apnea, glial apoptosis and neuronal plasticity in the arousal pathway of victims of sids
TRANSCRIPT

www.elsevier.com/locate/forsciint
Forensic Science International 149 (2005) 205–217
Apnea, glial apoptosis and neuronal plasticity in the arousal
pathway of victims of SIDS
T. Sawaguchia,*, I. Katob, P. Francoc, M. Sottiauxc, H. Kadhimc, S. Shimizud,J. Groswasserc, H. Togarib, M. Kobayashie, H. Nishidaf, A. Sawaguchia, A. Kahnc
aDepartment of Legal Medicine, Tokyo Women’s Medical University, 8-1 Kawada-cho, Shinjuku, 162-8666 Tokyo, JapanbDepartment of Paediatrics, Nagoya City University, 1 Aza Kawasumi, Mizuho-cho, Mizuho-ku, Nagoya, Aichi 467-8601, Japan
cBrussels Free University Children’s Hospital Reine Fabiola, Ave. J.J. Crocq 15, B-1020 Brussels, BelgiumdDepartment of Hygiene and Preventive Medicine, 8-1 Kawada-cho, Shinjuku, 162-8666 Tokyo, Japan
eDepartment of Pathology, Tokyo Women’s Medical University, 8-1 Kawada-cho, Shinjuku, 162-8666 Tokyo, JapanfMaternal and Perinatal Center, Tokyo Women’s Medical University, 8-1 Kawada-cho, Shinjuku, 162-8666 Tokyo, Japan
Available online 15 December 2004
Abstract
Of 27,000 infants whose sleep-wake characteristics were studied under the age of 6 months, 38 died unexpectedly 2–12
weeks after the sleep recording in a pediatric sleep laboratory. Of these infants, 26 died of sudden infant death syndrome (SIDS),
and 12 of definitely identified causes. The frequency and duration of sleep apneas were analysed. Sleep recordings and brainstem
histopathology were studied to elucidate the possible relationship between sleep apnea and neuropathological changes within
the arousal system. Immunohistochemical analyses were conducted using tryptophan hydroxylase (TrypH), a serotonin
synthesizing enzyme, and growth-associated phosphoprotein 43 (GAP43), a marker of synaptic plasticity. The terminal-
deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) method was used for apoptosis. The pathological and
physiological data were correlated for each infant.
In the SIDS victims, statistically significant positive correlations were seen between the number of TrypH-positive neurons in
the dorsal raphe nucleus of the midbrain and the duration of central apneas (p = 0.03), between the number of TUNEL-positive
glial cells in the pedunculopontine tegmental nucleus (PPTN) and the average number of spines in GAP43-positive neurons in
the PPTN (p = 0.04).
These findings in the dorsal raphe nucleus of the midbrain and PPTN, that play important roles in the arousal pathway suggest
a possible link between changes in arousal and SIDS.
# 2004 Elsevier Ireland Ltd. All rights reserved.
Keywords: Tryptophan-hydroxylase (TrypH); Terminal-deoxynucleotidyl transferase mediated dUTP nick end labeling (TUNEL); Growth
associated phosphoprotein 43 (GAP43); Apnea; Nucleus raphe; Periaqueductal gray matter; Pedunculopontine tegmental nucleus (PPTN)
* Corresponding author. Tel.: +81 3 5269 7300;
fax: +81 3 5269 7300.
E-mail address: [email protected]
(T. Sawaguchi).
0379-0738/$ – see front matter # 2004 Elsevier Ireland Ltd. All rights r
doi:10.1016/j.forsciint.2004.10.015
1. Introduction
Several etiological hypotheses have been proposed to
explain the mechanisms responsible for the sudden infant
death syndrome (SIDS), including pathological processes in
the brain stem which can cause apnea [1–3] or disturbances
of arousal [4–9].
eserved.
T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217206
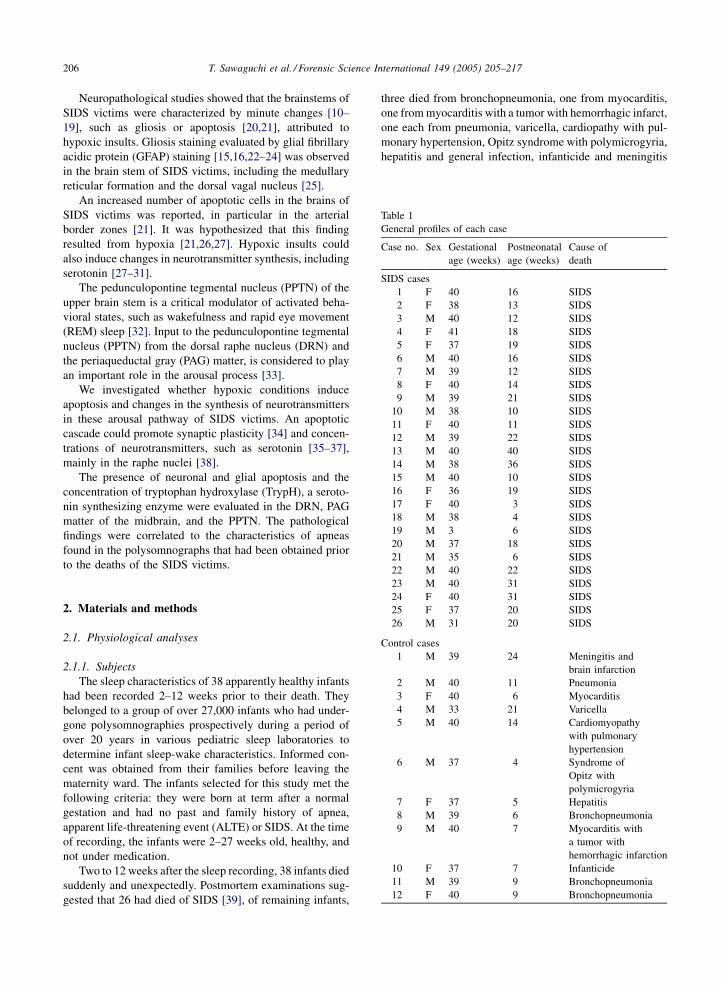
Table 1
General profiles of each case
Case no. Sex Gestational
age (weeks)
Postneonatal
age (weeks)
Cause of
death
SIDS cases
1 F 40 16 SIDS
2 F 38 13 SIDS
3 M 40 12 SIDS
4 F 41 18 SIDS
5 F 37 19 SIDS
6 M 40 16 SIDS
7 M 39 12 SIDS
8 F 40 14 SIDS
9 M 39 21 SIDS
10 M 38 10 SIDS
11 F 40 11 SIDS
12 M 39 22 SIDS
13 M 40 40 SIDS
14 M 38 36 SIDS
15 M 40 10 SIDS
16 F 36 19 SIDS
17 F 40 3 SIDS
18 M 38 4 SIDS
19 M 3 6 SIDS
20 M 37 18 SIDS
21 M 35 6 SIDS
22 M 40 22 SIDS
Neuropathological studies showed that the brainstems of
SIDS victims were characterized by minute changes [10–
19], such as gliosis or apoptosis [20,21], attributed to
hypoxic insults. Gliosis staining evaluated by glial fibrillary
acidic protein (GFAP) staining [15,16,22–24] was observed
in the brain stem of SIDS victims, including the medullary
reticular formation and the dorsal vagal nucleus [25].
An increased number of apoptotic cells in the brains of
SIDS victims was reported, in particular in the arterial
border zones [21]. It was hypothesized that this finding
resulted from hypoxia [21,26,27]. Hypoxic insults could
also induce changes in neurotransmitter synthesis, including
serotonin [27–31].
The pedunculopontine tegmental nucleus (PPTN) of the
upper brain stem is a critical modulator of activated beha-
vioral states, such as wakefulness and rapid eye movement
(REM) sleep [32]. Input to the pedunculopontine tegmental
nucleus (PPTN) from the dorsal raphe nucleus (DRN) and
the periaqueductal gray (PAG) matter, is considered to play
an important role in the arousal process [33].
We investigated whether hypoxic conditions induce
apoptosis and changes in the synthesis of neurotransmitters
in these arousal pathway of SIDS victims. An apoptotic
cascade could promote synaptic plasticity [34] and concen-
trations of neurotransmitters, such as serotonin [35–37],
mainly in the raphe nuclei [38].
The presence of neuronal and glial apoptosis and the
concentration of tryptophan hydroxylase (TrypH), a seroto-
nin synthesizing enzyme were evaluated in the DRN, PAG
matter of the midbrain, and the PPTN. The pathological
findings were correlated to the characteristics of apneas
found in the polysomnographs that had been obtained prior
to the deaths of the SIDS victims.
23 M 40 31 SIDS
24 F 40 31 SIDS
25 F 37 20 SIDS
26 M 31 20 SIDS
Control cases
1 M 39 24 Meningitis and
brain infarction
2 M 40 11 Pneumonia
3 F 40 6 Myocarditis
4 M 33 21 Varicella
5 M 40 14 Cardiomyopathy
with pulmonary
hypertension
6 M 37 4 Syndrome of
Opitz with
polymicrogyria
7 F 37 5 Hepatitis
8 M 39 6 Bronchopneumonia
9 M 40 7 Myocarditis with
a tumor with
hemorrhagic infarction
10 F 37 7 Infanticide
11 M 39 9 Bronchopneumonia
12 F 40 9 Bronchopneumonia
2. Materials and methods
2.1. Physiological analyses
2.1.1. Subjects
The sleep characteristics of 38 apparently healthy infants
had been recorded 2–12 weeks prior to their death. They
belonged to a group of over 27,000 infants who had under-
gone polysomnographies prospectively during a period of
over 20 years in various pediatric sleep laboratories to
determine infant sleep-wake characteristics. Informed con-
cent was obtained from their families before leaving the
maternity ward. The infants selected for this study met the
following criteria: they were born at term after a normal
gestation and had no past and family history of apnea,
apparent life-threatening event (ALTE) or SIDS. At the time
of recording, the infants were 2–27 weeks old, healthy, and
not under medication.
Two to 12 weeks after the sleep recording, 38 infants died
suddenly and unexpectedly. Postmortem examinations sug-
gested that 26 had died of SIDS [39], of remaining infants,
three died from bronchopneumonia, one from myocarditis,
one from myocarditis with a tumor with hemorrhagic infarct,
one each from pneumonia, varicella, cardiopathy with pul-
monary hypertension, Opitz syndrome with polymicrogyria,
hepatitis and general infection, infanticide and meningitis

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217 207
complicated by brain infarction. These 12 infants form the
control group. The general profiles of the subjects are shown
in Table 1.
2.1.2. Polysomnography
Eight-hour overnight sleep studies were conducted in a
sleep laboratory, following standard techniques [40–43].
The recordings were made in a quiet and darkened room
at an ambient temperature between 20 and 23 8C. All infants
slept in supine position without restraints. Recording started
around 9:00 p.m. The infants were observed continuously
during recording and were fed on demand. Their behavior
and any nursing intervention were recorded manually. Paci-
fiers were withheld during the recording. The following
variables were recorded simultaneously: two channels scalp
electroencephalograms from unilateral central and occipital
areas, horizontal and vertical electroculograms and an elec-
trocardiogram. As for variables of respiration, thoracic
respiratory movements were measured by impedance and
airflow with thermistors taped under both nostrils and on the
side of the mouth. Oxygen saturation was recorded con-
tinuously by a transcutaneous sensor (Nellcor, USA). Gross
body movements were measured with an actigram placed on
one arm. The data were collected on a computerized infant
sleep recorder (Alice recording system III, Healthdyne,
USA). Electromyographic recording of the mentalis muscle
was not done.
2.1.3. Method and standard of analysis
Based on the polygraphic recordings, sleep stages and
sleep apneas (when lasting 3 s or longer) were rated accord-
ing to standard definitions [40–45]. Arousals from sleep
were determined by direct observation by the technicians,
and by the recordings of breathing and heart rate changes as
well as eye movements. Arousals were also accompanied by
activation of the actigrams and movement artifacts were
identified on the cardiac and saturation recordings. Apneas
were designated as central apnea when flat tracings were
obtained simultaneously from the strain gauges and the
thermistors. Periodic breathing was defined as a succession
of more than two central apneas separated by a period of
less than 20 s. Obstructive apneas were defined as a flat
tracing recorded from the thermistors with continuous
deflections from the strain gauges. A mixed apnea was
scored when a central apnea was followed immediately by
an obstructive episode. Mixed apneas were scored together
with the obstructive episodes. The frequency of central and
obstructive apneas was measured by dividing the total
number of each apnea by the total sleep time in minutes
and multiplied by 60. The type, frequency (number per hour
of sleep) and duration (in seconds) of sleep apneas were
computed. All recordings were analyzed visually by 2
independent scorers without knowledge of the subject’s
sex and attribution to either the SIDS or the control group.
Discrepancies between scorers were discussed before the
data was computed.
2.2. Pathological analyses
2.2.1. Subjects
A total of 48 paraffin blocks of brain stems were collected
from the autopsied brain of the 38 infants who died unex-
pectedly: 7 blocks from the midbrain, 22 from the pons, and
19 from the medulla oblongata. The maximum time that
elapsed between the estimated time of death and the post-
mortem examination was 24 h.
2.2.2. Neurohistological examination
Hematoxylin-Eosin (HE) stain was used for the standard
staining method for neurohistological examination.
2.2.3. Immunohistochemical examination
The blocks were subjected to immunohistochemical
and nuclear histochemical analysis, using an anti-TrypH
monoclonal antibody (Oncogene, USA; 1:100), and an
anti-GAP43 monoclonal antibody (YLEM, Italy; 1:20).
For the analysis of apoptosis, the TUNEL method (In Situ
Cell Death Detection Kit, Boehringer Mannheim, Ger-
many) [46–49] was employed. The 4-mm-thick sections
from each block were pre-incubated with 1 mM of an
EDTA solution (pH, 8.0), using a microwave oven (Pana-
sonic) at 800 W for 10 min to stain GAP43. After blocking
intrinsic peroxidase by 3% hydrogen peroxide for 5 min
and washing, the sections were incubated with the antibody
for 3 days for TrypH and overnight for GAP43, both at
4 8C. Finally, TrypH and GAP43 were immunohistochemi-
cally examined with the aid of a LSAB2 kit (Dako)
followed by the DAB reaction. Prior to the TUNEL
method, the sections were pretreated by microwave irra-
diation in a 0.01 M citric acid buffer (pH, 6.0) at 800 W for
5 min in a microwave device (Panasonic); then the TUNEL
reaction was performed in the same manner as described
previously [47,48].
2.2.4. Quantification of immunohistochemical and nuclear
histochemical presentations
Measurements were made in the PAG matter and dorsal
raphe nucleus of the midbrain, and the PPTN (compact part:
PPTNc, dissipated parts: PPTNd, and both: PPTNt). The
numbers of TrypH-positive neurons, spines per GAP43-
positive neuron and TUNEL-positive glial cells were
counted manually. Each counting procedure in an area
625 mm � 102 mm was repeated five times at different
overlapping sites and an average was recorded. The density
was expressed as a percentage of the number of reaction-
positive neurons or glial cells divided by the number of total
neurons or glial cells. For the GAP43-positive dendritic
spines, the average number per neuron was calculated and
recorded.
The pathological measurements were made twice by the
same pathologist and data with large standard deviations
were recounted or rejected.
T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217208
2.3. Data analysis
2.3.1. Double-blind analyses
The scorers of the sleep recordings and the pathologist
were not aware of the causes of the infants’ deaths. The
scorers of the sleep recordings had not contact with the
pathologist.
2.3.2. Matching the physiological and pathological data
For each infant, the type and scores of apneas were correla-
ted with the neuropathological findings obtained from the
postmortem studies. Correlation was analyzed and Pearson’s
correlation coefficient was computed by SPSS ver. 8.0.
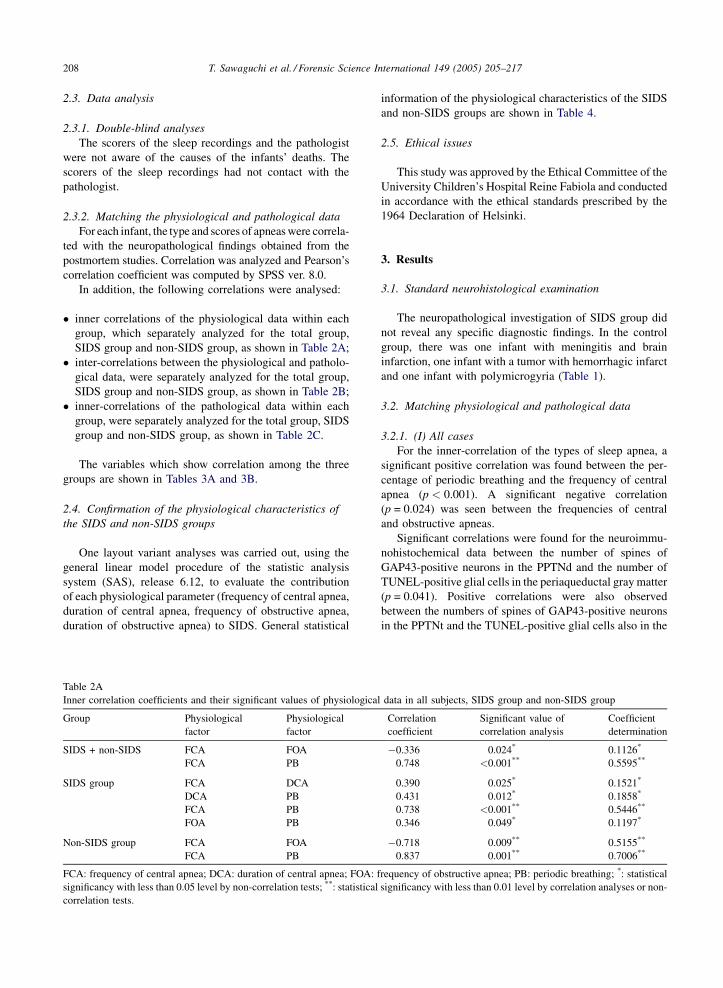
In addition, the following correlations were analysed:
� in
Ta
In
Gr
SI
SI
No
FC
sig
co
ner correlations of the physiological data within each
group, which separately analyzed for the total group,
SIDS group and non-SIDS group, as shown in Table 2A;
� in
ter-correlations between the physiological and patholo-gical data, were separately analyzed for the total group,
SIDS group and non-SIDS group, as shown in Table 2B;
� in
ner-correlations of the pathological data within eachgroup, were separately analyzed for the total group, SIDS
group and non-SIDS group, as shown in Table 2C.
The variables which show correlation among the three
groups are shown in Tables 3A and 3B.
2.4. Confirmation of the physiological characteristics of
the SIDS and non-SIDS groups
One layout variant analyses was carried out, using the
general linear model procedure of the statistic analysis
system (SAS), release 6.12, to evaluate the contribution
of each physiological parameter (frequency of central apnea,
duration of central apnea, frequency of obstructive apnea,
duration of obstructive apnea) to SIDS. General statistical
ble 2A
ner correlation coefficients and their significant values of physiological
oup Physiological
factor
Physiological
factor
DS + non-SIDS FCA FOA
FCA PB
DS group FCA DCA
DCA PB
FCA PB
FOA PB
n-SIDS group FCA FOA
FCA PB
A: frequency of central apnea; DCA: duration of central apnea; FOA: f
nificancy with less than 0.05 level by non-correlation tests; **: statistical
rrelation tests.
information of the physiological characteristics of the SIDS
and non-SIDS groups are shown in Table 4.
2.5. Ethical issues
This study was approved by the Ethical Committee of the
University Children’s Hospital Reine Fabiola and conducted
in accordance with the ethical standards prescribed by the
1964 Declaration of Helsinki.
3. Results
3.1. Standard neurohistological examination
The neuropathological investigation of SIDS group did
not reveal any specific diagnostic findings. In the control
group, there was one infant with meningitis and brain
infarction, one infant with a tumor with hemorrhagic infarct
and one infant with polymicrogyria (Table 1).
3.2. Matching physiological and pathological data
3.2.1. (I) All cases
For the inner-correlation of the types of sleep apnea, a
significant positive correlation was found between the per-
centage of periodic breathing and the frequency of central
apnea (p < 0.001). A significant negative correlation
(p = 0.024) was seen between the frequencies of central
and obstructive apneas.
Significant correlations were found for the neuroimmu-
nohistochemical data between the number of spines of
GAP43-positive neurons in the PPTNd and the number of
TUNEL-positive glial cells in the periaqueductal gray matter
(p = 0.041). Positive correlations were also observed
between the numbers of spines of GAP43-positive neurons
in the PPTNt and the TUNEL-positive glial cells also in the
data in all subjects, SIDS group and non-SIDS group
Correlation
coefficient
Significant value of
correlation analysis
Coefficient
determination
�0.336 0.024* 0.1126*
0.748 <0.001** 0.5595**
0.390 0.025* 0.1521*
0.431 0.012* 0.1858*
0.738 <0.001** 0.5446**
0.346 0.049* 0.1197*
�0.718 0.009** 0.5155**
0.837 0.001** 0.7006**
requency of obstructive apnea; PB: periodic breathing; *: statistical
significancy with less than 0.01 level by correlation analyses or non-
T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217 209
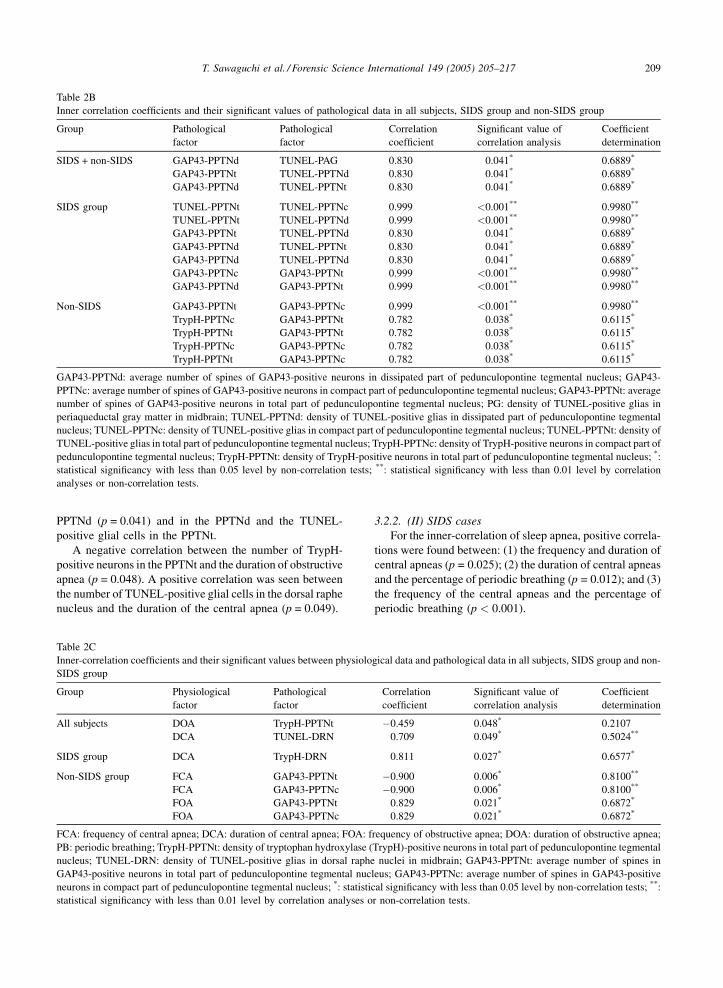
Table 2B
Inner correlation coefficients and their significant values of pathological data in all subjects, SIDS group and non-SIDS group
Group Pathological
factor
Pathological
factor
Correlation
coefficient
Significant value of
correlation analysis
Coefficient
determination
SIDS + non-SIDS GAP43-PPTNd TUNEL-PAG 0.830 0.041* 0.6889*
GAP43-PPTNt TUNEL-PPTNd 0.830 0.041* 0.6889*
GAP43-PPTNd TUNEL-PPTNt 0.830 0.041* 0.6889*
SIDS group TUNEL-PPTNt TUNEL-PPTNc 0.999 <0.001** 0.9980**
TUNEL-PPTNt TUNEL-PPTNd 0.999 <0.001** 0.9980**
GAP43-PPTNt TUNEL-PPTNd 0.830 0.041* 0.6889*
GAP43-PPTNd TUNEL-PPTNt 0.830 0.041* 0.6889*
GAP43-PPTNd TUNEL-PPTNd 0.830 0.041* 0.6889*
GAP43-PPTNc GAP43-PPTNt 0.999 <0.001** 0.9980**
GAP43-PPTNd GAP43-PPTNt 0.999 <0.001** 0.9980**
Non-SIDS GAP43-PPTNt GAP43-PPTNc 0.999 <0.001** 0.9980**
TrypH-PPTNc GAP43-PPTNt 0.782 0.038* 0.6115*
TrypH-PPTNt GAP43-PPTNt 0.782 0.038* 0.6115*
TrypH-PPTNc GAP43-PPTNc 0.782 0.038* 0.6115*
TrypH-PPTNt GAP43-PPTNc 0.782 0.038* 0.6115*
GAP43-PPTNd: average number of spines of GAP43-positive neurons in dissipated part of pedunculopontine tegmental nucleus; GAP43-
PPTNc: average number of spines of GAP43-positive neurons in compact part of pedunculopontine tegmental nucleus; GAP43-PPTNt: average
number of spines of GAP43-positive neurons in total part of pedunculopontine tegmental nucleus; PG: density of TUNEL-positive glias in
periaqueductal gray matter in midbrain; TUNEL-PPTNd: density of TUNEL-positive glias in dissipated part of pedunculopontine tegmental
nucleus; TUNEL-PPTNc: density of TUNEL-positive glias in compact part of pedunculopontine tegmental nucleus; TUNEL-PPTNt: density of
TUNEL-positive glias in total part of pedunculopontine tegmental nucleus; TrypH-PPTNc: density of TrypH-positive neurons in compact part of
pedunculopontine tegmental nucleus; TrypH-PPTNt: density of TrypH-positive neurons in total part of pedunculopontine tegmental nucleus; *:
statistical significancy with less than 0.05 level by non-correlation tests; **: statistical significancy with less than 0.01 level by correlation
analyses or non-correlation tests.
PPTNd (p = 0.041) and in the PPTNd and the TUNEL-
positive glial cells in the PPTNt.
A negative correlation between the number of TrypH-
positive neurons in the PPTNt and the duration of obstructive
apnea (p = 0.048). A positive correlation was seen between
the number of TUNEL-positive glial cells in the dorsal raphe
nucleus and the duration of the central apnea (p = 0.049).
Table 2C
Inner-correlation coefficients and their significant values between physiolo
SIDS group
Group Physiological
factor
Pathological
factor
All subjects DOA TrypH-PPTNt
DCA TUNEL-DRN
SIDS group DCA TrypH-DRN
Non-SIDS group FCA GAP43-PPTNt
FCA GAP43-PPTNc
FOA GAP43-PPTNt
FOA GAP43-PPTNc
FCA: frequency of central apnea; DCA: duration of central apnea; FOA: f
PB: periodic breathing; TrypH-PPTNt: density of tryptophan hydroxylase (
nucleus; TUNEL-DRN: density of TUNEL-positive glias in dorsal raphe
GAP43-positive neurons in total part of pedunculopontine tegmental nuc
neurons in compact part of pedunculopontine tegmental nucleus; *: statistic
statistical significancy with less than 0.01 level by correlation analyses o
3.2.2. (II) SIDS cases
For the inner-correlation of sleep apnea, positive correla-
tions were found between: (1) the frequency and duration of
central apneas (p = 0.025); (2) the duration of central apneas
and the percentage of periodic breathing (p = 0.012); and (3)
the frequency of the central apneas and the percentage of
periodic breathing (p < 0.001).
gical data and pathological data in all subjects, SIDS group and non-
Correlation
coefficient
Significant value of
correlation analysis
Coefficient
determination
�0.459 0.048* 0.2107
0.709 0.049* 0.5024**
0.811 0.027* 0.6577*
�0.900 0.006* 0.8100**
�0.900 0.006* 0.8100**
0.829 0.021* 0.6872*
0.829 0.021* 0.6872*
requency of obstructive apnea; DOA: duration of obstructive apnea;
TrypH)-positive neurons in total part of pedunculopontine tegmental
nuclei in midbrain; GAP43-PPTNt: average number of spines in
leus; GAP43-PPTNc: average number of spines in GAP43-positive
al significancy with less than 0.05 level by non-correlation tests; **:
r non-correlation tests.
T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217210
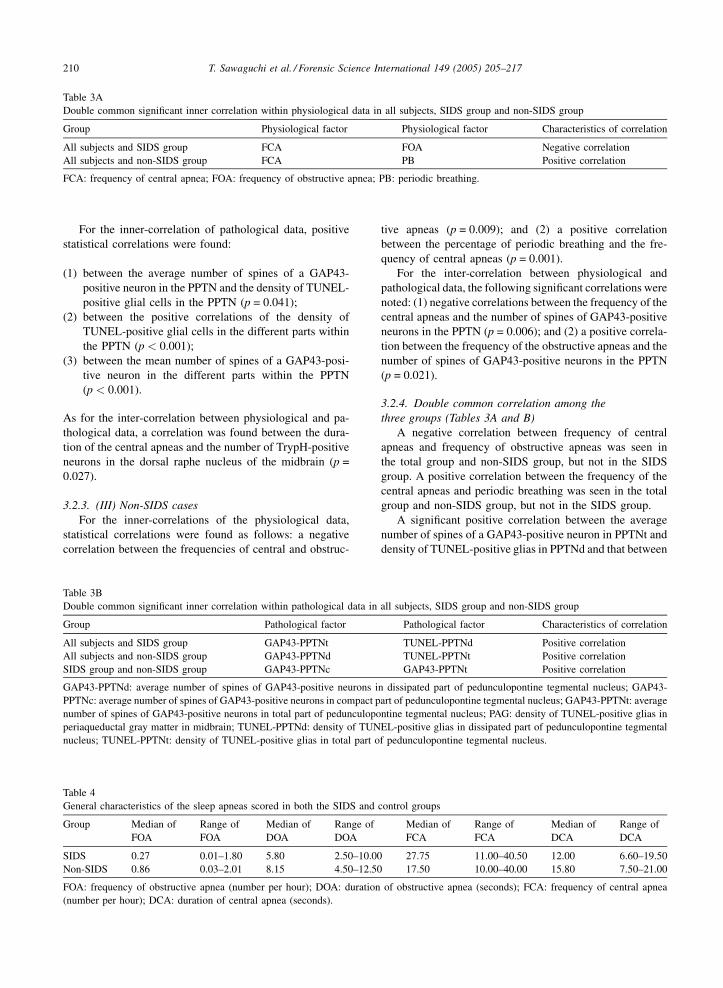
Table 3A
Double common significant inner correlation within physiological data in all subjects, SIDS group and non-SIDS group
Group Physiological factor Physiological factor Characteristics of correlation
All subjects and SIDS group FCA FOA Negative correlation
All subjects and non-SIDS group FCA PB Positive correlation
FCA: frequency of central apnea; FOA: frequency of obstructive apnea; PB: periodic breathing.
For the inner-correlation of pathological data, positive
statistical correlations were found:
(1) b
Tabl
Gen
Grou
SID
Non
FOA
(num
Tabl
Dou
Grou
All
All
SID
GAP
PPT
num
peria
nucl
etween the average number of spines of a GAP43-
positive neuron in the PPTN and the density of TUNEL-
positive glial cells in the PPTN (p = 0.041);
(2) b
etween the positive correlations of the density ofTUNEL-positive glial cells in the different parts within
the PPTN (p < 0.001);
(3) b
etween the mean number of spines of a GAP43-posi-tive neuron in the different parts within the PPTN
(p < 0.001).
As for the inter-correlation between physiological and pa-
thological data, a correlation was found between the dura-
tion of the central apneas and the number of TrypH-positive
neurons in the dorsal raphe nucleus of the midbrain (p =
0.027).
3.2.3. (III) Non-SIDS cases
For the inner-correlations of the physiological data,
statistical correlations were found as follows: a negative
correlation between the frequencies of central and obstruc-
e 4
eral characteristics of the sleep apneas scored in both the SIDS and
p Median of
FOA
Range of
FOA
Median of
DOA
Range of
DOA
S 0.27 0.01–1.80 5.80 2.50–10.00
-SIDS 0.86 0.03–2.01 8.15 4.50–12.50
: frequency of obstructive apnea (number per hour); DOA: duration
ber per hour); DCA: duration of central apnea (seconds).
e 3B
ble common significant inner correlation within pathological data in
p Pathological factor
subjects and SIDS group GAP43-PPTNt
subjects and non-SIDS group GAP43-PPTNd
S group and non-SIDS group GAP43-PPTNc
43-PPTNd: average number of spines of GAP43-positive neurons i
Nc: average number of spines of GAP43-positive neurons in compact p
ber of spines of GAP43-positive neurons in total part of pedunculopo
queductal gray matter in midbrain; TUNEL-PPTNd: density of TUN
eus; TUNEL-PPTNt: density of TUNEL-positive glias in total part o
tive apneas (p = 0.009); and (2) a positive correlation
between the percentage of periodic breathing and the fre-
quency of central apneas (p = 0.001).
For the inter-correlation between physiological and
pathological data, the following significant correlations were
noted: (1) negative correlations between the frequency of the
central apneas and the number of spines of GAP43-positive
neurons in the PPTN (p = 0.006); and (2) a positive correla-
tion between the frequency of the obstructive apneas and the
number of spines of GAP43-positive neurons in the PPTN
(p = 0.021).
3.2.4. Double common correlation among the
three groups (Tables 3A and B)
A negative correlation between frequency of central
apneas and frequency of obstructive apneas was seen in
the total group and non-SIDS group, but not in the SIDS
group. A positive correlation between the frequency of the
central apneas and periodic breathing was seen in the total
group and non-SIDS group, but not in the SIDS group.
A significant positive correlation between the average
number of spines of a GAP43-positive neuron in PPTNt and
density of TUNEL-positive glias in PPTNd and that between
control groups
Median of
FCA
Range of
FCA
Median of
DCA
Range of
DCA
27.75 11.00–40.50 12.00 6.60–19.50
17.50 10.00–40.00 15.80 7.50–21.00
of obstructive apnea (seconds); FCA: frequency of central apnea
all subjects, SIDS group and non-SIDS group
Pathological factor Characteristics of correlation
TUNEL-PPTNd Positive correlation
TUNEL-PPTNt Positive correlation
GAP43-PPTNt Positive correlation
n dissipated part of pedunculopontine tegmental nucleus; GAP43-
art of pedunculopontine tegmental nucleus; GAP43-PPTNt: average
ntine tegmental nucleus; PAG: density of TUNEL-positive glias in
EL-positive glias in dissipated part of pedunculopontine tegmental
f pedunculopontine tegmental nucleus.

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217 211
Fig. 1. TUNEL-positive glias in a SIDS case.
the mean number of spines of a GAP43-positive neuron in
PPTNd and the density of TUNEL-positive glias in PPTNt
were seen in the total group and SIDS group only, but not in
the non-SIDS group. A significant positive correlation
between the average number of spines of a GAP43-positive
neuron in the PPTNc and that in the PPTNt was seen only in
the SIDS and non-SIDS groups, but not in the total group.
3.2.5. (V) Single correlation in the SIDS groups
The single correlations seen only in the group of SIDS
victims were: (1) between the frequency and duration of the
central apneas (p = 0.025); (2) between the duration of the
central apneas and the percentage of periodic breathing
(p = 0.012); (3) between the frequency of the obstructive
apneas and the percentage of periodic breathing (p = 0.049);
(4) between the duration of the central apneas and the
number of TrypH-positive neurons in the dorsal raphe
nucleus (p = 0.027); (5) as an inter-correlation of
TUNEL-positive glial cells within the PPTN (p < 0.001);
(6) between the number of spines of GAP43-positive neu-
rons in the PPTNd or PPTNt and the number of TUNEL-
positive glial cells in the PPTNt or PPTNd (p = 0.041); and
(7) an inter-correlation between the number of spines of
GAP43-positive neurons in the PPTNd and those in the
PPTNt (p < 0.001).
3.3. Physiological characteristics of the SIDS and non-
SIDS groups
The physiological data of SIDS and non-SIDS groups are
shown in Table 4. The medians and range of frequency of the
obstructive apneas, the duration of the obstructive apneas
and that of central apneas in the non-SIDS group were
slightly larger than those in the SIDS group. In the non-
SIDS group the median and range of frequency of the central
apneas was slightly smaller than in the SIDS group.
The results of variant analyses of the 38 infants who died
showed the following significant associations with SIDS: the
frequency of the obstructive apneas (p = 0.001), duration of
the obstructive apneas (p = 0.026) and duration of the central
apneas (p = 0.049).
3.4. Pathological findings
The positivity of TUNEL and the immunohistochemical
positivities of GAP43 and TrypH in the brain of SIDS victim
are shown in Figs. 1–3. Glial apoptosis detected by the
TUNEL method in Fig. 1, GAP43 positive neuronal cells and
spines in Fig. 2 and TrypH positive neuronal cells in Fig. 3.
4. Discussion
Compared to the control subjects, the future SIDS vic-
tims were characterized by longer episodes of central apnea
as well as more frequent and longer obstructive sleep apneas.
Similar findings have been reported previously [40,41].
The observed correlation found between the pathology of
the brain stem and the apneas suggest that future SIDS
victims are characterized by changes in the arousal process
and breathing controls.
In the arousal pathway of the SIDS victims, a significant
correlation was seen between the central apnea character-
istics and TrypH-positive neurons in the DRN, as well as
between apoptosis and plasticity. Similar observations were
not found in the non-SIDS infants.
T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217212
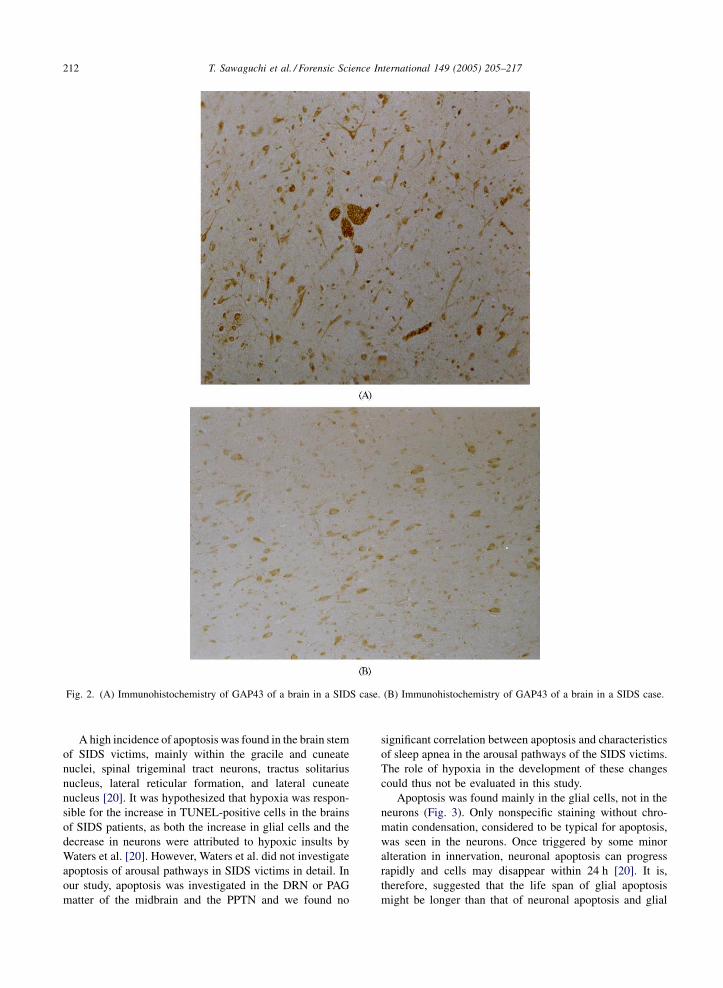
Fig. 2. (A) Immunohistochemistry of GAP43 of a brain in a SIDS case. (B) Immunohistochemistry of GAP43 of a brain in a SIDS case.
A high incidence of apoptosis was found in the brain stem
of SIDS victims, mainly within the gracile and cuneate
nuclei, spinal trigeminal tract neurons, tractus solitarius
nucleus, lateral reticular formation, and lateral cuneate
nucleus [20]. It was hypothesized that hypoxia was respon-
sible for the increase in TUNEL-positive cells in the brains
of SIDS patients, as both the increase in glial cells and the
decrease in neurons were attributed to hypoxic insults by
Waters et al. [20]. However, Waters et al. did not investigate
apoptosis of arousal pathways in SIDS victims in detail. In
our study, apoptosis was investigated in the DRN or PAG
matter of the midbrain and the PPTN and we found no
significant correlation between apoptosis and characteristics
of sleep apnea in the arousal pathways of the SIDS victims.
The role of hypoxia in the development of these changes
could thus not be evaluated in this study.
Apoptosis was found mainly in the glial cells, not in the
neurons (Fig. 3). Only nonspecific staining without chro-
matin condensation, considered to be typical for apoptosis,
was seen in the neurons. Once triggered by some minor
alteration in innervation, neuronal apoptosis can progress
rapidly and cells may disappear within 24 h [20]. It is,
therefore, suggested that the life span of glial apoptosis
might be longer than that of neuronal apoptosis and glial

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217 213
Fig. 3. Immunohistochemistry of Tryptophan-hydroxylase of a brain in a SIDS case.
apoptosis could be observed after the disappearance of
neuronal apoptosis.
Further investigations are required to determine whether
the apoptotic findings within the DRN result directly from
hypoxic exposure. As in most developing systems, an
excessive number of myelinating cells are generated during
myelination and redundant oligodendrocytes undergo a type
of degeneration known as programmed cell death or apop-
tosis [51]. Marked delay in myelination, i.e., hypomyelina-
tion, has been reported in victims of SIDS [52,53]. An
increase in the apoptosis of oligodendrocytes (oligodendro-
glias) may lead to hypomyelination in SIDS. The apoptosis
of glial cells observed in the present study could be asso-
ciated with these cellular changes.
The presence of gliosis [15,16,22–24] and apoptosis
[20,21] found in the brain stems of the SIDS victims has
been reported previously.
In a recent study in SIDS victims by Heyler et al. [50]
glial reaction in the thalamic and hypothalamic centers
associated with arousal was not observed.
Petito and Roberts speculated that apoptosis is a mean of
controlling the number of astrocytes during gliosis [28]. The
potential involvement of apoptosis in delayed neuronal death
following ischemia is likewise undetermined [54]. The
residual neurons from pontosubicular necrosis (PNS) caused
by hypoxic insults to the fetal or neonatal pontine nuclei and
subiculum differ from typical ischemic necrosis: they resem-
ble those involved in apoptosis [55]. Such findings can be
found in young children who succumbed suddenly [55].
In the present study, a significant correlation was found
between the duration of the central apnea and the number of
TUNEL-positive glial cells in the DRN. This correlation was
seen for the general population studied, but was not con-
firmed in the SIDS group, probably because of the small
number of subjects. Hypoxemic changes to the glial cells
(astrocytes) could be associated with the development of
prolonged obstructive sleep apneas [15,56].
Significant intercorrelations were measured between the
number of spines of GAP43-positive neurons in the PPTN
and the TUNEL-positive glial cells in the PPTN of SIDS
victims, as in all the others in the group. Inner-correlations
were found with apoptosis within the PPTN. This is intri-
guing because the PPTN have important roles not only for
the expression of elements in the REM sleep stage but also
for the arousal reaction [57]. Main projection neuron of the
PPTN is cholinergic and it is innervated by the aminergic
neurons. A malfunction of the PPTN could be caused by an
imbalance in the choline/monoamine system in the brain
stem [58,59]. The three diffuse projection systems arising
from the brain stem (the noradrenergic projection originat-
ing in the locus coeruleus, the serotonergic projection from
the DRN, and the cholinergic projection from neurons
gathering in the laterodorsal tegmental nucleus) are var-
iously related to the PPTN function as controllers of sleep
and wakefulness [60]. The PPTNc is considered to be the
area of strong cholinergic projection, and the PPTNc coin-
cides with the mesencephalic locomotor region [28]. The
projections from the medial preoptic nucleus and nucleus
accumbens to the mesencephalic locomotor region include
other neurotransmitters and the efferent projection from the
hippocampus is serotonergic [61–66]. TrypH-positive neu-
rons reflect the action of serotonergic neurons. In the SIDS
victims, TrypH-positive neuronal cells were found in the
PAG, DRN, superior central nucleus, nucleus raphe magnus,
and nucleus raphe obscurus. The percentage of TrypH-
positive cells was reduced in the caudal raphe in SIDS
[67]. Therefore, a reduction in the number of TrypH-positive
cells could be related that in the infants at risk for SIDS, and
a depression in serotonin synthesis could be associated with
compromised arousal responses.

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217214
In the SIDS infants, a significant positive correlation was
found between the duration of the central apneas and the
number of TrypH-positive neurons in the dorsal raphe
nucleus. The activity of TrypH was shown to increase
following a chronic exposure of hypoxia but not after an
acute exposure [28]. This observation could explain the
positive correlation between TrypH-positive neurons and
apnea in the SIDS group.
‘‘Plasticity of the brain’’ refers to functional changes
induced by external or internal factors, maintained even after
those factors disappear [68]. Plasticity of the brain is recog-
nized not only in neurons but also in glial cells. Neuronal
plasticity has been divided into two categories: plasticity of
synaptic transmission and plasticity of synaptic combina-
tions. The former represents functional changes, not neces-
sarily associated with morphometric changes. The latter
requires observable morphometric changes (e.g., the number
of synapses, the site of synapses, the number of dendritic
spines, the quality of receptors, and the number of recep-
tors). These morphometric changes occur not only in the
presynaptic but also in the postsynaptic sites, such as
dendritic spines [68].
GAP43 is a major protein of neuronal growth cones and
presynaptic terminals [69–73]. It is a candidate for involve-
ment in both axonal growth and synaptic plasticity. It has
been shown that in several neuronal systems, the GAP43
expression is greater in those neurons that are extending
axons, during the development or regeneration of injured
axons found in intact adult neurons. GAP43, best character-
ized as growth-associated proteins, can inhibit phosphati-
dylinositol phosphate kinase. It can be phosphorylated by
protein kinase C and affect neurotransmitter release.
SIDS might be caused by a specific neuronal changes that
could occur primarily before the apneic events results.
Because the correlation between the average number of
spines per a Gap-43 positive neuron in PPTNt or PPTNc
and the frequency of the central or obstructive apneas was
found in the non-SIDS group only. No correlation was found
between brainstem plasticity and sleep apnea characteristics
in SIDS victims.
In the present study, a positive correlation between the
number of TrypH-positive neurons and the number of spines
of GAP43-positive neurons both in the PPTNt was seen in
the non-SIDS group but not in the SIDS group. A significant
correlation was found for both the serotonergic neuron and
plasticity in the non-SIDS group, but not in the SIDS group.
This observation could suggest a possible neurotransmitter
imbalance in SIDS infants.
The so-called programmed cell death, which is often
synonymous with apoptosis, participates significantly to
brain plasticity, particularly in the process of growth and
development. Double mapping of apoptosis detected by the
in situ labeling method and the detection of immature
neurons by GAP43-positive staining, revealed that apoptotic
cell death occurs primarily among GAP43-positive neurons
in the olfactory epithelium [74]. A significant positive
correlation between the number of spines of GAP43-positive
neurons in the PPTNd and the number of TUNEL-positive
apoptotic glial cells in the PPTNd was confirmed in the SIDS
group only. The PPTN is an important part of the arousal
pathway. In this correlational relationship between glial
apoptosis and neuronal plasticity, the causal relationship
was not clear but in the background of this correlation some
process of reprogramming or remodeling might possibly
exist. Because of the lack of correlation between the hypoxic
events, sleep apneas and apoptosis, reprogramming or remo-
deling might be cause of glial apoptosis in the SIDS infants
in this study.
The correlation between plasticity and characteristics of
sleep apnea was found only in the non-SIDS group. Plas-
ticity is shown as the average number of spines per a GAP43
positive neuron in PPTNt or PPTNc. The frequency of the
central apnea correlated negatively with plasticity and fre-
quency of obstructive apnea, and corrleated positively with
plasticity. In other words, the average number of spines per a
GAP43 positive neuron in PPTNd correlated positively with
the density of TUNEL-positive glia in PPTNd in the SIDS
group only. This correlation suggests the reprogramming or
remodeling in PPTNd in SIDS. Therefore, the remodeling in
PPTN could positively promote the central apnea and inhibit
obstructive apnea.
The physiologic role of apoptosis has been invoked to
explain brain development [75]. Apoptosis may be a
mechanism for maintaining cell numbers at a critical set
point determined by a complex interaction of cellular and
extracellular factors, including growth factors and other
cellular biological factors [76]. Its role in reactive cellular
changes in the brain is still under investigation [76]. The final
correlation between apoptosis and plasticity in the arousal
pathway may suggest a possible reprogramming of neuronal
and glial cell death associated with an imbalance in neuro-
informational transfer through this pathway.
In this study, correlation between the duration of the
obstructive apnea and the pathological findings in PPTN,
and the correlation between the duration of the central apnea
and the pathological findings in DRN were found. An
association between the obstructive apnea and PPTN, and
that between the central apnea and DRN could be suggested.
Central apnea could be associated with the arousal pathway
within the DRN in the midbrain. PAG matter in the midbrain
can act not only as the superior center of the respiratory
nuclei in the medulla oblongata but also as part of the
defense areas in the visceral alerting response close to the
arousal reaction. DRN is in the same pathway as PAG matter.
Therefore, dual projection to respiration and arousal reaction
in the pathway including DRN and PAG matter could also be
associated with central apnea.
Such neuronal pathways with dual projection controlling
respiration and arousal reactions could be major contribution
in the pathophysiology of SIDS.
In conclusion, the correlation found between the brain
stem changes and apnea suggest the possibility of character-

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217 215
istic changes in the arousal process and apnea. Future studies
should be conducted to confirm the relationship between the
observed changes in the brainstem of SIDS victims, a
reduced propensity to be arouse from sleep and a frequency
and duration of sleep apneas.
Acknowledgments
This study was supported by Health Sciences Research
Grants for Research on Children and Families from the
Japanese Ministry of Health and Welfare, Okamoto-Itoe
Award and Satake Takako Award from Tokyo Women’s
Medical University. The authors sincerely thank Prof. C.
De Prez for the kind help extended to us at her laboratory and
Prof. H. Sasaki, Prof. H. Oda, Dr. M. Mizuguchi and Dr. S.
Otsuka for their advice in editing this manuscript.
References
[1] A. Steinschneider, Prolonged apnea and the sudden infant
death syndrome, clinical and laboratory observations, Pedia-
trics 50 (1972) 62–64.
[2] A. Steinschneider, S.L. Weinstein, E. Diamond, The sudden
infant death syndrome and apnea/obstruction during neonatal
sleep and feeding, Pediatrics 70 (1982) 858–863.
[3] D.H. Kelly, D.C. Shannon, K. O’Connell, Care of infants of
near-miss sudden infant death syndrome, Pediatrics 61 (1978)
511–514.
[4] K. Mcculloch, R.T. Brouillette, A.J. Guzzetta, Arousal
responses in near-miss sudden infants death syndrome and
normal infants, J. Pediatr. 101 (1982) 911–917.
[5] A. Van der Hal, A.M. Rodriguez, C.W. Sargent, Hypoxic and
hypercapneic arousal responses and prediction of subsequent
apnea of infancy, Pediatrics 75 (1985) 848–854.
[6] C. Hunt, Sudden infant death syndrome, in: R.C. Beckerman,
R.T. Brouillette, C.E. Hunt (Eds.), In Respiratory Control
Disorders in Infants and Children, Williams & Wilkins, Balti-
more, 1992, pp. 190–211.
[7] T. Hoppenbrouwers, J.E. Hodgman, L. Cabal, Obstructive
apnea, associated patterns of movement, heart rate, and oxy-
genation in infants at low and increased risk for SIDS, Pediatr.
Pulmonol. 15 (1993) 1–12.
[8] P. Franco, J. Groswasser, M. Sottiaux, E. Broadfield, A. Kahn,
Prone sleep and decreased cardiorespiratory response to audi-
tory stimulation in healthy infants, Pediatrics 97 (1996) 174–
178.
[9] K.A. Waters, A. Gonzalez, C. Jean, A. Morielli, R.T. Brouill-
ette, Face-straight down and face-near-straight-down positions
in healthy, prone-sleeping infants, J. Pediatr. 128 (1996) 616–
625.
[10] S. Takashima, T. Mito, L.E. Becker, Neuronal development in
the medullary reticular formation in sudden infant death
syndrome and premature infants, Neuropediatrics 16 (1985)
76–79.
[11] S. Takashima, L.E. Becker, Developmental abnormalities of
medullary ‘‘Respiratory Centers’’ in sudden infant death syn-
drome, Exp. Neurol. 90 (1985) 580–587.
[12] L.E. Becker, Neural maturation delay as a link in the chain of
events leading to SIDS, Can. J. Neurol. Sci. 17 (1990) 361–
371.
[13] S. Takashima, L.E. Becker, Delayed dendritic development of
catecholaminergic neurons in the ventrolateral medulla of
children who died of sudden infant death syndrome, Neuro-
pediatrics 22 (1991) 97–99.
[14] S. Takashima, D. Armstrong, L.E. Becker, J. Huber, Cerebral
white matter lesions in sudden infant death syndrome, Pedia-
trics 62 (1978) 155–159.
[15] S. Takashima, D. Armstrong, L.E. Becker, C. Bryan, Cerebral
hypoperfusion in the sudden infant death syndrome? Brain
stem gliosis and vasculature Ann Neurol. 4 (1978) 257–262.
[16] S. Takashima, Developmental changes of glial fibrillary acidic
protein and myelin basic protein in perinatal leukomalacia;
relationship to a predisposing factor, Brain Dev. 6 (1984)
444–450.
[17] R.L. Naeye, Hypoxemia and the sudden infant death syn-
drome, Science 186 (1974) 837–838.
[18] R.L. Naeye, Sudden infant death syndrome, Sci. Am. 242
(1980) 56–62.
[19] R.L. Naeye, J.M. Olsson, J.W. Combs, New brain stem and
bone marrow abnormalities in victims of the sudden infant
death syndrome, J. Perinatol. 9 (1989) 180–183.
[20] K.A. Waters, B. Meehan, J.Q. Huang Roy, A. Gravel, J.
Michaud, A. Cote, Neuronal apoptosis in sudden infant death
syndrome, Pediatr. Res. 45 (1999) 166–172.
[21] A. Cote, K. A. Waters, B. Meehan, Neuronal apoptosis in water-
shed areas in SIDS victims: evidence for hypotensive episodes
before death? Pediatric Res. 45 (5) (part 2 of 2) (1999) 27A.
[22] H.C. Kinney, P.C. Burger, F.E. Harrell, R.P. Hudson, Reactive
gliosis in the medulla oblongata of victims of the sudden infant
death syndrome, Pediatrics 72 (1983) 181–187.
[23] H.C. Kinney, J.J. Filiano, Brainstem research in sudden infant
death syndrome, J. Neuropathol. Exp. Neurol. 15 (1988) 240–
250.
[24] H.C. Kinney, J.J. Filiano, R.M. Harper, The neuropathology of
the sudden infant death syndrome: a review, J. Neuropathol.
Exp. Neurol. 51 (1992) 115–126.
[25] S. Takashima, T. Mito, H. Yamanouchi, Developmental brain-
stem pathology in sudden infant death syndrome, Acta Pae-
diatrica Japonica 36 (1994) 317–320.
[26] T.O. Rognum, O.D. Saugstad, S. Oyasaeter, B. Olaisen, Ele-
vated levels of hypoxanthine in sudden infant death syndrome,
Pediatrics 82 (1983) 615–618.
[27] I. Ferrer, A. Tortosa, A. Macaya, A. Sierra, D. Moreno, F.
Munell, R. Blanco, W. Squier, Evidence of nuclear DNA
fragmentation following hypoxia-ischemia in the infant rat
brain, and transient forebrain ischemia in the adult gerbil,
Brain Pathol. 4 (1994) 115–122.
[28] C.K. Petito, B. Roberts, Evidence of apoptotic cell death in
HIV encephalitis, Am. J. Pathol. 146 (1995) 1121–1130.
[29] S.H. Feinsilver, R. Wong, D.M. Raybin, Adaptions of neuro-
transmitter synthesis to chronic hypoxia in cell culture, Bio-
chimica et Biophisica Acta 928 (1987) 56–62.
[30] A. Vaccari, S. Brotman, J. Cimino, P.S. Timiras, Adaptive
changes induced by high altitude in the development of brain
monoamine enzymes, Neurochem. Res. 3 (1978) 295–311.
[31] T. Hedner, P. Lundborg, J. Engel, Effect of hypoxia on
monoamine synthesis in brains of developing rats. III. Various
O2 levels, Biol. Neonate 34 (1978) 55–60.

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217216
[32] D.B. Rye, Contributions of the pedunculopontine region to
normal and altered REM sleep, Sleep 20 (1997) 757–788.
[33] W.W. Blessing, The Lower Brainstem and Bodily Homeostasis,
Oxford University Press, New York & Oxford, 1997, 269-321.
[34] M.P. Mattson, W. Duan, ‘‘Apoptotic’’ biochemical cascades in
synaptic compartments: roles in adaptive plasticity and neu-
rodegenerative disorders, J. Neurosci. Res. 58 (1999) 152–166.
[35] A. Kirkwood, Serotonergic control of developmental plasti-
city, Proc. Nat. Acad. Sci. 97 (2000) 1951–1952.
[36] E.C. Azmitia, Serotonin neurons, neuroplasticity, and home-
ostasis of neural tissue, Neuropsychopharmacology 21 (2
Suppl.) (1999) 33–45.
[37] L. Kojic, Q. Gu, R.M. Douglas, M.S. Cynader, Serotonin
facilities synaptic plasticity in kitten visual cortex: an in vitro
study, Brain Res. 101 (1997) 299–304.
[38] R. Nieuwenhuys, J. Voogd, C.V. Huijen, The human central
nervous system synopsis and atlas, Springer-Verlag, Berlin,
1998, pp. 186–204, 302–303, 318.
[39] M. Willinger, S. James, C. Catz, Defining the sudden infant
death syndrome (SIDS), Pediatr. Pathol. 11 (1991) 677–
684.
[40] A. Kahn, J. Groswasser, E. Rebuffat, M. Sottiaux, D. Blum, M.
Foester, P. Franco, A. Bochner, M. Alexander, A. Bachy, P.
Richard, M. Verghote, D. LePolain, J.L. Wayenberg, Sleep and
cardioresoiratory characteristics of infant victims of sudden
death: a prospective case-control study, Sleep 15 (1992) 287–
292.
[41] A. Kahn, D. Blum, E. Rebuffat, M. Sottiaux, J. Levitt, A.
Bochner, M. Alexander, J. Grosswasser, M.F. Muller, Poly-
somnographic studies of infants who subsequently died of
sudden infant death syndrome, Pediatrics 82 (1988) 721–727.
[42] C. Guilleminault, M. Souquet, Sleep states and related pathol-
ogy, in: R. Korobkin, C. Guilleminault (Eds.), Advances in
Perinatal Neurology (1), Spectrum Publications, New York,
1979, pp. 225–247.
[43] C. Guilleminault, M. Souquet, R.L. Ariagno, R. Korobkin, F.B.
Simmons, Five cases of near-miss sudden infant death syn-
drome and development of obstructive apnea syndrome, Pedia-
trics 73 (1984) 71–78.
[44] D.H. Kelly, H. Golub, D. Carley, D.C. Shannon, Pneumograms
in infants who subsequently died of sudden infant death
syndrome, J. Pediatr. 109 (1986) 249–254.
[45] D.H. Kelly, D.C. Shannon, Periodic breathing in infants with
near-miss sudden infant death syndrome, Pediatrics 63 (1979)
355–360.
[46] Y. Gavrieli, Y. Sherman, S.A. Ben-Sasson, Identification of
programmed cell death in situ specific labeling of nuclear DNA
fragmentation, J. Cell. Biol. 119 (1992) 493–501.
[47] T. Sawaguchi, B. Jasani, M. Kobayashi, B. Knight, Post-
mortem analysis of apoptotic changes associated with human
skin bruises 108 (2000) 187–203.
[48] T. Sawaguchi, B. Jasani, B. Knight, Identification of apoptotic
cell nuclei in postmortem skin biopsies using the TUNEL
reaction: effect of postmortem time interval, Acta Crim. Japan
63 (1997) 39–43.
[49] S.V. Pravdenkova, A.G. Basnakian, S.J. James, B.J. Anderson,
DNA fragmentation and nuclear endonuclease activity in rat
brain after severe closed head injury, Brain Res. 729 (1996)
151–155.
[50] G. Heyler, G. Stoltenburg-Didinger, F. Landeghem, T. Baja-
nowski, Histopathology of thalamic and hypothalamic centers
of arousal in sudden infant death (SID) victims, Pediatric Res.
45 (5) (part 2 of 2) (1999) 21A.
[51] J. Brockhaus, S. Ilschner, R.B. Banati, H. Kettermann, Mem-
brane properties of ameboid microglial cells in the corpus
callosum slice from early postnatal mice, J. Neurosci. 16
(1993) 4412–4421.
[52] J.C. Kinney, B.A. Brody, D.M. Finkelstein, G.F. Vawter, F.
Mandell, F.H. Giles, Delayed central nervous system myelina-
tion in the sudden infant death syndrome, J. Neuropathol. Exp.
Neurol. 50 (1991) 29–48.
[53] R.L. Naeye, J.M. Olsson, J.W. Combs, Bew brain stem and
bone marrow abnormalities in victims of sudden infant death
syndrome, J. Perinatol. 9 (1989) 180–183.
[54] N. Kawai, Neuronal death and survival in the brain, Sci. Living
Body 47 (1996) 33–38.
[55] T. Mito, S. Takashima, E. Ohama, Brainstem pathology in the
severe motor and intellectual disabilities patients with sudden
death, Annual Report of Study of situation, long term con-
valescence and functional improvement of severe psycho-
somatic disorder by Ministry of Health and Welfare, 1997,
pp. 33–37.
[56] L.E. Becker, S. Takashima, Chronic hypoventilation and
development of brain stem gliosis, Neuropediatrics 16
(1985) 19–23.
[57] D.B. Rey, Contribution of the pedunculopontine region to
normal and altered REM sleep, Sleep 20 (1997) 757–788.
[58] J. Kohyama, M. Shimohira, Y. Iwakawa, Maturation of moti-
lity and motor inhibition in rapid-eye-movement sleep, J.
Pediatr. 130 (1997) 117–122.
[59] J. Kohyama, Sleep as a window on the developing brain, Curr.
Prob. Pediatr. 28 (1998) 73–92.
[60] Y. Kayama, Y. Koyama, Brainstem neural mechanisms of sleep
and wakefulness, Eur. Urol. 33 (Suppl. 3) (1998) 12–15.
[61] A.J.J. Pijnenburg, W.M.M. Honig, J.A.M. Van der Heyden,
J.M. Van Rossum, Effects of chemical stimulation of the
mesolimbic dopamine system upon locomotor activity, Eur.
J. Pharmacol. 35 (1976) 45–58.
[62] J. Ciriello, M.M. Caverson, Direct pathway from neurons in
the ventrolateral medulla relaying cardiovascular afferent
information to the supraoptic nucleus in cat, Brain Res. 292
(1984) 221–228.
[63] G.J. Mogenson, Limbic-motor integration with emphasis on
initiation of exploratory and goal-directed locomotion, in: R.
Bandler (Ed.), Modulation of Sensorimotor Activity During
Alterations in Behavioral States, Liss, New York, 1984, pp.
121–137.
[64] G.J. Mogenson, L.W. Swanson, M. Wu, Evidence that projec-
tions from substantia innominata to zona incerta mesencepha-
lic locomotor region contribute to locomotor activity, Brain
Res. 334 (1984) 65–76.
[65] L.W. Swanson, G.J. Mogenson, C.R. Gerfen, P. Robinson,
Evidence for a projection from the lateral preoptic area and
substantia innominata to the Mesencephalic Locomotor
Region’ in the rat, Brain Res. 295 (1984) 161–178.
[66] L.W. Swanson, G.J. Mogenson, R.B. Simerly, M. Wu, Anato-
mical and electrophysiological evidence for a projection from
the medial preoptic area to the ‘mesencephalic and subthala-
mic locomotor regions’ in the rat, Brain Res. 405 (1987) 108–
122.
[67] H.C. Kinney, J.J. Filiano, W.F. White, Medullary serotonergic
network deficiency in the sudden infant death syndrome:

T. Sawaguchi et al. / Forensic Science International 149 (2005) 205–217 217
review of a 15-year study of a single dataset, J. Neuropathol.
Exp. Neurol. 60 (2001) 228–247.
[68] J. Senba, Brain and homeostasis, Hoso Daigaku Kyoikush-
inkokai, Tokyo, 1998, pp. 22–23, 79–94, 192–193.
[69] R.L. Neve, E.A. Finch, E.D. Bird, L.I. Benowitz, Growth-
associated protein GAP43 is expressed selectively in associa-
tive regions of the adult human brain, Proc. Natl. Acad. Sci. 85
(1998) 3638–3642.
[70] S.M. De la Monte, H.J. Federoff, S.C. Ng, E. Grabczyk, M.C.
Fishman, GAP-43 gene expression during development: Per-
sistence in a distinctive set of neurons in the mature central
nervous system, Dev. Brain Res. 46 (1989) 161–168.
[71] J.H.P. Skene, Axonal growth associated proteins, Annu. Rev.
Neurosci. 12 (1989) 127–156.
[72] L.I. Benowitz, N.I. Perrone-Bizzozero, The relationship of
GAP43 to the development and plasticity of synaptic connec-
tions, Ann. N.Y. Acad. Sci. 627 (1991) 58–74.
[73] K.H. Pfenninger, B.A. De la Houssaye, S.M. Helmke, S.
Quiroga, Growth-regulated proteins and neuronal plasticity,
Mol. Neurobiol. 5 (1991) 143–151.
[74] T.J. Mahalik, Apparent apoptotic cell death in the olfactory
epithelium of adult rodents: death occurs at different devel-
opmental stages, J. Comp. Neurol. 372 (1996) 457–464.
[75] M.C. Raff, B.A. Barres, J.F. Burne, H.S. Coles, Y. Ishizaki, M.D.
Jacobson, Programmed cell death and the control of cell survival:
lessons from the nervous system, Science 262 (1993) 695–700.
[76] D.W. Dickson, Apoptosis in the brain, Am. J. Pathol. 146
(1995) 1040–1044.