association of the irf5 risk haplotype with high serum interferon-α activity in systemic lupus...
TRANSCRIPT

Association of the IRF5 Risk Haplotype With High SerumInterferon-α Activity in Systemic Lupus Erythematosus Patients
Timothy B. Niewold, MD1, Jennifer A. Kelly, MPH2, Marie H. Flesch, MS2, Luis R. Espinoza,MD3, John B. Harley, MD, PhD4, and Mary K. Crow, MD5
1University of Chicago, Chicago, Illinois
2Oklahoma Medical Research Foundation, Oklahoma City
3Louisiana State University, New Orleans
4Oklahoma Medical Research Foundation, Oklahoma City VAMC, and University of Oklahoma, OklahomaCity
5Hospital for Special Surgery, New York, New York
AbstractObjective—A haplotype of the interferon regulatory factor 5 (IRF5) gene has been associated withthe risk of developing systemic lupus erythematosus (SLE), and our previous studies havedemonstrated that high levels of serum interferon-α (IFNα) activity are a heritable risk factor forSLE. The aim of this study was to determine whether the IRF5 SLE risk haplotype mediates the riskof SLE by predisposing patients to the development of high levels of serum IFNα activity.
Methods—IFNα levels in 199 SLE patients of European and Hispanic ancestry were measured witha sensitive functional reporter cell assay. The rs2004640, rs3807306, rs10488631, and rs2280714single-nucleotide polymorphisms (SNPs) in IRF5 were genotyped in these patients. Haplotypes werecategorized as SLE risk, neutral, or protective based on published data.
Results—SLE patients with risk/risk and risk/neutral IRF5 genotypes had higher serum IFNαactivity than did those with protective/protective and neutral/protective genotypes (P = 0.025). Thisdifferential effect of IRF5 genotype on serum IFNα levels was driven largely by SLE patients whowere positive for either anti–RNA binding protein (anti-RBP) or anti–double-stranded DNA (anti-dsDNA) autoantibodies (P = 0.012 for risk/risk or risk/neutral versus protective/protective or neutral/protective). The rs3807306 genotype was independently associated with high serum IFNα in thisautoantibody group. We found no difference in IFNα activity according to IRF5 genotype in patientslacking either type of autoantibody or in patients positive for both classes of autoantibody.
Address correspondence and reprint requests to Timothy B. Niewold, MD, University of Chicago, Section of Rheumatology, 5841 SouthMaryland Avenue, Mail Code 0930, Chicago, IL 60637. E-mail: [email protected]. Harley has received consulting fees, speaking fees, and/or honoraria from Bio-Rad Industries, Inc., and IVAX Diagnostics, Inc. (morethan $10,000 each), and owns stock or stock options in IVAX Diagnostics, Inc. Dr. Crow has received consulting fees, speaking fees,and/or honoraria from Genentech, Novo Nordisk, and ZymoGenetics (less than $10,000 each), owns stock or stock options in XDx, Inc.,and has a patent pending for an interferon assay.Author Contributions: Dr. Niewold had full access to all of the data in the study and takes responsibility for the integrity of the dataand the accuracy of the data analysis.Study design. Niewold, Harley, Crow.Acquisition of data. Niewold, Kelly, Flesch, Espinoza, Harley.Analysis and interpretation of data. Niewold, Harley, Crow.Manuscript preparation. Niewold, Harley, Crow.Statistical analysis. Niewold.
NIH Public AccessAuthor ManuscriptArthritis Rheum. Author manuscript; available in PMC 2009 August 1.
Published in final edited form as:Arthritis Rheum. 2008 August ; 58(8): 2481–2487. doi:10.1002/art.23613.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Conclusion—The IRF5 SLE risk haplotype is associated with higher serum IFNα activity in SLEpatients, and this effect is most prominent in patients positive for either anti-RBP or anti-dsDNAautoantibodies. This study demonstrates the biologic relevance of the SLE risk haplotype of IRF5 atthe protein level.
The pathogenesis of systemic lupus erythematosus (SLE) is likely driven by a combination ofgenetic risk factors and environmental events that lead to an irreversible break in immunologicself tolerance. Interferon-α (IFNα) is a pleiotropic type I interferon with the potential to breakself tolerance by activating antigen-presenting cells after uptake of self material (1). Serumlevels of IFNα are elevated in SLE patients (2,3) and are correlated with disease activity (4).Additionally, a number of patients treated with recombinant human IFNα for malignancy andchronic viral hepatitis have developed de novo SLE, which typically resolves after the IFNαis discontinued (5,6). These data suggest a potential role of IFNα in susceptibility to SLE. Inprevious studies, we demonstrated that abnormally high serum IFNα activity is common inSLE families, both in healthy and SLE-affected members, as compared with unrelatedindividuals (7). These data implicate high levels of serum IFNα as a heritable SLE risk factor;however, the causative genes underlying this risk factor are not known.
Interferon regulatory factor 5 (IRF-5) is a transcription factor that can induce transcription ofthe IFNα transcript itself as well as many IFNα-induced genes (8). Genetic association studiesexamining single-nucleotide polymorphisms (SNPs) in IRF5 have defined distinct haplotypesthat confer either susceptibility to, or protection from, SLE in persons of European and Hispanicancestry (9–11). These haplotypes are characterized by functional changes in IRF5 messengerRNA (mRNA) (10,11), which presumably will alter IRF5-mediated transcription, resulting inthe risk of SLE. The SLE risk haplotypes of IRF5 could potentially alter serum IFNα activityin vivo. We hypothesized that the IRF5 SLE risk haplotype predisposes to high serum IFNαactivity, which subsequently results in the risk of SLE. In this study, we examined serumIFNα activity and IRF5 genotypes in SLE patients to determine whether the IRF5 genotypeinfluences IFNα activity in vivo in patients with SLE.
Patients and MethodsPatients and samples
Serum, plasma, and genomic DNA samples were obtained from the Hospital for SpecialSurgery (HSS) Lupus Family Registry, the HSS Lupus Registry, and the Lupus Family Registryand Repository (LFRR) at the Oklahoma Medical Research Foundation (OMRF). A total of199 SLE patients of European American and Hispanic American ancestry were studied,including 108 patients from the HSS registries (58 of European ancestry and 50 of Hispanicancestry) and 91 patients from the LFRR (65 of European ancestry and 26 of Hispanicancestry). The patients of Hispanic ancestry in both registries are self-identified as HispanicAmerican, with ancestral origins in Mexico, South America, and the Caribbean. The allelefrequencies and haplotype structures of IRF5 did not differ significantly between patients ofEuropean ancestry and patients of Hispanic ancestry, and both populations are presented in thestudy in aggregate. Control samples from 141 healthy subjects were obtained commerciallyfrom healthy blood donors (n = 59), as well as from healthy unrelated individuals who haddonated to the 2 registries as control subjects (50 from the HSS registries and 32 from theLFRR). The study was approved by the institutional review boards at all institutions, andinformed consent was obtained from all subjects in the study.
Measurement of autoantibodiesAntibodies to double-stranded DNA (dsDNA) and RNA binding proteins (anti-Ro, anti-La,anti-Sm, and anti-RNP, collectively referred to as anti-RBP antibodies) were measured in
Niewold et al. Page 2
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

samples from the HSS registries by the Clinical Laboratory at the HSS, using enzyme-linkedimmunosorbent assay (ELISA) methods. Antibodies were measured in samples from the LFRRby the Clinical Immunology Laboratory at the OMRF, using precipitin techniques. Standardclinical laboratory cutoff points from each laboratory were used to categorize samples aspositive or negative, and a sample with a positive result for 1 or more of the anti-Ro, anti-La,and anti-Sm, or anti-RNP antibodies was considered anti-RBP positive in this study.
Stimulation of reporter cellsELISAs for the detection of IFNα in human serum have been complicated by poor correlationof the results with those of functional assays (12), possibly due to detection of a similar epitopeon a non-IFNα protein or a stable, but biologically inactive, IFNα breakdown product (12). Wehave developed and validated a sensitive and reproducible bioassay for the detection of serumIFNα activity (7,13). In this assay, reporter cells are used to measure the ability of patient serato cause IFN-induced gene expression. The reporter cells (WISH cells) (no. CCL-25; AmericanType Culture Collection, Rockville, MD) are an epithelial-derived cell line that is highlyresponsive to IFNα.
WISH cells were cultured in minimum essential medium (Invitrogen, Carlsbad, CA) withEarle's salts, 10% fetal bovine serum, 10 mM HEPES buffer, 2 mM L-glutamine, 100 units/mlof penicillin, and 100 μg/ml of streptomycin. Cells were then plated in 96-well culture platesat a density of 5 × 105/ml and incubated with 50% patient plasma or serum for 6 hours.Recombinant IFNα was used as a positive control, and healthy sera and culture media wereused as negative controls. After 6 hours, the cells were lysed.
The ability of sera to cause IFN-induced gene expression was largely abrogated by the additionof monoclonal anti-IFNα antibody (Chemicon, Temecula, CA), confirming that IFNα was themajor active type I IFN causing the IFN-induced gene expression (7). In a set of samples withlow levels of IFNα activity, the addition of recombinant IFNα (BioSource International,Camarillo, CA) resulted in IFNα-induced gene expression proportional to the amount ofIFNα added, excluding any frequent significant endogenous inhibitors of IFNα in the samples(7). Further details on the validation of the assay are described elsewhere (7,13).
Purification of total cellular mRNA and synthesis of complementary DNA (cDNA)Total cellular mRNA was purified from the WISH cell lysates using the Qiagen TurbocaptureRNA purification kit (Qiagen, Chatsworth, CA) according to the manufacturer's protocol. ThecDNA was prepared from the mRNA using the Invitrogen oligo(dT) primer and SuperscriptIII reverse transcriptase system (Invitrogen).
Real-time polymerase chain reaction (PCR) quantification of mRNA-derived cDNATen microliters of a 1:40 dilution of the cDNA was then quantified using real-time PCR withthe Bio-Rad SYBR Green fluorophore system (Bio-Rad, Hercules, CA). Forward and reverseprimers for the genes encoding myxovirus resistance 1 (MX-1), RNA-dependent protein kinase(PKR), and IFN-induced protein with tetratricopeptide repeats 1 (IFIT-1), which are known tobe highly and specifically induced by IFNα (14), were used in the reaction: for MX1, 5′-TACCAGGACTACGAGATTG-3′ (forward) and 5′-TGCCAGGAAGGTCTATTAG-3′(reverse); for PKR, 5′-CTTCCATCTGACTCAGGTTT-3′ (forward) and 5′-TGCTTCTGACGGTATGTATTA-3′ (reverse), and for IFIT1, 5′-CTCCTTGGGTTCGTCTATAAATTG-3′ (forward) and 5′-AGTCAGCAGCCAGTCTCAG-3′ (reverse). The housekeeping gene GAPDH was alsoamplified to control for background gene expression: 5′-CAACGGATTTGGTCGTATT-3′(forward) and 5′-GATGGCAACAATATCCACTT-3′ (reverse). Each sample and control was
Niewold et al. Page 3
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

run in duplicate. Melt curves were analyzed to ensure the specificity of the amplified product,and standard curves were generated for each PCR experiment.
Real-time PCR data analysisThe amount of PCR product of the IFNα-induced gene was normalized to the amount of productfor the housekeeping gene GAPDH in the same sample. The relative expression of each of the3 IFN-induced genes tested was calculated as the fold increase compared with its expressionin WISH cells cultured in medium alone. Healthy unrelated donor sera were tested in the WISHassay to establish a normal value for IFNα activity, and the mean and SD of the relativeexpression of the IFNα-induced gene induced by healthy donor sera in the WISH assay werecalculated. The ability of serum samples from the patients to cause IFN-induced geneexpression in the reporter cells was then compared with the mean and SD induced by healthyunrelated donor sera. To accomplish this, the number of SDs of the relative expression abovethe mean in healthy donors was calculated for each gene. The mean relative expression of thegene in cells exposed to healthy donor sera was subtracted from the mean relative expressionof the gene in cells exposed to experimental sera, and then the remainder was divided by theSD value for the same gene in healthy donors, which gave the relative number of SDs abovethat in healthy donors. This number was generated for each of the 3 genes and then summedto yield the final score. This normalization by the SD value in healthy donors was neededbecause each gene is up-regulated to a different degree following stimulation, and withoutnormalization, data from the most highly up-regulated gene would dominate in the data set.Despite this difficulty, we think that measurement of 3 genes is a more robust indicator ofdownstream coordinate IFNα pathway activation than is measurement of just 1 highly inducedtranscript.
GenotypingIndividuals in the HSS registries were genotyped at the rs2004640, rs3807306, rs2070197,rs10488631, and rs2280714 SNPs using ABI TaqMan Assays-by-Design primers and probeson an ABI 7900HT PCR sequencer (Applied Biosystems, Foster City, CA). Individuals in theLFRR registry were genotyped at the rs2004640, rs3807306, rs10488631, rs10954213, andrs2280714 SNPs using the Illumina GoldenGate system (Illumina, San Diego, CA). SNPgenotyping was performed with >98% completeness in both registries. All tested SNPsconformed to Hardy-Weinberg equilibrium; P values for departure from Hardy-Weinbergequilibrium were as follows: for rs2004640, P = 0.94; for rs3807306, P = 0.83; for rs2070197,P = 1.0; for rs10488631, P = 0.92; for rs2280714, P = 0.95; and for rs10954213, P = 1.0. Therewere no significant differences in allele frequencies between the 2 study sites; P values for thedifference in allele frequency by site were as follows: for rs2004640, P = 0.67; for rs3807306,P = 0.27; for rs10488631, P = 0.74; and for rs2280714, P = 0.49 (by chi-square analysis).
Statistical analysisTwo-sided Fisher's exact test (sum of small P's method for observed ≥ expected) was used toanalyze the categorical data, and the Mann-Whitney nonparametric t-test was used to comparequantitative data. Haploview 4.0 software was used for haplotype analysis and for calculationof D′ and r2 values. Haplotypes observed in the patients were categorized as risk, neutral, orprotective as they related to SLE susceptibility, based on previous genetic association studiesin SLE patients of European and Hispanic ancestry (10,15).
Niewold et al. Page 4
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

ResultsIRF5 genotyping results
All individuals were genotyped at the rs2004640, rs3807306, rs10488631, and rs2280714SNPs in IRF5. Haplotype structures and frequencies based on these 4 SNPs in our cohort werevery similar to published data (10), as illustrated in Figure 1. The rs2070197 SNP wasgenotyped only in the HSS cohort. SNP rs10488631 was a perfect proxy for rs2070197 (D′ =1, r2 = 1), and the latter SNP was not genotyped in the LFRR patients. SNP rs10954213 is afunctional polyadenylation site variant associated with SLE risk (10), and this SNP wasgenotyped in the LFRR cohort. The rs2280714 and rs10954213 SNPs were in high linkagedisequilibrium (D′ = 1, r2 = 0.79), which is consistent with previous studies (10). The rs2280714and rs10488631 SNPs were used as proxies for rs10954213 as it relates to SLE risk, andcombinations of the 4 markers tested in all patients captured >98% of the variation atrs10954213.
Association of IRF5 SLE risk genotypes with higher levels of serum IFNα activity ascompared with protective genotypes
Among SLE patients stratified by genotype at single SNPs in IRF5, there were no significantdifferences in serum IFNα activity (P > 0.35 for all single-marker analyses). When IRF5genotypes were analyzed as combinations of risk, neutral, and protective haplotypes, SLEpatients with the risk/risk or risk/neutral IRF5 genotypes had higher serum IFNα activity thandid those with the protective/protective or protective/neutral genotypes (P = 0.025). Patientswith neutral/neutral or risk/protective genotypes demonstrated an intermediate level of serumIFNα activity, as shown in Figure 2. There was a trend toward higher IFNα activity in the risk/risk and risk/neutral group as compared with the neutral/neutral and risk/protective genotypegroup (P = 0.075). The 3 genotype groups presented in Figure 2B were formed by combininggenotypes that had the most similar median values for serum IFNα activity; median serumIFNα activity values for each genotype group were as follows protective/protective = 1.94,neutral/protective = 2.35, neutral/neutral = 3.14, risk/protective = 3.13, risk/neutral = 5.81, andrisk/risk = 9.59.
Most pronounced differential effect of IRF5 genotype on serum IFNα activity in SLE patientswith either anti-RBP or anti-dsDNA antibodies
In previous studies, we showed that anti-RBP autoantibodies, such as Ro, La, Sm, and RNP,as well as anti-dsDNA autoantibodies are independently associated with high levels of serumIFNα activity in SLE patients (7). Anti-RBP and anti-dsDNA antibodies influence serumIFNα in an additive manner, and patients who are positive for both classes of autoantibodies(double-positive) have higher mean levels of serum IFNα than those who are positive for eitheranti-RBP or anti-dsDNA alone (single-positive) (7). The 2 single-positive groups (RBP+/dsDNA− and RBP−/dsDNA+) have similar mean serum IFNα activity (7).
When SLE patients in the present study were stratified by double-negative, single-positive, ordouble-positive autoantibody status and IRF5 genotype, the greatest difference in serumIFNα activity by IRF5 genotype was seen in the single-positive autoantibody group, as shownin Figure 3. Double-positive patients had significantly higher serum IFNα activity as a groupthan single-positive patients (P = 0.015); however, there were no significant differences inserum IFNα activity by IRF5 genotype in the double-positive group. The strong influence ofdouble-positive autoantibody status appeared to obscure the influence of IRF5 genotype,although the apparent lack of influence of IRF5 genotype in this group could also be due tothe fact that there were fewer patients in the double-positive group to drive the statisticalsignificance (42 double-positive patients versus 82 single-positive patients). There were nosignificant differences in serum IFNα activity by IRF5 genotype in the double-negative patient
Niewold et al. Page 5
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

group (P = 0.46 for protective/protective plus neutral/protective versus risk/neutral plus risk/risk), despite having similar numbers of patients as the single-positive group. There were nodifferences in haplotype or genotype frequencies between the different autoantibody strata,suggesting that these 2 variables are independent (P for dependence = 0.29).
The median level of serum IFNα activity by antibody status and genotype in the double-negative and single-positive groups were as follows: 0.54, 0.88, and 1.01 in the double-negativegroup with the protective/protective or neutral/protective genotype, the neutral/neutral or risk/protective genotype, and the risk/neutral or risk/risk genotype, respectively, and 2.55, 5.42,and 19.52 in the single-positive group with the protective/protective or neutral/protectivegenotype, the neutral/neutral or risk/protective genotype, and the risk/neutral or risk/riskgenotype, respectively. Thus, the double-negative risk/risk plus risk/neutral group had 1.8times higher median serum IFNα activity than the double-negative protective/protective plusprotective/neutral group. The single-positive risk/risk plus risk/neutral group had 7.6 timeshigher median serum IFNα activity than the single-positive protective/protective plus neutral/protective group. This would suggest a roughly 4-fold multiplicative interaction between singleautoantibody positivity and IRF5 genotype on the median serum IFNα activity when comparingthe double-negative and single-positive groups. Variances are wide in the data set, however,as shown in Figure 3, so this interaction estimate should be interpreted with caution.
Two-marker analysis based on rs3807306 and rs10488631After completion of our study, new data were published suggesting that a novel insertion/deletion in the promoter region could be a major contributor to the SLE risk signal arising fromthe 5′ region of IRF5 (15). In their study, Sigurdsson et al (15) found the most parsimoniousdesignation of SLE risk using the promoter insertion/deletion as a marker for SLE risk in the5′ region of the gene, and the rs10488631 SNP as a marker for SLE risk in the 3′ region of thegene. The risk variant of the promoter insertion/deletion was highly correlated with the A alleleof the rs3807306 SNP (r2 = 0.61), which was also genotyped in the present study.
Given these data, we undertook a reanalysis of our serum IFNα activity data using thers3807306 risk allele to designate the contribution of the 5′ region of IRF5 to SLE risk and thers10488631 allele as a 3′ marker of SLE risk. As shown in Figure 4, serum IFNα activity variedsignificantly by IRF5 genotype at these 2 SNPs in the single-positive autoantibody group, withan increasing number of risk alleles corresponding to higher serum IFNα activity. Interestingly,in the single-positive autoantibody group, the rs3807306 SNP shows an independentassociation with high levels of IFNα in the absence of the rs10488631 risk variant (P = 0.03for AA/TT versus CC/TT genotypes at rs3807306/rs10488631). In our cohort, the SLE riskvariant of rs10488631 was always found with the SLE risk variant of rs3807306, so anindependent influence of rs10488631 and rs3807306 could not be tested, although the highestlevels of serum IFNα activity were seen in the presence of both SLE risk variants.
DiscussionWe have previously demonstrated familial aggregation of high serum IFNα activity in SLEfamilies, suggesting that high serum IFNα is a heritable SLE risk factor (7). In this study, wedemonstrated that the SLE risk haplotype of IRF5 is associated with higher serum IFNα activityin SLE patients than the protective haplotypes. These data suggest that IRF5 haplotypescontribute to the heritability of serum IFNα activity and play a role in the variance in IFNαactivity observed between different SLE patients. Thus, it seems likely that IRF5 mediates SLErisk, at least in part, by modulating serum IFNα activity, which provides biologic relevance atthe protein level for the SLE risk haplotype of IRF5. The protective/protective and protective/neutral genotype groups showed similar levels of serum IFNα activity, as did the risk/risk andrisk/neutral genotype groups, and the risk/protective genotype group had similar serum IFNα
Niewold et al. Page 6
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

activity as the neutral/neutral genotype group. These data suggest that risk and protective IRF5haplotypes may have a dominant influence on serum IFNα activity when combined with aneutral haplotype and that the risk/protective haplotype combination may have a balancingeffect on serum IFNα activity.
We demonstrated that separation of patients by autoantibody status is important, since thedifferential effect of IRF5 genotype on serum IFNα activity was detectable only in patientswho were positive for either anti-RBP or anti-dsDNA and may have been obscured by higherbackground IFNα activity in patients who were positive for both categories of autoantibodies.It is interesting that the risk/risk and risk/neutral genotypes did not show significantly higherserum IFNα activity than the protective genotypes in the double-negative autoantibody group,despite having similar numbers of patients as the single-positive group. An effect of IRF5genotype on IFNα activity in the double-negative patients is not ruled out by this study,however, since IFNα levels were much lower in this group in general than in the single-positiveor double-positive patients, and greater numbers of patients may be required in order to detecta significant difference. The number of patients in the double-positive autoantibody group wassmall, and we were not able to exclude an association of IRF5 genotype with serum IFNα inthis group.
The increased ratio of median serum IFNα levels between the single-positive protective/protective and neutral/protective genotypes and the risk/risk and risk/neutral genotypes ascompared with the double-negative patients suggests the possibility of a gene–autoantibodyinteraction. In vitro models have shown that the addition of sera containing anti-RBP or anti-dsDNA antibodies to dendritic cells in culture results in brisk production of IFNα (16). Thismay result from the nucleic acid contained within these autoantibody–immune complexestriggering endosomal Toll-like receptors (TLRs) after uptake into cells via Fc receptors. Ifautoantibodies in SLE patient sera are required to see a differential effect of IRF5 genotype onserum IFNα activity, then IRF5 may be operative in SLE downstream of the activation ofendosomal TLRs by nucleic acid–containing autoantibody–immune complexes. If IFNα is theprotein mediator of SLE risk due to IRF5 genotype, then we would expect that if data fromexisting case–control genetic association studies are reanalyzed according to the autoantibodystrata used in the present study, the IRF5 SLE risk haplotype would show a higher odds ratiofor disease in single-positive patients than in double-negative patients. This may or may notbe the case in the double-positive autoantibody group, but either result in this group would alsobe of high interest.
Emerging data suggest that an insertion/deletion polymorphism in the promoter region of IRF5may play an important role in SLE susceptibility (15). When we reanalyzed our serum IFNαactivity data using the model proposed in the present study and using rs3807306 as a proxy forthe promoter insertion/deletion, we again saw significant differences in serum IFNα activityby IRF5 genotype in the single-positive autoantibody group. Linkage is strong between thepromoter insertion/deletion and previously described SLE risk variants in the 5′ region of thegene, including both rs3807306 and rs2004640, and therefore, the findings of this reanalysisshare many similarities with the findings of our initial analysis. However, the 2-marker analysisby rs3807306/rs10488631 genotype categorized the 6% frequency TCTA haplotype as arelatively lower SLE risk haplotype than did our previous analysis, and this new analysisallowed for an association of genotype in the 5′ region of IRF5 with high levels of IFNα, whichis independent of 3′ risk alleles. This result could suggest that the promoter insertion/deletionplays a larger role in serum IFNα activity than does the 5′ splice-site variation; however, futurein vitro and mechanistic studies are required to more definitively address this question.
High levels of serum IFNα activity show complex inheritance as a trait, and modeling thenumber of factors involved using relative recurrence-to-risk ratios suggests that 3–4
Niewold et al. Page 7
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

independent factors will be operative in a given SLE patient (7). Therefore, while the effect ofIRF5 genotype on serum IFNα activity appears modest and limited to certain autoantibodygroups in this study, we expect that a number of other genetic factors will also be important.It is likely that combinations of high IFNα–predisposing genetic variants will be required tofully manifest the high serum IFNα trait, and the other genetic factors that underlie high levelsof serum IFNα as an SLE risk factor are as yet unknown.
AcknowledgementsWe would like to thank Karen Onel, MD, and Kenan Onel MD, PhD, for their contribution in establishing the Hospitalfor Special Surgery Lupus Family Registry, and Gail Bruner, BSN, for her assistance in obtaining data and materialsfrom the Lupus Family Registry and Repository at Oklahoma Medical Research Foundation.
Dr. Niewold's work was supported by the NIH (grant T32-AR-07517 and Clinical Research Loan Repayment grantAI-071651 from the National Institute of Allergy and Infectious Diseases) and by an Arthritis Foundation PostdoctoralFellowship award. Dr. Espinoza's work was supported by the NIH (grant AR-053646). Dr. Harley's work was supportedby the Lupus Family Registry and Repository (NIH grant AR-62277) and by research grants from the NIH (AR-42460,AI-53747, AI-31584, DE-15223, RR-20143, AI-24717, AI-62629, AR-48940, and AR-49084), the US Departmentof Veterans Affairs, the Alliance for Lupus Research, and Rheuminations, Inc. Dr. Crow's work was supported byresearch grants from the NIH (R01-AI-059893 from the National Institute of Allergy and Infectious Diseases), theAlliance for Lupus Research, the Mary Kirkland Center for Lupus Research, and the Lupus Research Institute. TheHospital for Special Surgery Lupus Family Registry was supported by the Toys “R” Us Foundation and the SLEFoundation, Inc.
References1. Blanco P, Palucka AK, Gill M, Pascual V, Banchereau J. Induction of dendritic cell differentiation by
IFN-α in systemic lupus erythematosus. Science 2001;294:1540–3. [PubMed: 11711679]2. Hooks JJ, Moutsopoulos HM, Geis SA, Stahl NI, Decker JL, Notkins AL. Immune interferon in the
circulation of patients with autoimmune disease. N Engl J Med 1979;301:5–8. [PubMed: 449915]3. Baechler EC, Batliwalla FM, Karypis G, Gaffney PM, Ortmann WA, Espe KJ, et al. Interferon-
inducible gene expression signature in peripheral blood cells of patients with severe lupus. Proc NatlAcad Sci U S A 2003;100:2610–5. [PubMed: 12604793]
4. Kirou KA, Lee C, George S, Louca K, Peterson MG, Crow MK. Activation of the interferon-α pathwayidentifies a subgroup of systemic lupus erythematosus patients with distinct serologic features andactive disease. Arthritis Rheum 2005;52:1491–503. [PubMed: 15880830]
5. Ronnblom LE, Alm GV, Oberg KE. Possible induction of systemic lupus erythematosus by interferon-α treatment in a patient with a malignant carcinoid tumour. J Intern Med 1990;227:207–10. [PubMed:1690258]
6. Niewold TB, Swedler WI. Systemic lupus erythematosus arising during interferon-α therapy forcryoglobulinemic vasculitis associated with hepatitis C. Clin Rheumatol 2005;24:178–81. [PubMed:15565395]
7. Niewold TB, Hua J, Lehman TJ, Harley JB, Crow MK. High serum IFN-α activity is a heritable riskfactor for systemic lupus erythematosus. Genes Immun 2007;8:492–502. [PubMed: 17581626]
8. Barnes BJ, Moore PA, Pitha PM. Virus-specific activation of a novel interferon regulatory factor,IRF-5, results in the induction of distinct interferon α genes. J Biol Chem 2001;276:23382–90.[PubMed: 11303025]
9. Sigurdsson S, Nordmark G, Goring HH, Lindroos K, Wiman AC, Sturfelt G, et al. Polymorphisms inthe tyrosine kinase 2 and interferon regulatory factor 5 genes are associated with systemic lupuserythematosus. Am J Hum Genet 2005;76:528–37. [PubMed: 15657875]
10. Graham RR, Kyogoku C, Sigurdsson S, Vlasova IA, Davies LR, Baechler EC, et al. Three functionalvariants of IFN regulatory factor 5 (IRF5) define risk and protective haplotypes for human lupus.Proc Natl Acad Sci U S A 2007;104:6758–63. [PubMed: 17412832]
11. Graham RR, Kozyrev SV, Baechler EC, Reddy MV, Plenge RM, Bauer JW, et al. A commonhaplotype of interferon regulatory factor 5 (IRF5) regulates splicing and expression and is associated
Niewold et al. Page 8
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

with increased risk of systemic lupus erythematosus. Nat Genet 2006;38:550–5. [PubMed:16642019]
12. Jabs WJ, Hennig C, Zawatzky R, Kirchner H. Failure to detect antiviral activity in serum and plasmaof healthy individuals displaying high activity in ELISA for IFN-α and IFN-β. J Interferon CytokineRes 1999;19:463–9. [PubMed: 10386858]
13. Hua J, Kirou K, Lee C, Crow MK. Functional assay of type I interferon in systemic lupuserythematosus plasma and association with anti–RNA binding protein autoantibodies. ArthritisRheum 2006;54:1906–16. [PubMed: 16736505]
14. Kirou KA, Lee C, George S, Louca K, Papagiannis IG, Peterson MG, et al. Coordinate overexpressionof interferon-α–induced genes in systemic lupus erythematosus. Arthritis Rheum 2004;50:3958–67.[PubMed: 15593221]
15. Sigurdsson S, Goring HH, Kristjansdottir G, Milani L, Nordmark G, Sandling J, et al. Comprehensiveevaluation of the genetic variants of interferon regulatory factor 5 (IRF5) reveals a novel 5 bp lengthpolymorphism as strong risk factor for systemic lupus erythematosus. Hum Molec Genet. 2007E-pub ahead of print
16. Lovgren T, Eloranta ML, Bave U, Alm GV, Ronnblom L. Induction of interferon-α production inplasmacytoid dendritic cells by immune complexes containing nucleic acid released by necrotic orlate apoptotic cells and lupus IgG. Arthritis Rheum 2004;50:1861–72. [PubMed: 15188363]
Niewold et al. Page 9
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
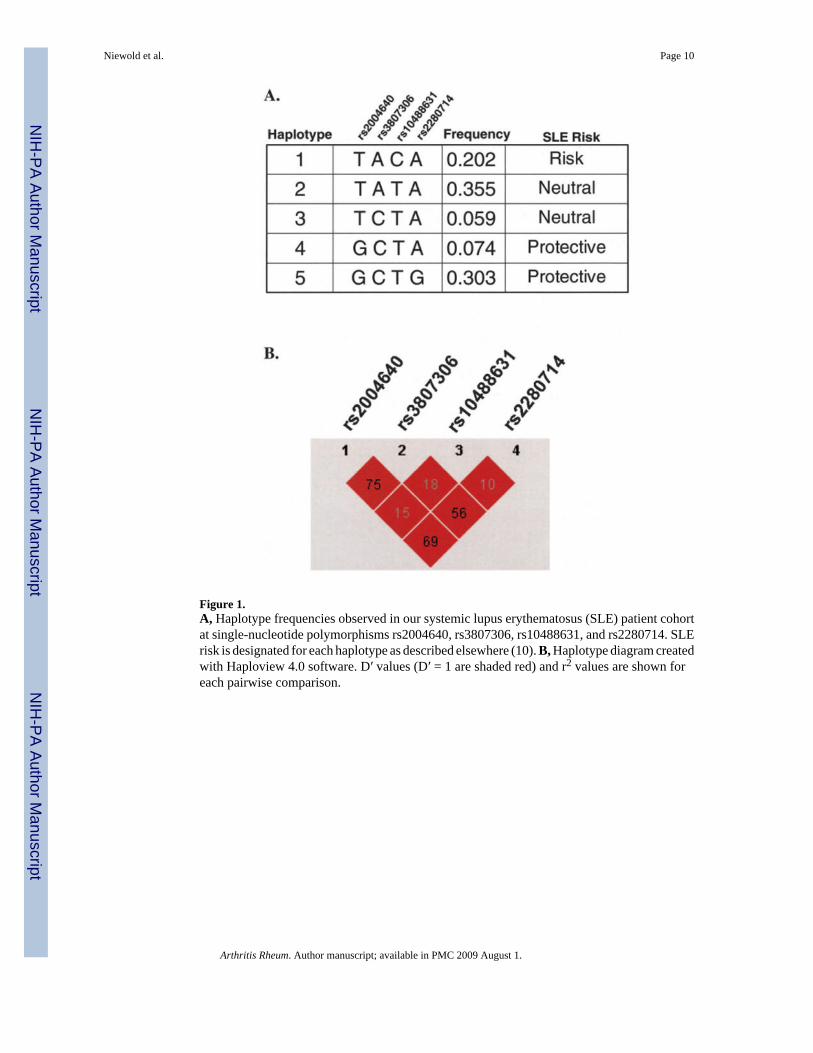
Figure 1.A, Haplotype frequencies observed in our systemic lupus erythematosus (SLE) patient cohortat single-nucleotide polymorphisms rs2004640, rs3807306, rs10488631, and rs2280714. SLErisk is designated for each haplotype as described elsewhere (10). B, Haplotype diagram createdwith Haploview 4.0 software. D′ values (D′ = 1 are shaded red) and r2 values are shown foreach pairwise comparison.
Niewold et al. Page 10
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
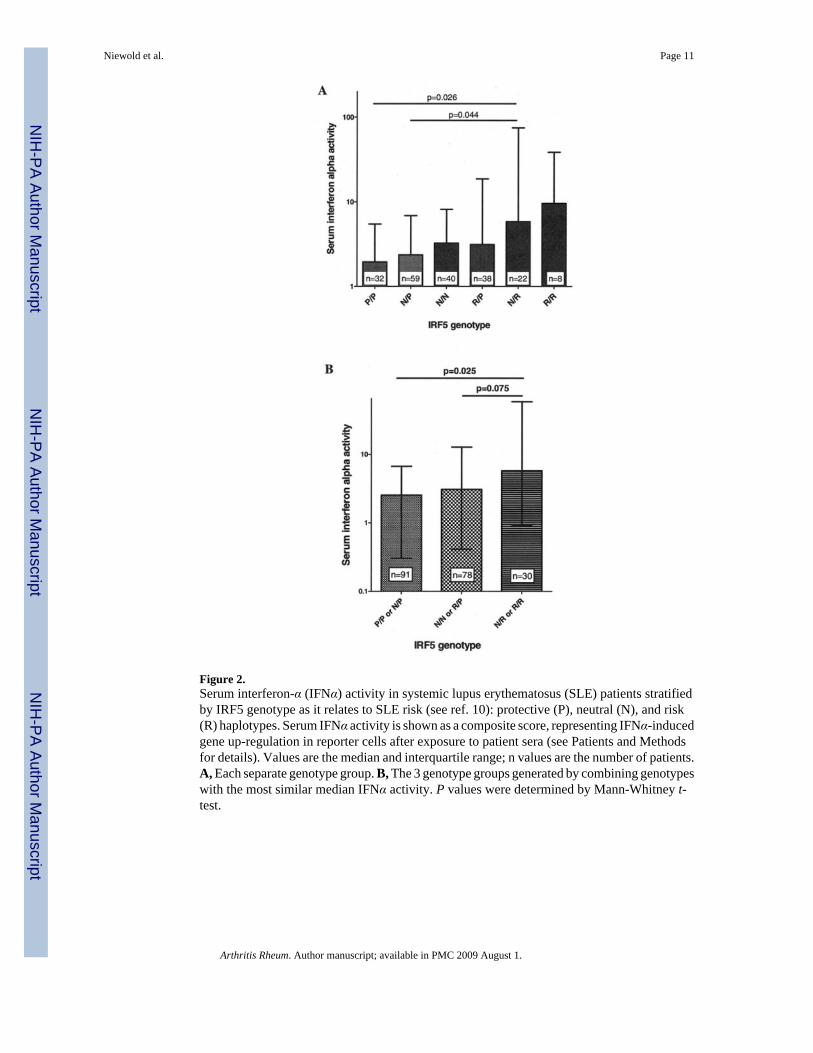
Figure 2.Serum interferon-α (IFNα) activity in systemic lupus erythematosus (SLE) patients stratifiedby IRF5 genotype as it relates to SLE risk (see ref. 10): protective (P), neutral (N), and risk(R) haplotypes. Serum IFNα activity is shown as a composite score, representing IFNα-inducedgene up-regulation in reporter cells after exposure to patient sera (see Patients and Methodsfor details). Values are the median and interquartile range; n values are the number of patients.A, Each separate genotype group. B, The 3 genotype groups generated by combining genotypeswith the most similar median IFNα activity. P values were determined by Mann-Whitney t-test.
Niewold et al. Page 11
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
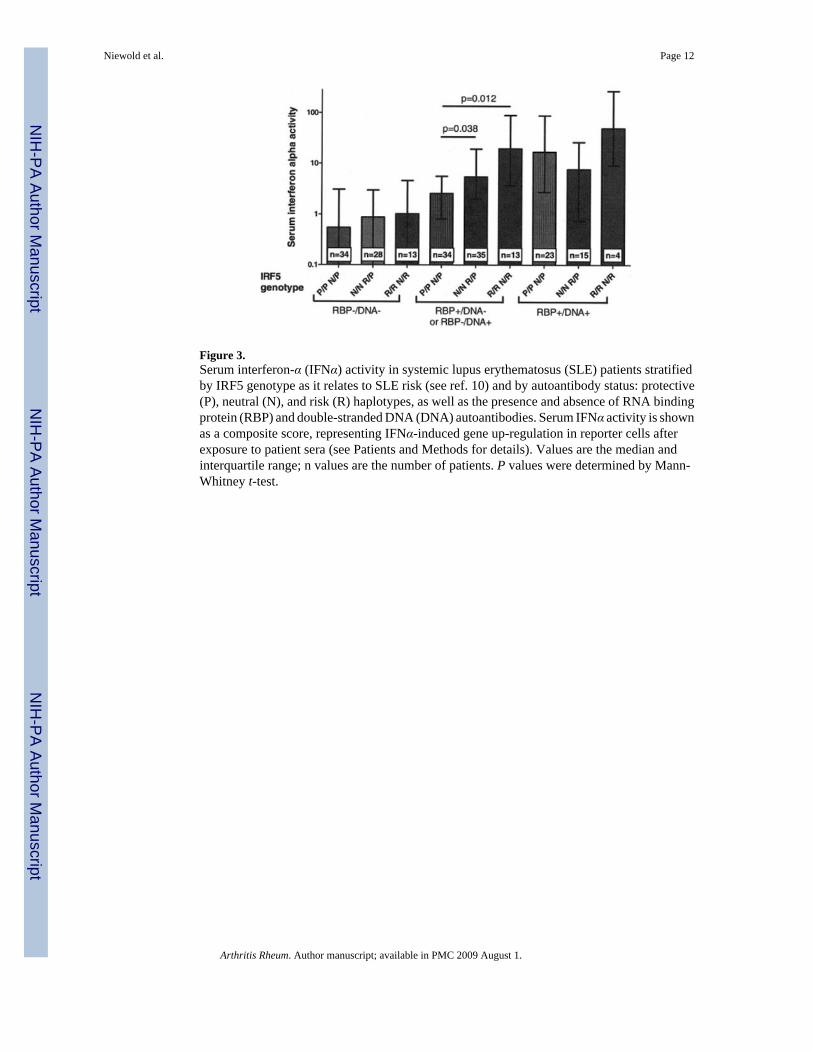
Figure 3.Serum interferon-α (IFNα) activity in systemic lupus erythematosus (SLE) patients stratifiedby IRF5 genotype as it relates to SLE risk (see ref. 10) and by autoantibody status: protective(P), neutral (N), and risk (R) haplotypes, as well as the presence and absence of RNA bindingprotein (RBP) and double-stranded DNA (DNA) autoantibodies. Serum IFNα activity is shownas a composite score, representing IFNα-induced gene up-regulation in reporter cells afterexposure to patient sera (see Patients and Methods for details). Values are the median andinterquartile range; n values are the number of patients. P values were determined by Mann-Whitney t-test.
Niewold et al. Page 12
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
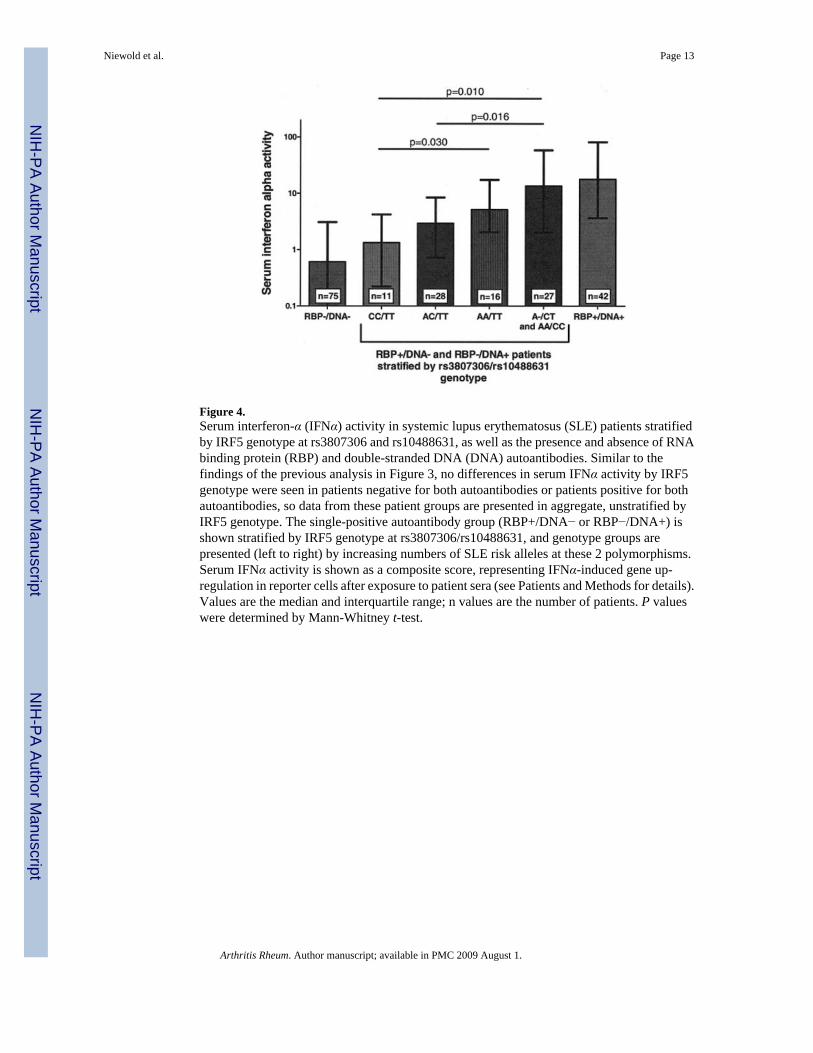
Figure 4.Serum interferon-α (IFNα) activity in systemic lupus erythematosus (SLE) patients stratifiedby IRF5 genotype at rs3807306 and rs10488631, as well as the presence and absence of RNAbinding protein (RBP) and double-stranded DNA (DNA) autoantibodies. Similar to thefindings of the previous analysis in Figure 3, no differences in serum IFNα activity by IRF5genotype were seen in patients negative for both autoantibodies or patients positive for bothautoantibodies, so data from these patient groups are presented in aggregate, unstratified byIRF5 genotype. The single-positive autoantibody group (RBP+/DNA− or RBP−/DNA+) isshown stratified by IRF5 genotype at rs3807306/rs10488631, and genotype groups arepresented (left to right) by increasing numbers of SLE risk alleles at these 2 polymorphisms.Serum IFNα activity is shown as a composite score, representing IFNα-induced gene up-regulation in reporter cells after exposure to patient sera (see Patients and Methods for details).Values are the median and interquartile range; n values are the number of patients. P valueswere determined by Mann-Whitney t-test.
Niewold et al. Page 13
Arthritis Rheum. Author manuscript; available in PMC 2009 August 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript