association of il17rb gene polymorphism with asthma
TRANSCRIPT

DOI 10.1378/chest.08-1595 2009;135;1173-1180; Prepublished online December 31, 2008;Chest
Choon-Sik Park and Hyoung Doo ShinYoung-Mok Lee, Ki-Up Kim, Soo-Taek Uh, Ju Ock Na, Yong-Hoon Kim,Kim, Hun Soo Chang, TaiYoun Rhim, Jong-Sook Park, An-Soo Jang, Ji-Sun Jung, Byung Lae Park, Hyun Sub Cheong, Joon Seol Bae, Ji-Hye With Asthma
Gene PolymorphismIL-17RBAssociation of
http://www.chestjournal.org/content/135/5/1173.full.html
and services can be found online on the World Wide Web at: The online version of this article, along with updated information
) ISSN:0012-3692http://www.chestjournal.org/site/misc/reprints.xhtml(of the copyright holder.may be reproduced or distributed without the prior written permission Northbrook IL 60062. All rights reserved. No part of this article or PDFby the American College of Chest Physicians, 3300 Dundee Road,
2007Physicians. It has been published monthly since 1935. Copyright CHEST is the official journal of the American College of Chest
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from

Association of IL-17RB GenePolymorphism With Asthma*
Ji-Sun Jung, BS;† Byung Lae Park, PhD;† Hyun Sub Cheong, MS;Joon Seol Bae, PhD; Ji-Hye Kim, MS; Hun Soo Chang, PhD;TaiYoun Rhim, PhD; Jong-Sook Park, MD; An-Soo Jang, MD;Young-Mok Lee, MD; Ki-Up Kim, MD; Soo-Taek Uh, MD; Ju Ock Na, MD;Yong-Hoon Kim, MD; Choon-Sik Park, MD, PhD; and Hyoung Doo Shin, PhD
Background: Interleukin (IL)-17E is a member of the IL-17 family, which induces IL-4, IL-5,IL-13, and eotaxin in experimental animals via IL-17 receptor B (IL-17RB). The activation ofIL-17RB amplifies allergic-type inflammatory responses by inducing Jun kinase (or JNK), p38mitogen-activated protein kinase (or MAPK), and nuclear factor-�B.Objectives: We examined the association of polymorphisms in the IL-17RB gene with asthmasusceptibility and investigated the effects of those polymorphisms on the transcription of variousIL-17RB isoforms.Methods: In total, 954 asthmatic patients or 265 healthy control subjects were screened for polymor-phisms in IL-17RB by single-base extension. The messenger RNA expression IL-17RB in B-cell linesderived from patients was also measured by reverse transcription-polymerase chain reaction.Results: Direct sequencing of 24 unrelated Korean DNA samples revealed 18 genetic variants, includingfour insertion/deletions and 14 single-nucleotide polymorphisms (SNPs). Six of the SNPs (�1465G>A,�5661G>A, �6297T>C �Y123Y�, �13797C>T, �18661C>T, and �18965G>A) were used to screen alarger group of subjects. Intronic polymorphism �5661G>A was significantly associated with thedevelopment of asthma (p � 0.001); moreover, a minor allele of IL-17RB �5661G>A appeared at a lowerfrequency in the asthmatic patients than in the healthy control subjects (0.13 vs 0.19, respectively). TheIL-17RB messenger RNA expression in B cells homozygous for IL-17RB� 5661GG was significantlyhigher than that in B cells homozygous for IL-17RB�5661AA (p � 0.002).Conclusions: A rare allele of IL-17RB �5661G>A may have a protective role against the development ofasthma via regulation at the level of transcription. The SNPs identified in this study may be used to developmarkers to assess the risk of asthma. (CHEST 2009; 135:1173–1180)
Abbreviations: AHR � airway hyperreactivity; GAPDH � glyceraldehyde-3-phosphate dehydrogenase; IL � interleukin; IL-17RB � interleukin-17 receptor B; LD � linkage disequilibrium; OR � odds ratio; PC20 � provocative concentration of metha-choline causing a 20% fall in FEV1; PCR � polymerase chain reaction; RT � reverse transcription; SNP � single-nucleotidepolymorphism; Th � T helper
A sthma and its phenotypes are complex traitsinduced by interactions between the surround-
ing environment and multiple disease susceptibil-ity factors.1 Asthma has been recognized as aT-helper (Th) type 2-related disease with a cyto-kine response profile that includes interleukin(IL)-4, IL-5, and IL-13.2,3 These cytokines play animportant role in the coordination and persistenceof the airway inflammatory process in asthmapatients.4
Treatment with or transgenic expression of IL-17E (IL-25) in mice was found to induce Th2-type
cytokines, notably IL-5 and IL-13, and chemokines,such as eotaxin (CCL11), along with eosinophilia,elevated levels of IgE, mucus secretion, and airwayhyperreactivity (AHR).5–7 Furthermore, the block-ade of IL-25 in an experimental model of allergicasthma prevented AHR.8 IL-25R, which is alsocalled IL-17 receptor B (IL-17RB) IL-17Rh1, andEvi27, is a 56-kd single-transmembrane protein withhomology to IL-17R. Th2 cytokines induce theexpression of IL-17RB in antigen-presenting cells.9The activation of IL-17RB induces activation of Junkinase (or JNK), p38 mitogen-activated protein kinase
CHEST Original ResearchASTHMA
www.chestjournal.org CHEST / 135 / 5 / MAY, 2009 1173
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from

(or MAPK), and nuclear factor-�B,10 which regulatethe expression of the genes responsible for the inflam-matory and remodeling processes in asthma.11 Letuveet al12 have demonstrated that human primary lungfibroblasts constitutively express IL-17BR. This sug-gests that the IL-17E derived from Th2 T cells, mast cells,and epithelial cells is capable of amplifying allergic-typeinflammatory responses through its effect on othercell types via IL-17RB.13,14 Human IL-17RB(mendelian inheritance in man No. 605458) islocated on chromosome 3p21.1.15 The region aroundchromosome 3p21 is associated with the risk of asth-ma16 and atopy.17 Given the biological effects of IL-17RB and its linkage to chromosome 3p21, geneticvariants of IL-17RB may play a role in Th2-mediatedimmune reactions, including asthma.
Materials and Methods
Subjects
Subjects were recruited from the Genome Research Center forAllergy and Respiratory Diseases at Soonchunhyang University,Bucheon, Seoul, Korea, and Chunan Hospital, Chunan, Korea.All patients had received a diagnosis of asthma by a physician andmet the definition of asthma set forth in the Global Initiative forAsthma guidelines.18 All patients had a history of dyspnea andwheezing during the previous 12 months plus one of the follow-ing: (1) a � 15% increase in FEV1 or a � 12% increase plus 200mL after the inhalation of a short-acting bronchodilator; (2) aprovocative concentration of methacholine causing a 20% fall inFEV1 (PC20) of � 10 mg/mL; and (3) a � 20% increase in FEV1
after 2 weeks of treatment with inhaled steroids and long-actingbronchodilators. The healthy subjects were recruited from thespouses of the patients and from members of the generalpopulation who answered negatively to a screening questionnairefor respiratory symptoms19 and had an FEV1 of � 75% predicted,a PC20 of � 10 mg/mL, and normal findings on a simple chestradiogram. Twenty-four common inhaled allergens (eg, dustmites �Dermatophagoides farinae and Dermatophagoides ptero-nyssinus�, cat fur, dog fur, cockroaches, grasses, trees, andragweed pollen) were used for a skin-prick test. Total IgE levelwas measured (ImmunoCAP System; Pharmacia Diagnostics;Uppsala, Sweden). Atopy was defined as having a wheal reactionequal to or greater than that from histamine or 3 mm in diameter.The subjects and study protocols were approved by the Institu-tional Review Board of Soonchunhyang University Hospital.
Sequence Analysis of Human IL-17RB
The entire IL-17RB gene, including the promoter region (approxi-mately 1.5 kb), was sequenced from 24 DNA samples to identifysingle-nucleotide polymorphisms (SNPs) �ABI PRISM 3700 DNAAnalyzer; Applied Biosystems; Foster City, CA�. The primer sets usedfor amplification and sequencing were designed based on sequenceinformation from GenBank (NC_000003). Information regarding theprimers is given online in supplementary Table 1 (http://www.snp-genetics.com/user/additional_list.asp).
Genotyping of IL-17RB Polymorphisms
Among the 18 polymorphisms discovered, six (�1465G�A in thepromoter, �5661G�A in intron 4, �6297T�C �Y123Y� in exon 5,�13797C�T in intron 10, and �18661C�T and �18965G�A inthe 3-UTR) were selected based on the following general criteria:�10% of the minor allele frequency and a polymorphism amongtight linkage disequilibriums (LDs) �D �1; r2 � 0.85� (supplemen-tary Tables 2 and 3 �http://www.snp-genetics.com/user/additional_list.asp�). For the purpose of genotyping the six SNPs, TaqManprimers and probes were designed.20 A design system (PrimerExpress; Applied Biosystems) was used to design both the polymer-ase chain reaction (PCR) primers and MGB TaqMan probes(supplementary Table 4). One allelic probe was labeled with 6-carboxyfluorescein dye, whereas the other was labeled with thefluorescent dye VIC (Perkin-Elmer Applied Biosystems). PCRswere run using TaqMan Universal Master Mix (Applied Biosystems)containing PCR primers and TaqMan MGB probes. The reactionswere performed in a 384-well format using a total reaction volumeof 5 L with 20 ng of genomic DNA. The plates were then placedin a thermal cycler (PE 9700 Thermal Cycler; PerkinElmer;Waltham, MA) and heated to 50°C for 2 min and 95°C for 10 min,followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. The plateswere then transferred to a sequencer (Prism 7900HT SequenceDetection System; Applied Biosystems), and the fluorescence inten-sity was read. The fluorescence data were analyzed using automatedsoftware (SDS, version 2.1; Applied Biosystems). There were 24samples used in a duplicate genotyping test for a positive control,and 12 blanks were used for a negative control.
Semi-Quantitation of Messenger RNA Expression by ReverseTranscription-PCR of Epstein-Barr Virus Transformation of theB Cells
B-cell lines were established from the peripheral blood mono-nuclear cells by transformation with Epstein-Barr virus from thesupernatant of the B95–8 cell line (VR-1492 �American TypeCulture Collection, CRL-10624�). Total RNA was extracted usinga reagent (TRI Reagent; Molecular Research Center; Cincinnati,
*From the Genome Research Center for Allergy and RespiratoryDisease (Drs. Chang, J.-S. Park, Jang, and C.-S. Park, and Ms.Jung and Ms. J.-H. Kim), Soonchunhyang University BucheonHospital, Bucheon, Korea; the Division of Allergy and Respira-tory Medicine (Drs. Lee, K.-U. Kim, and Uh), SoonchunhyangUniversity Seoul Hospital, Seoul, Korea; the Division of Allergyand Respiratory Medicine (Drs. Na and Y.-H. Kim), Soonchun-hyang University Chunan Hospital, Chunan, Korea; HanyangUniversity (Dr. Rhim), Seoul, Korea; the Department of GeneticEpidemiology (Drs. B.L. Park and Bae, and Mr. Cheong), SNPGenetics, Inc, Seoul, Korea; and the Department of Life Science(Dr. Shin), Sogang University, Seoul, Korea.†Ms. Jung and Dr. B.L. Park contributed equally to this work asthe first coauthors.This study was supported by the grants of the Korea Health 21R&D Project (Ministry of Health, Welfare and Family Affairs,Republic of Korea, A010249). B-cell lines were provided fromthe biobank of Soonchunhyang University Bucheon Hospital.The authors have reported to the ACCP that no significantconflicts of interest exist with any companies/organizations whoseproducts or services may be discussed in this article.Manuscript received June 27, 2008; revision accepted November14, 2008.Reproduction of this article is prohibited without written permissionfrom the American College of Chest Physicians (www.chestjournal.org/site/misc/reprints.xhtml).Correspondence to: Choon-Sik Park, MD, PhD, Division ofAllergy and Respiratory Medicine, Department of Internal Med-icine, Soonchunhyang University Bucheon Hospital, 1174, Jung-Dong, Wonmi-Gu, Bucheon-Si, Kyeonggi-Do, Korea 420-020;e-mail: [email protected]: 10.1378/chest.08-1595
1174 Original Research
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from

OH) and a modified version of the guanidium thiocyanate-phenolchloroform extraction method.21 DNA contamination was elimi-nated by treatment with 1 L of RNase-free DNase I for 15 minat 37°C. The DNase I-treated RNA was then heated to 65°C for5 min with 1 L of oligo-deoxythymidine-15 primer (500 g/mL)and 10 mmol/L deoxyribonucleoside triphosphates, then quicklychilled on ice. Pellets were incubated at 42°C for 2 min with 0.1mmol/L dithiothreitol and 1 L (200 U/L) of a reverse tran-scriptase (SuperScript II Reverse Transcriptase; Invitrogen;Carlsbad, CA) at 42°C for 50 min, and then heat inactivated at70°C for 15 min. After reverse transcription (RT), complemen-tary DNA was dispensed into tubes containing primers specificfor human IL-17RB and glyceraldehyde-3-phosphate dehydroge-nase (GAPDH) genes. The primer sequences were summarizedin Table 2.
Amplification was performed using the following program: 30cycles of 1 min at 95°C, 1 min at 52°C, and 1 min at 72°C, withan initial denaturation at 95°C for 3 min and a final extension at72°C for 7 min. The amplified products (546 and 300 base pairsfor IL-17RB and GAPDH, respectively) underwent electrophore-sis on a 1% agarose gel and were visualized by ethidium bromidestaining. The PCR products were then analyzed by direct se-quencing using the PCR primers and an analyzer (ABI PRISM3700 DNA Analyzer). The sequences were compared using theBLAST search program from the National Institutes of Health.
Statistical Analysis
LD was measured as the Lewontin D and r2.22,23 Thehaplotype of each individual was inferred using the EM algorithm
(PHASE, version 2.0) developed by Stephens et al.24 Theassociation of the various IL-17RB genotypes and haplotypes withthe risk of asthma was analyzed using logistic regression models,whereas controlling for age (continuous value), sex (male, 0;female, 1), smoking status (nonsmoker, 0; ex-smoker, 1; smoker,2), and atopy (positive, 0; negative, 1) as covariates. All statisticalanalyses were performed using two statistical software packages(SAS, version 9.1; SAS Institute; Cary, NC; or SPSS, version 11;SPSS Inc; Chicago, IL). The p values were corrected for multiplecomparisons using the Bonferroni correction test. The RT-PCRintensity data are expressed as the mean � SEM. The Mann-Whitney U test was applied to compare differences betweensamples. A p value � 0.05 was considered to be significant.
Results
Characteristics of the Study Subjects
The clinical characteristics of the study subjects aresummarized in Table 1. We found significant differ-ences in mean age, prevalence of smoking, and preva-lence of atopy between the healthy control subjects andasthmatic patients (p � 0.001). FVC, FEV1, and PC20values were significantly lower in asthmatic patientsthan in healthy control subjects (p � 0.001). IgE levelsand eosinophil numbers were significantly higher inasthmatic patients (p � 0.001).
Association of SNPs in IL-17RB With Risk ofAsthma
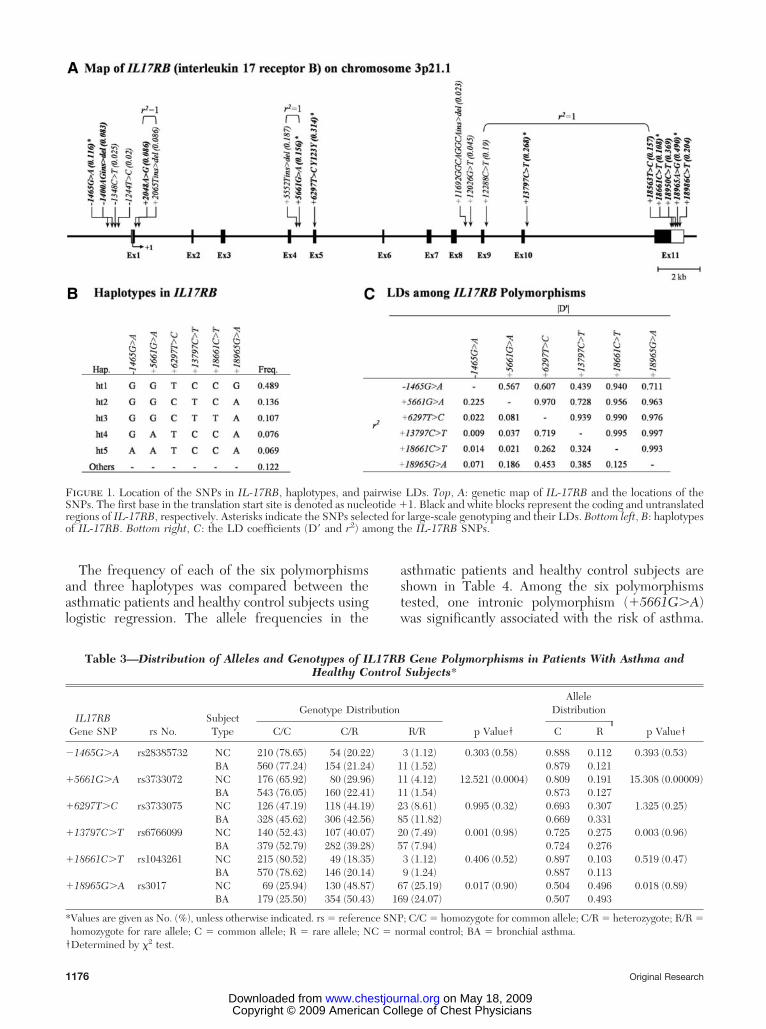
Direct sequencing of 24 unrelated Korean DNAsamples using 17 primer sets revealed 18 geneticvariants (four insertion/deletions and 14 SNPs) withinthe exons and flanking regions of IL-17RB, as follows:four SNPs in the promoter region; one SNP in exon 5;eight intronic SNPs; and five SNPs in the 3-UTR (Fig1, top, A, and supplementary Table 2). Pairwise com-parisons of the SNPs revealed three sets of absoluteLDs (D � 1 and r2 � 1; �2048A�G:�2065Tins�del, �5552Tins/del:�5661G�A, and �12288C�T:�18563T�C); several complete LDs (D � 1 andr2 � 1) were also found (Fig 1, bottom right, C). Six SNPswere selected for large-scale genotyping. Haplotypes ofIL-17RB were constructed using software (PHASE),and three major haplotypes with � 10% of minorallele frequency were used for further statisticalanalysis. The genotype distributions of the sixSNPs were in Hardy-Weinberg equilibrium for allsubjects (p � 0.05) �Table 3�.
Table 1—The Clinical Profiles for Asthmatic Patientsand Healthy Control Subjects*
Clinical Profiles
Healthy ControlSubjects
(n � 265)
AsthmaticPatients
(n � 954)
Age, yr 42 (8–80) 45 (8–76)†Age at onset, yr . 37 (1–75)Male sex, % 42.6 38.6Smoking status, %
Smoker 18.4 16.4Ex-smoker 11.4 16.9†
Atopy, % 33.2 57.8†FVC, % predicted 93.3 � 0.8 83.1 � 0.6†FEV1, % predicted 102.3 � 0.9 79.0 � 0.7†PC20, mg/mL 24.2 � 0.2 4.17 � 0.2†BMI, kg/m2 23.8 � 0.2 24.2 � 0.1Blood eosinophils, % 2.5 � 0.1 5.5 � 0.2†Total IgE concentration,
IU/mL118.7 � 10.5 385.9 � 31.4†
*Values are given as No. (range) or mean � SE, unless otherwiseindicated. BMI � body mass index.
†p � 0.001 (obtained using a t test or 2 test).
Table 2—PCR Primer Sequences Used for Semi-Quantification of Messenger RNA
Gene Locus Position Definition Primer Sequence (5-3)
IL-17RB � 2117/� 2134 Exon 2 Forward CGTTCAATGTGGCTCTGA� 10358/� 10374 Exon 7 Reverse CAGGGGAGTGGTTGTGA
GAPDH � 2151/� 2170 Exon 5 Forward CGTCTTCACCACCATGGAGA� 2806/� 2825 Exon 8 Reverse CGGCCATCACGCCACAGTTT
www.chestjournal.org CHEST / 135 / 5 / MAY, 2009 1175
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from
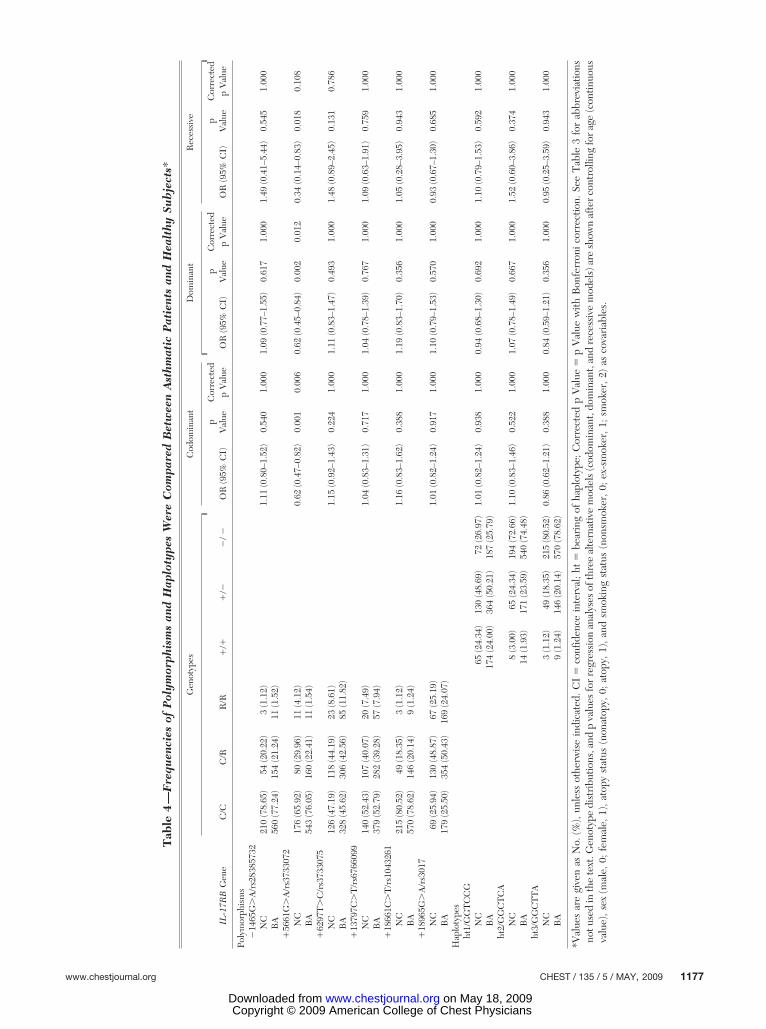
The frequency of each of the six polymorphismsand three haplotypes was compared between theasthmatic patients and healthy control subjects usinglogistic regression. The allele frequencies in the
asthmatic patients and healthy control subjects areshown in Table 4. Among the six polymorphismstested, one intronic polymorphism (�5661G�A)was significantly associated with the risk of asthma.
Figure 1. Location of the SNPs in IL-17RB, haplotypes, and pairwise LDs. Top, A: genetic map of IL-17RB and the locations of theSNPs. The first base in the translation start site is denoted as nucleotide �1. Black and white blocks represent the coding and untranslatedregions of IL-17RB, respectively. Asterisks indicate the SNPs selected for large-scale genotyping and their LDs. Bottom left, B: haplotypesof IL-17RB. Bottom right, C: the LD coefficients (D and r2) among the IL-17RB SNPs.
Table 3—Distribution of Alleles and Genotypes of IL17RB Gene Polymorphisms in Patients With Asthma andHealthy Control Subjects*
IL17RBGene SNP rs No.
SubjectType
Genotype Distribution
p Value†
AlleleDistribution
p Value†C/C C/R R/R C R
�1465G�A rs28385732 NC 210 (78.65) 54 (20.22) 3 (1.12) 0.303 (0.58) 0.888 0.112 0.393 (0.53)BA 560 (77.24) 154 (21.24) 11 (1.52) 0.879 0.121
�5661G�A rs3733072 NC 176 (65.92) 80 (29.96) 11 (4.12) 12.521 (0.0004) 0.809 0.191 15.308 (0.00009)BA 543 (76.05) 160 (22.41) 11 (1.54) 0.873 0.127
�6297T�C rs3733075 NC 126 (47.19) 118 (44.19) 23 (8.61) 0.995 (0.32) 0.693 0.307 1.325 (0.25)BA 328 (45.62) 306 (42.56) 85 (11.82) 0.669 0.331
�13797C�T rs6766099 NC 140 (52.43) 107 (40.07) 20 (7.49) 0.001 (0.98) 0.725 0.275 0.003 (0.96)BA 379 (52.79) 282 (39.28) 57 (7.94) 0.724 0.276
�18661C�T rs1043261 NC 215 (80.52) 49 (18.35) 3 (1.12) 0.406 (0.52) 0.897 0.103 0.519 (0.47)BA 570 (78.62) 146 (20.14) 9 (1.24) 0.887 0.113
�18965G�A rs3017 NC 69 (25.94) 130 (48.87) 67 (25.19) 0.017 (0.90) 0.504 0.496 0.018 (0.89)BA 179 (25.50) 354 (50.43) 169 (24.07) 0.507 0.493
*Values are given as No. (%), unless otherwise indicated. rs � reference SNP; C/C � homozygote for common allele; C/R � heterozygote; R/R �homozygote for rare allele; C � common allele; R � rare allele; NC � normal control; BA � bronchial asthma.
†Determined by 2 test.
1176 Original Research
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from
Tab
le4
—F
requ
enci
esof
Pol
ymor
phis
ms
and
Hap
loty
pes
Wer
eC
ompa
red
Bet
wee
nA
sthm
atic
Pat
ient
san
dH
ealt
hySu
bje
cts*
IL-1
7RB
Gen
e
Gen
otyp
esC
odom
inan
tD
omin
ant
Rec
essi
ve
C/C
C/R
R/R
�/�
�/�
�/�
OR
(95%
CI)
pV
alue
Cor
rect
edp
Val
ueO
R(9
5%C
I)p
Val
ueC
orre
cted
pV
alue
OR
(95%
CI)
pV
alue
Cor
rect
edp
Val
ue
Poly
mor
phis
ms
�14
65G
�A
/rs2
8385
732
NC
210
(78.
65)
54(2
0.22
)3
(1.1
2)1.
11(0
.80–
1.52
)0.
540
1.00
01.
09(0
.77–
1.55
)0.
617
1.00
01.
49(0
.41–
5.44
)0.
545
1.00
0B
A56
0(7
7.24
)15
4(2
1.24
)11
(1.5
2)�
5661
G�
A/r
s373
3072
NC
176
(65.
92)
80(2
9.96
)11
(4.1
2)0.
62(0
.47–
0.82
)0.
001
0.00
60.
62(0
.45–
0.84
)0.
002
0.01
20.
34(0
.14–
0.83
)0.
018
0.10
8B
A54
3(7
6.05
)16
0(2
2.41
)11
(1.5
4)�
6297
T�
C/r
s373
3075
NC
126
(47.
19)
118
(44.
19)
23(8
.61)
1.15
(0.9
2–1.
43)
0.22
41.
000
1.11
(0.8
3–1.
47)
0.49
31.
000
1.48
(0.8
9–2.
45)
0.13
10.
786
BA
328
(45.
62)
306
(42.
56)
85(1
1.82
)�
1379
7C�
T/r
s676
6099
NC
140
(52.
43)
107
(40.
07)
20(7
.49)
1.04
(0.8
3–1.
31)
0.71
71.
000
1.04
(0.7
8–1.
39)
0.76
71.
000
1.09
(0.6
3–1.
91)
0.75
91.
000
BA
379
(52.
79)
282
(39.
28)
57(7
.94)
�18
661C
�T
/rs1
0432
61N
C21
5(8
0.52
)49
(18.
35)
3(1
.12)
1.16
(0.8
3–1.
62)
0.38
81.
000
1.19
(0.8
3–1.
70)
0.35
61.
000
1.05
(0.2
8–3.
95)
0.94
31.
000
BA
570
(78.
62)
146
(20.
14)
9(1
.24)
�18
965G
�A
/rs3
017
NC
69(2
5.94
)13
0(4
8.87
)67
(25.
19)
1.01
(0.8
2–1.
24)
0.91
71.
000
1.10
(0.7
9–1.
53)
0.57
01.
000
0.93
(0.6
7–1.
30)
0.68
51.
000
BA
179
(25.
50)
354
(50.
43)
169
(24.
07)
Hap
loty
pes
ht1/
GG
TC
CG
NC
65(2
4.34
)13
0(4
8.69
)72
(26.
97)
1.01
(0.8
2–1.
24)
0.93
81.
000
0.94
(0.6
8–1.
30)
0.69
21.
000
1.10
(0.7
9–1.
53)
0.59
21.
000
BA
174
(24.
00)
364
(50.
21)
187
(25.
79)
ht2/
GG
CT
CA
NC
8(3
.00)
65(2
4.34
)19
4(7
2.66
)1.
10(0
.83–
1.46
)0.
522
1.00
01.
07(0
.78–
1.49
)0.
667
1.00
01.
52(0
.60–
3.86
)0.
374
1.00
0B
A14
(1.9
3)17
1(2
3.59
)54
0(7
4.48
)ht
3/G
GC
TT
AN
C3
(1.1
2)49
(18.
35)
215
(80.
52)
0.86
(0.6
2–1.
21)
0.38
81.
000
0.84
(0.5
9–1.
21)
0.35
61.
000
0.95
(0.2
5–3.
59)
0.94
31.
000
BA
9(1
.24)
146
(20.
14)
570
(78.
62)
*Val
ues
are
give
nas
No.
(%),
unle
ssot
herw
ise
indi
cate
d.C
I�
conf
iden
cein
terv
al;h
t�be
arin
gof
hapl
otyp
e;C
orre
cted
pV
alue
�p
Val
uew
ithB
onfe
rron
icor
rect
ion.
See
Tab
le3
for
abbr
evia
tions
notu
sed
inth
ete
xt.G
enot
ype
dist
ribu
tions
,and
pva
lues
for
regr
essi
onan
alys
esof
thre
eal
tern
ativ
em
odel
s(c
odom
inan
t,do
min
ant,
and
rece
ssiv
em
odel
s)ar
esh
own
afte
rco
ntro
lling
for
age
(con
tinuo
usva
lue)
,sex
(mal
e,0;
fem
ale,
1),a
topy
stat
us(n
onat
opy,
0;at
opy,
1),a
ndsm
okin
gst
atus
(non
smok
er,0
;ex-
smok
er,1
;sm
oker
,2)
asco
vari
able
s.
www.chestjournal.org CHEST / 135 / 5 / MAY, 2009 1177
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from
The frequency of minor allele (�IL-17RB�5661G�A) in the asthmatic group was lower thanthat in the healthy control group (0.13 vs 0.19,respectively). The frequency of a rare allele (IL-17RB�5661G�A) was significantly lower in theasthmatic patients, regardless of the analysis mode(codominant mode: odds ratio �OR�, 0.62; p � 0.001;dominant mode, OR, 0.62; p � 0.002; recessivemode, OR, 0.34; p � 0.018). After the Bonferronicorrection, the difference remained significant(p � 0.006 in the codominant mode; p � 0.012 inthe dominant mode). Thus, a rare allele of IL-17RB�5661G�A may have a protective effectagainst the development of asthma.
Comparison of the Messenger RNA Expression ofIL-17RB Between �IL-17RB�5661GG and AA
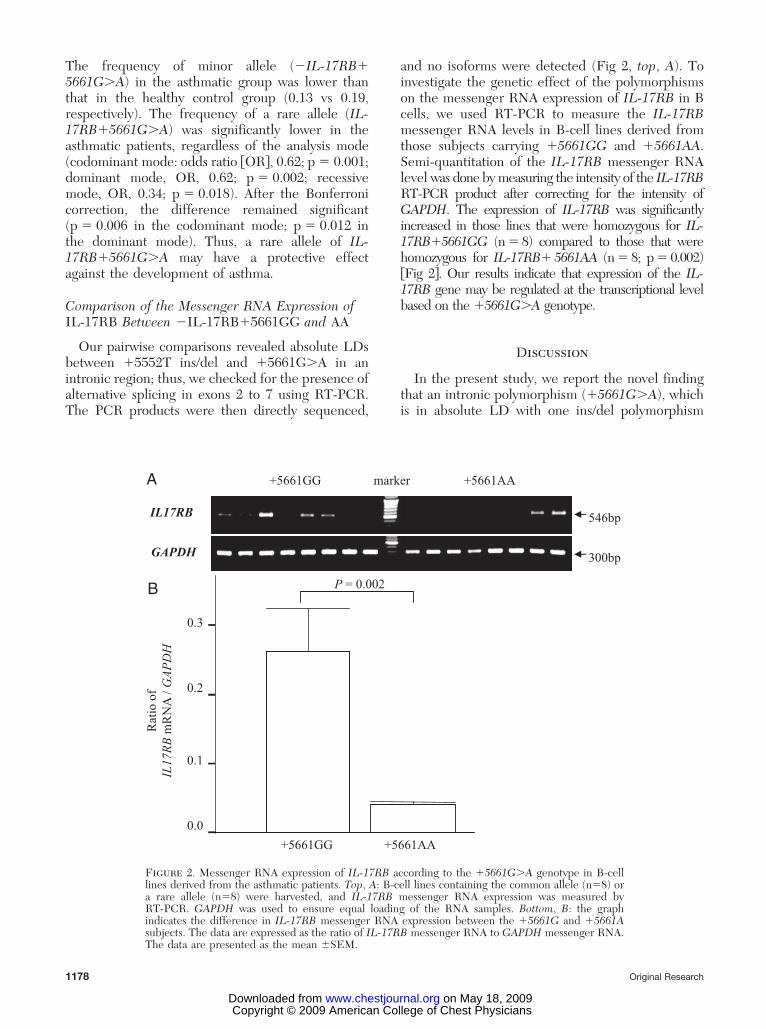
Our pairwise comparisons revealed absolute LDsbetween �5552T ins/del and �5661G�A in anintronic region; thus, we checked for the presence ofalternative splicing in exons 2 to 7 using RT-PCR.The PCR products were then directly sequenced,
and no isoforms were detected (Fig 2, top, A). Toinvestigate the genetic effect of the polymorphismson the messenger RNA expression of IL-17RB in Bcells, we used RT-PCR to measure the IL-17RBmessenger RNA levels in B-cell lines derived fromthose subjects carrying �5661GG and �5661AA.Semi-quantitation of the IL-17RB messenger RNAlevel was done by measuring the intensity of the IL-17RBRT-PCR product after correcting for the intensity ofGAPDH. The expression of IL-17RB was significantlyincreased in those lines that were homozygous for IL-17RB�5661GG (n � 8) compared to those that werehomozygous for IL-17RB� 5661AA (n � 8; p � 0.002)�Fig 2�. Our results indicate that expression of the IL-17RB gene may be regulated at the transcriptional levelbased on the �5661G�A genotype.
Discussion
In the present study, we report the novel findingthat an intronic polymorphism (�5661G�A), whichis in absolute LD with one ins/del polymorphism
A
B
Rat
io o
f
IL17RB
mR
NA
/ GAPDH
0.0
0.1
0.2
0.3
IL17RB
GAPDH
marker AA1665+GG1665+
546bp
300bp
P = 0.002
+5661GG +5661AA
Figure 2. Messenger RNA expression of IL-17RB according to the �5661G�A genotype in B-celllines derived from the asthmatic patients. Top, A: B-cell lines containing the common allele (n�8) ora rare allele (n�8) were harvested, and IL-17RB messenger RNA expression was measured byRT-PCR. GAPDH was used to ensure equal loading of the RNA samples. Bottom, B: the graphindicates the difference in IL-17RB messenger RNA expression between the �5661G and �5661Asubjects. The data are expressed as the ratio of IL-17RB messenger RNA to GAPDH messenger RNA.The data are presented as the mean �SEM.
1178 Original Research
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from

(�5552Tins�del), is significantly associated with thedevelopment of asthma (p � 0.001). Whole-genomeanalyses have shown that chromosome region 3p21–24, which contains a gene cluster of CC chemokinereceptors including CCR3, CCR2, and CCR5, islinked to asthma.25 One genetic marker on chromo-some 3p (D3S2409) was closely linked to the asth-matic phenotype in a founder population.16 Theseresults suggest that polymorphisms in IL-17RB havea genetic effect on the susceptibility of an individualto asthma.
Genomic variation, in the form of single-nucleotidesubstitutions or small insertions and deletions, inintronic regions was not previously thought tochange the sequence of a protein; however, someevidence indicates that such variation can causesplicing abnormalities, which may result in humandisease.26 To determine the genetic effect of IL-17RB �5661G�A or IL-17RB �5552T ins�del inintron 4 on splicing, we analyzed B-cell lines byRT-PCR using primers specific for exons 2 to 7. Theproducts were then sequenced and confirmed asIL-17RB messenger RNA. Our results revealed noalternative form of IL-17RB in B-cell lines derivedfrom those asthmatic subjects carrying either geno-type of IL-17RB �5661G�A (Fig 2, top, A). Con-sidering that splicing abnormalities are often tissuespecific, it is important to choose appropriate targetcells. IL-17RB is not expressed in peripheral bloodleukocytes, lymphoid organs, and most cell lines.IL-17RB, which is the receptor for IL-17E, is up-regulated in inflammatory tissues but is not presentat a high copy number on eosinophils or B cells.Because IL-17RB expression is limited to anti-CD3and anti-CD28 antibody-costimulated memory Thcells, antigen-presenting cells, and cultured primaryfibroblasts,9,12,27,28 no RT-PCR products were pro-duced from the peripheral blood mononuclear cellsof the asthmatic patients in the present study (datanot shown). However, IL-17RB is up-regulated innumerous B, T, and myeloid leukemic cell lines,including M1 (myeloid leukemia), EL-4 (thymoma),and WEHI231 (pre-B-cell lymphoma).29 Thisprompted us to use B-cell lines derived from theasthmatic patients for the semi-quantitative mea-surement of IL-17RB expression. The messengerRNA expression of IL-17RB in the B-cell lines wassignificantly higher in those cells that were homozy-gous for IL-17RB �5661GG compared to those thatwere homozygous for IL-17RB �5661AA (Fig 2).These results indicate that the expression of theIL-17RB gene may be regulated at the transcrip-tional level based on the �5661G�A genotype.
IL-17RB binds strongly to IL-17E and weakly toIL-17B, but does not bind IL-17 or IL-17C.30 Theactivation of IL-17RB induces Jun kinase, p38 mitogen-
activated protein kinase, and nuclear factor-�B activa-tion,10 and the production of monocyte chemotacticprotein-1, monocyte inflammatory protein-1�, IL-6,and IL-8 by eosinophils.11 It was recently demonstrat-ed12 that human primary lung fibroblasts constitutivelyexpress IL-17RB and that IL-17E stimulates the pro-duction of the eosinophil-associated mediators CCL-5,CCL-11, granulocyte macrophage colony-stimulatingfactor, and CXCL-8. This indicates that IL-17E maycontribute to the induction and maintenance of allergicinflammation in the airways via IL-17RB. In addition,the IL-17E produced by innate effector eosinophils andbasophils may augment allergic inflammation by en-hancing the maintenance and functions of adaptive Th2memory cells.13 The blockade of IL-25 in an experi-mental model of allergic asthma prevented AHR,which is a critical feature of clinical asthma. Theadministration of anti-IL-25 monoclonal antibodiesduring the sensitization phase resulted in significantlyreduced levels of IL-5 and IL-13 production, eosino-phil infiltration, goblet cell hyperplasia, and serum IgE,and prevented AHR.8 Thus, genetic variants of IL-17RB may play a role in Th2-mediated immune reac-tions in asthmatic patients. It is currently unclear howan intronic polymorphism can induce a phenotypicchange. IL-17RB �5661G�A and IL-17RB �5552Tins�del may induce exon skipping, enhance the use ofcryptic splice sites, or alter the ratio of alternativelyspliced isoforms. Further study is needed to elucidatethe functions of these intronic variants.
In summary, we examined the genetic associationof IL-17RB with the risk of asthma. Our resultsrevealed that the IL-17RB �5661G�A locus has adominant and protective genetic effect on the devel-opment of asthma. The results of this study may behelpful in understanding the function of polymor-phisms in IL-17RB in asthma development and maylead to the production of new asthma drugs.
References1 Sengler C, Lau S, Wahn U, et al. Interactions between genes
and environmental factors in asthma and atopy: new devel-opments. Respir Res 2002; 3:7
2 Robinson DS, Hamid Q, Ying S, et al. Predominant TH2-likebronchoalveolar T-lymphocyte population in atopic asthma.N Engl J Med 1992; 326:298–304
3 Broide DH, Lotz M, Cuomo AJ, et al. Cytokines in symptomaticasthma airways. J Allergy Clin Immunol 1992; 89:958–967
4 Cohn L, Elias JA, Chupp GL. Asthma: mechanisms of diseasepersistence and progression. Annu Rev Immunol 2004; 22:789–815
5 Fort MM, Cheung J, Yen D, et al. IL-25 induces IL-4, IL-5,and IL-13 and Th2-associated pathologies in vivo. Immunity2001; 15:985–995
6 Hurst SD, Muchamuel T, Gorman DM, et al. New IL-17family members promote Th1 or Th2 responses in the lung:in vivo function of the novel cytokine IL-25. J Immunol 2002;169:443–453
www.chestjournal.org CHEST / 135 / 5 / MAY, 2009 1179
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from

7 Kim MR, Manoukian R, Yeh R, et al. Transgenic overexpres-sion of human IL-17E results in eosinophilia, B-lymphocytehyperplasia, and altered antibody production. Blood 2002;100:2330–2340
8 Ballantyne SJ, Barlow JL, Jolin HE, et al. Blocking IL-25prevents airway hyperresponsiveness in allergic asthma. J AllergyClin Immunol 2007; 120:1324–1331
9 Gratchev A, Kzhyshkowska J, Duperrier K, et al. The recep-tor for interleukin-17E is induced by Th2 cytokines inantigen-presenting cells. Scand J Immunol 2004; 60:233–237
10 Lee J, Ho WH, Maruoka M, et al. IL-17E, a novel proinflam-matory ligand for the IL-17 receptor homolog IL-17Rh1.J Biol Chem 2001; 276:1660–1664
11 Wong CK, Cheung PF, Ip WK, et al. Interleukin-25-inducedchemokines and interleukin-6 release from eosinophils ismediated by p38 mitogen-activated protein kinase, c-JunN-terminal kinase, and nuclear factor-�B. Am J Respir CellMol Biol 2005; 33:186–194
12 Letuve S, Lajoie-Kadoch S, Audusseau S, et al. IL-17Eupregulates the expression of proinflammatory cytokines inlung fibroblasts. J Allergy Clin Immunol 2006; 117:590–596
13 Wang YH, Angkasekwinai P, Lu N, et al. IL-25 augments type2 immune responses by enhancing the expansion and func-tions of TSLP-DC-activated Th2 memory cells. J Exp Med2007; 204:1837–1847
14 Angkasekwinai P, Park H, Wang YH, et al. Interleukin 25promotes the initiation of proallergic type 2 responses. J ExpMed 2007; 204:1509–1517
15 Shi Y, Ullrich SJ, Zhang J, et al. A novel cytokine receptor-ligand pair: identification, molecular characterization, and invivo immunomodulatory activity. J Biol Chem 2000; 275:19167–19176
16 Ober C, Tsalenko A, Parry R, et al. A second-generationgenomewide screen for asthma-susceptibility alleles in afounder population. Am J Hum Genet 2000; 67:1154–1162
17 Webb BT, van den Oord E, Akkari A, et al. Quantitativelinkage genome scan for atopy in a large collection ofCaucasian families. Hum Genet 2007; 121:83–92
18 Global Initiative for Asthma. Global strategy for asthmamanagement and prevention: NHLBI/WHO workshop re-port. Bethesda, MD. National Institutes of Health NationalHeart, Lung, and blood Institute, 1995
19 Ferris BG. Epidemiology Standardization Project (AmericanThoracic Society). Am Rev Respir Dis 1978; 118:1–120
20 Livak KJ. Allelic discrimination using fluorogenic probes andthe 5 nuclease assay. Genet Anal 1999; 14:143–149
21 Chomczynski P, Sacchi N. Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction. Anal Biochem 1987; 162:156–159
22 Hedrick PW. Gametic disequilibrium measures: proceed withcaution. Genetics 1987; 117:331–341
23 Kaplan N, Weir BS. Expected behavior of conditional linkagedisequilibrium. Am J Hum Genet 1992; 51:333–343
24 Stephens M, Smith NJ, Donnelly P. A new statistical methodfor haplotype reconstruction from population data. Am JHum Genet 2001; 68:978–989
25 Maho A, Bensimon A, Vassart G, et al. Mapping of theCCXCR1, CX3CR1, CCBP2 and CCR9 genes to the CCRcluster within the 3p213 region of the human genome.Cytogenet Cell Genet 1999; 87:265–268
26 Pagani F, Baralle FE. Genomic variants in exons and introns:identifying the splicing spoilers. Nat Rev Genet 2004; 5:389–396
27 Ikeda K, Nakajima H, Suzuki K, et al. Mast cells produceinterleukin-25 upon FcERI-mediated activation. Blood 2003;101:3594–3596
28 Wong CK, Li PW, Lam CW. Intracellular JNK, p38 MAPKand NF-�B regulate IL-25 induced release of cytokines andchemokines from costimulated T helper lymphocytes. Immu-nol Lett 2007; 112:82–91
29 Tian E, Sawyer JR, Largaespada DA, et al. Evi27 encodes anovel membrane protein with homology to the IL17 receptor.Oncogene 2000; 19:2098–2109
30 Moseley TA, Haudenschild DR, Rose L, et al. Interleukin-17family and IL-17 receptors. Cytokine Growth Factor Rev2003; 14:155–174
1180 Original Research
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from

DOI 10.1378/chest.08-1595; Prepublished online December 31, 2008; 2009;135; 1173-1180Chest
Choon-Sik Park and Hyoung Doo ShinYoung-Mok Lee, Ki-Up Kim, Soo-Taek Uh, Ju Ock Na, Yong-Hoon Kim,Kim, Hun Soo Chang, TaiYoun Rhim, Jong-Sook Park, An-Soo Jang,
Ji-Sun Jung, Byung Lae Park, Hyun Sub Cheong, Joon Seol Bae, Ji-Hye Gene Polymorphism With AsthmaIL-17RBAssociation of
May 18, 2009This information is current as of
& ServicesUpdated Information
http://www.chestjournal.org/content/135/5/1173.full.htmlhigh-resolution figures, can be found at:Updated Information and services, including
References
html#ref-list-1http://www.chestjournal.org/content/135/5/1173.full.accessed free at:This article cites 29 articles, 10 of which can be
Open AccessoptionFreely available online through CHEST open access
Permissions & Licensing
http://www.chestjournal.org/site/misc/reprints.xhtml(figures, tables) or in its entirety can be found online at: Information about reproducing this article in parts
Reprints http://www.chestjournal.org/site/misc/reprints.xhtml
Information about ordering reprints can be found online:
Email alerting service
online article.article. sign up in the box at the top right corner of the Receive free email alerts when new articles cit this
formatImages in PowerPoint
format. See any online article figure for directions.downloaded for teaching purposes in PowerPoint slide Figures that appear in CHEST articles can be
Copyright © 2009 American College of Chest Physicians on May 18, 2009www.chestjournal.orgDownloaded from