author query form
TRANSCRIPT

Dear Author,Here are the proofs of your article.
• You can submit your corrections online, via e-mail or by fax.• For online submission please insert your corrections in the online correction form. Always
indicate the line number to which the correction refers.• You can also insert your corrections in the proof PDF and email the annotated PDF.• For fax submission, please ensure that your corrections are clearly legible. Use a fine black
pen and write the correction in the margin, not too close to the edge of the page.• Remember to note the journal title, article number, and your name when sending your
response via e-mail or fax.• Check the metadata sheet to make sure that the header information, especially author names
and the corresponding affiliations are correctly shown.• Check the questions that may have arisen during copy editing and insert your answers/
corrections.• Check that the text is complete and that all figures, tables and their legends are included. Also
check the accuracy of special characters, equations, and electronic supplementary material ifapplicable. If necessary refer to the Edited manuscript.
• The publication of inaccurate data such as dosages and units can have serious consequences.Please take particular care that all such details are correct.
• Please do not make changes that involve only matters of style. We have generally introducedforms that follow the journal’s style.Substantial changes in content, e.g., new results, corrected values, title and authorship are notallowed without the approval of the responsible editor. In such a case, please contact theEditorial Office and return his/her consent together with the proof.
• If we do not receive your corrections within 48 hours, we will send you a reminder.• Your article will be published Online First approximately one week after receipt of your
corrected proofs. This is the official first publication citable with the DOI. Further changesare, therefore, not possible.
• The printed version will follow in a forthcoming issue.
Please noteAfter online publication, subscribers (personal/institutional) to this journal will have access to thecomplete article via the DOI using the URL: http://dx.doi.org/[DOI].If you would like to know when your article has been published online, take advantage of our freealert service. For registration and further information go to: http://www.springerlink.com.Due to the electronic nature of the procedure, the manuscript and the original figures will only bereturned to you on special request. When you return your corrections, please inform us if you wouldlike to have these documents returned.
Metadata of the article that will be visualized in OnlineFirst
ArticleTitle Cells derived from normal or cancer breast tissue exhibit different growth properties when deprived of arginineArticle Sub-Title
Article CopyRight Springer Science+Business Media, LLC(This will be the copyright line in the final PDF)
Journal Name Medical Oncology
Corresponding Author Family Name AlojParticle
Given Name Salvatore M.Suffix
Division Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano”
Organization University of Naples Federico II
Address Naples, 80131, Italy
Division Istituto per la Endocrinologia ed Oncologia Sperimentale, National ResearchCouncil
Organization University of Naples Federico II
Address Naples, Italy
Email [email protected]
Author Family Name ChiavielloParticle
Given Name AngelaSuffix
Division Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano”
Organization University of Naples Federico II
Address Naples, 80131, Italy
Division Istituto per la Endocrinologia ed Oncologia Sperimentale, National ResearchCouncil
Organization University of Naples Federico II
Address Naples, Italy
Email [email protected]
Author Family Name PacielloParticle
Given Name IdaSuffix
Division Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano”
Organization University of Naples Federico II
Address Naples, 80131, Italy
Division Istituto per la Endocrinologia ed Oncologia Sperimentale, National ResearchCouncil
Organization University of Naples Federico II
Address Naples, Italy
Email [email protected]
Author Family Name VenezianiParticle

Given Name Bianca MariaSuffix
Division Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano”
Organization University of Naples Federico II
Address Naples, 80131, Italy
Division Istituto per la Endocrinologia ed Oncologia Sperimentale, National ResearchCouncil
Organization University of Naples Federico II
Address Naples, Italy
Email [email protected]
Author Family Name PalumboParticle
Given Name GiuseppeSuffix
Division Dipartimento di Biologia e Patologia Cellulare e Molecolare “L. Califano”
Organization University of Naples Federico II
Address Naples, 80131, Italy
Division Istituto per la Endocrinologia ed Oncologia Sperimentale, National ResearchCouncil
Organization University of Naples Federico II
Address Naples, Italy
Email [email protected]
Schedule
Received 22 July 2011
Revised
Accepted 28 November 2011
Abstract Arginine deprivation impairs cell proliferation more strong in cancer than in normal cells; thus, it has beenproposed that such an effect could be exploited for cancer therapy. We have compared the effect of argininedeprivation on normal and cancer cells, studying growth rate, morphology, and protein expression patternsin immortalized human MCF10a cells and in MCF7 cells. Arginine deprivation forces MCF10a cells intoirreversible senescence while the vast majority of MCF7 cells become quiescent and resume normal growthfollowing arginine re-addition. Arginine deprivation induced a significant burst of p21cip1 in both cell linesthat were reversible in MCF7 and irreversible in MCF10 cells. In the latter cells, p21cip1 increase wasaccompanied by a time-dependent increase of p16INK4A. Similar effects could be obtained by treating bothcell types with α-difluoro-methyl-ornithine, but not with Nω-hydroxy-L-arginine, drugs that interferespecifically but differently with the major pathways of arginine metabolism. Our data suggest thatderangement in polyamine synthesis is the main consequence of arginine starvation.
Keywords (separated by '-') Breast cancer - Arginine - Senescence - MCF10a - MCF7 - α-Difluoro-methyl-ornithine - Nω-hydroxy-L-arginine
Footnote Information

Author Query Form
Please ensure you fill out your response to the queries raised below
and return this form along with your corrections
Dear Author
During the process of typesetting your article, the following queries have arisen. Please
check your typeset proof carefully against the queries listed below and mark the
necessary changes either directly on the proof/online grid or in the „Author‟s response‟
area provided below
Query Details required Author’s response
1. Please check and confirm that the
authors and their respective affiliations
have been correctly identified and
amend if necessary.
2. Please confirm the section headings are
correctly identified.
3. The term “Discovered Pharmacia
scanner” has been changed as “Discover
Pharmacia scanner.” Please check and
approve.
Journal: 12032
Article: 130

UNCORRECTEDPROOF
ORIGINAL PAPER1
2 Cells derived from normal or cancer breast tissue exhibit different
3 growth properties when deprived of arginine
4 Angela Chiaviello • Ida Paciello • Bianca Maria Veneziani •
5 Giuseppe Palumbo • Salvatore M. Aloj
6 Received: 22 July 2011 / Accepted: 28 November 20117 � Springer Science+Business Media, LLC 2011
8 Abstract Arginine deprivation impairs cell proliferation
9 more strong in cancer than in normal cells; thus, it has been
10 proposed that such an effect could be exploited for cancer
11 therapy. We have compared the effect of arginine depri-
12 vation on normal and cancer cells, studying growth rate,
13 morphology, and protein expression patterns in immortal-
14 ized human MCF10a cells and in MCF7 cells. Arginine
15 deprivation forces MCF10a cells into irreversible senes-
16 cence while the vast majority of MCF7 cells become qui-
17 escent and resume normal growth following arginine
18 re-addition. Arginine deprivation induced a significant
19 burst of p21cip1 in both cell lines that were reversible in
20 MCF7 and irreversible in MCF10 cells. In the latter cells,
21 p21cip1 increase was accompanied by a time-dependent
22 increase of p16INK4A. Similar effects could be obtained by
23treating both cell types with a-difluoro-methyl-ornithine,
24but not with Nx-hydroxy-L-arginine, drugs that interfere
25specifically but differently with the major pathways of
26arginine metabolism. Our data suggest that derangement in
27polyamine synthesis is the main consequence of arginine
28starvation.
29
30Keywords Breast cancer � Arginine � Senescence �
31MCF10a � MCF7 � a-Difluoro-methyl-ornithine �
32Nx-hydroxy-L-arginine
33Introduction
34Several studies have investigated the effects of the amino
35acid arginine on the in vitro proliferation of cells derived
36from normal or cancer tissue. There is a consensus that
37arginine is not an essential amino acid, but it is rated as
38‘‘semi-essential’’ or ‘‘conditionally non-essential’’ amino
39acid. Indeed, arginine could be synthesized from citrulline
40through the sequential action of the cytosolic enzymes
41argininosuccinate synthetase (ASS) and argininosuccinate
42lyase; however, citrulline itself is made, to a large extent
43from arginine, as a by-product of the reaction catalyzed by
44the nitric oxide synthetase (NOS) family of enzymes. Thus,
45it is conceivable that minimal metabolic derangements and/
46or the developmental stage can turn the cell into a strict
47arginine auxotroph.
48In addition to its role as a building block in protein
49synthesis, arginine is the precursor of several metabolic
50pathways leading to the biosynthesis of molecules relevant
51to cell proliferation such as nitric oxide (NO) and poly-
52amines (spermidine, spermine, and putrescine) as well as
53the polyamine analog agmatine. All these molecules are
54relevant to the process of tumorigenesis as shown by
A1 A. Chiaviello � I. Paciello � B. M. Veneziani � G. Palumbo �
A2 S. M. Aloj (&)
A3 Dipartimento di Biologia e Patologia Cellulare e Molecolare
A4 ‘‘L. Califano’’, University of Naples Federico II, 80131 Naples,
A5 Italy
A6 e-mail: [email protected]
A7 A. Chiaviello
A8 e-mail: [email protected]
A9 I. Paciello
A10 e-mail: [email protected]
A11 B. M. Veneziani
A12 e-mail: [email protected]
A13 G. Palumbo
A14 e-mail: [email protected]
A15 A. Chiaviello � I. Paciello � B. M. Veneziani � G. Palumbo �
A16 S. M. Aloj
A17 Istituto per la Endocrinologia ed Oncologia Sperimentale,
A18 National Research Council, University of Naples Federico II,
A19 Naples, Italy
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Med Oncol
DOI 10.1007/s12032-011-0130-7
Au
tho
r P
ro
of

UNCORRECTEDPROOF
55 arginine deprivation [1]. It has been reported that L-argi-
56 nine removal from the culture medium causes different
57 fates spanning from growth arrest to cell death [2]
58 depending, to a large extent, on the level of expression of
59 ASS [1]. Exposing cells to arginase, which causes relative
60 arginine deficiency, induces similar effects [3].
61 Arginine deprivation seems to affect cancer cells more
62 severely, which die more quickly than cells with a non-
63 malignant phenotype [2]; thus, it is not surprising that
64 several attempts have been made to exploit arginine
65 deprivation as an anticancer therapy. However, this
66 approach has not significant success. Indeed, the role
67 played by arginine in cell metabolism and growth is rather
68 complicated by the observation that excess arginine in the
69 diet produces metabolic imbalance. It has been proposed
70 that combining an arginine-rich diet with cancer chemo-
71 therapy has an enhanced therapeutic effect [4]. Szende
72 et al. [5] have reported that D-arginine, administered orally
73 at a daily dose of 500 mg/kg to rats bearing Yoshida’s
74 sarcoma, significantly inhibited tumor growth and intra-
75 peritoneal injection of a similar dose into mice bearing
76 Ehrlich carcinoma inhibited growth of the tumor, albeit not
77 significantly.
78 L-arginine metabolism follows at least two major path-
79 ways: the oxidative deaminase pathway and the NADPH-
80 dependent enzyme nitric oxide synthase (NOS) generating
81 nitric oxide (NO) pathway. The arginase pathway yields
82 ornithine and polyamines that interact with the genomic
83 DNA, regulating its transcription among other effects on
84 proliferation [6]. The availability of intracellular L-arginine
85 is also a rate-limiting factor in nitric oxide (NO) production
86 [7]. NO participates in many cellular metabolic processes,
87 including cell proliferation.
88 In this study, we have attempted to explore the effect of
89 arginine deprivation on the proliferation of two related cell
90 lines: MCF10a and MCF7. MCF10a cells are diploid,
91 normal-like human breast cells that spontaneously became
92 immortal in culture [8]. MCF7 is a well-established human
93 breast tumor cell line that was originally developed in 1973
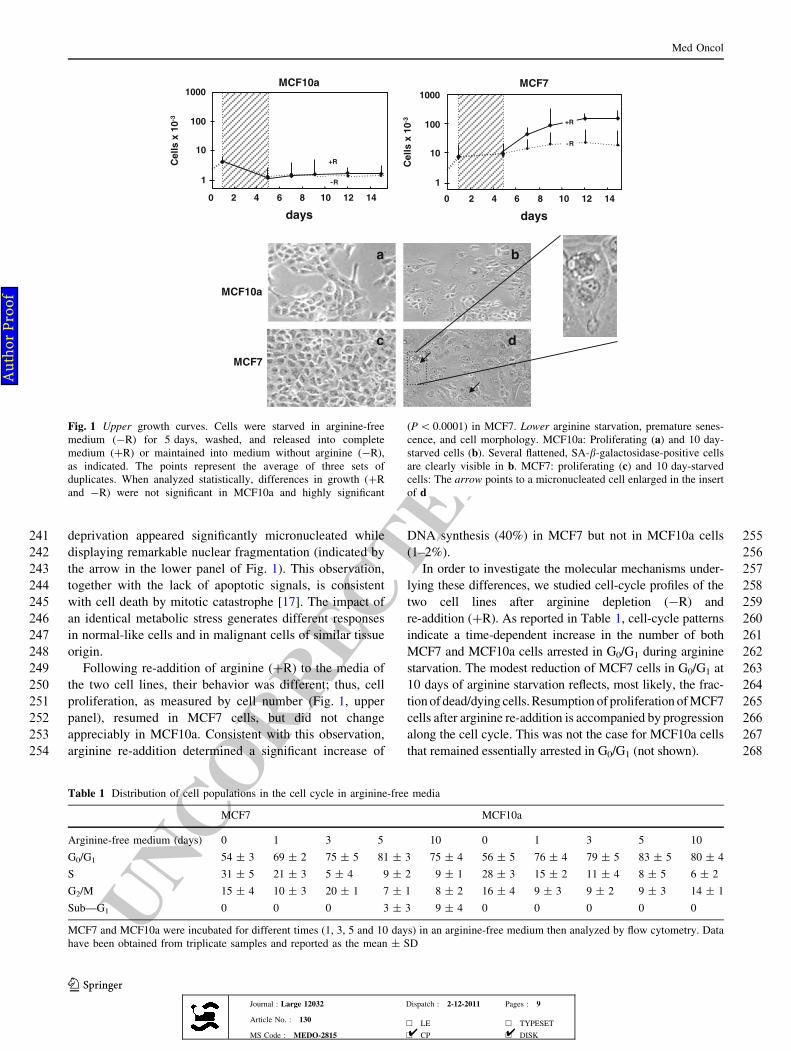
94 [9]. The aim was to determine whether arginine deprivation
95 would have any selective growth inhibition of normal-like,
96 versus malignant cells of similar tissue origin and which of
97 the major pathways of arginine metabolism is affected by
98 such deprivation.
99 Materials and methods
100 Cell cultures and media
101 MCF7 and MCF10a human mammary cell lines were
102 obtained from the American Type Culture Collection
103 (Rockville, MD) and were both grown in the presence of
104L-glutamine (2 mM), penicillin (100 units/mL), strepto-
105mycin (100 lg/mL), and 10% fetal calf serum. The
106chemically modified media were DMEM for MCF7 cells
107and a mixture (1:1) of DMEM and Coon’s modified F12
108medium for MCF10a cells. Both cell lines were p53?/?.
109The MCF7 cells are p16INK4 null. All media and cell cul-
110ture reagents were purchased from Life Technologies
111(Milan, Italy). Arginine-free medium was prepared
112according to Tesseraud et al. [10].
113Drugs
114a-Difluoromethylornithine (DFMO), an inhibitor of ornithine
115decarboxylase (ODC), was supplied by Alexis (UK). DFMO
116solution was prepared in water to obtain 10 mg/mL stock.
117Final concentration of DFMO was 5 mM in all experiments.
118The arginase inhibitor Nx-hydroxy-L-arginine (NOHA)
119was purchased from Calbiochem, UK. It was dissolved in
120water to obtain 5 mg/mL stock solution. Final concentra-
121tion of NOHA was 1 mM in all experiments.
122During incubations with drugs, cell media containing
123either DFMO or NOHA (5 and 1 mM, respectively) were
124replaced every second day.
125Sample treatment schedules
126Experiments performed in arginine-free media
127Usually, 5 9 104 MCF7 or MCF10a cells were seeded in
12860-mm dishes in complete medium. Twenty-four hours
129later, they were washed and incubated in arginine-free
130medium (-R). After 5 days, cells were washed and
131released into fresh complete medium (?R) or maintained
132into arginine-free medium for the needed time.
133Experiments performed in the presence of DFMO
134or NOHA
135The effect 5 mM DFMO or 1 mM NOHA (5 days incu-
136bation) on cell proliferation was performed on 1 9 105
137cells seeded in 6 wells plates.
138Cell-cycle profiles, protein expression, and onset of
139premature senescence were analyzed either in cells incu-
140bated for 5 days with the drugs or after cells released into
141drug-free media.
142The data from triplicate samples are reported as the
143mean ± SD.
144Thymidine incorporation
145[3H]-thymidine incorporation experiments were performed
146in triplicates in 24 multi-well plates according to Love-
147Schimenti et al. [11]. Measured volumes of samples were
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPROOF
148 used to quantify the protein concentrations [12], while
149 appropriate aliquots were employed to measure the effi-
150 ciency of thymidine incorporation by Harvester 96
151 (Skatron Instruments, Lier, Norway).
152 Flow cytometry
153 MCF10a and MCF7 cells (4 9 105) were seeded in 10-cm
154 plates and cultivated in arginine-free media (-R) or with
155 the drugs as reported above. Data have been obtained from
156 triplicate samples and reported as the mean ± SD. Cyto-
157 fluorimetric analyses were done according to Crescenzi
158 et al. [13] at established times. The orange fluorescence of
159 propidium iodide was detected in a linear scale using a
160 CyAn ADP Flow Cytometer (DAKO Cytomation, UK) and
161 Summit Software. About 30,000 events were collected and
162 recorded for each sample.
163 Apoptosis assays
164 Annexin V test: Detection of apoptotic cells with mem-
165 brane alterations (phosphatidylserine translocation) was
166 performed cytofluorimetrically using the Annexin-V-Alexa
167 568 kit (Roche, Milan Italy) according to the manufac-
168 turer’s instructions.
169 Caspases 3 and 7: The activity of Caspase 3 and 7 was
170 monitored by using the Caspase-Glo� 3/7 Assay kit
171 (Promega, Milan Italy).
172 Senescence-associated b-galactosidase (SA-b-Gal)
173 assay
174 Cell senescence was evaluated by assessing senescence-
175 associated-b-galactosidase activity [14]. To measure the
176 sudden appearance of premature senescence in our condi-
177 tions, 7 9 104 (either MCF7 or MCF10a) cells were see-
178 ded, deprived of arginine, or incubated with drugs as
179 reported above. At established times, cells were fixed for
180 5 min (room temperature) in 3% formaldehyde, washed,
181 and incubated at 37�C with fresh senescence-associated-b-
182 Gal-staining solution constituted of 1 mg/mL 5-bromo-4-
183 chloro-3-indolyl b-D-galactoside, 40 mM citric acid/
184 sodium phosphate pH 6.0, 5 mM potassium ferrocyanide,
185 5 mM potassium ferricyanide, 150 mM NaCl, and 2 mM
186 MgCl2. In such conditions, staining was maximal at
187 12–16 h. As expected, all b-galactosidase positive (senes-
188 cent) cells presented flat morphology. However, not all
189 flattened cells were positive to staining.
190 Western blot analysis
191 Cell extracts were obtained as previously described [13]
192 and analyzed by polyacrylamide gels [15]. Molecular
193weight standards were from New England Biolabs
194(Beverly, MA). Separated proteins were blotted onto
195nitrocellulose (Hybond-C pure, Amersham Italy), washed,
196stained with specific primary antibodies (all from Santa
197Cruz, CA), and incubated with horseradish peroxidase-
198conjugated secondary antisera (Biorad, Italy). Filters were
199developed using an electro-chemiluminescent Western
200blotting detection reagent (Amersham, Italy); profiles were
201acquired and quantified by scanning with a Discover
202Pharmacia scanner equipped with a Sun Spark Classic
203Workstation. The anti-Bcl-2 (100SC509), anti-p21cip1
204(C-19), anti-p53 (DO-1), anti-HSP70 (K-20), anti-p16
205INK4A (C-20), and actin (C-2) antibodies were from Santa
206Cruz Biotechnology (Santa Cruz, CA, USA). Anti-tubulin
207(MCA77G) was from Serotec (Kidlington, UK). Gel
208loadings were internally normalized by probing the filters
209with anti-tubulin or anti-actin antibodies.
210Statistical analysis
211All data are expressed as mean ± SD. Significance was
212assessed by the Student’s t test for unpaired data for
213comparisons between two means. Statistical significance
214was defined as *P\ 0.01; **P\ 0.001; ***P\ 0.0001.
215Results and discussion
216Exposure of growing MCF10a cells or MCF7 cells to
217arginine-free medium for up to 5 days caused growth arrest
218in both cell lines (Fig. 1, upper panels). These cells nor-
219mally grow in media containing different levels of arginine
220or arginine precursors (e.g., proline). In fact, MCF10a grow
221in a medium that contains high levels of arginine (about
222250 mg/L and about 15 mg/L of proline), whereas MCF7
223cells, which express high levels ASS [16], require far less
224arginine (about 85 mg/L and no proline). The upper panels
225of Fig. 1 suggest that after 5 days of arginine deprivation,
226MCF10a cells were decreased, and MCF7 slightly aug-
227mented. However, the differences are not significant. A few
228MCF7 cells were dead, as suggested by the appearance of a
229small but reproducible sub-G1 fraction (see below)
230amounting to\3% at 5 days and\10% at 10 days. Cell
231death could be attributed to a non-apoptotic mechanism
232since neither caspase 3 nor caspase 7 were activated. The
233cytofluorimetric Annexin V test, performed at day 5, was
234negative in both cell lines (data not shown).
235Both cell lines showed different, yet profound, mor-
236phological changes (Fig. 1, lower panel): MCF10a cells
237acquired a flat phenotype and became positive for
238SA-b-galactosidase staining, consistent with premature
239senescence, whereas MCF7 cells remained negative to
240SA-b-galactosidase staining. MCF7 cells killed by arginine
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of
UNCORRECTEDPROOF
241 deprivation appeared significantly micronucleated while
242 displaying remarkable nuclear fragmentation (indicated by
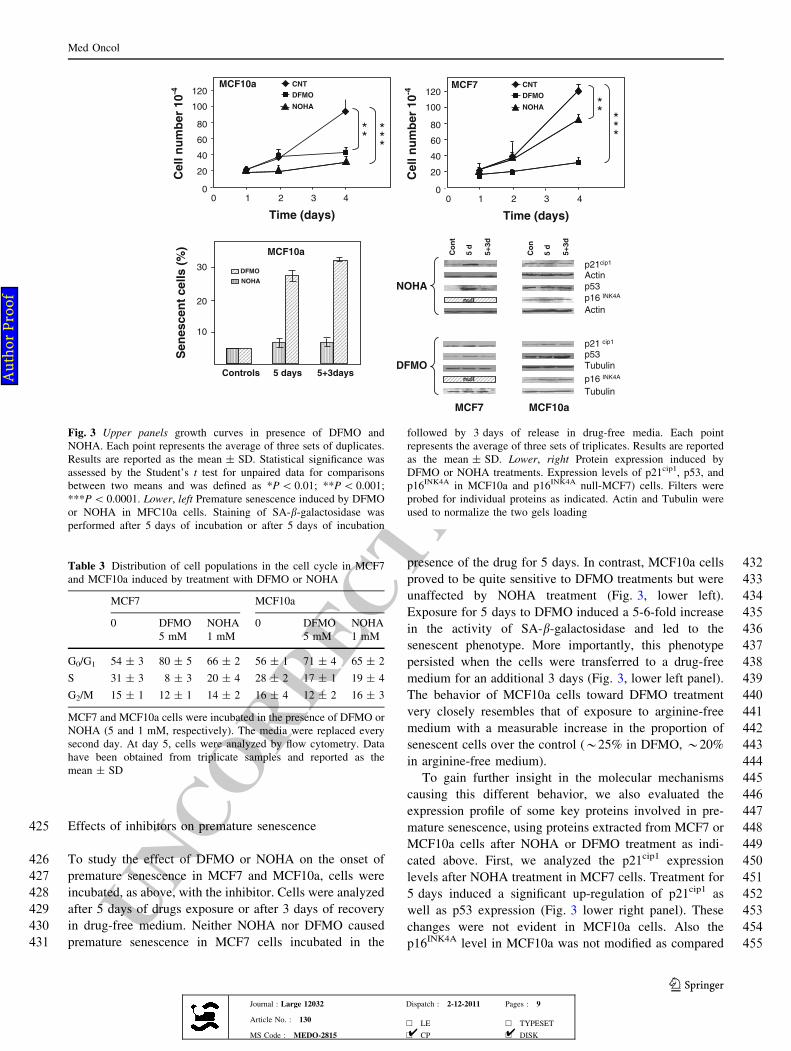
243 the arrow in the lower panel of Fig. 1). This observation,
244 together with the lack of apoptotic signals, is consistent
245 with cell death by mitotic catastrophe [17]. The impact of
246 an identical metabolic stress generates different responses
247 in normal-like cells and in malignant cells of similar tissue
248 origin.
249 Following re-addition of arginine (?R) to the media of
250 the two cell lines, their behavior was different; thus, cell
251 proliferation, as measured by cell number (Fig. 1, upper
252 panel), resumed in MCF7 cells, but did not change
253 appreciably in MCF10a. Consistent with this observation,
254 arginine re-addition determined a significant increase of
255DNA synthesis (40%) in MCF7 but not in MCF10a cells
256(1–2%).
257In order to investigate the molecular mechanisms under-
258lying these differences, we studied cell-cycle profiles of the
259two cell lines after arginine depletion (-R) and
260re-addition (?R). As reported in Table 1, cell-cycle patterns
261indicate a time-dependent increase in the number of both
262MCF7 and MCF10a cells arrested in G0/G1 during arginine
263starvation. The modest reduction of MCF7 cells in G0/G1 at
26410 days of arginine starvation reflects, most likely, the frac-
265tionof dead/dying cells.Resumption of proliferation ofMCF7
266cells after arginine re-addition is accompanied by progression
267along the cell cycle. This was not the case for MCF10a cells
268that remained essentially arrested in G0/G1 (not shown).
0 2 4 6 8 10 12 14
Cell
s x
10
-3
MCF10a
days
1000
100
10
1
+R
-R
0 2 4 6 8 10 12 14
MCF7
Cell
s x
10
-3
days
1000
100
10
1
+R
-R
MCF10a
MCF7
a b
c d
Fig. 1 Upper growth curves. Cells were starved in arginine-free
medium (-R) for 5 days, washed, and released into complete
medium (?R) or maintained into medium without arginine (-R),
as indicated. The points represent the average of three sets of
duplicates. When analyzed statistically, differences in growth (?R
and -R) were not significant in MCF10a and highly significant
(P\ 0.0001) in MCF7. Lower arginine starvation, premature senes-
cence, and cell morphology. MCF10a: Proliferating (a) and 10 day-
starved cells (b). Several flattened, SA-b-galactosidase-positive cells
are clearly visible in b. MCF7: proliferating (c) and 10 day-starved
cells: The arrow points to a micronucleated cell enlarged in the insert
of d
Table 1 Distribution of cell populations in the cell cycle in arginine-free media
MCF7 MCF10a
Arginine-free medium (days) 0 1 3 5 10 0 1 3 5 10
G0/G1 54 ± 3 69 ± 2 75 ± 5 81 ± 3 75 ± 4 56 ± 5 76 ± 4 79 ± 5 83 ± 5 80 ± 4
S 31 ± 5 21 ± 3 5 ± 4 9 ± 2 9 ± 1 28 ± 3 15 ± 2 11 ± 4 8 ± 5 6 ± 2
G2/M 15 ± 4 10 ± 3 20 ± 1 7 ± 1 8 ± 2 16 ± 4 9 ± 3 9 ± 2 9 ± 3 14 ± 1
Sub—G1 0 0 0 3 ± 3 9 ± 4 0 0 0 0 0
MCF7 and MCF10a were incubated for different times (1, 3, 5 and 10 days) in an arginine-free medium then analyzed by flow cytometry. Data
have been obtained from triplicate samples and reported as the mean ± SD
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPROOF
269 Mechanisms of cell-cycle arrest and senescence
270 Differences in the response to arginine deprivation
271 prompted us to investigate the molecular mechanisms
272 triggered in the two cell lines. Attention has been focused
273 on the senescent phenotype, which indicates a major dif-
274 ference between MCF10a and MCF7 cells in response to
275 metabolic stress. DNA-damaging stresses and oncogene
276 activation, as well as agents that induce telomere shorten-
277 ing, can also channel a cell toward a senescence program.
278 This is initiated by the activation of various cell-cycle
279 inhibitors and requires the functions of several proteins,
280 including p53, the CDKN1A gene product WAF1 (also
281 known as p21cip1), the CDKN2A gene product, INK4A
282 (also known as p16INK4A), and others.
283 Analysis of proteins involved in transduction of senes-
284 cence [18] inducing signals has focused on cyclin depen-
285 dent kinase (CDK) inhibitors. The expression of p21cip1 has
286 been measured in MCF10a and MCF7 cells exposed to
287 medium without and with arginine (Fig. 2). Exposure to
288 arginine-free medium (-R) caused a significant burst of
289 p21cip1 in both cell lines, which declined rapidly while
290 arginine starvation continued and growth was arrested.
291 Previous reports showed that during the decline of p21cip1,
292 the expression levels of p16INK4A, another CDK inhibitor,
293 gradually increases in senescent cells assuming, with time,
294 a pivotal role in the maintenance of the senescence pro-
295 gram [18]. Consistent with these observations, the levels of
296 p16INK4A increased in a time-dependent manner in
297 MCF10a cells (Fig. 2). In addition, the finding that the
298 p21cip1 decline could not be reversed is illustrated by
299 the sustained increase of p16INK4A that prevents the loss of
300 the senescent phenotype upon arginine restoration.
301 Protein p21cip1 is a downstream effector of p53. During
302 the first 5 days of starvation, the expression of p53 in
303 MCF10a cells resembles that of p21cip1, with the notable
304 difference that p53 did not disappear at longer times.
305Arginine starvation also induced up-regulation of Bcl-2 in
306MCF10a, which was reversed after restoration of arginine.
307This finding is in line with the fate of MCF10a cells that
308proceed to premature senescence upon arginine starvation
309and is in agreement with recent studies showing that
310senescence may be accompanied by Bcl-2 up-regulation.
311Indeed, Bcl-2 protein expression is increased in naturally
312senescent human fibroblasts as well as in cells in which a
313senescence-like growth arrest is induced by different
314stresses [19]. Finally, we observed that arginine starvation
315caused a slightly delayed expression of heat shock protein
31670 (HSP70), which is known to protect cells against
317apoptosis [20]. Possibly, the sustained over-expression of
318this protein may be considered as an indicator of cell auto-
319protective activity against errors induced by different
320stimuli, including nutrient deficiency. This fact agrees with
321the view that HSP70, a protein that supports p53 stability
322under stress conditions contributes to the recovery of
323damaged proteins [21]. It is noteworthy that the HSP70
324expression, increased by prolonged arginine absence, was
325not reversed by arginine restoration. This is not at all
326surprising since the senescent phenotype of MCF10a cells
327is not reversed by arginine addition to the culture medium.
328In contrast to the MCF10a cells, senescent features did
329not appear in MCF7cells (Fig. 1, panel d and Table 2),
330although they undergo growth arrest when exposed to
331arginine-free medium (-R). Expression profiles of p21cip1
332and p53 diverged from those observed in MCF10a cells in
333that they appear to increase continuously up to 10 days of
334starvation (Fig. 2, right panel). If starvation was limited to
3355 days only, the addition of arginine promptly reversed this
336behavior.
337These results are in line with cytofluorimetric data
338showing that arginine starvation induced a reversible,
339p21cip1-mediated, G0/G1 arrest (Table 1). It is to be noted
340that the effects caused by arginine withdrawal on p16INK4
341null-MCF7 cells are not limited to (reversible) growth
days days
Bcl-2
MCF7
p53
HSP 70
p21 cip1
Actin
- R
3
510510
3
510510
p16 INK4 A
MCF10a
+ R
- R
3
51050+ R 3
51050
null
Actin
Fig. 2 Protein expression
following subtraction (-R) and
re-addition of arginine (?R).
Expression levels of p21cip1,
p53, HSP70 (upper panel),
Bcl-2, and p16INK4A (lower
panel) in MCF10a and MCF7
cells. The rightmost lanes refer
to extracts of cells starved of
arginine for 5 days and
re-exposed to arginine for 3
following days. Actin was used
to normalize gels loading
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPROOF
342 arrest but include also the appearance of a sub-G1 fraction
343 (dead cells and/or fragments thereof) which, albeit small, is
344 measurable and reproducible in MCF7 and not evident in
345 MCF10a. The size of this fraction increases prolonging
346 starvation to 10 days (Table 1). Arginine starvation
347 dependent apoptosis in MCF7 cells appears to be ruled out
348 since Annexin test was negative (not shown). Necrosis
349 could not be ruled out; however, the occurrence of multi-
350 nucleated cells (inset of Fig. 1) would suggest that the
351 small fraction (10% or less of total cells) of damaged
352 MCF7 cells may have undergone mitotic catastrophe [22].
353 a-Difluoromethyl-ornithine (DFMO) and Nx-hydroxy-
354 L-arginine (NOHA) inhibit MCF10a and MCF7 cells
355 proliferation
356 The observation of the effects of arginine depletion reported
357 above prompted us to investigate whether or not these effects
358 could be mimicked by drugs that interfere with the major
359 arginine catabolic pathways summarized in the following
360 simplified scheme:
361
362
Arginine
NOHA
Ornithine
NO
Arginase
NOS NOS
ODCPolyamine
363
364 DFMO is an inhibitor of ornithine decarboxylase (ODC)
365 known to cause cytostatic effects by reducing the cellular
366 polyamines pool [23]. The drug has been also used to treat
367 malignancies and found to be successful only in patients
368 expressing high ODC levels [24].
369 Arginine metabolism may also proceed to NO biosyn-
370 thesis. This biosynthetic pathway in mammals involves the
371 oxidation of arginine to NO and citrulline. This reaction is
372 catalyzed by a particular class of nitric oxide synthases
373(NOS) and requires the formation of an intermediate,
374namely Nx-hydroxy-L-arginine (NOHA). This compound
375negatively regulates arginase activity so that it can inhibit
376cell proliferation of cancer cells characterized by high
377levels of this enzyme. In addition, it has been shown that
378treatment of various tumor cell lines with NOHA causes
379significant cellular effects including arrest of cell cycle,
380changes in mitochondrial permeability, and even cell death
381[25]. Besides the inhibitory action of NOHA on arginase
382activity, this compound potently regulates the production
383of NO since a massive presence of this intermediate may
384exert positive pressure on NO synthesis by mass action.
385We have studied the effects of DFMO and NOHA, on the
386proliferation rates of MCF10a and MCF7 cells. The con-
387centration of DFMO (5 mM) and NOHA (1 mM) has been
388chosen on the basis of our own experience and literature
389data [23, 25]. Both drugs inhibited cell proliferation, albeit
390with minor differences (Fig. 3, upper panels). These data
391were also confirmed by studying cell-cycle profiles. Treat-
392ment with NOHA for 5 days caused G0/G1 arrest in both
393cell lines. The accumulation amounted to *12 and *9%
394over the controls for MCF7 and MCF10a, respectively, and
395was accompanied by a concomitant loss of cells from the S
396phase (about 10% in both cell lines, Table 3).
397DFMO (5 days) appeared a more potent inhibitor of cell-
398cycle progression for both cell lines. Thus, theODC inhibitor
399forced a greater proportion (*25% over control) of MCF7
400cells into the G0/G1 phase while the number of cells in the S
401phase was reduced by a factor *4 (Table 3). Similarly, a
402larger fraction (about 15% over the control) of MCF10 cells
403treated with DFMO was arrested in the G0/G1 phase with a
404concomitant decrease in the S population (Table 3).
405This finding is in agreement with recent data showing
406that DFMO may induce G0/G1 arrest in both normal and
407transformed cells [23]. Switching cells incubated for
4085 days with NOHA to drug-free media for 3 days resulted
409in a full recovery of growth rate for both cell lines. DFMO
410treatment and release, following the identical sequence,
411had a similar effect on MCF7 cells; however, this was not
412the case for MCF10a cells, which were unable to recover
413from the DFMO-induced cell-cycle arrest (not shown).
414Wedid not use a direct inhibitor ofNOproduction, such as
415the L-arginine derivatives NG-nitro-L-arginine (L-NOARG)
416or NG-nitro-L-arginine methyl ester (L-NAME), which have
417been used to inhibit constitutive NO synthase (NOS) in dif-
418ferent biological systems [26]. Indeed, Reisser et al. [27]
419have shown that L-NAME also inhibited the activity of
420arginase of tumor cells in culture. More important, they
421showed that L-NAME inhibition of arginase activity per-
422sisted after treatment ceased. This irreversible effect on
423arginase commands caution when L-NAME is used on the
424assumption that it inhibits only NO synthase.
Table 2 SA-b-galactosidase activity in MCF7 and MCF10a cells
induced by arginine deprivation and re-addition
MCF7 MCF10a
Arginine-free medium
(days)
10 10 10 10
Arginine re-addition
(days)
– 10 – 10
Ability to grow – ? – –
Senescent Cells (%) 4 ± 0.5 1 ± 0.1 20 ± 2.8 18 ± 3.5
Both cell lines were incubated for 10 days in an arginine-free medium
or washed and released into fresh complete medium (?arginine) for
additional 10 days then analyzed for SA-b-galactosidase staining
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of
UNCORRECTEDPROOF
425 Effects of inhibitors on premature senescence
426 To study the effect of DFMO or NOHA on the onset of
427 premature senescence in MCF7 and MCF10a, cells were
428 incubated, as above, with the inhibitor. Cells were analyzed
429 after 5 days of drugs exposure or after 3 days of recovery
430 in drug-free medium. Neither NOHA nor DFMO caused
431 premature senescence in MCF7 cells incubated in the
432presence of the drug for 5 days. In contrast, MCF10a cells
433proved to be quite sensitive to DFMO treatments but were
434unaffected by NOHA treatment (Fig. 3, lower left).
435Exposure for 5 days to DFMO induced a 5-6-fold increase
436in the activity of SA-b-galactosidase and led to the
437senescent phenotype. More importantly, this phenotype
438persisted when the cells were transferred to a drug-free
439medium for an additional 3 days (Fig. 3, lower left panel).
440The behavior of MCF10a cells toward DFMO treatment
441very closely resembles that of exposure to arginine-free
442medium with a measurable increase in the proportion of
443senescent cells over the control (*25% in DFMO, *20%
444in arginine-free medium).
445To gain further insight in the molecular mechanisms
446causing this different behavior, we also evaluated the
447expression profile of some key proteins involved in pre-
448mature senescence, using proteins extracted from MCF7 or
449MCF10a cells after NOHA or DFMO treatment as indi-
450cated above. First, we analyzed the p21cip1 expression
451levels after NOHA treatment in MCF7 cells. Treatment for
4525 days induced a significant up-regulation of p21cip1 as
453well as p53 expression (Fig. 3 lower right panel). These
454changes were not evident in MCF10a cells. Also the
455p16INK4A level in MCF10a was not modified as compared
MCF10a
Time (days)
1 2 3 4
20
40
60
80
100
120
00
Cell
nu
mb
er
10
-4 CNT
DFMO
NOHA
MCF7
Time (days)
1 2 3 4
CNT
DFMO
NOHA
0
20
40
60
80
100
120
0
Cell
nu
mb
er
10
-4
Controls
DFMO
NOHA
5 days 5+3days
MCF10a
10
20
30
**
***
** *
**
Co
nt
5 d
5+
3d
NOHA
MCF10a
p16 INK4A
Actinp21cip1
p53
Actin
null
MCF7
Tubulin
Tubulin
p53
DFMO
p21 cip1
p16 INK4A
Co
n
5 d
5+
3d
null
Se
ne
sc
en
t c
ell
s (
%)
Fig. 3 Upper panels growth curves in presence of DFMO and
NOHA. Each point represents the average of three sets of duplicates.
Results are reported as the mean ± SD. Statistical significance was
assessed by the Student’s t test for unpaired data for comparisons
between two means and was defined as *P\ 0.01; **P\ 0.001;
***P\ 0.0001. Lower, left Premature senescence induced by DFMO
or NOHA in MFC10a cells. Staining of SA-b-galactosidase was
performed after 5 days of incubation or after 5 days of incubation
followed by 3 days of release in drug-free media. Each point
represents the average of three sets of triplicates. Results are reported
as the mean ± SD. Lower, right Protein expression induced by
DFMO or NOHA treatments. Expression levels of p21cip1, p53, and
p16INK4A in MCF10a and p16INK4A null-MCF7) cells. Filters were
probed for individual proteins as indicated. Actin and Tubulin were
used to normalize the two gels loading
Table 3 Distribution of cell populations in the cell cycle in MCF7
and MCF10a induced by treatment with DFMO or NOHA
MCF7 MCF10a
0 DFMO
5 mM
NOHA
1 mM
0 DFMO
5 mM
NOHA
1 mM
G0/G1 54 ± 3 80 ± 5 66 ± 2 56 ± 1 71 ± 4 65 ± 2
S 31 ± 3 8 ± 3 20 ± 4 28 ± 2 17 ± 1 19 ± 4
G2/M 15 ± 1 12 ± 1 14 ± 2 16 ± 4 12 ± 2 16 ± 3
MCF7 and MCF10a cells were incubated in the presence of DFMO or
NOHA (5 and 1 mM, respectively). The media were replaced every
second day. At day 5, cells were analyzed by flow cytometry. Data
have been obtained from triplicate samples and reported as the
mean ± SD
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPROOF
456 to controls (MCF7 are p16INK4A null). It is noteworthy that
457 all changes observed in MCF7 protein expression disap-
458 peared after switching cells to drug-free media for a min-
459 imum of 3 days (Fig. 3, lower right panel). This divergent
460 effect is not easy to explain but can be ascribed to the
461 dissimilar arginase and ODC activities described previ-
462 ously in various cells including breast cancer cells [28].
463 DFMO treatment appeared to have only a moderate
464 effect on p21cip1 and p53 profiles in both MCF7 and
465 MCF10a cells and of p16 INK4A in MCF10a cells. It appears
466 that all changes observed in the normal-like MCF10a cells
467 when cultivated in the absence of arginine could also be
468 obtained by treatment with a specific inhibitor of poly-
469 amine synthesis, suggesting that inhibition of this pathway
470 is responsible for the divergent behavior of normal and
471 transformed cells.
472 In conclusion, we have observed that in two cell lines,
473 whose arginine requirement for normal growth is sub-
474 stantially different, prolonged arginine deprivation causes
475 growth arrest. Re-addition of arginine to the culture med-
476 ium elicits quite different response in the two cell lines;
477 thus, whereas MCF7 cancer cells resume growth, MCF10a
478 cells do not. It is to be noted that for optimal growth
479 MCF10a cells require much higher concentration of argi-
480 nine in the culture medium (see above) than MCF7 cells. It
481 is conceivable that the different dependence upon arginine
482 of the two cell lines stems from the fact that MCF7 cells
483 express high levels of ASS. The observation that pharma-
484 cological inhibition of ODC elicits effects similar to argi-
485 nine deprivation suggests that derangement of polyamine
486 synthesis is the critical arm of arginine metabolism altered
487 in both cell lines.
488 The assumption that arginine deprivation has a strong
489 inhibitory effect on proliferation of cancer cells may not be
490 always justified. In fact, this effect can be less dramatic on
491 cancer cells expressing high levels of ASS (1), which are
492 less dependent upon arginine supply, as well as on cancer
493 cells expressing active estrogen receptors (such as breast
494 MCF7 or lung A549 cells). The presence of estrogen-
495 responsive elements in the promoter region of the bcl-2
496 gene [29] makes these cells refractory to pro-apoptotic
497 stresses [29, 30] in vivo and in vitro when grown in serum-
498 containing media. Although the results of this study refer to
499 cell lines grown in vitro, they may be construed as a caveat
500 if arginine metabolism is considered a target for cancer
501 therapy.
502 Acknowledgments The project was supported by MERIT/MIUR503 project (cod. E61J10000200001) and in part by an Industrial Italian504 Research plan (PON01_02433). We are deeply indebted to doctors505 Jan and Edie Wolff (National Institutes of Health, Bethesda, MD,506 USA) for critical review of the manuscript.
507References
5081. Delage B, Fennel DA, Nicholson L, McNeish I, Lemoine NR,509Crook T, Szolarek PW. Arginine deprivation and argininosucci-510nate synthetase expression in the treatment of cancer. Int J511Cancer. 2010;126:2762–72.5122. Scott L, Lamb J, Smith S, Wheatley DN. Single amino acid513(arginine) deprivation: rapid and selective death of cultured514transformed, malignant cells. Br J Cancer. 2000;83:800–10.5153. Umeda M, Diringer D, Heidelberger C. Inhibition of the growth516of cultured cells by arginase and soluble proteins from mouse517skin. Isr J Med Sci. 1968;4:1216–22.5184. Brittenden J, Heys SD, Eremin O. L-arginine and malignant519disease: a potential therapeutic role? Eur J Surg Oncol. 1994;20:520189–92.5215. Szende B, Tyihak E, Trezl L. Role of arginine and its methylated522derivatives in cancer biology and treatment. Cancer Cell Int.5232001;1:3–7.5246. Pegg AE, McCann PP. Polyamine metabolism and function. Am525J Physiol. 1982;243:C212–21.5267. Mori M, Gotoh T. Regulation of nitric oxide production by527arginine metabolic enzymes. Biochem Biophys Res Commun.5282000;275:715–9.5298. Soule HD, Maloney TM, Wolman SR, Peterso WDJ, Brent R,530McGrath CM, Russo J, Pauley RJ, Jones RF, Brooks SC. Isolation531and characterization of a spontaneously immortalized human532breast epithelial cell line, MCF-10. Cancer Res. 1990;50:6075–86.5339. Soule HD, Vazquez J, Long A, Albert S, Brennan M. A human534cell line from a pleural effusion derived from a breast carcinoma.535J Nat Cancer Inst. 1973;51:1409–16.53610. Tesseraud S, Bigot K, Taouis M. Amino acid availability regu-537lates S6K1 and protein synthesis in avian insulin-insensitive QM7538myoblasts. FEBS Lett. 2003;540:176–80.53911. Love-Schimenti CD, Gibson DF, Ratnam AV, Bikle DD. Anti-540estrogen potentiation of anti-proliferative effects of vitamin D3541analogues in breast cancer cells. Cancer Res. 1996;56:2789–94.54212. Bradford MM. A rapid and sensitive method for the quantitation543of microgram quantities of protein utilizing the principle of544protein-dye binding. Anal Biochem. 1976;72:248–54.54513. Crescenzi E, Chiaviello A, Canti G, Reddi E, Veneziani BM,546Palumbo G. Low doses of cisplatin or gemcitabine plus Photo-547frin/photodynamic therapy: disjointed cell cycle phase-related548activity accounts for synergistic outcome in metastatic non-small549cell lung cancer cells:(H1299). Mol Cancer Ther. 2006;5:776–85.55014. Crescenzi E, Palumbo G, Brady HJ. Roscovitine modulates DNA551repair and senescence: implications for combination chemother-552apy. Clin Cancer Res. 2005;11:8158–71.55315. Laemmli UK. Cleavage of structural proteins during the assembly554of the head of bacteriophage T4. Nature. 1971;227:680–5.55516. Lee JB, Shim YJ, Shin YJ, Jeong SY, Oh J, Park GH, Lee KH,556Min BH. Arginine deiminase enhances MCF-7 cell radiosensi-557tivity by inducing changes in the expression of cell cycle-related558proteins. Mol Cells. 2008;25:305–11.55917. Okada H, Mak TW. Pathways of apoptotic and non-apoptotic560death in tumour cells. Nat Rev Cancer. 2004;4:592–603.56118. Roninson IB, Broude EV, Chang BD. If not apoptosis, then what?562Treatment- induced senescence and mitotic catastrophe in tumor563cells. Drug Resist Updat. 2001;4:303–13.56419. Tombor B, Rundell K, Oltvai ZN. Bcl-2 promotes premature565senescence induced by oncogenic Ras. Biochem Biophys Res566Commun. 2003;303:800–7.56720. Bivik C, Rosdahl I, Ollinger K. Hsp70 protects against UVB568induced apoptosis by preventing release of cathepsins and
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of

UNCORRECTEDPROOF
569 cytochrome c in human melanocytes. Carcinogenesis. 2007;28:570 537–44.571 21. Walerych D, Olszewski MB, Gutkowska M, Helwak A, Zylicz572 M, Zylicz A. Hsp70 molecular chaperones are required to support573 p53 tumor suppressor activity under stress conditions. Oncogene.574 2009;28:4284–94.575 22. Castedo M, Perfettini JL, Roumier T, Andreau K, Medema R,576 Kroemer G. Cell death by mitotic catastrophe: a molecular def-577 inition. Oncogene. 2004;23:2825–37.578 23. Koomoa DL, Yco LP, Borsics T, Wallick CJ, Bachmann AS.579 Ornithine decarboxylase inhibition by alpha-difluor-580 omethylornithine activates opposing signaling pathways via581 phosphorylation of both Akt/protein kinase B and p27Kip1 in582 neuroblastoma. Cancer Res. 2008;68:9825–31.583 24. Shantz LM, Levin VA. Regulation of ornithine decarboxylase584 during oncogenic transformation: mechanisms and therapeutic585 potential. Aminoacids. 2007;33:213–23.586 25. Singh R, Pervin S, Chaudhuri G. Caspase-8-mediated BID587 cleavage and release of mitochondrial cytochrome c during588 Nomega-hydroxy-L-arginine-induced apoptosis in MDA-MB-468589 cells. antagonistic effects of L-ornithine. J Biol Chem. 2002;277:590 37630–6.
59126. Pfeiffer S, Leopold E, Schmidt K, Brunner F, Mayer B. Inhibition592of nitric oxide synthesis by NG-nitro-L-arginine methyl ester593(L-NAME): requirement for bioactivation to the free acid,594NG-nitro-L-arginine. Br J Pharmacol. 1996;118:1433–40.59527. Reisser D, Onier-Cherix N, Jean-Francois J. Arginase activity is596inhibited by l -NAME, both in vitro and in vivo. J Enzyme Inhib597Med Chem. 2002;4:267–70.59828. Hu X, Washington S, Verderame MF, Manni A. Interaction599between polyamines and the mitogen-activated protein kinase600pathway in the regulation of cell cycle variables in breast cancer601cells. Cancer Res. 2005;65:11026–133.60229. Perillo B, Sasso A, Abbondanza C, Palumbo G. 17beta-estradiol603inhibits apoptosis in MCF-7 cells, inducing bcl-2 expression via604two estrogen-responsive elements present in the coding sequence.605Mol Cell Biol. 2000;20:2890–901.60630. Stabile LP, Davis AL, Gubish CT, Hopkins TM, Luketich JD,607Christie N, Finkelstein S, Siegfried JM. Human non-small cell608lung tumors and cells (A549) derived from normal lung express609both estrogen receptor a and b and show biological responses to610estrogen. Cancer Res. 2002;62:2141–50.
611
Med Oncol
123Journal : Large 12032 Dispatch : 2-12-2011 Pages : 9
Article No. : 130h LE h TYPESET
MS Code : MEDO-2815 h CP h DISK4 4
Au
tho
r P
ro
of