binary structure of the two-domain (3r)-hydroxyacyl-coa dehydrogenase from rat peroxisomal...
TRANSCRIPT

Structure, Vol. 11, 87–97, January, 2003, 2003 Elsevier Science Ltd. All rights reserved. PII S0969-2126(02)00931-0
Binary Structure of the Two-Domain (3R)-Hydroxyacyl-CoA Dehydrogenase from Rat PeroxisomalMultifunctional Enzyme Type 2 at 2.38 A Resolution
mutually homologous (3R )-hydroxyacyl-CoA dehydrog-enases [1], mammalian MFE-2 contains only a singlecopy of this activity, followed by trans-2-enoyl-CoA hy-dratase-2 and a sterol carrier protein type 2 (SCP-2)-like domain. The last domain is not present in the fungal
Antti M. Haapalainen,1 M. Kristian Koski,1
Yong-Mei Qin,2 J. Kalervo Hiltunen,1
and Tuomo Glumoff1,*1Biocenter Oulu and Department of BiochemistryUniversity of OuluP.O. Box 3000 enzyme. The two NH2-terminal (3R )-hydroxyacyl-CoA
dehydrogenases in the yeast polypeptide are namedFIN-90014 University of OuluFinland dehydrogenase A and dehydrogenase B [1]. These de-
hydrogenases have different substrate specificities; A is2 Department of Biochemistry and MolecularBiology active toward medium and long chain (3R )-hydroxyacyl-
CoAs, and B shows the highest catalytic rate with shortCollege of Life SciencesPeking University chain (C4) substrates [11]. In vitro studies described that
the mammalian dehydrogenase region catalyzes the oxi-Beijing 100871China dation of 3-hydroxyl on straight chain and 2-methyl
branched chain fatty acyl-CoAs with broad chain lengthspecificity as well as 24 hydroxylated intermediates inbile acid synthesis [2, 5, 12, 13]. Mammalian MFE-2 wasSummaryinitially identified as 17�-hydroxysteroid dehydrogenase(17�-HSD) type 4, oxidizing 17�-estradiol and 5-andro-The crystal structure of (3R)-hydroxyacyl-CoA dehy-stene-3�,17�-diol [10, 12]. In view of the observationdrogenase of rat peroxisomal multifunctional enzymethat the yeast MFE-2 is also in vitro a 17�-HSD [14], thetype 2 (MFE-2) was solved at 2.38 A resolution. Thepotential physiological role of MFE-2 as a 17�-HSD iscatalytic entity reveals an �/� short chain alcohol de-unclear. The role of MFE-2 in fatty acid oxidation ishydrogenase/reductase (SDR) fold and the conforma-further supported by the observation that MFE-1, whichtion of the bound nicotinamide adenine dinucleotidealso catalyzes the second and third reactions of the(NAD�) found in other SDR enzymes. Of great interestperoxisomal � oxidation cycle, but with opposite chiralis the separate COOH-terminal domain, which is notspecificity, is not responsible for the degradation of veryseen in other SDR structures. This domain completeslong chain fatty acids (VLCFA) [15].the active site cavity of the neighboring monomer and
The deficiency of MFE-2 protein leads to very severeextends dimeric interactions. Peroxisomal diseasesclinical presentations [16]. Abnormalities include neona-that arise because of point mutations in the dehydrog-tal hypotonia, craniofacial dysmorphia, developmentalenase-coding region of the MFE-2 gene can be mappeddelay, neuronal migration defects, or demyelination,to changes in amino acids involved in NAD� bindingwhich results in early death, mostly within the first yearand protein dimerization.of life. A number of different mutations in the gene cod-ing for MFE-2 were found in humans, resulting in the
Introduction total or partial loss of its enzymatic activity [16–19].Patients with MFE-2 deficiency had elevated plasma
� oxidation of fatty acids is one of the central metabolic levels of branched chain fatty acids, bile acid intermedi-pathways in eukaryotic cells, which, in mammals, can ates, and VLCFAs, an observation also seen in MFE-2-proceed both in mitochondria and peroxisomes. The � deficient mice [20].oxidation pathway is often described as a spiral con- The (3R )-hydroxyacyl-CoA dehydrogenase part ofsisting of four reactions, with trans-2-enoyl-coenzyme MFE-2 belongs to the short chain alcohol dehydroge-A (CoA), 3-hydroxyacyl-CoA, and 3-ketoacyl-CoA as in- nase/reductase (SDR) superfamily [21]. This is a largetermediates. Multifunctional enzyme type 2 (MFE-2; EC and diverse group of NAD(H)- or NADP(H)-dependent1.1.1.62) catalyzes the second and third reactions of the oxidoreductases with low amino acid sequence similar-peroxisomal pathway; it has been cloned and character- ity found in bacteria, plants, and animals [21–27]. X-rayized from many species [1–6]. The three COOH-terminal crystal structures of close to 20 members of the familyamino acids of MFE-2 make up a peroxisomal targeting have been determined, revealing a one-domain foldingsignal type 1 (PTS1) [7, 8], which is sufficient for translo- pattern. Most of them are homotetrameric or homodi-cation into peroxisomes [9]. The highest mRNA level of meric by quaternary structure [28–33], with the excep-human MFE-2 was observed in liver and then in heart, tion of porcine carbonyl reductase, which is monomericprostate, and testis [10]. but has the basic �/� SDR fold [34]. The two most-
In contrast to the “classic” (3S )-hydroxyacyl-CoA-uti- conserved motifs are the Gly-X-X-X-Gly-X-Gly se-lizing route, MFE-2 accepts (3R )-hydroxyacyl-CoA es- quence in the site for cofactor binding [35, 36] and theters, which can be dehydrated to trans-2-enoyl or oxi- active site Ser-Tyr-Lys triad [21, 29, 37, 38].dized in an NAD(H)-dependent manner to their 3-keto In the present work the crystal structure of the (3R )-counterparts. Unlike the yeast MFE-2, which has two
Key words: 17�-HSD; MFE-2; NAD(H); � oxidation; peroxisome;SDR*Correspondence: [email protected]

Structure88
hydroxyacyl-CoA dehydrogenase region of rat MFE-2, 1X, 1X, 1X, where X represents a crossover connectionbetween the strands (Figure 3). Each of the four ���in complex with NAD�, was solved at 2.38 A resolution.
A novel feature of the determined X-ray structure is motifs along the central sheet exhibits a right-handedcrossover. Additional � sheet is formed by the � strandsthat the COOH-terminal residues constitute a separate
domain with a previously uncharacterized fold, with one �F, �G, and �H from monomer A and �I from monomerB and vice versa (Figure 2). While the �A–�F segmentmonomer lining the active site cavity of the other one.
Moreover, the COOH-terminal � strands from two differ- constitutes the classic Rossmann fold associated withthe binding of the NAD�, the �F–�I segment participatesent monomers establish a � sheet, which further in-
creases dimeric interactions. in quaternary association. The COOH-terminal � helices,�CT1, �CT2, and �H, contribute to the domain that isnot present in the other SDR proteins.Results
Identification of (3R )-Hydroxyacyl-CoA The NAD(H) Binding Site and the Mode of BindingThe bound NAD� molecule is in an extended conforma-Dehydrogenase from Rat MFE-2
Multiple-amino acid sequence alignment of the sever- tion and sits on a surface created by the COOH-terminalends of the � strands in the Rossmann fold (Figures 3al peroxisomal MFE-2s revealed nonconserved amino
acid residues (His311–Ser321 in the rat sequence) be- and 4). The nicotinamide part of NAD� is buried nearthe putative active site, while the adenine moiety locatestween the dehydrogenase and hydratase-2 regions (Fig-
ure 1). On the basis of this information, the plasmid close to the NH2-terminal part of the Rossmann fold.The adenine and nicotinamide rings are oriented roughlypET3d::dh�SCP-2L� encoding Met1–Asp319 of rat
MFE-2 was constructed. This construct lacks the DNA perpendicular to the planes of the respective riboses,resulting in an anti conformation for the adenine ringencoding hydratase-2 and the SCP-2-like domain and
was named dh�SCP-2L�. Also, the NH2-terminal frag- and a syn conformation for the nicotinamide ring (Figure4). Both ribose rings reveal a 2E (C2�-endo) puckering.ments of MFE-2, including Met1–Thr312 and Met1–
Ala316 (Figure 1), were isolated from rat liver with (3R )- A parameter frequently used to express the degree ofextension of the dinucleotide is the distance betweenhydroxyacyl-CoA dehydrogenase activity [2, 39]. The
dh�SCP-2L� was expressed in Escherichia coli and pu- C6 of the adenine and C2 of the nicotinamide in the synconformation [40]. This value for the binary complexrified chromatographically to apparent homogeneity.
Mass spectrometric analysis gave a molecular mass of dh�SCP-2L� is 14.3 A, which agrees well with thedistances reported for the other crystal structures ofof 34365.0 g/mol for the protein, corresponding to the
calculated mass of the polypeptide lacking the initial SDR proteins.Amino acid residues of the conserved SDR motifs,methionine (34365.1 g/mol). Purified dh�SCP-2L� had
a kcat value of 3.7 s�1, comparable to the kcat value of 3.4 such as Gly16, Gly20, Gly22, Ser151, Tyr164, and Lys168are in close contact with NAD�. Furthermore, Asp40s�1 obtained for the full-length recombinant rat MFE-2
under the assay conditions used. forms hydrogen bonds with the O2� and O3� hydroxylgroups of the adenine ribose moiety of the dinucleotide(Figure 4), with the bond lengths of 2.5 A and 2.6 A,Crystallization and Structure Determination
For the structure determination, dh�SCP-2L� was pro- respectively. This region regulates the specificity towardNAD(H) over NADP(H), since the side chain of this aspar-duced as a selenomethionine (SeMet)-labeled protein
with a molecular mass of 34602.0 g/mol, indicating com- tate would come into unacceptably close contact with2�-phosphate group of the adenosine moiety of NADP(H).plete labeling, with all five methionines replaced by
SeMet. Well-diffracting triangle-shaped crystals were In the NADP(H)-preferring enzymes, the aspartate resi-due is replaced by threonine or alanine.obtained, and the structure was solved by the multi-
wavelength anomalous dispersion (MAD) method. De-tails of the structure determination are presented in Ta- Quaternary Structure and the Catalytic Triad
When purified dh�SCP-2L� was applied to a size exclu-ble 1 and in Experimental Procedures.sion column, the elution volume corresponded to thesize of a homodimer. This was also the case in theOverall Structure and Topology
The crystal structure reveals two homodimers per asym- crystal, where the tertiary structures of the monomersare virtually identical (Figure 2). However, a loop of 12metric unit. The main body of the dimer is oval shaped,
with overall dimensions of 80 A � 55 A � 45 A. A ribbon amino acids between � strand �F and � helix �FG inmonomer B cannot be recognized, possibly because ofdiagram of the structure is shown in Figure 2, and a
schematic description of the structure is provided in the different crystal contacts in that area. In addition,Gly51, located just before � helix �C in monomer B,Figure 3. Both monomers of dh�SCP-2L� have an �/�
doubly-wound structure. Each monomer is built of a was not included in the model because of fragmentedelectron density. Otherwise, the polypeptide is continu-core of � sheet consisting of six parallel (�A–�F) �
strands sandwiched between two arrays of three parallel ous in both monomers: from Ser3 to Asp303 and fromPro4 to Asp303 in monomers A and B, respectively. The� helices located on each side of the � sheet (�B, �C,
and �G; �D, �E, and �F). This folding pattern is referred monomers are related to each other by a noncrystallo-graphic 2-fold axis (� 180, as calculated from theto as a basic �/� SDR fold. The two longest helices, �E
and �F, form the interfacial four-helix bundle typical of superposition of all C� atoms), and they superimposewell, with an rms deviation of 0.21 A and no conforma-dimeric and tetrameric SDR proteins. The topology of
the central sheet can be described as �1X, �1X, 3X, tional changes (Figure 5).

Crystal Structure of the SDR Region of MFE-289
Figure 1. Multiple Sequence Alignment of the (3R )-Hydroxyacyl-CoA Dehydrogenase from Eukaryotic MFE-2s
The rat dehydrogenase region was aligned with the human, pig, guinea pig, mouse, Candida tropicalis, and Saccharomyces cerevisiaecounterparts. The yeast MFE-2 has two dehydrogenases in the same polypeptide [1], and they are indicated in the figure as A and B,respectively. The alignment was performed with the BESTFIT program (EMBL, Heidelberg). The residues conserved throughout all sequencesare shaded black, and those with a high degree of sequence similarity are shaded either dark gray (80%–99%) or light gray (60%–79%). Thelocation and types of structural motifs were defined by PROMOTIF [65]. Cylinders, � helices; arrows, � strands. The roman numerals describethe type of � turn in the area, and 3�10 represents the 310 helix. The filled vertical arrows indicate the cleavage sites described in vivo resultingin separate dehydrogenase and hydratase-2 enzymes [39], and the plain vertical arrow shows the last COOH-terminal amino acid residue(Asp319) in the recombinant dh�SCP-2L�. The closed triangles indicate amino acid residues of the catalytic triad (see text), and the opentriangles point out the amino acid substitutions G16S, L21F, S177P, and V218L, as identified from patients with MFE-2 deficiency [17–19].The region marked with the dotted line above the rat sequence forms a loop that is seen neither in the nonperoxisomal dehydrogenases,such as 7�-HSD from E. coli, nor in the dehydrogenase B of the yeast MFE-2s.
The accessible surface areas [41] of a monomer and addition, the �E contains two kinks in both monomersat His123 and Ala133. In the �F-�F interface a few largea dimer with a probe of 1.4 A are 16,074 A2 and 22,686 A2,
respectively. A total of 40% of the subunit surfaces are hydrophobic side chains do not point toward the dimerinterface. Consequently, the two �F helices are closerused in the formation of the dimer. The association of
monomers into a dimer is accomplished by the COOH- to each other (4 A) than in the �E-�E interface.Figure 4 shows the arrangement of the active site interminal domain, as well as the four-helix bundle, which
is formed by the helices �E(A), �F(A), �E(B), and �F(B). the absence of substrate. Ser151, Tyr164, and Lys168form a triad essential for catalysis that is well conservedHydrophobic and aromatic interactions and hydrogen
bonds dominate the dimer interface. The �E and �F within the SDR family, emphasizing the common reac-tion mechanism [38, 42]. Both Tyr164 and Lys168 arehelices from one monomer are in contact with the corre-
sponding helices from the other monomer, such that the situated on an �F helix, and Ser151 is located in a smallhelix, �EF, between � strand �E and � helix �F. Thehelices from different monomers run nearly antiparallel.
The contact surface of the helix bundle includes resi- hydroxyl group of Tyr164, the proton acceptor in cataly-sis, is hydrogen bonded to the 2�-hydroxyl group of thedues Ser112–Val130 from �E and residues Gln161–
Thr177 from �F. In the �E-�E interface, large hydropho- nicotinamide ribose (2.6 A). Thus, the tyrosine is broughtto a close proximity of the nicotinamide ring (3.0 A),bic and polar side chains face each other, and the
backbone carbonyls are approximately 8 A apart. In which is the site of hydride transfer. The role of Lys168
Structure90
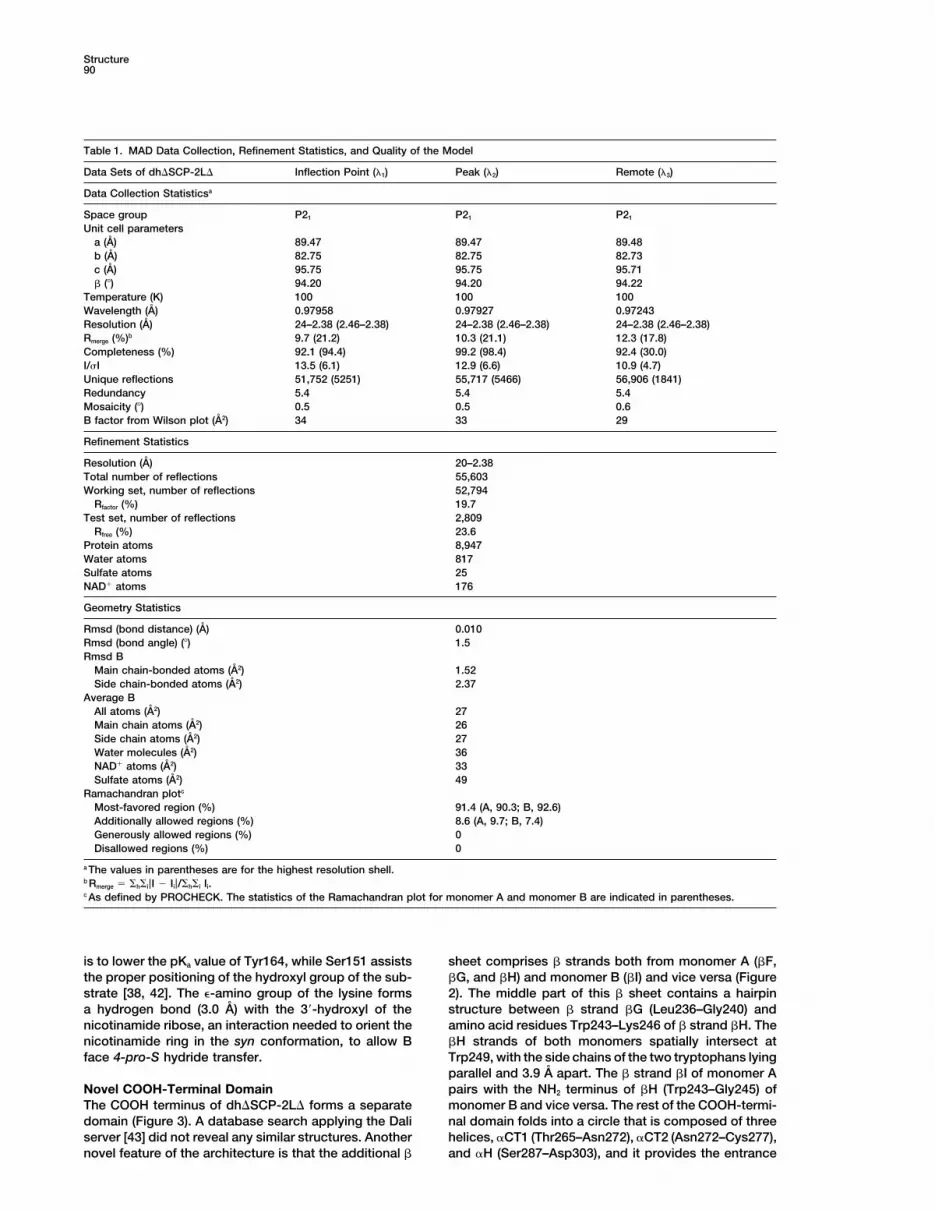
Table 1. MAD Data Collection, Refinement Statistics, and Quality of the Model
Data Sets of dh�SCP-2L� Inflection Point (�1) Peak (�2) Remote (�3)
Data Collection Statisticsa
Space group P21 P21 P21
Unit cell parametersa (A) 89.47 89.47 89.48b (A) 82.75 82.75 82.73c (A) 95.75 95.75 95.71� () 94.20 94.20 94.22
Temperature (K) 100 100 100Wavelength (A) 0.97958 0.97927 0.97243Resolution (A) 24–2.38 (2.46–2.38) 24–2.38 (2.46–2.38) 24–2.38 (2.46–2.38)Rmerge (%)b 9.7 (21.2) 10.3 (21.1) 12.3 (17.8)Completeness (%) 92.1 (94.4) 99.2 (98.4) 92.4 (30.0)I/�I 13.5 (6.1) 12.9 (6.6) 10.9 (4.7)Unique reflections 51,752 (5251) 55,717 (5466) 56,906 (1841)Redundancy 5.4 5.4 5.4Mosaicity () 0.5 0.5 0.6B factor from Wilson plot (A2) 34 33 29
Refinement Statistics
Resolution (A) 20–2.38Total number of reflections 55,603Working set, number of reflections 52,794
Rfactor (%) 19.7Test set, number of reflections 2,809
Rfree (%) 23.6Protein atoms 8,947Water atoms 817Sulfate atoms 25NAD� atoms 176
Geometry Statistics
Rmsd (bond distance) (A) 0.010Rmsd (bond angle) () 1.5Rmsd B
Main chain-bonded atoms (A2) 1.52Side chain-bonded atoms (A2) 2.37
Average BAll atoms (A2) 27Main chain atoms (A2) 26Side chain atoms (A2) 27Water molecules (A2) 36NAD� atoms (A2) 33Sulfate atoms (A2) 49
Ramachandran plotc
Most-favored region (%) 91.4 (A, 90.3; B, 92.6)Additionally allowed regions (%) 8.6 (A, 9.7; B, 7.4)Generously allowed regions (%) 0Disallowed regions (%) 0
a The values in parentheses are for the highest resolution shell.b Rmerge h i|I � Ii|/ h i Ii.c As defined by PROCHECK. The statistics of the Ramachandran plot for monomer A and monomer B are indicated in parentheses.
is to lower the pKa value of Tyr164, while Ser151 assists sheet comprises � strands both from monomer A (�F,�G, and �H) and monomer B (�I) and vice versa (Figurethe proper positioning of the hydroxyl group of the sub-
strate [38, 42]. The �-amino group of the lysine forms 2). The middle part of this � sheet contains a hairpinstructure between � strand �G (Leu236–Gly240) anda hydrogen bond (3.0 A) with the 3�-hydroxyl of the
nicotinamide ribose, an interaction needed to orient the amino acid residues Trp243–Lys246 of � strand �H. The�H strands of both monomers spatially intersect atnicotinamide ring in the syn conformation, to allow B
face 4-pro-S hydride transfer. Trp249, with the side chains of the two tryptophans lyingparallel and 3.9 A apart. The � strand �I of monomer Apairs with the NH2 terminus of �H (Trp243–Gly245) ofNovel COOH-Terminal Domain
The COOH terminus of dh�SCP-2L� forms a separate monomer B and vice versa. The rest of the COOH-termi-nal domain folds into a circle that is composed of threedomain (Figure 3). A database search applying the Dali
server [43] did not reveal any similar structures. Another helices, �CT1 (Thr265–Asn272), �CT2 (Asn272–Cys277),and �H (Ser287–Asp303), and it provides the entrancenovel feature of the architecture is that the additional �

Crystal Structure of the SDR Region of MFE-291
Figure 2. Stereo View of the Binary Structure of dh�SCP-2L�
The structure is drawn as ribbons; monomer A, blue; monomer B, orange. The ligand bound to both monomers is nicotinamide adeninedinucleotide (NAD�). The catalytically active entity is formed by two chemically identical monomers, where the COOH-terminal domain ofmonomer A (C1) interacts with the other monomer in the opposite side of the dimer and vice versa. The arrow indicates the site (Trp249)where the �H � strands from different monomers intersect. The local 2-fold axis runs nearly perpendicular to the long parallel � helicesbetween the monomers at the dimer interface. The figure was made with PyMOL.
(Figure 6) heading toward the amino acid residues criti- COOH-terminal domain in addition to the SDR fold do-main. A database search with Dali [43] for structuralcal for catalysis. The � helix �H of one monomer inter-
acts with the helices �EF and �FG in the other one, with homologs of dh�SCP-2L� gave statistically significantsimilarity (Z scores �3.6) for 48 known structures. Fivethe contact distances ranging from 3.6 A to 4.0 A. Both
�EF and �FG surround the active site pocket, and the of the structures (Protein Data Bank codes 1bdb, 1fds,1b16, 1hu4, and 1dhr), which are members of the SDR�EF contains Ser151 of the catalytic triad. Moreover,
the � turn type IV preceding helix �CT1 could provide an superfamily, have a COOH-terminal extension beyondthe conserved SDR fold. Of these five, only alcohol dehy-interaction site for CoA of a substrate, since it consists of
positively charged amino acid residues Arg258, Lys259, drogenase from Drosophila lebanonensis (Protein DataBank code 1b16) has a short stretch of 11 amino acidand Arg260.residues that interacts with the neighboring subunit [33].The extension in dh�SCP-2L� is much longer, about 60Discussionresidues, with a folding pattern found neither in pre-viously characterized SDR nor in other proteins. Thedh�SCP-2L� described here is the first characterized
two-domain member of the SDR superfamily having a unique COOH-terminal domain in dh�SCP-2L� plays
Figure 3. Detailed Representation of theMonomer in Dimeric dh�SCP-2L�
(A) Folding topologies found in the crystalstructure of dh�SCP-2L�. � helices, circles;� strands, triangles. The cylinders indicatethe shortest � helices in dh�SCP-2L�, whichare not conserved among the SDR proteins.The labeling is done according to the com-monly used nomenclature for the membersof the SDR superfamily [66]. N, amino termi-nus; C, carboxyl terminus.(B) Secondary-structural elements. The arrowindicates a loop found in dehydrogenases ac-cepting long chain (3R )-hydroxyacyl-CoAs assubstrates. � strand �G is located behind the� helix �F. The COOH-terminal amino acidresidues form a separate domain that com-pletes the tunnel heading for the active sitecavity in the other monomer. For clarity, the �
strands are indicated with lowercase letters,and the � helices are indicated with capitalletters.

Structure92
Figure 4. Electron Density around NAD�
The figure shows an Fo � Fc map, contoured at 2.5 �, just before including the NAD�. Ser151, Tyr164, and Lys168 form the catalytic triadneeded in the hydride transfer reaction. In the structure of dh�SCP-2L�, the close contact of Asp40 with the hydroxyl groups of the adenineribose does not leave enough space for the phosphate group in NADP(H). This guides specificity toward NAD(H). The figure was preparedwith Swiss-PdbViewer [67].
multiple roles: it strengthens the interactions between �F, �G, and �H of the other (Figure 2). The � strand �Falso contributes to the central � sheet in the dinucleotidesubunits, forms part of the active site pocket, and pro-
vides putative interaction sites with the CoA moiety of binding site, in such a way that the NH2- and COOH-terminal ends of �F are part of the central and COOH-a ligand.
Most of the SDR proteins are dimeric or tetrameric terminal � sheets, respectively.Among the lipid-metabolizing SDR enzymes, only twoby quaternary structure. The dimeric interface in the
homodimeric SDRs consists of a four-helix bundle, crystal structures have previously been described incomplex with a fatty acyl group, namely, enoyl-ACPwhich is comprised of the long parallel � helices, �E and
�F, of two neighboring subunits [33]. All the structures reductase from Mycobacterium tuberculosis (ProteinData Bank code 1bvr) [44] and type II 3-hydroxyacyl-of homotetrameric SDRs determined so far exhibit two
main subunit interfaces; the four-helix bundle is respon- CoA dehydrogenase/amyloid-� binding alcohol dehy-drogenase (HADH II/ABAD) from rat (Protein Data Banksible for dimerization, and two dimers interact with each
other via their COOH-terminal regions [29, 31]. The di- code 1e3w) [45]. In enoyl-ACP reductase the enoyl tailof trans-2-hexadecenoyl-(N-acetylcysteamine)-thioes-meric interactions in dh�SCP-2L� are more extensive
than in other dimeric SDR enzymes and involve both ter enters the active site from the same direction asNAD�, from the NH2-terminal part of the dinucleotidethe conserved four-helix bundle and a novel COOH-
terminal domain. The dimeric interactions by this COOH- binding domain [44]. In contrast to enoyl-ACP reduc-tase, in HADH II/ABAD, the substrate 3-ketobutyrateterminal domain are accomplished by the antiparallel
two-stranded � sheet, which is composed of the �H reaches the active site through an open space locatednext to the nicotinamide ring [45]. The authors sug-strands of both monomers and �H, which is in contact
with helices �EF and �FG of the neighboring subunit. gested that the unique two-stranded � sheet betweenthe two helices of the four-helix bundle is the interactionAn additional interaction mode between monomers in-
cludes another � sheet, which is comprised of the strand site for a CoA molecule. Comparison of dh�SCP-2L�with these two structures reveals differences that sug-�I in the COOH-terminal domain of one monomer and
Figure 5. Superimposition of the Backbone Structures of the Two Monomers, A and B, in the Crystal Structure of Dimeric dh�SCP-2L�
Monomer A, blue; monomer B, red. NAD(H) is included as a stick model. The two monomers align with an overall rms deviation of 0.21 A, ascalculated from all C� atoms. The arrows indicate the regions that are visible in monomer A, but not in B (Gly51 and Ala196–Glu207). Thedh�SCP-2L� is in the same orientation as that shown in Figure 3.

Crystal Structure of the SDR Region of MFE-293
Figure 6. Interaction of the COOH-TerminalDomain with the Neighboring Monomer in theDimeric dh�SCP-2L�
Negative surface potentials, red; positive sur-face potentials, blue; neutral potentials, white.(A) Entrance of the tunnel, as indicated by thearrow, heading toward the active site cavity.The positive clusters are putative interactionsites for the phosphate groups of a fatty acyl-CoA substrate molecule.(B) Same view as in (A), except that onemonomer is shown as secondary-structuralelements to enable a better view to the inte-rior of the tunnel. The figures were producedwith Swiss-PdbViewer [67].
gest a possibility for a different substrate binding mode. fore, more liganded structures are needed to resolve theproperties behind the stereospecificity and substrate-First, with respect to enoyl-ACP reductase, the loop
between �B and �C in the dh�SCP-2L� structure (Figure interacting regions in the lipid-metabolizing SDR en-zymes.3) is positioned such that the entry from this side would
be blocked. Second, the proposed CoA interaction sitein the HADH II/ABAD structure does not exist indh�SCP-2L�. Therefore, in dh�SCP-2L�, a substrate Biological Implicationscan enter the active site in a different manner, througha novel COOH-terminal domain (Figure 6). In this mode MFE-2 is a peroxisomal multifunctional enzyme that cat-
alyzes the central steps in the � oxidation cycle. It isof interaction, the positive cluster between �I and �CT1in dh�SCP-2L� interacts with the pyrophosphates in already needed at the 13th gestational week in neuronal
migration and, possibly, myelination [47]. In adults,CoA. A salt bridge between a phosphate group and theside chain of a lysine or arginine is one of the few com- MFE-2 also participates in the synthesis of bile acids
and hormone inactivation. Among the single enzymemon features in the binding mode of CoA in otherwisedistinctly different CoA binding proteins [46]. deficiencies, MFE-2 deficiency is very frequent and re-
sembles the peroxisomal assembly disorders with neu-It has been pointed out that dehydrogenases showingthe catalytic preference for short chain fatty acyl-CoAs rological disturbances and an impaired lipid metabolism
as the most striking changes. Some of the inborn errors,lack a glycine-rich loop [11, 14], present in dh�SCP-2L�(Leu41–Ser52) at the bottom of the active site cavity such as deletions and point mutations, have been as-
signed to the dehydrogenase coding region of theclose to bound NAD� (Figures 1 and 3). In the dh�SCP-2L� structure there would not be space for substrates MFE-2 gene [16]. These mutations result in elevated
plasma levels of very long chain fatty acids, pristaniclonger than 16 carbon atoms without interaction withthis loop, one possible factor in the accommodation of acid, and bile acid intermediates. Peroxisomes were
present in the patients’ fibroblasts, although in reducedbulky substrates. However, this loop cannot be the onlydeterminant in the accommodation of a substrate, since number and in larger size. The loss or change of MFE-2
expression in colon, breast, and prostate cancer cellin vitro enzymatic assays, as well as analyses of meta-bolic profiles in patients with MFE-2 deficiency [16] and lines has been demonstrated, revealing, in the case of
colon and breast cancers, allele loss at 5q21, which ismice ablated for the MFE-2 gene [20], indicate a role ofdh�SCP-2L� not only in the oxidation of very long chain the location where the MFE-2 gene has been assigned
[48–51].(3R )-hydroxyacyl-CoA (more than 20 carbon atoms), butalso for the (24R,25R)-stereoisomer of 24-tetrahydroxy- To understand the molecular basis for the diverse
functions of MFE-2, we have crystallized and deter-cholestanoicacyl-CoA.The conformation of the bound NAD� in relation to the mined the three-dimensional structure of the NAD(H)-
dependent dehydrogenase region of MFE-2. The crystalconserved catalytic triad, Ser151, Tyr164, and Lys168, indh�SCP-2L� allows the identification of the reaction structure allows for the rational interpretation of the
physiological effect of mutant enzymes characterizedmechanism as the one that involves B face 4-pro-Shydride transfer, as in other SDR enzymes [38, 42]. In from patients. The reported Gly16Ser [17] and Leu21Phe
[18] mutants (Figure 1) interfere with the binding of NAD�vitro characterization of MFE-2 and/or the NH2-terminaldehydrogenase showed strict stereospecificity, accept- (Figure 7), since serine removes the critical turn and
phenylalanine will occupy the space normally accepteding only the (R ) isomer of fatty acyl-CoAs [2, 5]. Whileshowing the highest catalytic efficiency (kcat/Km) with by NAD�. Val218Leu [19] mutant moves the side chain
of Leu21 in another position, thus affecting the bindingacetoacetyl-CoA and17�-estradiol, HADH II/ABAD,however, additionally catalyzed both (S ) and (R ) isomers of NAD�, as well. Ser177Pro [19] mutant would break
the helical structure and disturb the dimeric interactionsof the 3-hydroxyl substrate, showing a slight preferencetoward (3S )-hydroxybutyrate [45]. Considering the sub- (Figure 7). In addition, the structure describes a loop
region for accommodating substrates of variable sizestrate binding to enoyl-ACP reductase, HADH II/ABAD,and dh�SCP-2L�, it appears that SDR proteins can bind and length and the role of separate COOH-terminal do-
mains for substrate binding and quaternary association.acyl thioester substrates in different manners. There-

Structure94
48 mM Na2HPO4, 22 mM KH2PO4, 9 mM NaCl, 0.4% (w/v) glucose,2 mM MgSO4, 0.1 mM CaCl2, 8 mM (NH4)2SO4, 1� BME vitaminsolution (GIBCO BRL), and 0.5 mM L-SeMet (CalBiochem-NovaBio-chem)], centrifuged again, and resuspended in 20 ml of SeMet-MM.This 20 ml culture was then used as an inoculum for 2 liters ofprewarmed (30C) SeMet-MM medium. The cells were allowed togrow under aerobic conditions until an A600 of 0.9 was reached. Thetemperature was shifted from 37C to 20C, and the expression ofdh�SCP-2L� was induced by the addition of isopropyl-1-thio-�-D-galactopyranoside (IPTG) to a final concentration of 1 mM. After 20hr of additional incubation the bacterial cells were harvested,washed with cold buffer (120 mM NaCl, 16 mM potassium phosphate[pH 7.4], and 0.02% NaN3), and stored at �70C until used.
The bacterial cell pellet (4.6 g wet weight) was resuspended in 50ml of 25 mM Tris, 5 mM ethylene diaminetetraacetic acid (EDTA),50 mM NaCl, and 1 mM ditiothreitol (DTT) (pH 7.8) (buffer A). Afterthe cell wall had been digested with lysozyme (100 �g/ml) for 20min at 22C, the viscosity of the cell lysate was reduced with DNaseI (25 �g/ml) and RNase A (25 �g/ml) in the presence of 10 mM MgCl2.After further 20 min incubation at room temperature, the cell debriswas sedimented by centrifugation (33000 � g, 45 min, 4C). Thesupernatant was applied to an anion exchange Q Sepharose column
Figure 7. Mapping the Location of Amino Acid Residues Whose (2.5 cm � 13 cm; Amersham Pharmacia Biotech AB) equilibratedMutations Have Been Reported to Abolish the (3R)-Hydroxyacyl- with buffer A. The nonbound protein fraction containing dh�SCP-CoA Activity of MFE-2 2L� was concentrated and dialyzed against 20 mM 2-(N-morpho-
lino)ethanesulfonic acid (MES), 30 mM NaCl, 5 mM EDTA, and 1 mMMutations shown are Gly16Ser, Leu21Phe, Ser177Pro, andDTT (pH 6.0) (buffer B) and applied to a cation exchange ResourceVal218Leu [17–19]. Wild-type amino acids, dark gray. The figure wasS column (6 ml; Amersham Pharmacia Biotech AB) equilibrated withproduced with Swiss-PdbViewer [67].the same buffer. The bound proteins were eluted in a NaCl gradientincreasing linearly from 30 mM to 100 mM for 20 column volumesat a flow rate of 1.0 ml/minute, and fractions of 1.0 ml were collected.Experimental ProceduresPeak activity fractions were pooled, concentrated, and applied toa size exclusion chromatography column Superdex 200 HR 10/30Strains(Amersham Pharmacia Biotech AB) and equilibrated with 20 mMAll bacterial manipulations were performed with E. coli strain DH5�.piperazine-N,N�-bis(2-ethanesulfonic acid) (PIPES), 300 mM NaCl,1 mM EDTA, 5 mM DTT, and 1 mM NaN3 (pH 7.0) (buffer C). ProteinsCloning of (3R )-Hydroxyacyl-CoA Dehydrogenasewere eluted at a flow rate of 500 �l/min. The total yield of SeMet-from Rat MFE-2labeled dh�SCP-2L� from 4.6 g of bacterial cells was 1.6 mg.Standard molecular biology methods were performed according to
The native dh�SCP-2L� was overproduced as described [11] andprocedures by Sambrook and coworkers [52]. Total RNA from thepurified as the SeMet-labeled dh�SCP-2L�. Protein purification waslivers of Wistar rats (Laboratory Animal Centre of the University ofmonitored by SDS-PAGE. Electrospray mass spectrometric analy-Oulu, Oulu, Finland) was isolated by the use of a REX total RNAses and enzyme activity measurements were performed on the re-isolation kit in accordance with the manufacturer’s instructionscombinant protein to confirm the identity of dh�SCP-2L�. Both(USB). The open reading frame of rat MFE-2 cDNA was obtainednative dh�SCP-2L� and SeMet-labeled dh�SCP-2L� were analyzedfrom isolated total RNA by reverse transcription with Moloney mu-with electrospray mass spectrometry to determine the degree ofrine leukemia virus reverse transcriptase (Life Technologies), andSeMet labeling. The activity of both dh�SCP-2L� and the full-lengththe coding sequence for (3R )-hydroxyacyl-CoA dehydrogenase wasrecombinant rat MFE-2 were measured in the direction of oxidationamplified by PCR with a 5� primer (cacttccATGGCTTCGCCTCTof (3R )-hydroxyacyl-CoA to 3-ketoacyl-CoA [11].GAGGTTCGACG [lowercase sequences indicate mismatches to the
The SeMet dh�SCP-2L� obtained from the size exclusion columnMFE-2 sequence]) and 3� primer (catctggatcctcaATCTGCAGATGCwas concentrated to 3.9 mg/ml by centrifugal filtration, and TritonCACTTGACCG), which introduced an NcoI site at the 5� end andX-100 (Riedel-de-Haen; product number 56029) and NAD� (Sigma-BamHI at the 3� end (underlined in the primers). The PCR productAldrich) were added to final concentrations of 0.23 mM and 10 mM,was subsequently subcloned into pUC18 vector with the Sure Clonerespectively. Initial crystallization screening was performed by theLigation kit (Amersham Pharmacia Biotech AB), and the nucleotidesparse matrix screening method of Jancarik and Kim [53]. Crystalssequence of inserted DNA was verified to ensure that no mutationswere grown at 21C by the hanging drop vapor diffusion method.had been introduced. The embedded NcoI and BamHI restrictionCrystals suitable for X-ray analysis were obtained after four weeksendonuclease sites in the insert allowed the release of a 968 bpfrom 21% (w/v) polyethyleneglycol 6000, 0.2 M Li2SO4, and 0.1 Mfragment and subsequent cloning into the pET-3d vector (Novagen),Na-Cacodylate (pH 5.5). The triangle-shaped crystal of about 0.4 �which gave a plasmid, pET3d::dh�SCP-2L�, encoding amino acid0.2 � 0.05 mm3 belonged to the monoclinic space group P21 withresidues 1–319 of rat MFE-2. The plasmid was then used for expres-the following cell dimensions: a 89.47, b 82.75, c 95.75, andsion in E. coli B834(DE3) cells (Met-auxotrophic).� 94.20. Assuming four monomers per asymmetric unit, the Vm
value of the crystal is 2.55 A3/Da, which is within the most probableOverproduction, Purification, and Crystallizationrange of values for proteins [54] and also indicates the solventof Selenomethionine-Labeled dh�SCP-2L�content of 51%.E. coli strains of B834(DE3) were made competent for transformation
by the CaCl2 technique [52] and transformed with pET3d::dh�SCP-2L�. Since carbenicillin resistance was the selection marker of the Data Collection and Processing
The dh�SCP-2L� crystal was transferred to cryoprotecting motherstrains, 50 �g/ml carbenicillin was added to all media to maintainselection for cells carrying pET3d::dh�SCP-2L�. After transfor- liquor, which was made by adding glycerol to the original mother
liquor in the presence of Triton X-100 and NAD�, to a final concentra-mation, a single, freshly grown colony of E. coli B834(DE3)(pET3d::dh�SCP-2L�) was picked from an LB plate, placed into 50 tion of 16% (w/v). Glycerol was allowed to diffuse into the crystal
for 5 min before the crystal was flash-frozen at 100 K.ml of LB medium, and grown overnight at 37C. The cells from thisovernight culture were collected by centrifugation (4100 � g, 30 The MAD experiment was performed on beamline BM30 (ESRF,
Grenoble). The X-ray fluorescence from the crystal was measuredmin, RT), washed with 20 ml of SeMet minimal medium [SeMet-MM,

Crystal Structure of the SDR Region of MFE-295
as a function of the incident X-ray energy in the vicinity of the Se tion of the peroxisomal 3-hydroxyacyl-CoA dehydrogenasesfrom rat liver. Relationship between the different dehydroge-� edge. The wavelengths chosen for the data collection were 12,657
eV (�1 0.9796 A) and 12,661 eV (�2 0.9793 A), corresponding to nases and evidence that fatty acids and the C27 bile acids di-and tri-hydroxycoprostanic acids are metabolized by separatethe minimum f� and maximum f″, respectively. A third, remote energy
was selected at 12,749 eV (�3 0.9724 A). The MAD data were multifunctional proteins. Eur. J. Biochem. 240, 660–666.3. Jiang, L.L., Kobayashi, A., Matsuura, H., Fukushima, H., andcollected from a single crystal with a strategy of strict inverse beam
settings [55], 1 rotation per frame, and a MAR345 detector. All three Hashimoto, T. (1996). Purification and properties of human D-3-hydroxyacyl-CoA dehydratase: medium-chain enoyl-CoA hy-wavelengths were collected successively in portions of forty frames,
consisting of 20 frames from φ and 20 frames from φ � 180. dratase is D-3-hydroxyacyl-CoA dehydratase. J. Biochem. 120,624–632.On this source the crystal diffracted beyond 2.0 A resolution, but
practical limitations forced us to compromise it to 2.38 A. Images 4. Leenders, F., Tesdorpf, J.G., Markus, M., Engel, T., Seedorf, U.,and Adamski, J. (1996). Porcine 80-kDa protein reveals intrinsicwere processed with DENZO and reflections were merged with
SCALEPACK from the HKL suite [56]. The final statistics for the 17�-hydroxysteroid dehydrogenase, fatty acyl-CoA-hydratase/dehydrogenase, and sterol transfer activities. J. Biol. Chem.processing of the MAD data set are given in Table 1.271, 5438–5442.
5. Qin, Y.-M., Poutanen, M.H., Helander, H.M., Kvist, A.-P., Siivari,Phasing, Model Building, and RefinementK.M., Schmitz, W., Conzelmann, E., Hellman, U., and Hiltunen,The initial experimental phases were obtained and refined with theJ.K. (1997). Peroxisomal multifunctional enzyme of �-oxidationprogram SOLVE [57] from 12 of the 20 expected selenium atomsmetabolizing D-3-hydroxyacyl-CoA esters in rat liver: molecularper asymmetric unit with data between 20 A and 2.6 A. Maps calcu-cloning, expression and characterization. Biochem. J. 321,lated from these phases, however, were not interpretable. Three21–28.solutions of the self-rotation function, calculated with GLRF [58],
6. Hiltunen, J.K., and Qin, Y.-M. (2000). �-oxidation—strategies forwere found (� 180, φ 0, and � 86; � 180, φ 27, andthe metabolism of a wide variety of acyl-CoA esters. Biochim.� 23; � 180, φ 160, and � 31; peak heights were 10.2 �,Biophys. Acta 1484, 117–128.2.1 �, and 2.0 �, respectively). When applied to the list of heavy-
7. Erdmann, R., and Blobel, G. (1996). Identification of Pex13p, aatom sites obtained from SOLVE, the program FINDNCS [59] wasperoxisomal membrane receptor for the PTS1 recognition fac-able to both confirm the orientation of the noncrystallographic 2-foldtor. J. Cell Biol. 135, 111–121.axis (� 180, φ 0, and � 86) found from the self-rotation
8. Subramani, S. (1996). Protein translocation into peroxisomes.function and to provide the translational component. The phasesJ. Biol. Chem. 271, 32483–32486.were then further improved by the application of phase extension
9. Markus, M., Husen, B., Leenders, F., Jungblut, P.W., Hall, P.F.,from 2.6 A to 2.38 A, 2-fold averaging, and solvent flattening, in aand Adamski, J. (1995). The organelles containing porcinesingle run with the AUTOMASK option of the program DM [60]. Only17�-estradiol dehydrogenase are peroxisomes. Eur. J. Cell Biol.the local 2-fold axis that related two dimers together, covering the68, 263–267.whole asymmetric unit, was used in the DM run. The correlation
10. Adamski, J., Normand, T., Leenders, F., Monte, D., Begue, A.,coefficient between masked map areas before averaging was 0.49,Stehelin, D., Jungblut, P.W., and de Launoit, Y. (1995). Molecularimproving to 0.95 after the last cycle. This allowed the calculationcloning of a novel widely expressed human 80 kDa 17�-hydroxy-of better quality maps, which were used as a starting point forsteroid dehydrogenase IV. Biochem. J. 311, 437–443.autobuilding with MAID [61]. The resulting model was then further
11. Qin, Y.-M., Marttila, M.S., Haapalainen, A.M., Siivari, K.M., Glu-improved with model building in O [62]. Further iterations of refine-moff, T., and Hiltunen, J.K. (1999). Yeast peroxisomal multifunc-ment with CNS [41] against SeMet data collected at the peak (�2),tional enzyme: (3R)-hydroxyacyl-CoA dehydrogenase domainsincluding simulated annealing, resulted in the final model, as de-A and B are required for optimal growth on oleic acid. J. Biol.scribed in Table 1. Refinement was carried out with noncrystallo-Chem. 274, 28619–28625.graphic symmetry (NCS) restraints. Water molecules were added to
12. Leenders, F., Adamski, J., Husen, B., Thole, H.H., and Jungblut,the model, where the difference density exceeded 3 � and the peaksP.W. (1994). Molecular cloning and amino acid sequence of thewere within a reasonable distance to hydrogen bonding partnersporcine 17�-estradiol dehydrogenase. Eur. J. Biochem. 222,on the enzyme (�3.6 A). While water molecules were added, the221–227.NAD� that was bound in each monomer was included in the refine-
13. Novikov, D., Dieuaide-Noubhani, M., Vermeesch, J.R., Fournier,ment steps, as well. Throughout the refinement, the structural integ-B., Mannaerts, G.P., and Van Veldhoven, P.P. (1997). The humanrity was monitored with the WHAT IF [63] and PROCHECK [64]peroxisomal multifunctional protein involved in bile acid synthesis:
programs.activity measurement, deficiency in Zellweger syndrome andchromosome mapping. Biochim. Biophys. Acta 1360, 229–240.
Acknowledgments 14. Qin, Y.-M., Poutanen, M.H., and Novikov, D.K. (2000). Substratespecificities of peroxisomal members of short-chain alcohol
This work was supported by grants from the Academy of Finland and dehydrogenase superfamily: expression and characterizationthe Sigrid Juselius Foundation. We acknowledge ESRF Grenoble for of dehydrogenase part of Candida tropicalis multifunctional en-the opportunity to collect data at beamline BM30. We thank Dr. zyme. J. Lipid Res. 41, 93–98.Michael Merckel for advice on using MAID, Professor Rik Wierenga 15. Qi, C., Zhu, Y., Pan, J., Usuda, N., Maeda, N., Yeldandi, A.V.,for valuable comments during the study, Dr. Lloyd Ruddock for Rao, M.S., Hashimoto, T., and Reddy, J.K. (1999). Absence ofcritical comments on the manuscript, and Eeva-Liisa Stefanius, spontaneous peroxisome proliferation in enoyl-CoA hydratase/Marika Kamps and Ville Ratas for excellent technical assistance. L-3-hydroxyacyl-CoA dehydrogenase-deficient mouse liver. Fur-
ther support for the role of fatty acyl CoA oxidase in PPAR�Received: June 20, 2002 ligand metabolism. J. Biol. Chem. 274, 15775–15780.Revised: October 24, 2002 16. Moller, G., van Grunsven, E.G., Wanders, R.J., and Adamski, J.Accepted: October 28, 2002 (2001). Molecular basis of D-bifunctional protein deficiency.
Mol. Cell. Endocrinol. 171, 61–70.References 17. van Grunsven, E.G., van Berkel, E., Ijlst, L., Vreken, P., de Klerk,
J.B., Adamski, J., Lemonde, H., Clayton, P.T., Cuebas, D.A.,1. Hiltunen, J.K., Wenzel, B., Beyer, A., Erdmann, R., Fossa, A., and Wanders, R.J. (1998). Peroxisomal D-hydroxyacyl-CoA de-
and Kunau, W.H. (1992). Peroxisomal multifunctional �-oxida- hydrogenase deficiency: resolution of the enzyme defect andtion protein of Saccharomyces cerevisiae. Molecular analysis of its molecular basis in bifunctional protein deficiency. Proc. Natl.the fox2 gene and gene product. J. Biol. Chem. 267, 6646–6653. Acad. Sci. USA 95, 2128–2133.
2. Dieuaide-Noubhani, M., Novikov, D., Baumgart, E., Vanhooren, 18. van Grunsven, E.G., van Berkel, E., Mooijer, P.A., Watkins, P.A.,J.C., Fransen, M., Goethals, M., Vandekerckhove, J., Van Veld- Moser, H.W., Suzuki, Y., Jiang, L.L., Hashimoto, T., Hoefler,
G., Adamski, J., et al. (1999). Peroxisomal bifunctional proteinhoven, P.P., and Mannaerts, G.P. (1996). Further characteriza-

Structure96
deficiency revisited: resolution of its true enzymatic and molec- lishes differential effects on short-chain dehydrogenase/reduc-tase reactions. Biochemistry 36, 34–40.ular basis. Am. J. Hum. Genet. 64, 99–107.
19. Paton, B.C., and Pollard, A.N. (2000). Molecular changes in the 38. Benach, J., Atrian, S., Gonzalez-Duarte, R., and Ladenstein,R. (1999). The catalytic reaction and inhibition mechanism ofD-bifunctional protein cDNA sequence in Australasian patients
belonging to the bifunctional protein complementation group. Drosophila alcohol dehydrogenase: observation of an enzyme-bound NAD-ketone adduct at 1.4 A resolution by X-ray crystal-Cell Biochem. Biophys. 32, 247–251.
20. Baes, M., Huyghe, S., Carmeliet, P., Declercq, P.E., Collen, D., lography. J. Mol. Biol. 289, 335–355.39. Dieuaide-Noubhani, M., Novikov, D., Vandekerckhove, J., Veld-Mannaerts, G.P., and van Veldhoven, P.P. (2000). Inactivation
of the peroxisomal multifunctional protein-2 in mice impedes hoven, P.P., and Mannaerts, G.P. (1997). Identification and char-acterization of the 2-enoyl-CoA hydratases involved in peroxi-the degradation of not only 2-methyl-branched fatty acids and
bile acid intermediates but also of very long chain fatty acids. somal �-oxidation in rat liver. Biochem. J. 321, 253–259.40. Rossmann, M.G., Liljas, A., Branden, C.-I., and Banaszak, L.J.J. Biol. Chem. 275, 16329–16336.
21. Jornvall, H., Persson, B., Krook, M., Atrian, S., Gonzalez-Duarte, (1975). In The Enzymes, P.D. Boyer, ed. (New York: AcademicPress), pp. 61–102.R., Jeffery, J., and Ghosh, D. (1995). Short-chain dehydroge-
nases/reductases (SDR). Biochemistry 34, 6003–6013. 41. Brunger, A.T. (1998). Crystallography and NMR system: a new22. Baker, M.E. (1991). Genealogy of regulation of human sex and software system for macromolecular structure determination.
adrenal function, prostaglandin action, snapdragon and petunia Acta Crystallogr. 54, 905–921.flower colors, antibiotics, and nitrogen fixation: functional diver- 42. Winberg, J.O., Brendskag, M.K., Sylte, I., Lindstad, R.I., andsity from two ancestral dehydrogenases. Steroids 56, 354–360. McKinley-McKee, J.S. (1999). The catalytic triad in Drosophila
23. Krozowski, Z. (1992). 11�-hydroxysteroid dehydrogenase and alcohol dehydrogenase: pH, temperature and molecular model-the short-chain alcohol dehydrogenase (SCAD) superfamily. ing studies. J. Mol. Biol. 294, 601–616.Mol. Cell. Endocrinol. 84, 25–31. 43. Holm, L., and Sander, C. (1998). Touring protein fold space with
24. Bailey, T.L., Baker, M.E., and Elkan, C.P. (1997). An artificial Dali/FSSP. Nucleic Acids Res. 26, 316–319.intelligence approach to motif discovery in protein sequences: 44. Rozwarski, D.A., Vilcheze, C., Sugantino, M., Bittman, R., andapplication to steroid dehydrogenases. J. Steroid Biochem. Mol. Sacchettini, J.C. (1999). Crystal structure of the MycobacteriumBiol. 62, 29–44. tuberculosis enoyl-ACP reductase, InhA, in complex with NAD�
25. Oppermann, U.C., Persson, B., Filling, C., and Jornvall, H. (1997). and a C16 fatty acyl substrate. J. Biol. Chem. 274, 15582–15589.Structure-function relationships of SDR hydroxysteroid dehy- 45. Powell, A.J., Read, J.A., Banfield, M.J., Gunn-Moore, F., Yan,drogenases. Adv. Exp. Med. Biol. 414, 403–415. S.D., Lustbader, J., Stern, A.R., Stern, D.M., and Brady, R.L.
26. Jornvall, H., Hoog, J.O., and Persson, B. (1999). SDR and MDR: (2000). Recognition of structurally diverse substrates by typecompleted genome sequences show these protein families to II 3-hydroxyacyl-CoA dehydrogenase (HADH II)/amyloid-betabe large, of old origin, and of complex nature. FEBS Lett. 445, binding alcohol dehydrogenase (ABAD). J. Mol. Biol. 303,261–264. 311–327.
27. Oppermann, U.C., Filling, C., and Jornvall, H. (2001). Forms and 46. Engel, C., and Wierenga, R. (1996). The diverse world of coen-functions of human SDR enzymes. Chem. Biol. Interact. 130– zyme A binding proteins. Curr. Opin. Struct. Biol. 6, 790–797.132, 699–705. 47. Itoh, M., Suzuki, Y., and Takashima, S. (1999). A novel peroxi-
28. Varughese, K.I., Skinner, M.M., Whiteley, J.M., Matthews, D.A., somal enzyme, D-3-hydroxyacyl-CoA dehydratase/D-3-hydroxy-and Xuong, N.H. (1992). Crystal structure of rat liver dihydropter- acyl-CoA dehydrogenase bifunctional protein: its expression inidine reductase. Proc. Natl. Acad. Sci. USA 89, 6080–6084. the developing human brain. Microsc. Res. Tech. 45, 383–388.
29. Ghosh, D., Wawrzak, Z., Weeks, C.M., Duax, W.L., and Erman, 48. Krazeisen, A., Moller, G., Richter, T., Kremmer, E., and Adamski,M. (1994). The refined three-dimensional structure of 3�,20�- J. (1999). Human 17�-hydroxysteroid dehydrogenase type IV:hydroxysteroid dehydrogenase and possible roles of the resi- inhibition by phytoestrogens and loss of expression in breastdues conserved in short-chain dehydrogenases. Structure 2, cancer. J. Mol. Med. 77, 8–13.629–640. 49. Nishisho, I., Nakamura, Y., Miyoshi, Y., Miki, Y., Ando, H., Horii,
30. Ghosh, D., Pletnev, V.Z., Zhu, D.W., Wawrzak, Z., Duax, W.L., A., Koyama, K., Utsunomiya, J., Baba, S., and Hedge, P. (1991).Pangborn, W., Labrie, F., and Lin, S.X. (1995). Structure of hu- Mutations of chromosome 5q21 genes in FAP and colorectalman estrogenic 17�-hydroxysteroid dehydrogenase at 2.20 A cancer patients. Science 253, 665–669.resolution. Structure 3, 503–513. 50. Thomson, A.M., Morris, R.-G., Wallace, M., Wyllie, A.H., Steel,
31. Tanaka, N., Nonaka, T., Nakanishi, M., Deyashiki, Y., Hara, A., C.M., and Carter, D.C. (1993). Allele loss from 5q21 (APC/MCC)and Mitsui, Y. (1996). Crystal structure of the ternary complex of
and 18q21 (DCC) and DCC mRNA expression in breast cancer.mouse lung carbonyl reductase at 1.8 A resolution: the structural
Br. J. Cancer 68, 61–68.origin of coenzyme specificity in the short-chain dehydroge-
51. Castagnetta, L.A., Carruba, G., Traina, A., Granata, O.M., Mar-nase/reductase family. Structure 4, 33–45.
kus, M., Pavone-Macaluso, M., Blomquist, C.H., and Adamski,32. Tanaka, N., Nonaka, T., Tanabe, T., Yoshimoto, T., Tsuru, D.,J. (1997). Expression of different 17�-hydroxysteroid dehydrog-and Mitsui, Y. (1996). Crystal structures of the binary and ternaryenase types and their activities in human prostate cancer cells.complexes of 7�-hydroxysteroid dehydrogenase from Escher-Endocrinology 38, 4876–4882.ichia coli. Biochemistry 35, 7715–7730.
52. Sambrook, J., Fritzch, E.F., and Maniatis, T. (1989). Molecular33. Benach, J., Atrian, S., Gonzalez-Duarte, R., and Ladenstein, R.Cloning: A Laboratory Manual (Cold Spring Harbor, NY: Cold(1998). The refined crystal structure of Drosophila lebanonensisSpring Harbor Laboratory Press).alcohol dehydrogenase at 1.9 A resolution. J. Mol. Biol. 282,
53. Jancarik, J., and Kim, S.-H. (1991). Sparse matrix sampling:383–399.screening method for crystallization of proteins. J. Appl. Crys-34. Ghosh, D., Sawicki, M., Pletnev, V., Erman, M., Ohno, S., Nakajin,tallogr. 24, 409–411.S., and Duax, W.L. (2001). Porcine carbonyl reductase. Struc-
54. Matthews, B.W. (1968). Solvent content of crystals. J. Mol. Biol.tural basis for a functional monomer in short chain dehydroge-33, 491–497.nases/reductases. J. Biol. Chem. 276, 18457–18463.
55. Hendrickson, W.A., and Ogata, C.M. (1997). Phase determina-35. Rossmann, M.G., Moras, D., and Olsen, K.W. (1974). Chemicaltion from multiwavelength anomalous diffraction measurements.and biological evolution of a nucleotide-binding protein. NatureIn Methods in Enzymology Part A, Volume 276, C.W. Carter, Jr.250, 194–199.and R.M. Sweet, eds. (New York: Academic Press), pp 494–523.36. Wierenga, R.K., Maeyer, M.C.H., and Hol, W.G.J. (1985). Interac-
56. Otwinowski, Z., and Minor, W. (1997). Processing of X-ray dif-tion of pyrophosphate moieties with �-helices in dinucleotidefraction data collected in oscillation mode. Methods Enzymol.binding protein. Biochemistry 24, 1346–1357.276, 307–326.37. Oppermann, U.C., Filling, C., Berndt, K.D., Persson, B., Benach,
57. Telwilliger, T.C., and Berendzen, J. (1999). Automated MAD andJ., Ladenstein, R., and Jornvall, H. (1997). Active site directedmutagenesis of 3�/17�-hydroxysteroid dehydrogenase estab- MIR structure solution. Acta Crystallogr. D 55, 849–861.

Crystal Structure of the SDR Region of MFE-297
58. Tong, L., and Rossmann, M.G. (1997). Rotation function calcula-tions with GLRF program. Methods Enzymol. 276, 594–611.
59. Lu, G. (1999). FINDNCS: a program to detect non-crystallogra-phy symmetries in protein crystals from heavy atoms sites. J.Appl. Crystallogr. 32, 365–368.
60. Cowtan, K.D., and Zhang, K.Y. (1999). Density modification formacromolecular phase improvement. Prog. Biophys. Mol. Biol.72, 245–270.
61. Levitt, D.G. (2001). A new software routine that automates thefitting of protein X-ray crystallographic electron-density maps.Acta Crystallogr. D 57, 1013–1019.
62. Jones, T.A., Zou, J.Y., Cowan, S.W., and Kjeldgaard, M. (1991).Improved methods for binding protein models in electron den-sity maps and the location of errors in these models. Acta Crys-tallogr. A 47, 110–119.
63. Vriend, G. (1990). WHAT IF: a molecular modeling and drugdesign program. J. Mol. Graph. 8, 52–56.
64. Laskowski, R.A., McArthur, M.W., Moss, D.S., and Thornton,J.M. (1993). PROCHECK: a program to check the stereochemi-cal quality of protein structures. J. Appl. Crystallogr. 26, 283–291.
65. Hutchinson, E.G., and Thornton, J.M. (1996). PROMOTIF—aprogram to identify and analyze structural motifs in proteins.Protein Sci. 5, 212–220.
66. Ghosh, D., Weeks, C.M., Grochulski, P., Duax, W.L., Erman, M.,Rimsay, R.L., and Orr, J.C. (1991). Three-dimensional structureof holo 3�,20�-hydroxysteroid dehydrogenase: a member of ashort-chain dehydrogenase family. Proc. Natl. Acad. Sci. USA88, 10064–10068.
67. Guex, N., and Peitsch, M.C. (1997). SWISS-MODEL and theSwiss-PdbViewer: an environment for comparative proteinmodeling. Electrophoresis 18, 2714–2723.
Accession Numbers
Atomic coordinates and the structure factors for the binary complexof dh�SCP-2L� have been deposited in the Protein Data Bank underaccession codes 1gz6 and r1gz6sf, respectively.