asexual genetic variability in agavaceae determined with inverse sequence-tagged repeats and...
TRANSCRIPT

Genetic Resources
Asexual Genetic Variability in AgavaceaeDetermined with Inverse Sequence-Tagged Repeatsand Amplification Fragment Length PolymorphismAnalysis
DIÓGENES INFANTE*, SANDY MOLINA, JHONNY R. DEMEY andELIZABETH GÁMEZUnidad de Biotecnología de Plantas, Instituto de Estudios Avanzados, Apdo. 17606Parque Central, Caracas 1015-A, Venezuela
Abstract. Agaves are succulent monocot plants rich in fibers, sugars and other importantcompounds. They are also valued as ornamental plants and for their ability to grow in poorsoils. In the present study, inverse sequence–tagged repeats (ISTR) and amplified fragmentlength polymorphism (AFLP) analysis were used to study genetic diversity in differentAgavaceae plant samples. Comparison of the banding patterns between the mother plantand rhizome-derived daughter plants showed that genetic variability is generated duringasexual reproduction in these species. Phylogenetic relationships among Agave specieswere obtained using unweighted pair-group method, arithmetic average (UPGMA) analysis.Genetic diversity through asexual propagation allows for genetic selection and improvementwithin these asexually propagated plants.
Key words: Agave, genetic diversity, molecular markers, clonal diversity, reproduction
Abbreviations: AFLP, amplified fragment length polymorphism; ISTR, inverse se-quence–tagged repeat; PCR, polymerase chain reaction; RAPD, random-amplified poly-morphic DNA; S-SAP, sequence-specific amplified polymorphism; UPGMA, unweightedpair-group method, arithmetic average.
Agavaceae clonal diversity Infante et al.Introduction
Agaves are succulent monocot plants that grow natively in semiarid, subtropical,and tropical regions from the southern United States to northern South Americaand throughout the Caribbean. They are rich in fiber, sugars and other importantcompounds, in addition to having value as ornamental plants, and they are capableof growing in poor soils.
Some agaves have three different reproductive mechanisms. The first is theproduction of rhizomes, which appear in early stages of plant development andconsist of subterraneous shoots that grow from meristems at the base of the stemand emerge, forming new individuals. During later stages of development inAgave fourcroydes (Piven et al., 2001), Agave littaea (Rocha et al., 2005) and
Plant Molecular Biology Reporter 24: 205–217, June 2006© 2006 International Society for Plant Molecular Biology. Printed in Canada.
*Author for correspondence. e-mail: [email protected]; tel: 58-0212-903-5104;fax: 58-0212-903-5092.

Agave microcantha (Arizaga and Ezcurra, 2002), the second and third reproductivemechanisms come into play. A long flower stalk (10-12 m high in henequen[A. fourcroydes]) appears, with flowers and bulbils developing from cells at theflower stem. The flower takes all of the sugars from the rest of the plant and forthis reason the plant dies shortly after the blossoming.
Several markers have been used in our work to study genetic diversity inagaves, amplified fragment length polymorphism (AFLP), markers based on theselective amplification by PCR of a subset of restriction fragments generated using2 different restriction enzymes (Vos et al., 1995); inverse sequence–tagged repeat(ISTR), markers based on selective PCR amplification of genomic DNA by usingprimers derived from copia-like repetitive elements. ISTRs are generated at random,produce a high number of bands by PCR without further manipulation of DNAafter isolation, and are highly reproducible (Rohde, 1996). The comparison ofboth markers in A. fourcroydes demonstrated that use of ISTR has greater poly-morphic information content, better capacity for quantification of the geneticdiversity, and higher discriminatory capacity than does AFLP, with the possibilityof detecting simultaneously several polymorphic markers with a single PCR reaction(Demey et al., 2004). AFLP analysis was used to evaluate genetic diversity incultivated henequen, which demonstrates differences in AFLP patterns in a naturalpopulation despite being propagated asexually. Comparison of the AFLP bandingpattern between 2 mother plants with their rhizome-derived daughter plants showedthat in this species genetic variability is introduced through asexual reproduction(Infante et al., 2003). This result was accompanied by differences in morphologicalparameters, conditions allowing for selection in an improvement program. For thispurpose, González et al. (2003) using somatic embryogenesis–derived plants,demonstrated the heritability of the AFLP pattern and of the morphological char-acteristics, which allowed for genetic improvement of henequen using asexualreproduction. To further study the origin of this asexual diversity, AFLP and ISTRwere used for genetic analysis of intraorganismal genetic heterogeneity. Differ-ences in banding pattern were found in different leaves, bulbils, and suckers thathad originated from the same mother plant (Demey et al., 2004) (unpublisheddata).
In this work, we studied within-clone variability in different Agave speciesusing AFLP and ISTR. We compared the molecular marker banding patternsbetween mother plants and sucker-derived daughter plants. Phylogenetic relation-ships among the Agave species studied were obtained using unweightedpair-group method, arithmetic average (UPGMA).
Materials and Methods
Plant material
Leaf samples were taken from mother and daughter plants from the followingAgave species: A. deserti, A. angustifolia, A. angustifolia var. marginata, A. americanavar. americana, A. fourcroydes var. Kitam ki, A. fourcroydes var. Sac ki, A.fourcroydes var. Yaax ki, A. sisalana, A. fourcroydes var. Mac ki, A. angustifoliavar. letonae, A. lechuguilla, and A. tequilana. Samples were obtained from theBotanical Garden, Caracas, Venezuela.
206 Infante et al.

DNA isolation
DNA was isolated from fresh young leaves following a previously publishedprocedure for DNA mini-extraction from agave leaves (Keb-Llanes et al., 2002).
AFLP and ISTR development
AFLPs were developed using a scaled-down modification of the procedure originallydescribed (Vos et al., 1995), as previously published (Infante et al., 2003). Weused the following selective primers: E-ACG in combination with M-CAC,M-CTC and M-CTT. These combinations (E-ACG × M-CAC, E-ACG × M-CTC,E-ACG × M-CTT) were selected because they were highly polymorphic.
ISTRs were obtained using the following primer combinations: F1A × B2B,F2 × B1A, F2 × B2B, and F3 × B2B (F indicates the forward primer, while Bindicates the backward primer). Selective AFLP primers and ISTR primers wereradiolabeled using [33P]γ -ATP. The amplification products were mixed with 8 μLof loading buffer, and 4 μL was then electrophoresed at constant power (65 W) ina 0.4-mm-thick, 6% denaturing polyacrylamide gel in 1× tris-borate-EDTA runningbuffer, until the blue band reached three-quarters of the gel length, as describedby Sambrook and Russell (2001). After electrophoresis, the gels were lifted ontochromatographic paper, covered with plastic wrap, and dried under a heated vacuum,and the bands were visualized by exposure to Kodak X-Omat film for varyingtimes from overnight to several days.
Data analysis
Autoradiograms were scanned, and bands were analyzed using the program CrossChecker (Buntjer, 2000). This program created a 0–1 matrix scoring for the presenceor absence of each band. After visual correction of the results, the data wereexported as a 0–1 matrix to the programs used for statistical analysis.
For characterization of the capacity of each primer to detect polymorphicloci, we use polymorphic information content and its estimation of bootstrap’s SE(n = 500) (Anderson et al., 1993) and the probability of identical match by chance(Wetton et al., 1987) with modifications (Demey et al., 2003). The genetic rela-tionship among Agave species was investigated using cluster analysis with the Neiand Li coefficient of similarity, depicted in the UPGMA dendrogram. Boot-strapping (n = 500) was used to estimate SEs and confidence intervals of themean distance in each node (Felsenstein, 1985; Nei and Kumar, 2000). All analyseswere computed with Info-Gen, version 1 (Balzarini et al., 2003); FreeTree, version0.9.1.50 (Pavlicek et al., 1999); and Treeview, version 1.6.0 (Page, 2000).
Results
ISTRs were obtained for all of the Agave species studied using 4 primer combina-tions, which yielded a total of 142 scorable polymorphic bands. Figure 1A showsthe banding pattern for the ISTR primer combination F3 × B2B. Genetic variationvalues were similar with all combinations. However, a higher discriminatory capacitywas obtained using the combination F2 × B1A, as shown in Table 1. Data from
Agavaceae clonal diversity 207
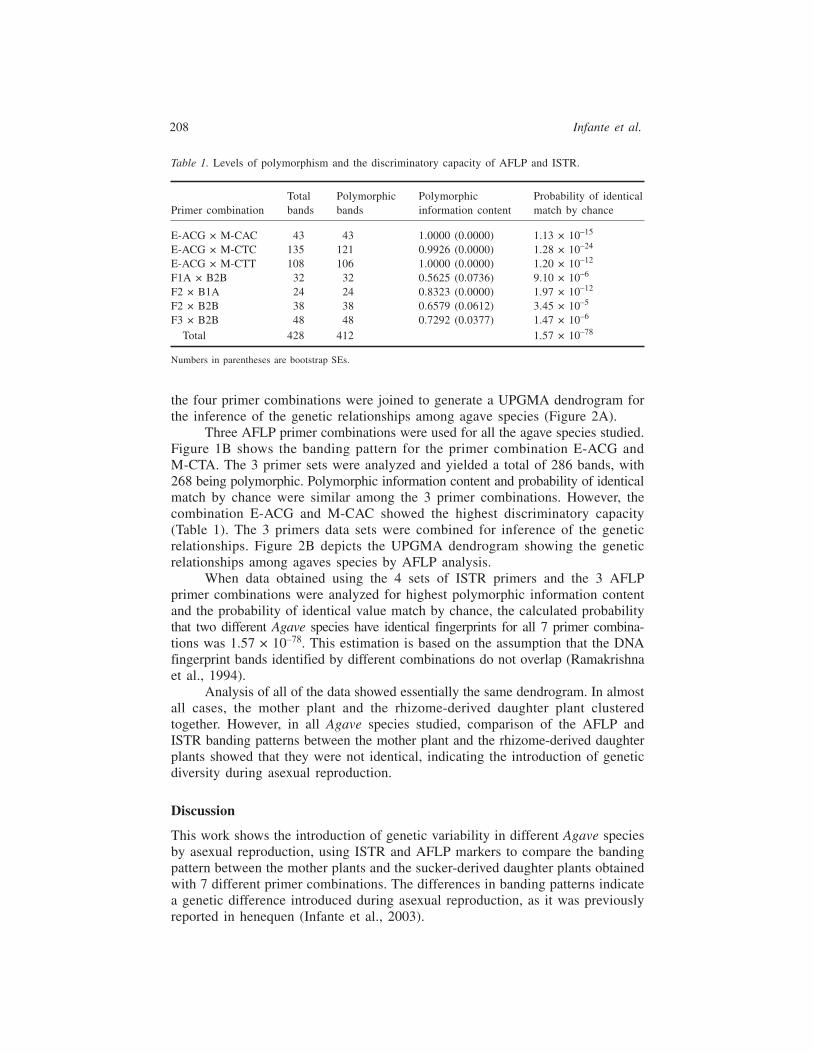
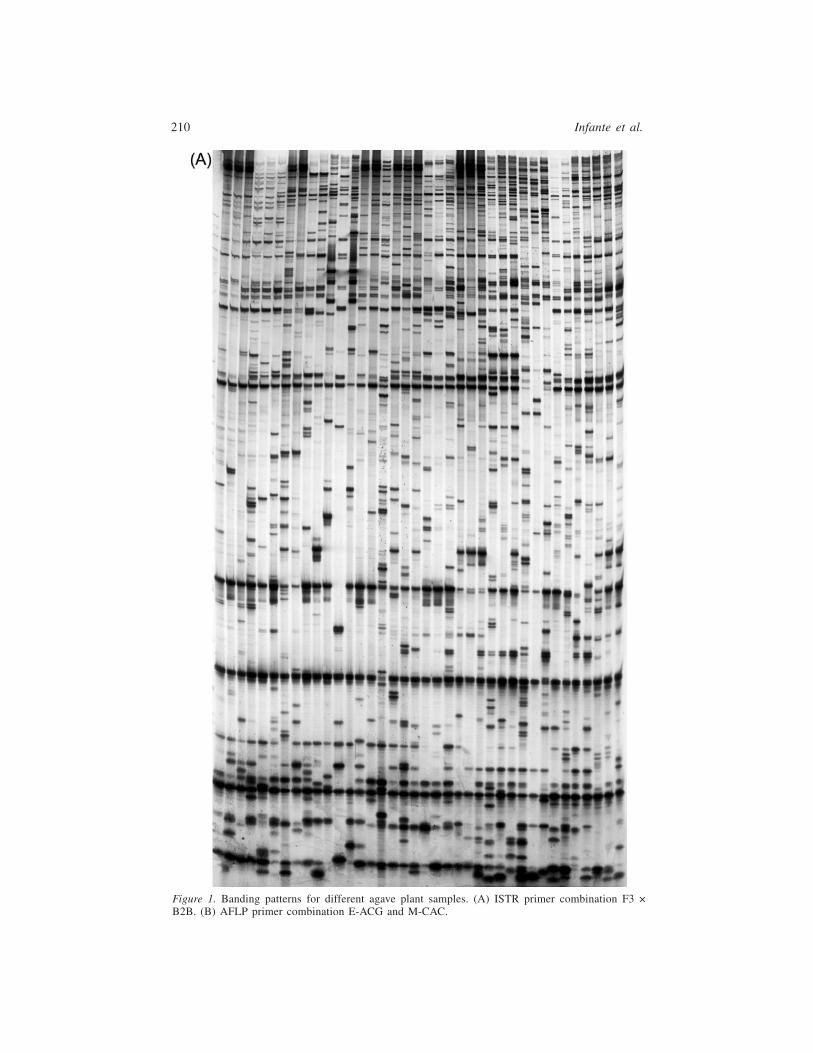
the four primer combinations were joined to generate a UPGMA dendrogram forthe inference of the genetic relationships among agave species (Figure 2A).
Three AFLP primer combinations were used for all the agave species studied.Figure 1B shows the banding pattern for the primer combination E-ACG andM-CTA. The 3 primer sets were analyzed and yielded a total of 286 bands, with268 being polymorphic. Polymorphic information content and probability of identicalmatch by chance were similar among the 3 primer combinations. However, thecombination E-ACG and M-CAC showed the highest discriminatory capacity(Table 1). The 3 primers data sets were combined for inference of the geneticrelationships. Figure 2B depicts the UPGMA dendrogram showing the geneticrelationships among agaves species by AFLP analysis.
When data obtained using the 4 sets of ISTR primers and the 3 AFLPprimer combinations were analyzed for highest polymorphic information contentand the probability of identical value match by chance, the calculated probabilitythat two different Agave species have identical fingerprints for all 7 primer combina-tions was 1.57 × 10–78. This estimation is based on the assumption that the DNAfingerprint bands identified by different combinations do not overlap (Ramakrishnaet al., 1994).
Analysis of all of the data showed essentially the same dendrogram. In almostall cases, the mother plant and the rhizome-derived daughter plant clusteredtogether. However, in all Agave species studied, comparison of the AFLP andISTR banding patterns between the mother plant and the rhizome-derived daughterplants showed that they were not identical, indicating the introduction of geneticdiversity during asexual reproduction.
Discussion
This work shows the introduction of genetic variability in different Agave speciesby asexual reproduction, using ISTR and AFLP markers to compare the bandingpattern between the mother plants and the sucker-derived daughter plants obtainedwith 7 different primer combinations. The differences in banding patterns indicatea genetic difference introduced during asexual reproduction, as it was previouslyreported in henequen (Infante et al., 2003).
208 Infante et al.
Primer combinationTotalbands
Polymorphicbands
Polymorphicinformation content
Probability of identicalmatch by chance
E-ACG × M-CAC 43 43 1.0000 (0.0000) 1.13 × 10–15
E-ACG × M-CTC 135 121 0.9926 (0.0000) 1.28 × 10–24
E-ACG × M-CTT 108 106 1.0000 (0.0000) 1.20 × 10–12
F1A × B2B 32 32 0.5625 (0.0736) 9.10 × 10–6
F2 × B1A 24 24 0.8323 (0.0000) 1.97 × 10–12
F2 × B2B 38 38 0.6579 (0.0612) 3.45 × 10–5
F3 × B2B 48 48 0.7292 (0.0377) 1.47 × 10–6
Total 428 412 1.57 × 10–78
Numbers in parentheses are bootstrap SEs.
Table 1. Levels of polymorphism and the discriminatory capacity of AFLP and ISTR.

Clonal genetic diversity of several plants species generated during asexualreproduction in the laboratory has been reported previously using AFLP analysis.In somatic embryogenesis–derived Arabidopsis thaliana, 66.6% of the studiedplants presented at least 1 variation when compared against the regenerated plantsobtained from the same explant, and the number of mutations per plant rangedfrom 1 to 18 (Polanco and Ruiz, 2002). In henequen, somaticembryogenesis–derived plants clustered with their mother plants in an AFLP anal-ysis and in a principal component analysis using morphological characters, but inthis case, the diversity was lower than in sucker-derived plants (González et al., 2003).Comparison of the AFLP banding pattern between mother Coffea arabica plantsand somatic embryogenesis–derived daughter plants showed that somaticembryogenesis per se introduced genetic diversity (Sanchez-Teyer et al., 2003).
After calculation of the mean probability, with the 4 ISTR and 3 AFLPprimer combinations, up to 1078 Agave individuals can be compared simulta-neously; thus, ISTR and AFLP are highly sensitive for detecting polymorphismsbetween mother and daughter plants within different Agave species. This indicatesthat the results obtained have a high degree of confidence, are not obtained bychance nor an artifact of the technique, and validate the use of the ISTR markersystem in Agave species.
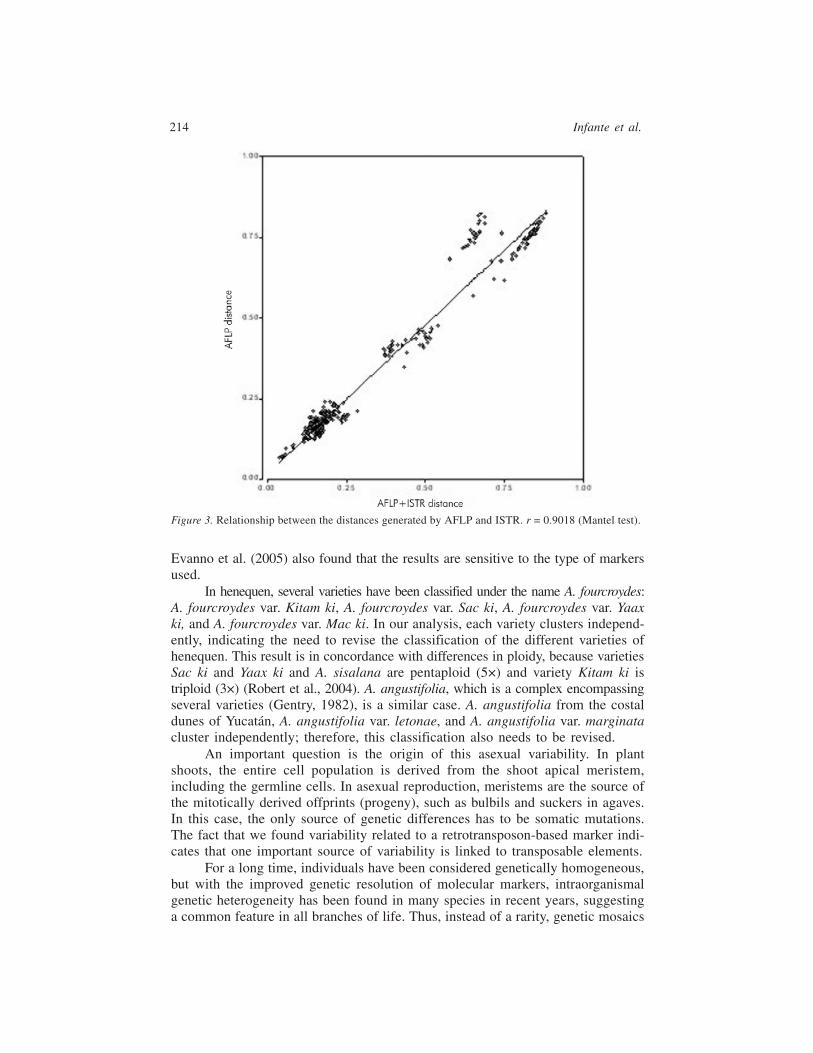
The Mantel test was used with the AFLP and ISTR data, as shown in Figure 3,obtaining a correlation of r = 0.9018; the data from each marker distributed overboth side of the middle of the graph, without overlap, indicating that each markerexplores a different region within the genome. Therefore, combining data setsdoes not significantly introduce a bias in the calculations. In another work, testing3 different markers systems with tomato and pepper, Tam et al. (2005) also foundthat the retrotransposon-based S-SAP marker did not overlap with AFLP markers.
A possible explanation for this finding is that AFLP that are generated withthe restriction enzymes MseI and EcoRI are frequently clustered in the centromericregions (Waugh et al., 1997), while transposons prefers heterochromatin regionsin the genome (Kidwell and Lisch, 1997). Lönnig and Saedler (2002), in theirreview of rearrangement induced by transposons, reference several works thatshow the existence of “hot spots” for the insertion of transposons. This result is inagreement with the fact that transposon-based ISTR and AFLP analyses coverdifferent regions in the genome of agaves.
After bootstrapping (n = 500), the clustering method using data from bothmarkers shows the mother and daughters plants clustering in the same group in allcases.
Many conclusions can be drawn from this tree. First, asexual genetic variabilityhas been found in A. tequilana, despite the fact that in a previous work, Gil-Vegaset al. (2001) found no variability using random-amplified polymorphic DNA(RAPD) analysis. A. tequilana is a diploid plant species that reproduces only bysuckers in plantations. This difference in findings may be explained by the factthat RAPD, AFLP, and ISTR explore different regions of the genome in A.tequilana, as we demonstrate with AFLP and ISTR for different agaves in thiswork. In this species, the region explored by RAPD is conserved and the regionexplored by the others markers is variable. Further, in a work aimed at identificationof genetically homogeneous groups of individuals using a Bayesian approach,
Agavaceae clonal diversity 209
210 Infante et al.
Figure 1. Banding patterns for different agave plant samples. (A) ISTR primer combination F3 ×B2B. (B) AFLP primer combination E-ACG and M-CAC.

Agavaceae clonal diversity 211
Figure 1. (concluded).

212 Infante et al.
Figure2.GeneticrelationshipamongAga
vespecies,depictedbyUPGMAtreeanalysis,withtwodifferentmarkersystems:(A)ISTR,(B)AFLP.

Agavaceae clonal diversity 213
Figure2.
(con
clud
ed).
Evanno et al. (2005) also found that the results are sensitive to the type of markersused.
In henequen, several varieties have been classified under the name A. fourcroydes:A. fourcroydes var. Kitam ki, A. fourcroydes var. Sac ki, A. fourcroydes var. Yaaxki, and A. fourcroydes var. Mac ki. In our analysis, each variety clusters independ-ently, indicating the need to revise the classification of the different varieties ofhenequen. This result is in concordance with differences in ploidy, because varietiesSac ki and Yaax ki and A. sisalana are pentaploid (5×) and variety Kitam ki istriploid (3×) (Robert et al., 2004). A. angustifolia, which is a complex encompassingseveral varieties (Gentry, 1982), is a similar case. A. angustifolia from the costaldunes of Yucatán, A. angustifolia var. letonae, and A. angustifolia var. marginatacluster independently; therefore, this classification also needs to be revised.
An important question is the origin of this asexual variability. In plantshoots, the entire cell population is derived from the shoot apical meristem,including the germline cells. In asexual reproduction, meristems are the source ofthe mitotically derived offprints (progeny), such as bulbils and suckers in agaves.In this case, the only source of genetic differences has to be somatic mutations.The fact that we found variability related to a retrotransposon-based marker indi-cates that one important source of variability is linked to transposable elements.
For a long time, individuals have been considered genetically homogeneous,but with the improved genetic resolution of molecular markers, intraorganismalgenetic heterogeneity has been found in many species in recent years, suggestinga common feature in all branches of life. Thus, instead of a rarity, genetic mosaics
214 Infante et al.
Figure 3. Relationship between the distances generated by AFLP and ISTR. r = 0.9018 (Mantel test).

are probably the rule, while genetic homogeneity is the exception (Pineda-Krchand Lehtilä, 2004). Here we found asexual genetic variability among differentAgave species, which opens the possibility for genetic improvement of theseplants. The high adaptability and degree of invasiveness displayed by agavescould be an advantage of their asexual genetic variability.
Acknowledgments
We thank Wolfgang Rohde and Dieter Becker (Max Planck Institute for PlantBreeding Research, Köln, Germany) for their support in marker development, andAna M. Kelly, Manoj Kumar, and Carolina Celis for critically reviewing themanuscript.
Note Added in Proof
In a recent work, Gil-Vegas et al. (2006) also demonstrated the existence of asexualgenetic variability in A. tequilana using AFLP, in contradiction with their previousfinding with RAPD, also supporting the fact that different marker systems, e.g.,AFLP and RAPD, examine distinct regions in the genome.
References
Anderson JA, Churchill GA, Autrique JE, Tanksley SD, and Sorrells ME (1993) Opti-mizing parental selection for genetic linkage maps. Genome 36: 181-186.
Arizaga S and Ezcurra E (2002) Propagation mechanisms in Agave macroacantha (Agavaceae),a tropical arid-land succulent rosette. Am J Bot 89: 632-641.
Balzarini M, Di Rienzo J, and Casanoves F (2003) Info-Gen: Un ambiente integrado parael analisis de datos genéticos. In: II Mtg Caribbean and Central American Region, IntBiometrics Soc, 2003, Puerto Rico.
Buntjer J (2000) CrossChecker fingerprint analysis software, version 2.9. WageningenUniversity and Research Center, Wageningen, the Netherlands.
Demey JR, Gamez E, Molina S, and Infante D (2004) Comparative study of the discrimi-nating capacity of AFLP and ISTR markers for genetic analysis of Agave fourcroydes.Plant Mol Biol Rep 22: 29-35.
Demey JR, Zambrano AY, Fuenmayor F, and Segovia V (2003) Relación entre las característicasmoleculares y morfológicas en una colección de Yuca. Interciencia 28: 684-689.
Evanno G, Regnaut S, and Goudet J (2005) Detecting the number of clusters of individualsusing the software structure: a simulation study. Mol Ecol 14: 2611-2620.
Felsenstein J (1985) Confidents limits on phylogenies: an approach using the bootstrap.Evolution 39: 783-791.
Gentry HS (1982) Agaves of continental North America, University of Arizona Press,Tucson AZ.
Gil-Vegas K, González-Chavira M, Martinez de la Vega O, Simpson J, and Vandermark G(2001) Analysis of genetic diversity in Agave tequilana var. Azul using RAPD markers.Euphytica 119: 335-341.
Gil-Vegas K, Diaz C, Nava-Cedillo A, and Simpson J (2006) AFLP analysis of Agavetequilana varieties. Plant Sci 170: 904-909.
González G, Alemán S, and Infante D (2003) Asexual genetic variability in Agave fourcroydes II:Selection among individuals in a clonally propagated population. Plant Sci 165: 595-601.
Agavaceae clonal diversity 215

Infante D, González G, Peraza-Echeverría L, and Keb-Llanes M (2003) Asexual geneticvariability in Agave fourcroydes. Plant Sci 164: 223-230.
Keb-Llanes M, Gerardo González, Chi-Manzanero B, and Infante D (2002) A rapid andsimple method for small scale DNA extraction in Agavaceae and other tropical plants.Plant Mol Biol Rep 20: 299a-299e.
Kidwell MG and Lisch D (1997) Transposable elements as sources of variation in animalsand plants. Proc Natl Acad Sci USA 94: 7704-7711.
Lönnig WE and Saedler H (2002) Chromosome rearrangements and transposable elements.Annu Rev Genet 36: 389-410.
Nei M and Kumar S (2000) Molecular evolution and phylogenetics, Oxford UniversityPress, New York.
Page R (2000) TREEVIEW 1.6.0. An application to display phylogenetic trees. Comp AppBiosci 12: 357-358.
Pavlicek A, Hrda S, and Flegr J (1999) FreeTree — Freeware program for construction ofphylogenetic trees on the basis of distance data and bootstrap/jackknife analysis of thetree robustness. Application in the RAPD analysis of the genus Frenkelia. Folia Biol(Praha) 45: 97-99.
Pineda-Krch M and Lehtilä K (2004) Challenging the genetically homogeneous individual.J Evol Biol 17: 1192-1194.
Piven NM, Barredo-Pool FA, Borges-Argaez IC, Herrera-Alamillo MA, Mayo-MosquedaA, Herrera-Herrera JL, and Robert ML (2001) Reproductive biology of henequen(Agave fourcroydes) and its wild ancestor Agave angustifolia (Agavaceae). i. Gametophytedevelopment. Am J Bot 88: 1966-1976.
Polanco C and Ruiz ML (2002) AFLP analysis of somaclonal variation in Arabidopsisthaliana regenerated plants. Plant Sci 162: 817-824.
Ramakrishna W, Lagu WD, Gupta VS, and Ranjekar PK (1994) DNA fingerprinting inrice using oligonucleotide probes specific for simple repetitive DNA sequences. TheorAppl Genet 88: 402-406.
Robert ML, Hanson L, Bennet MD, and Leitch I (2004) El tamaño del genoma de los agaveses proporcional a su nivel de ploidía. In: IV Int Simp Agavaceae and Nonlinaceae,2004 Nov 20-25, Centro de Investigación Científica de Yucatán, Mérida, México.
Rocha M, Valera A, and Eguiarte LE (2005) Reproductive ecology of five sympatricAgave littaea (Agavaceae) species in central Mexico. Am J Bot 92: 1330-1341.
Rohde W (1996) Inverse sequence–tagged repeat (ISTR) analysis, a novel and universalPCR-based technique for genome analysis in the plant and animal kingdom. J GenetBreed 50: 249-261.
Sambrook J and Russell D (2001) Molecular cloning, a laboratory manual, 3rd ed, ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
Sanchez-Teyer LF, Quiroz-Figueroa F, Loyola-Vargas V, and Infante D (2003) Cul-ture-induced variation in plants of Coffea arabica cv. caturra rojo, regenerated by di-rect and indirect somatic embryogenesis. Mol Biotechnol 23: 107-115.
Tam SM, Mhiri C, Vogelaar A, Kerkveld M, Pearce SR, and Grandbastien MA (2005)Comparative analyses of genetic diversities within tomato and pepper collectionsdetected by retrotransposon-based SSAP, AFLP, and SSR. Theor Appl Genet 110:819-831.
Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters A, Pot J,Peleman J, Kuiper M, and Zabeau M (1995) AFLP: a new technique for DNA finger-printing. Nucleic Acids Res 23: 4407-4414.
Waugh R, McLean K, Flavell AJ, Pearce SR, Kumar A, Thomas BBT, and Powell W(1997) Genetic distribution of Bare-1-like retrotransposable elements in the barley
216 Infante et al.

genome revealed by sequence-specific amplification polymorphisms (S-SAP). MolGenet Genomics 253: 687-694.
Wetton JH, Carter RE, Parkin DT, and Walters D (1987) Demographic study of a wildhouse sparrow population by DNA fingerprinting. Nature 327: 147-149.
Agavaceae clonal diversity 217