atrioventricular ring reentry in embryonic mouse hearts
TRANSCRIPT

Atrioventricular Ring Reentry in Embryonic Mouse HeartsMiguel Valderrábano, MD*; Fuhua Chen, MD*; Amish S. Dave, MD, PhD; Scott T. Lamp, MS;
Thomas S. Klitzner, MD; James N. Weiss, MD
Background—During development, AV conduction switches from base-to-apex to apex-to-base conduction afteremergence of the conduction system. We hypothesize that after this transition, the bulk of the AV ring, although nolonger required for AV conduction, remains transiently able to conduct, providing a potential arrhythmia substrate. Westudied AV conduction during this transition and its sensitivity to autonomic modulation.
Methods and Results—Simultaneous voltage and Ca2� mapping with RH-237 and Rhod-2 was performed with 2 CCDcameras in embryonic mouse hearts (n�43). Additionally, isolated calcium mapping was performed in 309 hearts withfluo-3AM. Propagation patterns in voltage and Ca2� mapping coincided. Arrhythmias were uncommon under basalconditions, with AV block in 14 (4%) and junctional rhythms in 4 (1%). Arrhythmias increased after stimulation withisoproterenol—junctional rhythm in 9 (3%) and ventricular rhythms in 22 (6%)—although AV block decreased (3hearts, 1%). Adding carbachol after isoproterenol caused dissociated antegrade and retrograde AV ring conduction in30 (8.6%) of E10.5 and E11.5 hearts, occurring preferentially in the right and left sides of the ring, respectively. In 2cases, reentry occurred circumferentially around the AV ring, perpendicular to normal propagation. Reentry persistedfor multiple beats, lasting from 3 to 22 minutes. No episodes of AV ring reentry occurred in E9.5 hearts.
Conclusions—AV ring reentry can occur by spatial dissociation of antegrade and retrograde conduction during combinedadrenergic and muscarinic receptor stimulation. Critical maturation (�E9.5) seems to be required to sustain reentry.(Circulation. 2006;114:543-549.)
Key Words: atrioventricular node � conduction � tachycardia
Atrioventricular conduction undergoes major remodeling duringdevelopment. The early heart tube exhibits a peristaltic, con-
tinuous propagation from the primitive atria to the primitive ventri-cles. After looping, the atria and ventricles become distinguishable,and AV conduction occurs through the AV ring (AVR), with acontinuous propagation sequence from the atria via the AVR to thebase of the ventricles, followed by the apex, and finally the outflowtract. Later on, with the emergence of the conduction system, theelectrical impulse is delivered from the primitive AV node to theapex, which then propagates to the rest of the ventricular tissue.1–3
At that point, AV conduction through the bulk of the AVR is nolonger necessary for ventricular activation and yet remains (at leasttransiently) able to conduct. We hypothesize that this redundancymay provide a substrate for reentry because different electrophysi-ological properties of AV conduction via the nascent AV node andremaining conduction via the AVR are to be expected.
Clinical Perspective p 549
AV node conduction is heavily regulated by autonomictone. We have recently shown that maturation of sympathetic
receptors precedes that of the parasympathetic receptors.4
Here, we show that during simultaneous sympathetic andparasympathetic receptor stimulation, reentry in the embry-onic AVR could be induced. Spatial dissociation of antegradeand retrograde conduction led to sustained reentry that inmost cases required participation of atrial tissue. In 1 case, theentire reentrant circuit appeared to be confined to the AVR.These findings support the hypothesis that the AVR, withoutparticipation of a mature AV node, can sustain reentry overan extremely small spatial scale. Speculations in relation tothe adult form of AV nodal reentrant tachycardia (AVNRT)also are proposed.
MethodsDissection of Embryonic Mouse HeartPregnant mice with different embryonic age groups were firstsedated by inhalation of isoflurane and then killed by cervicaldislocation.5 The uterus was dissected, and whole embryos wereexposed. Embryonic mouse hearts were then isolated with a dissect-ing microscope. During dissection, the embryonic mice were bathedin modified oxygenated Tyrode’s solution containing (mmol/L) 136
Received April 14, 2006; revision received May 24, 2006; accepted June 9, 2006.From the Departments of Medicine (Cardiology) (M.V., A.S.D., S.T.L., J.N.W.) and Pediatrics (F.C., T.S.K.) and the Cardiovascular Research
Laboratory (M.V., F.C., A.S.D., S.T.L., T.S.K., J.N.W.), David Geffen School of Medicine at UCLA, Los Angeles, Calif. Dr Valderrabano is currentlyat the Methodist DeBakey Heart Center, Houston, Tex.
The online-only Data Supplement is available with this article at http://circ.ahajournals.org/cgi/content/full/CIRCULATIONAHA.106.633727/DC1.*Drs Valderrábano and Chen contributed equally to this study.Correspondence to James N Weiss, MD, David Geffen School of Medicine at UCLA, 675 Charles Young Dr So, 3645 MRL, Los Angeles, CA
90095–1760. E-mail [email protected]© 2006 American Heart Association, Inc.
Circulation is available at http://www.circulationaha.org DOI: 10.1161/CIRCULATIONAHA.106.633727
543 by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from

NaCl, 5.4 KCl, 0.1 CaCl2, 0.33 NaH2PO4, 1 MgCl2, 10 HEPES, and10 glucose, pH 7.3. The same solution was used as the standard bathsolution in experiments with 1.8 mmol/L CaCl2.
Optical MappingIsolated E9.5 to E11.5 embryonic mouse hearts (n�43: E9.5 hearts,n�14; E10.5 hearts, n�13: and E11.5 hearts, n�16) were incubatedwith 5 �mol/L Rhod-2–acetoxymethyl ester dissolved in dimethylsulfoxide and pluronic F-127 for 40 minutes, followed by a 5-minuteincubation with 5 �mol/L RH-237. The embryonic mouse heartswere then washed in Tyrode’s solution before transfer to anexperimental chamber (T�37°C) on a modified inverted micro-scope. Green light (520�20 nm) from a high-intensity light-emittingdiode (Luxeon, Calgary, Canada) was delivered via the microscopelens into the hearts. Fluorescence was collected through the lens andsplit with a 630-nm dichroic mirror. Passed light (�630 nm) wasfiltered through a 710-nm long-pass filter (voltage signal); reflectedlight (�630 nm) was filtered through a 585�20-nm filter (Ca2�
signal). All filters were obtained from Chroma (Rockingham, Vt).Figure 1 shows a schematic of the setup. Ca2� and voltage signalswere simultaneously recorded with 2 electron-multiplier CCD cam-eras operating at 200 to 500 frames per second (PhotometricsCascade 128�, Tucson, Ariz) with a spatial resolution of 128�128pixels. Spontaneous activations were mapped before and after theaddition of Tyrode’s solution containing the �-adrenergic agonistisoproterenol (ISO; 1 �mol/L) and/or the muscarinic receptor agonistcarbachol (CCh; 10 �mol/L). Diacetylmonoxyme was used as anelectromechanical uncoupler to reduce motion artifact in 22 speci-mens. Experiments were conducted at 37°C. For technical simplicity,taking advantage of the higher signal-to-noise ratios of Ca2� indica-tors and the uniform correspondence of voltage and Ca2� propagationmaps, additional experiments (n�309) were conducted using iso-lated Ca2� mapping with fluo-3AM with either a single PhotometricsCascade camera as above or with another CCD camera (model LCL811K, Watec America, Las Vegas, Nev) using an ATI video capturecard (30 frames per second). A few experiments were conducted atroom temperature to visualize activations with the slower frame rate.
Data were then analyzed with custom software. After spatiotem-poral filtering, the onset of action potentials or Ca2� transients wasdetected by finding frames with a fluorescence that crossed themedian fluorescence value going up; the offset was assigned toframes whose fluorescence crossed the median going down. Actionpotential duration (APD) and Ca2� transient duration were measuredas the time delay between onset and offset. Propagation wave frontwas assigned to points at the onset of action potentials or Ca2�
transients and wave back to points at the offset. AV Ca2� transientconduction time was measured by marking the time differencebetween the activation of the atria and ventricle. All fluorophoreswere purchased from Molecular Probes (Eugene, Ore). ISO, CCh,and other chemicals were from Sigma Chemical Co (St Louis, Mo).The Student t test was used to compare means, and a value ofP�0.05 was considered statistically significant.
The authors had full access to the data and take full responsibilityfor their integrity. All authors have read and agree to the manuscriptas written.
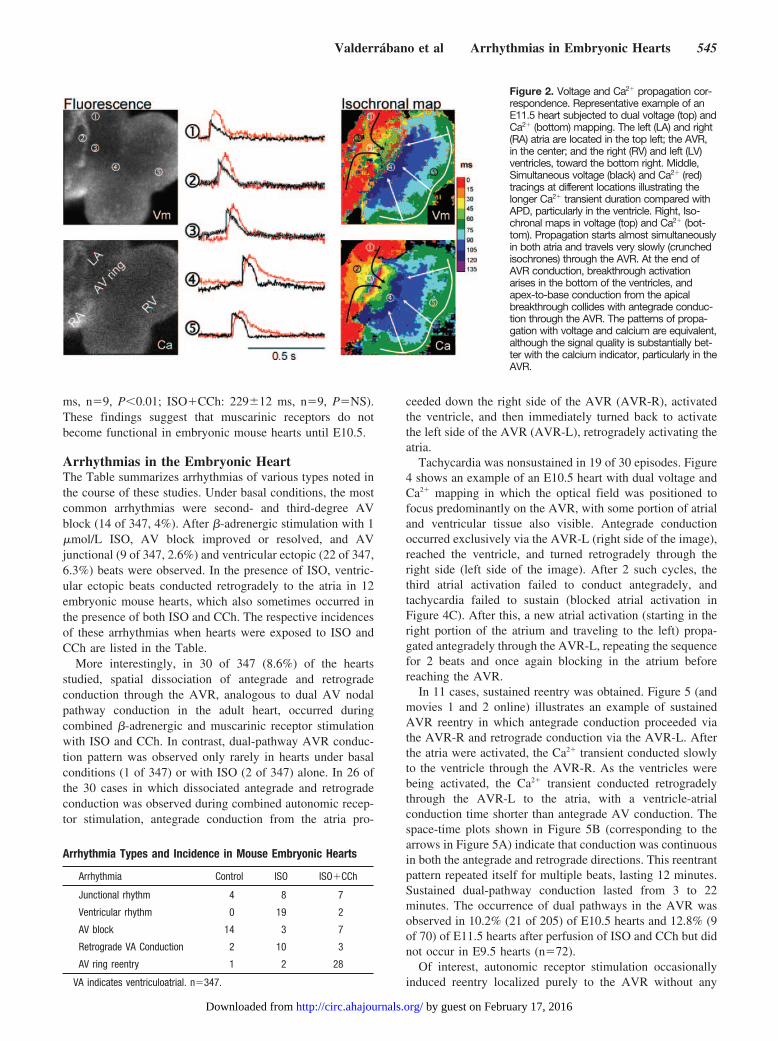
ResultsSimultaneous Voltage and Ca2� RecordingsActivation sequence and propagation patterns were equiva-lent between voltage and Ca2� maps in all 43 specimenssubjected to dual mapping. Figures 2 and 3 show examples.The mean ventricular APD was 166�34 ms. The meanventricular Ca2� transient duration in the ventricle measuredat 50% repolarization was 375�58 ms (P�0.001). AtrialAPDs measured 145�41 ms; atrial Ca2� transient durationmeasured 183�25 ms (P�0.05). In E9.5 hearts, AVconduction proceeded via the AVR in a base-to-apexpattern. In E10.5 and E11.5 hearts, however, the earliestventricular activation occurred at the apex, consistent witha maturing conduction system, as suggested previously (Figure3 for E9.5 and E10.5, Figure 2 for E11.5).1,2,6,7 The closeagreement between activation sequences in voltage and Ca2�
maps7 validates the use of Ca2� transients to map activationsequence.
Autonomic Modulation of Voltage and Ca2�
In response to 1 �mol/L ISO, heart rate increased from122�24 to 140�26 bpm (n�99), and AV conduction timeshortened from 234�6 to 205�5 ms (n�63), indicating thepresence of functional �-adrenergic receptors throughout theatria, AVR, and ventricles. APD prolongation accompaniedthe increases in heart rate induced by ISO, from 166�34 to240�29 ms (P�0.03). However, the Ca2� transient durationdid not change significantly after ISO (375�58 versus358�62 ms; P�0.05). APD decreased and Ca2� transientduration increased again after the addition of CCh, but thedifferences did not reach statistical significance comparedwith baseline or ISO values. In another group of embryonichearts exposed to ISO, 10 �mol/L CCh reversed the changesin AV conduction time and heart rate induced by ISO inE10.5 (basal: 282�11 ms, n�13; ISO: 212�9 ms, n�12;P�0.001; ISO�CCh: 280�14 ms, n�12, P�0.001) andE11.5 hearts (basal: 295�15 ms, n�13; ISO: 218�18 ms,n�13, P�0.01; ISO�CCh: 286�18 ms, n�13, P�0.01) butnot in E9.5 hearts (basal: 285�14 ms, n�9; ISO: 222�14
Figure 1. Experimental setup. Green lightfrom a light-emitting diode was filteredand delivered to the mapping field via adichroic mirror through a microscopelens. Reflected light passed through thefirst dichroic mirror and was split by asecond dichroic mirror. Wavelengths�630 nm were reflected, filtered at 585nm, and collected by 1 CCD camera reg-istering calcium signals (Rhod-2 emis-sion). Wavelengths �630 nm passed thedichroic mirror, were filtered through along-pass 710-nm filter, and were col-lected by the second CCD camera regis-tering voltage signals (RH-237 emission).
544 Circulation August 8, 2006
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from
ms, n�9, P�0.01; ISO�CCh: 229�12 ms, n�9, P�NS).These findings suggest that muscarinic receptors do notbecome functional in embryonic mouse hearts until E10.5.
Arrhythmias in the Embryonic HeartThe Table summarizes arrhythmias of various types noted inthe course of these studies. Under basal conditions, the mostcommon arrhythmias were second- and third-degree AVblock (14 of 347, 4%). After �-adrenergic stimulation with 1�mol/L ISO, AV block improved or resolved, and AVjunctional (9 of 347, 2.6%) and ventricular ectopic (22 of 347,6.3%) beats were observed. In the presence of ISO, ventric-ular ectopic beats conducted retrogradely to the atria in 12embryonic mouse hearts, which also sometimes occurred inthe presence of both ISO and CCh. The respective incidencesof these arrhythmias when hearts were exposed to ISO andCCh are listed in the Table.
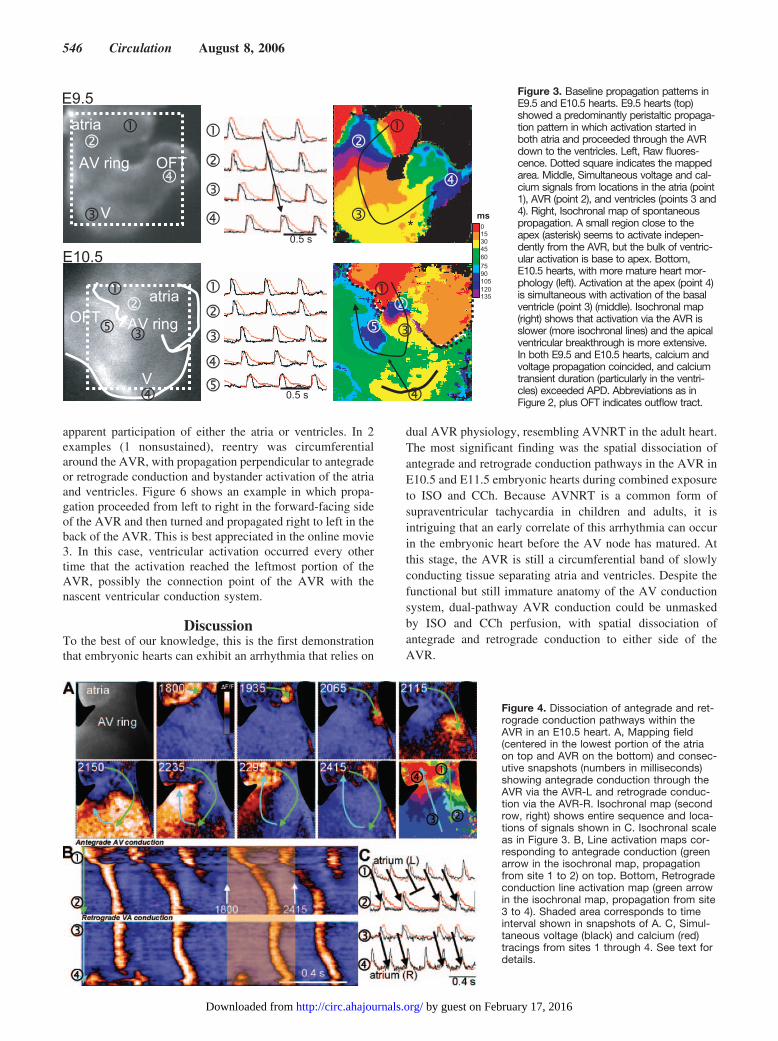
More interestingly, in 30 of 347 (8.6%) of the heartsstudied, spatial dissociation of antegrade and retrogradeconduction through the AVR, analogous to dual AV nodalpathway conduction in the adult heart, occurred duringcombined �-adrenergic and muscarinic receptor stimulationwith ISO and CCh. In contrast, dual-pathway AVR conduc-tion pattern was observed only rarely in hearts under basalconditions (1 of 347) or with ISO (2 of 347) alone. In 26 ofthe 30 cases in which dissociated antegrade and retrogradeconduction was observed during combined autonomic recep-tor stimulation, antegrade conduction from the atria pro-
ceeded down the right side of the AVR (AVR-R), activatedthe ventricle, and then immediately turned back to activatethe left side of the AVR (AVR-L), retrogradely activating theatria.
Tachycardia was nonsustained in 19 of 30 episodes. Figure4 shows an example of an E10.5 heart with dual voltage andCa2� mapping in which the optical field was positioned tofocus predominantly on the AVR, with some portion of atrialand ventricular tissue also visible. Antegrade conductionoccurred exclusively via the AVR-L (right side of the image),reached the ventricle, and turned retrogradely through theright side (left side of the image). After 2 such cycles, thethird atrial activation failed to conduct antegradely, andtachycardia failed to sustain (blocked atrial activation inFigure 4C). After this, a new atrial activation (starting in theright portion of the atrium and traveling to the left) propa-gated antegradely through the AVR-L, repeating the sequencefor 2 beats and once again blocking in the atrium beforereaching the AVR.
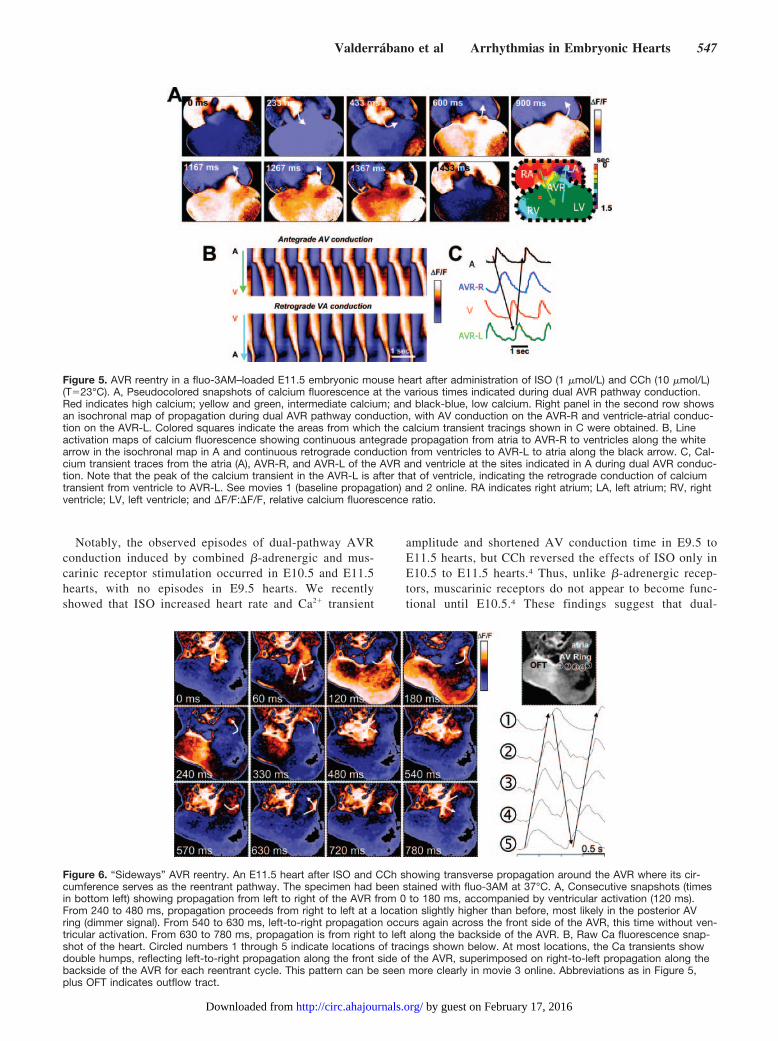
In 11 cases, sustained reentry was obtained. Figure 5 (andmovies 1 and 2 online) illustrates an example of sustainedAVR reentry in which antegrade conduction proceeded viathe AVR-R and retrograde conduction via the AVR-L. Afterthe atria were activated, the Ca2� transient conducted slowlyto the ventricle through the AVR-R. As the ventricles werebeing activated, the Ca2� transient conducted retrogradelythrough the AVR-L to the atria, with a ventricle-atrialconduction time shorter than antegrade AV conduction. Thespace-time plots shown in Figure 5B (corresponding to thearrows in Figure 5A) indicate that conduction was continuousin both the antegrade and retrograde directions. This reentrantpattern repeated itself for multiple beats, lasting 12 minutes.Sustained dual-pathway conduction lasted from 3 to 22minutes. The occurrence of dual pathways in the AVR wasobserved in 10.2% (21 of 205) of E10.5 hearts and 12.8% (9of 70) of E11.5 hearts after perfusion of ISO and CCh but didnot occur in E9.5 hearts (n�72).
Of interest, autonomic receptor stimulation occasionallyinduced reentry localized purely to the AVR without any
Arrhythmia Types and Incidence in Mouse Embryonic Hearts
Arrhythmia Control ISO ISO�CCh
Junctional rhythm 4 8 7
Ventricular rhythm 0 19 2
AV block 14 3 7
Retrograde VA Conduction 2 10 3
AV ring reentry 1 2 28
VA indicates ventriculoatrial. n�347.
Figure 2. Voltage and Ca2� propagation cor-respondence. Representative example of anE11.5 heart subjected to dual voltage (top) andCa2� (bottom) mapping. The left (LA) and right(RA) atria are located in the top left; the AVR,in the center; and the right (RV) and left (LV)ventricles, toward the bottom right. Middle,Simultaneous voltage (black) and Ca2� (red)tracings at different locations illustrating thelonger Ca2� transient duration compared withAPD, particularly in the ventricle. Right, Iso-chronal maps in voltage (top) and Ca2� (bot-tom). Propagation starts almost simultaneouslyin both atria and travels very slowly (crunchedisochrones) through the AVR. At the end ofAVR conduction, breakthrough activationarises in the bottom of the ventricles, andapex-to-base conduction from the apicalbreakthrough collides with antegrade conduc-tion through the AVR. The patterns of propa-gation with voltage and calcium are equivalent,although the signal quality is substantially bet-ter with the calcium indicator, particularly in theAVR.
Valderrábano et al Arrhythmias in Embryonic Hearts 545
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from
apparent participation of either the atria or ventricles. In 2examples (1 nonsustained), reentry was circumferentialaround the AVR, with propagation perpendicular to antegradeor retrograde conduction and bystander activation of the atriaand ventricles. Figure 6 shows an example in which propa-gation proceeded from left to right in the forward-facing sideof the AVR and then turned and propagated right to left in theback of the AVR. This is best appreciated in the online movie3. In this case, ventricular activation occurred every othertime that the activation reached the leftmost portion of theAVR, possibly the connection point of the AVR with thenascent ventricular conduction system.
DiscussionTo the best of our knowledge, this is the first demonstrationthat embryonic hearts can exhibit an arrhythmia that relies on
dual AVR physiology, resembling AVNRT in the adult heart.The most significant finding was the spatial dissociation ofantegrade and retrograde conduction pathways in the AVR inE10.5 and E11.5 embryonic hearts during combined exposureto ISO and CCh. Because AVNRT is a common form ofsupraventricular tachycardia in children and adults, it isintriguing that an early correlate of this arrhythmia can occurin the embryonic heart before the AV node has matured. Atthis stage, the AVR is still a circumferential band of slowlyconducting tissue separating atria and ventricles. Despite thefunctional but still immature anatomy of the AV conductionsystem, dual-pathway AVR conduction could be unmaskedby ISO and CCh perfusion, with spatial dissociation ofantegrade and retrograde conduction to either side of theAVR.
Figure 3. Baseline propagation patterns inE9.5 and E10.5 hearts. E9.5 hearts (top)showed a predominantly peristaltic propaga-tion pattern in which activation started inboth atria and proceeded through the AVRdown to the ventricles. Left, Raw fluores-cence. Dotted square indicates the mappedarea. Middle, Simultaneous voltage and cal-cium signals from locations in the atria (point1), AVR (point 2), and ventricles (points 3 and4). Right, Isochronal map of spontaneouspropagation. A small region close to theapex (asterisk) seems to activate indepen-dently from the AVR, but the bulk of ventric-ular activation is base to apex. Bottom,E10.5 hearts, with more mature heart mor-phology (left). Activation at the apex (point 4)is simultaneous with activation of the basalventricle (point 3) (middle). Isochronal map(right) shows that activation via the AVR isslower (more isochronal lines) and the apicalventricular breakthrough is more extensive.In both E9.5 and E10.5 hearts, calcium andvoltage propagation coincided, and calciumtransient duration (particularly in the ventri-cles) exceeded APD. Abbreviations as inFigure 2, plus OFT indicates outflow tract.
Figure 4. Dissociation of antegrade and ret-rograde conduction pathways within theAVR in an E10.5 heart. A, Mapping field(centered in the lowest portion of the atriaon top and AVR on the bottom) and consec-utive snapshots (numbers in milliseconds)showing antegrade conduction through theAVR via the AVR-L and retrograde conduc-tion via the AVR-R. Isochronal map (secondrow, right) shows entire sequence and loca-tions of signals shown in C. Isochronal scaleas in Figure 3. B, Line activation maps cor-responding to antegrade conduction (greenarrow in the isochronal map, propagationfrom site 1 to 2) on top. Bottom, Retrogradeconduction line activation map (green arrowin the isochronal map, propagation from site3 to 4). Shaded area corresponds to timeinterval shown in snapshots of A. C, Simul-taneous voltage (black) and calcium (red)tracings from sites 1 through 4. See text fordetails.
546 Circulation August 8, 2006
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from
Notably, the observed episodes of dual-pathway AVRconduction induced by combined �-adrenergic and mus-carinic receptor stimulation occurred in E10.5 and E11.5hearts, with no episodes in E9.5 hearts. We recentlyshowed that ISO increased heart rate and Ca2� transient
amplitude and shortened AV conduction time in E9.5 toE11.5 hearts, but CCh reversed the effects of ISO only inE10.5 to E11.5 hearts.4 Thus, unlike �-adrenergic recep-tors, muscarinic receptors do not appear to become func-tional until E10.5.4 These findings suggest that dual-
Figure 5. AVR reentry in a fluo-3AM–loaded E11.5 embryonic mouse heart after administration of ISO (1 �mol/L) and CCh (10 �mol/L)(T�23°C). A, Pseudocolored snapshots of calcium fluorescence at the various times indicated during dual AVR pathway conduction.Red indicates high calcium; yellow and green, intermediate calcium; and black-blue, low calcium. Right panel in the second row showsan isochronal map of propagation during dual AVR pathway conduction, with AV conduction on the AVR-R and ventricle-atrial conduc-tion on the AVR-L. Colored squares indicate the areas from which the calcium transient tracings shown in C were obtained. B, Lineactivation maps of calcium fluorescence showing continuous antegrade propagation from atria to AVR-R to ventricles along the whitearrow in the isochronal map in A and continuous retrograde conduction from ventricles to AVR-L to atria along the black arrow. C, Cal-cium transient traces from the atria (A), AVR-R, and AVR-L of the AVR and ventricle at the sites indicated in A during dual AVR conduc-tion. Note that the peak of the calcium transient in the AVR-L is after that of ventricle, indicating the retrograde conduction of calciumtransient from ventricle to AVR-L. See movies 1 (baseline propagation) and 2 online. RA indicates right atrium; LA, left atrium; RV, rightventricle; LV, left ventricle; and �F/F:�F/F, relative calcium fluorescence ratio.
Figure 6. “Sideways” AVR reentry. An E11.5 heart after ISO and CCh showing transverse propagation around the AVR where its cir-cumference serves as the reentrant pathway. The specimen had been stained with fluo-3AM at 37°C. A, Consecutive snapshots (timesin bottom left) showing propagation from left to right of the AVR from 0 to 180 ms, accompanied by ventricular activation (120 ms).From 240 to 480 ms, propagation proceeds from right to left at a location slightly higher than before, most likely in the posterior AVring (dimmer signal). From 540 to 630 ms, left-to-right propagation occurs again across the front side of the AVR, this time without ven-tricular activation. From 630 to 780 ms, propagation is from right to left along the backside of the AVR. B, Raw Ca fluorescence snap-shot of the heart. Circled numbers 1 through 5 indicate locations of tracings shown below. At most locations, the Ca transients showdouble humps, reflecting left-to-right propagation along the front side of the AVR, superimposed on right-to-left propagation along thebackside of the AVR for each reentrant cycle. This pattern can be seen more clearly in movie 3 online. Abbreviations as in Figure 5,plus OFT indicates outflow tract.
Valderrábano et al Arrhythmias in Embryonic Hearts 547
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from

pathway AVR conduction may require the presence offunctional muscarinic receptors. One possibility is that theexpression of muscarinic receptors in the AVR is stillheterogeneous at E10.5 to E11.5, so CCh only reverses theISO-enhanced conduction in certain regions, creating aheterogeneous substrate facilitating dissociation of AVRconduction pathways. In E9.5 hearts, the lack of musca-rinic receptors may preclude combined autonomic receptorstimulation from creating a sufficiently heterogeneoussubstrate for dual-pathway AVR. Alternatively, the AVRat E9.5 may be too small to support dual pathways. Finally,it is possible that AVR reentry also depends in some wayon a functional AV node and ventricular conductionsystem, which is not yet present in E9.5 hearts, as indicatedby the activation sequence (Figure 2).
Correlations between our findings and the pathophysi-ology of adult AVNRT remain speculative, and we canonly hypothesize how the specific activation patterns inembryos might relate to those of AVNRT. The embryo-genesis of nodal and perinodal tissues that form thesubstrates of AVNRT is unclear. After looping of the hearttube, the primitive atria and ventricles appear as distinctprotuberances, whereas the AVR area remains smoothlytubular. This AVR area carries the electrical impulse fromatria to ventricles, proceeding sequentially in a peristalticfashion. The primitive AV node forms in the AV junctionbetween the 2 ventricles as a localized bundle identifiablein rabbits by neurofilament 160 (NF-160) staining,3 whichis contiguous with the ventricular conduction system.8 Thedevelopment of the conduction system is correlated withan apex-to-base activation of the ventricles.1–3 However,before the AV node is formed, conduction between theatria and the ventricles occurs through the AVR. Thisimplies that a transition between AVR conduction and AVnode conduction must take place by which AV propagationvia the AVR is replaced by propagation through the AVnode. Once that occurs, conduction through the AVRbecomes superfluous. To electrically disconnect the AVRfrom the ventricles, propagation through the AVR has tobe uncoupled from the ventricles and funneled to the AVnode for it to become the sole AV connection. During thistransition, progressive uncoupling of the AVR and theventricles occurs. The AVR, however, remains connectedto the nascent AV node.
Despite the high success rate of curative radiofrequencyablation, the anatomic substrate that sustains AVNRTremains unclear. Since the initial demonstration of func-tional dual AV nodal conduction by Moe et al9 and Mendezand Moe,10 the specific anatomic locations of the conduc-tion pathways have been the subject of debate. Moreover,their embryologic origin remains unexplored. Optical andmultielectrode mapping data in a rabbit model have sug-gested that perinodal atrial tissue participates in the reen-trant circuit.11–13 We hypothesize that the AVR could formthese inputs to the AV node, which in turn could serve inthe adult as part of the perinodal reentrant circuit duringAVNRT.
Finally, we cannot distinguish whether these dual path-ways were functionally induced or anatomically based.
The observation that in sinus rhythm, conduction throughboth sides of the AVR appeared uniform may favor afunctional mechanism. More detailed studies are needed todetermine the basis for the dual AVR pathways in theseembryonic hearts.
In this study, we found that arrhythmias are relativelyuncommon in embryonic mouse heart under basal condi-tions but can be elicited by �-adrenergic and muscarinicreceptor stimulation. An important caveat, however, is thatwe studied the embryonic hearts in vitro rather than invivo. Whether similar arrhythmias occur in vivo embry-onic hearts under physiological conditions remains to bedetermined.
ConclusionThe present study is, to the best of our knowledge, the first todemonstrate that dual-pathway AVR conduction can occur inembryonic hearts before the AV node has developed into adiscrete anatomically defined structure.
AcknowledgmentsThe authors thank L. Pan, J. Parker, and T. Duong for their technicalassistance.
Sources of FundingThe study was funded by the American Heart Association, WesternStates Affiliate (beginning grant-in-aid 0565149y to M.V.), the NIH(P01 HL-078931 to J.N.W.), and the Laubisch and KawataEndowments.
DisclosuresNone.
References1. Rentschler S, Vaidya DM, Tamaddon H, Degenhardt K, Sassoon D,
Morley GE, Jalife J, Fishman GI. Visualization and functional character-ization of the developing murine cardiac conduction system. Devel-opment. 2001;128:1785–1792.
2. Chuck ET, Freeman DM, Watanabe M, Rosenbaum DS. Changing acti-vation sequence in the embryonic chick heart: implications for the devel-opment of the His-Purkinje system. Circ Res. 1997;81:470–476.
3. Rothenberg F, Nikolski VP, Watanabe M, Efimov IR. Electrophysiologyand anatomy of embryonic rabbit hearts before and after septation.Am J Physiol Heart Circ Physiol. 2005;288:H344–H351.
4. Chen F, Klitzner TS, Weiss JN. Autonomic regulation of calciumcycling in developing embryonic mouse hearts. Cell Calcium. 2006;39:375–385.
5. Reuter H, Henderson SA, Han T, Mottino GA, Frank JS, Ross RS,Goldhaber JI, Philipson KD. Cardiac excitation-contraction couplingin the absence of Na(�)-Ca2� exchange. Cell Calcium. 2003;34:19 –26.
548 Circulation August 8, 2006
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from

6. Sedmera D, Reckova M, Bigelow MR, Dealmeida A, Stanley CP,Mikawa T, Gourdie RG, Thompson RP. Developmental transitions inelectrical activation patterns in chick embryonic heart. Anat Rec A DiscovMol Cell Evol Biol. 2004;280:1001–1009.
7. Chuck ET, Meyers K, France D, Creazzo TL, Morley GE. Transitions inventricular activation revealed by two-dimensional optical mapping. AnatRec A Discov Mol Cell Evol Biol. 2004;280:990–1000.
8. Rothenberg F, Efimov IR. Three-dimensional anatomy of the conductionsystem of the early embryonic rabbit heart. Anat Rec A Discov Mol CellEvol Biol. 2006;288:3–7 .
9. Moe GK, Preston JB, Burlington H. Physiologic evidence for a dual A-Vtransmission system. Circ Res. 1956;4:357–375.
10. Mendez C, Moe GK. Demonstration of a dual A-V nodal conductionsystem in the isolated rabbit heart. Circ Res. 1966;19:378–393.
11. Nikolski V, Efimov I. Fluorescent imaging of a dual-pathway atrioven-tricular-nodal conduction system. Circ Res. 2001;88:E23–E30.
12. Nikolski VP, Jones SA, Lancaster MK, Boyett MR, Efimov IR. Cx43 anddual-pathway electrophysiology of the atrioventricular node and atrioven-tricular nodal reentry. Circ Res. 2003;92:469–75.
13. Loh P, Ho SY, Kawara T, Hauer RN, Janse MJ, Breithardt G, de BakkerJM. Reentrant circuits in the canine atrioventricular node during atrial andventricular echoes: electrophysiological and histological correlation. Cir-culation. 2003;108:231–8.
CLINICAL PERSPECTIVEThe embryogenesis of the structures involved in AV conduction is unclear. It is known that at an initial heart tube stage,propagation proceeds sequentially in a peristaltic fashion from the primitive atria to the ventricles, which are activated frombase to apex. With maturation of the conduction system, however, the ventricular activation sequence reverses becausePurkinje fibers deliver propagation to the apex first, which then proceeds toward the base. We reasoned that aftermaturation of the conduction system, the so-called peristaltic activation mechanism from atria to ventricles becomesredundant and can provide a substrate for reentry. We mapped propagation in embryonic mouse hearts at different stagesof development during which this transition occurs. By using dual autonomic modulation, both sympathetic andparasympathetic, we show that reentry in the AV ring can occur, during which spatial dissociation of antegrade andretrograde conduction pathways is present, similar to what happens in the clinically common AV nodal reentry. Wespeculate that the primitive AV ring may evolve to participate in the perinodal tissue that contributes to this common adultarrhythmia.
Valderrábano et al Arrhythmias in Embryonic Hearts 549
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from

James N. WeissMiguel Valderrábano, Fuhua Chen, Amish S. Dave, Scott T. Lamp, Thomas S. Klitzner and
Atrioventricular Ring Reentry in Embryonic Mouse Hearts
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2006 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/CIRCULATIONAHA.106.6337272006;114:543-549; originally published online July 31, 2006;Circulation.
http://circ.ahajournals.org/content/114/6/543World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org/content/suppl/2006/07/31/CIRCULATIONAHA.106.633727.DC1.htmlData Supplement (unedited) at:
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on February 17, 2016http://circ.ahajournals.org/Downloaded from