article in press
TRANSCRIPT

A
tee7ipcA1aeesvs©
K
7
Available online at www.sciencedirect.com
Theriogenology xx (2010) xxx
0d
ARTICLE IN PRESS
Heat shock protein 70 gene expression in equine blastocysts afterexposure of oocytes to high temperatures in vitro or in vivo after
exercise of donor mares
C.J. Mortensena,c,*, Y.-H. Choib, N.H. Inga, D.C. Kraemerb, M.M. Vogelsanga,Katrin Hinrichsb
a Department of Animal Science, College of Agriculture and Life Sciences, Texas A&M University, College Station, TX 77843-2471b Department of Veterinary Physiology and Pharmacology, College of Veterinary Medicine and Biomedical Sciences, Texas A&M University,
College Station, TX 77843-4466c Department of Animal Sciences, University of Florida, 459 Shealy Dr. P.O. Box 110910, Gainesville, FL 32611-0910
Received 2 October 2009; received in revised form 12 February 2010; accepted 22 February 2010
bstract
Heat above homeothermy can be detrimental to embryonic development, and cells may produce heat shock proteins to try to mitigatehese effects. The authors examined the developmental competence of equine oocytes after a single heat exposure (42 °C, 2 or 4 h) duringarly or late stages of in vitro maturation. Rates of nuclear maturation, cleavage after intracytoplasmic sperm injection, and advancedmbryonic development (morula or blastocyst) were compared to those for unexposed controls. Concentrations of heat shock protein0 (HSPA1A) mRNA were determined by real-time RT-PCR in resulting blastocysts, and were compared to those for embryos derivedn vivo from control or exercised mares. Exposure of oocytes to heat at the onset of in vitro maturation did not affect any measured endoint. However, exposure to 42 °C late in maturation culture reduced rates of oocyte nuclear maturation for both the 2 h (43/105 (43%)ompared to control 70/103 (68%); P � 0.01), and 4 h (47/106 (44%) compared to control 60/103 (59%); P � 0.05) groups.dditionally, late heat exposure reduced development to morulae and blastocyst stages after intracytoplasmic sperm injection (ICSI;8/89 (20%) compared to control 43/128 (34%); P � 0.05). Seven days after oocyte heat exposure, resultant blastocysts had a higherbundance of HSPA1A gene transcripts, relative to those for 18S rRNA. In vitro-produced embryos and lower-quality in vivo-producedmbryos had significantly higher relative HSPA1A mRNA (lower 18S rRNA) concentrations than did higher-quality in vivo-producedmbryos. The authors concluded that equine oocytes were sensitive to heat during late in vitro maturation, and responded to thermalhock with an increased ratio of HSPA1A:18S gene expression that was measurable in the resulting blastocyst. Embryos produced initro (including controls) had increased levels of HSPA1A mRNA relative to 18S rRNA compared to in vivo-produced embryos,uggesting a response to environmental insult.
2010 Elsevier Inc. All rights reserved.
eywords: Oocyte maturation; In vitro development; Embryo development; HSPA1A; Horse
www.theriojournal.com
1
c
* Corresponding Author. Tel.: �1 352 392 0133; fax: �1 352 392652.pE-mail address: [email protected] (C.J. Mortensen).
093-691X/$ – see front matter © 2010 Elsevier Inc. All rights reserved.oi:10.1016/j.theriogenology.2010.02.020
. Introduction
Oocyte maturation involves a series of nuclear andytoplasmic events that result in acquisition of the ca-
acity for fertilization, followed by embryo develop-
mhHRbdn(wbdt
adtaiaqItoeisi
fimnsi[ftslfedc
sslpoma
hssptHrt
ttawtmeovwaewkp
tooHcdist
2
2e
rapbcbuwHT
2 C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
ent. The maturing oocyte appears to be susceptible toeat-induced damage, both in vitro and in vivo [1–4].eat can have a direct effect on oocyte quality [3,5].oth and Hansen [3] reported that the majority ofovine oocytes exposed to temperatures of 40 or 41 °Curing the first 12 h of in vitro maturation (IVM) didot fully mature. Oocytes were arrested in metaphase IMI) and a higher proportion of heat-exposed oocytesere apoptotic. Ju et al. [6] reported that exposure ofovine oocytes to 42 °C for 4 h at the end of IVMecreased in vitro blastocyst formation after fertiliza-ion.
Potential effects of heat exposure on oocyte qualityre especially important in the horse, as assisted repro-uction techniques such as oocyte transfer and embryoransfer allow production of embryos from horses thatre in work [7]. Exercised horses have significant risesn core body temperature [8,9]. We previously reportedsignificant reduction in both embryo recovery rate anduality of recovered embryos in exercised mares [10].t is unclear whether these changes in embryo produc-ion and quality were due to direct effects of heat on theocyte or embryo, or rather to systemic effects of ex-rcise on the mare. To our knowledge, there is nonformation available on the direct effect of heat expo-ure on oocyte meiotic and developmental competencen the horse.
Oocytes, similar to somatic cells, have cellular de-ense mechanisms to protect and repair damage [11]. Its hypothesized that heat shock proteins, which act asolecular chaperones, are one of the primary mecha-
isms by which cells can survive injury. In response totress, cells can use stored (constitutive) or newly-nduced heat shock proteins. Ellis and Hemmingsen12] describe molecular chaperones as mediating theolding of polypeptides and potentially assembling cer-ain proteins into oligomeric structures. During heathock, non-specific protein interactions can occur thatead to the formation of non-functional proteins. Theamily of heat shock proteins acting as molecular chap-rones is thought to prevent the formation of interme-iate structures that might lead to a non-functionalonformation of proteins.
Heat shock protein 70 is highly conserved acrosspecies. It is one of the most widely studied cellulartress proteins. Under normal conditions, steady-stateevels of heat shock protein 70 functions as ATP-de-endent molecular chaperones by assisting the foldingf newly synthesized polypeptides, the assembly ofultiprotein complexes, and the transport of proteins
cross cellular membranes [13]. The inducible form of g
eat shock protein 70 (HSPA1A) is expressed in re-ponse to a variety of cellular stressors, including heathock. It acts by binding other proteins to preventrotein aggregation and maintain proteins in a compe-ent state for either refolding or degradation [14].SPA1A has been detected in mouse [15,16], rat [17],
abbit [18], porcine [19], and bovine [20,21] embryoshat have been exposed to thermal stress.
The relationship of heat treatment of mature oocyteso their subsequent development and gene transcrip-ional activity is poorly understood [6,22]. Fiorenza etl. [23] reported that when murine dictyate oocytesere exposed to temperatures of 43 °C for 30 min,
here was increased HSPA1A gene transcription. In theouse, HSPA1A gene expression was detectable as
arly as the single cell [23,24] and 2-cell stages [25]. Inther species, HSPA1A expression in older, more de-eloped embryos has been reported [20,22]. HSPA1Aas the most abundant inducible heat shock protein
fter thermal stress in Day 10 rat and Day 8.5 micembryos [26]. Therefore, the equine form of HSPA1Aas chosen as the target gene of this study. To ournowledge, no reports are available on embryonic ex-ression of heat shock proteins in the horse.
The aims of the present study were to: 1) evaluatehe effect of a single acute heat exposure on equineocytes in early or late stages of IVM culture on mei-tic and developmental competence, and 2) investigateSPA1A expression in in vitro-derived equine blasto-
ysts exposed or not exposed to increased temperaturesuring oocyte maturation, and in embryos producedn vivo from control or exercised mares. For theecond aim, 18S rRNA was measured in comparisono HSPA1A mRNA and input RNA levels.
. Materials and methods
.1. Effects of a single acute heat exposure onquine oocyte meiotic and developmental competence
Oocyte collection from abattoir-derived equine ova-ies (3–4 h transport time at 27–32 °C) was conducteds described previously [27]. Collection of ovaries waserformed outside the summer months to avoid possi-le heat stress to ovary donors. Briefly, oocytes wereollected by opening visible follicles with a scalpellade, and scraping the granulosa layer of each folliclesing a 0.5 cm bone curette. The contents of the curetteere washed into individual Petri dishes with 25 mMepes-buffered TCM199 with Hanks salts (Gibco Lifeechnologies, Grand Island, NY, USA) plus 25 �g/mL
entamycin (Gibco).
tedctcccoacamt
Twbm(ocpl(woniaoo(Ba
bdeaisjp2c
oip
NepwUrtbmtsoopdwmmcum
IsmmcfttpndaLiFC°
Dm[maecobd
3C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
Individual petri dishes were searched using a dissec-ion microscope at 10–20 x magnification for the pres-nce of an oocyte. Classification of oocytes was con-ucted as previously described [28]. Oocyte-cumulusomplexes were classified as Cp or Ex depending onhe expansion of both mural granulosa and cumulusells. Those oocytes with both a compact cumulus andompact mural granulosa were classified as Cp. Oo-ytes with any sign of expansion of either the cumulusr the mural granulosa were classified as Ex. Degener-ting oocytes (having a misshapen, shrunken, or denseytoplasm) were discarded. Because of their relativebundance compared to that of Cp oocytes, their higheraturation rate, and equivalent developmental compe-
ence [29], only Ex oocytes were used in these studies.Oocytes were washed once in EH medium (40%
CM199 with Earle’s salts (Gibco), 40% TCM199ith Hanks salts’ and 25 mM Hepes, and 20% fetalovine serum (FBS, Gibco)) and placed in the sameedium in 1.1 mL autosampler borosilicate glass vials
National Scientific Co., Duluth, GA, USA), as previ-usly described [27]. Each vial contained 27–50 oo-ytes. The vial was sealed with a lid and parafilm torevent leakage, wrapped in aluminum foil to limitight exposure, and laid on its side at room temperature22–27 °C) for 13–16 h before the onset of IVM. Thisas done to aid scheduling of onset of maturation:ocytes were recovered in the afternoon, held over-ight, and then placed in maturation culture the follow-ng morning. This treatment has been shown not toffect meiotic or developmental competence of equineocytes [27]. Before placement into culture for IVM,ocytes were washed twice in maturation mediumTCM199 with Earle’s salts, 5 mU/mL FSH (Siouxiochemical Inc., Sioux Center, IA, USA), 10% FBSnd 25 �g/mL gentamycin).
The standard-incubation control IVM treatment foroth trials began at 0700. This consisted of culture inroplets of maturation medium under light white min-ral oil (Sigma Chemical Co., St. Louis, MO, USA) atratio of 10 �L medium per oocyte at 38.2 °C for 24 h
n a humidified atmosphere of 5% CO2 in air, as de-cribed previously [30]. In Trial 1, oocytes were sub-ected to heat for 2 or 4 h at the onset of the 24 h IVMeriod. In Trial 2, oocytes were subjected to heat after0 h (4 h treatment) or 22 h (2 h treatment) of IVMulture. Five replicates were performed in each trial.
To subject the oocytes to heat treatment, in Trial 1,ocytes were placed in equilibrated maturation mediumn a covered borosilicate glass vial and submerged in a
rogrammable water bath (Lindberg/Blue, Ashville, aC, USA) at 42 °C, for 2 or 4 h. To control for possibleffects of handling, groups of oocytes were similarlyackaged and were submerged for 2 or 4 h in a 38.2 °Cater bath (Thermo Electron Corp., Waltham, MA,SA). After treatment in the water baths, oocytes were
emoved from the vials, placed in droplets of matura-ion medium under oil as described above, and incu-ated for an additional 20 or 22 h for a total 24 haturation period. In Trial 2, oocytes were similarly
reated under the same water bath conditions as de-cribed above; however, in this case exposure to heatccurred after 20 or 22 h maturation culture, for a totalf 2 or 4 h exposure at the end of the 24 h total cultureeriod. At the end of maturation culture, oocytes wereenuded by pipetting in 0.05% hyaluronidase. Oocytesithout a polar body but with an intact ooplasmicembrane were fixed in buffered formol saline,ounted on a slide with a 6.5 �L of 9:1 glycerol:PBS
ontaining 2.5 �g/mL Hoechst 33258, and examinedsing fluorescence microscopy to determine the chro-atin configuration.Oocytes exhibiting a polar body were subjected to
CSI. The ICSI procedures were conducted as de-cribed previously using Sp-CZB, CZB-M and CZB-Hedia (based on a Chatot, Ziomek, Bavister mediumodified to increase osmolarity [30]). Briefly, sperm
ells for ICSI were obtained by swim-up separationrom previously frozen equine semen. The diameter ofhe pipette used for sperm injection was 7–8 �m. Prioro sperm injection, 1 �L of sperm suspension waslaced in 3 �L of Sp-CZB containing 10% polyvi-ylpyrrolidone (Sigma) under mineral oil. Once an in-ividual sperm was selected, it was immobilized bypplying a few pulses with the Piezo drill (Prime TechTD., Ibaraki, Japan). Sperm injection was carried out
n a separate 50 �L drop of CZB-M containing 10%BS under oil, and injected oocytes were held inZB-H containing 10% FBS in 5% CO2 in air at 38.2
C until all manipulations were complete.After ICSI, injected oocytes were cultured in
MEM/F-12 medium (Sigma) with 10% FBS underineral oil at 38.2 °C in 5% CO2, 5% O2, and 90% N2
29] in groups of 2–5 oocytes, at a ratio of 1 �Ledium per oocyte. Medium was completely replaced
t 3 and 5 d of culture. Any uncleaved or retardedmbryos were removed at the time of each mediumhange. Presumptive embryos were cultured for a totalf 8 d and development to blastocyst was determinedy morphological examination at 7 and 8 d under aissection microscope at 63 X. Embryos not classified
s blastocysts morphologically were fixed and stained
wehpcpmbg
2eu
meema
prgeTbArebctscptttccblmeCti1o
s
mmotod
vavgttafVw7dMmt
fPi0IRUplwtd(m5sz
RimRCtHcR
4 C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
ith Hoechst 33258 as described above for oocytes, toxamine the number and status of nuclei. Embryosaving �64 nuclei and showing an outer layer of ap-arent trophoblast cells were considered to be blasto-ysts; whereas those with �32 nuclei but lacking ap-arent trophoblast organization were classified asorulae. Those embryos classified morphologically as
lastocysts were frozen in liquid nitrogen for HSPA1Aene expression analysis.
.2. Expression of HSPA1A and 18S rRNA genes inquine blastocysts, produced in vitro and in vivo,sing real time RT-PCR
In vivo-derived blastocysts from a previous experi-ent [10] were also used to: 1) detect HSPA1A mRNA
xpression differences in embryos recovered from ex-rcised mares versus control, and 2) compare HSPA1ARNA expression between in vivo-derived embryos
nd in vitro-produced embryos.For in vivo-derived embryos, horse use was ap-
roved by the Texas A&M University Institutional Ag-iculture Animal Care and Use Committee, usinguidelines set forth by the Federation of Animal Sci-nce Societies. Sixteen mares, two Arabian Horses, twohoroughbreds, and twelve American Quarter Horses,elonging to the Texas A&M University Department ofnimal Science, were utilized for this study. The mares
anged from 2–16 y of age, were randomly assigned toither treatment or control; after two consecutive em-ryo collection attempts, individual mares were given aycle without reproductive manipulation while transi-ioning to the other group, followed by two more con-ecutive embryo collection attempts. Additionally, theycle without reproductive manipulation served as aeriod of exercise conditioning for mares transitioningo the treatment group. Mares had no known reproduc-ive or physical abnormalities, were maintained on pas-ure throughout the duration of the study and daily fedoastal Bermuda grass hay and a 13% crude proteinommercial concentrate. Starting at least 2 wk beforereeding and continuing through the day of embryo col-ection, mares in the exercise group were worked for 30in between 1300 and 1500, six d/wk in a mechanical
xercise pen (free exerciser) at a long trot and canter.ontrol mares were kept under similar conditions, with
he exception of exercise. The mean rectal temperaturencrease from the beginning of exercise to completion was.97 °C, under environmental conditions of the majorityf days �30 °C and 50% humidity [10].
Mare follicular development was monitored by tran-
rectal ultrasonography. Once a follicle greater than 35 am in diameter was detected via ultrasonography,ares were inseminated with fresh stallion semen from
ne of two proven stallions. Embryos were collected byranscervical uterine flush on Day 7 after detection ofvulation. Embryos were graded on a 1–4 scale asescribed by Vanderwall [31].
Both in vitro- and in vivo-derived embryos wereitrified and stored individually in liquid nitrogen untilnalysis. Based on recent evidence, vitrification of initro and in vivo derived bovine embryos may yieldreater embryo survivability after warming than tradi-ional slow-cooling methods [32]. Vitrification of blas-ocysts followed the protocol described by Moussa etl. [33]. Briefly, individual blastocysts were washedour times in modified PBS (Emflush, Har-Vet, Springalley, WI, USA) with 1% FBS. Individual blastocystsere exposed to 7.5% dimethyl-sulfoxide (Sigma) plus.5% ethylene glycol (Sigma) for 3 min and in 18%imethyl-sulfoxide plus 18% ethylene glycol plus 0.4
sucrose for 1 min. Embryos were loaded into 0.25L French straws and plunged directly into liquid ni-
rogen.Prior to each real time RT-PCR experiment, isolated
rozen embryos were identified and thawed in modifiedBS. To isolate RNA, individual embryos were washed
n RNase-free PBS four times, and then placed into a.65 mL microcentrifuge tube (Sorrenson Biosciencenc., Salt Lake City, UT, USA) with 50 �L of Tripureeagent (Roche Applied Science, Indianapolis, IN,SA) and 10 �g glycogen (Sigma). To induce zonaellucida rupture, individual tubes were submerged iniquid nitrogen for a period of 5 min and thawed; thisas repeated four times. A DNase reaction was utilized
o purify RNA, and total RNA concentration wasetermined utilizing a nanodrop spectrophotometerNanodrop DN-1000®, Nanodrop Technologies, Wil-ington, DE, USA). To synthesize first strand cDNA,
00 ng of RNA was reverse-transcribed with Super-cript II (Invitrogen Corp., Carlsbad, CA, USA) en-yme and random octomer primer.
Individual embryos yielded enough total RNA (seeesults) to complete two runs of the real time RT-PCR
n triplicate for the two target genes. Quantification ofRNA was performed using an ABI 7900HT Fasteal-Time PCR System® (Applied Biosystems, Fosterity, CA, USA) with SYBR® Green (Applied Biosys-
ems) as the double-strand DNA-specific fluor forSPA1A cDNA. Normalization of HSPA1A mRNA
oncentration was performed using 18S ribosomalNA, which was measured in replicate reactions using
TAQMAN® probe (Applied Biosystems). Specific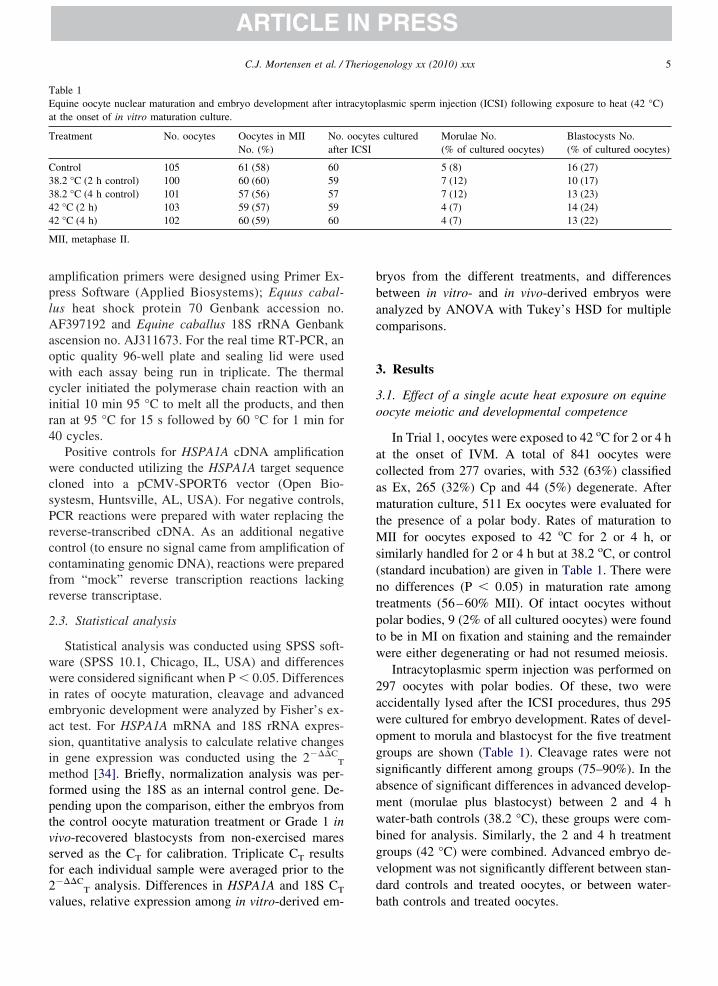
aplAaowcir4
wcsPrccfr
2
wwieasimfptvsf2v
bbac
3
3o
acamtMs(ntptw
2awogsamwbgvd
TEa
T
C3344
M
5C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
mplification primers were designed using Primer Ex-ress Software (Applied Biosystems); Equus cabal-us heat shock protein 70 Genbank accession no.F397192 and Equine caballus 18S rRNA Genbank
scension no. AJ311673. For the real time RT-PCR, anptic quality 96-well plate and sealing lid were usedith each assay being run in triplicate. The thermal
ycler initiated the polymerase chain reaction with annitial 10 min 95 °C to melt all the products, and thenan at 95 °C for 15 s followed by 60 °C for 1 min for0 cycles.
Positive controls for HSPA1A cDNA amplificationere conducted utilizing the HSPA1A target sequence
loned into a pCMV-SPORT6 vector (Open Bio-ystesm, Huntsville, AL, USA). For negative controls,CR reactions were prepared with water replacing theeverse-transcribed cDNA. As an additional negativeontrol (to ensure no signal came from amplification ofontaminating genomic DNA), reactions were preparedrom “mock” reverse transcription reactions lackingeverse transcriptase.
.3. Statistical analysis
Statistical analysis was conducted using SPSS soft-are (SPSS 10.1, Chicago, IL, USA) and differencesere considered significant when P � 0.05. Differences
n rates of oocyte maturation, cleavage and advancedmbryonic development were analyzed by Fisher’s ex-ct test. For HSPA1A mRNA and 18S rRNA expres-ion, quantitative analysis to calculate relative changesn gene expression was conducted using the 2���C
T
ethod [34]. Briefly, normalization analysis was per-ormed using the 18S as an internal control gene. De-ending upon the comparison, either the embryos fromhe control oocyte maturation treatment or Grade 1 inivo-recovered blastocysts from non-exercised mareserved as the CT for calibration. Triplicate CT resultsor each individual sample were averaged prior to the���C
T analysis. Differences in HSPA1A and 18S CT
able 1quine oocyte nuclear maturation and embryo development after intt the onset of in vitro maturation culture.
reatment No. oocytes Oocytes in MIINo. (%)
No.after
ontrol 105 61 (58) 608.2 °C (2 h control) 100 60 (60) 598.2 °C (4 h control) 101 57 (56) 572 °C (2 h) 103 59 (57) 592 °C (4 h) 102 60 (59) 60
II, metaphase II.
alues, relative expression among in vitro-derived em- b
ryos from the different treatments, and differencesetween in vitro- and in vivo-derived embryos werenalyzed by ANOVA with Tukey’s HSD for multipleomparisons.
. Results
.1. Effect of a single acute heat exposure on equineocyte meiotic and developmental competence
In Trial 1, oocytes were exposed to 42 oC for 2 or 4 ht the onset of IVM. A total of 841 oocytes wereollected from 277 ovaries, with 532 (63%) classifieds Ex, 265 (32%) Cp and 44 (5%) degenerate. Afteraturation culture, 511 Ex oocytes were evaluated for
he presence of a polar body. Rates of maturation toII for oocytes exposed to 42 oC for 2 or 4 h, or
imilarly handled for 2 or 4 h but at 38.2 oC, or controlstandard incubation) are given in Table 1. There wereo differences (P � 0.05) in maturation rate amongreatments (56–60% MII). Of intact oocytes withoutolar bodies, 9 (2% of all cultured oocytes) were foundo be in MI on fixation and staining and the remainderere either degenerating or had not resumed meiosis.Intracytoplasmic sperm injection was performed on
97 oocytes with polar bodies. Of these, two wereccidentally lysed after the ICSI procedures, thus 295ere cultured for embryo development. Rates of devel-pment to morula and blastocyst for the five treatmentroups are shown (Table 1). Cleavage rates were notignificantly different among groups (75–90%). In thebsence of significant differences in advanced develop-ent (morulae plus blastocyst) between 2 and 4 hater-bath controls (38.2 °C), these groups were com-ined for analysis. Similarly, the 2 and 4 h treatmentroups (42 °C) were combined. Advanced embryo de-elopment was not significantly different between stan-ard controls and treated oocytes, or between water-
lasmic sperm injection (ICSI) following exposure to heat (42 °C)
cultured Morulae No.(% of cultured oocytes)
Blastocysts No.(% of cultured oocytes)
5 (8) 16 (27)7 (12) 10 (17)7 (12) 13 (23)4 (7) 14 (24)4 (7) 13 (22)
racytop
oocytesICSI
ath controls and treated oocytes.
ao(dwd(irw0wa
2acm(adbbtatsmr
3eu
eite
Iesgp1Egep
Rjarvswta
rcsbGtdtbur
ttcip
TEt
T
C3344
MP � 0.
6 C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
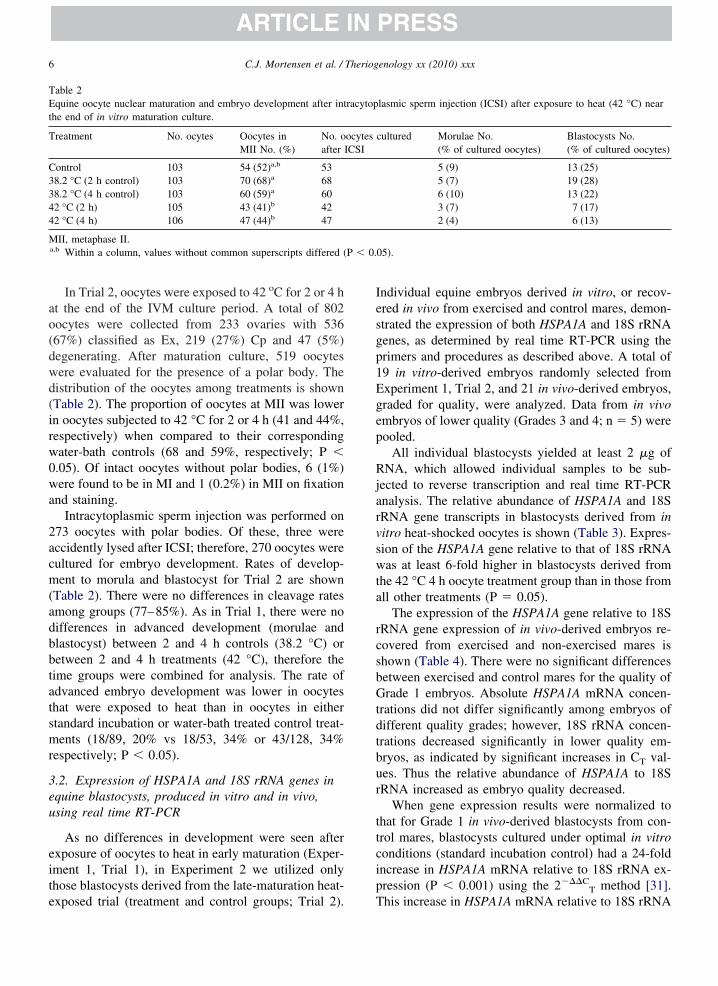
In Trial 2, oocytes were exposed to 42 oC for 2 or 4 ht the end of the IVM culture period. A total of 802ocytes were collected from 233 ovaries with 53667%) classified as Ex, 219 (27%) Cp and 47 (5%)egenerating. After maturation culture, 519 oocytesere evaluated for the presence of a polar body. Theistribution of the oocytes among treatments is shownTable 2). The proportion of oocytes at MII was lowern oocytes subjected to 42 °C for 2 or 4 h (41 and 44%,espectively) when compared to their correspondingater-bath controls (68 and 59%, respectively; P �.05). Of intact oocytes without polar bodies, 6 (1%)ere found to be in MI and 1 (0.2%) in MII on fixation
nd staining.Intracytoplasmic sperm injection was performed on
73 oocytes with polar bodies. Of these, three wereccidently lysed after ICSI; therefore, 270 oocytes wereultured for embryo development. Rates of develop-ent to morula and blastocyst for Trial 2 are shown
Table 2). There were no differences in cleavage ratesmong groups (77–85%). As in Trial 1, there were noifferences in advanced development (morulae andlastocyst) between 2 and 4 h controls (38.2 °C) oretween 2 and 4 h treatments (42 °C), therefore theime groups were combined for analysis. The rate ofdvanced embryo development was lower in oocyteshat were exposed to heat than in oocytes in eithertandard incubation or water-bath treated control treat-ents (18/89, 20% vs 18/53, 34% or 43/128, 34%
espectively; P � 0.05).
.2. Expression of HSPA1A and 18S rRNA genes inquine blastocysts, produced in vitro and in vivo,sing real time RT-PCR
As no differences in development were seen afterxposure of oocytes to heat in early maturation (Exper-ment 1, Trial 1), in Experiment 2 we utilized onlyhose blastocysts derived from the late-maturation heat-
able 2quine oocyte nuclear maturation and embryo development after int
he end of in vitro maturation culture.
reatment No. ocytes Oocytes inMII No. (%)
No. oafter I
ontrol 103 54 (52)a,b 538.2 °C (2 h control) 103 70 (68)a 688.2 °C (4 h control) 103 60 (59)a 602 °C (2 h) 105 43 (41)b 422 °C (4 h) 106 47 (44)b 47
II, metaphase II.a,b Within a column, values without common superscripts differed (
xposed trial (treatment and control groups; Trial 2). T
ndividual equine embryos derived in vitro, or recov-red in vivo from exercised and control mares, demon-trated the expression of both HSPA1A and 18S rRNAenes, as determined by real time RT-PCR using therimers and procedures as described above. A total of9 in vitro-derived embryos randomly selected fromxperiment 1, Trial 2, and 21 in vivo-derived embryos,raded for quality, were analyzed. Data from in vivombryos of lower quality (Grades 3 and 4; n � 5) wereooled.
All individual blastocysts yielded at least 2 �g ofNA, which allowed individual samples to be sub-
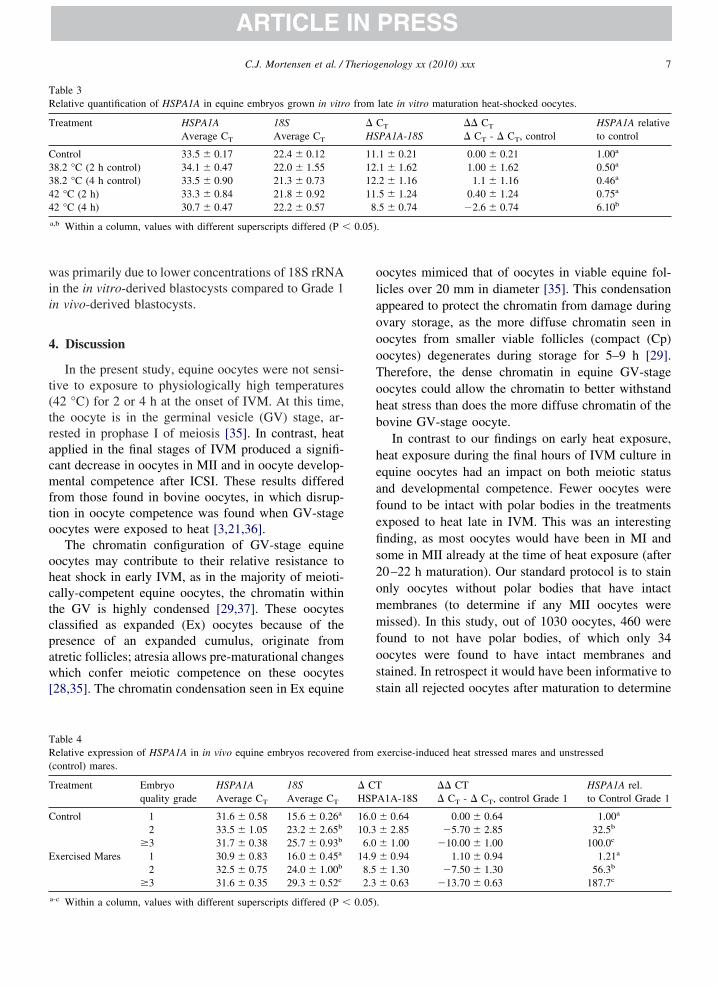
ected to reverse transcription and real time RT-PCRnalysis. The relative abundance of HSPA1A and 18SRNA gene transcripts in blastocysts derived from initro heat-shocked oocytes is shown (Table 3). Expres-ion of the HSPA1A gene relative to that of 18S rRNAas at least 6-fold higher in blastocysts derived from
he 42 °C 4 h oocyte treatment group than in those fromll other treatments (P � 0.05).
The expression of the HSPA1A gene relative to 18SRNA gene expression of in vivo-derived embryos re-overed from exercised and non-exercised mares ishown (Table 4). There were no significant differencesetween exercised and control mares for the quality ofrade 1 embryos. Absolute HSPA1A mRNA concen-
rations did not differ significantly among embryos ofifferent quality grades; however, 18S rRNA concen-rations decreased significantly in lower quality em-ryos, as indicated by significant increases in CT val-es. Thus the relative abundance of HSPA1A to 18SRNA increased as embryo quality decreased.
When gene expression results were normalized tohat for Grade 1 in vivo-derived blastocysts from con-rol mares, blastocysts cultured under optimal in vitroonditions (standard incubation control) had a 24-foldncrease in HSPA1A mRNA relative to 18S rRNA ex-ression (P � 0.001) using the 2���C
T method [31].
lasmic sperm injection (ICSI) after exposure to heat (42 °C) near
cultured Morulae No.(% of cultured oocytes)
Blastocysts No.(% of cultured oocytes)
5 (9) 13 (25)5 (7) 19 (28)6 (10) 13 (22)3 (7) 7 (17)2 (4) 6 (13)
05).
racytop
ocytesCSI
his increase in HSPA1A mRNA relative to 18S rRNA
wii
4
t(tracmfto
ohctcpaw[
olaoooTohb
heafefis2ommfoss
TR
T
C3344
0.05)
TR(
T
C
E
7C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
as primarily due to lower concentrations of 18S rRNAn the in vitro-derived blastocysts compared to Grade 1n vivo-derived blastocysts.
. Discussion
In the present study, equine oocytes were not sensi-ive to exposure to physiologically high temperatures42 °C) for 2 or 4 h at the onset of IVM. At this time,he oocyte is in the germinal vesicle (GV) stage, ar-ested in prophase I of meiosis [35]. In contrast, heatpplied in the final stages of IVM produced a signifi-ant decrease in oocytes in MII and in oocyte develop-ental competence after ICSI. These results differed
rom those found in bovine oocytes, in which disrup-ion in oocyte competence was found when GV-stageocytes were exposed to heat [3,21,36].
The chromatin configuration of GV-stage equineocytes may contribute to their relative resistance toeat shock in early IVM, as in the majority of meioti-ally-competent equine oocytes, the chromatin withinhe GV is highly condensed [29,37]. These oocyteslassified as expanded (Ex) oocytes because of theresence of an expanded cumulus, originate fromtretic follicles; atresia allows pre-maturational changeshich confer meiotic competence on these oocytes
28,35]. The chromatin condensation seen in Ex equine
able 3elative quantification of HSPA1A in equine embryos grown in vitr
reatment HSPA1AAverage CT
18SAverage CT
ontrol 33.5 � 0.17 22.4 � 0.128.2 °C (2 h control) 34.1 � 0.47 22.0 � 1.558.2 °C (4 h control) 33.5 � 0.90 21.3 � 0.732 °C (2 h) 33.3 � 0.84 21.8 � 0.922 °C (4 h) 30.7 � 0.47 22.2 � 0.57
a,b Within a column, values with different superscripts differed (P �
able 4elative expression of HSPA1A in in vivo equine embryos recovered
control) mares.
reatment Embryoquality grade
HSPA1AAverage CT
18SAverage CT
ontrol 1 31.6 � 0.58 15.6 � 0.26a
2 33.5 � 1.05 23.2 � 2.65b
�3 31.7 � 0.38 25.7 � 0.93b
xercised Mares 1 30.9 � 0.83 16.0 � 0.45a
2 32.5 � 0.75 24.0 � 1.00b
�3 31.6 � 0.35 29.3 � 0.52c
a-c Within a column, values with different superscripts differed (P � 0.05)
ocytes mimiced that of oocytes in viable equine fol-icles over 20 mm in diameter [35]. This condensationppeared to protect the chromatin from damage duringvary storage, as the more diffuse chromatin seen inocytes from smaller viable follicles (compact (Cp)ocytes) degenerates during storage for 5–9 h [29].herefore, the dense chromatin in equine GV-stageocytes could allow the chromatin to better withstandeat stress than does the more diffuse chromatin of theovine GV-stage oocyte.
In contrast to our findings on early heat exposure,eat exposure during the final hours of IVM culture inquine oocytes had an impact on both meiotic statusnd developmental competence. Fewer oocytes wereound to be intact with polar bodies in the treatmentsxposed to heat late in IVM. This was an interestingnding, as most oocytes would have been in MI andome in MII already at the time of heat exposure (after0–22 h maturation). Our standard protocol is to stainnly oocytes without polar bodies that have intactembranes (to determine if any MII oocytes wereissed). In this study, out of 1030 oocytes, 460 were
ound to not have polar bodies, of which only 34ocytes were found to have intact membranes andtained. In retrospect it would have been informative totain all rejected oocytes after maturation to determine
late in vitro maturation heat-shocked oocytes.
T
PA1A-18S�� CT
� CT - � CT, controlHSPA1A relativeto control
1 � 0.21 0.00 � 0.21 1.00a
1 � 1.62 1.00 � 1.62 0.50a
2 � 1.16 1.1 � 1.16 0.46a
5 � 1.24 0.40 � 1.24 0.75a
5 � 0.74 �2.6 � 0.74 6.10b
.
exercise-induced heat stressed mares and unstressed
A1A-18S�� CT� CT - � CT, control Grade 1
HSPA1A rel.to Control Grade 1
� 0.64 0.00 � 0.64 1.00a
� 2.85 �5.70 � 2.85 32.5b
� 1.00 �10.00 � 1.00 100.0c
� 0.94 1.10 � 0.94 1.21a
� 1.30 �7.50 � 1.30 56.3b
� 0.63 �13.70 � 0.63 187.7c
o from
� CHS
11.12.12.11.8.
from
� CTHSP
16.010.36.0
14.98.52.3
.

te
lbtdedbefuhcpI
oeticrb4eacmcsf
dgoflsHcCcfwemTmh
gb[amb
wofRTRztOemossm3ese
taec[ettb
lbcrgbf4sstse
8 C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
he chromatin status of those showing cellular degen-ration (lacking an intact ooplasmic membrane).
After ICSI, fewer oocytes treated with heat duringate IVM culture had advanced development (morula orlastocyst), than did oocytes in either standard incuba-ion or water-bath controls, indicating decreased oocyteevelopmental competence associated with late heatxposure. Similarly, Ju et al. [6] reported a significantecrease in proportion of bovine oocytes developing tolastocysts in vitro after exposure to 42 °C for 4 h at thend of a 20 h standard IVM culture. These authorsurther reported heat-treated (42 °C, 2 or 4 h) oocytesnderwent a change in spindle morphology. In anot-er report, porcine oocytes underwent chromosomalhanges and exhibited decreased developmental com-etence after exposure to 41.5 °C for up to 4 h after theVM period [38].
To our knowledge, this is the first report on analysisf expression of HSPA1A mRNA and 18S rRNA inquine embryos. We chose HSPA1A as our target geneo analyze early equine embryonic response to heat-nduced stress. Heat exposure applied during late oo-yte maturation increased HSPA1A gene expression,elative to that of 18S rRNA, in the subsequent equinelastocyst. Similarly, subjection of murine oocytes to a3 °C for 30 min induced measurable HSPA1A genexpression in early one-cell embryos [23]. Christians etl. [24] and Chandolia et al. [39] both reported in-reases in HSPA1A gene activity in heat-stressed earlyurine and bovine embryos respectively, and con-
luded that HSPA1A was the first major inducible heathock gene important for protecting embryonic cellsrom cellular stress.
High quality in vivo-recovered embryos showed noifferences in HSPA1A mRNA relative to 18S rRNAene expression, regardless of their source (exercisedr control mares). However, in our previous report, weound that exercised mares were more likely to produceower-quality embryos [10] and in this study we ob-erved significant increase in the relative expression ofSPA1A to 18S rRNA genes as embryo quality de-
reased. This was related to an increase in 18S rRNA
T values, thus indicating decreased 18S rRNA con-entrations. The lower concentrations of 18S rRNAound in Grade 2 and lower in vivo-recovered embryosere similar to those measured in in vitro-produced
mbryos. Absolute HSPA1A mRNA CT values re-ained relatively unchanged across quality grades.hese results were notable, because 18S rRNA, a com-only used “housekeeping gene”, has been reported to
ave the least variability among a host of housekeeping t
enes in IVF- or somatic-cell nuclear transfer-derivedovine blastocysts [40]. Bilodeau-Goeseels and Shultz41] stated that 28S and 18S rRNA together make uppproximately 70% of the total RNA content in a mam-alian cell, and are useful components on which to
ase estimates of RNA content in early embryos.In the current study, the expression of 18S rRNA
as not stable across embryos from different sources,r of different quality grades. Perhaps there is no per-ect “housekeeping gene” for assessing changes inNA content during development of the early embryo.he embryonic genome is initially silent and manyNAs including 18S rRNA are degraded after fertili-ation [41]. The embryonic genome activates aroundhe four-cell stage of embryonic development [42].thers have found housekeeping genes including glyc-
raldehyde-3-phosphate (GAPDH) and �-actin (ACTB)ay be unsuitable as reference genes in nuclear transfer
r IVF embryos comparisons, due to aberrant expres-ion [43]. In addition, Smith et al. [44] reported aignificant (40%) decrease in �-actin gene expressioneasured via real time RT-PCR in Day-7 quality Gradebovine embryos when compared to quality Grade 1
mbryos. Perhaps these should not be surprising, con-idering the extensive changes in gene expression inarly embryos.
The obvious question is why 18S rRNA concentra-ions were decreased in lower grade in vivo embryosnd in vitro produced embryos. Perhaps these embryosxperienced stresses that caused 18S rRNA to beleaved as part of a global reduction in translation45,46]. The preferential maintenance of HSPA1A genexpression would allow this gene product to help main-ain cellular function by acting as molecular chaperoneso stabilize or refold proteins damaged by heat, and bylocking apoptosis [14,47].
Alternatively, transcription of rRNA genes could beower in stressed embryos. A study of in vitro-producedovine embryos identified an 8-fold increase in rRNAontent between morulae and blastocysts [41]; thus aeduction in transcription of any gene would result in areatly lowered relative abundance in the resultinglastocyst. In the pig, in vivo-produced embryos wereound to activate rRNA genes synchronously at the-cell stage and activity continued to the blastocysttage [48]. However in vitro produced embryos of alltages had many fewer cells with active rRNA genes. Inhe present study, perhaps the lower 18S rRNA expres-ion in the in vitro-derived and lower quality in vivombryos reflected a developmental retardation, and
hese embryos had not quite achieved the developmen-
tT“sgrp
poomtoecHeiHqct
A
R
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
9C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
al-stage dependent up-regulation in rRNA expression.hese results supported previous findings that evenoptimal” culture conditions can perturb gene expres-ion in developing embryos [24,49–52]. Taken to-ether, this could explain our observations of lowerelative expression of 18S rRNA in in vitro, in com-arison to in vivo Grade 1 equine blastocysts.
We concluded that GV-stage equine oocytes ap-eared to be able to withstand a one-time heat exposuref 42 °C for up to 4 h in vitro. However, heat treatmentf late-stage maturing oocytes reduced numbers of nor-al MII oocytes, and reduced developmental compe-
ence, suggesting that heat may have a direct effect onocytes, resulting in reduced embryo production inxposed mares. We found that exposure of equine oo-ytes to 42 °C for 4 h at the end of maturation increasedSPA1A gene transcript levels relative to 18S rRNA
xpression in resulting in vitro-produced blastocysts. Inn vivo-derived blastocysts, increased expression of theSPA1A relative to the 18S rRNA occurred as embryouality decreased. All in vitro-produced equine blasto-ysts exhibited altered gene expression patterns relativeo normal in vivo-derived blastocysts.
cknowledgments
This study was funded in part by the Link Equineesearch Fund, Texas A&M University.
eferences
[1] Lenz RW, Ball GD, Leibfried ML, Ax RL, First NL. In vitromaturation and fertilization of bovine oocytes are temperature-dependent processes. Biol Reprod 1983;29:173–9.
[2] Hansen PJ, Drost M, Rivera RM, Paula-Lopes FF, al-KatananiYM, Krininger CE 3rd, Chase CC Jr. Adverse impact of heatstress on embryo production: causes and strategies for mitiga-tion. Theriogenology 2001;55:91–103.
[3] Roth Z, Hansen PJ. Disruption of nuclear maturation and rear-rangement of cytoskeletal elements in bovine oocytes exposedto heat shock during maturation. Reproduction 2005;129:235–44.
[4] Roth Z. Heat stress, the follicle, and its enclosed oocyte: mech-anisms and potential strategies to improve fertility in dairycows. Reprod Dom Anim 2008;(Suppl 43):238–44.
[5] De Rensis F, Scaramuzzi RJ. Heat stress and seasonal effects onreproduction in the dairy cow- a review. Theriogenology 2003;60:1139–51.
[6] Ju JC, Jiang S, Tseng JK, Parks JE, Yang X. Heat shock reducesdevelopmental competence and alters spindle configuration ofbovine oocytes. Theriogenology 2005;64:1677–89.
[7] Squires EL, Carnevale EM, McCue PM, Bruemmer JE. Embryotechnologies in the horse. Theriogenology 2003;59:151–70.
[8] Hodgson DR, Davis RE, McConaghy FF. Thermoregulation in
the horse in response to exercise. Br Vet J 1994;150:219–35.[9] McCutcheon LJ, Geor RJ. Influence of training on sweatingresponses during submaximal exercise in horses. J Appl Physiol2000;89:2463–71.
10] Mortensen CJ, Choi YH, Hinrichs K, Ing NH, Kraemer DC,Vogelsang SG, Vogelsang MM. Embryo recovery from exer-cised mares. Anim Reprod Sci 2009;110:237–44.
11] Lanska V, Chmelikova E, Sedmikova M, Petr J, Rajmon R,Jeseta M, Rozinek J. Expression of heat shock protein 70 in pigoocytes: heat shock response during oocyte growth. Anim Re-prod Sci 2006;96:154–64.
12] Ellis RJ, Hemmingsen SM. Molecular chaperones: proteinsessential for the biogenesis of some macromolecular structures.Trends Bioch Sci 1989;14:339–42.
13] Didelot C, Schmitt E, Brunet M, Maingret L, Parcellier A,Garrido C. Heat shock proteins: endogenous modulators ofapoptotic cell death. Handb Exp Pharmacol 2006;172:171–98.
14] Kampinga HH. Chaperones in preventing protein denaturationin living cells and protecting against cellular stress. Handb ExpPharmacol 2006;172:1–42.
15] Bensaude O, Babinet C, Morange M, Jacob F. Heat shockproteins, first major products of zygotic gene activity in mouseembryo. Nature 1983;305:331–33.
16] Thompson DM, Lu C, Green PJ, Parker R. tRNA cleavage is aconserved response to oxidative stress in eukaryotes. RNA2008;14:2095–103.
17] Mirkes PE. Hyperthermia-induced heat shock response andthermotolerance in postimplantation rat embryos. Dev Biol1987;119:115–22.
18] Heikkila JJ, Browder LW, Gedamu L, Nickells RW, Shultz GA.Heat-shock gene expression in animal embryonic systems. CanJ Genet Cytol 1986;28:1093–105.
19] Bernardini C, Fantinati P, Castellani G, Forni M, Zannoni A,Seren E, Bacci ML. Alteration of constitutive heat shock protein70 (HSC70) production by in vitro culture of porcine preim-planted embryos. Vet Res Commun 2003;(Suppl 27):575–8.
20] Putney DJ, Malayer JR, Gross TS, Thatcher WW, Hansen PJ,Drost M. Heat stress-induced alterations in the synthesis andsecretion of proteins and prostaglandins by cultured bovineconceptuses and uterine endometrium. Biol Reprod 1988;39:717–28.
21] Edwards JL, Hansen PJ. Differential responses of bovine oo-cytes and preimplantation embryos to heat shock. Mol ReprodDev 1997;46:138–45.
22] Curci A, Bevilacqua A, Mangia F. Lack of heat-shock responsein preovulatory mouse oocytes. Dev Biol 1987;123:154–60.
23] Fiorenza MT, Bevilacqua A, Canterini S, Torcia S, PontecorviM, Mangia F. Early transcriptional activation of the hsp70.1gene by osmotic stress in one-cell embryos of the mouse. BiolReprod 2004;70:1606–13.
24] Christians E, Campion E, Thompson EM, Renard JP. Expres-sion of the HSP70.1 gene, a landmark of early zygotic activityin the mouse embryo, is restricted to the first burst of transcrip-tion. Development 1995;121:113–22.
25] Fiorenza MT, Mangia F. Quantitative RT-PCR amplification ofRNA in single mouse oocytes and preimplantation embryos.Bio Tech 1998;24:618–23.
26] Hunt CR, Dix DJ, Sharma GG, Pandita RK, Gupta A, Funk M,Pandita TK. Genomic instability and enhanced radiosensitivityin Hsp70.1- and Hsp70.3 deficient mice. Mol Cell Biol 2004;24:899–911.
27] Choi YH, Love LB, Varner DD, Hinrichs K. Holding immature
equine oocytes in the absence of meiotic inhibitors: effect
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
10 C.J. Mortensen et al. / Theriogenology xx (2010) xxx
ARTICLE IN PRESS
on germinal vesicle chromatin and blastocyst developmentafter intracytoplasmic sperm injection. Theriogenology 2006;66:955–63.
28] Hinrichs K, Williams KA. Relationship among oocyte-cumulusmorphology, follicular atresia, initial chromatin configuration,and oocyte meiotic competence in the horse. Biol Reprod 1997;57:377–84.
29] Hinrichs K, Choi YH, Love LB, Varner DD, Love CC, Walck-enaer BE. Chromatin configuration within the germinal vesicleof horse oocytes: changes post mortem and relationship tomeiotic and developmental competence. Biol Reprod 2005;72:1142–50.
30] Choi YH, Chung YG, Walker SC, Westhusin ME, Hinrichs K.In vitro development of equine nuclear transfer embryos: effectsof oocyte maturation media and amino acid composition duringembryo culture. Zygote 2003;11:77–86.
31] Vanderwall DK. Early embryonic development and evaluationof equine embryo viability. Vet Clin North Am Equine Pract1996;12:61–83.
32] Yu XL, Deng W, Liu FJ, Li YH, Li XX, Zhang YL, Zan LS.Closed pulled straw vitrification of in vitro-produced and invivo-produced bovine embryos. Theriogenology 2010 Mar1;73(4):474–9. Epub 2009 Dec 3.
33] Moussa M, Bersinger I, Doligez P, Guignot F, Duchamp G,Vidament M, Mermillod P, Bruyas JF. In vitro comparisons oftwo cryopreservation techniques for equine embryos: slow-cooling and open pulled straw (OPS) vitrification. Theriogenol-ogy 2005;64:1619–32.
34] Livak KJ, Schmittgen TD. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2���C
T method.Methods 2001;25:402–8.
35] Hinrichs K, Schmidt AL. Meiotic competence in horse oocytes:interactions among chromatin configuration, follicle size and cu-mulus morphology, and season. Biol Reprod 2000;62:1402–8.
36] Payton RR, Romar R, Coy P, Saxton AM, Lawrence JL, Ed-wards JL. Susceptibility of bovine germinal vesicle-stage oo-cytes from antral follicles to direct effects of heat stress in vitro.Biol Reprod 2004;71:1303–8.
37] Hinrichs K, Schmidt AL, Friedman PP, Selgrath JP, MartinMG. In vitro maturation of horse oocytes: characterization ofchromatin configuration using fluorescence microscopy. BiolReprod 1993;48:363–70.
38] Ju JC, Tseng JK. Nuclear and cytoskeletal alterations of in vitromatured porcine oocytes under hyperthermia. Mol Reprod Dev2004;68:125–33.
39] Chandolia RK, Peltier MR, Tian W, Hansen PJ. Transcriptionalcontrol of development, protein synthesis, and heat-inducedheat shock protein 70 synthesis in 2-cell bovine embryos. Biol
Reprod 1999;61:1644–48.40] Ross PJ, Wang K, Beyhan Z, Kocabas A, Cibelli JB. House-keeping gene expression of bovine fertilized and cloned em-bryos. Reprod Fertil Dev 2009;21:195–6 (abstract).
41] Bilodeau-Goeseels S, Schultz GA. Changes in ribosomal ribo-nucleic acid content within in vitro-produced embryos. BiolReprod 1997;56:1323–29.
42] Brinkso SP, Ball BA, Ignotz GG, Thomas PG, Currie WB,Ellington JE. Initiation of transcription and nucleogenesis inequine embryos. Mol Reprod Dev 1995;42:298–302.
43] Bower NI, Moser RJ, Hill JR, Lehnert SA. Universal referencemethod for real-time PCR gene expression analysis of preim-plantation embryos. Biotechniques 2007;42:199–206.
44] Smith C, Berg D, Beaumont S, Standley NT, Wells DN, PfefferPL. Simultaneous gene quantitation of multiple genes in indi-vidual bovine nuclear transfer blastocysts. Reproduction 2007;133:231–42.
45] Holcik M, Sonenberg N. Translation control in stress and ap-optosis. Nat Rev Mol Cell Biol 2005;6:318–27.
46] Thompson EM, Legouy E, Christians E, Renard JP. Progressivematuration of chromatin structure regulates HSP70.1 gene ex-pression in the preimplantation mouse embryo. Development1995;121:3425–37.
47] Mosser DD, Caron AW, Bourget L, Meriin AB, Sherman MY,Morimoto RI, Massie B. The chaperone function of hsp70 isrequired for protection against stress-induced apoptosis. MolCell Biol 2000;20:7146–59.
48] Bjerregaard B, Pedersen HG, Jakobsen AS, Rickords LF, Lai L,Cheong HT, Samuel M, Prather RS, Strejcek F, Rasmussen ZR,Laurincik J, Niemann H, Maddox-Hyttel P, Thomsen PD. Ac-tivation of ribosomal RNA genes in porcine embryos producedin vitro or by somatic cell nuclear transfer. Mol Reprod Dev2007;74:35–41.
49] Rizos D, Lonergan P, Boland MP, Arroyo-Garcia R, Pintado B,de la Fuente J, Gutierrez-Adan A. Analysis of differential mes-senger RNA expression between bovine blastocysts produced indifferent culture systems: implications for blastocyst quality.Biol Reprod 2002;66:589–95.
50] Gardner DK, Lane M. Ex vivo early embryo development andeffects on gene expression and imprinting. Reprod Fertil Dev2005;17:361–70.
51] de Oliveira AT, Lopes RF, Rodrigues JL. Gene expression anddevelopmental competence of bovine embryos produced invitro under varying embryo density conditions. Theriogenology2005;64:1559–72.
52] McElroy SL, Kim JH, Kim S, Jeong YW, Lee EG, Park SM,Hossein MS, Koo OJ, Abdul Hashem MD, Jang G, Kang SK,Lee BC, Hwang WS. Effects of culture conditions and nucleartransfer protocols on blastocysts formation and mRNA expres-sion in pre-implantation porcine embryos. Theriogenology
2008;69:416–25.