attention in neglect and extinction: assessing the degree of correspondence between visual and...
TRANSCRIPT

JOURNAL OF CLINICAL AND EXPERIMENTAL NEUROPSYCHOLOGY2010, 32 (1), 71–80
© 2009 Psychology Press, an imprint of the Taylor & Francis Group, an Informa business
http://www.psypress.com/jcen DOI: 10.1080/13803390902838058
NCEN Attention in neglect and extinction: Assessing the degree of correspondence between visual and auditory
impairments using matched tasks
Neglect in Vision and Audition Doug J. K. Barrett,1,2 A. Mark Edmondson-Jones,2 and Deborah A. Hall2
1Nottingham Trent University, Nottingham, UK2MRC Institute of Hearing Research, University Park, Nottingham, UK
Claims have been made for associated degrees of impairment on both visual and auditory performance in unilat-eral neglect and extinction. Since this evidence is primarily based on different tests in each modality, it is difficultto properly quantify the degree of association between performance in vision and audition. The current studycompares visual and auditory extinction and temporal order judgments (TOJs) in two cases with clinical visualneglect. Stimuli in both modalities were precisely matched in their temporal and spatial parameters. The resultsreveal a mixed pattern of association between different auditory tests and their visual counterparts. This suggeststhat associations between visual and auditory neglect can occur but these are neither obligatory nor pervasive.Instead, our data support models of spatial impairment in neglect and extinction that acknowledge differences inthe contribution of spatial information to performance in each modality in responses to changing task demands.
Keywords: Matched spatial location; Temporal order; Dissociation.
INTRODUCTION
Unilateral neglect is a neurological syndrome associatedwith unilateral (predominantly right) cortical lesions. Itaffects up to two thirds of acute right hemisphere strokepatients (Parton, Malhotra, & Husain, 2004) andpresents as a lack of awareness of objects and events onthe side of space contralateral to the causal lesion (i.e.,on the left side for a patient with right hemisphere dam-age). These symptoms exist despite the absence of basicsensory deficits (e.g., hemianopia) and are often attrib-uted to an impairment of spatial representation and/orselective attention (e.g., E. Bisiach, 1993; Di Pellegrino,Basso, & Frassinetti, 1997; Farah, Wong, Monheit, &Morrow, 1989; Mesulam, 1999; Rorden, Mattingley,Karnath, & Driver, 1997). In some cases, contralesionalneglect only occurs in the presence of a simultaneousipsilesional object. This pattern of impairment, oftencalled extinction, is usually considered part of the neglect
syndrome (Parton et al., 2004). Despite the fact thatneglect and extinction are most commonly described anddiagnosed on the basis of visuospatial behavior, there isgrowing evidence that these deficits can occur in othersensory modalities (e.g., Cusack et al., 2000; Eramudugolla,Irvine, & Mattingley, 2007; Pavani, Husain, Ladavas, &Driver, 2004; Pavani, Ladavas, & Driver, 2003; Spierer,Meuli, & Clarke, 2007). Associations between visual,auditory, and tactile neglect suggest that the syndromereflects a perturbation of multimodal spatial selectionand attention (E. Bisiach et al., 2004; L. Bisiach,Cornacchia, & Vallar, 1984; Bueti, Costantini, Forster, &Aglioti, 2004).
Models ascribing visuospatial deficits in neglect andextinction to changes in the distribution of neural activa-tion representing stimuli presented in ipsilesional and cont-ralesional space provide a theoretical account forspatiotemporal deficits in other modalities (e.g., Cate &Behrmann, 2002; Kinsbourne, 1993; Rizzolati, Scandolara,
This work was supported by a Medical Research Council Career Development Grant awarded to the first author and formed part ofan ongoing program of research at the Medical Research Council’s Institute of Hearing Research, Nottingham, U.K. We would like tothank the consultants and staff at the Queen’s Medical Centre, Nottingham, who provided support as well as to the patients themselvesfor their participation.
Address correspondence to Doug J. K. Barrett, The Division of Psychology, Nottingham Trent University, Nottingham NG1 3LT,UK (E-mail: [email protected]).

72 BARRETT, EDMONDSON-JONES, HALL
Matelli, & Gentilucci, 1981). This has led researchers toextend models of visuospatial neglect and extinction toexplain deficits in audition and touch (e.g., Pavani et al.,2004; Pavani et al., 2003). This generalization is givenfurther support by behavioral (e.g., Driver & Spence,1994; Spence & Driver, 1997; Spence, Lloyd, McGlone,Nicholls, & Driver, 2000), neuroanatomical (seeGhazanfar & Schroeder, 2006, for a recent review), andneuroimaging (Lewis, Beauchamp, & DeYoe, 2000;Macaluso, Frith, & Driver, 2001) evidence and furtherindicates that spatial location and attention play animportant part in multimodal integration.
Despite the above, multimodal accounts of neglect andextinction remain somewhat contentious (Golay, Hauert,Greber, Schnider, & Ptak, 2005; Sinnett, Juncadella,Rafal, Azanon, & Soto-Faraco, 2007; Spierer et al.,2007). A number of studies have found dissociationsbetween visual and auditory attentional biases in patientsdiagnosed with visual neglect (De Renzi, Gentilini, &Barbieri, 1989; Soroker, Calamaro, Glicksohn, &Myslobodsky, 1997; Spierer et al., 2007; Zimmer,Lewald, & Karnath, 2003). These have been interpretedas evidence for selective damage to modality-specificmechanisms of spatial representation (e.g., De Renzi,Gentilini, & Pattachini, 1984; Soroker et al., 1997). Thisassertion emphasizes potential differences in the role ofspatial selection in perceptual systems characterized bydifferent coding principles (e.g., the spatiotopic visualand tonotopic auditory systems) and is consistent withevidence for modality-specific spatial cuing effects invision and audition (McDonald & Ward, 1999; Roberts,Summerfield, & Hall, 2006). An alternative explanationis that the reported dissociations between measures ofattentional bias may, in part, reflect differences in thespecificity and sensitivity of the visual and auditory testsemployed rather than the nature of the underlyingimpairment itself. Given the potential remedial utilityafforded by intact auditory-spatial representation inpatients with visual neglect (Golay et al., 2005), it isimportant to establish the degree of association betweenvisual and auditory impairment when potential measure-ment artifacts are controlled.
Much of the evidence for an association between visualand auditory attentional bias is based on tests that placedifferent demands on performance in each modality (e.g.,Pavani et al., 2004; Pavani, Meneghello, & Ladavas, 2001;Pavani et al., 2003). Neglect assessment typically utilizesclinical “pen and paper” tests that may include line bisec-tion, cancellation, and copying from sample tasks (e.g.,Wilson, Cockburn, & Halligan, 1987). These are presentedat the patient’s bedside, with the patient being given unlim-ited time to view the material and to make a manualresponse. Performance on these tasks draws heavily on vis-uomotor control (Parton et al., 2004). In contrast, auditoryneglect has typically been assessed using tasks in which thepatient is asked to localize briefly presented sounds and tomake either a verbal or a simple manual response that isorthogonal to left and right (e.g., Pavani et al., 2003; Pinek,Duhamel, Cave, & Brouchon, 1989).
Differences in task demands have also been con-founded by differences in the location to which visual
and auditory stimuli are presented. Most auditory taskshave used sounds presented over headphones (Bellmann,Meuli, & Clarke, 2001; Karnath, Zimmer, & Lewald,2002; Zimmer et al., 2003). These manipulate perceivedlocation using binaural cues (interaural timing differ-ences, ITDs, and interaural level differences, ILDs; e.g.,L. Bisiach et al., 1984; Zimmer et al., 2003) or monauralcues. A sound presented to only one ear has an infiniteILD (e.g., Sinnett et al., 2007). This produces soundsthat appear to arise from an intracranial position, a loca-tion for which there is no visual counterpart (Blauert &Lindemann, 1986). Binaural cues also elicit a weakspatial percept to which even normal listeners, withouttraining, can have difficulty orienting attention (Robertset al., 2006). The lack of a spatial coincidence betweenauditory stimuli presented over headphones and visualstimuli presented in extrapersonal space means that stim-uli in either modality arise from different locations. Dif-ferences between binaural and monaural sounds can alsomake it difficult to distinguish between the involvement ofspatial representation on one hand and the perceptiveextinction of information from one ear on the other.Extinction for monaural sounds presented to the left earand for binaural sounds lateralized to the left hemispacehave also been shown to dissociate, suggesting distinctunderlying neural processes in each case (Bellman et al.,2001; Spierer et al., 2007).
An alternative method of sound presentation is to usehead-related transfer functions. These generate virtualspace by simulating the linear distortions in the magni-tude and phase spectrum of the acoustic signal on itspath from the sound source to the eardrums, which arecaused by shadowing, reflection, and diffraction by thehead and outer ear. These filter characteristics areknown to disambiguate front–back confusions andimprove localization accuracy (Wenzel, Arruda, Kistler, &Wightman, 1993). Although this method improves theperception of auditory spatial location for sounds pre-sented over headphones, it is still not ideal as the soundsource is rarely perceived as external unless personal-ized transfer functions are used (Wenzel et al., 1993;Wightman & Kistler, 1989). Even when these or free-field sounds have been used to assess auditory neglect invirtual or real space, comparative visual stimuli haveeither been absent (Bender & Diamond, 1965; Pavani,Lavadas, & Driver, 2002; Soroker et al., 1997) or notmatched for egocentric location. In Eramudugollaet al.’s (2007) study, for example, visual stimuli were pre-sented at eccentricities of ±8.7° while auditory stimuliwere presented at ±90° from fixation. This lack ofexplicit matching for location across visual and auditorytests has implications for the assessment of any multimo-dal spatial metric in neglect and makes quantitative com-parisons across sensory modalities difficult (but seeFrassinetti et al., 2002, for a recent exception).
The current paper presents two case studies that exam-ine the degree of correspondence between visual and audi-tory neglect and extinction using tasks carefully designedto match the extrapersonal spatial and temporal parame-ters of stimuli in each modality. Unlike group studies,the case-by-case approach allows direct comparisons of

NEGLECT IN VISION AND AUDITION 73
unimodal performance that are independent of variabilityintroduced by differences in the site and extent of lesionsamong patients in the sample (Caramazza & Badecker,1989). Dissociations within a single case provide evidencefor the absolute or relative separation of neural processesunderlying individual performance under different testconditions (see Shallice, 1988, for a review of the logicunderlying single-case methodology). In the first task, wereport extinction indices that measure decrements in thedetection of dual compared to single objects as a functionof their egocentric location. To our knowledge, this is thefirst time that detection rates have been compared for vis-ual and auditory stimuli presented at identical locations inthe horizontal meridian (azimuth). In the second task, sen-sitivity to temporal asynchrony between two stimuli ismeasured using a temporal order judgment (TOJ) task.Previous TOJ results have revealed a temporal disadvan-tage for contralesional compared to ipsilesional stimuli (DiPellegrino et al., 1997; Eramudugolla et al., 2007; Karnathet al., 2002; Rorden et al., 1997). Our experiment extendsthese studies by evaluating spatiotemporal judgmentsusing visual and auditory stimuli that are matched pre-cisely in terms of their spatial and temporal characteristics.
METHOD
Participants
Two patients (F.D. & J.B.) were recruited from the strokerehabilitation ward at the Queen’s Medical Centre,Nottingham. F.D. suffered a right middle cerebral artery(MCA) territory infarct resulting in low attenuation inthe region of the superior right frontal lobe, caudate, andlentiform nucleus. J.B. had a hyperacute right MCA ter-ritory infarction with hyperdense right MCA branches.Hypodensity was also observed in the right insula andexternal capsules (radiographer’s reports). Patients wereassessed within one month of hospital admission (14 and31 days) and undertook a battery of clinical tests, includ-ing a star cancellation task, line bisection, and figuredrawing from sample. Each demonstrated a pattern ofvisuospatial performance consistent with a clinical diag-nosis of neglect (see Table 1 for a clinical description foreach patient). Both patients had intact hemifields, andpure-tone audiometry revealed hearing within the rangeof age-matched controls (Davis, 1995).
A total of 15 age-matched controls (mean = 66 years,SD = 2.5 years) were also recruited from the Nottinghamarea. Controls had no history of neurological impairment
and showed no symptoms of neglect assessed using thesame battery of tests as that administered to patients. Allcontrols had intact hemifields and hearing performancewithin the normal range for their age (Davis, 1995).
Stimuli and apparatus
Visual stimuli consisted of two geometric shapes: a redcircle and a blue square. Each was closed and subtended8.3° of visual angle. Auditory stimuli were two digitallycreated vowel sounds: “ah” and “ee.” Vowel pairs wereseparated in their fundamental frequency by four semi-tones so that they could be easily segregated from oneanother (Culling, Summerfield, & Marshall, 1994). Vow-els were presented at 69 dB(A) on single-target trials andat 66 dB(A) on dual-target trials to reduce the sound levelcues that would otherwise distinguish single- from dual-target trials. Before testing, each participant was familiar-ized with the visual and auditory stimuli to ensure theycould accurately identify and discriminate them.
The apparatus (see Figure 1) consisted of five smallBose Cube loudspeakers arranged horizontally at ear
TABLE 1 Patient information and clinical characteristics
Patient Gender Age (years)Time since stroke
onset (days) Visual fieldAuditory screen
(at 4,000 Hz; in dB)Line bisectiona
magnitude deviation (%)Star cancellationb
L/R omissions
F.D. F 85 31 Intact <60 7 16/3J.B. M 62 14 Intact <40 8 26/5
Note. F = female. M = male. L = left. R = right.aLine length = 172 mm; bTotal number of targets = 27 L and 27 R.
Figure 1. Schematic of the experimental apparatus. A projec-tor, screen, and speaker array were used to present visual andauditory stimuli to the same egocentric position. The displayhad five locations equally distributed in the horizontal planeand centered on the participant’s vertical midline. Visual andauditory stimuli were presented in separate blocks.

74 BARRETT, EDMONDSON-JONES, HALL
level. Loudspeakers were mounted on a flat metal frame(height 150 cm, width 150 cm) supported off the floor bytwo metal legs. Each loudspeaker was located at aneccentricity of 0°, 17°, and 34° to either side of the fixa-tion point, providing one position on the vertical mid-line, two in the left hemispace, and two in the right. Asheet of white fabric was stretched across the metal frameto cover the loudspeakers and to prevent any visual cuesabout their position. Participants sat in front of thisscreen at a viewing distance of 1,100 mm. Visual stimuliwere projected from a ceiling-mounted projector (NECWT610) onto the screen at precisely the same positions asthe loudspeakers. Visual targets were the same size as theloudspeakers, and visual and auditory stimuli were pre-sented at exactly the same location in extrapersonalspace. A fixation cross was projected immediately belowthe horizontal midline and remained on the screenthroughout each trial. Stimulus presentation was con-trolled using customized software running on a ViglenPC equipped with a standard graphics card and a Motu24 I/O sound card. Verbal responses were coded andentered into the computer by the experimenter.
Procedure
Ethical approval for the study was granted by the Not-tingham committee of the National Health ResearchEthics Board, and written consent was obtained from allparticipants. Testing comprised the clinical assessmentand the two experimental tasks. Clinical assessmentswere carried out on the ward while experimental taskswere conducted in a sound-attenuated booth at the Ear,Nose, and Throat department. For the patients, assess-ment and experimental tasks were carried out on consec-utive days with cancellation and visual confrontationtasks repeated on the final day of testing. Controls com-pleted all of their testing within a single session in thesound-attenuated booth.
Visual and auditory versions of the extinction and TOJtasks were presented in separate blocks, with the ordercounterbalanced across participants. Once the participantwas ready, trials were initiated by the experimenter andalways started with fixation.
In the extinction task, stimuli were presented singly orin a pair for one second. Single-target trials comprisedan equal number of each stimulus (e.g., “ah” and “ee”).
Dual-target trials always contained one of each stimulus(e.g., a blue square and a red circle) and were always pre-sented to locations separated by an angle of 34° (i.e.,with one loudspeaker position between them). Single tar-gets were presented to each of the five horizontal loca-tions. Dual targets were presented to three spatialconfigurations in which the pair was (a) located in theright hemispace (RH), (b) across the central meridian(CM) or (c) located in the left hemispace (LH; Figure 2).Stimulus type, location, and number of single- and dual-target trials (12 within each spatial configuration) weresampled with equal probability in each task. Trials werepresented in a random order. Participants were asked toreport the number of targets (one or two), but were notrequired to identify them. Responses were, therefore,orthogonal to the location of the stimuli in order tonegate any confounds associated with a spatially medi-ated response bias.
In the TOJ task, the pair of targets (e.g., blue squareand red circle) was always presented at an eccentricity of17° in the left and right hemifields (i.e., separated by afixed angle of 34°). The stimulus onset asynchrony(SOA) between the two targets could take the followingvalues: 1, 50, 100, 150, 200, 250, 300, and 400 ms. Theside and order in which each stimulus type was presentedwere counterbalanced. Six right- and six left-first targettrials occurred at each location for all SOAs. Thismethod of constant stimuli provided a spread of datathat was sufficient to plot individual psychometric func-tions. The offset of the pair of targets was simultaneous,and the first stimulus was always presented for a total ofone second. Both the SOA and the side on which the tar-get first appeared were randomized. Participants indi-cated verbally which of the two targets appeared first byreporting its identity. Again, participants were requiredto report a stimulus attribute (identity) that was orthog-onal to the location to which stimuli were presented(Shore & Spence, 2005).
Analyses: Extinction task
To assess the effect of eccentricity, the proportion ofcorrectly detected single-target trials was entered intocontingency tables for each participant. Trial outcome(0 = incorrect, 1 = correct) was entered as the depend-ent variable and eccentricity (–34°, –17°, 0°, 17°, 34°) as
Figure 2. The left-hand panel illustrates the dual-target conditions in the extinction task. Dual targets were presented to the left hemis-pace (LH), to the right hemispace (RH), and across the central meridian (CM). Single targets were presented to each of the five loca-tions. The visual angles correspond to the five locations at which stimuli were presented. The right-hand panel presents a schematic ofthe temporal order judgment (TOJ) task. Two stimuli were always presented at a fixed eccentricity of ±17°. Stimulus onset asynchronies(SOAs) varied between 0 and 400 ms.
NEGLECT IN VISION AND AUDITION 75
the independent variable. Due to the binary nature of theresponse, the data were analyzed using Somers’ d. This isa test of association between the outcome (frequency ofcorrect responses) and the ordinal independent varia-ble (eccentricity; see Siegel & Castellan, 1988).
To evaluate the ability to detect dual targets comparedto single targets, the data were transformed into anextinction index derived from that of Duncan et al.(1999). An index was calculated separately for LH, CM,and RH configurations by dividing the proportion ofcorrect responses on dual-target trials (PD) by the prod-uct of the proportion of correct responses for the singletrials (PS) at the corresponding two locations: PD/(PSleft × PSright); see Table 2. An index value equal tounity represents equivalent performance on both single-and dual-target trials, whilst an index value of less thanunity reflects relatively worse performance on dual-targettrials. To measure the degree of association between eachpatient’s visual and auditory extinction indices, stimulusmodality only was permuted across all locations. Ran-dom permutations were repeated 1,000,000 times in orderto estimate an empirical distribution for any differencesin the index under the null hypothesis. Reported p-valuesestimate the two-tailed probability that there is no differ-ence in the distribution of visual and auditory indicesover the three spatial configurations. Randomizationtests were also used to compare the patients’ extinctionindices with the observed indices for the control group. Inthis comparison the p-value reflects the two-tailed proba-bility that the patient’s performance is consistent withthat of the controls (see Figure 3).
Analyses: TOJ
The number of correctly named targets at each SOA wasrecoded to represent the proportion of “right-first”responses. These data were then analyzed using a probitanalysis (Finney, 1964), which produces a probabilityestimate for each participant of right-first responses as alinear function of SOA, transformed via a cumulative
standard normal distribution (McCullagh & Nelder,1983). The resulting probability functions were then usedto estimate each participant’s point of subjective simul-taneity (PSS): the point at which right-first and left-firstdecisions are equally probable (i.e., .5). PSS values devi-ating from zero reflect slowed responses for the ipsile-sional (PSS > 0) or contralesional stimulus (PSS < 0).Goodness-of-fit estimates (GoF) for the observed andestimated data for each individual were calculated usingthe deviance statistic (p-values < .05 suggest that thepsychometric function is a poor fit). For one case (J.B.auditory test), a single incorrect response at SOA of300 ms was omitted as an outlier on the basis that it wasgrossly inconsistent with performance at adjacent SOAsand reduced the GoF. The deviance statistic and the 95%confidence intervals around the PSS were calculatedusing Matlab’s “glmfit” routine (Version 7.5.0; Math-Works Inc., 2007; see Collett, 2002, for a description).For each individual, the sharpness of temporal acuitywas also estimated using the just noticeable difference(JND). This is calculated as half of the SOA betweenthe point on the probit function at which 25% and75% of targets were reported as right-first trials(Shore & Spence, 2005). Table 3 reports the statisticsassociated with the individual probit analyses for eachpatient.
RESULTS
Controls
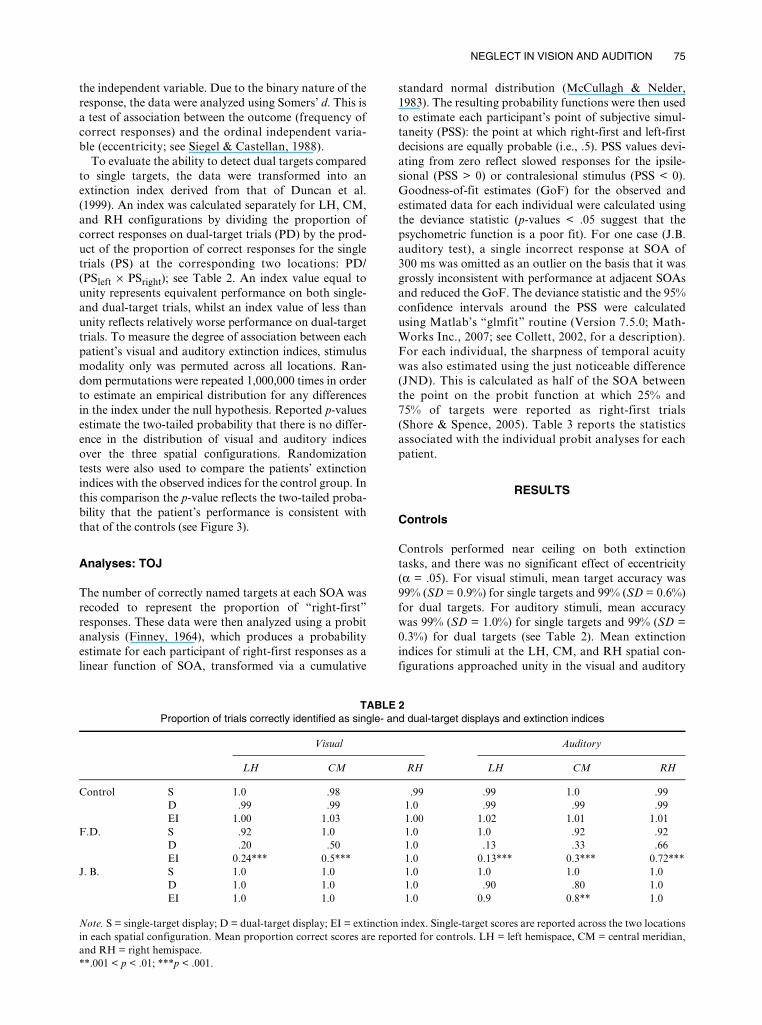
Controls performed near ceiling on both extinctiontasks, and there was no significant effect of eccentricity(α = .05). For visual stimuli, mean target accuracy was99% (SD = 0.9%) for single targets and 99% (SD = 0.6%)for dual targets. For auditory stimuli, mean accuracywas 99% (SD = 1.0%) for single targets and 99% (SD =0.3%) for dual targets (see Table 2). Mean extinctionindices for stimuli at the LH, CM, and RH spatial con-figurations approached unity in the visual and auditory
TABLE 2 Proportion of trials correctly identified as single- and dual-target displays and extinction indices
Visual Auditory
LH CM RH LH CM RH
Control S 1.0 .98 .99 .99 1.0 .99D .99 .99 1.0 .99 .99 .99EI 1.00 1.03 1.00 1.02 1.01 1.01
F.D. S .92 1.0 1.0 1.0 .92 .92D .20 .50 1.0 .13 .33 .66EI 0.24*** 0.5*** 1.0 0.13*** 0.3*** 0.72***
J. B. S 1.0 1.0 1.0 1.0 1.0 1.0D 1.0 1.0 1.0 .90 .80 1.0EI 1.0 1.0 1.0 0.9 0.8** 1.0
Note. S = single-target display; D = dual-target display; EI = extinction index. Single-target scores are reported across the two locationsin each spatial configuration. Mean proportion correct scores are reported for controls. LH = left hemispace, CM = central meridian,and RH = right hemispace.**.001 < p < .01; ***p < .001.
76 BARRETT, EDMONDSON-JONES, HALL
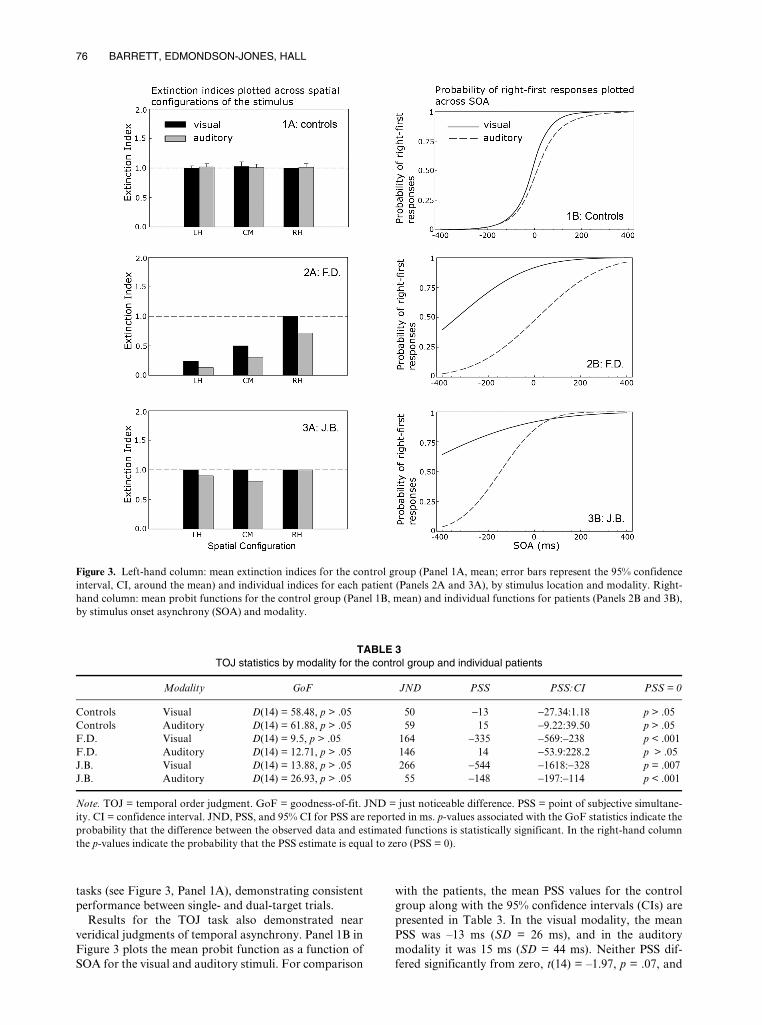
tasks (see Figure 3, Panel 1A), demonstrating consistentperformance between single- and dual-target trials.
Results for the TOJ task also demonstrated nearveridical judgments of temporal asynchrony. Panel 1B inFigure 3 plots the mean probit function as a function ofSOA for the visual and auditory stimuli. For comparison
with the patients, the mean PSS values for the controlgroup along with the 95% confidence intervals (CIs) arepresented in Table 3. In the visual modality, the meanPSS was –13 ms (SD = 26 ms), and in the auditorymodality it was 15 ms (SD = 44 ms). Neither PSS dif-fered significantly from zero, t(14) = –1.97, p = .07, and
Figure 3. Left-hand column: mean extinction indices for the control group (Panel 1A, mean; error bars represent the 95% confidenceinterval, CI, around the mean) and individual indices for each patient (Panels 2A and 3A), by stimulus location and modality. Right-hand column: mean probit functions for the control group (Panel 1B, mean) and individual functions for patients (Panels 2B and 3B),by stimulus onset asynchrony (SOA) and modality.
TABLE 3 TOJ statistics by modality for the control group and individual patients
Modality GoF JND PSS PSS:CI PSS = 0
Controls Visual D(14) = 58.48, p > .05 50 −13 −27.34:1.18 p > .05Controls Auditory D(14) = 61.88, p > .05 59 15 −9.22:39.50 p > .05F.D. Visual D(14) = 9.5, p > .05 164 −335 −569:−238 p < .001F.D. Auditory D(14) = 12.71, p > .05 146 14 −53.9:228.2 p > .05J.B. Visual D(14) = 13.88, p > .05 266 −544 −1618:−328 p = .007J.B. Auditory D(14) = 26.93, p > .05 55 −148 −197:−114 p < .001
Note. TOJ = temporal order judgment. GoF = goodness-of-fit. JND = just noticeable difference. PSS = point of subjective simultane-ity. CI = confidence interval. JND, PSS, and 95% CI for PSS are reported in ms. p-values associated with the GoF statistics indicate theprobability that the difference between the observed data and estimated functions is statistically significant. In the right-hand columnthe p-values indicate the probability that the PSS estimate is equal to zero (PSS = 0).

NEGLECT IN VISION AND AUDITION 77
t(14) = 1.33, p = .20, respectively. The steep slope of theprobit function indicates good temporal acuity with avisual JND of 50 ms and an auditory JND of 59 ms.
Case: F.D.
Table 2 shows that F.D.’s single-target performance inthe extinction task was close to ceiling at all three spatialconfigurations (mean = 97%). No effect of eccentricityupon performance was observed in either the visual orthe auditory modality (p > .1). In contrast, extinctionindices of < 1 revealed a significant reduction in accu-racy on dual- compared to single-target trials on visualand auditory trials (Figure 3, Panel 3A). This pattern isconsistent with extinction in dual-target trials. With theexception of dual visual targets presented to the RH,F.D.’s performance was significantly worse than that ofthe controls at all locations (p < .001), decreasing monot-onically as the stimuli moved from the RH to the LHspatial configurations. Statistical comparisons of visualand auditory extinction indices at each location revealedno difference (p = .15) indicating comparable rates ofextinction in both modalities.
In the TOJ task, F.D. showed a general reduction oftemporal acuity (Figure 3 Panel 3B). Goodness-of-fit forthe observed data and estimated function were accepta-ble (see Table 3). The estimated mean fell outside the95% CI around the control mean. F.D.’s visual PSSrevealed a large bias away from the contralesional target.Visual targets presented on the left had to precede thoseon the right by an estimated 335 ms before their onsetswere judged to be simultaneous. In contrast there was noevidence for an auditory spatiotemporal bias, with theauditory PSS close to that of the controls (14 ms). F.D.’sdata, therefore, reveal a multimodal impairment indetecting simultaneous visual or auditory stimuli in theextinction task and a dissociation between visual andauditory judgments of temporal order. Importantly, thistask by modality interaction occurs for stimuli presentedto identical egocentric locations (i.e., ± 17°).
Case: J.B.
J.B.’s detection accuracy for single targets in the extinc-tion task was 100% (see Table 2). In the visual modality,extinction indices of unity demonstrated equally goodperformance for single- and dual-target trials at all loca-tions (Figure 3, Panel 4A). Auditory extinction indicesfor stimulus pairs presented to LH and CM revealed aslight decrement in dual- compared to single-targetdetection rates at these locations. Although responses tocentrally presented auditory targets were reliably lessaccurate than those of controls (p < .05), visual andauditory performance was statistically consistent at thislocation (p > .2). This patient, therefore, demonstratesno reliable difference in performance across the twomodalities.
In the TOJ task, JND estimates in the visual (266 ms)and auditory (96 ms) tasks showed impoverished tempo-ral acuity compared to controls (Figure 3, Panel 4B). The
probit function on the visual task was particularly shal-low and had to be extrapolated beyond the SOA range inorder to estimate the PSS (–544 ms). In the auditorytask, the deviation of the PSS from zero was muchsmaller (–148 ms), but in the same direction (95% CIsaround the visual and auditory PSS did not overlaprevealing a reliable reduction in auditory compared to vis-ual spatiotemporal bias; see Table 3). Despite this, bothvisual and auditory PSS values fall outside the 95% CIaround the control mean, revealing a multimodal spatio-temporal bias towards the ipsilesional stimulus. J.B’sdata, therefore, reveal a multimodal impairment of tempo-ral order judgments for asynchronous visual and auditorystimuli in the absence of multimodal extinction for the sameobjects when these are simultaneously presented.
DISCUSSION
Research investigating the degree of correspondencebetween visual and auditory attentional bias has pro-duced mixed findings (e.g., De Renzi et al., 1989; Pavaniet al., 2004; Pavani et al., 2003; Sinnett et al., 2007;Soroker et al., 1997; Zimmer et al., 2003). Inconsisten-cies in estimates of the degree of association between vis-ual and auditory neglect and extinction may in part bedue to methodological factors: first, the use of tests thateach afford different measurement sensitivity in eachsensory modality, and, second, the grouping of dataacross individuals who vary in the site and extent of theircausal lesion. The current study addresses these method-ological considerations by comparing visual and audi-tory neglect symptoms separately for two patients usingtests designed explicitly to match spatial and temporalstimulus parameters in each modality. Despite a com-mon clinical diagnosis of visual neglect using pen andpaper tests, both patients showed a unique pattern ofimpairment across two laboratory tasks commonly usedto assess unimodal neglect and extinction. Our resultsreveal within-patient variability in the presence and degreeof association between visual and auditory inattention aswell as highlighting task-dependent variability in the waythese are expressed.
In the extinction task, F.D. showed reduced awarenessof contralesional visual and auditory stimuli when acompeting stimulus was present. This bias increasedacross a leftward spatial gradient that is consistent withgradient models of neglect and extinction (Cate &Behrmann, 2002; Kinsbourne, 1993; Pouget & Driver,2000). This suggests the balance of competition betweenstimuli with a fixed angle of separation is mediated byboth their relative and egocentric locations. The lack ofany significant difference between extinction indices ineach modality suggests that performance in this patientwas affected by a common gradient of inattention. Thisis consistent with a multimodal attentional impairmentfor visual and auditory information represented within acommon spatial metric (e.g., Frassinetti et al., 2002;Pavani et al., 2002, 2003). F.D.’s TOJ data, in contrast,reveal intact auditory performance in the presence of avisual deficit. In the auditory TOJ, F.D. performed

78 BARRETT, EDMONDSON-JONES, HALL
within normal limits, although the shallow function indi-cated a general loss of temporal acuity. In the visualTOJ, F.D.’s data revealed a large spatiotemporal biasagainst the contralesional object. This is consistent withprior entry models of neglect and extinction, whichattribute spatiotemporal deficits to a disruption in thecompetitive interactions between ipsi- and contralesionalstimuli (Di Pellegrino et al., 1997; Karnath et al., 2002;Rorden et al., 1997). F.D.’s data, therefore, reveal anassociation between visual and auditory deficits on onetask (extinction) and a clear dissociation between visualand auditory deficits on another (TOJ). Data for J.B.also reveal a dissociation between the extinction andTOJ tasks. In this patient, intact performance on the vis-ual and auditory extinction tasks contrasts with a multi-modal deficit in the TOJ for the same objects presentedto the same location.
The pattern of dissociations above caution against theassumption that the same test presented in differentmodalities will tap identical attentional resources. Previ-ous studies comparing associations between visual andauditory attentional bias using extinction and TOJ taskshave found conflicting results (e.g., Eramudugolla et al.,2007; Sinnett et al., 2007) and highlight the variabilitytypically observed in comparisons of visual and audi-tory inattention using different measures. The use of vis-ual and auditory stimuli precisely matched in theirspatial and temporal parameters in this study suggeststhat this variation is attributable to the way individualpatterns of impairment disrupt spatial and temporalselection in particular tasks rather than to confoundsassociated with the presentation of stimuli in eachmodality to different (intracranial and extrapersonal)locations (e.g., Sinnett et al., 2007). The strength ofassociation between visual and auditory deficits in ourpatients depends upon whether the presentation of stim-uli is simultaneous or asynchronous. Recent evidencefrom transcranial magnetic stimulation studies hasrevealed modality-specific substrates of visual and audi-tory time perception (Bueti, Bahrami, & Walsh, 2008).Spatial and temporal deficits in neglect and extinctionoften coincide (Becchio & Berone, 2006; Danckert et al.,2007), and it is likely that associations between visualand auditory deficits will reflect the contribution ofthese to task performance in each modality. In audition,temporal asynchronies have been shown to be a power-ful determinant of scene segregation. Common onsetsprovide a cue for grouping while asynchronous onsetspromote stimulus segregation (e.g., Hukin & Darwin,1995). This temporal segregation is thought to occurprior to localization (Hill & Darwin, 1996; Woods &Colburn, 1992) and would have provided a nonspatialmechanism for segregating sounds in the TOJ but notthe extinction task. Asking respondents to report theidentity rather than the position of the first target in theTOJ task might have accentuated this nonspatial effect,distinguishing our data from evidence of an ipsilesionalbias in TOJ for auditory targets identified by their spa-tial location (Eramudugolla et al., 2007; Karnath et al.,2002). Task-dependent variability in the effect of audi-tory spatial cues has previously been reported by
McDonald and Ward (1999) and has been interpreted asevidence for mechanisms of auditory attention thatoperate independently of spatial location. This can becontrasted with the obligatory role of spatial informa-tion in delimiting object boundaries in the visual system(Baylis & Driver, 1993; Kim & Cave, 2001; Kramer,Weber, & Watson, 1997; Lamy & Tsal, 2000) and pro-vides a potential account for the (relative and absolute)dissociation of visual and auditory TOJs in our patients.An important implication of this account is that theassumption of equivalence that has informed much ofthe research investigating visual and auditory atten-tional biases inadequately describes modality-specificvariation in the contribution of spatial information toperformance on measures of neglect and extinction.
In conclusion, the current results suggest that associa-tions between visual and auditory attentional deficits canoccur but these are neither obligatory nor pervasive.Associations varied within patients and across tasks andwere apparent despite the use of visual and auditorystimuli that were precisely matched in their spatial andtemporal parameters. An important implication of thisresult is that measures of association between visual andauditory inattention will vary according to the type oftask involved and the extent that performance in eachmodality is mediated by spatial information. This arguesagainst the characterization of attention as either a uni-modal or a supramodal resource and instead supports amore complex model of task-specific impairment thatmay vary across modalities in neglect. Research hasrevealed both unimodal and multimodal responses tospatial information across a distributed network of corti-cal areas (Bushara, Weeks, Ishii, Catalan, & Tian, 1999;Lewis et al., 2000; Macaluso, Frith, & Driver, 2000),with neurons responding to spatially convergent inputfrom different modalities found in the superior temporaland intraparietal sulci as well as the prefrontal cortices(Beauchamp, 2005; Gifford & Cohen, 2004; Maier, Neu-hoff, Logothetis, & Ghazanfar, 2004). Neglect symptomscan arise from damage to many different cortical andsubcortical regions (Coulthard, Parton, & Husain, 2007;Halligan, Fink, Marshall, & Vallar, 2003; Vallar, 2001),and it is likely that individual patterns of impairment willreflect the contribution of these to different levels of spa-tial description (Ghazanfar & Schroeder, 2006; Karnath,2001; Marshall, Halligan, & Robertson, 1993; Mesulam,1981). Accordingly the type of deficit observed when thisdistributed system is compromised will depend on the siteand extent of the causal lesion as well as the nature of spa-tial representation required by the task. By using matchedtasks to explore the manifestation of these deficits acrossdifferent sensory modalities, confounds associated withdifferent measures of performance can be reduced. Thisapproach supplements evidence from existing investiga-tions of neglect and extinction in order to specify more pre-cisely the way spatial selection operates across differentsensory modalities and tasks.
Original manuscript received 2 December 2008Revised manuscript accepted 19 February 2009
First published online 29 May 2009

NEGLECT IN VISION AND AUDITION 79
REFERENCES
Baylis, G. C., & Driver, J. (1993). Visual attention and objects:Evidence for a hierarchical coding of location. Journal ofExperimental Psychology: Human Perception and Perform-ance, 19, 451–470.
Beauchamp, M. S. (2005). See me, hear me touch me: Multisen-sory integration in lateral occipital-temporal cortex. CurrentOpinion in Neurobiology, 15, 145–153.
Becchio, C., & Berone, C. (2006). Abnormal temporal dynamicsin unilateral spatial neglect. Neuropsychologia, 44, 2775–2782.
Bellman, A., Meuli, R., & Clarke, S. (2001). Two types of audi-tory neglect. Brain, 124, 676–687.
Bender, M. B., & Diamond, S. P. (1965). An analysis ofauditory perceptual defects with observations on thelocalization of the dysfunction. Brain, 88, 675–686.
Bisiach, E. (1993). Mental representation in unilateral neglectand related disorders: The twentieth Bartlett Memorial Lec-ture. Quarterly Journal of Experimental Psychology, 46A,435–461.
Bisiach, E., McIntosh, R. D., Dijkerman, C., McClements, K. I.,Colombo, M., & Milner, D. (2004). Visual and tactile lengthmatching in spatial neglect. Cortex, 40, 651–657.
Bisiach, L., Cornacchia, R., & Vallar, G. (1984). Disorders ofperceived auditory lateralisation after lesions of the righthemisphere. Brain, 107, 37–52.
Blauert, J., & Lindemann, W. (1986). Spatial mapping of intrac-ranial auditory events for various degrees of interauralcoherence. Journal of the Acoustic Society of America, 79,806–813.
Bueti, D., Costantini, M., Forster, B., & Aglioti, S. M. (2004).Uni- and cross-modal temporal modulation of tactile extinc-tion in right brain damaged patients. Neuropsychologia, 42,1689–1696.
Bueti, D., Bahrami, B., & Walsh, V. (2008a). Sensory andassociation cortex in time perception. Journal of CognitiveNeuroscience, 20, 1045–1062.
Bushara, K. O., Weeks, R. A., Ishii, K., Catalan, M.-J., & Tian, B.(1999). Modality specific frontal and parietal areas forauditory and visual spatial localization in humans. NatureNeuroscience, 2, 759–765.
Caramazza, A., & Badecker, W. (1989). Patient classification inneuropsychological research. Brain and Cognition, 10, 256–295.
Cate, A., & Behrmann, M. (2002). Spatial and temporal influ-ences on extinction. Neuropsychologia, 40, 2206–2225.
Collett, D. (2002). Modeling binary data (2nd ed.). Boca Raton,FL: Chapman & Hall/CRC Press.
Coulthard, E., Parton, A., & Husain, M. (2007). The modulararchitecture of the neglect syndrome: Implications for actioncontrol in visual neglect. Neuropsychologia, 45, 1982–1984.
Culling, J. F., Summerfield, A. Q., & Marshall, D. H. (1994).Effects of simulated reverberation on the use of binauralcues and fundamental-frequency differences for separatingconcurrent vowels. Speech Communication, 14, 71–95.
Cusack, R., Carlyon, R. P., & Robertson, I. H. (2000). Neglectbetween but not within auditory objects. Journal of Cogni-tive Neuroscience, 12, 1056–1065.
Danckert, J., Ferber, S., Pun, C., Broderick, C., Stiemer, C., Rock,S., et al. (2007). Neglected time: Impaired temporal percep-tion of multisecond intervals in unilateral neglect. Journal ofCognitive Neuroscience, 19, 1706–1720.
Davis, A. (1995). Hearing in adults: The prevalence and distribu-tion of hearing impairment and reporting hearing disability inthe MRC Institute of Hearing Research’s national study ofhearing. London: Whurr Publishers.
De Renzi, E., Gentilini, M., & Barbieri, C. (1989). Auditoryneglect. Journal of Neurology, Neurosurgery and Psychiatry,52, 613–617.
De Renzi, E., Gentilini, M., & Pattachini, F. (1984). Auditoryextinction following right hemisphere damage. Neuropsych-ologia, 22, 733–744.
Di Pellegrino, G., Basso, G., & Frassinetti, F. (1997). Spatialextinction on double asynchronous stimulation. Neuropsy-chologia, 35, 1215–1223.
Driver, J., & Spence, C. J. (1994). Spatial synergies betweenauditory and visual attention. In T. Inui & J. L. McClelland(Eds.), Attention and Performance XVI: Information integra-tion in perception and communication (pp. 209–235).Cambridge, MA: MIT Press.
Duncan, J., Bundesen, C., Olson, A., Humphreys, G., Chavda,S., & Shibuya, H. (1999). Systematic analysis of deficits invisual attention. Journal of Experimental Psychology:General, 128, 450–478.
Eramudugolla, R., Irvine, D. R. F., & Mattingley, J. B.(2007). Association of auditory and visual symptoms ofunilateral neglect. Neuropsychologia, 45, 2631–2637.
Farah, M. J., Wong, A. B., Monheit, M. A., & Morrow, L. A.(1989). Parietal lobe mechanisms of spatial attention: Modal-ity specific or supramodal? Neuropsychologia, 27, 461–470.
Finney, D. J. (1964). Probit analysis: Statistical treatment of thesigmoid curve. London: Cambridge University Press.
Frassinetti, F., Pavani, F., & Ladavas, E. (2002). Acousticalvision of neglected stimuli: Interaction among spatiallyconverging audiovisual inputs in neglect patients. Journalof Cognitive Neuroscience, 14, 62–69.
Ghazanfar, A. A., & Schroeder, C. E. (2006). Is neocortexessentially multisensory? Trends in Cognitive Sciences, 10,276–285.
Gifford, G. W., & Cohen, Y. E. (2004). Effect of a central fixa-tion light on auditory responses in area LIP. Journal ofNeurophysiology, 91, 2929–2933.
Golay, L., Hauert, C. A., Greber, C., Schnider., A., & Ptak, R.(2005). Dynamic modulation of visual detection by auditorycues in spatial neglect. Neuropsychologia, 43, 1258–1265.
Halligan, P. W., Fink, G. R., Marshall, J. C., & Vallar, G.(2003). Spatial cognition: Evidence from visual neglect.Trends in Cognitive Sciences, 7, 125–133.
Hill, N. I., & Darwin, C. J. (1996). Lateralisation of a perturbedharmonic: Effects of onset asynchrony and mistuning. Jour-nal of the Acoustical Society of America, 100, 2352–2364.
Hukin, R. W., & Darwin, C. J. (1995). Comparison of the effectof onset asynchrony on auditory grouping in pitch matchingand vowel identification. Perception and Psychophysics, 52,91–196.
Karnath, H.-O. (2001). New insights into the functions of thesuperior temporal cortex. Nature Neuroscience, 2, 568–576.
Karnath, H.-O., Zimmer, U., & Lewald, J. (2002). Impairedperception of temporal order in auditory extinction. Neu-ropsychologia, 40, 1977–1982.
Kim, M.-S., & Cave, K. R. (2001). Perceptual grouping via spa-tial selection in a focussed attention task. Vision Research,41, 611–624.
Kinsbourne, M. (1993). Orientation bias model of unilateralneglect: Evidence from attentional gradients within hemis-pace. In I. H. M. Robertson (Ed.), Unilateral neglect, clinicaland experimental studies (pp. 63–86). Hove, UK: PsychologyPress.
Kramer, A. F., Weber, T. A., & Watson, S. E. (1997). Object-based attentional selection—grouped arrays or spatially invar-iant representations? Comment on Vecera and Farah (1994).Journal of Experimental Psychology: General, 126, 3–13.
Lamy, D., & Tsal, Y. (2000). Object features, object locationsand object files: Which does selective attention activate andwhen? Journal of Experimental Psychology: Human Percep-tion and Performance, 26, 1387–1400.
Lewis, J. W., Beauchamp, M. S., & DeYoe, E. A. (2000). Acomparison of visual and auditory motion processing inhuman cerebral cortex. Cerebral Cortex, 10, 873–888.
Macaluso, E., Frith, C. D., & Driver, J. (2000). Selective spatialattention in vision and touch: Unimodal and multimodalmechanisms revealed by PET. Journal of Neurophysiology,63, 3062–3085.
Macaluso, E., Frith, C. D., & Driver, J. (2001). Multimodalmechanisms of attention related to rates of spatial shifting invision and touch. Experimental Brain Research, 137, 445–454.
Maier, J. X., Neuhoff, J. G., Logothetis, N. K., &Ghazanfar, A. A. (2004). Multisensory integration oflooming signals in rhesus monkeys. Neuron, 43, 177–181.

80 BARRETT, EDMONDSON-JONES, HALL
Marshall, J. C., Halligan, P. W., & Robertson, I. H. (1993).Contemporary theories of unilateral neglect: A criticalreview. In I. H., Robertson & J. C. Marshall (Eds.), Unilat-eral neglect: Clinical and experimental studies (pp. 311–329).Hove, UK: Psychology Press.
McCullagh, P., & Nelder, J. A. (1983). Generalized linearmodels. London: Chapman & Hall.
McDonald, J. B., & Ward, M. (1999). Spatial relevance deter-mines facilitatory and inhibitory effects of auditory covertspatial orienting. Journal of Experimental Psychology:Human Perception and Performance, 25, 1234–1252.
Mesulam, M. M. (1981). A cortical network for directedattention and unilateral neglect. Annals of Neurology, 10,309–325.
Mesulam, M. M. (1999). Spatial attention and neglect: Parietal,frontal and cingulated contributions to mental representa-tion and attentional targeting of salient extrapersonalevents. Philosophical Transactions of the Royal Society ofLondon. Series B, Biological Sciences, 354, 1325–1346.
Parton, A., Malhotra, P., & Husain, M. (2004). Hemispatialneglect. Journal of Neurosurgery and Psychiatry, 75, 13–21.
Pavani, F., Husain, M., Ladavas, E., & Driver, J. (2004). Audi-tory deficits in visuospatial neglect patients. Cortex, 40,347–365.
Pavani, F., Meneghello, F., & Ladavas, E. (2001). Deficit ofauditory space perception in patients with visuospatialneglect. Neuropsychologia, 39, 1401–1409.
Pavani, F., Lavadas, E., & Driver, J. (2002). Selective deficit ofauditory localisation in patients with visuospatial neglect.Neuropsychologia, 40, 291–301.
Pavani, F., Ladavas, E., & Driver, J. (2003). Auditory and mul-tisensory aspects of visuospatial neglect. Trends in CognitiveSciences, 7, 407–414.
Pinek, B., Duhamel, J., Cave, C., & Brouchon, M. (1989).Audiospatial deficits in humans: Differential effects associ-ated with left versus right hemisphere parietal damage.Cortex, 25, 175–186.
Pouget, A., & Driver, J. (2000). Object-centered visual neglect,or relative egocentric neglect? Journal of Cognitive Neuro-science, 12, 542–545.
Rizzolatti, G., Scandolara, C., Matelli, M., & Gentilucci, M.(1981). Afferent properties of periarcuate neurons inmacaque monkeys. Behavioural Brain Research, 2, 147–163.
Roberts, K. L., Summerfield, A. Q., & Hall, D. A. (2006).Presentation modality influences behavioural measures ofalerting, orienting, and executive control. Journal of theInternational Neuropsychological Society, 12, 485–492.
Rorden, C., Mattingley, J. B., Karnath, O.-H., & Driver, J.(1997). Visual extinction and prior entry: Impaired percep-tion of temporal order with intact motor perception afterunilateral parietal damage. Neuropsychologia, 35, 421–433.
Shallice, T. (1988). From neuropsychology to mental structure.Cambridge, UK: Cambridge University Press.
Shore, D. I., & Spence, C. S. (2005). Prior entry. In L. Itti, G.Rees, & J. K. Tsotsos (Eds.), Neurobiology of attention(pp. 89–95). Burlington, MA: Elsevier Academic Press.
Siegel, S., & Castellan, N. J. (1988). Nonparametric statistics forthe behavioral sciences. New York: McGraw-Hill.
Sinnett, S., Juncadella, M., Rafal, R. D., Azanon, E., & Soto-Faraco, S. (2007). A dissociation between visual andauditory hemi-inattention: Evidence from temporal orderjudgements. Neuropsychologia, 45, 552–560.
Soroker, N., Calamaro, N., Glicksohn, J., & Myslobodsky, M. S.(1997). Auditory inattention in right-hemisphere-damagedpatients with and without visual neglect. Neuropsychologia,35, 249–256.
Spence, C., & Driver, J. (1997). Audiovisual links in exogenouscovert spatial orienting. Perception and Psychophysics, 59,1–22.
Spence, C., Lloyd, D., McGlone, F., Nicholls, M. E. R., &Driver, J. (2000). Inhibition of return is supramodal: A dem-onstration between all possible pairings of vision touch andaudition. Experimental Brain Research, 134, 42–48.
Spierer, L., Meuli, R., & Clarke, S. (2007). Extinction of audi-tory stimuli in hemineglect: Space versus ear. Neuropsycho-logia, 45, 540–551.
Vallar, G. (2001). Extrapersonal visual unilateral spatialneglect and its neuroanatomy. NeuroImage, 14, S52–S58.
Wenzel, E. M., Arruda, M., Kistler, D. J., & Wightman, F. L.(1993). Localization using nonindividualized head-relatedtransfer functions. Journal of the Acoustical Society ofAmerica, 94, 111–123.
Wightman, F. L., & Kistler, D. J. (1989). Headphone simula-tion of free-field listening. I: Stimulus synthesis. Journalof the Acoustical Society of America, 85, 858–867.
Wilson, B., Cockburn, J., & Halligan, P. W. (1987). BehaviouralInattention Test. Bury St Edmonds, UK: Thames Valley.
Woods, W. A., & Colburn, S. (1992). Test of a model ofauditory object formation using intensity and interauraltime difference discriminations. Journal of the AcousticalSociety of America, 91, 2894–2902.
Zimmer, U., Lewald, J., & Karnath, H.-O. (2003). Disturbedsound lateralization in patients with spatial neglect.Journal of Cognitive Neuroscience, 15, 694–703.