 4)
mais pas selon MD (W = 11053 ; p = 0,31 ; figure 4). En fin de compte, les adultes tendaient à
avoir une hétérozygotie supérieure à celle observée chez les jeunes avec une possible
réduction de la variance du stade juvénile au stade adulte.
Tableau 3 : Déviations absolues de la médiane des différents indices d'hétérozygotie.
L’homogénéité des déviations à la médiane a été testée au moyen d'un test de Brown-Forsythe
dont le score ainsi que la probabilité associée sont respectivement représentés par B et p.
Juvéniles Adultes B p
SH 0,24 0,24 3,34 0,07
MD 0,07 0,07 0,57 0,57
HL 0,19 0,15 3,84 0,05
IR 0,21 0,18 3,86 0,05
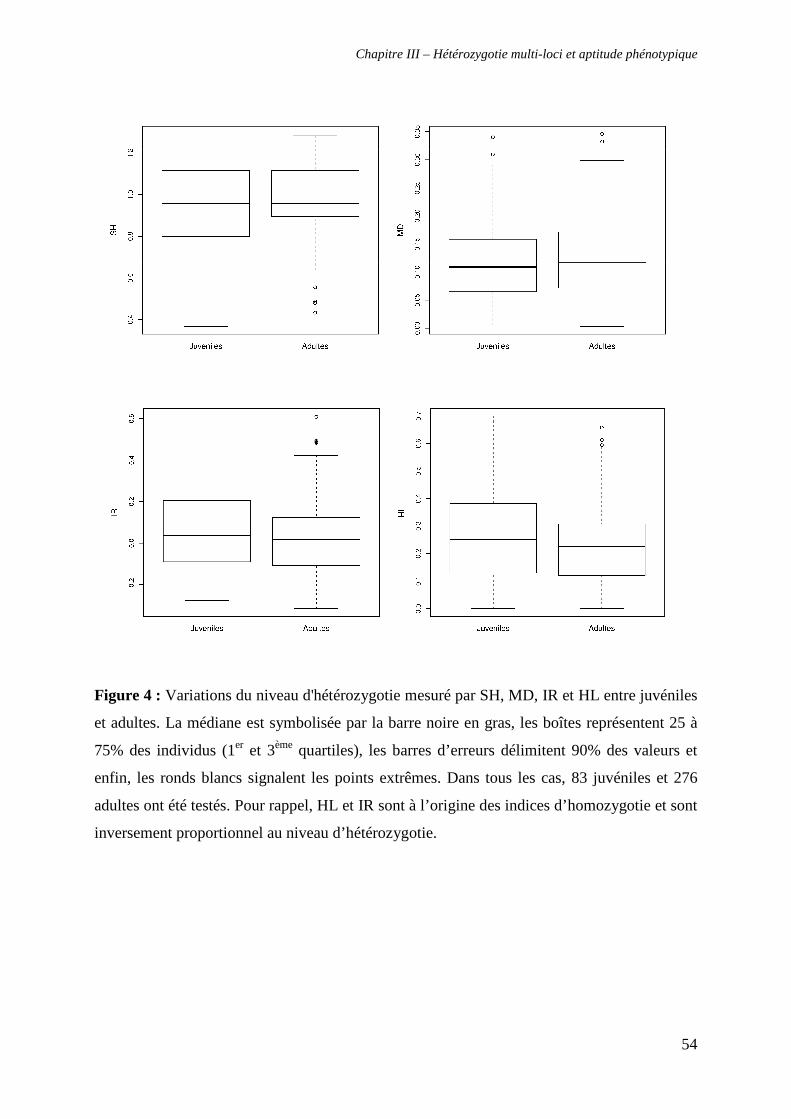
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
54
Figure 4 : Variations du niveau d'hétérozygotie mesuré par SH, MD, IR et HL entre juvéniles
et adultes. La médiane est symbolisée par la barre noire en gras, les boîtes représentent 25 à
75% des individus (1er et 3ème quartiles), les barres d’erreurs délimitent 90% des valeurs et
enfin, les ronds blancs signalent les points extrêmes. Dans tous les cas, 83 juvéniles et 276
adultes ont été testés. Pour rappel, HL et IR sont à l’origine des indices d’homozygotie et sont
inversement proportionnel au niveau d’hétérozygotie.

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
55
b. Hétérozygotie et asymétrie fluctuante
i. Erreur de mesure et asymétrie
Que ce soit pour les ailes ou les tarses, l’interaction entre le réplicat de la mesure et le
côté était significative, indiquant que l’erreur de mesure ne surpassait pas l’asymétrie (ailes :
F275, 1103 = 7,62 ; p < 0,0001 et tarses : F275, 1103 = 1,96 ; p < 0,0001). Globalement, l’erreur de
mesure était faible (2,50 < EM < 6,50 %) excepté pour les tarses droits où elle était un peu
plus importante (22,59%). Néanmoins, la répétabilité dans les deux cas étant bonne (0,77 < R
< 0,98), la suite des analyses a donc été réalisée sur la moyenne des doubles mesures.
ii. Antisymétrie (AS)
Pour l’ensemble des individus, la différence signée entre côté droit et côté gauche ne
suivait pas une loi normale, aussi bien pour les ailes que pour les tarses (tableau 4) et ce
même après transformation (logarithme ou Box-Cox). La présence d’AS a donc été détectée
chez les mâles pour les ailes et chez les femelles pour les tarses (tableau 4). En ce qui
concerne les mâles, un individu présentait un score extrême (figure 5a). L’analyse a donc été
reconduite après avoir exclu cet individu, l’écart entre l’aile droite et l’aile gauche pouvant
dans ce cas être expliquée par le mauvais état des plumes (notamment au niveau des rémiges
primaires). Cependant, bien que la distribution en ait été améliorée (figure 5b), celle-ci ne
suivait toujours pas une loi normale (W = 0,98 ; p = 0,04). Dans le cas des femelles, rien ne
semblait désigner l’existence de points extrêmes pouvant potentiellement biaiser les analyses
(figure 6). Les origines de la forme leptokurtique de ces distributions étant variées (Palmer &
Strobeck 1992, 2003), seuls les tarses pour les mâles et les ailes pour les femelles ont été
conservés pour les analyses subséquentes.
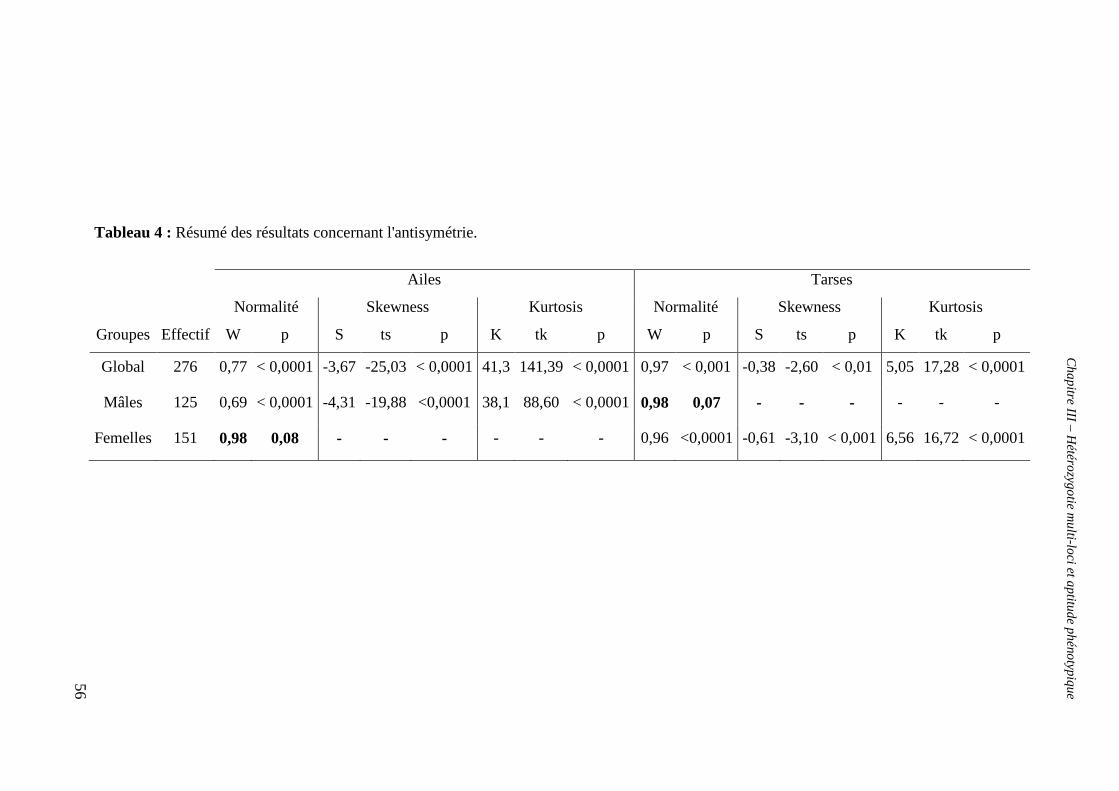
Ch
ap
itre III –
Hé
téro
zygo
tie m
ulti-lo
ci et a
ptitu
de
ph
éno
typiq
ue
56
Tableau 4 : Résumé des résultats concernant l'antisymétrie.
Ailes Tarses
Groupes Effectif
Normalité Skewness Kurtosis Normalité Skewness Kurtosis
W p S ts p K tk p W p S ts p K tk p
Global 276 0,77 < 0,0001 -3,67 -25,03 < 0,0001 41,3 141,39 < 0,0001 0,97 < 0,001 -0,38 -2,60 < 0,01 5,05 17,28 < 0,0001
Mâles 125 0,69 < 0,0001 -4,31 -19,88 <0,0001 38,1 88,60 < 0,0001 0,98 0,07 - - - - - -
Femelles 151 0,98 0,08 - - - - - - 0,96 <0,0001 -0,61 -3,10 < 0,001 6,56 16,72 < 0,0001
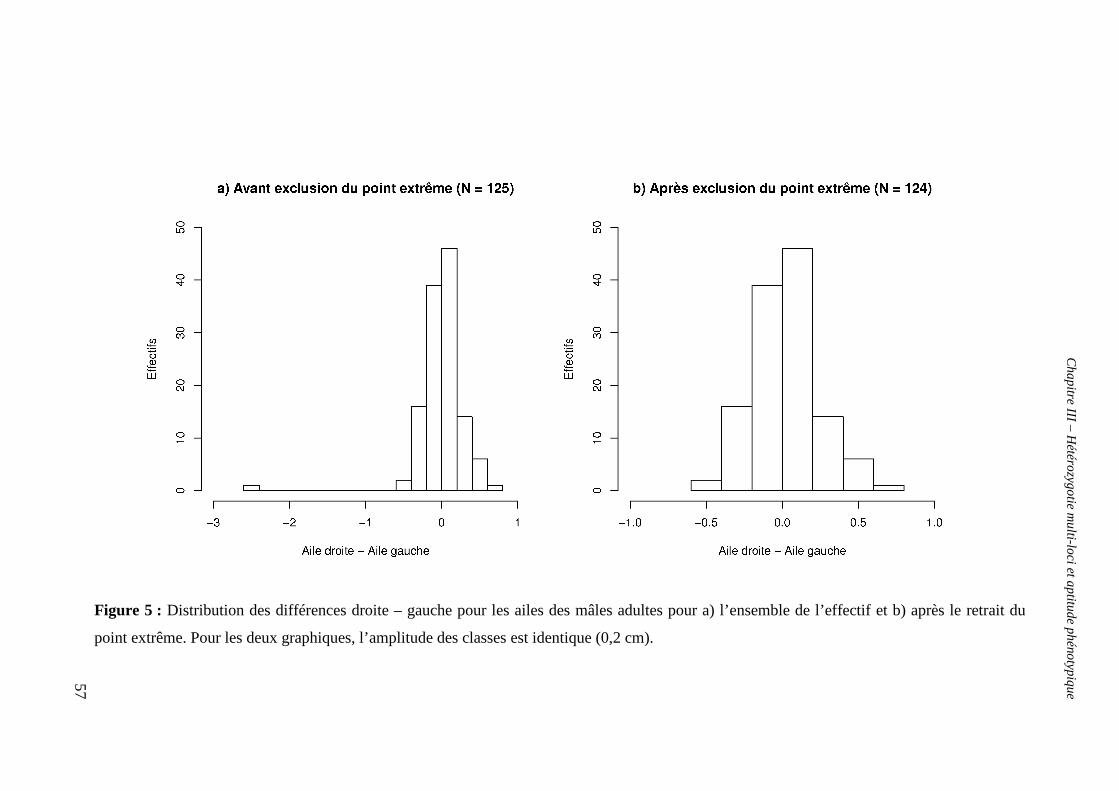
Ch
ap
itre III –
Hé
téro
zygo
tie m
ulti-lo
ci et a
ptitu
de
ph
éno
typiq
ue
57
Figure 5 : Distribution des différences droite – gauche pour les ailes des mâles adultes pour a) l’ensemble de l’effectif et b) après le retrait du
point extrême. Pour les deux graphiques, l’amplitude des classes est identique (0,2 cm).
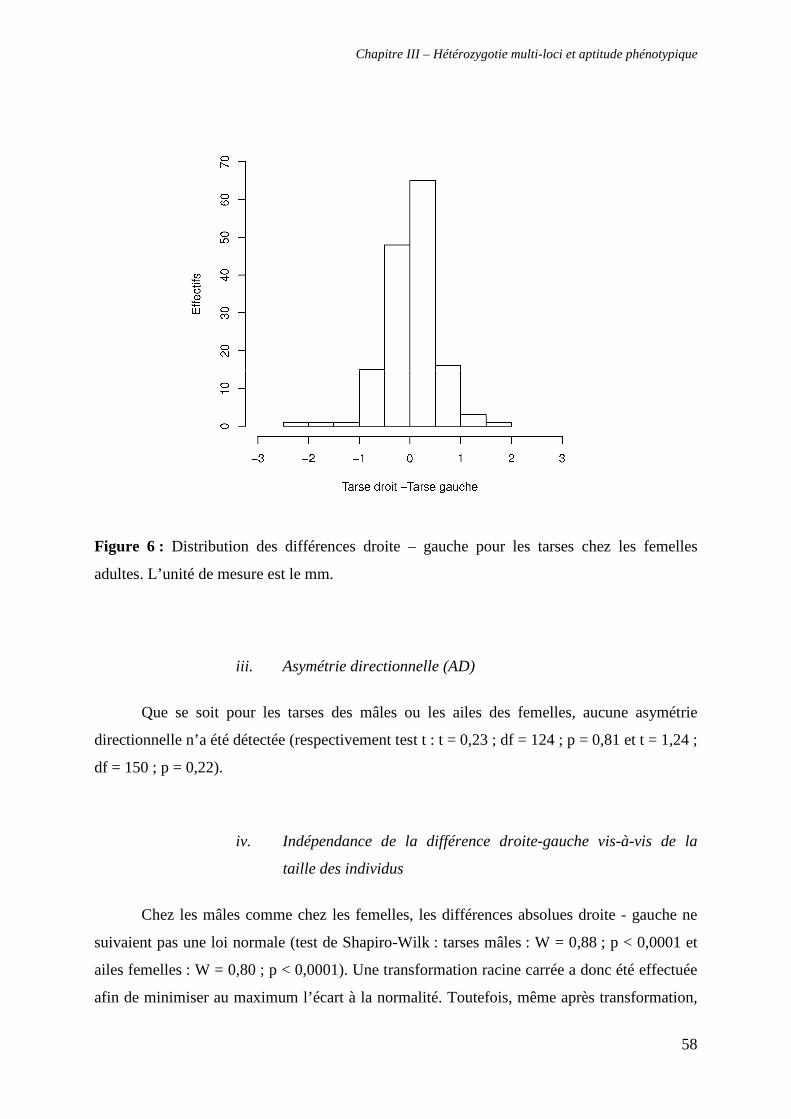
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
58
Figure 6 : Distribution des différences droite – gauche pour les tarses chez les femelles
adultes. L’unité de mesure est le mm.
iii. Asymétrie directionnelle (AD)
Que se soit pour les tarses des mâles ou les ailes des femelles, aucune asymétrie
directionnelle n’a été détectée (respectivement test t : t = 0,23 ; df = 124 ; p = 0,81 et t = 1,24 ;
df = 150 ; p = 0,22).
iv. Indépendance de la différence droite-gauche vis-à-vis de la
taille des individus
Chez les mâles comme chez les femelles, les différences absolues droite - gauche ne
suivaient pas une loi normale (test de Shapiro-Wilk : tarses mâles : W = 0,88 ; p < 0,0001 et
ailes femelles : W = 0,80 ; p < 0,0001). Une transformation racine carrée a donc été effectuée
afin de minimiser au maximum l’écart à la normalité. Toutefois, même après transformation,

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
59
la normalité n’était toujours pas respectée (tarses mâles : W = 0,98 ; p = 0,03 et ailes
femelles : W = 0,97 ; p < 0,0001). Les deux autres variables représentant la moyenne des
caractères considérés (moyenne des tarses pour les mâles et moyenne des ailes pour les
femelles) étant distribuées normalement, les régressions entre les différences absolues (|D –
G|) contre moyenne du caractère ( [D + G] / 2) ont tout de même été réalisées avec néanmoins
un contrôle de la distribution des résidus. Pour les tarses des mâles, les résidus étaient
distribués normalement (W = 0,99 ; p = 0,32) et la régression était significative (R² = 0,053 ;
123 df ; p < 0,01). Dans ce cas, les deux indices d’AF (FA1 et FA4) ont dû être standardisés
par la moyenne du caractère pour les analyses suivantes. En ce qui concerne les ailes des
femelles, la taille corporelle ne semblait pas influencer l’asymétrie (R² = -0,006 ; 149 df ; p =
0,76). A noter cependant, que les résidus de cette régression n’étaient pas distribués
normalement.
v. Test de la relation entre hétérozygotie et asymétrie fluctuante
α. Chez les mâles
Pour cette analyse, comme nous l’avons vu précédemment, les indices d’AF, FA1 et
FA4, ont dû être standardisés par la moyenne du caractère considéré, en l’occurrence la taille
moyenne des tarses. Parmi les indices que ce soit pour l’AF (FA1 et FA4 standardisés) ou
l’hétérozygotie (SH, MD, IR et HL), seules les distributions de FA4 et IR suivaient une loi
normale (tableau 5). De ce fait, une transformation racine carrée a été appliquée aux autres
indices afin de tenter de les normaliser. Ceci a permis de restaurer la normalité de MD et de
réduire l’écart pour FA1 (tableau 5). En ce qui concernait SH et HL, cette transformation
n’améliorant pas leur distribution, la variable d’origine a été conservée (tableau 5).
Aucune des huit régressions envisageables avec chacun des quatre indices
d’hétérozygotie et des deux indices d’AF était significative (tableau6, figure 7). Dans tous les
cas, les résidus des régressions suivaient bien une loi normale (tableau 6).
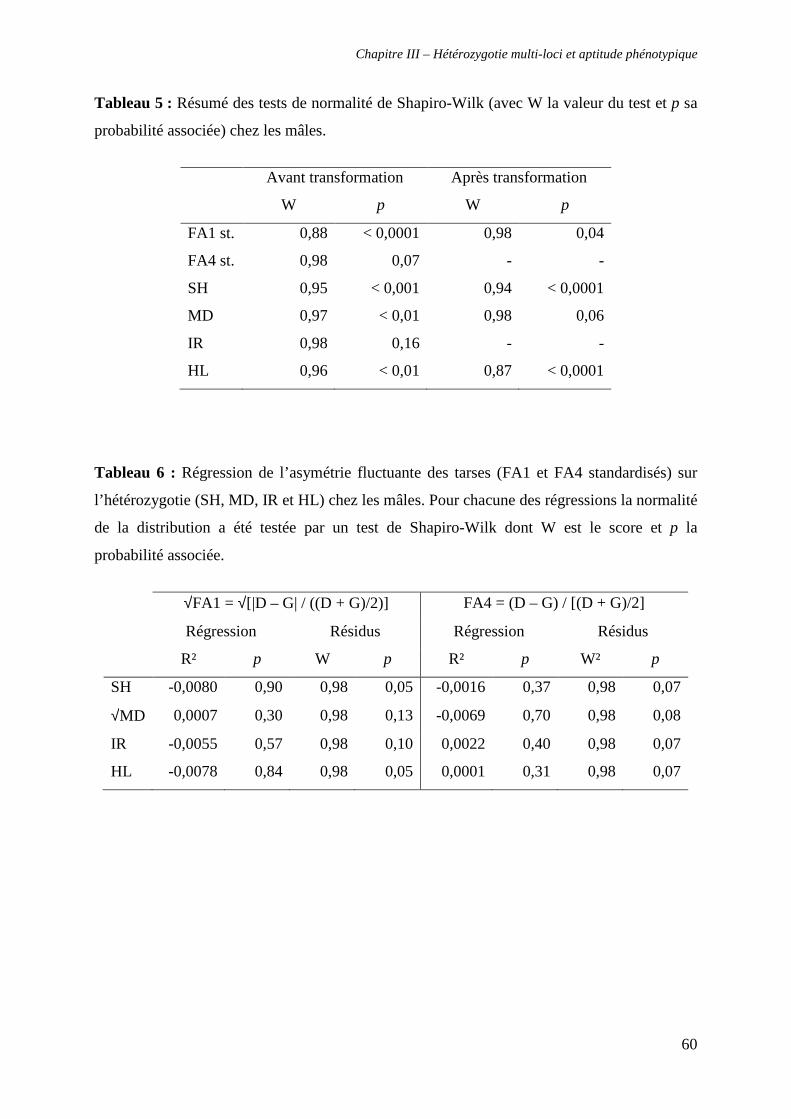
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
60
Tableau 5 : Résumé des tests de normalité de Shapiro-Wilk (avec W la valeur du test et p sa
probabilité associée) chez les mâles.
Avant transformation Après transformation
W p W p
FA1 st. 0,88 < 0,0001 0,98 0,04
FA4 st. 0,98 0,07 - -
SH 0,95 < 0,001 0,94 < 0,0001
MD 0,97 < 0,01 0,98 0,06
IR 0,98 0,16 - -
HL 0,96 < 0,01 0,87 < 0,0001
Tableau 6 : Régression de l’asymétrie fluctuante des tarses (FA1 et FA4 standardisés) sur
l’hétérozygotie (SH, MD, IR et HL) chez les mâles. Pour chacune des régressions la normalité
de la distribution a été testée par un test de Shapiro-Wilk dont W est le score et p la
probabilité associée.
√FA1 = √[|D – G| / ((D + G)/2)] FA4 = (D – G) / [(D + G)/2]
Régression Résidus Régression Résidus
R² p W p R² p W² p
SH -0,0080 0,90 0,98 0,05 -0,0016 0,37 0,98 0,07
√MD 0,0007 0,30 0,98 0,13 -0,0069 0,70 0,98 0,08
IR -0,0055 0,57 0,98 0,10 0,0022 0,40 0,98 0,07
HL -0,0078 0,84 0,98 0,05 0,0001 0,31 0,98 0,07
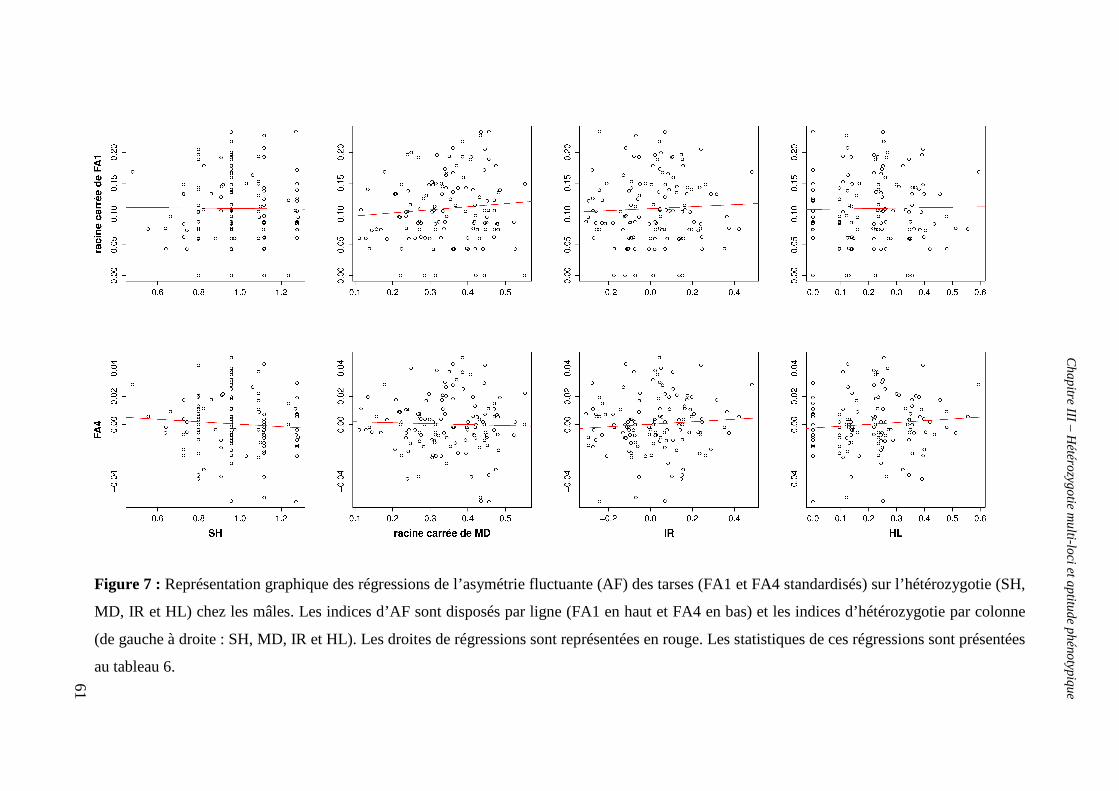
Ch
ap
itre III –
Hé
téro
zygo
tie m
ulti-lo
ci et a
ptitu
de
ph
éno
typiq
ue
61
Figure 7 : Représentation graphique des régressions de l’asymétrie fluctuante (AF) des tarses (FA1 et FA4 standardisés) sur l’hétérozygotie (SH,
MD, IR et HL) chez les mâles. Les indices d’AF sont disposés par ligne (FA1 en haut et FA4 en bas) et les indices d’hétérozygotie par colonne
(de gauche à droite : SH, MD, IR et HL). Les droites de régressions sont représentées en rouge. Les statistiques de ces régressions sont présentées
au tableau 6.

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
62
β. Chez les femelles
Parmi les indices que ce soit pour l’AF (FA1 et FA4) ou l’hétérozygotie (SH, MD, IR
et HL), seule la distribution de FA4 suivait une loi normale (tableau 7). De ce fait, une
transformation racine carrée a été appliquée aux autres indices afin de tenter de les normaliser.
Ceci a permis de restaurer la normalité de MD et de réduire l’écart pour FA1 et IR (tableau 7).
En ce qui concernait SH et HL, cette transformation n’améliorant pas leur distribution, la
variable d’origine a été conservée (tableau 7).
Sur les huit régressions envisageables avec chacun des quatre indices d’hétérozygotie
et des deux indices d’AF, aucune était significative (tableau 8, figure 8). Les résidus issus des
régressions incluant FA1 ne suivaient pas une loi normale contrairement à ceux issus des
régressions impliquant FA4 (tableau 8).
Tableau 7 : Résumé des tests de normalité de Shapiro-Wilk (avec W la valeur du test et p sa
probabilité associée) chez les femelles.
Avant transformation Après transformation
W p W p
FA1. 0,88 < 0,0001 0,95 < 0,0001
FA4 0,98 0,08 - -
SH 0,94 < 0,0001 0,93 < 0,0001
MD 0,97 < 0,01 1 0,60
IR 0,97 < 0,01 0,95 < 0,01
HL 0,95 < 0,0001 0,90 < 0,0001
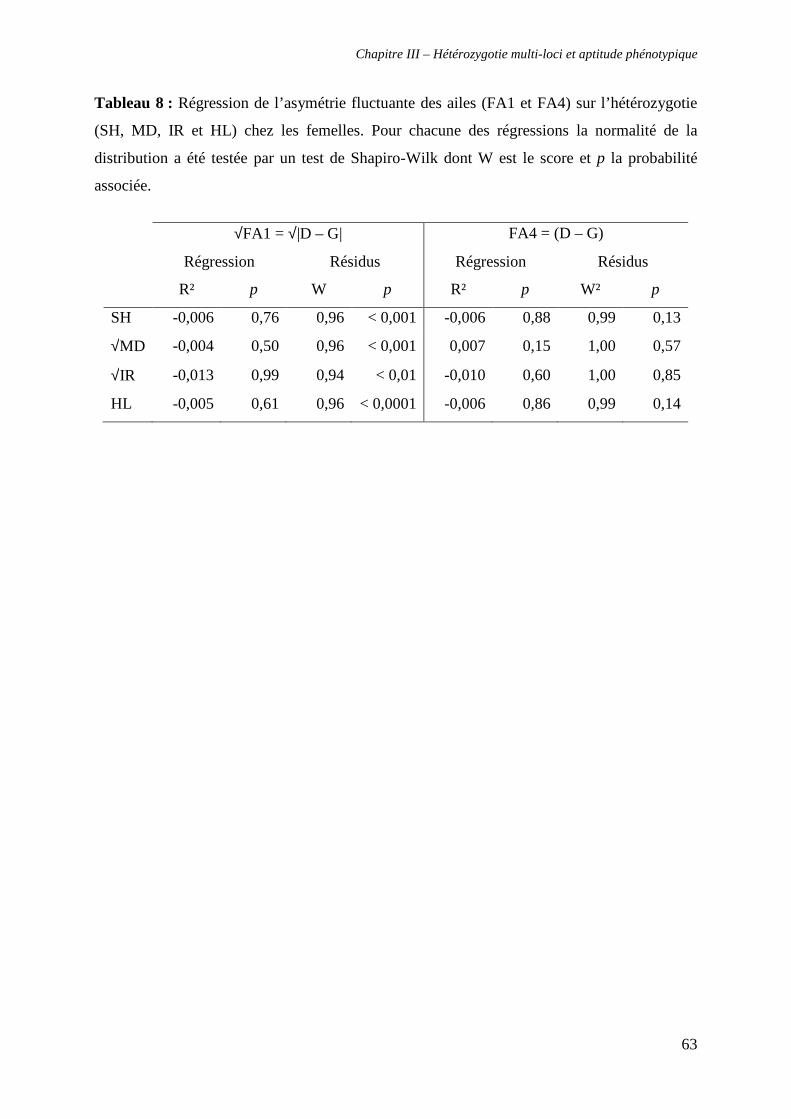
Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
63
Tableau 8 : Régression de l’asymétrie fluctuante des ailes (FA1 et FA4) sur l’hétérozygotie
(SH, MD, IR et HL) chez les femelles. Pour chacune des régressions la normalité de la
distribution a été testée par un test de Shapiro-Wilk dont W est le score et p la probabilité
associée.
√FA1 = √|D – G| FA4 = (D – G)
Régression Résidus Régression Résidus
R² p W p R² p W² p
SH -0,006 0,76 0,96 < 0,001 -0,006 0,88 0,99 0,13
√MD -0,004 0,50 0,96 < 0,001 0,007 0,15 1,00 0,57
√IR -0,013 0,99 0,94 < 0,01 -0,010 0,60 1,00 0,85
HL -0,005 0,61 0,96 < 0,0001 -0,006 0,86 0,99 0,14
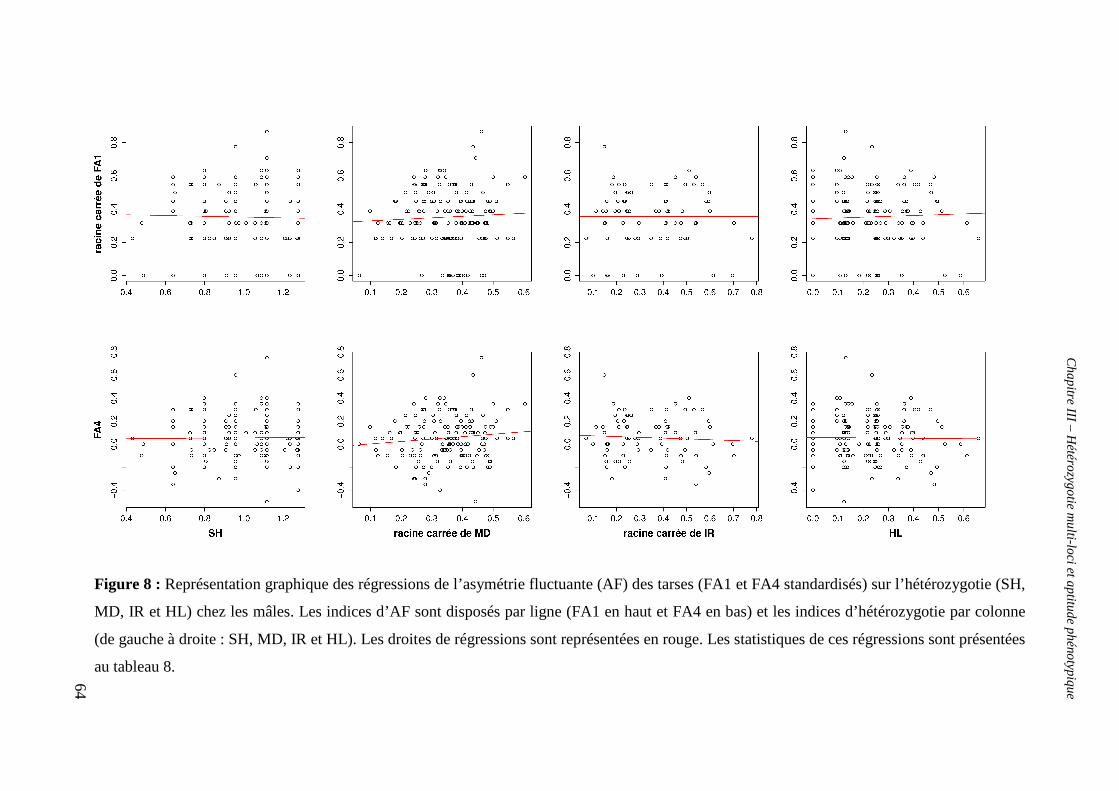
Ch
ap
itre III –
Hé
téro
zygo
tie m
ulti-lo
ci et a
ptitu
de
ph
éno
typiq
ue
64
Figure 8 : Représentation graphique des régressions de l’asymétrie fluctuante (AF) des tarses (FA1 et FA4 standardisés) sur l’hétérozygotie (SH,
MD, IR et HL) chez les mâles. Les indices d’AF sont disposés par ligne (FA1 en haut et FA4 en bas) et les indices d’hétérozygotie par colonne
(de gauche à droite : SH, MD, IR et HL). Les droites de régressions sont représentées en rouge. Les statistiques de ces régressions sont présentées
au tableau 8.

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
65
IV. DISCUSSION
Dans ce chapitre, nous avons analysé la relation potentielle entre l’hétérozygotie de
l’individu et son aptitude phénotypique en utilisant pour les deux facettes plusieurs indices
afin de maximiser l’efficacité de cette détection. Quel que soit l’indice d’hétérozygotie
employé ou le facteur proximal représentant la qualité de l’individu, les relations entre ces
deux paramètres semblaient majoritairement rares et de faible amplitude. En effet, bien que la
littérature soit abondante en la matière, une méta-analyse récente a montré que sur 481 effets
recensés (issus de 58 articles publiés) seulement 24% étaient significatifs (Chapman et al.
2009). De ce fait, l’utilisation de plusieurs indices en parallèle afin de maximiser les chances
de détection est une pratique largement répandue (pour un exemple se référer au tableau 1 de
Coltman & Slate 2003). Toutefois, cela n’a pas permis dans notre cas, une multiplication des
résultats positifs.
Globalement, notre étude soulevait plusieurs interrogations que ce soit d’un point de
vu des résultats mais également des techniques employées et des choix d’analyses.
Hétérozygotie et aptitude phénotypique : un bilan léger
Dans un premier temps, la qualité des individus a été testée au travers de leur capacité
à détenir un territoire. Effectivement, en milieu tropical, la survie des individus est supérieure
à celle observée en zone tempérée, en conséquence de quoi, les territoires libres constituent
une ressource limitée d’où la forte compétition pour leur accession (Greenberg & Gradwohl
1997, Morton et al. 2000, Stutchbury & Morton 2008). Néanmoins, ce trait ne semblait pas
être en relation avec l’hétérozygotie. Par contre, la survie des juvéniles semblerait être
affectée par la diversité génétique. Chez les marmottes des Alpes, Cohas et al. (2009) ont
également mis en évidence une diminution de la variance de l’hétérozygotie avec l’âge mais
ceci était également accompagné d’une augmentation du niveau moyen de diversité génétique
individuelle. La réduction de la variabilité du niveau d’hétérozygotie, observée entre les
juvéniles et les adultes, supposait alors l’existence d’une "purge" des juvéniles de qualité
inférieure sous l’effet de pressions sélectives plus fortes dans les premiers stades de
développement ("filter effect" de Koehn & Gaffney 1984, David 1998). Toutefois, dans notre
cas, une tendance à la diminution de la variabilité de l’hétérozygotie a effectivement été
détectée accompagnée d’une augmentation du niveau médian. Toutefois, dans le cas de la

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
66
variance, il ne s’agissait que de tendances. Si ce point était confirmé, notamment par
l’utilisation d’un plus grand nombre de marqueurs et d’un échantillon de taille supérieure
(surtout pour les juvéniles), celui-ci suggèrerait l’existence d’une mortalité des juvéniles les
plus homozygotes par rapport au niveau médian de la population. Cependant, même dans ce
cas de figure, ce résultat nécessitait tout de même d’être tempéré étant donné qu’il ne
s’agissait ici que d’une analyse corrélative. En effet, la survie des juvéniles per se n’a pas été
testée directement mais elle a été estimée, ou du moins explicitée, au travers du passage du
stade juvénile au stade adulte. De ce fait, l’analyse devrait non seulement être réitérée sur un
échantillon plus important en augmentant le nombre de loci testés mais elle mériterait d’être
également complétée par un suivi populationnel afin de définir les probabilités de survie de
chaque groupe.
La seconde partie de notre analyse visait à évaluer l’existence d’une relation entre la
stabilité du développement des individus, au travers de l’asymétrie fluctuante (AF) et leur
niveau d’hétérozygotie. Toutefois, aucune relation n’a pu être mise en évidence. Cependant,
la détection d’une forme de distributions leptokurtique (cf. figure 2) pour les ailes des mâles
et les tarses des femelles soulève quelques interrogations. D’un point de vue théorique, cette
forme particulière peut être causée par i) la présence de points extrêmes, ii) d’une erreur de
mesure variable, iii) d’un mélange entre AF et AS, iv) de l’hétérogénéité de la dépendance de
la taille du caractère considéré ou v) de l’hétérogénéité due à une véritable instabilité de
développement (Palmer & Strobeck, 2003). Pour ce qui est du premier point, cette vérification
a déjà été réalisée. Dans le cas des femelles, le biais pourrait être la conséquence de l’erreur
de mesure plus importante pour les tarses droits. Toutefois, même à 22% (la moyenne sur
l’étude étant à 8,59%), cette erreur de mesure restait inférieure à ce qui a pu être recensé dans
la littérature comme par exemple, dans l’étude de Merilä & Björklund (1995) où l’erreur de
mesure était en moyenne de 44% (Kruuk et al. 2003). D’autre part, même s’il s’avérait que ce
phénomène puisse trouver son origine dans un problème imputable à la prise de mesure
comme une erreur de mesure différente entre expérimentateurs par exemple, cela devrait
toucher de façon homogène les mâles et les femelles et ce pour les deux caractères
indistinctement. Ceci n’étant pas le cas, la conclusion la plus parcimonieuse serait que ces
particularités pour les tarses des femelles et les ailes des mâles soient imputables aux
hypothèses iii) à v) et seraient donc chacune à tester. La première étape pour ces vérifications
serait d’identifier l’origine du mélange et/ou de l’hétérogénéité observée.
Globalement, l’effet de l’hétérozygotie aux marqueurs que nous avons testés ne
semblait pas avoir d’influence ni sur l’AF ni sur les stratégies d’approvisionnement qui chez

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
67
les tourterelles à queue carrée apparaissaient comme étant une bonne valeur approchée de la
qualité individuelle. Par contre, la survie des juvéniles apparaitrait comme en partie
dépendante de leur génotype.
D’un point de vue général, certains des estimateurs l’AF ou les indices d’hétérozygotie
peuvent être et sont souvent discutés dans la littérature ce qui pourrait potentiellement
nuancer certains des résultats négatifs obtenus ici.
Aptitude phénotypique et hétérozygotie : le choix des bons indices
Dans un premier temps, l’AF, d’un point de vue théorique, offre l’avantage d’être un
estimateur dont la valeur "idéale" est connue (Palmer 1996) et l’acquisition des données ne
nécessite pas de long suivi en capture - marquage – recapture (Lens et al. 2002b). Toutefois,
la méthode d’analyse nécessite une attention particulière afin d’éviter toute possibilité de
mauvaise interprétation due à la présence d’autres formes d’asymétrie notamment en ce qui
concerne la taille de l’échantillon, pouvant limiter le pouvoir statistique (Van Dongen 1999)
mais également pour l’erreur de mesure (Palmer & Strobeck 1986, Palmer 1994, van Dongen
1999, Lens et al. 2002b, Kruuk et al. 2003). En ce qui concerne notre analyse, le traitement
statistique a été réalisé en intégrant ces problèmes potentiels. Cependant, un élément clé dans
cette réalisation était manquant à savoir l’établissement de la relation entre l’AF et l’aptitude
phénotypique. En effet, l’absence d’un patron général (commun à tous les organismes)
concernant la relation entre AF et aptitude phénotypique ou plus précisément le fait que
certaines études soient parvenues à démontrer l’existence d’une relation entre ces deux
paramètres ne permet pas de conclure qu’en règle générale l’AF est représentative de la
qualité de l’individu (van Dongen et al. 1999, Lens et al. 2002b, van Dongen 2006). De même
que l’absence de relation ne signifie pas que celle-ci n’existe pas, par exemple en étant trop
faible pour être détectée (Lens et al. 2002a, Slate & Pemberton 2002, Kruuk et al. 2003). De
ce fait, avant de conclure sur l’existence ou non d’une relation entre AF et hétérozygotie, la
question serait de savoir si l’AF représente ou non une bonne valeur approchée de l’aptitude
phénotypique chez Z. aurita. Pour tester cela, une évaluation de la relation entre l’AF et un
paramètre tel que la survie, ou toute autre mesure directe de l’aptitude phénotypique, serait
nécessaire.

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
68
Dans le cas de l’hétérozygotie, plusieurs indices ont été publiés. Avec notre jeu de
données, nous avons pu constater que SH, IR et HL sont tous les trois très corrélés comme
cela a déjà été montré dans le cadre d’une méta-analyse récente (Chapman et al. 2009).
Globalement, ces trois indices se comportaient de manière équivalente, même si HL semblait
être plus sensible que SH et IR comme le définissait Aparicio et al. (2006). Finalement, parmi
les quatre indices testés, seul MD (mean d² standardisé) se distinguait par une variabilité plus
importante. Cependant, bien que corrélé aux trois autres, tous les résultats issus d’une analyse
l’incluant étaient non significatifs. Ceci soulève la question de la validité de cet indice dans ce
type d’analyse, question abondamment discutée dans la littérature. En effet, l’utilité et
l’efficacité de mean d² dans le cadre d’une mise en évidence de croisements consanguins a
souvent été remise en question (Hedrick et al. 2001, Coltman & Slate 2003). En fait, cet
indice serait plus approprié pour estimer la divergence entre deux lignées sur un temps
important et serait plus adapté à l’évaluation de l’hétérozygotie dans le cas où les complexes
de gènes coadaptés seraient rompus par métissage avec des individus provenant d’autres
populations ("admixture model", Coulson et al. 1998, Neff 2004b). En fin de compte, les
conditions pour lesquelles mean d² serait adéquat semblent être relativement restrictives (un
modèle mixte de populations, un taux de mutation faible, des tailles de sous-populations
importantes et un temps de divergence élevés, Tsitrone et al. 2001). Selon Goudet & Keller
(2002), le produit du taux de mutation du marqueur considéré µ et de la taille des sous-
populations considérées N doit être supérieur à 1 pour que mean d² soit plus corrélé à
l’aptitude phénotypique que ne l’est la proportion H de loci d’hétérozygotes en elle-même.
Toutefois, comme l’ont fait remarquer les auteurs, ceci nécessite que les sous-populations
comptent plus de 10 000 individus, le taux de mutation observé dans le cas de marqueurs
microsatellites en milieu naturel étant compris entre 10-4 et 10-3 mutations par génération
(Weber & Wong 1993). De plus, dans ce cas de figure, l’effet de la consanguinité serait
rapidement (cinq générations) dilué par le mélange de ces populations (Goudet & Keller
2002). Globalement, tant que le processus de mutation des microsatellites n’est pas précisé, le
débat concernant l’utilité et l’acuité de cet indice restera sujet à controverse (Slate et al. 2000,
Slate & Pemberton 2002, Chapman et al. 2009).
D’un point de vue plus général, un élément particulier peut laisser perplexe. Chapman
et al. (2009) ont détecté, dans une revue récente de la littérature, un biais de publication en
faveur des relations hétérozygotie – aptitude phénotypique significatives au détriment de
travaux présentant des effets moindres ou pas de relation du tout. Le problème majeur dans ce

Chapitre III – Hétérozygotie multi-loci et aptitude phénotypique
69
cadre de travail est que la base même du support génétique dont la plupart des études se
servent à l’heure actuelle est un support neutre de la sélection et surtout représentant une
infime partie d’un génome pouvant être excessivement complexe. Dans quelle mesure et avec
quelle certitude est-il possible de conclure à un effet de l’hétérozygotie sur l’aptitude
phénotypique globale d’un organisme ? Dans la recherche de relation entre hétérozygotie et
aptitude phénotypique, une hypothèse de base est implicitement formulée à savoir que la
diversité génétique mesurée à un nombre variable de loci est une bonne estimation du niveau
de consanguinité de l’individu (ou d’hétérozygotie globale). Toutefois, ce présupposé
semblerait être loin d’être soutenu par l’ensemble de la communauté scientifique. En effet,
l’hétérozygotie mesurée à l’aide de plus d’une dizaine de marqueurs de types microsatellites
serait trop faiblement corrélée avec le degré de consanguinité pour être également corrélée
avec l’aptitude phénotypique (Balloux et al. 2004), ce qui pourrait apporter un élément de
réponse au fait que les corrélations observées sont généralement faibles (Coltman & Slate
2003). De même, une étude récente a montré qu’en fonction des microsatellites utilisés pour
l’analyse, les résultats étaient ou non significatifs (Masters et al. 2009). Ce dernier exemple
montre bien la fragilité de la relation entre hétérozygotie et aptitude phénotypique mais
renforce également le fait que la diversité génétique mesurée à quelques loci n’est en rien
représentative de l’ensemble du génotype (excepté dans certaines conditions, voir Ruiz-López
et al. 2009).
Finalement, quelle que soit la technique employée, ses conditions d’application ainsi
que ses limites doivent être prises en compte. Certaines d’entre elles passent comme un effet
de mode, parfois au détriment de leur validité scientifique. Dans le cas de l’asymétrie
fluctuante, l’enthousiasme pour ce nouveau paradigme a peut-être été tellement grand que
finalement certaines études ont été publiées uniquement parce que par chance, dans la masse
d’analyses effectuées, un résultat a été trouvé (Palmer 1996, Simmons et al. 1999). Par un
effet boule de neige, et surtout par le fait que les résultats négatifs soient rarement publiés
(Kotze et al. 2004), la littérature est devenue plus qu’abondante jusqu’à noyer les
informations les plus pertinentes. Comme nous l’avons déjà évoqué au premier chapitre, le
seul remède connu pour pallier ce genre de problème est de bien considérer la validité de sa
méthode ainsi que celle des analyses statistiques (Simmons et al. 1999).

70
DISCUSSION GENERALE
ET
PERSPECTIVES D’ETUDES

71
DISCUSSION GENERALE
ET
PERSPECTIVES D’ETUDES
Cette étude représentait le point de départ des investigations sur la tourterelle à queue
carrée qui, comme cela a déjà été mentionné précédemment, n’avait jamais fait l’objet
d’études autres que morphologiques, comportementales et démographiques (Nellis & Dewey
1978, Nellis et al. 1984, Burger et al. 1989, Wiley 1991, Rivera-Milán 1992, 1996, 1997,
1999, 2001, Carlier & Lefebvre 1996, Dolman et al. 1996, Lefebvre et al. 1996, 1997, Rivera-
Milán & Vásquez 2000, Goldberg et al. 2001, Seferta et al. 2001, Rivera-Milán & Schaffner
2002, Griffin et al. 2005, Sol et al. 2005). De ce fait, un panel d’outils méthodologiques tant
au niveau morphométriques que génétiques a donc dû être mis en place (chapitre I) afin de
pouvoir débuter d’autres analyses spécifiques (chapitre II & III). Néanmoins, au cours du
cheminement de ces trois années de travail, ces aspects méthodologiques ont finalement
revêtu une part plus importante que celle que nous pensions leur accorder dans ce manuscrit.
La méthodologie à la base de la recherche
En parcourant de nouveau le préambule de cette thèse, la partie méthodologique devait
seulement occuper le premier chapitre. Toutefois, a posteriori, ces aspects s’étendent au-delà
de ce chapitre, non pas que cela ait été là notre principale préoccupation mais juste que
finalement, aux détours de certaines questions, nous nous sommes rendus compte de
l’importance de ces tenants et aboutissants. Finalement, le message présenté ici n’est en rien
un jugement de valeur vis-à-vis des études présentant des incohérences méthodologiques mais
plutôt un signal d’alarme car bon nombre d’avertissements passent souvent inaperçus soit
dans le domaine considéré, soit dans des revues de disciplines que nous n’avons pas
l’habitude de consulter (en sciences économiques notamment pour les aspects statistiques).

Discussion générale et perspectives d’études
72
Dans le cas de l’analyse discriminante par exemple, cette technique ainsi que les problèmes
que nous avons abordés la concernant (annexe 1), relatifs entre autre aux méthodes de
validations ainsi qu’à la réduction des variables, sont connus, et ce depuis fort longtemps, par
les mathématiciens "purs" ainsi que par les économistes (Eisenbeis 1977). En ce qui concerne
la littérature en ornithologie, nous avons pu constater que certains auteurs maîtrisaient bien le
sujet et tenaient compte de ce qui était réalisé dans d’autres disciplines. Le problème est qu’au
fil du temps, ces "bonnes pratiques" ont plus ou moins disparu. Diverses hypothèses sont
possibles, notamment le fait que l’utilisation de logiciels statistiques permette d’exécuter
diverses procédures complexes avec une grande simplicité. Toutefois, si l’utilisateur ne
dispose pas d’un minimum de connaissances statistiques, cela peut aboutir à des conclusions
erronées ou à la mauvaise utilisation d’une procédure. Ceci est notamment le cas pour
l’analyse discriminante. En fonction du logiciel employé, les techniques de validation
diffèrent. Par ignorance, certains auteurs auraient finalement cité leurs prédécesseurs sans
connaître spécifiquement les détails techniques de la procédure, parfois même sans vérifier la
source initiale, juste en se basant sur la validité d’une étude déjà publiée.
Néanmoins, un autre principe de la recherche scientifique est que rien n’est jamais
figé, ni gravé ad vitam æternam dans le marbre notamment en fonction des techniques
employées. Ceci est finalement bien illustré par l’étude du polymorphisme de ressources chez
Z. aurita sur l’île de la Barbade où une partie de la population se nourrit et défend un territoire
alors que d’autres tourterelles s’agrègent sur des sites de stockage de grains pour se nourrir en
groupe (chapitre II, annexe 2). Cette étude trouve ses fondements dans les travaux de Sol et
al. (2005) dans lesquels les tourterelles étaient sexées sur la base d’un supposé dimorphisme
sexuel au niveau du plumage de ces oiseaux. De plus, aucun réplicat était effectué pour les
deux sites d’études (un site territorial et un site de nourrissage en groupe). Notre intention à
l’égard de cette publication n’était en rien de la pointer du doigt en jugeant le travail effectué
mais au contraire d’améliorer la connaissance de ce sujet et, éventuellement de rectifier le tir.
Par l’ajout de réplicats et l’introduction de techniques moléculaires (sexage moléculaire et
microsatellites), la vision que nous avions des stratégies d’approvisionnement des tourterelles
à queue carrée de la Barbade a évolué. Bien que nous ne sachions toujours pas définir avec
précision cette situation, l’hypothèse du polymorphisme de ressource a néanmoins pu être
écartée. Cette étude est une illustration de l’intérêt d’une métaréplication à savoir améliorer la
connaissance en réexaminant une problématique avec de nouvelles méthodes (Johnson 2002,
2006, Kelly 2006). Toutefois, comme le souligne Palmer (2000), les lignes éditoriales de
revues scientifiques ne semblent pas encourager la publication d’études réplicatives excepté

Discussion générale et perspectives d’études
73
dans le cas où les conclusions peuvent créer une émulation. En d’autres termes, les points
cruciaux de la publication, du moins dans les meilleures revues (Palmer 2000), sont le niveau
de significativité des tests statistiques mais surtout l’originalité des résultats. De ce fait, une
étude réplicative peut être publiée dans le cas où les résultats obtenus infirment les travaux
précédents. Par ce système, vantant l’originalité et la forte significativité des études, bon
nombre de travaux présentant des effets faibles, non significatifs et négatifs sont rejetés. La
difficulté à publier des résultats issus d’études réplicatives ou des travaux tendant à contredire
la "théorie du moment" contribue à créer un biais de publication (Palmer 2000, Møller &
Jennions 2001, Jennions & Møller 2002a, 2002b, Koricheva 2003) que nous avons déjà
évoqué au cours du troisième chapitre pour la relation entre hétérozygotie, aptitude
phénotypique et asymétrie fluctuante (Coltman & Slate 2003, Chapman et al. 2009). Le
problème majeur est que finalement, les études publiées ne reflètent pas les connaissances que
l’ensemble de la communauté scientifique pourrait avoir. Par extension, les revues
bibliographiques ne dévoilent qu’une partie cachée de l’iceberg. Pour ce qui est des méta-
analyses, donnant en fin de compte une vision plus précise que l’analyse de la littérature,
celles-ci devraient systématiquement intégrer une correction, telle que la méthode "trim &
fill" préconisée par Jennions & Møller (2002a) afin de limiter les effets de ce biais (Møller &
Jennions 2001, Jennions & Møller 2002b, Tomkins & Kotiaho 2004, Chapman et al. 2009).
Globalement, outre les aspects purement méthodologiques à soigner lors de l’élaboration du
protocole expérimental puis au moment de l’analyse des données, ce sont les mentalités de
l’ensemble de la communauté scientifique et surtout des éditeurs de revues scientifiques qui
devraient être modifiées vis-à-vis des résultats négatifs ou non significatifs. A partir du
moment où l’ensemble du protocole et du traitement des données est correctement effectué,
un résultat négatif ou non-significatif est un résultat avant tout, méritant d’être publié au
même titre qu’un résultat significatif. Bien que ceci soit foncièrement accepté par tous, peu
d’articles ont été publiés dans les journaux développés à cet effet tels que Journal of Negative
Results - Ecology & Evolutionary Biology, créé en 2004 (Kotze et al. 2004). Même si un réel
enjeu de connaissance existe, ce journal a publié en moyenne, depuis sa création, moins de
deux articles de recherche par an (neuf articles au total sur cinq volumes publiés).
Dans le cadre de nos travaux, une attention particulière a été portée aux méthodes afin
de construire des bases solides pour pouvoir mener une étude chez Zenaida aurita sur le long
terme. Pour cela, divers outils et techniques ont été adaptés afin d’être utilisés en routine
(chapitre I). Parmi ceux-ci, les outils moléculaires permettant le sexage ainsi que les
microsatellites pour le génotypage des tourterelles à queue carrée sont sans doute le point

Discussion générale et perspectives d’études
74
crucial de cette étude. L’apologie de ces méthodes n’est sans nul doute plus à faire car elles
font aujourd’hui partie intégrante de la caisse à outils du biologiste, que ce soit pour
l’écologie ou la biologie évolutive. Ces méthodes moléculaires seules ou associées aux
mesures morphométriques permettent d’élargir l’horizon quant aux différents thèmes
potentiellement abordables sur une espèce monogame insulaire, largement répartie au sein de
l’archipel d’où elle est endémique. Les possibilités de recherche étant multiples, je me
restreindrai ici à seulement deux axes, le premier en génétique des populations et le second en
écologie comportementale, disciplines pour lesquelles l’apport de la génétique a sans doute
été déterminant dans la progression des connaissances.
La tourterelle à queue carrée dans l’arc caribéen
La tourterelle à queue carrée est, comme cela a été présenté dans le chapitre I, une
espèce commune dans les Antilles, avec une présence plus ou moins marquée de la Floride au
sud des Petites Antilles (Allen 1950, Emlen 1977, Evans 1990, Raffaele et al. 2006). Sa large
répartition en fait une espèce idéale pour l’étude de l’impact des différences biotiques et
abiotiques sur la différenciation morphologique et génétique. En effet, chacune des îles
possède une identité propre en termes "d’environnements compétitifs" ce qui, sous l’effet de
la sélection, ne favorise pas le même type d’individus (Grant 2001). Ainsi, sous l’effet d’une
réduction plus ou moins importante du flux de gènes entre les différents isolats constituant
l’archipel, une différenciation génétique et/ou morphologique plus ou moins prononcée
devrait être observée entre les populations de tourterelles à queue carrée issues de ces
différentes îles. Des travaux préliminaires réalisés par deux étudiantes (master 1ère et 2ème
année) comparant les tourterelles de la Barbade et celles de la Guadeloupe ont d’ores et déjà
mis en évidence une différenciation tant aux niveaux morphologique que génétique grâce aux
outils moléculaires dont nous disposons (Harrang 2008, Colin 2009). Des données concernant
Sainte Lucie devraient rapidement pouvoir être également intégrées afin de préciser l’analyse.
Deux autres îles ont également été prospectées pour l’échantillonnage en 2009 : Saint Martin
et la Jamaïque. Toutefois, trop peu d’individus ont pu être capturés pour être utilisés
(respectivement deux et trois tourterelles). Dans ce cadre, une extension de la zone
d’échantillonnage devra être envisagée. Cependant, ceci nécessite non seulement des moyens
financiers mais également des moyens humains car, finalement, la situation à la Barbade (i.e.
le fait que les tourterelles soient relativement anthropophiles), facilite tout de même

Discussion générale et perspectives d’études
75
grandement la question du baguage, ce qui n’est à l’évidence pas le cas dans les îles comme
Saint Martin et la Jamaïque.
Par ailleurs, une recherche annexe visant à rechercher des parasites sanguins chez les
tourterelles à queue carrée a également été effectuée sur les individus de la Barbade et de la
Guadeloupe. Aucun des individus de la Barbade était porteur de parasites tels que
Plasmodium sp. et/ou Haemoproteus sp. (plus de 150 individus testés) contrairement aux
individus de la Guadeloupe chez lesquels environ 50% de prévalence a été observée pour une
des populations testées (données non publiées). A la Barbade, deux lignées d’Haemoproteus
sp. ont été relevées (Fallon et al. 2005, Svensson & Ricklefs 2009) et plusieurs espèces vivant
en sympatrie avec les tourterelles à queue carrée en sont porteuses telles que la colombe à
queue noire, Columbina passerina, le sucrier à ventre jaune, Coereba flaveola, le sporophile
cici, Tiaris bicolor, le sporophile de la Barbade, Loxigilla barbadensis et le quiscale merle,
Quiscalus lugubris (Fallon et al. 2005, Svensson & Ricklefs 2009). Celle-ci est également
présente dans toutes les Petites Antilles dont la Guadeloupe (Fallon et al. 2005). Comme cela
a déjà été montré chez le moineau domestique, Passer domesticus (Loiseau et al. 2008), un
allèle particulier du CMH (Complexe Majeur d’Histocompatibilité) conférant une résistance à
cette lignée d’Haemoproteus sp. pourrait être présent à la Barbade mais absent ou rare à la
Guadeloupe. Ainsi, le développement de marqueurs du CMH pourrait être ici un atout et
ceux-ci seraient également utilisables dans un tout autre contexte qui est celui de la sélection
sexuelle.
Le choix du partenaire : un retour aux origines
L’idée originale de l’étude de la monogamie et notamment du choix du partenaire chez
la tourterelle à queue carrée n’a certes pas été abordée expérimentalement dans l’ensemble du
manuscrit, mais des travaux préliminaires sur les couples, également entrepris par deux
étudiantes (master 1ère et 2ème année), ont été réalisés (Slaski 2008, Doyen 2009).
Globalement, les associations entre mâles et femelles ne semblaient pas se faire selon des
traits simples (voir néanmoins les résultats obtenus pour les ailes et la queue, Doyen 2009).
D’autres caractères tels que les taches iridescentes sur le plumage de part et d’autre du cou
des individus pourraient également être testés mais supposent de pouvoir établir une méthode
de mesure compatible avec les conditions du terrain. Un élément que nous n’avons jamais
dissocié du phénotype au fil de ce manuscrit, le génotype, est également une piste à suivre.

Discussion générale et perspectives d’études
76
D’un point de vue du choix du partenaire, chaque sexe bénéficie à choisir un partenaire ayant
un patrimoine génétique différent afin d’optimiser l’hétérozygotie ou du moins minimiser la
consanguinité de leur descendance (Brown 1997, 1999, Kempenaers 2007). Cependant, dans
ce cas de figure, les marqueurs les plus appropriés devraient être soumis à la sélection
naturelle ce qui n’est, a priori, pas le cas des marqueurs microsatellites (voir néanmoins
Selkoe & Toonen 2006 pour une remarque à ce sujet). Par contre, les marqueurs du CMH,
évoqués précédemment, peuvent jouer un rôle important dans le choix du partenaire compte
tenu de leur implication dans le système immunitaire (pour des exemples récents se référer à
Eizaguirre et al. 2009 et Bos et al. 2009). Toutefois, le développement de ces marqueurs
nécessite du temps et des compétences. Ce travail n’ayant été entrepris que très récemment,
aucun résultat préliminaire ne peut être abordé ici.
To be continued…
Dans l’ensemble, nul ne doute aujourd’hui de l’utilité des outils moléculaires compte
tenu de leur apport incontestable en écologie et en biologie évolutive. Néanmoins, la
génétique seule n’explique pas tout et finalement l’environnement agit comme un sculpteur
façonnant une statue correspondant à son univers. Sortie de son contexte écologique aucune
relation liant le phénotype et le génotype dans un contexte évolutif ne peut être généralisée.
Les techniques qui ont été soit développées ici ou tout simplement adaptées et/ou testées chez
la tourterelle à queue carrée constituent des bases solides pour un projet de grande ampleur tel
que celui qui a été initialement pensé et qui est voué à prendre d’autant plus d’envergure que
d’autres outils, tels que les marqueurs du CMH, seront efficients. Bien qu’aucune prédiction
quant à la pérennité des recherches ne puisse être formulée, cette étude est promise à
foisonner en un questionnement illimité. Maintenant, compte tenu du fait que les outils sont
disponibles (du moins en grande majorité), le travail de terrain que ce soit au niveau des
captures mais également des observations pour le dénombrement des couples serait à
améliorer, notamment par un suivi systématique des individus identifiés comme tel. En effet,
l’intégralité des travaux doit pouvoir s’appuyer sur un nombre important de tourterelles
baguées chaque année pour conforter la robustesse de nos analyses. Ceci constitue donc un
point important à confirmer mais également à améliorer afin de pouvoir obtenir des suivis de
populations et plus précisément de couples sur les sites territoriaux de la Barbade.

77
REFERENCES
BIBLIOGRAPHIQUES

Références bibliographiques
78
- A -
Acevedo-Whitehouse, K., Petetti, L., Duignan, P. & Castinel, A. 2009. Hookworm infection, anaemia and genetic variability of the New Zealand sea lion. Proceedings of the Royal Society of London B, 276, 3523-3529.
Alarcos, S., de la Cruz, C., Solís, E., Valencia, J. & García-Baquero, M. J. 2007. Sex determination of Iberian Azure-winged magpies Cyanopica cyanus cooki by discriminant analysis of external measurements. Ringing & Migration, 23, 211-216.
Allen, R. P. 1950. Record of Zenaida dove on Florida mainland. The Auk, 67, 237.
Allen, P. J., Amos, W., Pomeroy, P. P. & Twiss, S. D. 1995. Microsatellite variation in grey seals (Halichoerus grypus) shows evidence of genetic differentiation between two british breeding colonies. Molecular Ecology, 4, 653-662.
Amos, W., Worthington Wilmer, J., Fullard, K., Burg, T. M., Croxall, J. P., Bloch, D. & Coulson, T. 2001. The influence of parental relatedness on reproductive success. Proceedings of the Royal Society of London B, 268, 2021-2027.
Aparicio, J. M., Ortego, J. & Cordero, P. J. 2006. What should we weight to estimate heterozygosity, alleles or loci? Molecular Ecology, 15, 4659-4665.
Aristote. 350 av. J. C. De Partibus Animalium.
Arizaga, J., Aldalur, A., Herrero, A. & Galicia, D. 2008. Sex differentiation of Yellow-legged gull (Larus michahellis lusitanius): the use of biometrics, bill morphometrics and wing tip coloration. Waterbirds, 31, 211-219.
Arnould, J. P. Y., Dann, P. & Cullen, J. M. 2004. Determining the sex of Little penguins (Eudyptula minor) in northern Bass Strait using morphometric measurements. Emu, 104, 261-265.
Avise, J. C. 2006. The ontogeny of molecular. A review of Molecular Ecology by J. R. Freeland. Molecular Ecology, 15, 2687-2689.
Ayala, F. J. & Fitch, W. M. 1997. Genetics and the origin of species: an introduction. Proceedings of the National Academy of Sciences of the United States of America, 94, 7691-7697.
- B -
Baker, A. J., Piersma, T. & Greenslade, A. D. 1999. Molecular vs. phenotypic sexing in Red knots. The Condor, 101, 887-893.
Balloux, F., Amos, W. & Coulson, T. 2004. Does heterozygosity estimate inbreeding in real populations? Molecular Ecology, 13, 3021-3031.

Références bibliographiques
79
Bancroft, G. T. 1984. Growth and sexual dimorphism of the Boat-tailed grackle. The Condor, 86, 423-432.
Barré, N., de Garine Wichatitsky, M., Lecoq, R. & Maillard, J.-C. 2003. Contribution to the knowledge of the New Caledonian Imperial pigeon Ducula goliath (Gray 1859) with emphasis on sexual dimorphism. Notornis, 50, 155-160.
Bateman, A. J. 1948. Intra-sexual selection in Drosophila. Heredity, 2, 349-368.
Beebee, T. J. C. & Rowe, G. 2008. An Introduction to Molecular Ecology (2nd edition). New York, USA: Oxford University Press.
Beltran, S., Cézilly, F. & Boissier, J. 2008. Genetic dissimilarity between mates, but not male heterozygosity, influences divorce in schistosomes. Plos One, 3, e3328.
Berlin, K. E., Simon, J. C., Pratt, T. K., Baker, P. E. & Kowalsky, J. R. 2001. Age and sex determination of the Maui parrotbill. Journal of Field Ornithology, 72, 12-21.
Bermudez-Humarán, L. G., Chávez-Zamarripa, P., Guzmán-Velasco, A., Leal-Garza, C. H. & Montes de Oca-Luna, R. 2002. Loss of restriction site DdeI, used for avian molecular sexing, in Oreophasis derbianus. Reproduction in Domestic Animals, 37, 321-323.
Bertellotti, M., Tella, J. L., Godoy, J. A., Blanco, G., Forero, M. G., Donázar, J. A. & Ceballos, O. 2002. Determining sex of Magellanic penguins using molecular procedures and discriminant functions. Waterbirds, 25, 479-484.
Bigler, W. J., Hoff, G. L. & Scribner, L. A. 1977. Survival of Mourning doves unaffected by with drawing blood samples. Bird Banding, 48, 168.
Bishop, J. M., O'Ryan, C. & Jarvis, J. U. M. 2007. Social common mole-rats enhance outbreeding via extra-pair mating. Biology Letters, 3, 176-179.
Blakesley, J. A., Franklin, A. B. & Gutiérrez, R. J. 1990. Sexual dimorphism in Northern Spotted owls from northwest California. Journal of Field Ornithology, 61, 320-327.
Bluso, J. D., Ackerman, J. T., Takekawe, J. Y. & Yee, J. L. 2006. Sexing Forster's terns using morphometric measurements. Waterbirds, 29, 512-517.
Bond, A. B., Wilson, K.-J. & Diamond, J. 1991. Sexual dimorphism in the kea Nestor notabilis. Emu, 91, 12-19.
Borges, R. M. 2005. Ernst Mayr and evolutionary biology: polemics and synthesis. Current Science, 89, 947-954.
Bos, D. H., Williams, R. N., Gopurenko, D., Bulut, Z. & Dewoody, J. A. 2009. Condition-dependent mate choice and a reproductive disadvantage for MHC-divergent male Tiger salamanders. Molecular Ecology, 18, 3307-3315.
Bosch, M. 1996. Sexual size dimorphism and determination of sex in Yellow-legged gulls. Journal of Field Ornithology, 67, 534-541.

Références bibliographiques
80
Bourgeois, K., Curé, C., Legrand, J., Gómez-Díaz, Vidal, E., Aubin, T. & Mathevon, N. 2007. Morphological versus acoustic analysis: what is the most efficient method for sexing Yelkouan shearwaters Puffinus yelkouan? Journal of Ornithology, 148, 261-269.
Bretagnolle, V. & Thibault, J.-C. 1995. Method for sexing fledglings in Cory's shearwaters and comments on sex-ratio variation. The Auk, 112, 785-790.
Brown, J. L. 1969. Territorial behavior and population regulation in birds. The Wilson Bulletin, 81, 293-329.
Brown, J. L. 1997. A theory of mate choice based on heterozygosity. Behavioral Ecology, 8, 60-65.
Brown, J. L. 1999. The new heterozygosity theory of mate choice and the MHC. Genetica, 104, 215-221.
Brown, M. B. & Forsythe, A. B. 1974. Robust tests for equality of variances. Journal of the American Statistical Association, 69, 364-367.
Brucy, L., Dehorter, O. & Fiquet, P. 2007. Guide de saisie des données issues du baguage d'oiseaux sauvages. pp. 45: Muséum National d'Histoire Naturelle - Centre de Recherches par le Baguage des Populations d’Oiseaux, Paris, France.
Buffon, G.-L. L., Comte de. 1749-1789. Histoire Naturelle, Générale et Particulière, avec la Description du Cabinet du Roy. Paris, France: Imprimerie Royale.
Burger, J., Gochfeld, M., Saliva, J. E., Gochfeld, D., Gochfeld, D. & Morales, H. 1989. Antipredator behaviour in nesting Zenaida doves (Zenaida aurita): Parental investment or offspring vulnerability. Behaviour, 111, 129-143.
Burke, T., Seidler, R. & Smith, H. 1992. Editorial. Molecular Ecology, 1, 1.
- C -
Campos, F., Hernández, M., Arizaga, J., Miranda, R. & Amezcua, A. 2005. Sex differentiation of Corn buntings Miliaria calandra wintering in northern Spain. Ringing & Migration, 22, 159-162.
Carlier, P. & Lefebvre, L. 1996. Differences in individual learning between group-foraging and territorial Zenaida doves. Behaviour, 133, 1197-1207.
Castoro, P. L. & Guhl, A. M. 1958. Pairing behavior of pigeons related to aggressiveness and territory. The Wilson Bulletin, 70, 57-69.
Castro, M. S., Recco-Pimentel, S. M. & Rocha, G. T. 2003. Sexual dimorphism in Ramphastos toco and Ramphastos dicolorus (Piciformes, Aves). Revista De Biologia Tropical, 51, 241-245.

Références bibliographiques
81
Cézilly, F. 2006. Le Paradoxe de l'Hippocampe. Une Histoire Naturelle de la Monogamie. Paris, France: Buchet / Chastel.
Chapman, J. R., Nakagawa, S., Coltman, D. W., Slate, J. & Sheldon, B. C. 2009. A quantitative review of heterozygosity-fitness correlations in animal populations. Molecular Ecology, 18, 2746-2765.
Chardine, J. W. & Morris, R. D. 1989. Sexual size dimorphism and assortative mating in the Brown noddy. The Condor, 91, 868-874.
Chochi, M., Nizuma, Y. & Takagi, M. 2002. Sexual differences in the external measurements of Black-tailed gulls breeding on Rishiri Island, Japan. Ornithological Science, 1, 163-166.
Clarke, G. M. 1993. The genetic basis of developmental stability. I. Relationships between stability, heterozygosity, and genomic coadaptation. Genetica, 89, 15-23.
Cohas, A., Bonenfant, C., Kempenaers, B. & Allainé, D. 2009. Age-specific effect of heterozygosity on survival in Alpine marmots, Marmota marmota. Molecular Ecology, 18, 1491-1503.
Cohas, A., Yoccoz, N. G. & Allainé, D. 2007. Extra-pair paternity in Alpine marmots, Marmota marmota: genetic quality and genetic diversity effects. Behavioral Ecology and Sociobiology, 61, 1081-1092.
Cohas, A., Yoccoz, N. G., Da Silva, A., Goossens, B. & Allainé, D. 2006. Extra-pair paternity in the monogamous Alpine marmot (Marmota marmota): the roles of social setting and female mate choice. Behavioral Ecology and Sociobiology, 59, 597-605.
Colin, E. 2009. Différenciation morphologique inter-île chez la tourterelle à queue carrée, Zenaida aurita. Mémoire de Master 1ère année, Université de Bourgogne, Dijon, France.
Coltman, D. W., Bowen, W. D. & Wright, J. M. 1998. Birth weight and neonatal survival of Harbour seal pups are positively correlated with genetic variation measured by microsatellites. Proceedings of the Royal Society of London B, 265, 803-809.
Coltman, D. W., Pilkington, J. G., Smith, J. A. & Pemberton, J. M. 1999. Parasite-mediated selection against inbred Soay sheep in a free-living, island population. Evolution, 53, 1259-1267.
Coltman, D. W. & Slate, J. 2003. Microsatellite measures of inbreeding: a meta-analysis. Evolution, 57, 971-983.
Copello, S., Quintana, F. & Somoza, G. 2006. Sex determination and sexual size-dimorphism in Southern Giant-petrels (Macronectes giganteus) from Patagonia, Argentina. Emu, 106, 141-146.

Références bibliographiques
82
Coulson, T. N., Albon, S., Slate, J. & Pemberton, J. 1999. Microsatellite loci reveal sex-dependent responses to inbreeding and outbreeding in Red deer calves. Evolution, 53, 1951-1960.
Coulson, T. N., Pemberton, J. M., Albon, S. D., Beaumont, M., Marshall, T. C., Slate, J., Guinness, F. E. & Clutton-Brock, T. H. 1998. Microsatellites reveal heterosis in Red deer. Proceedings of the Royal Society of London B, 265, 489-495.
Cronk, Q. C. B. 1997. Islands: stability, diversity, conservation. Biodiversity and Conservation, 6, 477-493.
Crow, J. F. 1998. Perspectives on genetics: anecdotal, historical, and critical commentaries - Ninety years ago: the beginning of hybrid maize. Genetics, 148, 923-928.
Cucco, M., Lingua, G., Bocchio, D., Acquarone, C. & Malacarne, G. 1999. Sex identification in the moorhen (Gallinula chloropus) by flow cytometry and morphometric analysis. Italian Journal of Zoology, 66, 1-6.
- D -
da Silva, A., Gaillard, J.-M., Yoccoz, N. G., Hewison, A. J. M., Galan, M., Coulson, T., Allainé, D., Vial, L., Delorme, D., van Laere, G., Klein, F. & Luikart, G. 2008. Heterozygosity-fitness correlations revealed by neutral and candidate gene markers in Roe deer from a long-term study. Evolution, 63, 403-417.
Darwin, C. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life (First Edition). London, UK: Murray, J.
Dávalos, L. M. 2004. Phylogeny and biogeography of Caribbean mammals. Biological Journal of the Linnean Society, 81, 373-394.
David, P. 1998. Heterozygosity-fitness correlations: new perspectives on old problems. Heredity, 80, 531-537.
Debat, V. & David, P. 2001. Mapping phenotypes: canalization, plasticity and developmental stability. Trends in Ecology & Evolution, 16, 555-561.
Defoe, D. 1719. The Life and strange Surprizing Adventures of Robinson Crusoe of York, Mariner: Who lived Eight and Twenty Years, all Alone in an Un-inhabited Island on the Coast of America, Near the Mouth of the Great River of Oroonoque; Having Been Cast on Shore by Shipwreck, Where-in all the Men Perished but Himself. With An Account How he Was at Last as Strangely Deliver'd by Pyrates. Written by Himself. London, UK.
del Hoyo, J., Elliott, A. & Sargatal, J. 1997. Handbook of the Birds of the World. Barcelona, Spain.

Références bibliographiques
83
del mar Delgado, M. & Penteriani, V. 2004. Gender determination of Eurasian Eagle-owls (Bubo bubo) by morphology. Journal of Raptor Research, 38, 375-377.
Dengo, G. & Case, J. E. 1990. The Geology of North America. Volume H: The Caribbean Region. Boulder, USA: The Geological Society of America.
Desrochers, A. 1990. Sex determination of Black-capped chickadees with discriminant analysis. Journal of Field Ornithology, 61, 79-84.
Devlin, C. M., Diamond, A. W. & Saunders, G. W. 2004. Sexing Arctic terns in the field and laboratory. Waterbirds, 27, 314-320.
Dobzhansky, T. 1937. Genetics and the Origin of Species New York, USA: Columbia University Press.
Dobzhansky, T. 1973. Nothing makes sense except in the light of evolution. The American Biology Teacher, 35, 125-129.
Dolman, C. S., Templeton, J. & Lefebvre, L. 1996. Mode of foraging competition is related to tutor preference in Zenaida aurita. Journal of Comparative Psychology 110, 45-54.
Donohue, K. C. & Dufty, A. M., Jr. 2006. Sex determination of Red-tailed hawks (Buteo jamaicensis calurus) using DNA analysis and morphometrics. Journal of Field Ornithology, 77, 74-79.
Donovan, S. K. & Jackson, T. A. 1994. Caribbean Geology: An Introduction. Kingston, Jamaica: University of the West Indies Publishers' Association.
Dorr, B., King, D. T., Harrel, J. B., Gerard, P. & Spalding, M. G. 2005. The use of culmen length to determine sex of the American White pelican. Waterbirds, 28, 102-106.
Dowling, D. K. & Mulder, R. A. 2006. Combined influence of maternal and paternal quality on sex allocation in Red-capped robins. Journal of Evolutionary Biology, 19, 440-449.
Doyen, G. 2009. Dimorphisme sexuel et homogamie chez une espèce monogame, la tourterelle à queue carrée, Zenaida aurita. Mémoire de Master 1ère année, Université de Bourgogne, Dijon, France.
Dubiec, A. & Zagalska-Neubauer, M. 2006. Molecular techniques for sex identification in birds. Biological Letters, 43, 3-12.
- E -
Eason, D., Millar, C. D., Cree, A., Halverson, J. L. & Lambert, D. M. 2001. A comparison of five methods for assignment of sex in the takahe (Aves: Porphyrio mantelli). Journal of Zoology, 253, 281-292.

Références bibliographiques
84
Eisenbeis, R. A. 1977. Pitfalls in the application of discriminant analysis in business, finance, and economics. The Journal of Finance, 32, 875-900.
Eizaguirre, C., Yeates, S. E., Lenz, T. L., Kalbe, M. & Milinski, M. 2009. MHC-based mate choice combines good genes and maintenance of MHC polymorphism. Molecular Ecology, 18, 3316-3329.
Ellegren, H. 1996. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proceedings of the Royal Society of London B, 263, 1635-1641.
Ellegren, H. 2008. Comparative genomics and the study of evolution by natural selection. Molecular Ecology, 17, 4586-4596.
Ellegren, H. & Fridolfsson, A.-K. 1997. Male-driven evolution of DNA sequences in birds. Nature, 17, 182-184.
Emlen, J. T. 1977. Land bird communities of Grand Bahamas Island: the structure and dynamics of an avifauna. Ornithological Monographs, 24, 1-143.
Eraud, C., Trouvé, C., Dano, S., Chastel, O. & Faivre, B. 2008. Competition for resources modulates cell-mediated immunity and stress hormone level in nestling Collared doves (Streptopelia decaocto). General and Comparative Endocrinology, 155, 542-551.
Evans, P. 1990. Birds of the Eastern Caribbean. Oxford, UK: Macmillan Publishers Limited.
Evans, D. R., Cavanagh, P. M., French, T. W. & Blodget, B. G. 1995. Identifying the sex of Massachusetts Herring gulls by linear measurements. Journal of Field Ornithology, 66, 128-132.
- F -
Fallon, S. M., Bermingham, E. & Ricklefs, R. E. 2005. Host specialization and geographic localization of avian malaria parasites: a regional analysis in the Lesser Antilles. The American Naturalist, 165, 466-480.
Figuerola, J., Garcia, L., Green, A. J., Ibañez, F., Mañez, M., del Valle, J. L., Garrido, H., Arroyo, J. L. & Rodríguez, R. 2006. Sex determination in Glossy ibis chicks based on morphological characters. Ardeola, 53, 229-235.
Fleming, T. L., Halverson, J. L. & Buchanan, J. B. 1996. Use of DNA analysis to identify sex of Northern Spotted owls (Strix occidentalis caurina). Journal of Raptor Research, 30, 118-122.
Fletcher, K. L. & Hamer, K. C. 2003. Sexing terns using biometrics: the advantage of within-pair comparisons. Bird Study, 50, 78-83.

Références bibliographiques
85
Flux, I. & Innes, J. 2001. A field technique for determining the sex of North Island kokapo (Callaeas cinerea wilsoni). Notornis, 48, 217-223.
Foerster, K., Delhey, K., Johnsen, A., Lifjeld, J. T. & Kempenaers, B. 2003. Females increase offspring heterozygosity and fitness through extra-pair matings. Nature 425, 714-717.
Fossøy, F., Johnsen, A. & Lifjeld, J. T. 2008. Multiple genetic benefits of female promiscuity in a socially monogamous passerine. Evolution, 62, 145-156.
Freeman-Gallant, C. R., Wheelwright, N. T., Meiklejohn, K. E. & Sollecito, S. V. 2006. Genetic similarity, extra-pair paternity, and offspring quality in Savannah sparrows (Passerculus sandwichensis). Behavioral Ecology, 17, 952-958.
Frey, S. J. K., Rimmer, C. C., McFarland, K. P. & Menu, S. 2008. Identification and sex determination of Bicknell's thrushes using morphometric data. Journal of Field Ornithology, 79, 408-420.
Fridolfsson, A.-K., Cheng, H., Copeland, N. G., Jenkins, N. A., Liu, H.-C., Raudsepp, T., Woodage, T., Chowdhary, B., Halverson, J. & Ellegren, H. 1998. Evolution of the avian sex chromosomes from an ancestral pair of autosomes. Proceedings of the National Academy of Sciences of the United States of America, 95, 8147-8152.
Fridolfsson, A. .-K. & Ellegren, H. 1999. A simple and universal method for molecular sexing of non-ratite birds. Journal of Avian Biology, 30, 116 - 121.
- G -
Galarza, A., Hidalgo, J., Ocio, G. & Rodriguez, P. 2008. Sexual size dimorphism and determination of sex in Atlantic Yellow-legged gulls Larus michahellis lusitanius from northern Spain. Ardeola, 55, 41-47.
Genovart, M., McMinn, M. & Bowler, D. 2003. A discriminant function for predicting sex in the Balearic shearwater. Waterbirds, 26, 72-76.
Gibbs, D., Barnes, E. & Cox, J. 2001. Pigeons and Doves: A Guide to the Pigeons and Doves of the World. pp. 615. New Haven, Connecticut, USA: Yale University Press.
Gill, S. A. & Vonhof, M. J. 2006. Sexing monochromatic birds in the field: cryptic sexual size dimorphism in Buff-breasted wrens (Thryotorus leucotis). Ornitología Neotropical, 17, 409-418.
Glahn, J. F. & McCoy, R. B. 1995. Measurements of wintering double-crested cormorants and discriminant models of sex. Journal of Field Ornithology, 66, 299-304.
Glor, R. E., Losos, J. B. & Larson, A. 2005. Out of Cuba: overwater dispersal and speciation among lizards in the Anolis carolinensis subgroup. Molecular Ecology, 14, 2419-2432.

Références bibliographiques
86
Goldberg, J. L., Grant, J. W. A. & Lefebvre, L. 2001. Effects of the temporal predictability and spatial clumping of food on the intensity of competitive aggression in the Zenaida dove. Behavioral Ecology, 12, 490-495.
Goldstein, D. B., Linares, A. R., Cavalli-Sforza, L. L. & Feldman, M. W. 1995. An evaluation of genetic distances for use with microsatellite loci. Genetics, 139, 463-471.
Goudet, J. & Keller, L. 2002. The correlation between inbreeding and fitness: does allele size matter? Trends in Ecology & Evolution, 17, 201-202.
Gowaty, P. A. 1996. Battle of the sexes and origins of monogamy. In: Partnerships in Birds (Ed. by Black, J. M.), pp. 21-52. Oxford, UK: Oxford University Press.
Grant, P. R. 2001. Reconstructing the evolution of birds on islands: 100 years of research. Oikos, 92, 385-403.
Grant, P. R. & Grant, B. R. 2008. How and Why Species Multiply. The Radiation of Darwin's Finches. Princeton, USA: Princeton University Press.
Grecian, V. D., Diamond, A. W. & Chardine, J. W. 2003. Sexing razorbills Alca torda breeding at Machias Seal Island, New Brunswick, Canada, using discriminant function analysis. Atlantic Seabirds, 5, 73-80.
Greenberg, R. & Gradwohl, J. 1997. Sexual roles in the dot-winged antwren (Microrhopias quixensis), a tropical forest passerine. The Auk, 100, 920-925.
Griffin, A. S., Savani, R. S., Hausmanis, K. & Lefebvre, L. 2005. Mixed-species aggregations in birds: Zenaida doves, Zenaida aurita, respond to alarm calls of Carib grackles, Quiscalus lugubris. Animal Behaviour, 70, 507-515.
Griffith, S. C., Owens, I. P. F. & Thuman, K. A. 2002. Extra pair paternity in birds: a review of interspecific variation and adaptive function. Molecular Ecology, 11, 2195-2212.
Griffiths, R., Daan, S. & Dijkstra, C. 1996. Sex identification in birds using two CHD genes. Proceedings of the Royal Society of London B, 263, 1251-1256.
Griffiths, R., Double, M. C., Orr, K. & Dawson, R. J. C. 1998. A DNA test to sex most birds. Molecular Ecology, 7, 1071-1075.
Griffiths, R. & Korn, R. M. 1997. A CHD1 gene is Z chromosome linked in the chicken Gallus domesticus. Gene, 197, 225-229.
Griffiths, R. & Tiwari, B. 1993. The isolation of molecular genetic markers for the identification of sex. Proceedings of the National Academy of Sciences of the United States of America, 90, 8324-8326.
Gruys, R. C. & Hannon, S. J. 1993. Sex determination of hunter-killed and depredated Willow ptarmigan using a discriminant-analysis. Journal of Field Ornithology, 64, 11-17.

Références bibliographiques
87
- H -
Haag-Liautard, C., Vitikainen, E., Keller, L. & Sundström, L. 2009. Fitness and the level of homozygosity in a social insect. Journal of Evolutionary Biology, 22, 134-142.
Hallgrimsson, G. T., Palsson, S. & Summers, R. W. 2008. Bill length: a reliable method for sexing Purple sandpipers. Journal of Field Ornithology, 79, 87-92.
Hanners, L. A. & Patton, S. R. 1985. Sexing Laughing gulls using external measurements and discriminant analysis. Journal of Field Ornithology, 56, 158-164.
Hansson, B., Bensch, S., Hasselquist, D. & Åkesson, M. 2001. Microsatellite diversity predicts recruitment of sibling great reed warblers. Proceedings of the Royal Society of London B, 268, 1287-1291.
Hardy, A. R., Hirons, G. J. M., Stanley, P. I. & Huson, L. W. 1981. Sexual dimorphism in size of Tawny owls (Strix aluco): a method for sexing in field studies. Ardea, 69, 181-184.
Harrang, E. 2008. Diversification morpho-anatomique et génétique chez un colombidé endémique des Antilles, la tourterelle à queue carrée (Zenaida aurita). Mémoire de stage de Master 2ème année, Université de Bourgogne, Dijon, France.
Hedges, S. B. 2001. Biogeography of the West Indies: An Overview. In: Biogeography of the West Indies: Patterns and Perspectives (Ed. by Woods, C. A. & Sergile, F. E.), pp. 15-33: CRC Press
Hedges, S. B., Couloux, A. & Vidal, N. 2009. Molecular phylogeny, classification, and biogeography of West Indian Racer snakes of the tribe Alsophiini (Squamata, Dipsadidae, Xenodontinae). Zootaxa, 2067, 1-28.
Hedges, S. B., Hass, C. A. & Maxson, L. R. 1992. Caribbean biogeography: molecular evidence for dispersal in west indian terrestrial vertebrates. Proceedings of the National Academy of Sciences of the United States of America, 89, 1909-1913.
Hedrick, P., Fredrickson, R. & Ellegren, H. 2001. Evaluation of đ², a microsatellite measure of inbreeding and outbreeding, in wolves with a known pedigree. Evolution, 55, 1256-1260.
Heinicke, M. P., Duellman, W. E. & Hedges, S. B. 2007. Major Caribbean and Central American frog faunas originated by ancient oceanic dispersal. Proceedings of the National Academy of Sciences of the United States of America, 104, 10092–10097.
Helander, B., Hailer, F. & Vilà, C. 2007. Morphological and genetic sex identification of White-tailed eagle Haliaeetus albicilla nestlings. Journal of Ornithology, 148, 435-442.

Références bibliographiques
88
Hermosell, I. G., Balbontin, J., Marzal, A., Reviriego, M. & de Lope, F. 2007. Sex determination in Barn swallows Hirundo rustica by means of discriminant analysis in two European populations. Ardeola, 54, 93-100.
Herring, G., Gawlik, D. E. & Beerens, J. M. 2008. Sex determination for the Great egret and White ibis. Waterbirds, 31, 298-303.
Hillis, D. M., Moritz, C. & Mable, B. K. 1996. Molecular Systematics, Second Edition. Massachusets, USA: Sunderland.
Hipkiss, T. 2007. Can migrating Tengmalm's owls Aegolius funereus be reliably sexed in autumn using simple morphometric measurements? Ringing & Migration, 23, 201-204.
Hochholdinger, F. & Hoecker, N. 2007. Towards the molecular basis of heterosis. Trends in Ecology & Evolution, 12, 427-432.
Hocken, A. G. & Russell, J. J. 2002. A method for determination of gender from bill measurements in Otago Blue penguins (Eudyptula minor). New Zealand Journal of Zoology, 29, 63-69.
Hoffman, J. I. 2009. A panel of new microsatellites loci for genetic studies of Antarctic Fur seals and other otariids. Conservation Genetics, 10, 989-992.
Hogan, F. E., Cooke, R., Burridge, C. P. & Norman, J. A. 2007. Optimizing the use of shed feathers for genetic analysis. Molecular Ecology, 8, 561-567.
Höglund, J., Piertney, S. B., Alatalo, R. V., Lindell, J., Lundberg, A. & Rintamäki, P. T. 2002. Inbreeding depression and male fitness in Black grouse. Proceedings of the Royal Society of London B, 269, 711-715.
Hower, L. M. & Hedges, S. B. 2003. Molecular phylogeny and biogeography of West Indian Teiid lizards of the genus Ameiva. Caribbean Journal of Science, 39, 298-306.
Hoysak, D. J. & Weatherhead, P. J. 1991. Sampling blood from birds: A technique and an assessment of its effect. The Condor, 93, 746-752.
Hui, W., Gel, Y. R. & Gastwirth, J. L. 2008. lawstat: An R package for law, public policy and biostatistics. Journal of Statistical Software, 28.
Hull, C. L. 1996. Morphometric indices for sexing adult Royal Eudyptes schlegeli and rockhopper E. chrysocome penguins at Macquarie Island. Marine Ornithology, 24, 23-27.

Références bibliographiques
89
- I -
Iko, N. M., Dinsmore, S. J. & Knopf, F. L. 2004. Evaluating the use of morphometric measurements from museum specimens for sex determination in Mountain plovers (Charadrius montanus). Western North American Naturalist, 64, 492-496.
- J -
Jakubas, D. & Wojczulanis, K. 2007. Predicting the sex of dovekies by discriminant analysis. Waterbirds, 30, 92-96.
Janicke, T., Ritz, M. S., Hahn, S. & Peter, H.-U. 2007. Sex recognition in Brown skuas: do accoustic signals matter? Journal of Ornithology, 148, 565-569.
Jeffrey, J. J., Fancy, S. G., Lindsey, G. D., Banko, P. C., Pratt, T. K. & Jacobi, J. D. 1993. Sex and age identification of palila. Journal of Field Ornithology, 64, 490-499.
Jehl, J. R., Henry, A. E. & Bond, S. I. 1998. Sexing Eared grebes by bill measurements. Colonial Waterbirds, 21, 98-100.
Jennions, M. D. & Møller, A. P. 2002a. Publication bias in ecology and evolution: an empirical assessment using the "trim and fill" method. Biological Reviews, 77, 211-222.
Jennions, M. D. & Møller, A. P. 2002b. Relationship fade with time: a meta-analysis of temporal trends in publication in ecology and evolution. Proceedings of the Royal Society of London B, 269, 43-48.
Jodice, P. G. R., Lanctot, R. B., Gill, V. A., Roby, D. D. & Hatch, S. A. 2000. Sexing adult Black-legged kittiwakes by DNA, behavior, and morphology. Waterbirds, 23, 405-415.
Johnson, D. H. 2002. The importance of replication in wildlife research. Journal of Wildlife Management, 66, 919-932.
Johnson, D. H. 2006. The many faces of replication. Crop Science 46, 2486-2491.
Johnson, K. P. & Clayton, D. H. 2000a. A molecular phylogeny of the dove genus Zenaida: mitochondrial and nuclear DNA sequences. The Condor, 102, 864-870.
Johnson, K. P. & Clayton, D. H. 2000b. Nuclear and mitochondrial genes contain similar phylogenetic signal for pigeons and doves (Aves: Columbiformes). Molecular Phylogenetics and Evolution, 14, 141-151.
Jouventin, P., Charmantier, A., Dubois, M.-P., Jarne, P. & Bried, J. 2007. Extra-pair paternity in the strongly monogamous Wandering albatross Diomedea exulans has no apparent benefits for females. Ibis, 149, 67-78.

Références bibliographiques
90
- K -
Kelly, C. D. 2006. Replicating empirical research in behavioral ecology: How and why it should be done but rarely ever is. The Quarterly Review of Biology, 81, 221-236.
Kempenaers, B. 2007. Mate choice and genetic quality: a review of the heterozygosity theory. Advances in the Study of Behavior, 37, 189-278.
Kenward, B., Rutz, C., Weir, A. A. S., Chapell, J. & Kacelnik, A. 2004. Morphology and sexual dimorphism of the New Caledonian crow Corvus moneduloides, with notes on its behaviour and ecology. Ibis, 146, 652-660.
Kesler, D. C., Lopes, I. F. & Haig, S. M. 2006. Sex determination of Pohnpei Micronesian kingfishers using morphological and molecular genetic techniques. Journal of Field Ornithology, 77, 229-232.
Kleven, O. & Lifjeld, J. T. 2005. No evidence for increased offspring heterozygosity from extrapair mating in the Reed bunting (Emberiza schoeniclus). Behavioral Ecology, 16, 561-565.
Kloskowski, J., Grela, P., Krogulec, J., Gaska, M. & Tchorzewski, M. 2006. Sexing Red-necked grebes Podiceps grisegena by molecular techniques and morphology. Acta Ornithologica, 41, 176-180.
Koehn, R. K. & Gaffney, P. M. 1984. Genetic heterozygosity and growth rate in Mytilus edulis. Marine Biology, 82, 1-7.
Koricheva, J. 2003. Non-significant results in ecology: a burden or a blessing disguise? Oikos, 102, 397-401.
Kotze, D. J., Johnson, C. A., O'Hara, R. B., Vepsäläinen, K. & Fowler, M. S. 2004. Editorial: the Journal of Negative Results in Ecology and Evolutionary Biology. Journal of Negative Results - Ecology & Evolutionary Biology, 1, 1-5.
Kruuk, L. E. B., Slate, J., Pemberton, J. & Clutton-Brock, T. H. 2003. Fluctuating asymmetry in a secondary sexual trait: no associations with individual fitness, environmental stress or inbreeding, and no heritability. Journal of Evolutionary Biology, 16, 101-113.
Krystkowiak, K., Adamski, T., Surma, M. & Kaczmarek, Z. 2009. Relationship between phenotypic and genetic diversity of parental genotypes and the specific combining ability and heterosis effects in wheat (Triticum aestivum L.). Euphytica, 165, 419-434.
- L -
Lack, D. 1968. Ecological Adaptations for Breeding in Birds. London, UK: Methuen & Co.

Références bibliographiques
91
Lambert, D. 1995. The new science of molecular ecology. New Zealand Journal of Ecology, 19, 93-96.
Leamy, L. J. & Klingenberg, C. P. 2005. The genetics and evolution of fluctuating asymmetry. Annual Review of Ecology Evolution and Systematics, 36, 1-21.
Lefebvre, L., Palameta, B. & Hatch, K. K. 1996. Is group-living associated with social learning? A comparative test of gregarious and a territorial columbid. Behaviour, 133, 241-261.
Lefebvre, L., Templeton, J., Brown, K. & Koelle, M. 1997. Carib grackles imitate conspecific and Zenaida dove tutors. Behaviour, 134, 1003-1017.
Lens, L., van Dongen, S., Kark, S. & Matthysen, E. 2002b. Fluctuating asymmetry as an indicator of fitness: can we bridge the gap between studies? Biological Reviews, 77, 27-38.
Lens, L., van Dongen, S. & Matthysen, A. E. 2002a. Fluctuating asymmetry as an early warning system in the critically endangered Taita trushes. Conservation Biology, 16, 479-487.
Lessells, C. M. & Boag, P. T. 1987. Unrepeatable repeatabilities: a common mistake. The Auk, 104, 116-121.
Lieutenant-Gosselin, M. & Bernatchez, L. 2006. Local heterozygosity-fitness correlations with global positive effects on fitness in Threespine stickleback. Evolution, 60, 1658-1668.
Lindstedt, E. R., Oh, K. P. & Badyaev, A. V. 2007. Ecological, social, and genetic contingency of extrapair behavior in a socially monogamous bird. Journal of Avian Biology, 38, 214-223.
Linné, C. von. 1758. Systema naturae per regna tria naturae :secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (10ème edition). Stockholm, Sweden: Impensis Direct. Laurentii Salvii.
Liordos, V. & Goutner, V. 2008. Sex determination of Great Cormorants (Phalacrocorax carbo sinensis) using morphometric measurements. Waterbirds, 31, 203-210.
Lislevand, T., Marthinsen, G. & Lifjeld, J. T. 2009. Sex differences in body size and body condition in breeding Temminck's stints Calidris temminckii. Journal of Ornithology, 150, 299-302.
Lo Valvo, M. 2001. Sexing adult Cory’s shearwater by discriminant analysis of body measurements on Linosa Island (sicilian channel), Italy. Waterbirds, 24, 169-174.
Loiseau, C., Zoorob, R., Garnier, S., Birard, J., Federici, P., Julliard, R. & Sorci, G. 2008. Antagonistic effects of a Mhc class I allele on malaria-infected house sparrows. Ecology Letters, 11, 258-265.

Références bibliographiques
92
López-Pujol, J., Zhang, F.-M. & Ge, S. 2008. No correlation between heterozygosity and vegetative fitness in the narrow endemic and critically endangered Clematis acerifolia (Ranunculaceae). Biochemical Genetics, 46, 433-445.
Lorentsen, S.-H. & Røv, N. 1994. Sex determination of Antarctic petrels Thalassoica antarctica by discriminant analysis of morphometric characters. Polar Biology, 14, 143-145.
Losos, J. B., Glor, R. E., Kolbe, J. J. & Nicholson, K. 2006. Adaptation, speciation and convergence: a hierarchical analysis of adaptative radiation in Caribbean Anolis lizards. Annals of the Missouri Botanical Garden 93, 24-33.
Losos, J. B. & Ricklefs, R. E. 2009. Adaptation and diversification on islands. Nature, 457, 830-836.
Lovette, I. J., Seutin, G., Ricklefs, R. E. & Bermingham, E. 1999. The assembly of an island fauna by natural invasion: sources and temporal patterns in the avian colonization of Barbados. Biological Invasions, 1, 33-41.
Lozano, G. A. 1994. Size, condition, and territory ownership in male tree swallows (Tachycineta bicolor). Canadian Journal of Zoology, 72, 330-333.
Lyons, E. J., Amos, W., Berkley, J. A., Mwangi, I., Shafi, M., Williams, T. N., Newton, C. R., Peshu, N., Marsh, K., Scott, J. A. G. & Hill, A. V. S. 2009. Homozygosity and risk of childhood death due to invasive bacterial disease. BMC Medical Genetics, 10, 55-63.
- M -
Ma, H. Y., Bi, J. Z., Shao, C. W., Chen, Y., Miao, G. D. & Chen, S. L. 2009. Development of 40 microsatellites markers in Spotted halibut (Verasper variegatus) and the cross-species amplification in Barbin flounder (Verasper moseri). Animal Genetics, 40, 575-582.
MacArthur, R. H. & Wilson, E. O. 1967. The Theory of Island Biogeography. Princeton, USA: Princeton University Press.
MacDougall-Shackleton, E. A., Derryberry, E. P., Foufopoulos, J., Dobson, A. P. & Hahn, T. P. 2005. Parasite-mediated heterozygosity advantage in an outbred songbird population. Biology Letters, 1, 105-107.
Magnacca, K. N., Foote, P. & O'Grady, P. M. 2008. A review of the endemic Hawaiian Drosophilidae and their host plants. Zootaxa, 1728, 1-58.
Mainguy, J., Côté, S. D. & Coltman, D. W. 2009. Multilocus heterozygosity, parental relatedness and individual fitness components in a wild Mountain goat, Oreamnos americanus population. Molecular Ecology, 18, 2297-2306.

Références bibliographiques
93
Malacalaza, V. E. & Hall, M. A. 1988. Sexing adult King cormorants (Phalacrocorax albiventer) by discriminant analysis. Colonial Waterbirds, 11, 32-37.
Mallory, M. L. & Forbes, M. R. 2005. Sex discrimination and measurement bias in Northern fulmars Fulmarus glacialis from the Canadian Arctic. Ardea, 93, 25-36.
Mariano-Jelicich, R., Madrid, E. & Faverol, M. 2007. Sexual dimorphism and diet segregation in the Black skimmer Rynchops niger. Ardea, 95, 115-124.
Marr, A. B., Keller, L. F. & Arcese, P. 2002. Heterosis and outbreeding depression in descendants of natural immigrants to an inbred population of Song sparrows (Melospiza melodia). Evolution, 56, 131-142.
Marshall, T. C. & Spalton, J. A. 2000. Simultaneous inbreeding and outbreeding depression in reintroduced Arabian oryx. Animal Conservation, 3, 241-248.
Martinez-Gómez, J. E. & Curry, R. L. 1998. Distinguishing sex of Socorro mockingbirds by body measurements. Ornitología Neotropical, 9, 103-110.
Masters, B. S., Hicks, B. G., Johnson, L. S. & Erb, L. A. 2003. Genotype and extra-pair paternity in the house wren: a rare-male effect? Proceedings of the Royal Society of London B, 270, 1393-1397.
Masters, B. S., Johnson, L. S., Johnson, B. G. P., Neely, C. J. & Williams, K. L. 2009. "Compatible alleles" and extra-pair paternity: conclusions depend on the microsatellite loci used. The Condor, 111, 365-369.
Mayr, E. 1940. Speciation phenomena in birds. The American Naturalist, 74, 249-278.
Mayr, E. 1942. Systematics and the Origin of Species, from the Viewpoint of a Zoologist. Cambridge, USA: Harward University Press.
McCaffery, B. J. & Ruthrauff, D. R. 2004. Spatial variation in shorebird nest success: implications for inference. Wader Study Group Bulletin 103, 67-70.
McCracken, K. G., Paton, D. C. & Afton, A. D. 2000. Sexual size dimorphism of the Musk duck. The Wilson Bulletin, 112, 457-466.
Meissner, W. & Pilacka, L. 2008. Sex identification of adult dunlins Calidris alpina alpina migrating in autumn trough Baltic region. Ornis Fennica, 85, 135-139.
Mendel, G. 1966. Versuche über pflanzen-hybriden. Verh Naturforsch Ver Brünn, 4, 3-47.
Menotti-Raymond, M. & O'Brien, S. J. 1993. Dating the genetic bottleneck of the African cheetah. Proceedings of the National Academy of Sciences of the United States of America, 90, 3172-3176.
Merilä, J. & Björklund, M. 1995. Fluctuating asymmetry and measurement error. Systematic Biology, 44, 97-101.

Références bibliographiques
94
Merilä, J., Sheldon, B. C. & Griffith, S. C. 2003. Heterotic effects on fitness in a wild bird population. Annales Zoologici Fennici, 40, 269-280.
Miles, L. G., Lance, S. L., Isberg, S. R., Moran, C. & Glenm, T. C. 2009. Cross-species amplification of microsatellites in crocodilians: assessment and applications for the futur. Conservation Genetics, 10, 935-954.
Millar, C. D., Lambert, D. M., Anderson, S. & Halverson, J. L. 1996. Molecular sexing of the communally breeding pukeko: an important ecological tool. Molecular Ecology, 5, 289-293.
Miller, W. J. & Wagner, F. H. 1955. Sexing mature columbiformes by cloacal characters. The Auk, 72, 279-285.
Mills, J. A. 1971. Sexing Red-billed gulls from standard measurements. Journal of Marine and Freshwater Research, 5, 326-328.
Mitton, J. B. 1994. Molecular approches to population biology. Annual Review of Ecology and Systematics, 25, 45-69.
Mizuta, T., Yamada, H., Lin, R.-S., Yodogawa, Y. & Okanoya, K. 2003. Sexing White-rumped munias in Taiwan, using morphology, DNA and distance calls. Ornithological Science, 2, 97-102.
Mock, D. W. 1985. An Introduction to the Neglected Mating System. In: Avian Monogamy (Ed. by Gowaty, P. A. & Mock, D. W.), pp. 1-10. Washington, D. C., USA: The American Ornithologists' Union.
Møller, A. P. & Jennions, M. D. 2001. Testing and adjusting for publication bias. Trends in Ecology & Evolution, 16, 580-586.
Monceau, K., Gaillard, M., Harrang, E., Santiago-Alarcon, D., Parker, P., Cézilly, F. & Wattier, R. 2009. Twenty-three polymorphic microsatellite markers for the Caribbean endemic Zenaida dove, Zenaida aurita, and its conservation in related Zenaida species. Conservation Genetics, 10, 1577-1581.
Monge-Nájera, J. 2008. Ecological biogeography: a review with emphasis on conservation and the neutral model. Gayana, 72, 102-112.
Morrison, J. L. & Maltbie, M. 1999. Methods for gender determination of Crested caracaras. Journal of Raptor Research, 33, 128-133.
Morton, E. S., Derrickson, K. C. & Stutchbury, B. J. M. 2000. Territory switching behavior in a sedentary tropical passerine, the Dusky antbird (Cercomacra tyrannina). Behavioral Ecology, 11, 648-653.
Murie, J. O., Davis, L. S. & McLean, I. G. 1991. Identifying the sex of Fiordland Crested penguins by morphometric characters. Notornis, 38, 233-238.

Références bibliographiques
95
- N -
Neff, B. D. 2004a. Stabilizing selection on genomic divergence in a wild fish population. Proceedings of the National Academy of Science of the United States of America, 101, 2381-2385.
Neff, B. D. 2004b. Mean d² and divergence time: transformations and standardizations. Journal of Heredity, 21, 165-171.
Nellis, D. W. & Dewey, R. A. 1978. Correct band size for Zenaida doves. North American Bird Bander, 3, 106.
Nellis, D. W., Dewey, R. A., Hewitt, M. A., Imsand, S., Philibosian, R. & Yntema, J. A. 1984. Population status of Zenaida doves and other columbids in the Virgin Islands. Journal of Wildlife Management 48, 889-894.
Nisbet, I. C. T., Bridge, E. S., Szczys, P. & Heidinger, B. J. 2007. Sexual dimorphism, female-female pairs, and test for assortative mating in Common terns. Waterbirds, 30, 169-179.
Nitta, J. H. & O'Grady, P. M. 2008. Mitochondrial phylogeny of the endemic Hawaiian craneflies (Diptera, Limoniidae, Dicranomyia): Implications for biogeography and species formation. Molecular Phylogenetics and Evolution, 46, 1182-1190.
Nizuma, Y., Takahashi, A., Kuroki, M. & Watanuki, Y. 1999. Sexing by external measurements of adult Rhinoceros auklets breeding on Teuri Island. Japanese Journal of Ornithology, 48, 145-150.
- O -
Oh, K. P. & Badyaev, A. V. 2006. Adaptative genetic complementarity in mate choice coexists with selection for elaborate sexual traits. Proceedings of the Royal Society of London B, 273, 1913-1919.
Ortego, J., Aparicio, J. M., Calabuig, G. & Cordero, P. J. 2007b. Risk of ectoparasitism and genetic diversity in a wild Lesser kestrel population. Molecular Ecology, 16, 3712-3720.
Ortego, J., Calabuig, G., Cordero, P. J. & Aparicio, J. M. 2007a. Egg production and individual genetic diversity in Lesser kestrels. Molecular Ecology, 16, 2383-2392.
Ortego, J., Zapero, L., Calabuig, G., Cordero, P. J. & Aparicio, J. M. 2009. Physiological response to stress in fledgling Lesser kestrels Falco naumanni: the role of physical condition, sex and individual genetic diversity. Ibis, 151, 559-567.

Références bibliographiques
96
- P -
Palma, L., Mira, S., Cardia, P., Beja, P., Guillemaud, T., Ferrand, N., Cancela, M. L. & da Fonseca, L. C. 2001. Sexing Bonelli's Eagle nestlings: Morphometrics versus molecular techniques. Journal of Raptor Research, 35, 187-193.
Palmer, A. R. 1994. Fluctuating asymmetry analyses: a primer. In: Developmental Instability: Its Origins and Evolutionary Implications (Ed. by Markow, T. A.), pp. 335-364. Dordrecht, Netherland: Kluwer.
Palmer, A. R. 1996. Waltzing with asymmetry. BioScience, 46, 518-532.
Palmer, A. R. 2000. Quasireplication and the contract of error: lessons from sex ratios, heritabilities and fluctuating asymmetry. Annual Review of Ecology and Systematics, 31, 441-480.
Palmer, A. R. & Strobeck, C. 1986. Fluctuating asymmetry: measurement, analysis, patterns. Annual Review of Ecology and Systematics, 17, 391-421.
Palmer, A. R. & Strobeck, C. 1992. Fluctuating asymmetry as a measure of developmental stability: implication of non-normal distributions and power of statistical tests. Acta Zoologica Fennica, 191, 57-72.
Palmer, A. R. & Strobeck, C. 2003. Fluctuating Asymmetry Analyses Revisited. In: Developmental Instability (DI): Causes and Consequences (Ed. by Polak, M.), pp. 279-319. New York, USA: Oxford University Press.
Pemberton, J. M., Slate, J., Bancroft, D. R. & Barrett, J. A. 1995. Nonamplifying alleles at microsatellite loci: a caution for parentage and population studies. Molecular Ecology, 4, 249-252.
Petren, K., Grant, B. R. & Grant, P. R. 1999. A phylogeny of Darwin's finches based on microsatellite DNA length variation. Proceedings of the Royal Society of London B, 266, 321-329.
Petrides, G. A. 1950. Notes on determination of sex and age in the woodcock and Mourning dove. The Auk, 67, 357-360.
Phillips, R. A. & Furness, R. W. 1997. Predicting the sex of Parasitic jaegers by discriminant analysis. Colonial Waterbirds, 20, 14-23.
Pike, T. W. 2005. Sex ratio manipulation in response to maternal condition in pigeons: evidence for pre-ovulatory follicle selection. Behavioral Ecology and Sociobiology, 58, 407-413.
Pitzer, S., Hull, J., Ernest, H. B. & Hull, A. C. 2008. Sex determination of three raptor species using morphology and molecular techniques. Journal of Field Ornithology, 79, 71-79.

Références bibliographiques
97
Powell, J. R. 1987. Perspectives on Genetics: Anecdotal, Historical, and Critical Commentaries - "In the Air"—Theodosius Dobzhansky's Genetics and the Origin of Species. Genetics, 117, 363-366.
Pramuk, J. B., Hass, C. A. & Hedges, S. B. 2001. Molecular phylogeny and biogeography of West Indian toads (Anura: Bufonidae). Molecular Phylogenetics and Evolution, 20, 294-301.
- Q -
Quintana, F., Somoza, G., Uhart, M., Cassará, C., Gandini, P. & Frere, E. 2003. Sex determination of adult Rock shags by molecular sexing and morphometric parameters. Journal of Field Ornithology, 74, 370-375.
- R -
R Development Core Team 2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
Raffaele, H., Wiley, J., Garrido, O., Keith, A. & Raffaele, J. 2006. Les oiseaux des Antilles. Waterloo, Canada: Michel Quintin.
Raymond, M. & Rousset, F. 1995. An exact test for population differentiation. Evolution, 49, 1280-1283.
Remisiewicz, M. & Wennerberg, L. 2006. Differential migration strategies of the Wood sandpiper (Tringa glareola) - genetic analyses reveal sex differences in morphology and spring migration phenology. Ornis Fennica, 83, 1-10.
Reynolds, S. J., Martin, G. R., Wallace, L. L., Wearn, C. P. & Hughes, B. J. 2008. Sexing Sooty terns on Ascension Island from morphometric measurements. Journal of Zoology, 274, 2-8.
Richardson, D. S., Komdeur, J. & Burke, T. 2004. Inbreeding in the Seychelles warbler: environment-dependent maternal effects. Evolution, 58, 2037-2048.
Ricklefs, R. E. & Bermingham, E. 2004. History and the species-area relationship in Lesser Antillean birds. The American Naturalist, 163, 227-239.
Ricklefs, R. E. & Bermingham, E. 2008. The West Indies as a laboratory of biogeography and evolution. Philosophical Transactions of the Royal Society of London B, 363, 2393-2413.
Rijks, J. M., Hoffman, J. I., Kuiken, T., Osterhaus, A. D. M. E. & Amos, W. 2008. Heterozygosity and lungworm burden in Harbour seals (Phoca vitulina). Heredity, 100, 587-593.

Références bibliographiques
98
Rivera-Milán, F. F. 1992. Distribution and relative abundance patterns of columbids in Puerto Rico. The Condor, 94, 224-238.
Rivera-Milán, F. F. 1996. Nest density and success of Columbids in Puerto Rico. The Condor, 98, 100-113.
Rivera-Milán, F. F. 1997. Seasonal and annual changes in the population density of Zenaida doves in the xerophytic forest of Guanica, Puerto Rico. Journal of Field Ornithology, 68, 259-272.
Rivera-Milán, F. F. 1999. Population dynamics of Zenaida doves in Cidra, Puerto Rico. Journal of Wildlife Management, 63, 232-244.
Rivera-Milán, F. F. 2001. Transect survey of columbid nests on Puerto Rico. The Condor, 103, 332-342.
Rivera-Milán, F. F. & Schaffner, F. C. 2002. Demography of Zenaida doves on Cayo Del Agua, Culebra, Puerto Rico. The Condor, 104, 587-597.
Rivera-Milán, F. F. & Vásquez, M. 2000. Demography of Zenaida doves in Puerto Rico. The Condor, 102, 385-391.
Robertson, B. C. & Gemmell, N. J. 2006. PCR-based sexing in conservation biology: wrong answers from an accurate methodology. Conservation Genetics, 7, 267-271.
Roca, A. L., Bar-Gal, G. K., Eizirik, E., Helgen, K. M., Maria, R., Springer, M. S., O’Brien, S. J. & Murphy, W. J. 2004. Mesozoic origin for West Indian insectivores. Nature, 429, 649-651.
Rodriguez, C., Bustamante, J., Martinez-Cruz, B. & Negro, J. J. 2005. Evaluation of methods for gender determination of Lesser kestrel nestlings. Journal of Raptor Research, 39, 127-133.
Rodriguez, E. F., Pugesek, B. H. & Diem, K. L. 1996. A sexing technique for California gulls breeding at Bamforth Lake, Wyoming. Journal of Field Ornithology, 67, 519-524.
Rodríguez-Durán, A. & Kunz, T. H. 2001. Biogeography of West Indian Bats: An Ecological Perspective. In: Biogeography of the West Indies: Patterns and Perspectives (Ed. by Woods, C. A. & Sergile, F. E.), pp. 355-368: CRC Press
Rossiter, S. J., Jones, G., Ransome, R. D. & Barratt, E. M. 2001. Outbreeding increases offspring survival in wild Greater Horseshoe bats (Rhinolophus ferrumequinum). Proceedings of the Royal Society of London B, 268, 1055-1061.
Rubega, M. A. 1996. Sexual size dimorphism in Red-necked phalaropes and functional significance of non sexual bill structure variation for feeding performance. Journal of Morphology, 228, 45-60.

Références bibliographiques
99
Rubenstein, D. R. 2007. Female extrapair mate choice in a cooperative breeder: trading sex for help and increasing offspring heterozygosity. Proceedings of the Royal Society of London B, 274, 1895-1903.
Ruiz-López, M. J., Roldán, E. R. S., Espeso, G. & Gomendio, M. 2009. Pedigrees and microsatellites among endangered ungulates: what do they tell us? Molecular Ecology, 18, 1352-1364.
Ryder, J. P. 1978. Sexing Ring-billed gulls externally. Bird Banding, 49, 218-222.
- S -
Santiago-Alarcon, D. & Parker, P. G. 2007. Sexual size dimorphism and morphological evidence supporting the recognition of two subspecies in the Galápagos dove. The Condor, 109, 132-141.
Santiago-Alarcon, D., Tanksley, S. M. & Parker, P. 2006. Morphological variation and genetic structure of Galápagos dove (Zenaida galapagoensis) populations: issues in conservation for the Galápagos bird fauna. The Wilson Journal of Ornithology, 118, 194-207.
Sato, A., O'Huigin, C., Figuerola, F., Grant, P. R., Grant, B. R., Tichy, H. & Klein, J. 1999. Phylogeny of Darwin’s finches as revealed by mtDNA sequences. Proceedings of the National Academy of Sciences of the United States of America, 96, 5101-5106.
Schnell, G. D., Worthen, G. L. & Douglas, M. E. 1985. Morphometric assessment of sexual dimorphism in skeletal elements of California gulls. The Condor, 87, 484-493.
Schroeder, J., Lourenco, P. M., van der Velde, M., Hooijmeijer, J., Both, C. & Piersma, T. 2008. Sexual dimorphism in plumage and size in Black-tailed godwits Limosa limosa limosa. Ardea, 96, 25-37.
Seddon, N., Amos, W., Mulder, R. A. & Tobias, J. A. 2004. Male heterozygosity predicts territory size, song structure and reproductive success in a cooperatively breeding bird. Proceedings of the Royal Society of London B, 271, 1823-1829.
Seferta, A., Guay, P. J., Marzinotto, E. & Lefebvre, L. 2001. Learning differences between feral pigeons and Zenaida doves: the role of neophobia and human proximity. Ethology, 107, 281-293.
Selkoe, K. A. & Toonen, R. J. 2006. Microsatellites for ecologists: a practical guide to using and evaluating microsatellite markers. Ecology Letters, 9, 615-629.
Setiawan, A. N., Darby, J. T. & Lambert, D. M. 2004. The use of morphometric measurements to sex Yellow-eyed penguins. Waterbirds, 27, 96-101.
Seutin, G., White, B. N. & Boag, P. T. 1991. Preservation of avian blood tissue samples for DNA analyses. Canadian Journal of Zoology, 69, 82-90.

Références bibliographiques
100
Shamis, J. D. & Forrester, D. J. 1977. Haematozoan parasites of Mourning doves in Florida. Journal of Wildlife Diseases, 13, 349-355.
Shealer, D. A. & Cleary, C. M. 2007. Sex determination of adult black terns by DNA and morphometrics: tests of sample size, temporal stability and geographic specificity in the classification accuracy of discriminant functions models. Waterbirds, 30, 180-188.
Sheldon, L. D., Chin, E. H., Gill, S. A., Schmaltz, G., Newman, A. E. M. & Soma, K. K. 2008. Effects of blood collection on wild birds: an update. Journal of Avian Biology, 39, 369-378.
Shull, G. H. 1946. Hybrid seed corn. Science, 103, 547-550.
Shull, G. H. 1948. What is "heterosis"? Genetics, 33, 439-446.
Sikora, A. & Dubiec, A. 2007. Sex identification of Jack snipe Lymnocryptes minimus by discriminant analysis of morphometric measurements. Ardea, 95, 125-133.
Simmons, L. W., Tomkins, J. L., Kotiaho, J. S. & Hunt, J. 1999. Fluctuating paradigm. Proceedings of the Royal Society of London B, 266, 593-595.
Skúlason, S. & Smith, T. B. 1995. Resource polymorphisms in vertebrates. Trends in Ecology & Evolution, 10, 366-370.
Slaski, A. 2008. Dimorphisme sexuel et homogamie chez une espèce monogame tropicale, la tourterelle à queue carrée. Mémoire de Master 2ème année, Université de Bourgogne, Dijon, France.
Slate, J., Kruuk, L. E. B., Marshall, T. C., Pemberton, J. M. & Clutton-Brock, T. H. 2000. Inbreeding depression influences lifetime breeding success in a wild population of red deer (Cervus elaphus). Proceedings of the Royal Society of London B, 267, 1657-1662.
Slate, J. & Pemberton, J. M. 2002. Comparing molecular measures for detecting inbreeding depression. Journal of Evolutionary Biology, 15, 20-31.
Small, M. F., Baccus, J. T., Mink, J. N. & Roberson, J. A. 2005. Hematologic responses in captive White-winged doves (Zenaida asiatica) induced by various radiotransmitter attachments. Journal of Wildlife Diseases, 41, 387-394.
Smith, J. N. M. & Arcese, P. 1989. How fit are floaters? Consequences of alternative territorial behaviors in a nonmigratory sparrow. The American Naturalist, 133, 830-845.
Smith, T. B. & Skúlason, S. 1996. Evolutionary significance of resource polymorphisms in fishes, amphibians, and birds. Annual Review of Ecology and Systematics, 27, 111-133.
Snyder, B. F. & Gowaty, P. A. 2007. A reappraisal of Bateman's classic study of intrasexual selection. Evolution, 61, 2457-2468.

Références bibliographiques
101
Sohmers, S. H. & Gustafson, R. 1993. Plants and Flowers of Hawaii. Honolulu, Hawaii: University of Hawaii Press.
Sol, D., Elie, M., Marcoux, M., Chrostovsky, E., Porcher, C. & Lefebvre, L. 2005. Ecological mechanisms of resource polymorphism in Zenaida doves of Barbados. Ecology, 86, 2397-2407.
Speed, R. C. 1994. Barbados and the Lesser Antilles. In: Caribbean Geology: An Introduction, pp. 179-192. Kingston, Jamaica: University of the West Indies Publishers's Association.
Stutchbury, B. J. M. 1992. Experimental evidence that bright coloration is not important for territory defense in Purple martins. Behavioral Ecology and Sociobiology, 31, 27-33.
Stutchbury, B. J. M. & Morton, E. S. 2001. Behavioral Ecology of Tropical Birds. San Diego, USA: Academic Press.
Stutchbury, B. J. M. & Morton, E. S. 2008. Recent advances in the behavioral ecology of tropical birds. The Wilson Journal of Ornithology, 121, 26-37.
Svagelj, W. S. & Quintana, F. 2007. Sexual size dimorphism and sex determination by morphometric measurements in breeding Imperial shags (Phalacrocorax atriceps). Waterbirds, 30, 97-102.
Svensson, L. M. E. & Ricklefs, R. E. 2009. Low diversity and high intra-island variation in prevalence of avian Haemoproteus parasites on Barbados, Lesser Antilles. Parasitology, 136, 1121-1131.
Swanson, D. A. & Rappole, J. H. 1992. Determining the sex of adult White-winged doves by cloacal characteristics. North American Bird Bander, 17, 137-139.
Sweeney, J. J. & Tatner, P. 1996. Sexing wrens Troglodytes troglodytes indigenus using morphological measurements and discriminant analysis. Bird Study, 43, 342-350.
- T -
Taberlet, P. & Bouvet, J. 1991. A single plucked feather as a source of DNA for bird genetic studies. The Auk, 108, 959-960.
Taberlet, P., Griffin, S., Goossens, B., Questiau, S., Manceau, V., Escaravage, N., Waits, L. P. & Bouvet, J. 1996. Reliable genotyping of samples with very low DNA quantities using PCR. Nucleic Acids Research, 24, 3189-3194.
Tallmon, D. A. & Mills, L. S. 2004. Edge effects and isolation: Red-backed voles revisited. Conservation Biology, 18, 1658-1664.

Références bibliographiques
102
Tarvin, K. A., Webster, M. S., Tuttle, E. M. & Pruett-Jones, S. 2005. Genetic similarity of social mates predicts the level of extrapair paternity in Splendid Fairy-wrens. Animal Behaviour, 70, 945-955.
Thalmann, S., Baker, B., Hindell, M., Double, M. C. & Gales, R. 2007. Using biometric measurements to determine gender of Flesh-footed shearwaters, and their application as a tool in long-line by-catch management and ecological field studies. Emu, 107, 231-238.
Thompson, B. 1995. Stepwise regression and stepwise discriminant analysis need not apply here: a guidelines editorial. . Educational and Psychological Measurements 55, 525-534.
Tigreros, N., South, A., Fedina, T. & Lewis, S. 2009. Does fertilization by proxy occur in Tribolium beetles? A replicated study of a novel mechanism of sperm transfer. Animal Behaviour, 77, 555-557.
Tomkins, J. L. & Kotiaho, J. S. 2004. Publication bias in meta-analysis: seeing the wood for the trees. Oikos, 104, 194-196.
Traxler, B., Brem, G., Iler, M. & Achmann, R. 2000. Polymorphic DNA microsatellites in the domestic pigeon, Columba livia var. domestica. Molecular Ecology, 9, 366-368.
Tsitrone, A., Rousset, F. & David, P. 2001. Heterosis, marker mutational processes and population inbreeding history. Genetics, 159, 1845-1859.
- U -
Ura, T., Azuma, N., Hayama, S. & Higashi, S. 2005. Sexual dimorphism of Latham's snipe (Gallinago hardwickii). Emu, 105, 259-262.
Urfi, A. J. & Kalam, A. 2006. Sexual size dimorphism and mating pattern in the Painted stork (Mycteria leucocephala). Waterbirds, 29, 489-496.
- V -
van de Pol, M., Oosterbeek, K., Rutten, A. L., Ens, B. J., Tinbergen, J. M. & Verhulst, S. 2009. Biometric sex discrimination is unreliable when sexual dimorphism varies within and between years: an example in Eurasian Oystercatchers Haematopus ostralegus. Ibis, 151, 171-180.
van Dongen, S. 1999. Accuracy and power in fluctuating asymmetry studies: effects of sample size and number of within-subject repeats. Journal of Evolutionary Biology, 12, 547-550.
van Dongen, S. 2006. Fluctuating asymmetry and developmental instability in evolutionary biology: past, present and future. Journal of Evolutionary Biology, 19, 1727-1743.

Références bibliographiques
103
van Dongen, S. & Lens, L. 2000. The evolutionary potential of developmental instability. Journal of Evolutionary Biology, 13, 326-335.
van Dongen, S., Sprengers, E., Löfstedt, C. & Matthysen, E. 1999. Fitness components of male and female Winter moths (Operophtera brumata L.) (Lepidoptera, Geometridae) relative to measures of body size and asymmetry. Behavioral Ecology, 10, 659-665.
van Franeker, J. A. & ter Braak, J. F. 1993. A generalized discriminant for sexing Fulmarine petrels from external measurements. The Auk, 110, 492-502.
Vögeli, M., Serrano, D., Tella, J. L., Mendez, M. & Godoy, J. A. 2007. Sex determination Dupont's lark Chersophilus duponti using molecular sexing and discriminant functions. Ardeola, 54, 69-79.
Volodin, I. A., Kaiser, M., Matrosova, V., Volodina, E. V., Klenova, A. V., Filatova, O. A. & Kholodova, M. 2008. The technique of noninvasive distant sexing for four monomorphic Dendrocygna whistling duck species by their loud whistles. Bioacoustics, 18, 277-290.
Volodin, I. A., Volodina, E. V., Klenova, A. V. & Filatova, O. A. 2005. Individual and sexual differences in the calls of the monomorphic White-faced Whistling duck Dendrocygna viduata. Acta Ornithologica, 40, 43-52.
- W -
Wallace, A. R. 1869. The Malay Archipelago: The land of the Orang-utan, and the Bird of Paradise. A Narrative of Travel, with Sketches of Man and Nature London, UK: Macmillan & Co.
Wallace, A. R. 1876. The Geographical Distribution of Animals with a Study of the Relations of Living and Extinct Faunas as Elucidating the Past Changes of the Earth's Surface. London, UK: Macmillan & Co.
Wallace, A. R. 1880. Island Life; or, the Phenomena and Causes of Insular Faunas and Floras, Including a Revision and Attempted Solution of the Problem of Geological Climates. London, UK: Macmillan & Co.
Watson, J. D. & Crick, F. H. 1953. Molecular structure of nucleic acids. Nature, 171, 737-738.
Wattier, R., Engel, C. R., Saumitou-Laprade, P. & Valero, M. 1998. Short allele dominance as a source of heterozygote deficiency at microsatellite loci: experimental evidence at the dinucleotide locus Gv1CT in Gracilaria gracilis (Rhodophyta). Molecular Ecology, 7, 1569-1573.
Weber, J. L. & Wong, C. 1993. Mutation of human short tandem repeats. Human Molecular Genetics, 2, 1123-1128.

Références bibliographiques
104
Weidinger, K. & van Franeker, J. A. 1998. Applicability of external measurements to sexing of the Cape petrel Daption capense at within-pair, within-population and beetween-population scales. Journal of Zoology, London, 245, 473-482.
Weismann, F. L. A. 1891. On Heredity. In: Essays upon Heredity (Ed. by Poulton, E. B., Schöndland, S. & Shipley, A. E.), pp. 67-106. Oxford: Clarendon Press.
Wheelwright, N. T., Trussell, G., Devine, J. P. & Anderson, R. 1994. Sexual dimorphism and population sex ratios in juvenile Savannah sparrows. Journal of Field Ornithology, 65, 520-529.
Whittaker, R. J. & Fernández-Palacios, J. M. 2007. Island Biogeography. Ecology, Evolution, and Conservation. New York, USA: Oxford University Press.
Wiley, J. W. 1991. Ecology and behavior of the Zenaida dove. Ornitología Neotropical, 2, 49-75.
Williams, B. K. & Titus, K. 1988. Assessment of sampling stability in ecological applications of discriminant analysis. Ecology, 69, 275-285.
Wilson, A. C. C., Massonnet, B., Simon, J.-C., Prunier-Leterme, N., Dolatti, L., Llewellyn, K. S., Figueroa, C. C., Ramirez, C. C., Blackman, R. L., Estoup, A. & Sunnucks, P. 2004. Cross-species amplification of microsatellite loci in aphids: assessment and application. Molecular Ecology Notes, 4, 104-109.
Winker, K. 1998. The concept of floater. Ornitología Neotropical, 9, 111-119.
Winker, K., Klicka, J. T. & Voelker, G. 1996. Sexual size dimorphism in birds from southern Veracruz, Mexico. 2. Thryothorus maculipectus and Henicorhina [Leucosticta] prostheleuca. Journal of Field Ornithology, 67, 236-251.
Winker, K., Voelker, G. A. & Klicka, J. T. 1994. A morphometric examination of sexual dimorphism in the Hylophilus, Xenops, and an Automolus from southern Veracruz, Mexico. Journal of Field Ornithology, 65, 307-323.
- Z -
Zenatello, M. & Kiss, J. B. 2005. Biometrics and sex identification of the Rose-coloured starling Sturnus roseus. Ringing & Migration, 22, 163-166.
Zwarts, L., Hulscher, J. B., Koopman, K. & Zegers, P. M. 1996. Discriminating the sex of oystercatchers Haematopus ostralegus. Ardea, 84A, 1-12.

105
ANNEXES

ANNEXE 1
Monceau, K., Cézilly, F. & Dechaume-Moncharmont, F.-X. Sexing birds using discriminant
function analysis based on morphometric measurements: A critical appraisal.
Manuscrit en préparation

SEXING BIRDS USING DISCRIMINANT FUNCTION ANALYSIS 1
BASED ON MORPHOMETRIC MEASUREMENTS : A CRITICAL APPRAISAL 2
3
MONCEAU K., CEZILLY , F. & DECHAUME-MONCHARMONT, F.-X. 4
5
Université de Bourgogne, 6
Equipe Ecologie Evolutive, UMR CNRS 5561 Biogéosciences 7
8
Abstract 9
Despite the existence of alternative techniques, sexing birds using discriminant function 10
analysis based on morphometric measures remains a popular method among field 11
ornithologists. However, behind the apparent standardization and relative simplicity of the 12
technique, lie several subtle differences and pitfalls that have been somewhat neglected in 13
many published studies. Using a large data set on the Zenaida dove (Zenaida aurita), we 14
compare three methods commonly used to estimate the proportion of correctly classified 15
males and females by DFA: resubstitution, jack-knife or sample splitting. Simulations based 16
of 525 adults molecularly sexed birds showed that these procedures may lead to opposite 17
conclusions, especially when sample size is small. We call for caution in the use of 18
resubstitution techniques which are clearly over-optimistic. We instead recommend the use of 19
jack-knife cross-validation procedure, and we stress out the importance of large data sample 20
size. Finally, we show that moderate measurement error only marginally affects the accuracy 21
of the discriminant score. 22
23
Keyword : Morphometrical sexing, discriminant function analysis, cross-validation, sample 24
size effect, measurement effect 25

INTRODUCTION 26
27
Sex-related differences in the ecology of behaviour of bird species are crucial to our 28
understanding of sexual selection and mating systems (Andersson 1994), and may also have 29
important consequences in terms of management and conservation (Zavalaga & Parades 1997, 30
Fernandez-Juricic et al. 2009). Therefore, distinguishing sexes is of paramount importance in 31
avian studies. However, although sexual dimorphism can be conspicuous, especially in the 32
case of polygynous species, differences between male and female birds are often tenuous, or 33
escape the human eye (Cuthill et al. 1999). Several techniques have therefore been developed 34
to alleviate this difficulty, such as anatomical examination (Petrides 1950, Miller & Wagner 35
1955, Shamis & Forrester 1977, Bancroft 1984, Malacalaza & Hall 1988, Swanson & 36
Rappole 1992, Lorentsen & Røv 1994, Small et al. 2005), vocalization analyses (Blakesley et 37
al. 1990, Murie et al. 1991, Bretagnolle & Thibault 1995, Lo Valvo 2001, Gill & Vonhof 38
2006, Bourgeois et al. 2007), or sex-specific behaviour analyses (Castoro & Guhl 1958, 39
Chardine & Morris 1989, Blakesley et al. 1990, Desrochers 1990, Hull 1996, Martínez-40
Gómez & Curry 1998, Flux & Innes 2001, Fletcher & Hamer 2003, Gill & Vonhof 2006, 41
Alarcos et al. 2007). Such techniques tend, however, to be time-consuming, and do not 42
always allow perfect sex assignation. Molecular techniques (Griffiths & Tiwari 1993; see 43
Dubiec & Zagalska-Neubauer 2006 for a recent review), on the other hand, are thought to be 44
more reliable, but require training and a licence to collect blood or tissues, and imply financial 45
costs to process samples. 46
Consequently, sexing based on morphometrics remains a popular technique among 47
field ornithologists. Indeed, small, albeit significant, differences in biometrics often exist 48
between males in females of monomorphic bird species, offering the possibility to 49
discriminate between sexes using statistical inference. To that end, various statistical analyses 50

have been used, such as logistic regression (Jeffrey et al. 1993, Iko et al. 2004, Ura et al. 51
2005, Gill & Vonhof 2006), principal component analysis (Rubega 1996, McCracken et al. 52
2000, Remisiewicz & Wennerberg 2006, Urfi & Kalam 2006, Shealer & Cleary 2007, 53
Schroeder et al. 2008), or generalised linear model (Hallgrimsson et al. 2008). However, 54
discriminant function analysis (DFA) remains the most commonly used procedure, and over 55
the last 40 years, more than one hundred studies on sex determination or sex dimorphism 56
using DFA have been published (source: Web of Science). DFA provides equations including 57
at least one component allowing the parting in two categories or more (Fisher 1936). 58
Classification of individuals of known sex in relation to morphological measurements using 59
DFA provides subsequently a quick and relatively easy determination of gender in individuals 60
of unknown sex. 61
However, caution should be exerted when resorting to morphometric measurements 62
and DFA to sex birds. In particular, both sample size (Toft & Shea 1983, Rotenberry & Wiens 63
1985) and measurement error (ME) (Lougheed et al. 1991) require particular attention. 64
Indeed, discriminating rate is known to increase with increasing sample size (Morrison 1984), 65
and various rules of thumb have been recommended to determine minimum sample size 66
(Morrison 1984). Following Bailey & Barnes (1990), ME can be defined as the variability of 67
a repeated measurement of a particular character taken on the same individual, relative to its 68
variability among individuals in a particular group. ME can arise from random or systematic 69
error that affects measurements, or systematic variation between two different measurers. 70
Repeated measurements on several subjects are thus required to quantify measurement error. 71
However, despite the fact that the need for repeated measures has been emphasized (see 72
Lessells & Boag 1987), few published studies using DFA mention ME, and its potential effect 73
on discrimination rate is rarely investigated (Flux & Innes 2001, Devlin et al. 2004, Kenward 74
et al. 2004; see however Mallory & Forbes 2005). However, differences within group or 75

among individuals can be biased due to ME, increasing the risk of type II error (Bailey & 76
Byrnes 1990, Lougheed et al. 1991, Yezerinac et al. 1992, Arnqvist & Mårtensson 1998). For 77
example, Francis & Mattlin (1986) have shown that discriminant power could fall from 89 % 78
to less than 50 % when a small amount of bias exists, due to measurement error. So far, the 79
simultaneous effects of sample size and ME on sex determination from morphometrics have 80
not been considered in avian studies. In addition, various statistical methods can be used to 81
estimate the proportion of correctly classified males and females by DFA, but their use is 82
rarely justified in the avian literature (see below). 83
Here, we address the problem using a twofold approach. First, using statistical 84
analyses and simulations, we analyze a large data set obtained in the Zenaida dove (Zenaida 85
aurita), to look at the relation between discriminant power and variation in sample size, 86
measurement error, and classification method. Second, we review the literature to assess the 87
proportion of published studies relying on adequate methodology, and provide 88
recommendations for future studies. 89
90
91
MATERIAL AND METHODS 92
93
SEXUAL DIMORPHISM AND SEX DETERMINATION IN THE ZENAIDA DOVE 94
95
The Zenaida dove is widely distributed through the Caribbean islands (Bahamas, 96
Greater and Lesser Antilles), the coast of the Yucatán Peninsula and adjacent islands. It 97
generally feeds on the ground in agricultural or urban areas, or in wood lands (Wiley 1991, 98
Rivera-Milán 1997, 1999, Sol et al. 2005). The species is sexually monomorphic, and 99

supposed differences in plumage coloration between males and females (del Hoyo et al. 1997, 100
Gibbs et al. 2001) have not received confirmation. 101
102
Captures and measurements 103
Zenaida doves were caught in four locations in the western part of the island of 104
Barbados: two locations where doves hold territories (Bellairs Research Institute and Sunset 105
Crest area, both in St James Parish) and two where they feed in large flocks at grain storage 106
facilities (Barbados Mills Compounds and Roberts Manufacturing Co. Ltd., both in St 107
Michael Parish). Captures were made from March to April in 2007 at the four locations using 108
walk-in baited drop traps (Goldberg et al. 2001, Sol et al. 2005). Birds were first banded with 109
an unique combination of colour plastic bands (A.C. Hughes Ltd., Hampton Hill, United 110
Kingdom) and one numbered aluminium ring from the National Museum of Natural Life 111
History of Paris. Then, the following characters were measured twice: three dimensions of the 112
bill at nostrils (length, depth and width), head plus bill length, left and right tarsus length, left 113
and right wing length, and tail (fig. 1). All measurements were made with a digital calliper 114
(precision: ± 0.2 mm), except for wings and tail length which were measured with a ruler 115
(precision: ± 1 mm). Blood sample (40 µl twice) for molecular sex determination was 116
obtained by puncture of the brachial vein and collected with sodium heparinised haematocrit 117
capillary tube (Hirschmann Laborgeräte, Germany). Samples were stored in 800 µl of a 118
special buffer containing 70% of absolute ethanol and 30% of Tris-EDTA buffer (TE, pH 8). 119
Weight was measured with a PESOLA® digital pocket scale MS 500, (Precision: ± 0.1 g, 120
Switzerland). All, birds were released where they were caught. An additional sample caught 121
in March 2006 was used only for testing molecular methods (see next section for details). 122
123
124

Molecular sexing 125
DNA extraction protocol was modified from Hillis et al. (1996). One hundred 126
microliters of blood in storage buffer were centrifuged at 4000 rpm for 1 min to pellet blood 127
cells. Supernatant was discarded before adding 200 µl of Queen’s Lysis buffer (Seutin et al. 128
1991) and three units of proteinase K. Samples were incubated at 55°C for at least three 129
hours. A double phenol-chloroform extraction was applied followed by another chloroform 130
extraction and then 150 µl of aqueous phase were transferred in 375 µl of -20°C cold 131
isopropanol for precipitating DNA. Jellyfishes (precipitated DNA) were transferred in a new 132
tube, washed three times with -20°C cold 70% ethanol, dried for 15 min at room temperature 133
and resuspended into 100 µl of Tris-EDTA buffer (pH 8). 134
Two sets of primers were initially tested for sexing Zenaida doves. The first one was 135
P2 (5′-TCTGCATCGCTAAATCCTTT-3’) and P8 (5′-CTCCCAAGGATGAGRAAYTG-3′) 136
defined by Griffiths et al. (1998). All individuals caught in 2006 were sexed with P2 / P8 at 137
least once and 21 (randomly chosen) were sexed twice to control. Then, 88 samples (also 138
randomly chosen) were tested with the second set of primers 2550F (5′-139
GTTACTGATTCGTCTACGAGA-3′) and 2718R (5′-ATTGAAATGATCCAGTGCTTG-3′) 140
defined by Fridolfsson & Ellegren (1999) for confirming results and evaluate PCR accuracy 141
with both set of primers. In all cases, PCR were performed using 3 µl of DNA mixed with 5 142
U/µl of HotMasterTM Taq polymerase (Eppendorf®), Taq HotMaster buffer 10 X 143
(Eppendorf®), dNTPs at 5mM, Pf and Pr both at 5 µM. PCR program was the same for both 144
set of primers: 30 at 94°C, 30 s at 94°C, 45 s at 46°C, 45s at 65°C and 5 min at 65°C. Then 145
PCR products were put on a 3% agarose gel and revealed with ethidium bromide under UV. 146
Results with P2/P8 were not obtained directly. After PCR, only one band (around 380 pb) was 147
visible for all individuals on 3% agarose gel. Two enzymes can be used Hae III or Dde I. The 148
last one was chosen because the restriction site has been demonstrated to be conserved on 149

CHD1-W in different genus (Griffiths & Tiwari 1993, Griffiths et al. 1996, Bermúdez-150
Humarán et al. 2002). Therefore, Dde I was applied on each PCR product: 2µl of PCR 151
product with 10µl of enzyme buffer, 9 µl of performed like this: 5h at 37°C and the last 10 152
min. at 65°C. Results were given after run on 3% agarose gel: two bands, were then visible 153
for one individual of each pair (female) and only one (Z). Sex identification with P2 / P8 154
coupled with DdeI (88) matched perfectly with sex identification using 2550F / 2718R. 155
Because 2550F / 2718R were easier to use than P2 / P8, birds caught in 2007 were all sexed 156
with those primers. 157
158
Analysis of sexual dimorphism 159
A total of 525 adults (294 females, 231 males) were used for morphometric analysis. 160
Normality (Shapiro-Wilk test) and homoscedascity (Levene test) were first checked. 161
Repeatability and measurement error were assessed using F ratios from type II ANOVA 162
(Lessells & Boag 1987, Bailey & Byrnes 1990, Yezerinac et al. 1992, Arnqvist & Mårtensson 163
1998, Señar 1999). As repeatability was satisfactory for all measurements (table 2), we used 164
the mean of first and second measurement of each parameter in subsequent analyses. 165
Each character was compared between males and females using a t test. Mean 166
different index (MDI) was used as an index of sexual dimorphism index and computed as: (Xf 167
/ Xm) x 100, with Xf et Xm corresponding to the means for the considered characters for 168
female and male respectively (Delestrade 2001, Helfenstein et al. 2004, Haggerty 2006). 169
170
Discriminant function analyses 171
We performed discriminant function analysis (DFA) on the 525 molecularly sexed adults 172
using the eight morphometrical parameters listed in table 2. Due to violation of the 173
assumption of homogeneity of variance-covariance matrices (box’s M test, χ²190 = 267.1, p = 174

0.0002), we performed quadratic DFA (Stevens 1992) using qda function from MASS 175
package (Venables & Ripley 2002) for R language (R Development Core Team 2009). 176
We compared three methods commonly used in bird sexing to estimate the proportion 177
of correctly classified individuals by a DFA: resubstitution, jack-knife or sample splitting. In 178
the first method, the sex of each individual is predicted using the discriminant function 179
calculated from the complete data set. With jack-knife (or leave-one-out) method, the sex of 180
an individual is predicted from the discriminant equation calculated after that individual has 181
been removed from the data set. This procedure was repeated until a sex was assigned to each 182
individual (Tabachnick & Fidell 1996). With sample splitting, the data set was randomly 183
divided in two sub-samples, and the training set (two third of the individuals) was used to 184
compute the discriminant function, while the remaining data were used to assess the accuracy 185
of the discriminant function (Picard & Berk 1990). 186
187
Simulations 188
We used simulations first to study the effect of the sample size on the estimated 189
accuracy of DFA classification. We defined 100 sample sizes ranging from 25 to 520 190
individuals, regularly spaced every five individuals. For each sample size, we randomly 191
selected 500 different sub-samples from the complete data set. For each one of these 50000 192
sub-samples, we performed DFA and assessed the discriminant rates based on resubstitution, 193
jack-knife or sample splitting methods. 194
We performed another series of simulations to address the effect of measurement 195
errors, using this time the complete data set (n = 525 individuals). For each bird, each 196
parameter was estimated as the mean of two measures. Measurement error corresponds to the 197
difference between the first and the second measure. This difference followed a normal 198
distribution centred around zero and of standard deviation σ. We simulated an increase in 199

measurement error by adding a random noise to every parameter. This random noise followed 200
a normal distribution with mean µ = 0 and standard deviation σ’ = k σ, proportional to the 201
corresponding standard deviation σ observed on the original data. We called measurement 202
noise coefficient, the coefficient of proportionality k. For a given simulation of data set, the 203
measurement noise coefficient k was the same for every parameter, but the standard deviation 204
σ was different for each parameter. Thus, the distributions used for the random generation of 205
the noise were different for each parameter. A coefficient k = 0 means that no random noise 206
has been added to the measures and we used the original data set. Note that it was not possible 207
to have an index lower than 0 because data accuracy in our simulations cannot exceed that of 208
data actually collected in the field. It was then only possible to simulate less accurate data. For 209
value of coefficient k, we simulated 500 data sets and performed a DFA on each data set. For 210
every DFA, the proportion of correctly sexed individuals was estimated using the jack-knife 211
procedure. 212
213
214
L ITERATURE SURVEY 215
A literature survey concerning studies involving DFA for sex identification or sexual 216
dimorphism in birds was done using Web of Science and additional references. From 1977 to 217
present, 85 papers were reviewed containing 105 analyses (see table 1). At the first reading, 218
papers differed in the DA procedure used for discriminant score evaluation in the use or not of 219
automated stepwise variable reduction. Due to this large variety, different kinds of 220
information were retained for subsequent analyses: the bird species and its family, the total 221
sample size, the discriminant rate, the validation process and the variable reduction (table 1). 222
Measurement error was also noted when it was mentioned. 223
224

RESULTS 225
226
Measurement error, repeatability and sexual dimorphism 227
228
Measurement error was low for all traits (less than 11%), except for bill depth for 229
which ME was around 22% (table 2). Males and females were significantly different for all 230
morphological traits and mean differences index (MDI) was above 95% for all characters 231
(table 2). 232
233
Discriminant function analysis 234
235
Effect of the method of validation and sample size 236
Consider the DFA based on the complete original data set (n = 525 adults). The 237
estimated proportions of correctly classified birds were only marginally different according to 238
the validation method. Resubstitution method estimated a proportion of correctly sexed adults 239
of 80.95% (87.41% of the females and 72.73% of the males). Jack-knife cross-validation led 240
to an estimated proportion of 80.38% (86.39% for the females and 72.73% for males). Sample 241
splitting method depends strongly on the random selection of training and test sets. A 242
bootstrapped estimate based of 500 iterations lead to a mean (+/- standard deviation) 243
proportion of correctly classified birds of 79.1 +/- 3.51 % (79.13 +/- 2.6 % of the females and 244
79.05 +/- 4.69 % of the males). 245
246
Effect of sample size 247
The differences between the three methods were far from negligible for small sample 248
size (figure 1). The estimated the proportions of correctly sexed birds were the most 249

optimistic when using the resubstitution method, and, more surprisingly, decreased with 250
increasing sample size (figure 1a). For small data set (n < 60 birds), the mean estimate was 251
greater than 90% of correctly classified adults. On the contrary, the mean estimates with jack-252
knife (figure 1b) or sample splitting (figure 1c) methods were consistent, and increased with 253
increasing sample size: the more birds were measured, the more accurate the discriminant 254
function was. A noticeable difference between these two methods was related to the variance 255
of the estimates. Variance was always much bigger with sample splitting, even for large data 256
set. 257
258
Effect of measurement error 259
When adding random noise to the data in order to simulate increasing measurement 260
error, the proportion of correctly classified individuals decreased (figure 2). However, this 261
decrease was quite slow. Setting a coefficient of proportionality k = 1 means that we added to 262
every variable a random noise with a standard deviation equal to the corresponding observed 263
standard deviation. With this added random noise, the measurement error of every parameter 264
approximately doubled. For example, the error for bill depth increased from 21.8% (raw data) 265
to 45.2% (with added noise), and the error for wing length increased from 1.25 % (raw data) 266
to 3.25% (with added noise). Despite this drop of accuracy in the measures, the mean 267
proportion of correctly classified individuals only decreased from 80.38% to 79.36%, a 268
proportional decrease of the original precision of less than 1.3% corresponding to 269
approximately five additional adults incorrectly classified. Moreover, in many simulations 270
increasing the measurement errors actually increased the discriminant score. 271
272
273
274

Literature survey 275
276
Thirty-one families including 77 different species were represented in our subset of the 277
existing literature. Sample size varied greatly from ten birds to 1891 (mean = 141.15) and 278
discriminant went from 63 to 100% (mean = 89.57%). Although a threshold of 30 individuals 279
is usually recommended, published studies using DFA did not systematically respect this 280
criteria (table 1). All types of validation procedure were represented, but not in equal 281
proportion: 43.5% relied on resubstitution (including cases were no particular method was 282
clearly mentioned), whereas jackknife and sample splitting accounted for 35.3% 21.2% of the 283
studies, respectively. Concerning stepwise variable reduction, 60% of the reviewed papers 284
used automatic procedure implemented in statistical software packages. Finally, measurement 285
error was mentioned in less than 6% of the papers. 286
287
288
DISCUSSION 289
290
Every study we have reviewed referred to the estimated proportion of correctly sexed 291
individuals in order to validate the discriminant equations they recommended for 292
morphometrical sexing. There are a number ways in which this proportion can be estimated, 293
and yet the choice of a particular validation method instead of another is sometimes perceived 294
as a minor question, more related to statistician debates than to field biology. For example, 295
jack-knife cross-validation and resubstitution techniques are believed to be largely consistent, 296
the latter being slightly more optimistic than the first one. On the contrary, our simulations 297
show that the validation techniques may have far from negligible effects on the analysis 298

predictions, and may even lead to completely opposite conclusions, especially for small to 299
medium sample size. 300
Mean and variance of estimated DFA rates have already be reported to be highly 301
variable according to the validation procedure used. It has been largely publicized in 302
statistical (Johnson & Wichern 1992, Huberty 1994, Manly 1994, Wehberg & Schumacher 303
2004) and applied (Eisenbeis 1977, Lance et al. 2000) literature that reclassification of the 304
original sampled used in construction the discriminant rule as a mean to estimate expected 305
error rates (resubstitution) lead to a biased and overly optimistic prediction. In one of the 306
oldest papers using DFA for birds sexing, Ryder (1978) explicitly raised the problem of 307
resubstitution and recommend not using it especially in case of comparison between studies. 308
But as the technique grew more popular within the ornithologist community, this advice has 309
been clearly overlooked. However, the fact that a package offers a particular validation 310
procedure is clearly not a sufficient reason to use it as a ‘black-box’ that makes a decision 311
without any explanation (Piraux & Palm 2001, Venables & Ripley 2002). 312
In accordance with previous studies (Huberty 1994, Lance et al 2000, Hastie et al. 313
2009), the bias tends to decrease as the size of the sample increases. Nevertheless, the bias 314
remained particularly sensitive to data of small or intermediate size (less than 200 individuals 315
in the present study). Only sub-sampling techniques (such jack-knife cross-validation or 316
sample splitting of the data into training and test sets) provide an unbiased estimate of the 317
model performance (Hand et al 2001, Piraux & Palm 2001, Wehberg & Schumacher 2004, 318
Hastie et al 2009). Estimates of the discriminant score based on jack-knife procedure and 319
sample splitting should therefore be preferred over apparent discriminant score based on 320
resubstitution. But these two sub-sampling techniques are not completely interchangeable. 321
When we split the sample and used only a portion of it, the resulting estimates was expected 322
to have high variance. Indeed, using a very large training set ensuring a precise computation 323

of the discriminant function, restrains the validation to a small test set. On the contrary, using 324
a large test set and a smaller training set, leads to a less precise computation of the 325
discriminant function. It is commonly recommended to randomly split the data in a large 326
training set of about two third of the original data set, letting the remaining third for the test 327
set (Picard & Berk 1990). Such procedure leads to a mean estimate of the proportion of 328
correctly classified individuals consistent with the mean estimate using jack-knife procedure. 329
However, the variance was much bigger, especially for large sample sizes. This is why jack-330
knife cross-validation should also be preferred against sample splitting procedure. 331
Assessment of discriminant performance is of crucial importance in practice, since it 332
gives a measure of the quality of the model ultimately chosen equation for sexing birds. Field 333
biologists are extremely sensitive to the proportion of misclassified birds using a published 334
discriminant equation or another. If the risk of error was reported to be high, one would prefer 335
to use alternative and sometimes more expensive, time consuming (behavioral or molecular 336
sexing) or (laparoscopy) invasive techniques. It is also possible that many authors restrained 337
themselves from publishing DFA with low discriminant score. Our simulations with data set 338
of small to medium size emphasize the large variance in the estimated proportion of 339
misclassified bird. Even with less-biased techniques such jack-knife cross-validation or 340
sample splitting, a low (or high) discriminant success can perfectly be obtained by chance. 341
In the discussion of many papers, the discriminant scores were also frequently 342
compared to previous studies, most of the times in order to claim that their new discriminant 343
function lead to better classification success, sometimes by increasing the previous one only 344
by few percents. It is obvious that such precise comparisons are totally meaningless if 345
different techniques are used to estimate the discriminant score. Even if the same validation 346
procedure has been followed, one should keep in mind that the estimates may be highly 347
variable in relation to the sample size. The absence of consensus about the validation 348

techniques and large disparity among predicted discriminant scores according to the most 349
commonly used techniques, as well as the effects of sample size, should be taken into account 350
in future meta-analyses. 351
Another use of discriminant scores comparison is also highly questionable. Some 352
authors compared apparent error (from resubmission procedure) and error estimated by 353
sample splitting in order to detect possible sampling bias (e.g. Hanners & Patton 1985). As 354
long as these two error rates were not too different, they concluded to the absence of sampling 355
bias. However, our simulations showed that important differences between these two 356
estimates are possible even in absence of particular sampling bias, especially for small data 357
sets. 358
Large sample sizes were previously requested for accurate prediction of the sex ratio 359
in natural population (Brennan et al 1991). Moreover, our simulations stressed out the crucial 360
importance of large sample sizes in DFA. Analysis lead not only to a more discriminant 361
equation with a higher jack-knifed discriminant score, but also a more accurate estimates of 362
this score because its variance shrinks with increased sample size (Wehberg & Schumacher 363
2004). Several authors carefully adjusted the number of birds measured and the number 364
morphological parameters in order to have a sample size at least three times bigger than the 365
number of variables used in the DFA (Williams & Titus 1988, McGarigal et al. 2000), but this 366
criterion is sometimes claimed to be arbitrary (James & McCulloch 1990). There is a large 367
variability of performance estimations among samples from different sizes. Especially 368
discriminant rate from small data sets might be highly unreliable (Isaksson et al. 2008). For 369
small sample sets relative to the number of variables, the analysis leads to poor discriminant 370
rate. This result is known as the curse of dimensionality (Bellman 1957), and is consistent 371
with previous observation (Marks & Dunn 1974). 372

Morphometrics measures are costly in time and stressful for the birds. Field biologists 373
have to limit themselves on a small set of morphological parameters easy to measure. It is 374
why many authors use automated stepwise procedures in order to identify a reasonable set of 375
variables (60 % of the papers listed table 1). A common procedure is to measure many 376
morphological variables on a large number of individual. Then stepwise discriminant function 377
analysis is applied to examine differences between sexes, and to rank the morphological 378
variables according to their "importance". But biologists should be extremely cautious with 379
the output of such automated analysis. Stepwise methods tend to capitalize on sampling error, 380
and tend to yield results that are not replicable (Thompson 1995). Moreover, there is no 381
reason for the stepwise procedures to lead to the best equation to sex their model species. At 382
best, one should consider the equation as a “suggested” reduced set of parameters, not 383
necessarily the most discriminating nor the simplest (Derksen & Keselman 1992). One should 384
then consider it as a selection of a subset of variables among others that does an adequate job 385
of prediction, and keep in mind that, even if predictive models can eventually be developed, 386
there is no guarantee that they will be useful for later studies (James & McCulloch 1990). 387
Automated stepwise procedures are not reliable for assessing the relative importance of 388
variables and they are not able to select from a set of variables those that are most influential. 389
Thus, several authors strongly recommend not using any stepwise procedure at all (James & 390
McCulloch 1990, Snyder 1991, Thompson 2001). The fact that some statistical packages offer 391
such routine is clearly not a sufficient reason to use them. Indeed, one peer-reviewed journal 392
even made it an editorial policy to summarily reject any article that uses the technique 393
(Babyak 2004). Several authors underlie the crucial importance of large data set and 394
recommend alternative procedures based on combination of variables into biologically 395
meaningful groups. Scientific judgment must play a leading role in variables selection (James 396
& McCulloch 1990, Babyak 2004). If stepwise procedures are nevertheless used, Manly 397

(1994) advises to check its validity by rerunning it several times with randomized subsamples 398
of individual to see how significant are the results obtained. 399
Most importantly, the greatest danger of the use of stepwise analysis is of leaping 400
directly from routine procedure to straightforward conclusions about biological causes. In any 401
case, the fact that some variables were selected by the stepwise procedure does not mean that 402
they are more important than non-selected predicators. 403
Another conclusion of this study is the relative moderate effects of measurement errors 404
on the discriminant score. A simulated large increase in the measurement errors in our data 405
only marginally increased the estimated proportion of misclassification. Nevertheless, an 406
estimation of measurement errors has been rarely done in the paper we have reviewed and we 407
greatly need more data in order to conclude on the importance of the effect of such errors on 408
the accuracy of DFA. 409
The aim of this paper was to underlie many pitfalls that should be avoided and make 410
simple recommendations already well known from professional statisticians but apparently 411
somewhat neglected by field biologists. Despite the fact that DFA is extremely popular in the 412
ornithology literature, and constitute a standard technique for sexing birds, it is far from a 413
monolithic technique. The large disparity in methods for discriminant score estimation, the 414
sensitivity to sample size, and, to a lesser extent to measurement error, raise reasonable 415
doubts about usefulness of inter-studies comparisons and/or meta-analyses. 416
417
ACKNOWLEDGEMENTS 418
We are particularly grateful to Mr Steve A. Devonish and Natural Heritage 419
Department for allowing us to conduct our study in Barbados. We thank the Centre de 420
Recherches par le Baguage des Populations d’Oiseaux (Paris, France), and particularly 421
Olivier Dehorter and Denis Couvet, for providing us with metal rings and authorization to 422

ring Zenaida doves in the Caribbean. We also wish to thank Alison Mayers and Wayne 423
Worrell for hospitality, Richard Haynes, Victor Small, Sylvester Rowe, and Pat Odle for 424
logistic assistance, Laure Cauchard, Jérôme Moreau, Sébastien Motreuil, and Georges Prato 425
for assistance in catching and ringing doves, and Maria Gaillard and Rémi Wattier for help 426
and advice on molecular sexing. KM was supported by a doctoral grant from the Ministère de 427
la Recherche et de l'Enseignement Supérieur. Financial support was partly provided a 428
research grant from ANR (programme blanc, "Monogamix"). 429
430
REFERENCES 431
Alarcos, S., de la Cruz, C., Solís, E., Valencia, J. & García-Baquero, M. J. 2007. Sex 432
determination of Iberian Azure-winged magpies Cyanopica cyanus cooki by 433
discriminant analysis of external measurements. Ringing & Migration, 23, 211-216. 434
Andersson, M. 1994. Sexual Selection. Princeton University Press. 435
Arizaga, J., Aldalur, A., Herrero, A. & Galicia, D. 2008. Sex differentiation of Yellow-legged 436
gull (Larus michahellis lusitanius): the use of biometrics, bill morphometrics and wing 437
tip coloration. Waterbirds, 31, 211-219. 438
Arnould, J. P. Y., Dann, P. & Cullen, J. M. 2004. Determining the sex of Little penguins 439
(Eudyptula minor) in northern Bass Strait using morphometric measurements. Emu, 440
104, 261-265. 441
Arnqvist, G. & Mårtensson, T. 1998. Measurement error in geometric morphometrics: 442
empirical strategies to assess and reduce its impact on measures of shape. Acta 443
Zoologica Academiae Scientiarum Hungaricae, 44, 73-96. 444
Babyak, M. A. 2004. What you see may not be what you get: a brief, nontechnical 445
introduction to overfitting in regression-type models. Psychosomatic Medicine, 66, 446
411-421. 447

Bailey, R. C. & Byrnes, J. 1990. A new, old method for assessing measurement error in both 448
univariate and multivariate morphometric studies. Systematic Zoology, 39, 124-130. 449
Baker, A. J., Piersma, T. & Greenslade, A. D. 1999. Molecular vs. phenotypic sexing in Red 450
knots. The Condor, 101, 887-893. 451
Balbontin, J., Ferrer, M. & Casado, E. 2001. Sex determination in Booted eagles (Hieraaetus 452
pennatus) using molecular procedures and discriminant function analysis. Journal of 453
Raptor Research, 35, 20-23. 454
Bancroft, G. T. 1984. Growth and sexual dimorphism of the Boat-tailed grackle. The Condor, 455
86, 423-432. 456
Barré, N., de Garine Wichatitsky, M., Lecoq, R. & Maillard, J.-C. 2003. Contribution to the 457
knowledge of the New Caledonian Imperial pigeon Ducula goliath (Gray 1859) with 458
emphasis on sexual dimorphism. Notornis, 50, 155-160. 459
Bellman, R. 1957. Dynammic Programming. Princeton, USA: Princeton University Press. 460
Berlin, K. E., Simon, J. C., Pratt, T. K., Baker, P. E. & Kowalsky, J. R. 2001. Age and sex 461
determination of the Maui parrotbill. Journal of Field Ornithology, 72, 12-21. 462
Bermudez-Humarán, L. G., Chávez-Zamarripa, P., Guzmán-Velasco, A., Leal-Garza, C. H. & 463
Montes de Oca-Luna, R. 2002. Loss of restriction site DdeI, used for avian molecular 464
sexing, in Oreophasis derbianus. Reproduction in Domestic Animals, 37, 321-323. 465
Bertellotti, M., Tella, J. L., Godoy, J. A., Blanco, G., Forero, M. G., Donázar, J. A. & 466
Ceballos, O. 2002. Determining sex of Magellanic penguins using molecular 467
procedures and discriminant functions. Waterbirds, 25, 479-484. 468
Blakesley, J. A., Franklin, A. B. & Gutiérrez, R. J. 1990. Sexual dimorphism in Northern 469
Spotted owls from northwest California. Journal of Field Ornithology, 61, 320-327. 470
Bluso, J. D., Ackerman, J. T., Takekawe, J. Y. & Yee, J. L. 2006. Sexing Forster's terns using 471
morphometric measurements. Waterbirds, 29, 512-517. 472

Boersma, P. D. & Davies, E. M. 1987. Sexing monomorphic birds by vent measurements. The 473
Auk, 104, 779-783. 474
Bond, A. B., Wilson, K.-J. & Diamond, J. 1991. Sexual dimorphism in the kea Nestor 475
notabilis. Emu, 91, 12-19. 476
Bosch, M. 1996. Sexual size dimorphism and determination of sex in Yellow-legged gulls. 477
Journal of Field Ornithology, 67, 534-541. 478
Bourgeois, K., Curé, C., Legrand, J., Gómez-Díaz, Vidal, E., Aubin, T. & Mathevon, N. 2007. 479
Morphological versus acoustic analysis: what is the most efficient method for sexing 480
Yelkouan shearwaters Puffinus yelkouan? Journal of Ornithology, 148, 261-269. 481
Brennan, L. A., Buchanan, J. B., Schick, C. T. & Herman, S. G. 1991. Estimating sex-ratios 482
with discriminant function-analysis - the influence of probability cutpoints and 483
sample-size. Journal of Field Ornithology, 62, 357-366. 484
Brennan, L. A., Buchanan, J. B., Schick, C. T., Herman, S. G. & Johnson, T. M. 1984. Sex 485
determination of Dunlins in winter plumage. Journal of Field Ornithology, 55, 343-486
348. 487
Bretagnolle, V. & Thibault, J.-C. 1995. Method for sexing fledglings in Cory's shearwaters 488
and comments on sex-ratio variation. The Auk, 112, 785-790. 489
Campos, F., Hernández, M., Arizaga, J., Miranda, R. & Amezcua, A. 2005. Sex 490
differentiation of Corn buntings Miliaria calandra wintering in northern Spain. 491
Ringing & Migration, 22, 159-162. 492
Casaux, R. & Baroni, A. 2000. Sexual size dimorphism in the Antarctic shag. Waterbirds, 23, 493
489-493. 494
Castoro, P. L. & Guhl, A. M. 1958. Pairing behavior of pigeons related to aggressiveness and 495
territory. The Wilson Bulletin, 70, 57-69. 496

Chardine, J. W. & Morris, R. D. 1989. Sexual size dimorphism and assortative mating in the 497
Brown noddy. The Condor, 91, 868-874. 498
Childress, B., Harper, D., Hughes, B. & Ferris, C. 2005. Sex determination in the lesser 499
flamingo (Phoenicopterus minor) using morphological measurements. Ostrich, 76, 500
148-153. 501
Chochi, M., Nizuma, Y. & Takagi, M. 2002. Sexual differences in the external measurements 502
of Black-tailed gulls breeding on Rishiri Island, Japan. Ornithological Science, 1, 163-503
166. 504
Clark, R. G., James, P. C. & Morari, J. B. 1991. Sexing adult and yearling American crows by 505
external measurements and discriminant analysis. Journal of Field Ornithology, 62, 506
132-138. 507
Copello, S., Quintana, F. & Somoza, G. 2006. Sex determination and sexual size-dimorphism 508
in Southern Giant-petrels (Macronectes giganteus) from Patagonia, Argentina. Emu, 509
106, 141-146. 510
Coulson, J. C., Thomas, C. S., Butterfield, J. E. L., Duncan, N., Monaghan, P. & Shedden, C. 511
1983. The use of head and bill length to sex live gulls Laridae. Ibis, 125, 549-557. 512
Coulter, M. C. 1986. Assortative mating and sexual dimorphism in the Common tern. Wilson 513
Bulletin, 98, 93-100. 514
Counsilman, J. J., Nee, K., Jalil, A. K. & Keng, W. L. 1994. Discriminant analysis of 515
morphometric characters as a means of sexing mynas. Journal of Field Ornithology, 516
65, 1-7. 517
Cucco, M., Lingua, G., Bocchio, D., Acquarone, C. & Malacarne, G. 1999. Sex identification 518
in the moorhen (Gallinula chloropus) by flow cytometry and morphometric analysis. 519
Italian Journal of Zoology, 66, 1-6. 520

Cuthill, I.C., Bennett, A.T.D., Partridge, J.C. & Maier, E.J. 1999. Plumage reflectance and the 521
objective assessment of avian sexual dichromatism. The American Naturalist, 153, 522
183-200. 523
del Hoyo, J., Elliott, A. & Sargatal, J. 1997. Handbook of the Birds of the World. Barcelona, 524
Spain. 525
del mar Delgado, M. & Penteriani, V. 2004. Gender determination of Eurasian Eagle-owls 526
(Bubo bubo) by morphology. Journal of Raptor Research, 38, 375-377. 527
Delestrade, A. 2001. Sexual size dimorphism and positive assortative mating in alpine 528
choughs. 529
Derksen, S. & Keselman, H. J. 1992. Backward, forward and stepwise automated subset 530
selection algorithms: frequency of obtaining authentic and noise variables. British 531
Journal of Mathematical and Statistical Psychology, 45. 532
Desrochers, A. 1990. Sex determination of Black-capped chickadees with discriminant 533
analysis. Journal of Field Ornithology, 61, 79-84. 534
Devlin, C. M., Diamond, A. W. & Saunders, G. W. 2004. Sexing Arctic terns in the field and 535
laboratory. Waterbirds, 27, 314-320. 536
Donohue, K. C. & Dufty, A. M., Jr. 2006. Sex determination of Red-tailed hawks (Buteo 537
jamaicensis calurus) using DNA analysis and morphometrics. Journal of Field 538
Ornithology, 77, 74-79. 539
Dorr, B., King, D. T., Harrel, J. B., Gerard, P. & Spalding, M. G. 2005. The use of culmen 540
length to determine sex of the American White pelican. Waterbirds, 28, 102-106. 541
Dubiec, A. & Zagalska-Neubauer, M. 2006. Molecular techniques for sex identification in 542
birds. Biological Letters, 43, 3-12. 543

Eason, D., Millar, C. D., Cree, A., Halverson, J. L. & Lambert, D. M. 2001. A comparison of 544
five methods for assignment of sex in the takahe (Aves: Porphyrio mantelli). Journal 545
of Zoology, 253, 281-292. 546
Eisenbeis, R. A. 1977. Pitfalls in the application of discriminant analysis in business, finance, 547
and economics. The Journal of Finance, 32, 875-900. 548
Evans, D. R., Cavanagh, P. M., French, T. W. & Blodget, B. G. 1995. Identifying the sex of 549
Massachusetts herring gulls by linear measurements. Journal of Field Ornithology, 66, 550
128-132. 551
Evans, D. R., Hoopes, E. M. & Griffin, C. R. 1993. Discriminating the sex of Laughing gulls 552
by linear measurements. Journal of Field Ornithology, 64, 472-476. 553
Fernandez-Juricic, E., del Nevo, A.J. & Poston, R. 2009. Identification of individual and 554
population-level variation in vocalizations of the endangered Southern Willow 555
Flycatcher (Empidonax traillii extimus). The Auk, 126, 89-99. 556
Ferrer, M. & Delecourt, C. 1992. Sex Identification in the Spanish Imperial eagle. Journal of 557
Field Ornithology, 63, 359-364. 558
Figuerola, J., Garcia, L., Green, A. J., Ibañez, F., Mañez, M., del Valle, J. L., Garrido, H., 559
Arroyo, J. L. & Rodríguez, R. 2006. Sex determination in Glossy ibis chicks based on 560
morphological characters. Ardeola, 53, 229-235. 561
Fisher, R. A. 1936. The use of multiple measurements in taxonomic problems. Annals of 562
Eugenics, 7, 179-188. 563
Fletcher, K. L. & Hamer, K. C. 2003. Sexing terns using biometrics: the advantage of within-564
pair comparisons. Bird Study, 50, 78-83. 565
Flux, I. & Innes, J. 2001. A field technique for determining the sex of North Island kokapo 566
(Callaeas cinerea wilsoni). Notornis, 48, 217-223. 567

Fox, G. A., Cooper, C. R. & Ryder, J. P. 1981. Predicting the sex of herring gulls by using 568
external measurements. Journal of Field Ornithology, 52, 1-9. 569
Francis, R. I. C. C. & Mattlin, R. H. 1986. A possible pitfall in the morphometric application 570
of discriminant analysis: measurement bias. Marine Biology, 93, 311-313. 571
Frey, S. J. K., Rimmer, C. C., McFarland, K. P. & Menu, S. 2008. Identification and sex 572
determination of Bicknell's thrushes using morphometric data. Journal of Field 573
Ornithology, 79, 408-420. 574
Fridolfsson, A.-K. & Ellegren, H. 1999. A simple and universal method for molecular sexing 575
of non-ratite birds. Journal of Avian Biology, 30, 116 - 121. 576
Genovart, M., McMinn, M. & Bowler, D. 2003. A discriminant function for predicting sex in 577
the Balearic shearwater. Waterbirds, 26, 72-76. 578
Gibbs, D., Barnes, E. & Cox, J. 2001. Pigeons and Doves: A Guide to the Pigeons and Doves 579
of the World. pp. 615. New Haven, Connecticut, USA: Yale University Press. 580
Gill, S. A. & Vonhof, M. J. 2006. Sexing monochromatic birds in the field: cryptic sexual size 581
dimorphism in Buff-breasted wrens (Thryotorus leucotis). Ornitología Neotropical, 582
17, 409-418. 583
Glahn, J. F. & McCoy, R. B. 1995. Measurements of wintering Double-crested cormorants 584
and discriminant models of sex. Journal of Field Ornithology, 66, 299-304. 585
Goldberg, J. L., Grant, J. W. A. & Lefebvre, L. 2001. Effects of the temporal predictability 586
and spatial clumping of food on the intensity of competitive aggression in the Zenaida 587
dove. Behavioral Ecology, 12, 490-495. 588
Grecian, V. D., Diamond, A. W. & Chardine, J. W. 2003. Sexing razorbills Alca torda 589
breeding at Machias Seal Island, New Brunswick, Canada, using discriminant function 590
analysis. Atlantic Seabirds, 5, 73-80. 591

Green, P. T. & Theobald, C. M. 1989. Sexing birds by discriminant analysis: furthers 592
considerations. Ibis, 131, 442-447. 593
Green, P. T. 1982. Sexing rooks Corvus frugilegus by discriminant analysis. Ibis, 124, 320-594
324. 595
Griffiths, R. & Tiwari, B. 1995. Sex of the last wild Spix's macaw. Nature, 375, 454. 596
Griffiths, R., Daan, S. & Dijkstra, C. 1996. Sex identification in birds using two CHD genes. 597
Proceedings of the Royal Society of London B, 263, 1251-1256. 598
Griffiths, R., Double, M. C., Orr, K. & Dawson, R. J. C. 1998. A DNA test to sex most birds. 599
Molecular Ecology, 7, 1071-1075. 600
Gruys, R. C. & Hannon, S. J. 1993. Sex determination of hunter-killed and depredated 601
Willow ptarmigan using a discriminant-analysis. Journal of Field Ornithology, 64, 11-602
17. 603
Haggerty, T. M. 2006. Sexual size dimorphism and assortative mating in Carolina wrens. 604
Journal of Field Ornithology, 77, 259-265. 605
Hallgrimsson, G. T., Palsson, S. & Summers, R. W. 2008. Bill length: a reliable method for 606
sexing Purple sandpipers. Journal of Field Ornithology, 79, 87-92. 607
Hand, D. J., Mannila, H. & Smyth, P. 2001. Principles of Data Mining. Cambridge, USA: 608
The Massachusetts Institute of Technology. 609
Hanners, L. A. & Patton, S. R. 1985. Sexing Laughing gulls using external measurements and 610
discriminant analysis. Journal of Field Ornithology, 56, 158-164. 611
Hardy, A. R., Hirons, G. J. M., Stanley, P. I. & Huson, L. W. 1981. Sexual dimorphism in 612
size of Tawny owls (Strix aluco): a method for sexing in field studies. Ardea, 69, 181-613
184. 614
Hastie, T., Tibshirani, R. & Friedman, J. 2009. The Elements of Statistical Learning Data 615
Mining, Inference, and Prediction. Berlin, Germany: Springer Verlag. 616

Hayward, G. D. & Hayward, P. H. 1991. Body measurements of Boreal owls in Idaho and a 617
discriminant model to determine sex of live specimens. The Wilson Bulletin, 103, 497-618
500. 619
Helander, B., Hailer, F. & Vilà, C. 2007. Morphological and genetic sex identification of 620
White-tailed eagle Haliaeetus albicilla nestlings. Journal of Ornithology, 148, 435-621
442. 622
Helfenstein, F., Danchin, E. & Wagner, R. H. 2004. Assortative mating and sexual size 623
dimorphism in Black-legged kittiwakes. Waterbirds, 27, 350-354. 624
Hermosell, I. G., Balbontin, J., Marzal, A., Reviriego, M. & de Lope, F. 2007. Sex 625
determination in Barn swallows Hirundo rustica by means of discriminant analysis in 626
two European populations. Ardeola, 54, 93-100. 627
Herring, G., Gawlik, D. E. & Beerens, J. M. 2008. Sex determination for the Great egret and 628
White ibis. Waterbirds, 31, 298-303. 629
Hillis, D. M., Moritz, C. & Mable, B. K. 1996. Molecular Systematics, Second Edition. 630
Massachusets, USA: Sunderland. 631
Hipkiss, T. 2007. Can migrating Tengmalm's owls Aegolius funereus be reliably sexed in 632
autumn using simple morphometric measurements? Ringing & Migration, 23, 201-633
204. 634
Huberty, C. J. 1994. Applied Discriminant Analysis. New York, USA: Wiley. 635
Hull, C. L. 1996. Morphometric indices for sexing adult Royal Eudyptes schlegeli and 636
rockhopper E. chrysocome penguins at Macquarie Island. Marine Ornithology, 24, 23-637
27. 638
Iko, N. M., Dinsmore, S. J. & Knopf, F. L. 2004. Evaluating the use of morphometric 639
measurements from museum specimens for sex determination in Mountain plovers 640
(Charadrius montanus). Western North American Naturalist, 64, 492-496. 641

Isaksson, A., Wallman, M., Göransson, H. & Gustafsson, M. G. 2008. Cross-validation and 642
bootstrapping are unreliable in small sample classification. Pattern Recognition 643
Letters, 19, 1960-1965. 644
Jakubas, D. & Wojczulanis, K. 2007. Predicting the sex of dovekies by discriminant analysis. 645
Waterbirds, 30, 92-96. 646
James, F. C. & McCulloch, C. E. 1990. Multivariate analysis in ecology and systematics: 647
panacea or Pandora's box? Annual Review of Ecology and Systematics, 21, 129-166. 648
Jeffrey, J. J., Fancy, S. G., Lindsey, G. D., Banko, P. C., Pratt, T. K. & Jacobi, J. D. 1993. Sex 649
and age identification of palila. Journal of Field Ornithology, 64, 490-499. 650
Jehl, J. R., Henry, A. E. & Bond, S. I. 1998. Sexing Eared grebes by bill measurements. 651
Colonial Waterbirds, 21, 98-100. 652
Jodice, P. G. R., Lanctot, R. B., Gill, V. A., Roby, D. D. & Hatch, S. A. 2000. Sexing adult 653
Black-legged kittiwakes by DNA, behavior, and morphology. Waterbirds, 23, 405-654
415. 655
Johnson, R. A. & Wichern, D. W. 1992. Applied Multivariate Statistical Analysis. 3rd ed. 656
Englewood Cliffs, USA: Prentice-Hall. 657
Kenward, B., Rutz, C., Weir, A. A. S., Chapell, J. & Kacelnik, A. 2004. Morphology and 658
sexual dimorphism of the New Caledonian crow Corvus moneduloides, with notes on 659
its behaviour and ecology. Ibis, 146, 652-660. 660
Kerry, K. R., Agnew, D. J., Clarke, J. R. & Else, G. D. 1992. Use of Morphometric 661
Parameters for the Determination of Sex of Adelie Penguins. Wildlife Research, 19, 662
657-664. 663
Kesler, D. C., Lopes, I. F. & Haig, S. M. 2006. Sex determination of Pohnpei Micronesian 664
kingfishers using morphological and molecular genetic techniques. Journal of Field 665
Ornithology, 77, 229-232. 666

Kloskowski, J., Grela, P., Krogulec, J., Gaska, M. & Tchorzewski, M. 2006. Sexing Red-667
necked grebes Podiceps grisegena by molecular techniques and morphology. Acta 668
Ornithologica, 41, 176-180. 669
Koffijberg, K. & van Eerden, M. R. 1995. Sexual dimorphism in the cormorant 670
Phalacrocorax carbo sinensis: possible implications for differences in structural size. 671
Ardea, 83, 37-46. 672
Lance, R. F., Kennedy, M. L. & Leberg, P. L. 2000. Classification bias in discriminant 673
function analyses used to evaluate putatively different taxa. Journal of Mammology 674
81, 245-249. 675
Lessells, C. M. & Boag, P. T. 1987. Unrepeatable repeatabilities: a common mistake. The 676
Auk, 104, 116-121. 677
Liordos, V. & Goutner, V. 2008. Sex determination of Great cormorants (Phalacrocorax 678
carbo sinensis) using morphometric measurements. Waterbirds, 31, 203-210. 679
Lislevand, T., Marthinsen, G. & Lifjeld, J. T. 2009. Sex differences in body size and body 680
condition in breeding Temminck's stints Calidris temminckii. Journal of Ornithology, 681
150, 299-302. 682
Lo Valvo, M. 2001. Sexing adult Cory's shearwater by discriminant analysis of body 683
mesurements on Linosa Island (sicilian channel), Italy. Waterbirds, 24, 169-174. 684
Lorentsen, S.-H. & Røv, N. 1994. Sex determination of Antarctic petrels Thalassoica 685
antarctica by discriminant analysis of morphometric characters. Polar Biology, 14, 686
143-145. 687
Lougheed, S. C., Arnold, T. W. & Bailey, R. C. 1991. Measurement error of external and 688
skeletal variables in birds and its effect on pricipal components. The Auk, 108, 432-689
436. 690

Malacalaza, V. E. & Hall, M. A. 1988. Sexing adult King cormorans (Phalacrocorax 691
albiventer) by discriminant analysis. Colonial Waterbirds, 11, 32-37. 692
Mallory, M. L. & Forbes, M. R. 2005. Sex discrimination and measurement bias in Northern 693
fulmars Fulmarus glacialis from the Canadian Arctic. Ardea, 93, 25-36. 694
Manly, B. F. J. 1994. Multivariate Statistical Methods: A Primer. 2nd ed. London, UK: 695
Chapman and Hall. 696
Mariano-Jelicich, R., Madrid, E. & Faverol, M. 2007. Sexual dimorphism and diet 697
segregation in the Black skimmer Rynchops niger. Ardea, 95, 115-124. 698
Marks, S. & Dunn, O. J. 1974. Discriminant functions when covariance matrices are unequal. 699
Journal of the American Statistical Association, 69, 555-559. 700
Maron, J. L. & Myers, J. P. 1984. A description and evaluation of two techniques for sexing 701
wintering sanderlings. Journal of Field Ornithology, 55, 336-342. 702
Martinez-Gómez, J. E. & Curry, R. L. 1998. Distinguishing sex of Socorro mockingbirds by 703
body measurements. Ornitología Neotropical, 9, 103-110. 704
McCloskey, J. T. & Thompson, J. E. 2000. Aging and sexing Common snipe using 705
discriminant analysis. Journal of Wildlife Management, 64, 960-969. 706
McCracken, K. G., Paton, D. C. & Afton, A. D. 2000. Sexual size dimorphism of the Musk 707
duck. The Wilson Bulletin, 112, 457-466. 708
McGarigal, K., Cushman, S. & Stafford, S. 2000. Multivariate Statistics for Wildlife and 709
Ecology Research. New York, USA: Springer Verlag. 710
Meissner, W. & Pilacka, L. 2008. Sex identification of adult dunlins Calidris alpina alpina 711
migrating in autumn trough Baltic region. Ornis Fennica, 85, 135-139. 712
Meissner, W. 2005. Sex determination of juvenile dunlins migrating through the Polish Baltic 713
region. Journal of Field Ornithology, 76, 368-372. 714

Migot, P. 1986. Le goéland argenté Larus argentatus argenteus Brehm en Bretagne: 715
caractéristiques biométriques des reproducteurs. Alauda, 54, 269-278. 716
Miller, W. J. & Wagner, F. H. 1955. Sexing mature columbiformes by cloacal characters. The 717
Auk, 72, 279-285. 718
Morrison, M. L. 1984. Influence of sample size on discriminant function analysis of habitat 719
use by birds. Journal of Field Ornithology, 55, 330-335. 720
Murie, J. O., Davis, L. S. & McLean, I. G. 1991. Identifying the sex of Fiordland Crested 721
penguins by morphometric characters. Notornis, 38, 233-238. 722
Palomares, L. E., Arroyo, B. E., Marchamalo, J., Sainz, J. J. & Voslamber, B. 1997. Sex- and 723
age-related biometric variation of Black-headed gulls Larus ridibundus in Western 724
European populations. Bird Study, 44, 310-317. 725
Petrides, G. A. 1950. Notes on determination of sex and age in the Woodcock and Mourning 726
dove. The Auk, 67, 357-360. 727
Picard, R. R. & Berk, K. N. 1990. Data splitting. American Statistician, 44, 140-147. 728
Piraux, F. & Palm, R. 2001. Etude empirique des estimateurs des taux d’erreur en analyse 729
discriminante. Revue de Statistique Appliquée, 49, 71-85. 730
R Development Core Team (2009) R: a language and environment for statistical computing. 731
R Foundation for Statistical Computing. Vienna, Austria. ISBN:3-900051-07-0. 732
http://www.R-project.org 733
Remisiewicz, M. & Wennerberg, L. 2006. Differential migration strategies of the Wood 734
sandpiper (Tringa glareola) - genetic analyses reveal sex differences in morphology 735
and spring migration phenology. Ornis Fennica, 83, 1-10. 736
Renner, M. & Davis, L. S. 1999. Sexing Little penguins Eudyptula minor from Cook Strait, 737
New Zealand using discriminant function analysis. Emu, 99, 74-79. 738

Renner, M., Valencia, J., Davis, L. S., Saez, D. & Cifuentes, O. 1998. Sexing adult Gentoo 739
penguins in Antarctica using morphometrics. Colonial Waterbirds, 21, 444-449. 740
Rivera-Milán, F. F. 1997. Seasonal and annual changes in the population density of Zenaida 741
doves in the xerophytic forest of Guanica, Puerto Rico. Journal of Field Ornithology, 742
68, 259-272. 743
Rivera-Milán, F. F. 1999. Population dynamics of Zenaida doves in Cidra, Puerto Rico. 744
Journal of Wildlife Management, 63, 232-244. 745
Rotenberry, J. T. & Wiens, J. A. 1985. Statistical power analysis and community-wide 746
patterns. The American Naturalist, 125, 164-168. 747
Rubega, M. A. 1996. Sexual size dimorphism in Red-necked phalaropes and functional 748
signifiance of non sexual bill structure variation for feeding performance. Journal of 749
Morphology, 228, 45-60. 750
Ryder, J. P. 1978. Sexing Ring-billed gulls externally. Bird Banding, 49, 218-222. 751
Santiago-Alarcon, D. & Parker, P. G. 2007. Sexual size dimorphism and morphological 752
evidence supporting the recognition of two subspecies in the Galápagos dove. The 753
Condor, 109, 132-141. 754
Schroeder, J., Lourenco, P. M., van der Velde, M., Hooijmeijer, J., Both, C. & Piersma, T. 755
2008. Sexual dimorphism in plumage and size in Black-tailed godwits Limosa limosa 756
limosa. Ardea, 96, 25-37. 757
Señar, J. C. 1999. La medición de la repetibilidad y el error de medida. Etologuia, 17, 53-64. 758
Seutin, G., White, B. N. & Boag, P. T. 1991. Preservation of avian blood tissue samples for 759
DNA analyses. Canadian Journal of Zoology, 69, 82-90. 760
Shamis, J. D. & Forrester, D. J. 1977. Haematozoan parasites of Mourning doves in Florida. 761
Journal of Wildlife Diseases, 13, 349-355. 762

Shealer, D. A. & Cleary, C. M. 2007. Sex determination of adult black terns by DNA and 763
morphometrics: tests of sample size, temporal stability and geographic specificity in 764
the classification accuracy of discriminant functions models. Waterbirds, 30, 180-188. 765
Shugart, G. W. 1977. A method for externally sexing gulls. Bird Banding, 48, 118-121. 766
Small, M. F., Baccus, J. T., Mink, J. N. & Roberson, J. A. 2005. Hematologic responses in 767
captive White-winged doves (Zenaida asiatica) induced by various radiotransmitter 768
attachments. Journal of Wildlife Diseases, 41, 387-394. 769
Snyder, P. 1991. Three reasons why stepwise regression methods should not be used by 770
researchers. In: Advances in educational research: Substantive findings, 771
methodological developments (Ed. by Thompson, B.), pp. 99-105. Greenwich, USA: 772
JAI. 773
Sol, D., Elie, M., Marcoux, M., Chrostovsky, E., Porcher, C. & Lefebvre, L. 2005. Ecological 774
mechanisms of resource polymorphism in zenaida doves of Barbados. Ecology, 86, 775
2397-2407. 776
Stevens, J. 1992. Applied Multivariate Statistics for Social Sciences. 2nd ed. Mahwah, USA: 777
Lawrance Erlbaum Associates Publishers. 778
Suthers, H. B. & Suthers, D. D. 1990. Aging and sexing Gray catbirds by external 779
characteristics. North American Bird Bander, 15, 45-52. 780
Swanson, D. A. & Rappole, J. H. 1992. Determining the sex of adult White-winged doves by 781
cloacal characteristics. North American Bird Bander, 17, 137-139. 782
Tabachnick, B. G. & Fidell, L. S. 1996. Using Multivariate Statistics (3rd ed.). New York, 783
USA: Harper Collins. 784
Thompson, B. 1995. Stepwise regression and stepwise discriminant analysis need not apply 785
here: a guidelines editorial. Educational and Psychological Measurements 55, 525-786
534. 787

Thompson, B. 2001. Significance, effect sizes, stepwise methods, and other issues: Strong 788
arguments move the field. The Journal of Experimental Education, 70, 80-93. 789
Toft, C. A. & Shea, P. J. 1983. Detecting community-wide patterns: estimating power 790
strengthens statistical inference. The American Naturalist, 122, 618-625. 791
Ura, T., Azuma, N., Hayama, S. & Higashi, S. 2005. Sexual dimorphism of Latham's snipe 792
(Gallinago hardwickii). Emu, 105, 259-262. 793
Urfi, A. J. & Kalam, A. 2006. Sexual size dimorphism and mating pattern in the Painted stork 794
(Mycteria leucocephala). Waterbirds, 29, 489-496. 795
Venables, W. N. & Ripley, B. D. 2002. Modern Applied Statistics with S. Fourth edition. 796
Berlin, Germany: Springer. 797
Wehberg, S. & Schumacher, M. 2004. A comparison of nonparametric error rate estimation in 798
classifications problems. Biometrical Journal, 46, 35-47. 799
Wiley, J. W. 1991. Ecology and behavior of the Zenaida dove. Ornitología Neotropical, 2, 800
49-75. 801
Williams, B. K. & Titus, K. 1988. Assessment of sampling stability in ecological applications 802
of discriminant analysis. Ecology, 69, 275-285. 803
Yezerinac, S. M., Lougheed, S. C. & Handford, P. 1992. Measurement error and 804
morphometric studies: statistical power and observer experience. Systematic Zoology, 805
41, 471-482. 806
Zavalaga, C. B. & Paredes, R. 1997. Sex determination in Humboldt penguins using 807
morphometric characters. Journal of Field Ornithology, 68, 102-112. 808
809
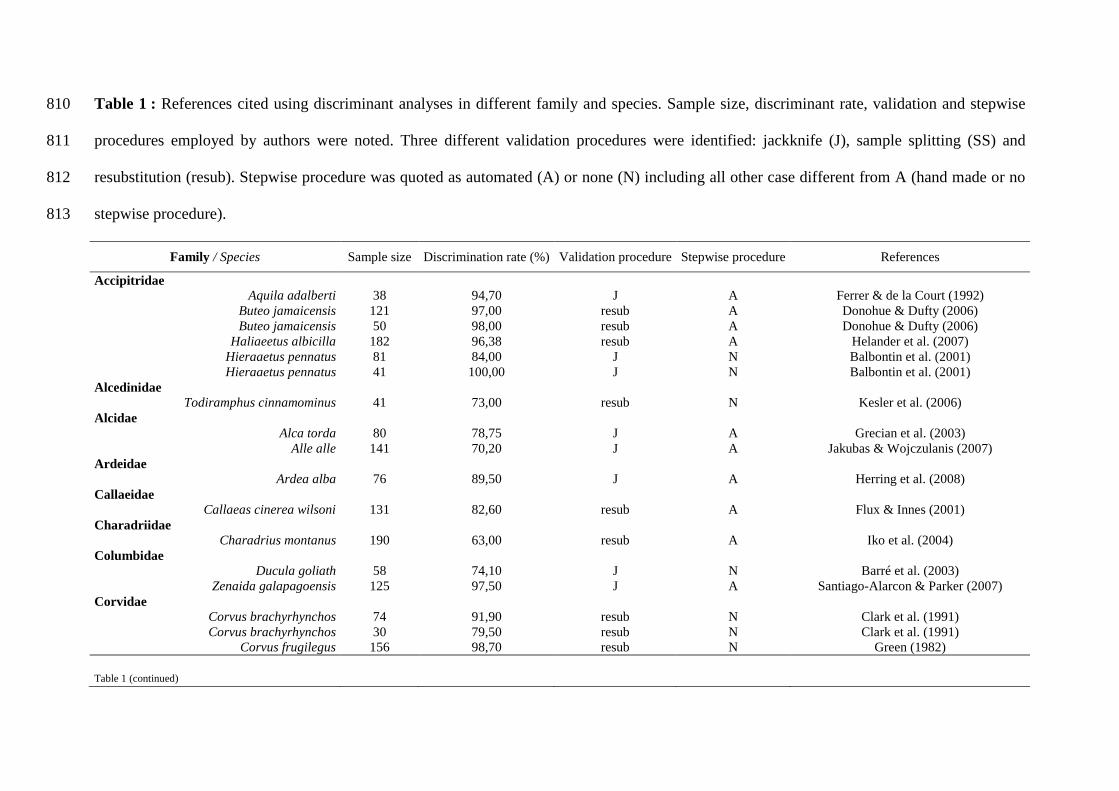
Table 1 : References cited using discriminant analyses in different family and species. Sample size, discriminant rate, validation and stepwise 810
procedures employed by authors were noted. Three different validation procedures were identified: jackknife (J), sample splitting (SS) and 811
resubstitution (resub). Stepwise procedure was quoted as automated (A) or none (N) including all other case different from A (hand made or no 812
stepwise procedure). 813
Family / Species Sample size Discrimination rate (%) Validation procedure Stepwise procedure References
Accipitridae Aquila adalberti 38 94,70 J A Ferrer & de la Court (1992)
Buteo jamaicensis 121 97,00 resub A Donohue & Dufty (2006) Buteo jamaicensis 50 98,00 resub A Donohue & Dufty (2006)
Haliaeetus albicilla 182 96,38 resub A Helander et al. (2007) Hieraaetus pennatus 81 84,00 J N Balbontin et al. (2001) Hieraaetus pennatus 41 100,00 J N Balbontin et al. (2001)
Alcedinidae Todiramphus cinnamominus 41 73,00 resub N Kesler et al. (2006)
Alcidae Alca torda 80 78,75 J A Grecian et al. (2003)
Alle alle 141 70,20 J A Jakubas & Wojczulanis (2007) Ardeidae
Ardea alba 76 89,50 J A Herring et al. (2008) Callaeidae
Callaeas cinerea wilsoni 131 82,60 resub A Flux & Innes (2001) Charadriidae
Charadrius montanus 190 63,00 resub A Iko et al. (2004) Columbidae
Ducula goliath 58 74,10 J N Barré et al. (2003) Zenaida galapagoensis 125 97,50 J A Santiago-Alarcon & Parker (2007)
Corvidae Corvus brachyrhynchos 74 91,90 resub N Clark et al. (1991) Corvus brachyrhynchos 30 79,50 resub N Clark et al. (1991)
Corvus frugilegus 156 98,70 resub N Green (1982) Table 1 (continued)
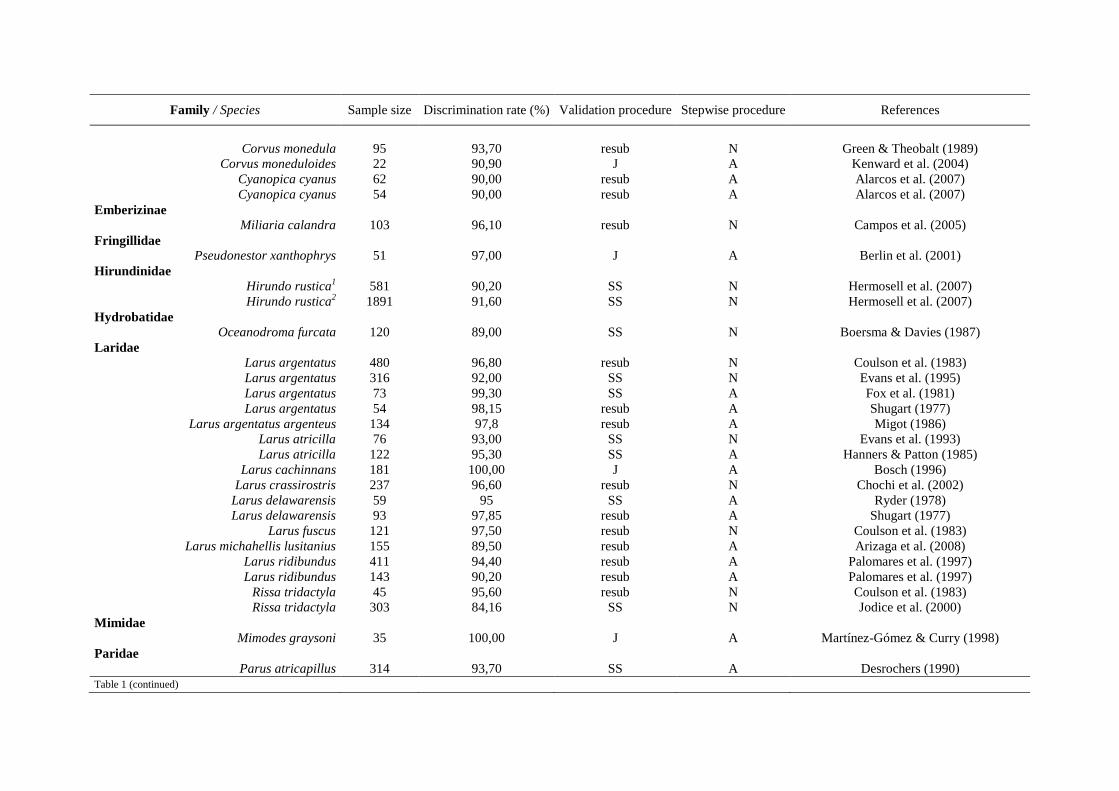
Family / Species Sample size Discrimination rate (%) Validation procedure Stepwise procedure References
Corvus monedula 95 93,70 resub N Green & Theobalt (1989)
Corvus moneduloides 22 90,90 J A Kenward et al. (2004) Cyanopica cyanus 62 90,00 resub A Alarcos et al. (2007) Cyanopica cyanus 54 90,00 resub A Alarcos et al. (2007)
Emberizinae Miliaria calandra 103 96,10 resub N Campos et al. (2005)
Fringillidae Pseudonestor xanthophrys 51 97,00 J A Berlin et al. (2001)
Hirundinidae Hirundo rustica1 581 90,20 SS N Hermosell et al. (2007) Hirundo rustica2 1891 91,60 SS N Hermosell et al. (2007)
Hydrobatidae Oceanodroma furcata 120 89,00 SS N Boersma & Davies (1987)
Laridae Larus argentatus 480 96,80 resub N Coulson et al. (1983) Larus argentatus 316 92,00 SS N Evans et al. (1995) Larus argentatus 73 99,30 SS A Fox et al. (1981) Larus argentatus 54 98,15 resub A Shugart (1977)
Larus argentatus argenteus 134 97,8 resub A Migot (1986) Larus atricilla 76 93,00 SS N Evans et al. (1993) Larus atricilla 122 95,30 SS A Hanners & Patton (1985)
Larus cachinnans 181 100,00 J A Bosch (1996) Larus crassirostris 237 96,60 resub N Chochi et al. (2002) Larus delawarensis 59 95 SS A Ryder (1978) Larus delawarensis 93 97,85 resub A Shugart (1977)
Larus fuscus 121 97,50 resub N Coulson et al. (1983) Larus michahellis lusitanius 155 89,50 resub A Arizaga et al. (2008)
Larus ridibundus 411 94,40 resub A Palomares et al. (1997) Larus ridibundus 143 90,20 resub A Palomares et al. (1997)
Rissa tridactyla 45 95,60 resub N Coulson et al. (1983) Rissa tridactyla 303 84,16 SS N Jodice et al. (2000)
Mimidae Mimodes graysoni 35 100,00 J A Martínez-Gómez & Curry (1998)
Paridae Parus atricapillus 314 93,70 SS A Desrochers (1990)
Table 1 (continued)
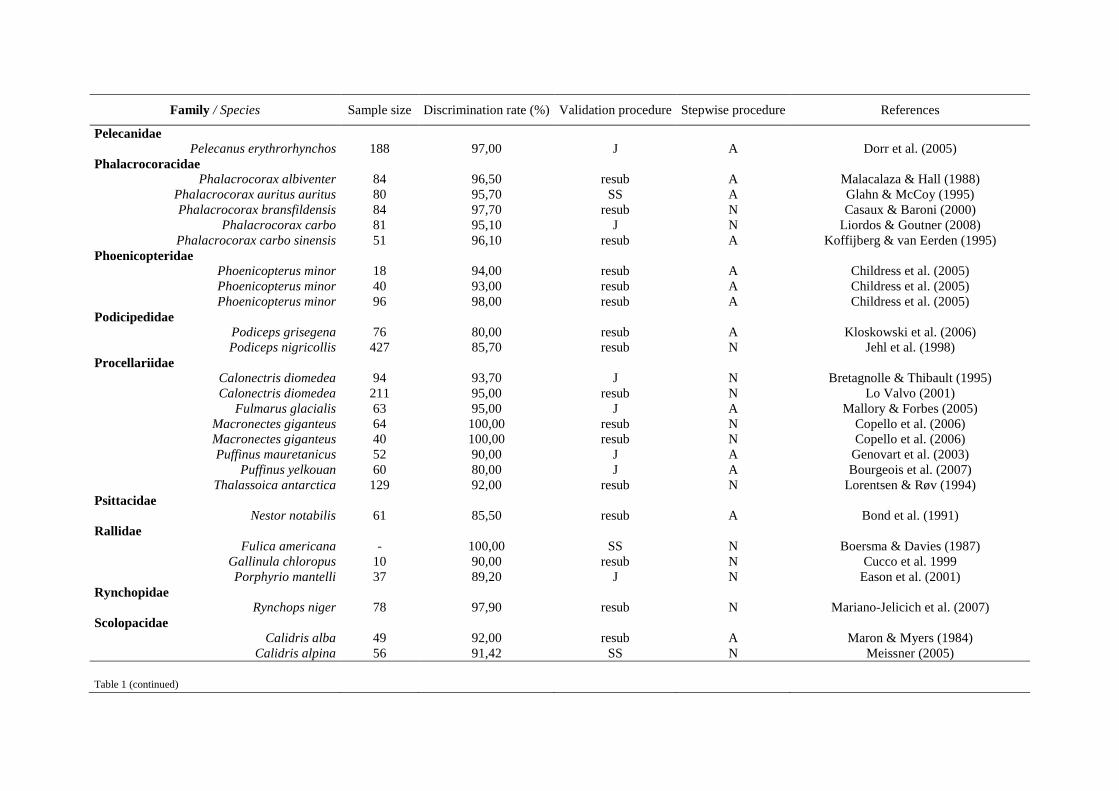
Family / Species Sample size Discrimination rate (%) Validation procedure Stepwise procedure References
Pelecanidae Pelecanus erythrorhynchos 188 97,00 J A Dorr et al. (2005)
Phalacrocoracidae Phalacrocorax albiventer 84 96,50 resub A Malacalaza & Hall (1988)
Phalacrocorax auritus auritus 80 95,70 SS A Glahn & McCoy (1995) Phalacrocorax bransfildensis 84 97,70 resub N Casaux & Baroni (2000)
Phalacrocorax carbo 81 95,10 J N Liordos & Goutner (2008) Phalacrocorax carbo sinensis 51 96,10 resub A Koffijberg & van Eerden (1995)
Phoenicopteridae Phoenicopterus minor 18 94,00 resub A Childress et al. (2005) Phoenicopterus minor 40 93,00 resub A Childress et al. (2005) Phoenicopterus minor 96 98,00 resub A Childress et al. (2005)
Podicipedidae Podiceps grisegena 76 80,00 resub A Kloskowski et al. (2006) Podiceps nigricollis 427 85,70 resub N Jehl et al. (1998)
Procellariidae Calonectris diomedea 94 93,70 J N Bretagnolle & Thibault (1995) Calonectris diomedea 211 95,00 resub N Lo Valvo (2001)
Fulmarus glacialis 63 95,00 J A Mallory & Forbes (2005) Macronectes giganteus 64 100,00 resub N Copello et al. (2006) Macronectes giganteus 40 100,00 resub N Copello et al. (2006) Puffinus mauretanicus 52 90,00 J A Genovart et al. (2003)
Puffinus yelkouan 60 80,00 J A Bourgeois et al. (2007) Thalassoica antarctica 129 92,00 resub N Lorentsen & Røv (1994)
Psittacidae Nestor notabilis 61 85,50 resub A Bond et al. (1991)
Rallidae Fulica americana - 100,00 SS N Boersma & Davies (1987)
Gallinula chloropus 10 90,00 resub N Cucco et al. 1999 Porphyrio mantelli 37 89,20 J N Eason et al. (2001)
Rynchopidae Rynchops niger 78 97,90 resub N Mariano-Jelicich et al. (2007)
Scolopacidae Calidris alba 49 92,00 resub A Maron & Myers (1984)
Calidris alpina 56 91,42 SS N Meissner (2005)
Table 1 (continued)
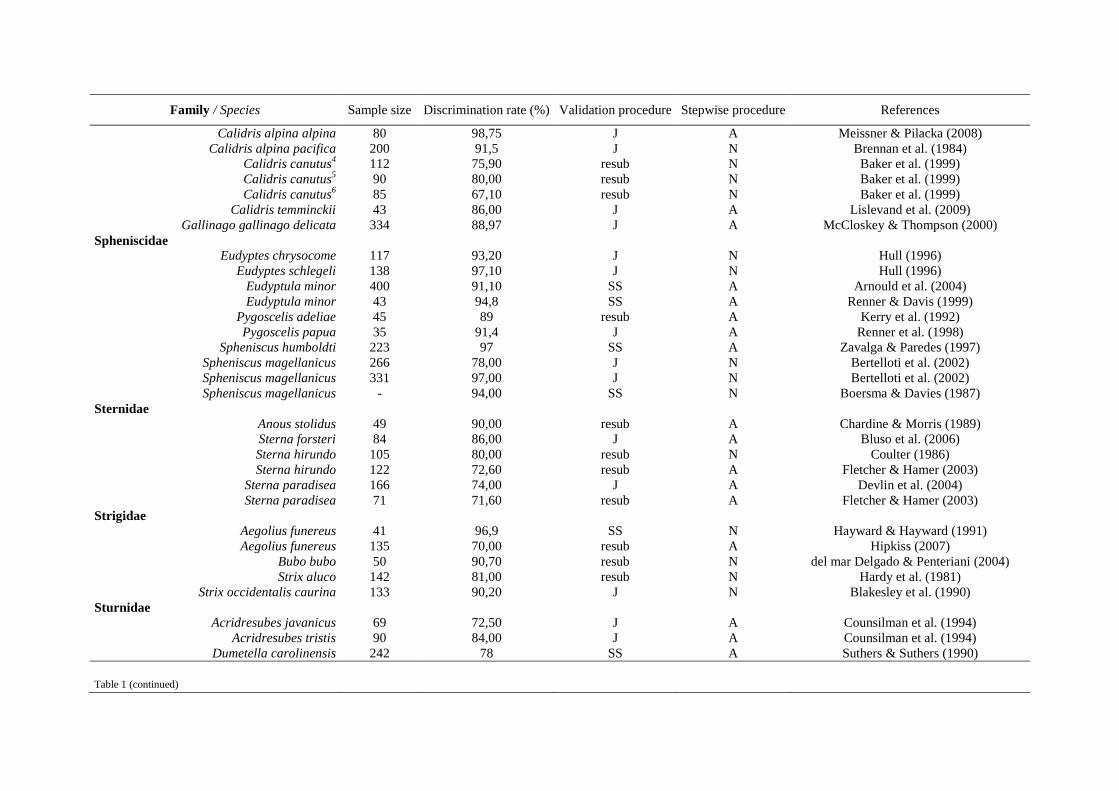
Family / Species Sample size Discrimination rate (%) Validation procedure Stepwise procedure References
Calidris alpina alpina 80 98,75 J A Meissner & Pilacka (2008) Calidris alpina pacifica 200 91,5 J N Brennan et al. (1984)
Calidris canutus4 112 75,90 resub N Baker et al. (1999) Calidris canutus5 90 80,00 resub N Baker et al. (1999) Calidris canutus6 85 67,10 resub N Baker et al. (1999)
Calidris temminckii 43 86,00 J A Lislevand et al. (2009) Gallinago gallinago delicata 334 88,97 J A McCloskey & Thompson (2000)
Spheniscidae Eudyptes chrysocome 117 93,20 J N Hull (1996)
Eudyptes schlegeli 138 97,10 J N Hull (1996) Eudyptula minor 400 91,10 SS A Arnould et al. (2004) Eudyptula minor 43 94,8 SS A Renner & Davis (1999)
Pygoscelis adeliae 45 89 resub A Kerry et al. (1992) Pygoscelis papua 35 91,4 J A Renner et al. (1998)
Spheniscus humboldti 223 97 SS A Zavalga & Paredes (1997) Spheniscus magellanicus 266 78,00 J N Bertelloti et al. (2002) Spheniscus magellanicus 331 97,00 J N Bertelloti et al. (2002) Spheniscus magellanicus - 94,00 SS N Boersma & Davies (1987)
Sternidae Anous stolidus 49 90,00 resub A Chardine & Morris (1989) Sterna forsteri 84 86,00 J A Bluso et al. (2006) Sterna hirundo 105 80,00 resub N Coulter (1986) Sterna hirundo 122 72,60 resub A Fletcher & Hamer (2003)
Sterna paradisea 166 74,00 J A Devlin et al. (2004) Sterna paradisea 71 71,60 resub A Fletcher & Hamer (2003)
Strigidae Aegolius funereus 41 96,9 SS N Hayward & Hayward (1991) Aegolius funereus 135 70,00 resub A Hipkiss (2007)
Bubo bubo 50 90,70 resub N del mar Delgado & Penteriani (2004) Strix aluco 142 81,00 resub N Hardy et al. (1981)
Strix occidentalis caurina 133 90,20 J N Blakesley et al. (1990) Sturnidae
Acridresubes javanicus 69 72,50 J A Counsilman et al. (1994) Acridresubes tristis 90 84,00 J A Counsilman et al. (1994)
Dumetella carolinensis 242 78 SS A Suthers & Suthers (1990) Table 1 (continued)
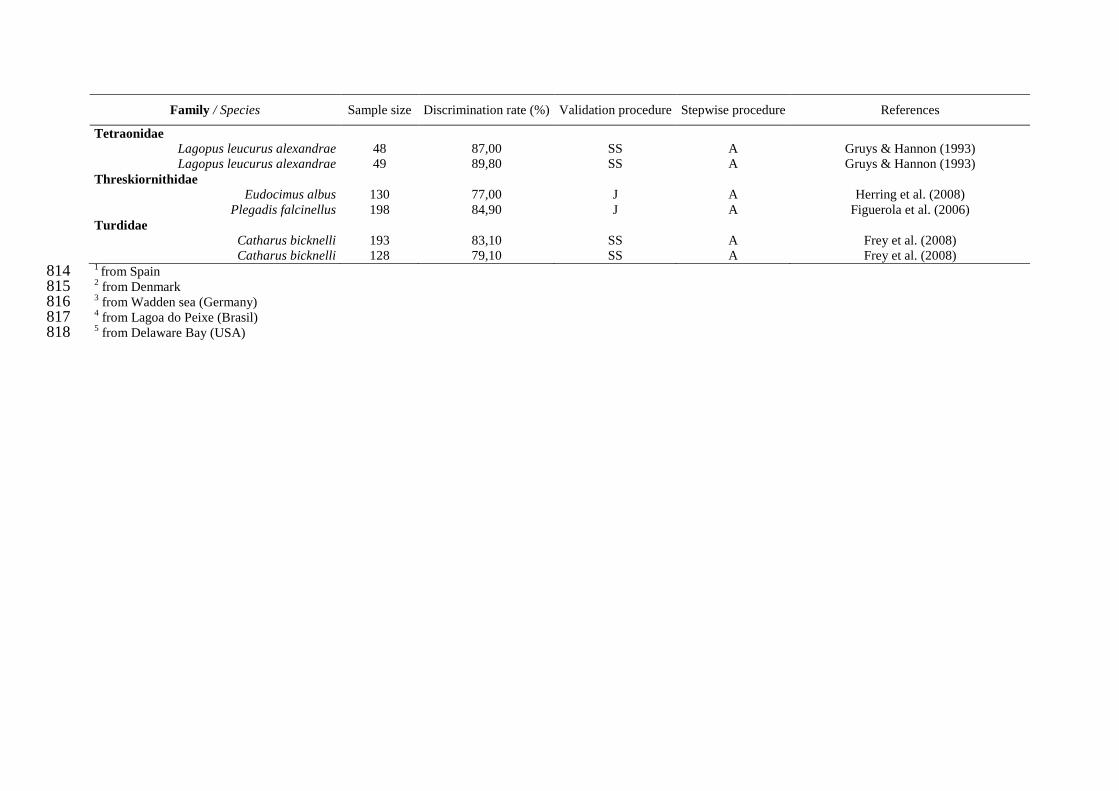
Family / Species Sample size Discrimination rate (%) Validation procedure Stepwise procedure References
Tetraonidae Lagopus leucurus alexandrae 48 87,00 SS A Gruys & Hannon (1993) Lagopus leucurus alexandrae 49 89,80 SS A Gruys & Hannon (1993)
Threskiornithidae Eudocimus albus 130 77,00 J A Herring et al. (2008)
Plegadis falcinellus 198 84,90 J A Figuerola et al. (2006) Turdidae
Catharus bicknelli 193 83,10 SS A Frey et al. (2008) Catharus bicknelli 128 79,10 SS A Frey et al. (2008)
1 from Spain 814 2 from Denmark 815 3 from Wadden sea (Germany) 816 4 from Lagoa do Peixe (Brasil) 817 5 from Delaware Bay (USA) 818
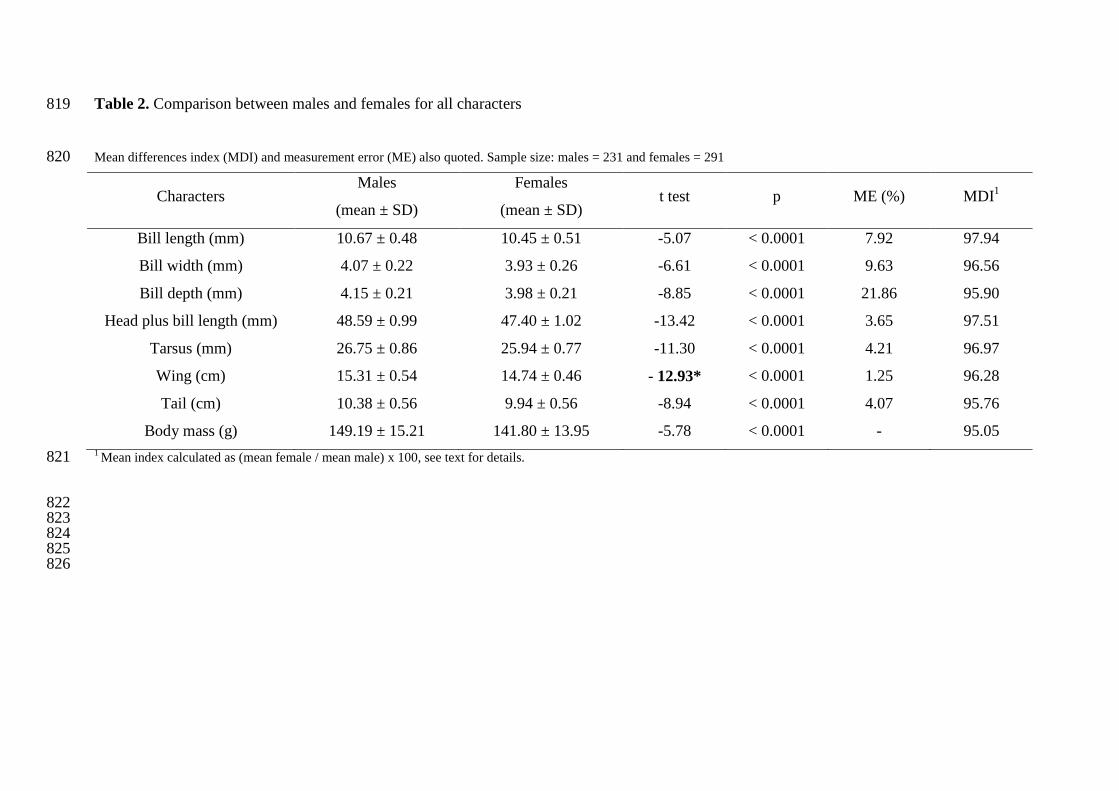
Table 2. Comparison between males and females for all characters 819
Mean differences index (MDI) and measurement error (ME) also quoted. Sample size: males = 231 and females = 291 820
Characters Males
(mean ± SD)
Females
(mean ± SD) t test p ME (%) MDI1
Bill length (mm) 10.67 ± 0.48 10.45 ± 0.51 -5.07 < 0.0001 7.92 97.94
Bill width (mm) 4.07 ± 0.22 3.93 ± 0.26 -6.61 < 0.0001 9.63 96.56
Bill depth (mm) 4.15 ± 0.21 3.98 ± 0.21 -8.85 < 0.0001 21.86 95.90
Head plus bill length (mm) 48.59 ± 0.99 47.40 ± 1.02 -13.42 < 0.0001 3.65 97.51
Tarsus (mm) 26.75 ± 0.86 25.94 ± 0.77 -11.30 < 0.0001 4.21 96.97
Wing (cm) 15.31 ± 0.54 14.74 ± 0.46 - 12.93* < 0.0001 1.25 96.28
Tail (cm) 10.38 ± 0.56 9.94 ± 0.56 -8.94 < 0.0001 4.07 95.76
Body mass (g) 149.19 ± 15.21 141.80 ± 13.95 -5.78 < 0.0001 - 95.05 1 Mean index calculated as (mean female / mean male) x 100, see text for details. 821
822 823 824 825 826

Caption for figures 827
828
Figure 1. Simulation of the effect of the sample size on the estimated proportion of correctly 829
classified individuals (discriminant rate). From the complete data set (n = 525 individuals), 830
smaller sub-samples were randomly selected (size ranging from 25 to 520 individuals, with 831
500 sub-samples per size). For each of these 50000 sub-samples, we performed a DFA and 832
we evaluated the discriminant rate according to three methods: (a) resubstitution, (b) jack-833
knife cross-validation, (c) sample splitting in which two third of the data set were used as 834
training sample and the remaining third as test sample. Each grey dot represents one analysis. 835
Each dot has been slightly randomly jittered to reduce overplotting. The thick line represents 836
the mean discriminant rate computed from the 500 DFA validations performed for each 837
sample size. The horizontal dotted line represents the discriminant rate estimated from the 838
complete data set. 839
840
Figure 2. Simulation of the effect of measurement errors on the jack-knifed estimate for 841
proportion of correctly classified individuals (discriminant rate). The complete data set 842
(n = 525 individuals) was used. We added to every parameter a random noise normally 843
distributed with standard deviation σ’ = kσ, proportional to the standard deviation σ of the 844
corresponding parameter. A noise coefficient k = 0 means that no noise has been added to the 845
original data. For each value of k, we simulated 500 data sets. Each grey dot represents one 846
DFA performed on one simulated data set. The thick line represents the mean discriminant 847
rate computed from the 500 DFA performed for every given value of k. 848
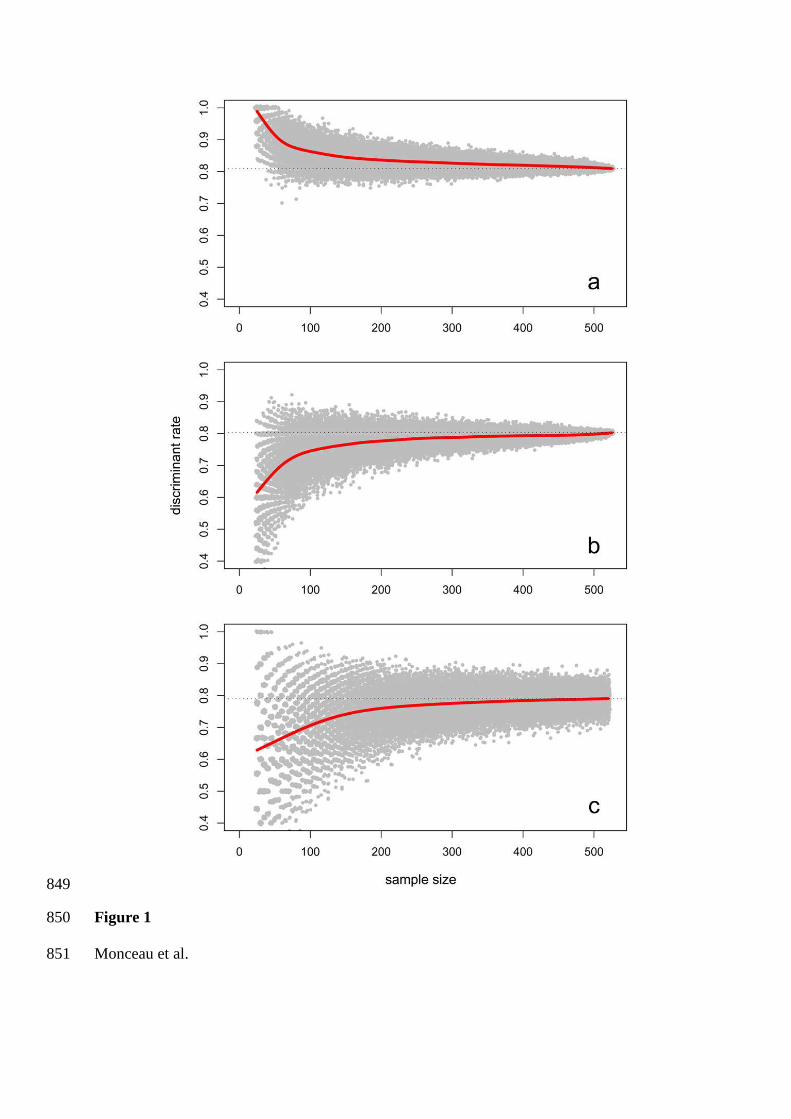
849
Figure 1 850
Monceau et al. 851
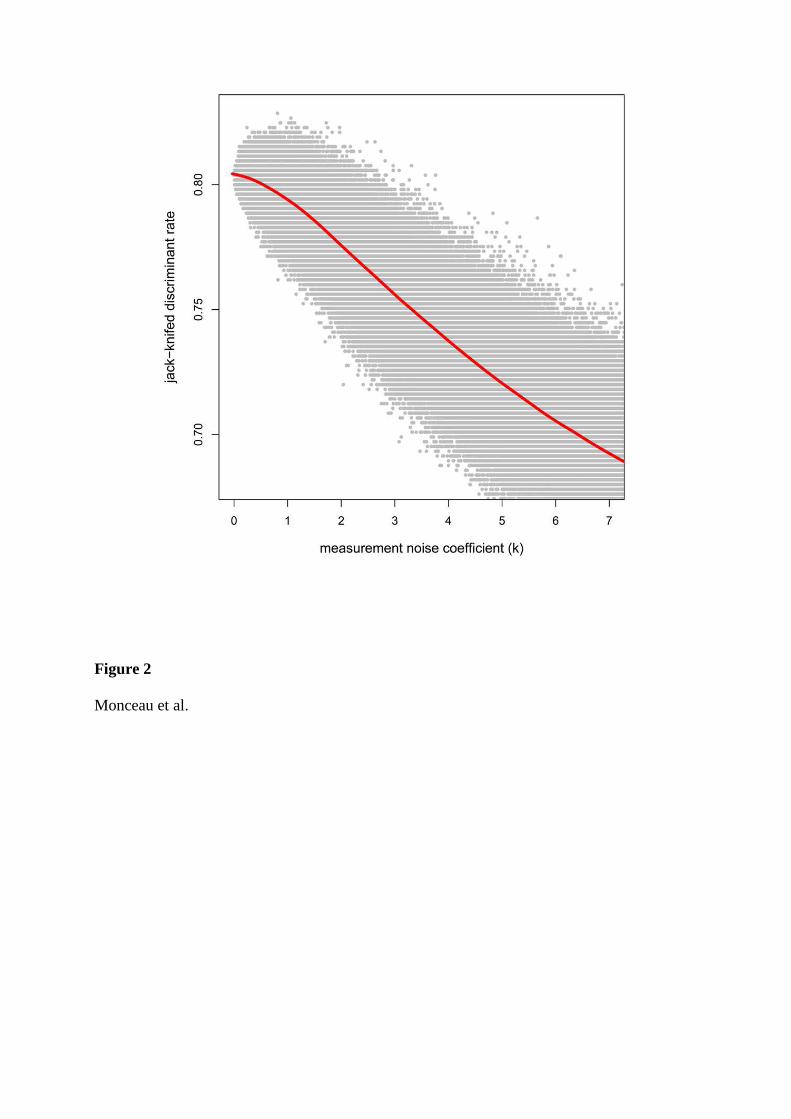
Figure 2
Monceau et al.

ANNEXE 2
Monceau, K., Gaillard, M., Harrang, E., Santiago-Alarcon, D., Parker, P., Cézilly, F. &
Wattier, R. 2009. Twenty-three polymorphic microsatellite markers for the Caribbean
endemic Zenaida dove, Zenaida aurita, and its conservation in related Zenaida species.
Conservation Genetics, 10, 1577-1581.

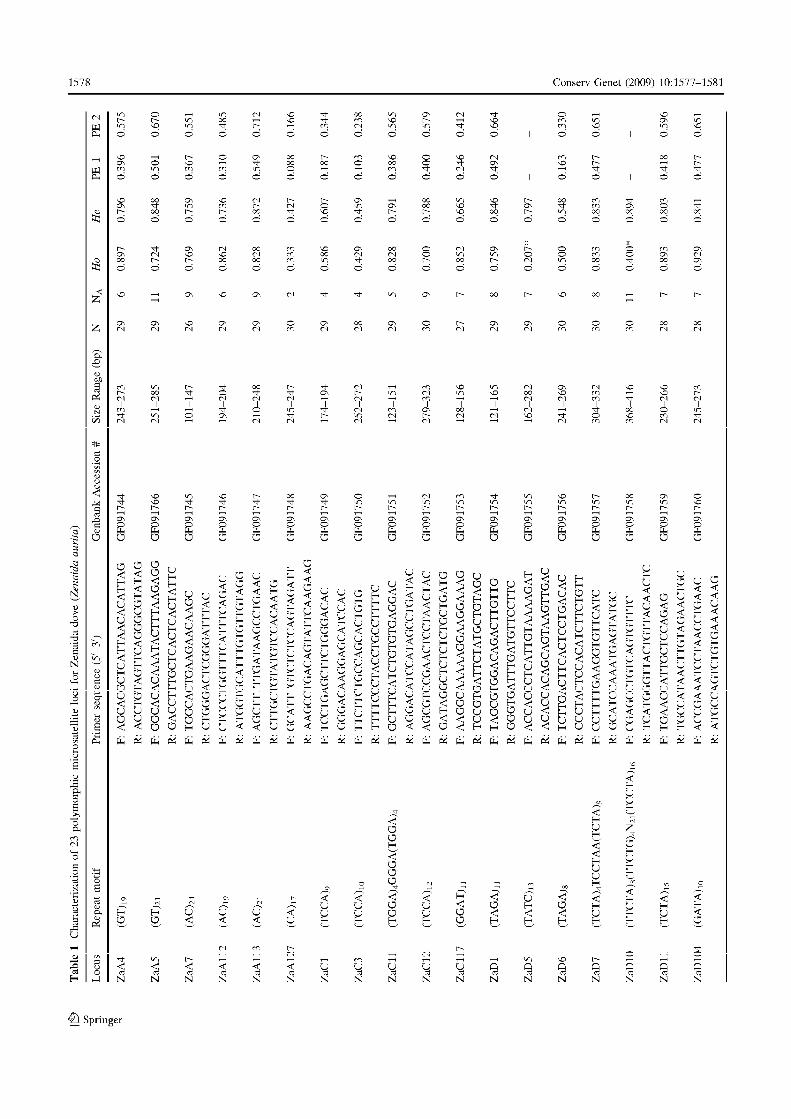
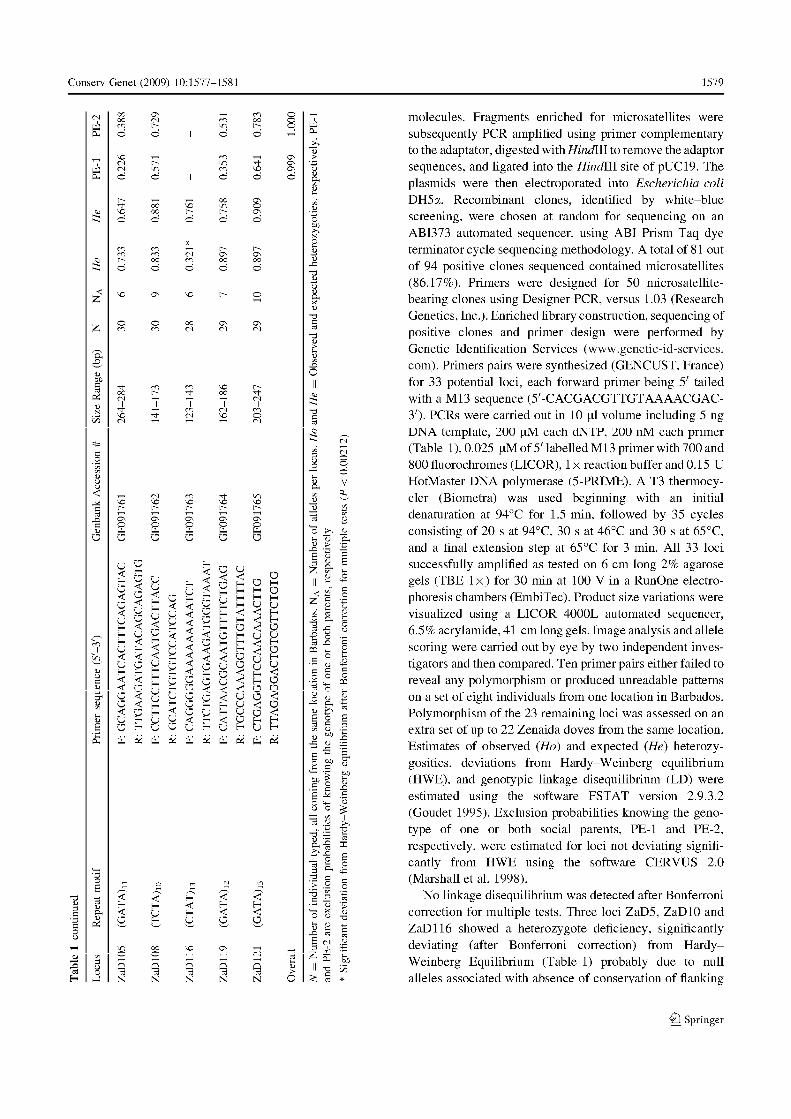
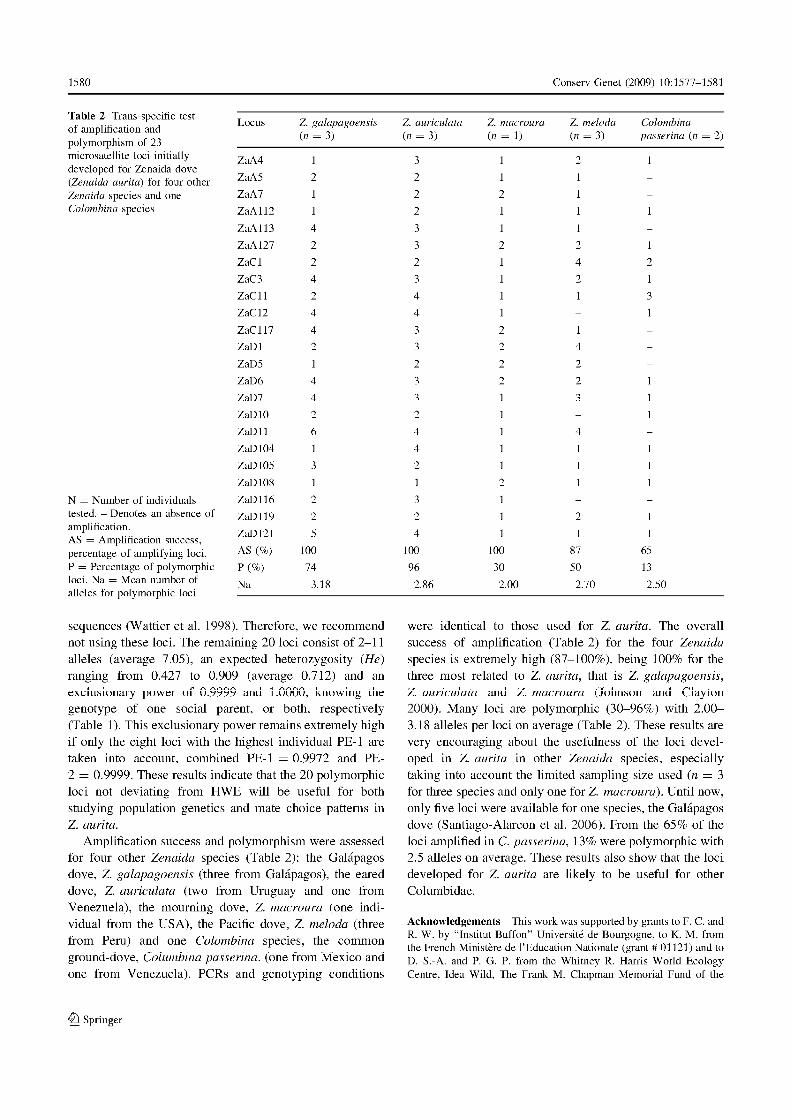


ANNEXE 3
Monceau, K., Dechaume-Moncharmont, F.-X., Wattier, R., Motreuil, S. & Cézilly, F.
Territoriality vs. flocking in the Zenaida dove, Zenaida aurita: Resource polymorphism
revisited using morphological and genetic analyses.
Manuscrit à soumettre à Ecology

Monceau et al. 40 pages
Resource use in Zenaida doves 1
Territoriality vs. flocking in the Zenaida dove, Zenaida aurita: 2
Resource polymorphism revisited using morphological and genetic analyses 3
4
KARINE MONCEAU, FRANÇOIS-XAVIER DECHAUME-MONCHARMONT, REMI WATTIER, 5
SEBASTIEN MOTREUIL, AND FRANK CEZILLY 6
7
Université de Bourgogne, 8
Equipe Ecologie Evolutive, UMR CNRS 5561 Biogéosciences. 9
10
11
Abstract. Evidence for resource polymorphism, i.e. inter- or intra-specific 12
differentiation in phenotypic characters in relation to resource use, has accumulated in fish, and 13
to a lesser extent, in amphibians, but remains scarce in birds. One noticeable exception, 14
however, is the case of Zenaida doves (Zenaida aurita) in Barbados. There, doves either 15
aggressively defend year-round territories or feed in flocks at grain-storage sites (where large 16
amounts of corn spillage are available), with little, if any, aggression. Sol et al. (2005; Ecology 17
86: 2397-2407) concluded that the existence of alternative foraging tactics in Zenaida doves 18
was a case of resource polymorphism, primarily driven by competition for territories, forcing 19
less competitive individuals to use suboptimal resources. We repeated and expanded the study 20
by Sol et al. (2005) through adding replications in space and time for each type of foraging 21
tactic. Compared to the previous study, the use of molecular makers allowed unambiguous 22
sexing of both adults and juveniles. We also used microsatellite markers to assess potential 23
genetic differentiation between flock-feeding and territorial doves. Our results confirm that, 24
irrespective of sex, territorial adults are larger than adults feeding in flocks, whereas the reverse 25
was observed in juveniles. Contrary to previous evidence, however, we found a consistent and 26

Monceau et al. 40 pages
significant excess of females among flock-feeding adults, whereas the sex ratio did not differ 27
from unity among territorial birds, and among both territorial and flock-feeding juveniles. In 28
addition, body condition of birds did not differ between birds feeding in flocks and birds 29
feeding on territories. Finally we found no evidence for genetic differentiation between birds 30
feeding in flocks and birds feeding on territories. We argue that resorting to the concept of 31
'resource polymorphism' might be inadequate to characterize the existence of alternative 32
foraging tactics in Zenaida doves in Barbados. Instead, we propose that the phenomenon is 33
better explained by alternative resource use during ontogeny and age and sex-related 34
differences in the benefits associated with holding a territory. The present study illustrates the 35
importance of metareplication in population biology and ecology. 36
Key words: alternative resource use, competition, genetic differentiation, 37
metareplication, morphometrics 38
39
INTRODUCTION 40
Understanding how variation in resource levels influences morphological diversity and 41
divergent natural selection is of primary interest in ecological and evolutionary research, as it 42
may ultimately result in speciation events (Schluter 2000, Bolnicki and Fitzpatrick 2007). 43
Various mechanisms may link resource use and morphology. In particular, both the invasion of 44
"open" niches in a heterogeneous environment and the relaxation of inter- or intra-specific 45
competition are thought to favour the evolution of resource polymorphism, i.e. inter- or intra-46
specific differentiation in phenotypic characters in relation to resource use (Skúlason and Smith 47
1995, Smith and Skúlason 1996, Pfennig et al. 2007). Although it has been suggested that 48
resource polymorphism may play an important role in population divergence and speciation in 49
vertebrates (Smith and Skúlason 1996), the extent of empirical evidence varies between 50
taxonomic groups. Whereas resource polymorphism has been regularly reported in fish 51
(Ruzzante et al. 2003, Proulx and Magnan 2004, Olsson et al. 2007, Parsons and Robinson 52

Monceau et al. 40 pages
2007, Svanbäck et al. 2008) and, to a lesser extent, in amphibians (Maerz et al. 2006, Pfennig 53
et al. 2007), its importance in birds remains poorly documented. 54
The few cited examples of resource polymorphism in birds involve the existence of 55
discrete morphs, based on bill size or shape (Smith and Temple 1982, Goss-Custard and Durell 56
1983, Smith 1990, Borras et al. 2008), or plumage colour (Rohwer 1990, Roulin 2004). One 57
noticeable exception, however, is the resource polymorphism reported by Lefebvre and 58
collaborators (Dolman et al 1996; Lefebvre 1996; Carlier and Lefebvre 1996, 1997; Sol et al 59
2005) in Zenaida doves (Zenaida aurita) on the island of Barbados (West Indies). Barbados 60
hosts a very limited avifauna and has practically no pristine habitats left due to intense 61
agriculture and development. On the island, Zenaida doves usually establish year-round 62
feeding territories that they defend against conspecifics and, to a lesser extent, other columbid 63
species (Columbina passerina, and more recently, Columba squamosa), while showing more 64
tolerance towards unrelated avian species (Loxigilla noctis, Molothrus bonariensis, Quiscalus 65
lugubris; Dolman et al 1996; F. Cézilly, unpublished data). In contrast, where food availability 66
is particularly high, doves gather in large numbers and feed in flocks with relatively little 67
aggression. This is especially the case at some isolated grain storage facilities on the island, 68
where large amounts of seed spillage attract Zenaida doves and a few other bird species. Based 69
on a comparison of morphological traits between flocking and territorial individuals and 70
additional experiments, Sol et al. (2005) concluded that the existence of alternative foraging 71
tactics in Zenaida doves in Barbados was a case of resource polymorphism driven by 72
competition for territories, where less competitive individuals, irrespective of age and sex, are 73
forced to exploit a sub-optimal resource. However, Sol et al's (2005) study was based on only 74
one territorial and one flock-feeding site, which limits its generality. In addition, their 75
conclusions on male and female differences, as well as the probable lack of genetic 76
differenciation between territorial and flock-feeding populations (see also Dolman et al 1996; 77

Monceau et al. 40 pages
Carlier and Lefebvre 1996, 1997), were based on observational and morphological data, not the 78
genetic analyses that would be much more appropriate. 79
The present study re-examines the evidence for resource polymorphism in Zenaida 80
doves in Barbados, as part of a long-term study on the behavioral ecology and population 81
biology of the species. In this paper, we expand on the earlier study of Sol et al. (2005) by 82
adding replications in space and time for each type of foraging tactic. In addition, we use 83
molecular makers to unambiguously sex all individuals of the four sites we studied, adults and 84
juveniles alike. Because resource polymorphism is expected to affect gene flow (Skúlason and 85
Smith 1995), we also measured the extent of genetic differentiation between territorial and 86
flock-feeding Zenaida doves using recently developed microsatellite markers (Monceau et al. 87
2009). Recent evidence suggests that rapid niche expansion and resource polymorphism can be 88
associated with genetic divergence at very small spatial scales in vertebrates (McCormack and 89
Smith 2008, Bergek and Björklund 2009), even in the presence of gene flow (Señar et al. 90
2006). 91
92
METHODS 93
ZENAIDA DOVES IN BARBADOS 94
95
The Zenaida dove is widely distributed through the Caribbean islands (Raffaele et al. 96
1998). Although its primary habitat consists of wooldands and scrub thickets (Wiley 1991, 97
Rivera-Milán 1997, 1999), on several islands, including Barbados, the species has largely 98
colonized open coastal areas and gardens. In Barbados, Zenaida doves typically feed from 99
seeds on the ground, alone or in pairs (Lefebvre et al. 1996), but may exploit opportunistically 100
alternative food sources, such as leftover bread or cooked rice. 101
Both sexes are similar in appearance, with males being slightly larger than females 102
(Wiley 1991, F. Cézilly, unpubl. data). Juveniles can easily be distinguished from adults 103

Monceau et al. 40 pages
through the absence of iridescent patches on each side of the neck (Sol et al. 2005), the 104
presence of grey-like first feathers, and high-pitched vocalizations. 105
Birds defend their territory against conspecifics by running or flying towards them, and 106
eventually attacking them with wing slaps and pecks. Escalated contests between territory 107
owners and intruders involve ritualized displays such as ground pecking and wing-raising. 108
During the latter, birds typically flick one wing contra-lateral to the opponent, while walking 109
parallel to each other (Goldberg et al. 2001). 110
111
Study area 112
For each foraging tactic, i.e. territorial feeding and flock-feeding, two distinct sites were 113
sampled over two consecutive years (2007 and 2008), thus resulting in replications in space 114
and time. Territorial birds were studied at the Bellairs Research Institute (St. James Parish 115
13°11'30.87"N / 59°38'21.55"W) and the adjacent Folkestone Park (collectively referred to as 116
'Bellairs' hereafter), and in the Sunset Crest area (13°10'55.06"N / 59°38'11.80"W), a 117
residential location consisting in small to medium-sized villas surrounded by gardens and parks 118
(referred to as 'Sunset Crest' hereafter). The distance between the two sites is about 1500 m. 119
Flock-feeding birds were studied at the Barbados Mills compound Deep Water Harbour (St 120
Michael Parish, 13° 6'38.99"N / 59°37'46.06"O, designated as Harbour hereafter), and at 121
Roberts Manufacturing Compagny Ltd. (Lower Estate, St. Michael Parish, 13° 7'52.46"N / 122
59°34'48.66"O; referred to as 'Roberts' hereafter). Both sites include large facilities for storage 123
of animal feed and grain. The territorial sites are about 8 km north from Harbour and northwest 124
from Roberts, whereas the distance between the latter two sites is about 5 km (Fig. 1). Our 125
study thus includes the two sites studied by Sol et al 2005, Bellairs and Deep Water Harbour, 126
as well as a replicate of each. Contrary to Sol et al (2005), we did not distinguish between 127
territory holders and floaters at the Bellairs and Sunset Crest sites. 128
129

Monceau et al. 40 pages
Captures 130
All captures were made from March to May in both 2007 and 2008, for a total of 857 131
birds, all ages confounded (586 in 2007 and 271 in 2008). Zenaida doves were caught using 132
walk-in baited drop traps (as used by Sol et al. 2005) and single-catch closing net bird traps. 133
Birds were banded with a unique combination of color plastic bands (A.C. Hughes Ltd., 134
Hampton Hill, United Kingdom) and one numbered aluminium ring from the Muséum 135
National d’Histoire Naturelle de Paris. Each banded bird was then measured (see below), 136
weighed to the nearest 0.1g (using a PESOLA® digital pocket scale MS 500), and blood 137
sampled (see below). All birds were released where they had been caught. 138
139
140
MORPHOLOGICAL DATA 141
142
Repeatability and measurement error 143
The following morphological characters were measured: bill length, depth and width at 144
nostrils, tarsus length (left and right), wing length (left and right). All measurements were 145
made using a digital calliper (accuracy: ± 0.2 mm), except for wing length which was 146
measured with a ruler (accuracy: ± 1 mm). All measurements were taken twice by the same 147
person. Repeatability (R) and measurement error (ME) were later assessed using ANOVA II 148
(Lessells and Boag 1987, Bailey and Byrnes 1990, Yezerinac et al. 1992, Sokal and Rohlf 149
1995, Arnqvist and Mårtensson 1998, Señar 1999, Zar 1999). Since repeatability was 150
significant and measurement error was moderate for both adults (R range: 0.79 – 0.98 and ME 151
range: 2 – 11 %, except bill depth 20%) and juveniles (R range: 0.76 – 0.99 and ME range: 152
0.48 – 5.5 %, except bill depth 24%), the means of double measurements were used in the 153
analyses. In addition, right and left measurements were significantly correlated for both tarsus 154
length (Pearson’s product-moment correlation, adults: r = 0.85, n = 783, P < 0.0001, juveniles: 155

Monceau et al. 40 pages
r = 0.93, n = 74, P < 0.0001) and wing length (Spearman’s correlation coefficient, adults: rs = 156
0.91, n = 783, P < 0.0001 and juveniles: rs = 0.92, n = 74, P < 0.0001). Therefore, the means of 157
the right and left measurements were used for analysis. 158
159
Morphological specialization for feeding 160
Following Sol et al. (2005), we assessed the extent of morphological specialization for 161
feeding from differences in bill shape between groups. A bill morphology index (BMI) was 162
computed from a principal components analysis (PCA) on standardized values of bill 163
measurements (bill length, depth and width), using wing length as control for body size. All 164
variables were log-transformed and standardized (Venables and Ripley 1999). Correlations 165
between the first contribution and components loading were then tested. As homoscedasticity 166
(Levene’s test) was verified in both adults and juveniles, the influence of sex, year, foraging 167
tactic (i.e. territorial vs. flock feeding), and site (nested in foraging tactic) on BMI was assessed 168
using a nested ANOVA I, coupled with a stepwise backward procedure based on likelihood 169
ratios comparison in order to retain only the main significant effects. Then, Tukey Honestly 170
Significant Differences (HSD) test was performed to identify whether groups differed from 171
each other. 172
173
Competitive ability – wing length 174
As wing-flicking appears to be an important component of territorial display (see 175
introduction), competitive ability was assessed from wing length. This character was normally 176
distributed in adults. However variances were not homogeneous, even after transformation, 177
when comparing between the two foraging tactics. A global linear model (GLM) including sex, 178
year, foraging tactic and site was then performed (model assumptions were verified through 179
analysing the distribution of residuals). In juveniles, homoscedasticity was verified, and a 180
nested ANOVA I was then performed with the same parameters than the one used in the 181

Monceau et al. 40 pages
analysis of bill morphology. Both GLM and ANOVA were coupled with stepwise backward 182
procedure and Tukey HSD post hoc analyses. 183
184
Overall body size comparison 185
Following Sol et al. (2005), a body size index (BSI) was computed from the first factor 186
of a PCA procedure including wing length, tarsus length, and bill length (log transformed 187
data). Contrary to Sol et al. (2005), body mass was not included in the PCA, as it should not be 188
considered as an indicator of body size (Alisauskas and Ankney 1990, Piersma and Davidson 189
1991, Green 2001). Correlations between the first contribution and components loading were 190
tested. As homoscedasticity (Levene’s test) was verified for both adults and young individuals, 191
the influence of sex, year, foraging tactic and site on BSI was compared between using a nested 192
ANOVA I (like BMI and wing length comparison) coupled with stepwise backward and Tukey 193
HSD post-hoc test. 194
To further assess the extent of morphological differentiation between individuals 195
feeding on territories and individuals feeding in flocks, we performed discriminant function 196
analyses (DFA), separately for each sex and age class. Quadratic discriminant analyses were 197
performed since the assumption of homogeneity of the variance-covariance matrices was not 198
met (Stevens 1992), except for adult males for which a linear discriminant function was used 199
(Box’s M test: adult males: Mbox = 6.89, χ² = 6.83, df = 6, P = 0.34; adult females: Mbox = 200
14.48, χ² = 14.36, df = 6, P = 0.03; juvenile males: Mbox = 21.45, χ² = 18.22, df = 6, P < 0.01 201
et juvenile females: Mbox = 15.14, χ² = 13.62, df = 6, P = 0.03). For each DFA, reliability was 202
assessed using the cross-validation procedure (Tabachnick and Fidell 1996). 203
204
Differences in payoffs between foraging tactics 205
Following Sol et al. (2005), we assessed potential differences in payoff between the two 206
foraging tactics through the comparison of body condition. Body condition was defined as 207

Monceau et al. 40 pages
body mass controlled by body size (Jacob et al. 1996), using BSI as a covariable (García-208
Berthou 2001). The effects of sex, years, foraging tactic and site were tested in an ANCOVA, 209
followed by stepwise backward procedure. Residuals from the regression of log of body mass 210
against log of BSI were used for post-hoc tests. 211
212
Data on morphology were analysed with software R version 2.8.0. (R Development 213
Core Team 2008) and the packages “car” for Levene’s test, “MASS” for linear regression, 214
quadratic and linear discriminant analyses, “nlme” for ANOVA II, and “stats” et “multcomp” 215
for GLM analyses and Tukey HSD test; all other current functions were provided by R basic 216
packages. 217
218
219
MOLECULAR ANALYSES 220
221
Blood samples (40 µL twice) were obtained by puncture of the brachial vein and 222
collected using sodium heparinised capillary tube (Hirschmann Laborgeräte, Germany). 223
Samples were kept in 800 µl of storage buffer (70% ethanol and 30% of Tris-EDTA buffer pH 224
8) and kept at 4°C for about two months before being processed. 225
226
DNA extraction 227
One hundred microliters of blood in storage buffer were centrifuged at 4000 rpm for 1 228
minute to pellet blood cells. The supernatant was discarded before adding 200 µL of Queen’s 229
Lysis buffer (Seutin et al. 1991). Samples were incubated at 55°C with 0.1 mg of proteinase K 230
overnight. DNA was then extracted based on a standard phenol-chloroform method, as 231
described in Hillis et al. (1996). DNA was resuspended into 100 µL of Tris-EDTA (pH 8). 232
233

Monceau et al. 40 pages
Molecular sex identification and sex ratios 234
Sex was identified through size variation of introns of the Chromo-Helicase-DNA 235
binding protein genes (CHD1-Z and CHD1-W) using 2550F/2718R primer pair (Fridolfsson 236
and Ellegren 1999). PCRs were carried out in 10 µL volume including ca 100 ng DNA 237
template, 200 µM each of dNTP, 200 nM each of primer, 1X reaction buffer and 0.45 U 238
HotMaster DNA polymerase (5-PRIME). A T3 thermocycler (Biometra) was used beginning 239
with an initial denaturation at 94°C for 1.5 min, followed by 35 cycles of 20 s at 94°C, 30 s at 240
46°C and 30 s at 65°C, and a final extension step at 65°C for 3 min. Intron size variation was 241
easily scored (CHD1-W = 450 pb and CHD1-Z = 750) on 6 cm long 3 % agarose gels (TBE 242
1X) run for 30 min at 100V. Accuracy of typing was assessed through verifying the 243
congruence of sexing results with another primer pair P2/P8 (Griffiths et al 1998) for a subset 244
of individuals (Monceau et al. in prep). 245
Sex ratio in adults and juveniles were assessed using binomial tests and then compared 246
between years, foraging tactics and sites using Chi square tests or Fisher’s exact tests. 247
248
Microsatellite genotyping 249
About 30 adult individuals were randomly chosen from each site for each year (247 250
individuals in total), resulting in eight sampling units allowing both spatial and temporal 251
comparisons. Polymorphism was scored at seven microsatellite markers (ZaD121, ZaD108, 252
ZaD11, ZaD119, ZaD104, ZaD105 and ZaD1) specifically developed for Z. aurita (Monceau 253
et al. 2009). PCR and genotyping were as described in Monceau et al. (2009). 254
255
Genetic differentiation 256
Linkage disequilibrium between loci was first investigated for each sampling units. 257
Then, departure from Hardy-Weinberg equilibrium was investigated using Fisher’s exact test 258
for each sampling unit, as well as for the all data set considered as a single unit, allowing for 259

Monceau et al. 40 pages
detection of a possible Wahlund effect. Both pairwise and overall genetic differentiation 260
between sampling units were evaluated using Fisher’s exact test (Raymond and Rousset 1995) 261
and FST-statistics (Weir and Cockerham 1984). An estimate of the effective migration rate 262
among sampling units (Nm) was calculated from the equation Nm = (1- FST)/4 FST after Wright 263
(1969). Analyses were processed using ARLEQUIN v. 2.000 software (Schneider et al. 1997). 264
Fisher’s exact test probabilities were approximated by Markov chain. Departure from null 265
value for FST was tested through permuting genotypes between sampling unit, and 0.05 266
significant level was adjusted with Bonferroni’s correction for multiple comparisons. Finally, 267
we evaluated whether genetic differentiation was more pronounced when comparing between 268
sites where birds used a different foraging tactic than when comparing between sites where 269
birds used the same foraging tactic. To that end, Fst values were calculated for all pairs of sites, 270
and compared using the Osx statistic implemented in FSTAT software (Goudet 1995), after 271
pooling data from the two years. 272
273
274
RESULTS 275
276
Differences in age and sex composition between foraging tactics 277
278
At each site, age composition did not differ between 2007 and 2008 (Fisher’s exact test: 279
Bellairs: P = 0.07, Roberts: P = 0.23, Harbour: P = 1 and Sunset Crest: P = 0.17). Data from 280
the two years were therefore pooled for subsequent analyses (Table 1). The proportion of 281
juveniles among all individuals captured was significantly lower than that of adults in all sites 282
(Binomial test, P < 0.0001 for each site). The proportion of juveniles differed significantly 283
between Bellairs and the three other sites (Chi² test: χ² = 37.06, df = 1, P < 0.0001), while there 284
was no difference between the two flock-feeding sites (χ² = 0.13, df = 1, P = 0.71). 285

Monceau et al. 40 pages
Sex ratio in juveniles did not differ between years at any site (Fisher's Exact Test, 286
Bellairs: P = 0.15, Sunset Crest: P = 0.64, Roberts: P = 1 and Harbour: P = 0.07), justifying 287
data pooling in subsequent analyses. A balanced sex ratio was observed in both territorial and 288
group-feeding sites (Table 2), without significant difference between replicates for each 289
foraging tactic (Fisher's Exact Test, territorial feeding: P = 0.56; flock-feeding: P = 0.41). 290
There was no significant difference in sex ratio between juveniles foraging on territories or in 291
flocks (Chi² test, χ² = 0.05, df = 1, P = 0.83). 292
Sex ratio in adults did not differ between years at any site (Chi² test, Bellairs: χ² = 0.73, 293
df = 1, P = 0.39, Sunset Crest: χ² = 0.06, df = 1, P = 0.80, Roberts: χ² = 3.47, df = 1, P = 0.06 294
and Harbour: χ² = 1.29, df = 1, P = 0.26), and data were pooled. A balanced sex ratio was 295
observed in the two territorial sites (Table 2) with no difference them (χ² = 0.28, df = 1, P = 296
0.60). In contrast, the sex ratio in both Roberts and Harbour was significantly female-biased 297
(Table 2), with no difference between sites (χ² = 0.23, df = 1, P = 0.63). Overall, there was a 298
significant difference in sex ratio between birds feeding in territories and birds feeding in 299
groups (χ² = 11.21, df = 1, P < 0.001). 300
301
302
Feeding specialization – bill morphology 303
In both adults and juveniles, BMI represented a large part of the total variance observed 304
(Table 3; juveniles: 54.21%; adults: 43.07%). Only foraging tactic had an influence on BMI in 305
juveniles (F1, 73= 6.91, P < 0.05, Tukey HSD P < 0.05), with bill being longer and thicker, but 306
thinner at the base, in juveniles feeding in groups compared to those captured on territories. In 307
adults, BMI differed between sexes (F1, 782= 281.42, P < 0.0001, Tukey HSD test, P < 0.0001), 308
with bill shape being globally thinner in females than in males in both territorial and group-309
feeding birds (F1, 782= 28.36, P < 0.0001, Tukey HSD test, P < 0.0001), while no significant 310
interaction was observed between sex and foraging tactic. ANOVA and Tukey HSD tests also 311

Monceau et al. 40 pages
revealed that flock-feeders’ bills were larger in 2008 than in 2007 (F1, 782= 20.52, P < 0.0001, 312
Tukey HSD P < 0.001) whereas the situation was reversed for territorial birds (2007 > 2008, 313
Tukey HSD P < 0.05). 314
315
Competitive ability – wing length 316
Neither sex, foraging tactic, or site had an influence on wing length in juveniles (Fig. 2a 317
left column sex: F1,73 = 0.470, P = 0.50; foraging tactic: F1,73 = 2.62, P = 0.11; site: F2,73 = 1.50, 318
P = 0.23 ). In adults, GLM and post hoc test (Tukey HSD test, all P values < 0.0001) revealed 319
that, within each foraging tactic, males had longer wings than females. Overall, territorial birds 320
had longer wings than group-feeding ones, and males feeding in groups had longer wings than 321
territorial females (Table 4 and Fig. 2a right column). For each comparison, no difference 322
between years was detected. 323
324
Overall morphological differentiation 325
Body size index (BSI) and variables included in PCA were all negatively correlated in 326
both juveniles and adults (Table 3). In both age categories, BSI represented a large part of total 327
variance observed (juveniles: 69.71%; adults: 52.28%). Juveniles feeding in groups were larger 328
than those caught on territories (F1, 73= 4.49, P < 0.05, Tukey HSD test, P < 0.05; Fig. 2b left 329
column). In adults, males were larger than females (F1, 782= 317.86, P < 0.0001, Tukey HSD 330
test, P < 0.0001; Fig. 2b right column) and, overall, territorial birds were larger than birds 331
feeding in groups (F1, 782= 5.83, P < 0.05, Tukey HSD test, P < 0.05; Fig. 2b right column). 332
ANOVA and Tukey HSD tests indicated that the significant interaction between site and year 333
of capture was due to the largest body size of adult birds feeding in groups in 2008 compared 334
to 2007 (F2, 782= 15.94, P < 0.0001; Tukey HSD test, P < 0.001; Fig. 2b right column). 335

Monceau et al. 40 pages
A posteriori classifications performed on the basis of the discriminant functions were 336
rather poor, correctly classifiying only 72.09% of adult males, 64.13% of adult females, 60% 337
of juvenile males, and 54.54% of juvenile females. 338
339
Resource payoffs - body condition 340
In juveniles, variation in body mass was only explained by variation in body size (F1, 341
73= 61.17, P < 0.0001), and was independent of foraging tactic and sex. A negative relationship 342
between body mass and BSI (Spearman’s rank correlation test ρ = - 0.58, P < 0.0001), 343
indicated that body mass increased with body size, since BSI was negatively correlated with all 344
its components (see Table 1 and Fig. 3a). In adults, BSI (F1, 782= 200.83, P < 0.0001) and, to a 345
lesser extent, the interaction between year and site (F1, 782= 4.77, P < 0.05) had a significant 346
effect on body mass. Body mass of adults increased with body size in the same way than 347
juveniles (Pearson’s product-moment correlation between body mass and BSI: r = - 0.45, P < 348
0.0001). Post hoc test also failed to identify any difference between groups. Overall, body 349
condition did not differ according to foraging tactic, sex, or age (Fig. 3b). 350
351
Genetic differentiation 352
Genotyping at seven microsatellite loci for 30 individuals for each sampling sites and 353
years (that is eight sampling units) revealed a high level of polymorphism with an average of 354
9.43 (6-14) alleles per locus (Appendix A). No evidence for linkage disequilibrium was 355
detected in any sampling unit. No multi-loci deviation from Hardy-Weinberg equilibrium 356
(HWE) was detected (Appendix A), in each sampling unit considered separately (Fisher’s 357
exact tests, all P ≥ 0.60) or when data from all sampling units were pooled (Fisher’s exact test, 358
P = 1.00). There was no difference in allele frequencies, either when comparing between pairs 359
of sampling units (Fisher’s exact test, all P ≥ 0.48), or overall (Fisher’s exact test: P = 0.70) 360
(Appendix B). Similarly, pairwise Fst-values between sampling units (-0.006 to 0.011) were 361

Monceau et al. 40 pages
small, and not significantly different from zero. A large (Slatkin 1985) absolute number of 362
migrants was estimated between sites, with a minimum of 46.2 effective migrants per 363
generation (Appendix B). Globally, the four sampled sites could be considered as one 364
genetically homogenous population in space as in time. 365
366
DISCUSSION 367
368
Metareplications, i.e. replications of entire studies addressing the same phenomenon in 369
the same species, remain scarce in ecology (Johnson 2002, McCaffery and Ruthrauff 2004, 370
Kelly 2006). However, a single study, even well designed and executed, can lead to spurious 371
results (Johnson 2006). Revisiting the evidence for the existence of a resource polymorphism 372
in Zenaida doves in Barbados provides a different picture from what was reported by Sol et al. 373
(2005) on the same population. 374
Although we were able to confirm the larger size of territorial adults, irrespective of 375
their sex, compared to adults feeding in flocks, other results differ from those obtained by Sol 376
et al. (2005). First, contrary to earlier findings, age and, especially, sex composition of doves 377
did significantly differ between the two foraging tactics. We found a larger proportion of 378
juveniles on territorial sites compared to areas where birds feed in flocks in the present study, 379
whereas Sol et al. (2005) observed no such difference. However, the effect is largely due to the 380
unusually high proportion of juveniles in Bellairs in 2007 in the present study. This might be 381
simply explained by stochastic differences in productivity between years. Indeed, variation in 382
breeding chronology in Zenaida doves has been observed between Caribbean islands (Nellis et 383
al. 1984, Wiley 1991), and between years for the same island (Nellis et al. 1984). 384
The difference between the present study and that of Sol et al. (2005) for what concerns 385
the proportions of males and females engaged in each foraging tactic, on the other hand, is 386
most probably due to the method used for sexing birds. Sol et al. (2005) relied on differences in 387

Monceau et al. 40 pages
the color of the back, supposedly less reddish in females than in males. In contrast, we relied 388
here on molecular markers to assign a sex to each captured dove. The second method is 389
generally considered to be more reliable (Griffiths and Tiwari 1993, van de Pol et al. 2009). 390
Consistency in female-biased sex-ratio in areas where birds feed in flock over two consecutive 391
years further indicates that the phenomenon is not an artifact. Disparate sex ratios in studies of 392
the closely-related mourning dove, Z. macroura, have been previously attributed to errors in 393
assigning genders from external appearance (Schulz et al. 1995). Relying on molecular markers 394
for sex identification may then provide a more reliable estimate of variation in sex-ratio 395
between Zenaida doves engaged in alternative foraging tactics. 396
Results between the present study and that of Sol et al. (2005) also differ for what 397
concerns variation in body condition between birds feeding on territories and birds feeding in 398
flocks. Whereas Sol et al. (2005) observed differences in body condition between birds caught 399
on territories and birds caught when feeding in groups, and a lower body condition in males 400
compared to females, no such effects were observed in the present study. The observed 401
interaction between year and site suggests, however, that body condition may fluctuate in time 402
and space at the population level. The observed discrepancy between the present study and that 403
of Sol et al. (2005) could be due to a difference in the calculation of BMI between the two 404
studies. Sol et al. (2005) included weight in their index of body size, whereas, following 405
recommendations from Piersma and Davidson (1991) and Green (2001), the same variable was 406
not included in the present study. However, adding body weight in the computation of the body 407
size index did not affect our conclusions (results not shown). Therefore, our results indicate 408
that the two types of foraging strategies have equal payoffs, at least as assessed from the body 409
condition of individuals, as previoulsy done by Sol et al. (2005). Indeed, feeding in flocks at 410
grain storage facilities may provide benefits and costs. At both Harbour and Roberts, birds 411
have access all day long to very large quantities of food, particularly maize. Maize is very rich 412
in lipids, and columbids use lipids better than carbohydrates as energy sources (Goodman and 413

Monceau et al. 40 pages
Griminger 1969, Sales and Janssens 2003a). Indeed, maize has very high digestibility and is 414
readily stored in the body of columbid species as fat (Hullar et al. 1999, Sales and Janssens 415
2003b). Fat provides the energy necessary for muscle function (George and Jyotti 1955) and is 416
an important component of crop milk (Shetty and Hedge 1991). In addition, birds might be less 417
vulnerable to predators when feeding in flocks than when feeding alone or in pairs on 418
territories. Evidence suggests that columbids feeding in large flocks benefit from both the 419
"dilution" (Foster and Treherne 1981) and "many eyes" (Pulliam 1973) effects, and, 420
consequently, achieve higher rates of food acquisition than when feeding alone or in small 421
groups (Phelan 1987, Cézilly and Brun 1989, Sadedin and Elgar 1998, Dias 2006). In addition, 422
we never saw any mangoose (Herpestes javanicus) or feral cats (Felix catus) at the two sites 423
where doves forage in flocks, whereas both predators are frequently seen in areas where doves 424
defend territories, and have been seen attacking and capturing doves (F. Cézilly pers. obs., J. 425
Moreau pers. com.; see also Nellis et al. 1984). On the other hand, non-selective culling 426
campaigns occasionally conducted at least at the Harbour site could result in high mortality 427
among birds feeding in flocks (L. Lefebrve pers. com.). To what extent the balance between 428
costs and benefits differ between the two foraging tactics is then difficult to establish, and even 429
more so in the absence of a more direct mesure of fitness such as survival rate or breeding 430
success. Future studies could then use radiotracking to follow individual birds feeding on 431
territories or in flocks, in order to provide information on their breeding activity, reproductive 432
output and mortality. 433
Finally, genetic analyses failed to reveal any significant level of genetic differentiation 434
between individuals feeding on territories and individuals feeding in groups, as previously 435
hypothesied by Sol et al. (2005) based on casual observations of birds moving between 436
territorial and flock feeding sites. Given the large sample size, the number of microsatellite loci 437
used in the present study and their degree of polymorphism, we are confident that even a low 438
level of genetic differentiation would have been detected if present. Studies on resource 439

Monceau et al. 40 pages
polymorphism have revealed a variable amount of gene flow among sympatric morphs (Smith 440
and Skúlason 1996, Dynes et al. 1999). However, there is some evidence that variation in 441
spatial distribution linked to variation in feeding habits can promote partial reproductive 442
isolation, non-random mating in vertebrates, and genetic differentiation in vertebrates. For 443
instance, Dynes et al. (1999) observed a significant genetic differentiation between pelagic and 444
littoral forms of brook charr (Salvelinus fontinalis) in a lake of moderate size and depth. 445
Similar results have been obtained for the Arctic char (S. alpinus) in Icelandic lakes (Gíslason 446
et al. 1999). More recently, Bergek and Björklund (2009) reported congruent morphological 447
and genetic differentiation at a microgeographic scale in the perch (Perca fluviatilis). In citril 448
finches (Serinus citronella), a strong genetic and morphological differentiation has been 449
observed at a reduced geographical sale (< 5km), in spite of current gene flow (Señar et al. 450
2006). 451
Taken together, our results suggest that the existence of alternative foraging tactics in 452
Zenaida doves in Barbados does not conform quite well to the concept of "resource 453
polymorphism", as ordinarily defined in the ecological literature (Skúlason and Smith 1995, 454
Smith and Skúlason 1996). Confirming earlier results (Sol et al. 2005), we found no evidence 455
for the existence of discrete morphs linked to each foraging strategy in Zenaida doves, as often 456
reported in true cases of resource polymorphism (Skúlason and Smith 1995, Smith and 457
Skúlason 1996, Whiteley 2007; but see Maerz et al. 2006). Sol et al. (2005) argued, however, 458
that birds feeding in flocks were mainly "floaters" and/or small sized individuals forced by 459
intraspecific competition to use lower rewarding resources. The argument was partly based on 460
the observation that individuals classified as "floaters" on territories were smaller than 461
territorial individuals, but did not differ in size from flock-feeding animals. Such a pattern may 462
arise, however, from a larger proportion of females among both "floaters" and group feeding 463
birds, as observed for the latter in the present study. In addition, territorial females could be 464

Monceau et al. 40 pages
more likely than males to engage in extraterritorial forays, particularly during their fertile 465
period (see Humbird and Neudorf 2008), and be incorrectly classified as "floaters". 466
But more importantly, the assumption that floating is an inferior strategy compared to 467
holding a territory might deserve further consideration (Brown and Long 2007). Freed from 468
spatial constraints, floaters might be better able than sedentary birds to find and exploit 469
patchily distributed food resources. Thus, a more parsimonious explanation for the existence of 470
alternative foraging tactics in Zenaida doves in Barbados, is that flock feeding is simply a 471
facultative strategy, conditioned by local food availability, and sex and age-related variation in 472
benefit associated with holding a territory. Juveniles and young adults with a low resource 473
holding potential and poorly developed parental abilities may benefit more in the short term 474
from exploiting a rich and predictable resource than from attempting to defend a territory. 475
Adult females may also be less able to defend a territory when unpaired, and engage more than 476
males in flock feeding, thus explaining the larger proportion of females among flock-feeders. 477
Difference in body size between birds caught when feeding alone or in pairs and birds caught 478
when feeding in flocks may then reflect not only variation in competitive ability but also 479
differences in age between adult birds. Indeed, evidence shows that wing length can increase 480
significantly with age for several years in birds (Pienkowski and Minton 1973, Merom et al. 481
1999, Pérez-Tris and Telleria 2001). If flock-feeding is an alternative strategy particularly 482
rewarding for young adults, we would expect that a significant proportion of flock-feeding 483
birds would eventually become territorial later on. Out of 400 birds ringed at Roberts or 484
Harbour, nine were resigthed on territorial sites (seven on one occasion, and two on two 485
occasions) one or two years later. 486
The present study illustrates both the importance of metareplication (Johnson 2002) and 487
the value of molecular tools in population biology and behavioral ecology (Hughes 1998, 488
Parker et al. 1998). In addition, it suggests that caution should be exerted when referring to 489

Monceau et al. 40 pages
well-defined ecological concepts such as "resource polymorphism" to interpret 490
intrapopulational variation in foraging strategy. 491
492
493
ACKNOWLEDGMENTS 494
495
We are particularly grateful to Mr Steve A. Devonish and Natural Heritage Department 496
for allowing us to conduct our study in Barbados. We thank the Centre de Recherches par le 497
Baguage des Populations d’Oiseaux (Paris, France), and particularly Olivier Dehorter and 498
Denis Couvet, for providing us with metal rings and authorization to ring Zenaida doves in the 499
Caribbeans. We also wish to thank Alison Mayers and Wayne Worrell for hospitality, Richard 500
Haynes, Victor Small, Sylvester Rowe, and Pat Odle for logistic assistance, and Nicole 501
Atherley, Laure Cauchard, Maria Gaillard, Estelle Harrang, Marc Lavoie, Kirk Mayers, Jérome 502
Moreau, Georges Prato, and Amélie Slaski for assistance in catching and ringing doves. We are 503
particularly grateful to Louis Lefebvre for very constructive and useful comments on an earlier 504
draft. We also thank Neeltje Boogert, Sarah Overrington, and Daniel Sol for useful discussions. 505
KM was supported by a doctoral grant from the Ministère de la Recherche et de 506
l'Enseignement Supérieur. Financial support was provided by CNRS and the Institut Buffon 507
(Université de Bourgogne). 508
509
REFERENCES 510
Alisauskas, R. T., and C. D. Ankney. 1990. Body size and fecundity in Lesser Snow geese. The 511
Auk 107:440-443. 512
Arnqvist, G., and T. Mårtensson. 1998. Measurement error in geometric morphometrics: 513
empirical strategies to assess and reduce its impact on measures of shape. Acta Zoologica 514
Academiae Scientiarum Hungaricae 44:73-96. 515

Monceau et al. 40 pages
Bailey, R. C., and J. Byrnes. 1990. A new, old method for assessing measurement error in both 516
univariate and multivariate morphometric studies. Systematic Zoology 39:124-130. 517
Bergek, S., and M. Björklund. 2009. Genetic and morphological divergence reveals local 518
subdivision of perch (Perca fluviatilis L.). Biological Journal of the Linnean Society 519
96:746-758. 520
Bolnicki, D. I., and B. M. Fitzpatrick. 2007. Sympatric speciation: Models and empirical 521
evidence. Annual Review of Ecology Evolution and Systematics 38:459-487. 522
Borras, A., J. Cabrera, and J.C. Senar. 2008. Local divergence between Mediterranean 523
crossbills occurring in tow different species of pine. Ardeola 55: 169-177. 524
Brown, D. R., and J. A. Long. 2007. What is a winter floater? Causes, consequences, and 525
implications for habitat selection. The Condor 109:548-565. 526
Carlier, P., and L. Lefebvre. 1996. Differences in individual learning between group-foraging 527
and territorial Zenaida doves. Behaviour 133:1197-1207. 528
Carlier, P., and L. Lefebvre. 1997. Ecological differences in social learning between adjacent, 529
mixing, populations of Zenaida doves. Ethology 103:772-784. 530
Cézilly, F., and B. Brun. 1989. Surveillance et picorage chez la tourterelle rieuse, Streptopelia 531
risoria: effets de la présence d'un congénère et de la dispersion des graines. Behaviour 532
110:146-160. 533
Dias, R. I. 2006. Effects of position and flock size on vigilance and foraging behaviour of the 534
Scaled dove Columbina squammata. Behavioural Processes 73:248-252. 535
Dolman, C. S., J. Templeton, and L. Lefebvre. 1996. Mode of foraging competition is related 536
to tutor preference in Zenaida aurita. Journal of Comparative Psychology 110:45-54. 537
Dynes, J., P. Magnan, L. Bernatchez, and M. A. Rodríguez. 1999. Genetic and morphological 538
variation between two forms of lacustrine Brook charr. Journal of Fish Biology 54:955-972. 539
Foster, W. A., and J. E. Treherne. 1981. Evidence for the dilution effect in the selfish herd 540
from fish predation on a marine insect. Nature 293:466-467. 541

Monceau et al. 40 pages
Fridolfsson, A., and H. Ellegren. 1999. A simple and universal method for molecular sexing of 542
non-ratite birds. Journal of Avian Biology 30:116 - 121. 543
Garcia-Berthou, E. 2001. On the misuse of residuals in ecology: testing regression residuals vs. 544
the analysis of covariance. Journal of Animal Ecology 70:708-711. 545
George, J. C., and D. Jyotti. 1955. The lipid content and its reduction in the muscle and liver 546
during long and sustained muscular activity. Journal of Animal Morphology and Physiology 547
2:2-9. 548
Gíslason, D., M. M. Ferguson, S. Skúlason, and S. S. Snorrason. 1999. Rapid and coupled 549
phenotypic and genetic divergence in Icelandic Arctic char (Salvelinus alpinus). Canadian 550
Journal of Fisheries and Aquatic Sciences 56:229-2234. 551
Goldberg, J. L., J. W. A. Grant, and L. Lefebvre. 2001. Effects of the temporal predictability 552
and spatial clumping of food on the intensity of competitive aggression in the Zenaida dove. 553
Behavioral Ecology 12:490-495. 554
Goodman, H. M., and P. Griminger. 1969. Effect of dietary energy source on racing 555
performance in the pigeon. Poultry Science 48:2058-2063. 556
Goss-Custard, J. D., and S. E. A. Le. V. dit, Durell. 1983. Individual and age differences in the 557
feeding ecology of oystercatchers Haematopus ostralegus wintering on the Exe Estuary, 558
Devon. Ibis 125:155-171. 559
Goudet, J. 1995. FSTAT (Version 1.2): A computer program to calculate F-statistics. Journal 560
of Heredity 86:485-486. 561
Green, A. J. 2001. Mass/length residuals: measures of body condition or generators of spurious 562
results? Ecology 82:1473-1483. 563
Griffiths, R., and B. Tiwari. 1993. The isolation of molecular genetic markers for the 564
identification of sex. Proceedings of the National Academy of Sciences of the United States 565
of America 90:8324-8326. 566

Monceau et al. 40 pages
Griffiths, R., M. C. Double, K. Orr, and R. J. C. Dawson. 1998. A DNA test to sex most birds. 567
Molecular Ecology 7:1071-1075. 568
Hillis, D. M., C. Moritz, and B. K. Mable. 1996. Molecular Systematics, Second Edition. 569
Sunderland, Massachusets, USA. 570
Hughes, C. 1998. Integrating molecular techniques with field methods in studies of social 571
behavior: A revolution results. Ecology 79:383-399. 572
Hullar, I., I. Meleg, S. Fekete, and R. Romvar. 1999. Studies of the energy content of pigeon 573
feeds I. Determination of digestibility and metabolizable energy content. Poultry Science 574
78:1757-1762. 575
Humbird, S. K., and D. L. H. Neudorf. 2008. The effects of food supplementation on 576
extraterritorial behavior in female Northern cardinals. The Condor 110:392-395. 577
Jacob, E. M., S. D. Marshall, and G. W. Uetz. 1996. Estimating fitness: a comparison of body 578
condition indices. Oikos 77:61-67. 579
Johnson, D. H. 2002. The importance of replication in wildlife research. Journal of Wildlife 580
Management 66:919-932. 581
Johnson, D. H. 2006. The many faces of replication. Crop Science 46:2486-2491. 582
Kelly, C. D. 2006. Replicating empirical research in behavioral ecology: How and why it 583
should be done but rarely ever is. The Quarterly Review of Biology 81:221-236. 584
Lefebvre, L., B. Palameta, and K. K. Hatch. 1996. Is group-living associated with social 585
learning? A comparative test of gregarious and a territorial columbid. Behaviour 133:241-586
261. 587
Lessells, C. M., and P. T. Boag. 1987. Unrepeatable repeatabilities: a common mistake. The 588
Auk 104:116-121. 589
Maerz, J. C., E. M. Myers, and D. C. Adams. 2006. Trophic polymorphism in a terrestrial 590
salamander. Evolutionary Ecology Research 8:23-35. 591

Monceau et al. 40 pages
McCaffery, B. J., and D. R. Ruthrauff. 2004. Spatial variation in shorebird nest success: 592
implications for inference. Wader Study Group Bulletin 103:67-70. 593
McCormack, J. E., and T. B. Smith. 2008. Niche expansion leads to small-scale adaptive 594
divergence along an elevation gradient in a medium-sized passerine bird. Proceedings of the 595
Royal Society of London B 275:2155-2164. 596
Merom, K., R. McCleery, and Y. Yom-Tov. 1999. Age-related changes in wing-length and 597
body mass in the Reed warbler Acrocephalus scirpaceus and Clamorous Reed warbler A. 598
stentoreus. Bird Study 46:249-255. 599
Monceau, K., M. Gaillard, E. Harrang, D. Santiago-Alarcon, P. Parker, F. Cézilly, and R. 600
Wattier. 2009. Twenty-three polymorphic microsatellite markers for the Caribbean endemic 601
Zenaida dove, Zenaida aurita, and its conservation in related Zenaida species. Conservation 602
Genetics 10:1577-1581. 603
Nellis, D. W., R. A. Dewey, M. A. Hewitt, S. Imsand, R. Philibosian, and J. A. Yntema. 1984. 604
Population status of Zenaida doves and other columbids in the Virgin Islands. Journal of 605
Wildlife Management 48:889-894. 606
Olsson, J., R. Svanbäck, and P. Eklöv. 2007. Effects of resource level and habitat type on 607
behavioral and morphological plasticity in Eurasian perch. Oecologia 152:48-56. 608
Parker, P. G., A. A. Snow, M. D. Schug, G. C. Booton, and P. A. Fuerst. 1998. What molecules 609
can tell us about populations: choosing and using a molecular marker. Ecology 79:361-382. 610
Parsons, K. J., and B. W. Robinson. 2007. Foraging performance of diet-induced morphotypes 611
in Pumpkinseed sunfish (Lepomis gibbosus) favours resource polymorphism. Journal of 612
Evolutionary Biology 20:673-684. 613
Pérez-Tris, J., and J. L. Telleria. 2001. Age-related variation in wing shape of migratory and 614
sedentary blackcaps Sylvia atricapilla. Journal of Avian Biology 32:207-213. 615
Pfennig, D. W., A. M. Rice, and R. A. Martin. 2007. Field and experimental evidence for 616
competition's role in phenotypic divergence. Evolution 61:257-271. 617

Monceau et al. 40 pages
Phelan, J. P. 1987. Some components of flocking behavior in the Rock dove (Columbia livia). 618
Journal of Field Ornithology 58:135-143. 619
Pienkowski, M. W., and C. D. T. Minton. 1973. Wing length changes of the knot with age and 620
time since moult. Bird Study 20:63-68. 621
Piersma, T., and N. C. Davidson. 1991. Confusions of mass and size. The Auk 108:441-443. 622
Proulx, R., and P. Magnan. 2004. Contribution of phenotypic plasticity and heredity to the 623
trophic polymorphism of lacustrine Brook charr (Salvelinus fontinalis M.). Evolutionary 624
Ecology Research 6:503-522. 625
Pulliam, H. R. 1973. On the advantages of flocking. Journal of Theoretical Biology 38:419-626
422. 627
R Development Core Team 2008. R: A language and environment for statistical computing. R 628
Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL 629
http://www.R-project.org. 630
Raffaele, H., J. Wiley, O. Garrido, A. Keith, and J. Raffaele. 1998. Birds of the West Indies, 631
London, UK. 632
Raymond, M., and F. Rousset. 1995. An exact test for population differentiation. Evolution 633
49:1280-1283. 634
Rivera-Milán, F. F. 1997. Seasonal and annual changes in the population density of Zenaida 635
doves in the xerophytic forest of Guanica, Puerto Rico. Journal of Field Ornithology 636
68:259-272. 637
Rivera-Milán, F. F. 1999. Population dynamics of Zenaida doves in Cidra, Puerto Rico. Journal 638
of Wildlife Management 63:232-244. 639
Rohwer, S. 1990. Foraging differences between white and dark morphs of the Pacific Reef 640
heron Egretta sacra. Ibis 132:21-26. 641
Roulin, A. 2004. Covariation between plumage colour polymorphism and diet in the Barn owl 642
Tyto alba. Ibis 146:509-517. 643

Monceau et al. 40 pages
Ruzzante, D. E., S. J. Walde, V. E. Cussac, P. J. Macchi, M. F. Alonso, and M. Battini. 2003. 644
Resource polymorphism in a Patagonian fish Percichthys trucha (Percichthyidae): 645
phenotypic evidence for interlake pattern variation. Biological Journal of the Limnean 646
Society 78:497-515. 647
Sadedin, S. R., and M. A. Elgar. 1998. The influence of flock size and geometry on the 648
scanning behaviour of Spotted Turtle doves, Streptopelia chinensis. Australian Journal of 649
Ecology 23:177-180. 650
Sales, J., and G. P. J. Janssens. 2003a. Nutrition of the domestic pigeon. World's Poultry 651
Science Journal 59:221-232. 652
Sales, J., and G. P. J. Janssens. 2003b. Methods to determine metabolizable energy and 653
digestibility of feed ingredients in the Domestic pigeon (Columba livia domestica). Poultry 654
Science 82:1457-1461. 655
Schluter, D. 2000. The Ecology of Adaptive Radiation. Oxford University Press, Oxford, UK. 656
Schneider, S., D. Roessli, and L. Excoffier. 2000. Arlequin: A software for population genetics 657
data analysis. Ver 2.000. Genetics and Biometry Lab, Dept. of Anthropology, University of 658
Geneva. 659
Schulz, J. H., S. L. Sheriff, Z. Q. He, C. E. Braun, R. D. Drobney, R. E. Tomlinson, D. D. 660
Dolton, and R. A. Montgomery. 1995. Accuracy of techniques used to assign Mourning 661
doves age and gender. Journal of Wildlife Management 59:759-765. 662
Señar, J. C. 1999. La medición de la repetibilidad y el error de medida. Etologuia 17:53-64. 663
Señar, J. C., A. Borras, J. Cabrera, T. Cabrera, and M. Björklund. 2006. Local differentiation in 664
the presence of gene flow in the Citril finch Serinus citronella. Biology Letters 2:85-87. 665
Seutin, G., B. N. White, and P. T. Boag. 1991. Preservation of avian blood tissue samples for 666
DNA analyses. Canadian Journal of Zoology 69:82-90. 667
Shetty, S., and S. N. Hedge. 1991. Changes in lipids of pigeon milk in the 1st week of its 668
secretion. Lipids 26:930-933. 669

Monceau et al. 40 pages
Skúlason, S., and T. B. Smith. 1995. Resource polymorphisms in vertebrates. Trends in 670
Ecology and Evolution 10:366-370. 671
Slatkin, M. 1985. Gene flow in natural-populations. Annual Review of Ecology and 672
Systematics 16:393-430. 673
Smith, T. B. 1990. Resource use by bill morphs of an African finch: evidence for intraspecific 674
competition. Ecology 71:1246-1257. 675
Smith, T. B., and S. A. Temple. 1982. Feeding habits and bill polymorphism in Hook-billed 676
kites. The Auk 99:197-207. 677
Smith, T. B., and S. Skulason. 1996. Evolutionary significance of resource polymorphisms in 678
fishes, amphibians, and birds. Annual Review of Ecology and Systematics 27:111-133. 679
Sokal, R. R., and F. J. Rohlf. 1995. Biometry, 3rd edition. W. H. Freeman and Company, New 680
York, USA. 681
Sol, D., M. Elie, M. Marcoux, E. Chrostovsky, C. Porcher, and L. Lefebvre. 2005. Ecological 682
mechanisms of resource polymorphism in Zenaida doves of Barbados. Ecology 86:2397-683
2407. 684
Stevens, J. 1992. Applied Multivariate Statistics for Social Sciences. 2nd. ed. Lawrance 685
Erlbaum Associates Publishers, Mahwah, USA. 686
Svanbäck, R., P. Eklöv, R. Fransson, and K. Holmgren. 2008. Intraspecific competition drives 687
multiple species resource polymorphism in fish communities. Oikos 117:114-124. 688
Tabachnick, B. G., and L. S. Fidell. 1996. Using Multivariate Statistics (3rd ed.). Harper 689
Collins, New York, USA. 690
van de Pol, M., K. Oosterbeek, A. L. Rutten, B. J. Ens, J. M. Tinbergen, and S. Verhulst. 2009. 691
Biometric sex discrimination is unreliable when sexual dimorphism varies within and 692
between years: an example in Eurasian oystercatchers Haematopus ostralegus. Ibis 693
151:171-180. 694

Monceau et al. 40 pages
Venables, W. N., and B. D. Ripley. 1999. Modern Applied Statistics with S-plus Springer-695
Verlag, New York, USA. 696
Weir, B. S., and C. C. Cockerham. 1984. Estimating F-statistics for the analysis of population-697
structure. Evolution 38:1358-1370. 698
Whiteley, A. R. 2007. Trophic polymorphism in a riverine fish: morphological, dietary, and 699
genetic analysis of mountain whitefish. Biological Journal of the Linnean Society 92:253-700
267. 701
Wiley, J. W. 1991. Ecology and behavior of the Zenaida dove. Ornitologia Neotropical 2:49-702
75. 703
Wright, S. 1969. Evolution and the Genetics o f Populations.Vol. 2. The Theory of Gene 704
Frequencies. University of Chicago Press, Chicago, USA. 705
Yezerinac, S. M., S. C. Lougheed, and P. Handford. 1992. Measurement error and 706
morphometric studies: statistical power and observer experience. Systematic Zoology 707
41:471-482. 708
Zar, J. H. 1999. Biostatistical Analysis, fourth edition. Prentice Hall, Upper Saddle River, 709
USA. 710
711
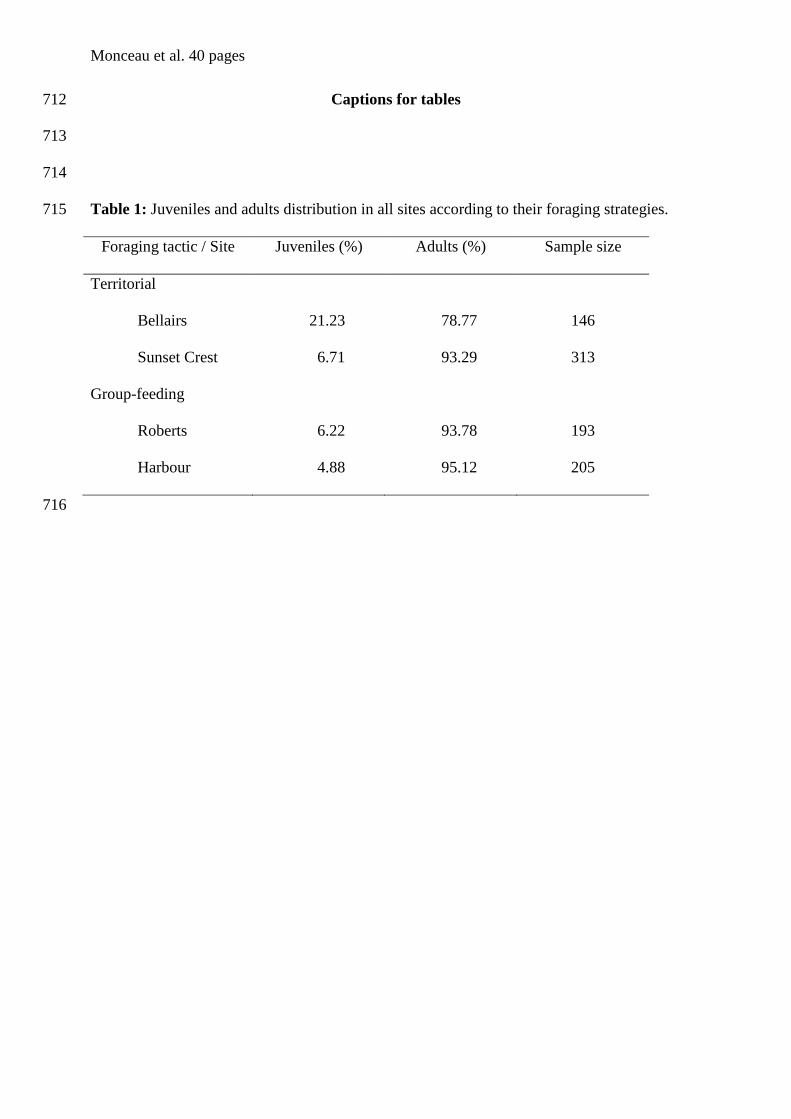
Monceau et al. 40 pages
Captions for tables 712
713
714
Table 1: Juveniles and adults distribution in all sites according to their foraging strategies. 715
Foraging tactic / Site Juveniles (%) Adults (%) Sample size
Territorial
Bellairs 21.23 78.77 146
Sunset Crest 6.71 93.29 313
Group-feeding
Roberts 6.22 93.78 193
Harbour 4.88 95.12 205
716
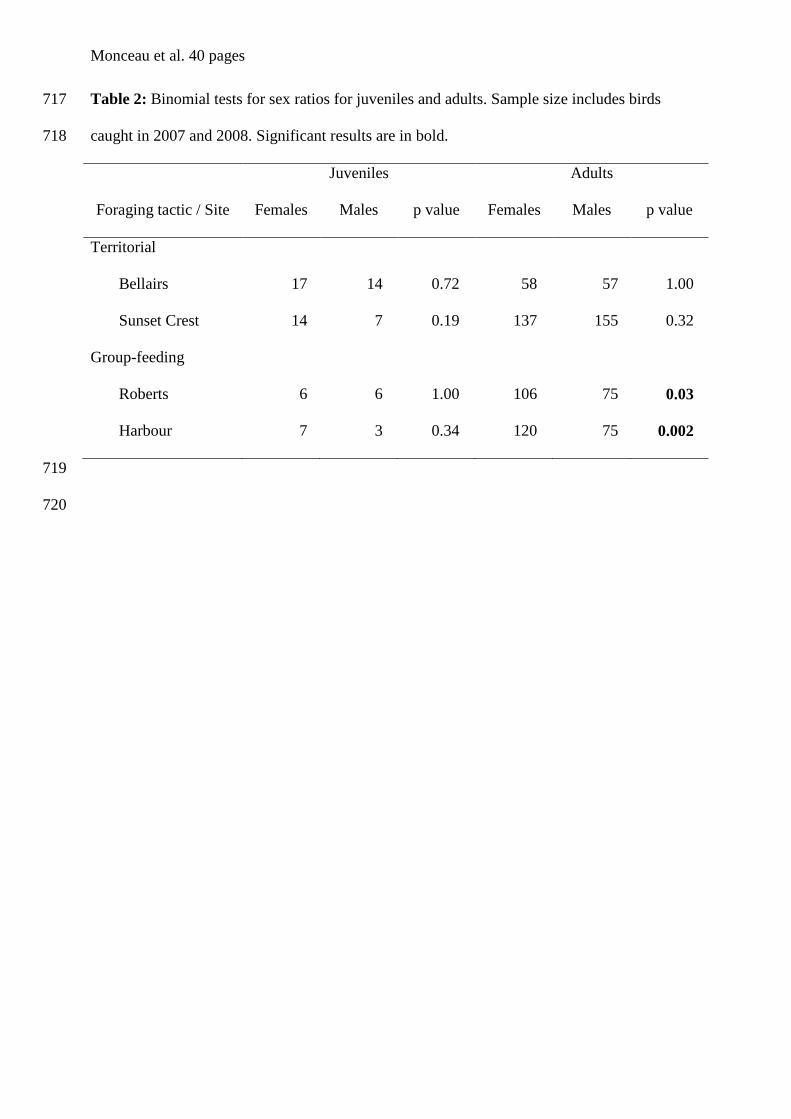
Monceau et al. 40 pages
Table 2: Binomial tests for sex ratios for juveniles and adults. Sample size includes birds 717
caught in 2007 and 2008. Significant results are in bold. 718
Juveniles Adults
Foraging tactic / Site Females Males p value Females Males p value
Territorial
Bellairs 17 14 0.72 58 57 1.00
Sunset Crest 14 7 0.19 137 155 0.32
Group-feeding
Roberts 6 6 1.00 106 75 0.03
Harbour 7 3 0.34 120 75 0.002
719
720
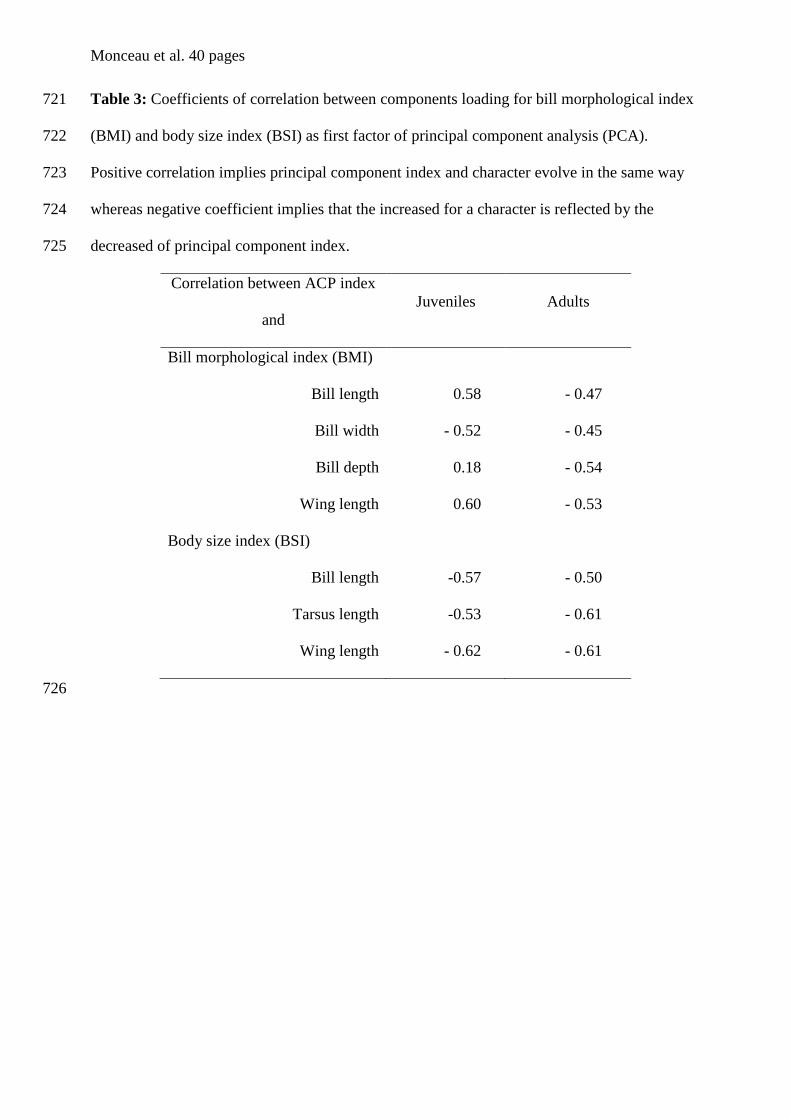
Monceau et al. 40 pages
Table 3: Coefficients of correlation between components loading for bill morphological index 721
(BMI) and body size index (BSI) as first factor of principal component analysis (PCA). 722
Positive correlation implies principal component index and character evolve in the same way 723
whereas negative coefficient implies that the increased for a character is reflected by the 724
decreased of principal component index. 725
Correlation between ACP index
and Juveniles Adults
Bill morphological index (BMI)
Bill length 0.58 - 0.47
Bill width - 0.52 - 0.45
Bill depth 0.18 - 0.54
Wing length 0.60 - 0.53
Body size index (BSI)
Bill length -0.57 - 0.50
Tarsus length -0.53 - 0.61
Wing length - 0.62 - 0.61
726
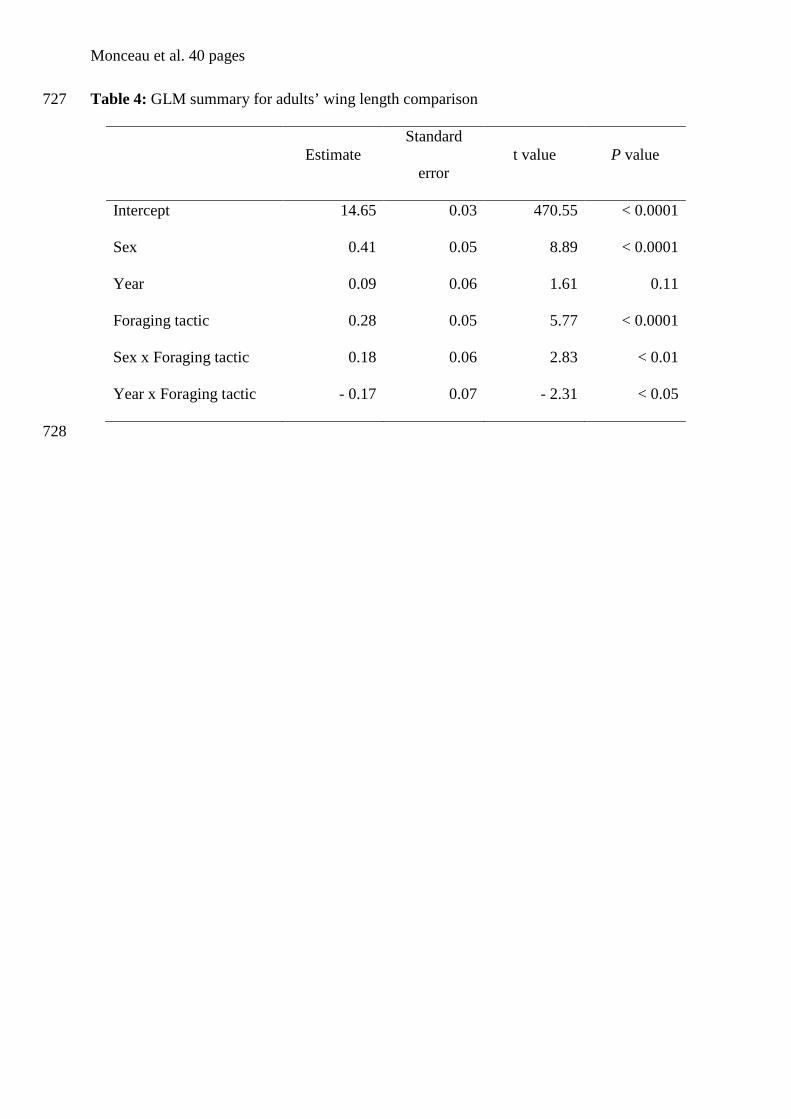
Monceau et al. 40 pages
Table 4: GLM summary for adults’ wing length comparison 727
Estimate Standard
error t value P value
Intercept 14.65 0.03 470.55 < 0.0001
Sex 0.41 0.05 8.89 < 0.0001
Year 0.09 0.06 1.61 0.11
Foraging tactic 0.28 0.05 5.77 < 0.0001
Sex x Foraging tactic 0.18 0.06 2.83 < 0.01
Year x Foraging tactic - 0.17 0.07 - 2.31 < 0.05
728

Monceau et al. 40 pages
Captions for figures 729
730
731
Figure 2: Study area. T in brackets specifies areas where Zenaida doves were territorial and 732
GF stand for group-feeding sites where they feed in flocks. 733
734
735
Figure 2: Comparison (box plot) between type of sites and sex for (a) wing length and (b) 736
body size index (BSI) for both juveniles (left column) and adults (right column). GF stand for 737
group-feeding areas and T for territorial sites. Juveniles sample size was divided as follow GF 738
females = 13, T females = 31, GF males = 9 and T males = 21. Adults individuals were divided 739
as follow: GF females = 226, T females = 195, GF males = 150 and T males = 212. Boxes 740
represented 50% of all values. Medians and means were respectively represented with plain 741
line and black point. Dash lines were standing for standard error and open circles represented 742
extreme values. To interpret BSI graphical representation, please refer to correlation table 743
(Table 1). 744
745
746
Figure 3: Body condition comparison between type of sites and years for a) juveniles and b) 747
adults. GF stand for group-feeding areas and T for territorial sites. Juveniles sample size was 748
divided as follow GF females = 13, T females = 31, GF males = 9 and T males = 21. Adults 749
individuals were divided as follow: GF females = 226, T females = 195, GF males = 150 and T 750
males = 212. Boxes represented 50% of all values. Medians were with plain line. Dash lines 751
were standing for standard error and open circles represented extreme values. 752
753
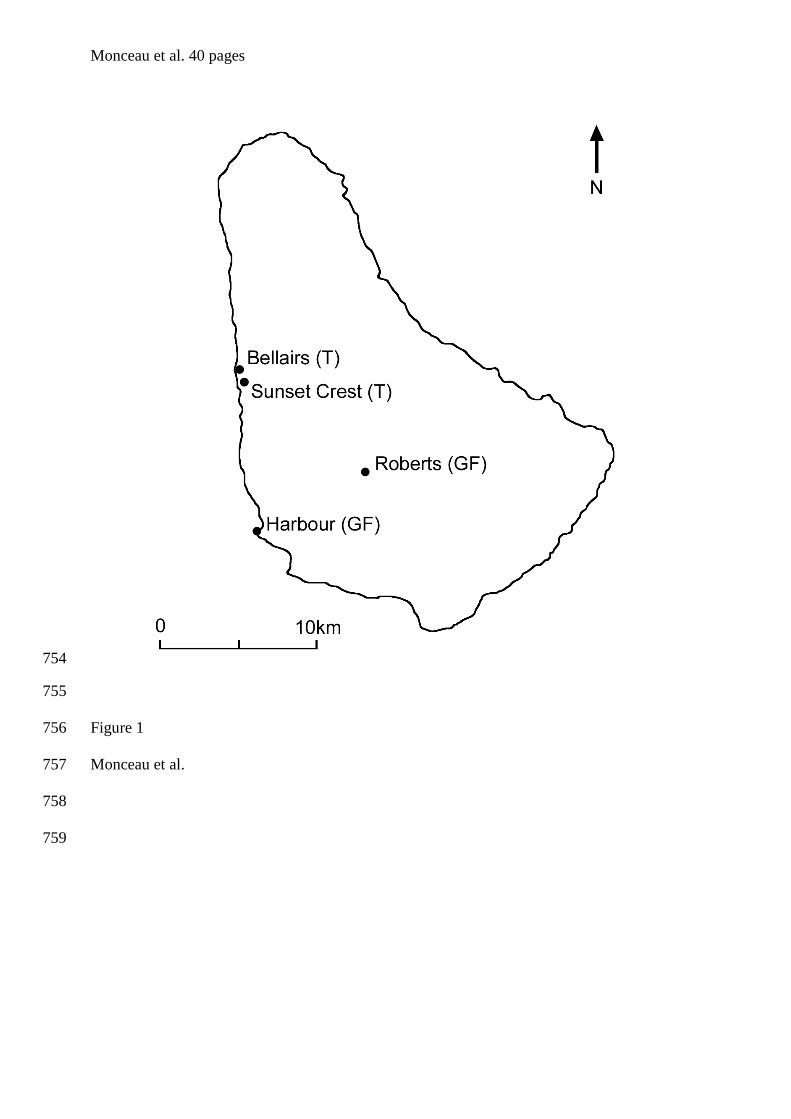
Monceau et al. 40 pages
754
755
Figure 1 756
Monceau et al. 757
758
759
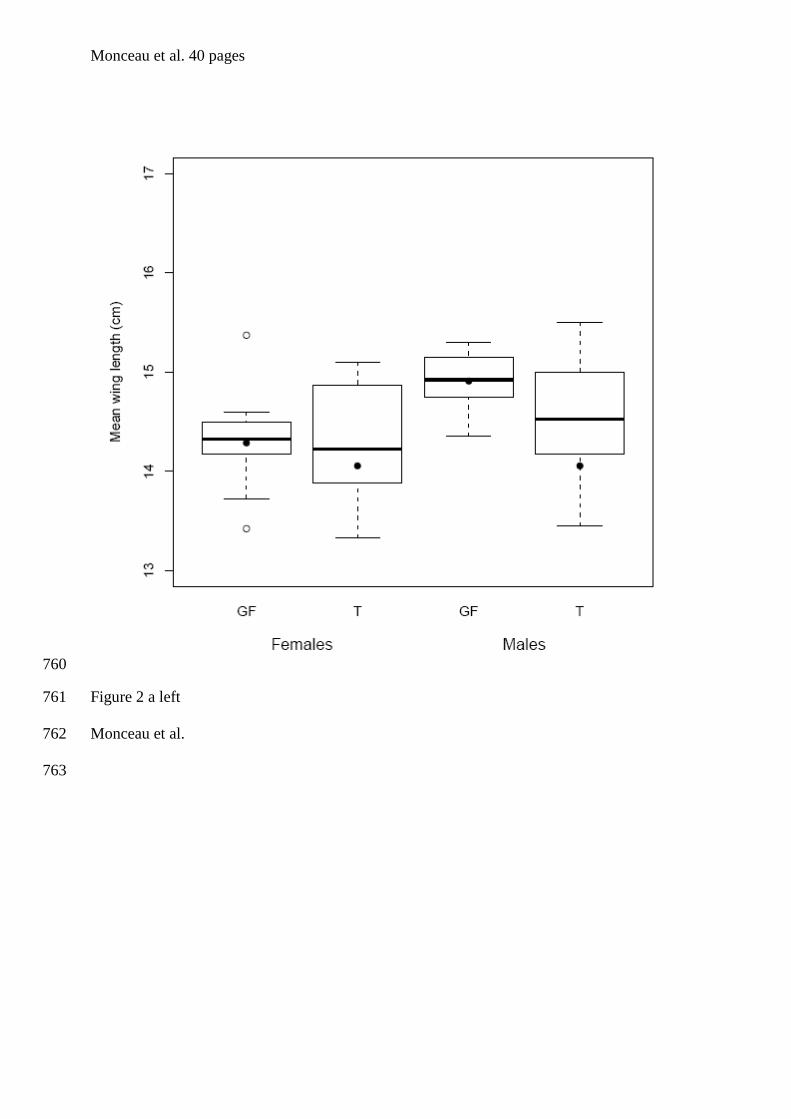
Monceau et al. 40 pages
760
Figure 2 a left 761
Monceau et al. 762
763
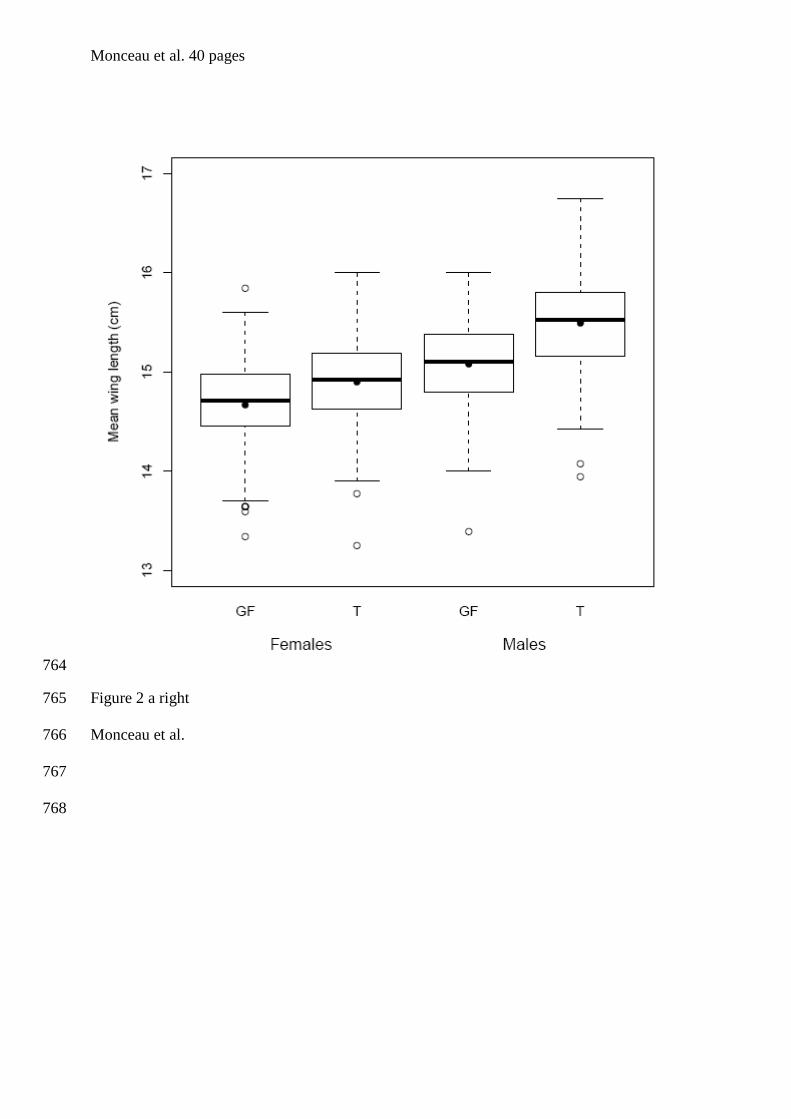
Monceau et al. 40 pages
764
Figure 2 a right 765
Monceau et al. 766
767
768
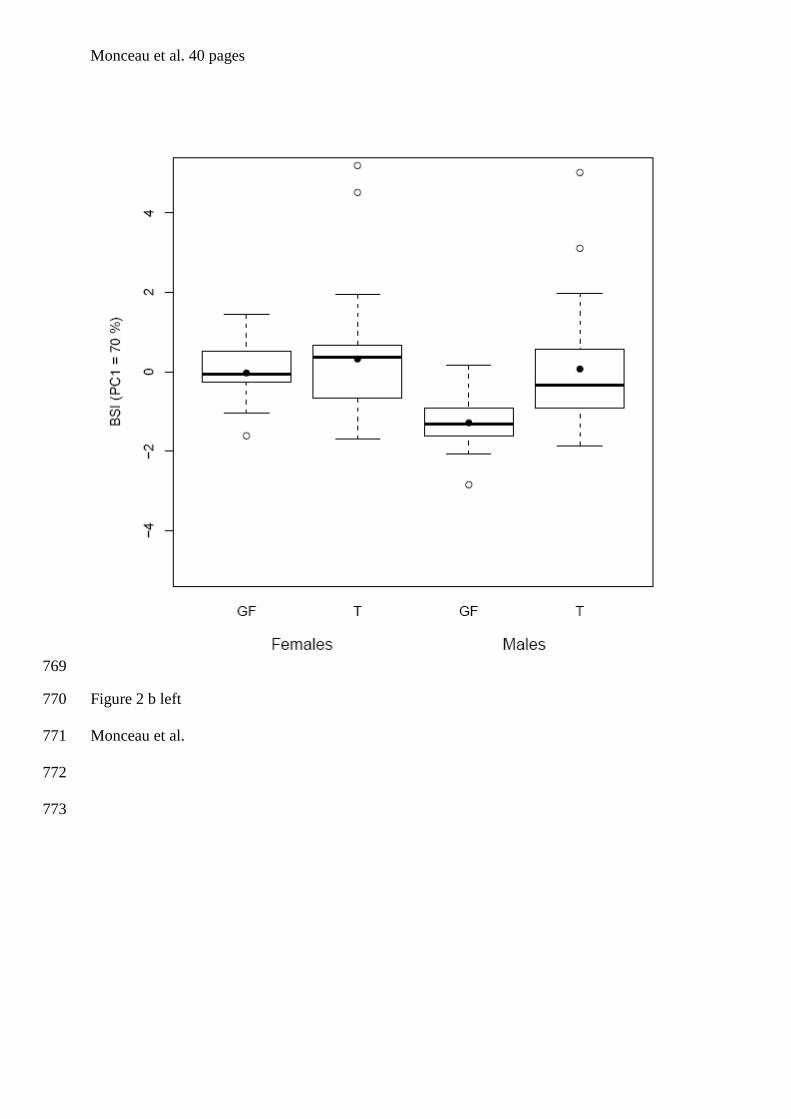
Monceau et al. 40 pages
769
Figure 2 b left 770
Monceau et al. 771
772
773
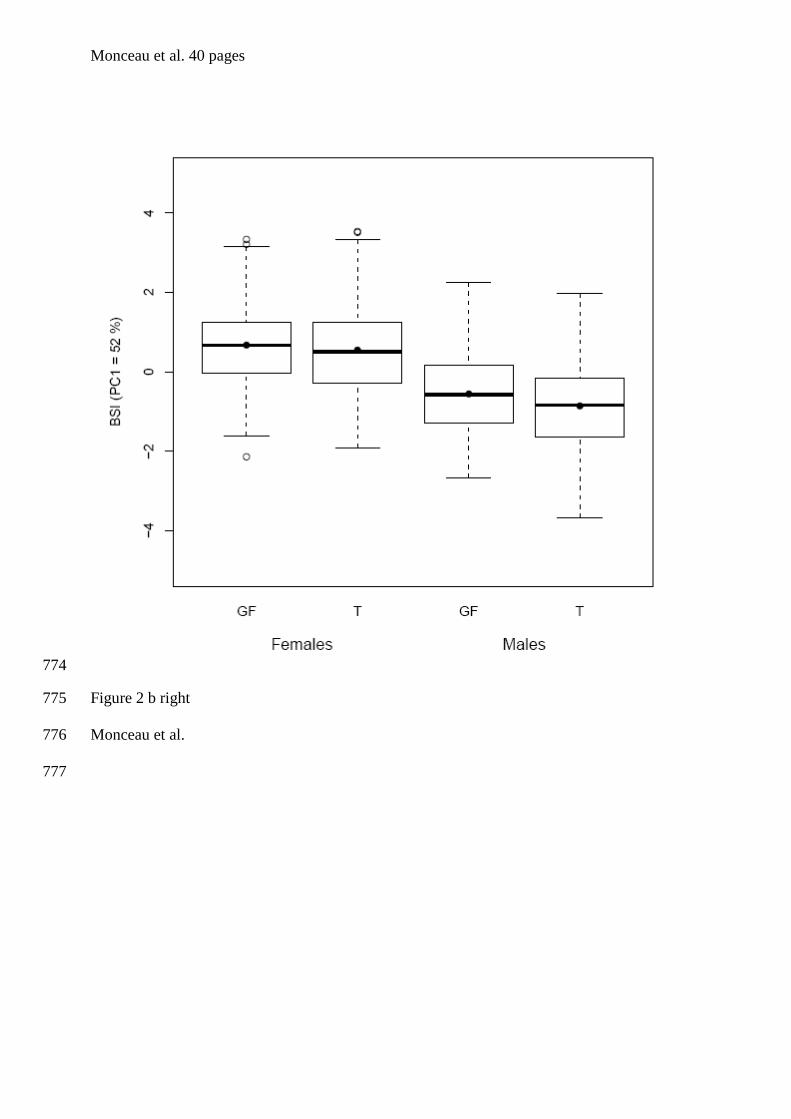
Monceau et al. 40 pages
774
Figure 2 b right 775
Monceau et al. 776
777
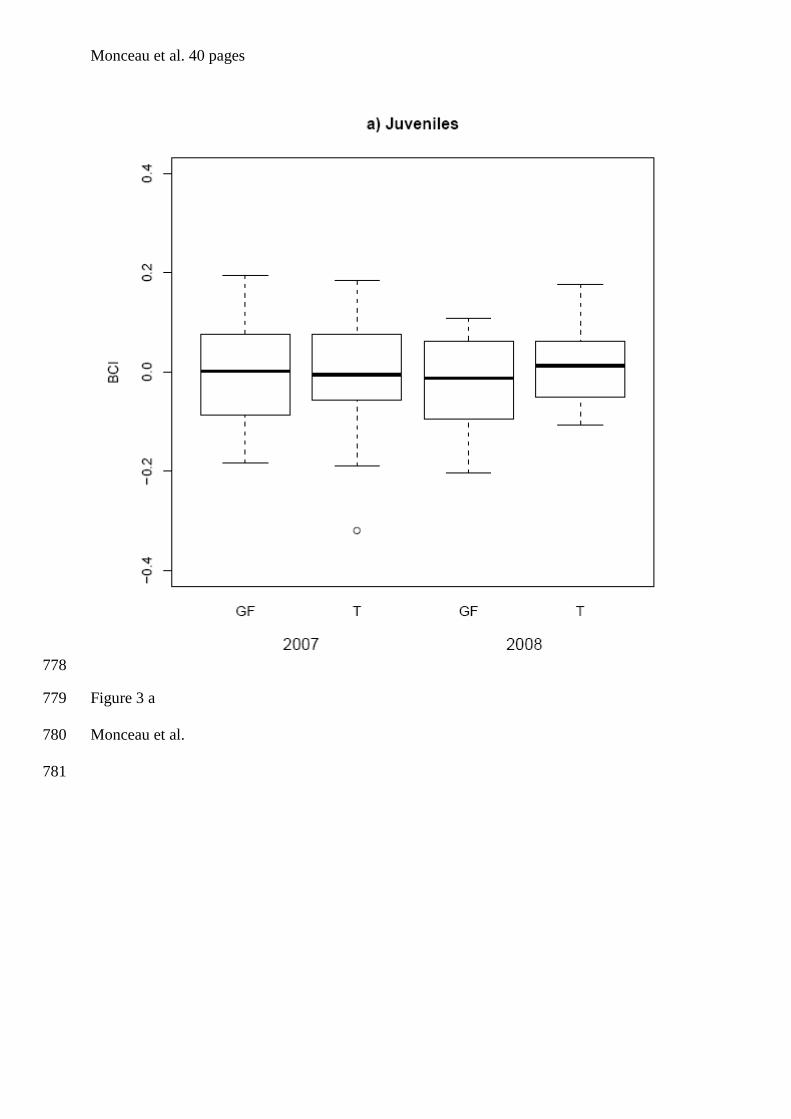
Monceau et al. 40 pages
778
Figure 3 a 779
Monceau et al. 780
781
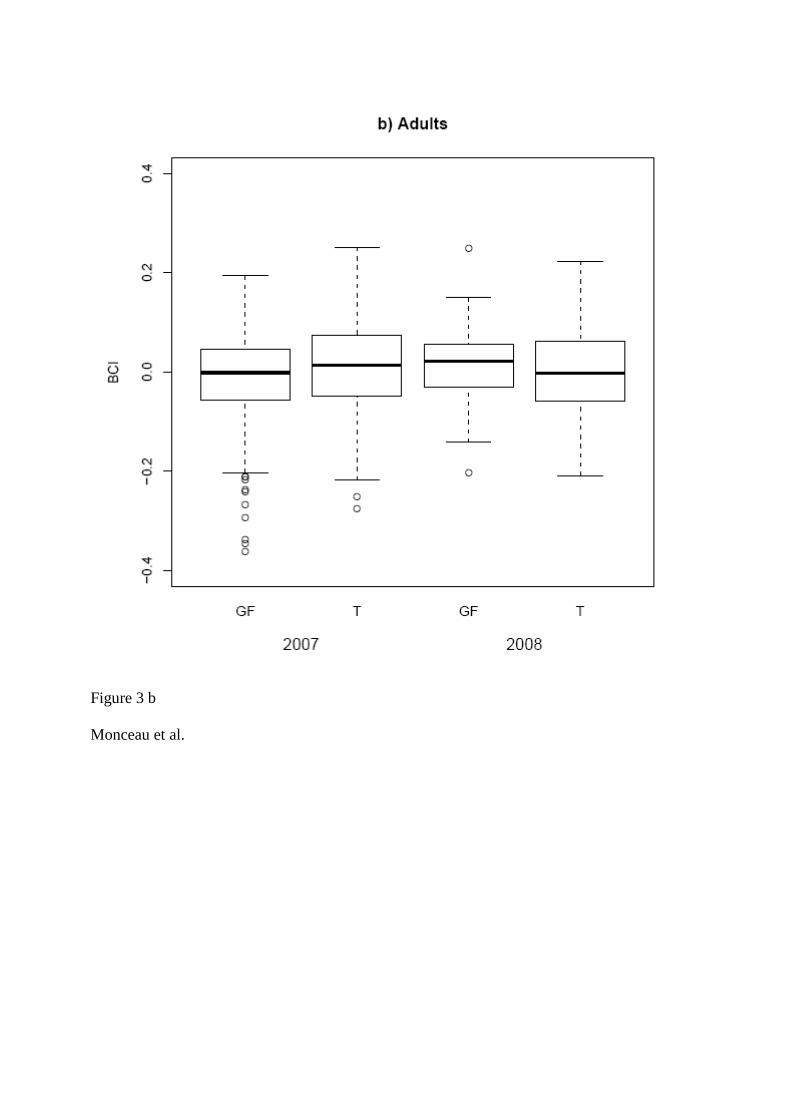
Figure 3 b
Monceau et al.
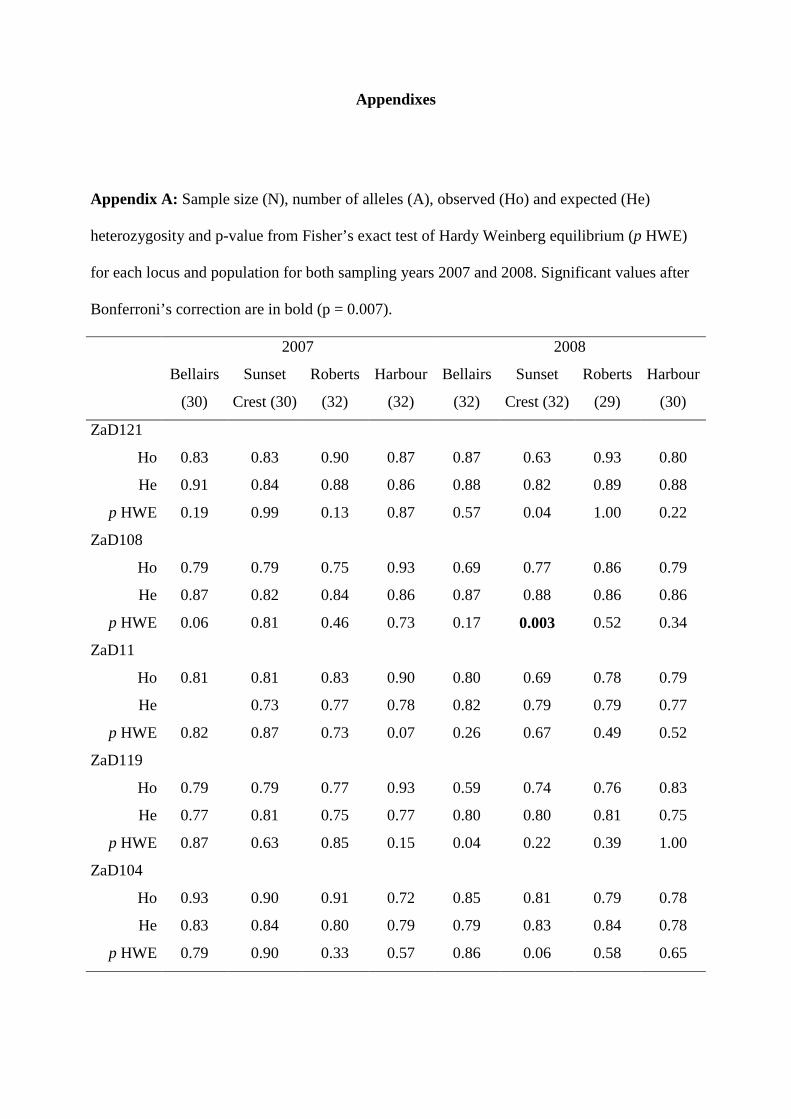
Appendixes
Appendix A: Sample size (N), number of alleles (A), observed (Ho) and expected (He)
heterozygosity and p-value from Fisher’s exact test of Hardy Weinberg equilibrium (p HWE)
for each locus and population for both sampling years 2007 and 2008. Significant values after
Bonferroni’s correction are in bold (p = 0.007).
2007 2008
Bellairs
(30)
Sunset
Crest (30)
Roberts
(32)
Harbour
(32)
Bellairs
(32)
Sunset
Crest (32)
Roberts
(29)
Harbour
(30)
ZaD121
Ho 0.83 0.83 0.90 0.87 0.87 0.63 0.93 0.80
He 0.91 0.84 0.88 0.86 0.88 0.82 0.89 0.88
p HWE 0.19 0.99 0.13 0.87 0.57 0.04 1.00 0.22
ZaD108
Ho 0.79 0.79 0.75 0.93 0.69 0.77 0.86 0.79
He 0.87 0.82 0.84 0.86 0.87 0.88 0.86 0.86
p HWE 0.06 0.81 0.46 0.73 0.17 0.003 0.52 0.34
ZaD11
Ho 0.81 0.81 0.83 0.90 0.80 0.69 0.78 0.79
He 0.73 0.77 0.78 0.82 0.79 0.79 0.77
p HWE 0.82 0.87 0.73 0.07 0.26 0.67 0.49 0.52
ZaD119
Ho 0.79 0.79 0.77 0.93 0.59 0.74 0.76 0.83
He 0.77 0.81 0.75 0.77 0.80 0.80 0.81 0.75
p HWE 0.87 0.63 0.85 0.15 0.04 0.22 0.39 1.00
ZaD104
Ho 0.93 0.90 0.91 0.72 0.85 0.81 0.79 0.78
He 0.83 0.84 0.80 0.79 0.79 0.83 0.84 0.78
p HWE 0.79 0.90 0.33 0.57 0.86 0.06 0.58 0.65
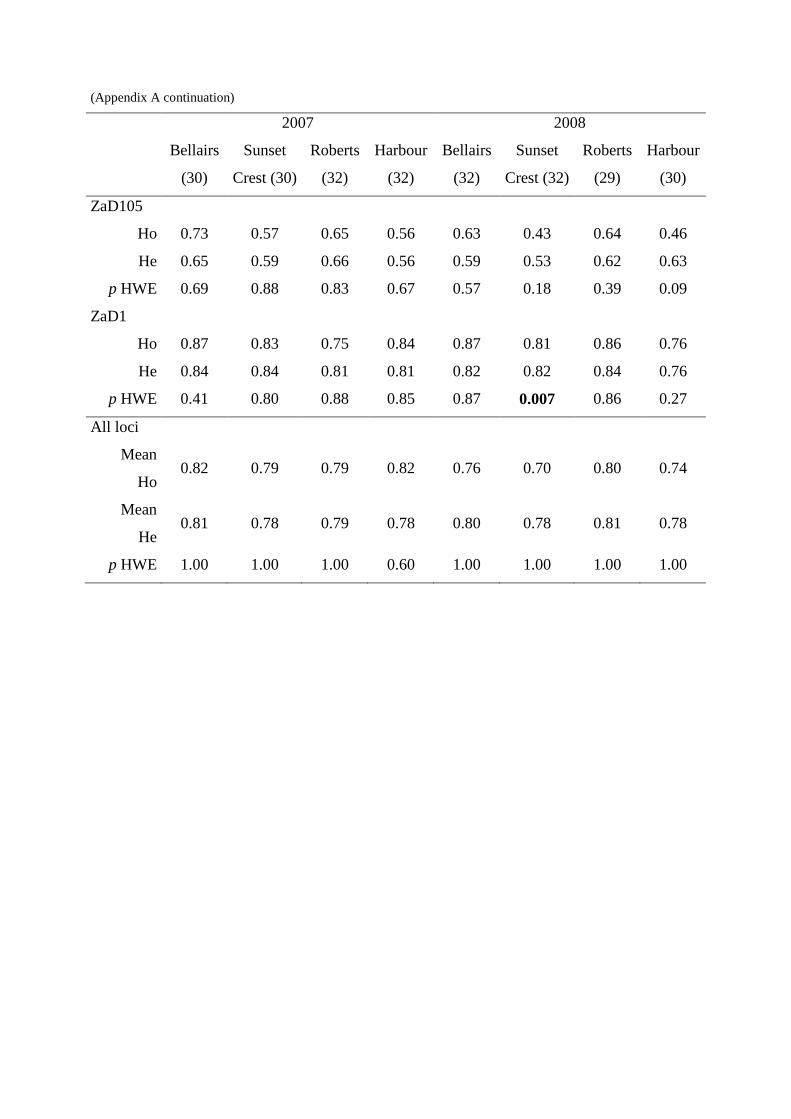
(Appendix A continuation)
2007 2008
Bellairs
(30)
Sunset
Crest (30)
Roberts
(32)
Harbour
(32)
Bellairs
(32)
Sunset
Crest (32)
Roberts
(29)
Harbour
(30)
ZaD105
Ho 0.73 0.57 0.65 0.56 0.63 0.43 0.64 0.46
He 0.65 0.59 0.66 0.56 0.59 0.53 0.62 0.63
p HWE 0.69 0.88 0.83 0.67 0.57 0.18 0.39 0.09
ZaD1
Ho 0.87 0.83 0.75 0.84 0.87 0.81 0.86 0.76
He 0.84 0.84 0.81 0.81 0.82 0.82 0.84 0.76
p HWE 0.41 0.80 0.88 0.85 0.87 0.007 0.86 0.27
All loci
Mean
Ho 0.82 0.79 0.79 0.82 0.76 0.70 0.80 0.74
Mean
He 0.81 0.78 0.79 0.78 0.80 0.78 0.81 0.78
p HWE 1.00 1.00 1.00 0.60 1.00 1.00 1.00 1.00
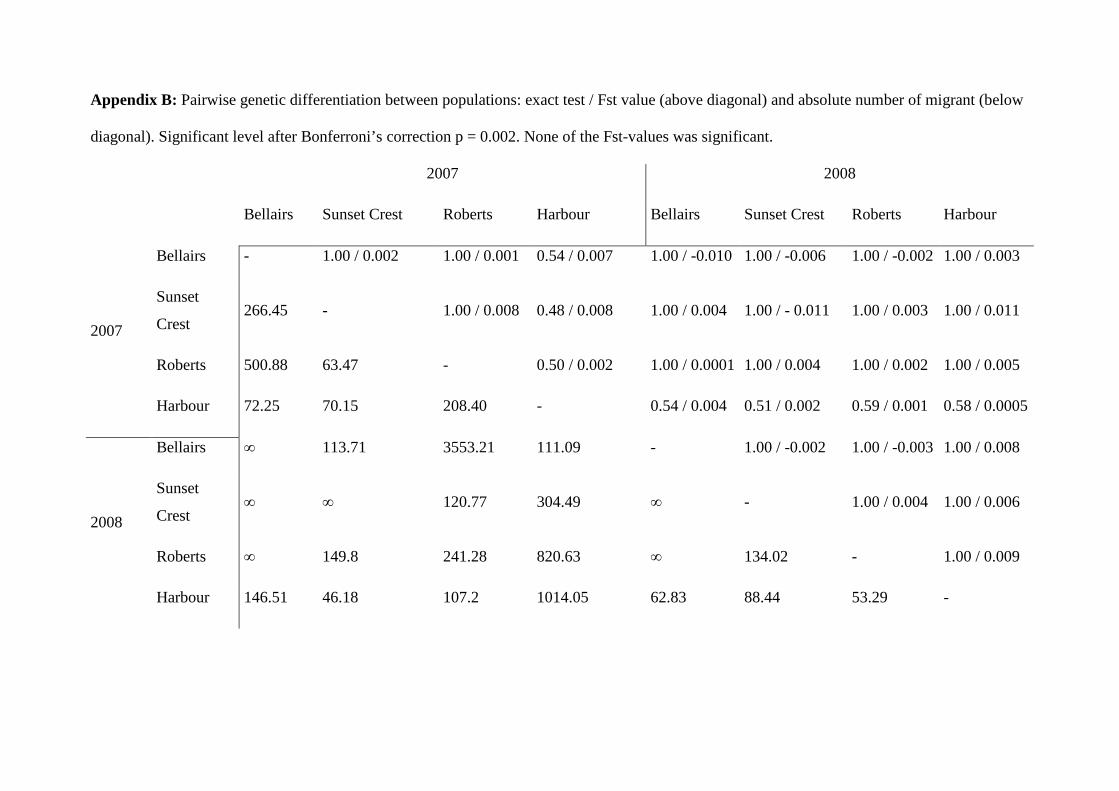
Appendix B: Pairwise genetic differentiation between populations: exact test / Fst value (above diagonal) and absolute number of migrant (below
diagonal). Significant level after Bonferroni’s correction p = 0.002. None of the Fst-values was significant.
2007 2008
Bellairs Sunset Crest Roberts Harbour Bellairs Sunset Crest Roberts Harbour
2007
Bellairs - 1.00 / 0.002 1.00 / 0.001 0.54 / 0.007 1.00 / -0.010 1.00 / -0.006 1.00 / -0.002 1.00 / 0.003
Sunset
Crest 266.45 - 1.00 / 0.008 0.48 / 0.008 1.00 / 0.004 1.00 / - 0.011 1.00 / 0.003 1.00 / 0.011
Roberts 500.88 63.47 - 0.50 / 0.002 1.00 / 0.0001 1.00 / 0.004 1.00 / 0.002 1.00 / 0.005
Harbour 72.25 70.15 208.40 - 0.54 / 0.004 0.51 / 0.002 0.59 / 0.001 0.58 / 0.0005
2008
Bellairs ∞ 113.71 3553.21 111.09 - 1.00 / -0.002 1.00 / -0.003 1.00 / 0.008
Sunset
Crest ∞ ∞ 120.77 304.49 ∞ - 1.00 / 0.004 1.00 / 0.006
Roberts ∞ 149.8 241.28 820.63 ∞ 134.02 - 1.00 / 0.009
Harbour 146.51 46.18 107.2 1014.05 62.83 88.44 53.29 -