antitumor immune response by cx3cl1 fractalkine gene transfer depends on both nk and t?cells
TRANSCRIPT

Antitumor immune response by CX3CL1 fractalkinegene transfer depends on both NK and T cells
Hong Xin1,2, Toshiaki Kikuchi1, Sita Andarini1, Shinya Ohkouchi1,Takuji Suzuki1, Toshihiro Nukiwa1, Huqun3, Koichi Hagiwara3,Tasuku Honjo4 and Yasuo Saijo1,2
1 Department of Respiratory Oncology and Molecular Medicine, Institute ofDevelopment, Aging, and Cancer, Tohoku University, Sendai, Japan
2 Department of Molecular Medicine, Tohoku University Graduate School of Medicine,Sendai, Japan
3 Department of Respiratory Medicine, Saitama Medical School, Saitama, Japan4 Department of Medical Chemistry, Kyoto University Graduate School of Medicine,Kyoto, Japan
The CX3C chemokine fractalkine (CX3CL1) exists as both a membrane-bound formpromoting firm cell-cell adhesion and a soluble form chemoattracting leukocytesexpressing its receptor CX3CR1. When adenoviral vector expressing mouse fractalkine(AdFKN) was transduced to the tumor cells, fractalkine was expressed as bothmembrane-bound form on the tumor cells and soluble form in the supernatant in vitro.Intratumoral injection of AdFKN (1�109PFU/tumor) into C26 and B16F10 tumorsresulted in marked reduction of tumor growth compared to control (C26: 86.5%,p<0.001; B16F10: 85.5%, p<0.001). Histological examination of tumor tissues revealedabundant infiltration of NK cells, dendritic cells, and CD8+ T lymphocytes 3 and/or6 days after treatment with AdFKN. Splenocytes frommice treated by AdFKN developedtumor-specific cytotoxic T cells, and thereby protected from rechallenging with parentaltumor cells. Antitumor effects by AdFKN were completely abrogated in both NK cell-depleted mice and CD8–/– mice, and partially blocked in CD4–/– mice. These dataindicated that fractalkine mediates antitumor effects by both NK cell-dependent andT cell-dependent mechanisms. This study suggests that fractalkine can be a suitablecandidate for immunogene therapy of cancer because fractalkine induces both innateand adaptive immunity.
Introduction
An idea that insufficient immune response enablestumor development to escape from the immune systemled to consideration of various types of immunotherapy
for human tumors [1–3]. Although immunotherapy hasbeen successfully used for treating tumors in animalmodel, only few types of tumors such as melanoma andrenal cell carcinoma responded to immunotherapy usingIL-2 and IFN-a [4–6]. Systemic administration ofcytokines often causes severe adverse effect at higherdoses that can induce efficient antitumor effects inseveral human tumors [7, 8].
An alternative strategy for stimulation of antitumorimmune response has been in vivo gene therapy, becausegene therapy can enable expression of the immunosti-mulatory molecules in only the tumor milieu, andtherefore can avoid systemic adverse effects [9].
Correspondence: Yasuo Saijo, Department of MolecularMedicine, Tohoku University Graduate School of Medicine,21 Seiryomachi Aobaku, Sendai 980-8575, JapanFax: +81-22-717-8230e-mail: [email protected]
Received 25/1/05Accepted 18/2/05
[DOI 10.1002/eji.200526042]
Key words:Chemokine
� Fractalkine � Genetherapy � NK cells
� T cells
Abbreviations: MOI: Multiplicity of infection �AdFKN: Adenoviral vector expressing mouse fractalkine
Eur. J. Immunol. 2005. 35: 1371–1380 Cellular immune response 1371
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Different cytokine genes have been employed forexperimental cancer immunogene therapy becausethese cytokines can stimulate proliferation of leukocytesand enhance the cytotoxic activity of effector cellsincluding NK cells and T lymphocytes [10–12].
Chemokines comprise families of molecules capableof chemoattracting various cell types to certain physio-logical and pathological sites, and also in tumor tissues[13]. Some chemokines induce angiogenesis and thuspromote tumor growth and metastasis [14, 15].However, tumor suppression by chemokine genetransfer into tumor cells has been demonstrated inanimal models [16–18]. Intratumoral injection ofadenoviral vector encoding macrophage-derived che-mokine (MDC, CCL22) resulted in marked tumorregression of Lewis lung carcinoma through strongchemoattracting activity of dendritic cells (DC), NKcells, and T lymphocytes [16]. Gene transfer of anotherCC chemokine, RANTES (CCL5) [17], or of theC chemokine lymphotactin (XCL1) [18] also exhibitedstrong antitumor activity in solid tumor models. Theseresults suggested that chemokines could mediate innateand adaptive antitumor immune response by recruitingmonocytes, DC, NK, and T lymphocytes into tumor sites.
To date more than 40 chemokines have beenidentified and classified on the spacing of N-terminalcysteine residues into four groups: C, CC, CXC, andCX3C. Fractalkine (CX3CL1) is the only member of theCX3C chemokine group and is structurally distinct fromother types of chemokines [19, 20]. Fractalkine exists asa membrane-bound molecule with the chemokinedomain at the end of a mucin-like stalk, and is releasedin soluble form through cleavage at a membrane-proximal site by the disintegrin-like metalloproteinase(ADM10) and tumor necrosis factor-a-converting en-zyme (ADAM17) [21, 22].
A soluble form of CX3CL1 induces migration of cellsexpressing its receptor CX3CR1, similar to other solublechemokines [23]. The interaction between fractalkineand its receptor CX3CR1 has a low dissociation rateresulting in very strong interactions between fractalkineand CX3CR1-expressing cells [24]. Thus, a membrane-anchored form of CX3CL1 mediates firm adhesionbetween CX3CL1-expressing cells and CX3CR1-expres-sing leukocytes. CX3CR1 is expressed on peripheralblood cytotoxic effector lymphocytes commonly armedwith intracellular perforin and granzyme B, whichinclude NK cells, cd T cells, terminally differentiatedCD8+ T cells, and a minor fraction of CD4+ cells inhumans [25]. On the other hand, CX3CR1 is expressedon monocytes, NK cells, and DC, but not on T cells inmice [26]. Therefore, it is likely that gene transfer ofCX3CL1 might induce strong antitumor activity throughrecruiting effector cells into tumor tissues and inducingfirm adhesion between tumor cells and effector cells.
In this study, we constructed an adenoviral vectorexpressing mouse fractalkine gene (AdFKN) anddemonstrated that fractalkine is expressed as bothmembrane-bound form and soluble form. Intratumoralinjection of AdFKN into B16F10 or C26 tumors resultedin striking inhibition of tumor growth in vivo. Injectionof AdFKN recruited numerous leukocytes into tumortissues, and led to strong antitumor responses by bothNK cells and T lymphocytes.
Results
Fractalkine expression in AdFKN-transducedtumor cells and cell growth
The expression of fractalkine mediated by the AdFKNwas confirmed in vitro by RT-PCR (Fig. 1A). RT-PCRanalysis revealed that 1,114-bp fragments correspondingto the fractalkine transcripts were amplified with thetotal cellular RNA from AdFKN-transduced B16F10 cells,but were not amplified with that from AdNull-trans-duced or nontransduced B16F10 cells (Fig. 1A). Expres-sion of b-actin mRNA was monitored as an internalcontrol in all B16F10 cells demonstrating the intactnessof the RNA. B16F10 cells infected with AdFKN revealedfractalkine protein expression in vitro by Western blotanalysis (Fig. 1B). Although 100-kDa fractalkine wasdetected in AdFKN-transduced B16F10 cells, recombi-nant fractalkine yielded an approximately 90-kDaprotein band because this recombinant protein consistsonly of the extracellular domain of fractalkine. Asexpected, there were no bands in control AdNull-transduced B16F10 as well as in nontransduced B16F10cells.
To assess surface expression of fractalkine, we usedflow cytometry to analyze the expression of membrane-bound fractalkine in AdFKN-transduced B16F10 cells(Fig. 1C). AdFKN-transduced B16F10 cells expressedfractalkine protein on the cell surface, whereas AdNull-transduced B16F10 or nontransduced B16F10 cells didnot at all. Expression of fractalkine transcripts, fractalk-ine protein, and membrane-bound fractalkine inAdFKN-transduced C26 cells was also confirmed byeach method (data not shown).
Neither AdFKN-transduced C26 nor B16F10 cells at amultiplicity of infection (MOI) of 100 altered cellgrowth compared to AdNull-transduced C26 orB16F10 cells, as detected by the cell viability assay(Alamar Blue; Biosource International, Camarillo, CA;data not shown).
Hong Xin et al. Eur. J. Immunol. 2005. 35: 1371–13801372
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
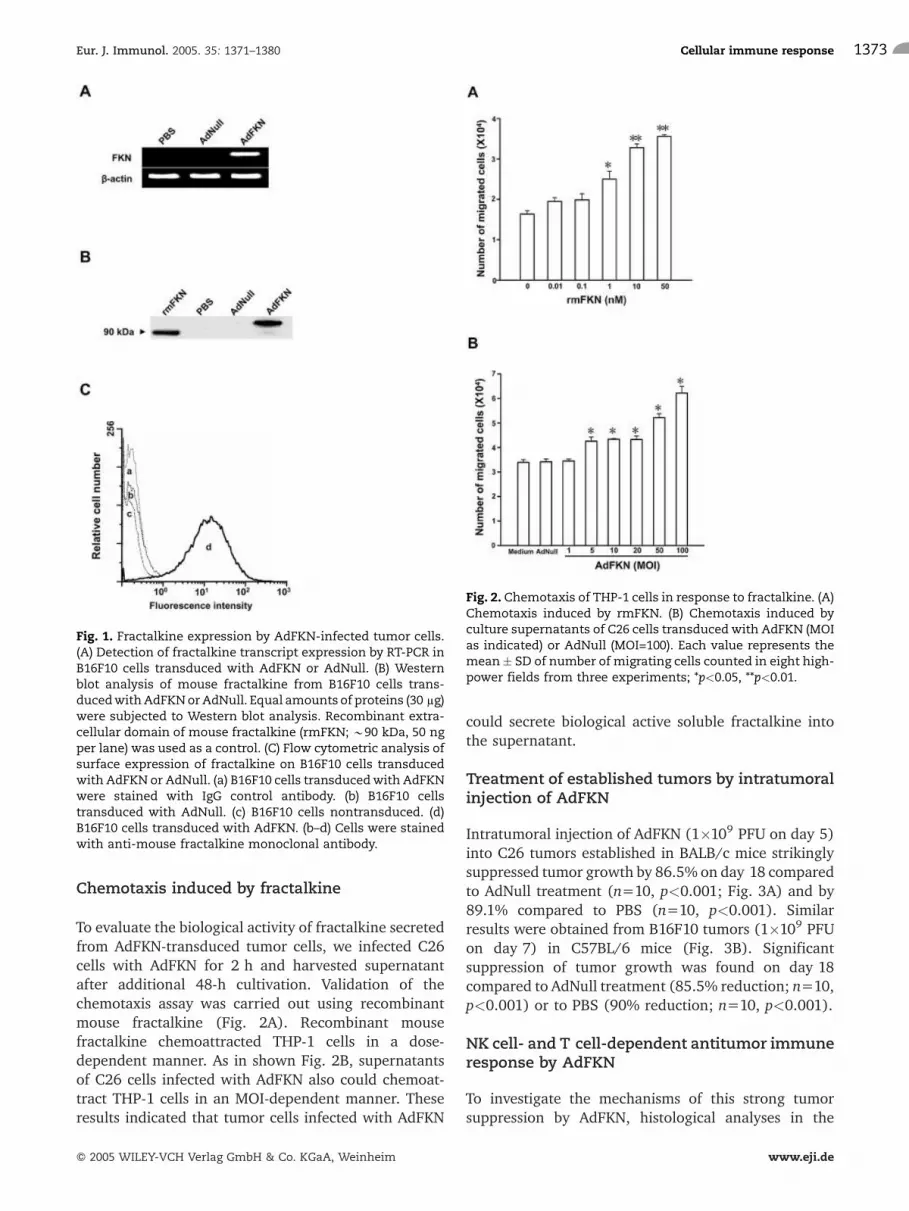
Chemotaxis induced by fractalkine
To evaluate the biological activity of fractalkine secretedfrom AdFKN-transduced tumor cells, we infected C26cells with AdFKN for 2 h and harvested supernatantafter additional 48-h cultivation. Validation of thechemotaxis assay was carried out using recombinantmouse fractalkine (Fig. 2A). Recombinant mousefractalkine chemoattracted THP-1 cells in a dose-dependent manner. As in shown Fig. 2B, supernatantsof C26 cells infected with AdFKN also could chemoat-tract THP-1 cells in an MOI-dependent manner. Theseresults indicated that tumor cells infected with AdFKN
could secrete biological active soluble fractalkine intothe supernatant.
Treatment of established tumors by intratumoralinjection of AdFKN
Intratumoral injection of AdFKN (1�109 PFU on day 5)into C26 tumors established in BALB/c mice strikinglysuppressed tumor growth by 86.5% on day 18 comparedto AdNull treatment (n=10, p<0.001; Fig. 3A) and by89.1% compared to PBS (n=10, p<0.001). Similarresults were obtained from B16F10 tumors (1�109 PFUon day 7) in C57BL/6 mice (Fig. 3B). Significantsuppression of tumor growth was found on day 18compared to AdNull treatment (85.5% reduction; n=10,p<0.001) or to PBS (90% reduction; n=10, p<0.001).
NK cell- and T cell-dependent antitumor immuneresponse by AdFKN
To investigate the mechanisms of this strong tumorsuppression by AdFKN, histological analyses in the
Fig. 2. Chemotaxis of THP-1 cells in response to fractalkine. (A)Chemotaxis induced by rmFKN. (B) Chemotaxis induced byculture supernatants of C26 cells transducedwith AdFKN (MOIas indicated) or AdNull (MOI=100). Each value represents themean� SD of number of migrating cells counted in eight high-power fields from three experiments; *p<0.05, **p<0.01.
Fig. 1. Fractalkine expression by AdFKN-infected tumor cells.(A) Detection of fractalkine transcript expression by RT-PCR inB16F10 cells transduced with AdFKN or AdNull. (B) Westernblot analysis of mouse fractalkine from B16F10 cells trans-ducedwithAdFKNorAdNull. Equal amounts of proteins (30 lg)were subjected to Western blot analysis. Recombinant extra-cellular domain of mouse fractalkine (rmFKN; *90 kDa, 50 ngper lane) was used as a control. (C) Flow cytometric analysis ofsurface expression of fractalkine on B16F10 cells transducedwith AdFKN or AdNull. (a) B16F10 cells transducedwith AdFKNwere stained with IgG control antibody. (b) B16F10 cellstransduced with AdNull. (c) B16F10 cells nontransduced. (d)B16F10 cells transduced with AdFKN. (b–d) Cells were stainedwith anti-mouse fractalkine monoclonal antibody.
Eur. J. Immunol. 2005. 35: 1371–1380 Cellular immune response 1373
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

tumors were performed on day 3 and 6 after intratu-moral injection of adenoviral vectors. Tumor necrosiswas observed in AdFKN-treated C26 tumors by hema-toxylin/eosin (H/E) staining on both day 3 and 6(Fig. 4A). On day 3, AdFKN-treated C26 tumors showedmarked infiltrations of NK cells and a slight increase ofCD4+ T lymphocytes, but no change of CD8+
T lymphocytes compared with AdNull-treated or PBS-treated tumors (Fig. 4A, B). Quantification of theseleukocytes revealed 4.7-fold infiltration of NK cells(p<0.001) and 1.4-fold increase of CD4+ T lymphocytes(p<0.01) compared to AdNull-treated tumors. Incontrast, tumor tissues on day 6 showed predominantinfiltration of CD8+ T lymphocytes (5.3-fold increasecompared to AdNull treatment, p<0.001). However, no
statistically significant increase of NK or CD4+
T lymphocytes was observed on day 6 (Fig. 4A, B).Increased infiltrations of CD11c into AdFKN-treated
tumors were observed on both day 3 (2.0-fold,p<0.001) and day 6 (3.1-fold, p<0.001) compared toAdNull (Fig. 4A, B). Similar results were obtained inB16F10 tumors (data not shown). These immuneeffector cells were observed not only in the peripheralarea but also in the central area of the tumor.
To functionally delineate the role of T cells in theantitumor immune responses by intratumoral injectionof AdFKN, B16F10 tumors in either CD4–/– or CD8–/–
mice were treated with AdFKN. As shown in Fig. 5A,antitumor responses in CD8–/– mice completely dis-appeared, whereas those in CD4–/– mice were partiallydiminished. In the control study, treatment by PBS didnot change the tumor growth in CD4–/– and CD8–/– mice(data not shown). Thus, the antitumor effects byfractalkine were predominantly dependent on CD8+
T cells and partially on CD4+ T cells. Using NK cell-depleted mice, we next investigated whether NK cellswere involved in antitumor immune response by AdFKN,because NK cells express the fractalkine receptorCX3CR1. As shown in Fig. 5B, while control IgGtreatment did not affect antitumor effects of AdFKN atall, antitumor response by AdFKN was completelyabolished by NK cell depletion. Taken together, bothNK cells and T cells play a key role for antitumor effectsof AdFKN.
Tumor-specific cytotoxic T cells induced byintratumoral injection of AdFKN
C26 tumors in BALB/c mice were treated by intratumor-al injection with AdFKN on day 5. Effector cells weregenerated from splenocytes obtained 9 days after theAdFKN treatment by co-culture with mitomycin C-treated C26 cells, and then assayed for cytotoxicityagainst C26 cells or BALB/c 3T3 cells as a control(Fig. 6A). Lymphocytes from mice treated with AdFKNexhibited significant cytotoxicity against the parentalC26 tumor cells, but no cytotoxicity was induced byAdNull or PBS (Fig. 6A). These CTL did not lyse againstirrelevant but syngeneic BALB/c 3T3 fibroblast cells,indicating that these CTL were tumor-specific. The sameresults were obtained with B16F10 cells (data notshown). In addition, splenocytes from mice treated withAdFKN exhibited significant enhancement of IFN-cproduction (469.29�2.62 pg/106 cells) compared withthose treated with AdNull (159.28�4.71 pg/106 cells)or with PBS (153.85�4.13 pg/106 cells) in co-culturewith C26 cells (n=5, p<0.001; Fig. 6B).
Fig. 3. Growth suppression of preexisting tumors by intratu-moral administration of AdFKN. (A) C26 cells (5�105) weresubcutaneously injected into BALB/c mice in the right flank(day 0). On day 5, established C26 tumors were treated byintratumoral administration of AdFKN (1�109 PFU/mouse, !),AdNull (1�109 PFU/mouse, *), and PBS (50 ll/mouse, *). (B)B16F10 cells (3�105) were subcutaneously injected into C57BL/6mice. Tumors were treated on day 7 as in (A). The datarepresent the means � SE of results from ten mice.
Hong Xin et al. Eur. J. Immunol. 2005. 35: 1371–13801374
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Suppression of and protection from of tumorgrowth by immunizationwith AdFKN-transducedtumor cells
Immunization with irradiated AdFKN-transduced C26cells protected four out of six mice from a rechallengingwith parental C26 cells (Fig. 7A). Although only threeout of six mice immunized with irradiated control C26cells or irradiated AdNull-transduced C26 cells devel-oped tumors, suppression of tumor growth on day 20was found compared to irradiated AdNull-C26 cell orC26 cell immunization (72.0% reduction, p<0.05 and64.3% reduction, p=0.06, respectively). Immunizationwith AdFKN-transduced B16F10 cells also protected twoout of six mice. In contrast, no mice immunized byirradiated B16F10 cells or irradiated AdNull-transducedB16F10 cells were tumor-free. Immunization withAdFKN-B16F10 cells resulted in significant slower tumorgrowth compared with control (70.0% reduction inAdNull-B16F10 and 67.1% in PBS-B16F10 cells, p<0.05;Fig. 7B).
CTL induction by immunization of irradiatedAdFKN-transduced tumors
Since the data have shown reduced tumorigenicity ordelayed progression of tumor cells after immunizationwith irradiated AdFKN-transduced C26 cells, we furtherinvestigated cytotoxic activity against C26 cells inimmunized mice. Splenocytes harvested from miceimmunized with irradiated AdFKN-transduced C26 cellsshowed significant lytic activity against C26 cells(p<0.001; Fig. 8A). In contrast, neither AdNull norPBS immunization evoked significant lytic activity(Fig. 8A). Higher IFN-c production was also observedin splenocytes harvested from mice immunized withAdFKN-transduced C26 cells (385.16�15.88 pg/106 cells) compared with AdNull (147.05�0.79 pg/106 cells) and with PBS (153.26�4.3 pg/106 cells,p<0.001; Fig. 8B).
Fig. 4. Infiltration of NK cells, DC, CD4+, and CD8+
lymphocytes into C26 tumors injected withAdFKN. C26 tumors established in BALB/c micewere treated by intratumoral injection of1�109 PFU of AdFKN, AdNull, or PBS alone onday 0. Tumors were removed on day 3 or 6 aftertreatment, and were stained using anti-asialoGM1, anti-CD4, anti-CD8, and anti-CD11c anti-body. (A) Histology of the C26 tumor stainedwithH/E, anti-asialo GM1, anti-CD4, anti-CD8, or anti-CD11c antibody on days 3 and 6 after treatment(magnification 400�). (B) Quantification of thenumbers of NK, CD4+, CD8+, or CD11c+ cells perten random high-power fields (400�) in tumortissues. The data represent the means � SD.Black bar: AdFKN treatment; gray bar: AdNulltreatment; white bar: PBS treatment.
Eur. J. Immunol. 2005. 35: 1371–1380 Cellular immune response 1375
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

Discussion
In this study, we demonstrated strong tumor growthinhibition by intratumoral injection of AdFKN in C26-and B16F10-bearing mice. Immunization by fractalkine-transduced tumor cells induced specific CTL thatspecifically killed target tumor cells directly or bysecreting the IFN-c and thus protected from rechallen-ging parental tumors. Along with immunohistochemicalanalysis of the tumor tissue and depletion of CD4, CD8,
or NK cells in mice, we conclude that the strong tumor-suppressive effects by fractalkine depend on both NKcells and T cells.
A chemokine family is another main regulator ofimmune effector cells as well as a number of cytokines[20, 27, 28]. Chemokines, secreted proteins of lowmolecular weight, provide important signals for migra-tion of leukocytes into pathological tissues. Therefore,chemokines can be suitable candidates for immunogenetherapy of cancers. In fact, several chemokine-basedimmunogene therapies demonstrated strong antitumoreffects in preclinical models [16–18]. Fractalkine, theonly member of the CX3C chemokine group, inducesmigration of leukocytes out of vessels into pathological
Fig. 5. T cell- and NK cell-dependent antitumor immuneresponse by AdFKN. (A) CD4–/–, CD8–/–, or wild-type C57BL/6mice transplanted subcutaneously with B16F10 cells (3�105/mouse)were treatedwith AdFKN (1�109 PFU/mouse).!: Wild-type mice treated with AdFKN;!: wild-type mice treated withPBS; *: CD4–/– mice treated with AdFKN; *: CD8–/– micetreated with AdFKN. (B) C57BL/6 mice received depletinganti-asialo GM1 or control antibodies. B16F10 tumors in NKcell-depleted mice were treated with AdFKN. !: Controlantibody-treated mice treated with AdFKN; !: wild-type micetreated with PBS; *: NK cell-depleted mice treated withAdFKN; *: NK cell-depleted mice treated with PBS. The datarepresent the means � SE of results from seven mice.
Fig. 6. Induction of tumor-specific CTL by intratumoraladministration of AdFKN. (A) C26 tumors established inBALB/c mice received injection of AdFKN (1�109 PFU), AdNull,or PBS alone (50 ll). Spleen cells were isolated 9 days aftertreatment. After restimulation in vitro for 5 days withmitomycin C-treated C26 cells, cytotoxicity of these spleno-cytes against C26 or BALB/c3T3 cells was determined by 6-h51Cr-release assay (n=3). The target cellswere C26 cells, and thesplenocytes treated with PBS (*), AdNull (*), or AdFKN (!).The target cells were BALB/c 3T3 cells, and the splenocytestreatedwith AdFKN (!). (B) The levels of IFN-c in supernatantsof splenocytes co-cultured with C26 or BALB/c3T3 cells afterstimulation of C26 cells were determined by ELISA kit. White:splenocytes from PBS-treated mice; gray: splenocytes treatedwith AdNull; black: splenocytes treated with AdFKN. The dataare means � SD of five samples.
Hong Xin et al. Eur. J. Immunol. 2005. 35: 1371–13801376
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

tissues such as inflammation, and mediates firmadhesion in an integrin-dependent and -independentmanner to bind their endothelial ligands with highaffinity through CX3CL1-CX3CR1 interactions [22, 24,29, 30]. In this context, we presumed that fractalkinecould induce a strong antitumor immune responsethrough its dual function in migration and adhesion ofleukocytes into tumor tissues.
We constructed AdFKN and confirmed that tumorcells infected with AdFKN expressed both membrane-bound form and biological active soluble form. CX3CL1gene was transferred to the tumor in vivo by directinjection of AdFKN into the tumor tissue that is relativelysimilar to the clinical setting in our model. Thistreatment modality strongly inhibited tumor growthby over 80% in both C26 and B16F10 tumors.
We demonstrated the evidence that antitumor effectsby fractalkine were dependent on both NK cells andT cells. Immunohistochemical staining of tumors treatedwith AdFKN in vivo revealed marked increase ofinfiltration of NK cells and slight increase of CD4+
T cells 3 days after the treatment, and subsequentlyapparent increase of CD8+ T lymphocytes but not NKcells or CD4+ T cells 6 days after the treatment.Fractalkine secreted into the tumor milieu directlyrecruited NK cells that expressed the fractalkine receptorinto tumor tissues and subsequently these recruited NKcells lysed tumor cells. Depletion of NK cells causing lossof the antitumor immune response confirmed the role ofNK cells in fractalkine-mediated antitumor effects.
DC, professional antigen-presenting cells, play apivotal role at the interphase of the innate and adaptiveimmune response [31, 32]. Since DC have been reportedto express both CX3CL1 and CX3CR1 [26], and CX3CL1expression to be up-regulated during the DC maturation[33], we investigated the infiltration of DC into tumortissues using CD11c antibody. We observed increased
Fig. 7. Induction of protective antitumor immunity byfractalkine. (A) BALB/c mice were immunized with1�106 irradiated AdFKN-transduced C26 cells (!), AdNull-transduced C26 cells (*), or nontransduced C26 cells (*) in theleft flank, and were challenged with parental C26 cells in theright flank after 2 weeks. (B) C57BL/6 mice were immunizedwith AdFKN-transduced B16F10 cells and rechallenged as in(A). The data represent the means � SE.
Fig. 8. Induction of CTL after immunization of AdFKN-transduced C26 cells. Two weeks after immunization withirradiatedC26 cells either transducedwithAdFKNorAdNull, ornontransduced, the splenocytes were harvested and restimu-lated with C26 cells. CTL assays (A) and IFN-c assay (B) wereperformed as in Fig. 6. The data represent the means � SD offive samples.
Eur. J. Immunol. 2005. 35: 1371–1380 Cellular immune response 1377
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

infiltration of CD11c+ cells into tumors on both day 3and day 6 after AdFKN treatment. These infiltratedimmature DC might efficiently capture tumor antigensfrom lysed tumor cells by NK cells, and possiblymaturation is induced by fractalkine. Subsequently,mature DC present tumor antigens and induce primingand activation of naive T cells, resulting in induction oftumor-specific adaptive immunity.
Based on CTL assay and IFN-c secretion in this study,it is likely that CD8+ T cells infiltrated into tumor tissuesare the tumor-specific CTL. The antitumor effect offractalkine was completely blocked in CD8–/– mice andpartially blocked in CD4–/– mice. These results suggestthat CD8+ T lymphocytes play an important role forfractalkine-mediated antitumor immune responses astumor-specific CTL. CD4+ T lymphocytes observed inthe tumor tissues could be T helper 1 cells and enhancecytotoxicity of CD8+ T cells through sensitizing tumorcells to CTL by IFN-c secretion as we demonstrated.Taken together, these experiments led us to concludethat both NK cells and T cells play a key role forantitumor effects by fractalkine.
With regard to the innate immunity versus adaptiveimmunity induced by AdFKN, Guo et al. [34] demon-strated CD4+, CD8+ T cell, and DC infiltration intoCX3CL1-tranduced Lewis lung carcinoma (LLC/CX3CL1), and concluded that LLC/CX3CL1 inducedantitumor immunity through chemoattraction andactivation of T cells and DC. In a subsequent study,they pointed out that NK cells also play an important rolein the antitumor immunity induced by LLC/CX3CL1cells [35]. On the other hand, in the EL-4 lymphomamodel, an antitumor effect by fractalkine was dependentonly on NK cells but not T cells, because this antitumoreffect was kept in T cell- and B cell-deficient Rag1–/–
mice but vanished in NK cell-deficient beige mice [36].In another study [37], no significant antitumor effect ofCX3CL1 was found using ex vivo gene transfer into OV-HM tumor cells by an adenoviral vector. This study is incontradiction to other reports [34–36].
We confirmed by in vivo observation a markedintratumoral infiltration of NK, DC, CD4+, and CD8+
T cells into the AdFKN-treated tumors, indicating that invivo genetic modification of tumor cells to expressfractalkine increased the number of tumor-reactiveCD4+ and CD8+ T cells in the tumor-bearing hosts.Magnitudes of NK cells and T cells on antitumor effectsmight be dependent on tumor types and/or treatmentprotocols. However, it is likely that both NK cells andT cells contribute to the antitumor immune response byfractalkine, since both NK and DC express CX3CR1 andcan induce tumor-specific CTL [26, 38, 39].
NK cell depletion also demonstrated completedisappearance of antitumor effects by AdFKN. Thisresult suggests that NK cells might play a role in
triggering adaptive immunity. Recent reports demon-strated that activated NK cells can promote maturationof immature DC [40]. Activated NK cells could inducematuration of DC, resulting in strong antitumor adaptiveimmunity.
In conclusion, we demonstrated strong antitumoreffects of fractalkine by intratumoral injection of AdFKNin solid tumor models. These antitumor effects weredependent on both NK cells and T cells. Fractalkine canbe a candidate for cancer gene therapy since fractalkinecan induce both innate and adaptive immunity againstsolid tumors.
Materials and methods
Cell culture and mice
Mouse colon adenocarcinoma cell line C26, mouse melanomacell line B16F10, BALB/c3T3 cells, and human monocyticleukemia cell line THP-1 were grown in RPMI 1640 mediumwith 10% fetal bovine serum. Female C57BL/6 and BALB/cmice were purchased from Charles River (Atsugi, Japan).Female CD4+ T cell-deficient and CD8+ T cell-deficient micethat had been backcrossed to the C57BL/6 background werepurchased from The Jackson Laboratory (Bar Harbor, ME).
Adenovirus vectors
The replication-deficient adenoviral vectors used in this studyare E1- and E3-deleted vectors based on human adenovirustype 5. An adenoviral vector was generated by homologousrecombination of pJM17 plasmid and shuttle plasmid vectorpCMV.SV2 containing an expression cassette under transcrip-tional control of the cytomegalovirus immediate-early en-hancer, followed by a mouse fractalkine cDNA [41]. Therecombinant adenovirus vectors were propagated using293 cells and purified by CsCl gradient centrifugation. Thetiters (expressed as PFU/ml) of viral stocks were quantified bya plaque-forming assay using 293 cells.
RT-PCR, Western blot, and flow cytometry
To determine the expression of fractalkine mRNA after AdFKNinfection, total cellular RNAwere extracted from the cells usingISOGEN (Nippon Gene Co., Tokyo, Japan) after 24 h infection.Total RNA (2 lg) were converted into cDNA by oligo(dT)12–18primers and Superscript II reverse transcriptase (GIBCO BRL,Carlsbad, CA). This cDNA was amplified with the followingprimers specific for either vector-derived fractalkine or controlb-actin transcripts: for fractalkine 50-GTCAGCACCTCGGCAT-GACGAAATG-3 and 50-CACTGGCACCAGGACGTATG-30; for b-actin 50-CTCTTTGATGTCACGCACGATTTC-30 and 50GTGGGC-CGCTCTAGGCACCAA-30.
The cells were lysed in RIPA buffer and protease inhibitors(Roche Molecular Biochemicals, Indianapolis, IN) after 48 hinfection with AdFKN. After determination of protein con-centration using the Bio-Rad Bradford protein assay (Bio-RadLaboratories Inc., Hercules, CA), 30 lg of total protein was
Hong Xin et al. Eur. J. Immunol. 2005. 35: 1371–13801378
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

loaded per lane on a 10% Bis-Tris Gel (Invitrogen, Carlsbad,CA) and transferred to a polyvinylidene difluoride membrane(Invitrogen). After blocking with 5% milk in Tris buffer/salinecontaining 0.1% Tween-20 at 4�C overnight, the membranewas incubated with rat anti-mouse fractalkine monoclonalantibody (R&D Systems, Minneapolis, MN) for 45 min at roomtemperature. Blots were stainedwith secondary antibody (goatanti-rat IgG-HR; Santa Cruz Biotechnology) for 45 min atroom temperature. The detection of specific signal wasperformed using the ECL detection system (AmershamPharmacia Biotech AB, Uppsala, Sweden).
After 48-h infection with AdFKN (MOI=100), cells werestained with a rat monoclonal antibody to mouse fractalkinefollowed by FITC-conjugated secondary antibody and ana-lyzed by flow cytometry (EPICS XL-MCL; Beckman Coulter,Hialeah, FL).
In vitro chemotaxis assay
Cell migration assay was carried out as described previously[42]. THP-1 cells (5�105 in 100 ll) were seeded in the upperwells of a Transwell plate (Costar, Cambridge, MA). The lowerwells were filled with 600 ll of supernatants of C26 cellsinfected with AdFKN at different MOI for 48 h. After 3-hincubation at 37�C, the cells that had migrated through thepolycarbonate membrane were harvested from the lowerchamber and counted under a microscope. The assay wasperformed in triplicate and the results were expressed as themean � standard deviation (SD).
Tumor studies in syngeneic mice and histologicalanalysis
In tumor treatment studies, tumor cells (5�105 C26 cells,3�105 B16F10 cells) were subcutaneously injected in the rightflank of female mice on day 0. When the tumors becamepalpable, AdFKN, AdNull (1�109 PFU/mouse), or PBS wasinjected intratumorally on day 5 (C26) or day 7 (B16F10).Each treatment group contained ten mice and studies wererepeated at least two times. To quantify the tumor growth, twoperpendicular diameters of the tumors were measured withcalipers three times per week. The tumor volume wascalculated using the formula: tumor volume (mm3) = 0.52� width2 � length. For tumor prevention studies, mice weresubcutaneously injected in the left flank with irradiated(50 Gy) B16F10 cells or C26 cells (1�106/mouse) infectedwith AdFKN or AdNull at MOI=100 on day –14. Two weeksafter the immunization (day 0), themice were challengedwithB16F10 cells or C26 cells in the right flank, and tumor volumewas calculated as described above. Each group contained sixmice, and studies were repeated three times.
For histological analysis, the tumors were excised andembedded with Tissue-Tek OCT embedding medium (SakuraFinetechnical Co. Ltd., Tokyo, Japan) as frozen sections 3 and6 days after treatment. After blocking of nonspecific stainingand endogenous peroxidase, sections were incubated over-night with optimal dilutions of the primary antibodiesincluding anti-mouse CD4 (RM4–5; BD PharMingen, SanDiego, CA), CD8 (KT15; Serotec Ltd., Oxford, UK), CD11c(HL3; BD PharMingen), rabbit anti-asialo GM1 (WAKO, Tokyo,
Japan), or isotype-matched IgG. The sections were thenincubated with biotin-labeled secondary antibody followed bystreptavidin-horseradish peroxidase, and applied with ABCreagents (Nichirei, Tokyo, Japan). The number of immunos-tained cells was determined by light microscopy at 400�magnification in ten randomly chosen fields for each section.
Tumor studies in CD4–/–, CD8–/–, and NK-depleted mice
AdFKN (1�109 PFU/mouse) was injected intratumorally intoB16F10 tumors on day 7 in CD4–/– or CD8–/– mice. For NK cellsdepletion, C57BL/6 mice and BALB/c mice were treated withasialo GM1 antiserum (200 lg/mouse) or same doses ofcontrol IgG by intraperitoneal injection six times on day –2, –1,0, 5, 10, and 15. The flow cytometric analysis using anti-NK1.1antibody (clone PK136; BD PharMingen) showed that theNK1.1+ population was less than 1%. These NK-depleted miceand control IgG-injected mice were transplanted with B16F10or C26 subcutaneously on day 0 and treated with AdFKN onday 7 or day 5. Each group contained seven mice.
Cytotoxic T lymphocyte assay
To assess the ability of AdFKN treatment and immunization toinduce tumor-specific CTL, splenocytes of tumor-bearing micetreated with AdFKN were isolated 9 days after the treatment,or 2 weeks after immunization. Effector cells were obtained byco-culturing splenocytes (5�106 cells) with mitomycin C-treated tumor cells in the presence of IL-2 (20 IU/ml).Cytotoxicity of effector cells was measured in a standard 6-h51Cr-release assay with C26 and B16F10 cells serving as targetcells. BALB/c 3T3 fibroblasts were used as negative control.The levels of mouse IFN-c released to the culture mediumduring the stimulation were assessed by enzyme-linkedimmunosorbent assay (ELISA) kit (R&D Systems).
References
1 Marincola, F. M., Jaffee, E. M., Hicklin, D. J. and Ferrone, S., Escape ofhuman solid tumors from T cell recognition: molecular mechanisms andfunctional significance. Adv. Immunol. 2000. 74: 181–273.
2 Rosenberg, S. A., Progress in human tumor immunology and immunother-apy. Nature 2001. 411: 380–384.
3 Ribas, A., Butterfield, L. H., Glaspy, J. A. and Economou, J. S., Currentdevelopments in cancer vaccines and cellular immunotherapy. J. Clin. Oncol.2003. 21: 2415–2432.
4 Keilholz, U. and Gore, M. E., Biochemotherapy for advanced melanoma.Semin. Oncol. 2002. 29: 456–461.
5 Rosenberg, S. A., Lotze, M. T., Muul, L. M., Chang, A. E., Avis, F. P.,Leitman, S., Linehan, W. M., Robertson, C. N., Lee, R. E. and Rubin, J. T.,A progress report on the treatment of 157 patients with advanced cancerusing lymphokine activated killer cells and interleukin-2 or high-doseinterleukin-2 alone. N. Engl. J. Med. 1987. 316: 889–897.
6 Fossa, S. D., Martinelli, G., Otto, U., Schneider, G., Wander, H., Oberling,F., Bauer, H.W., Achtnicht, U. andHoldener, E. E.,Recombinant interferonalfa-2a with or without vinblastin in metastatic renal cell carcinoma: resultsof a European multi-center phase III study. Ann. Oncol 1992. 3: 301–305.
7 Mier, J. W., Vachino, G., Klempner, M. S., Aronson, F. R., Noring, R.,Smith, S., Brandon, E. P., Laird, W. and Atkins, M. B., Inhibition ofinterleukin-2-induced tumor necrosis factor released by dexamethasone:prevention of an acquired neutrophil chemotaxis defect and differential
Eur. J. Immunol. 2005. 35: 1371–1380 Cellular immune response 1379
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

suppression of inteleukin-2-associated side effects. Blood 1990. 76:1933–1940.
8 Vogelzang, P. J., Bloom, S. M., Mier, J. W. and Atkins, M. B., Chestroentgenographic abnormalities in IL-2 recipients. Incidence and correlationwith clinical parameters. Chest 1992. 101: 746–752.
9 Dranoff, G., Cytokines in cancer pathogenesis and cancer therapy. Nat. Rev.Cancer 2004. 4: 11–22.
10 Uchiyama, A., Hoon, D. S., Morisaki, T., Kaneda, Y., Yuzuki, D. H. andMorton, D. L., Transfection of interleukin 2 gene into human melanomacells augments cellular immune response. Cancer Res. 1992. 53: 946–952.
11 Tahara, H., Zitvogel, L., Storkus, W. J., Zeh, H. J. 3rd., McKinney, T. G.,Schreiber, R. D., Gubler, U., Robbins, P. D. and Lotze, M. T., Effectiveeradication of established murine tumors with IL-12 gene therapy using apolycistronic retroviral vector. J. Immunol. 1995. 154: 6466–6474.
12 Natsume, A., Mizuno, M., Ryuke, Y. and Yoshida, J., Antitumor effect andcellular immunity activation bymurine interferon-beta gene transfer againstintracerebral glioma in mouse. Gene Ther. 1999. 6: 1626–1633.
13 Ono, S. J., Nakamura, T., Miyazaki, D., Ohbayashi, M., Dawson, M. andToda, M., Chemokines: roles in leukocyte development, trafficking, andeffector function. J. Allergy Clin. Immunol. 2003. 111: 1185–1199.
14 Arenberg, D. A., Kunkel, S. L., Polverini, P. J., Glass, M., Burdick, M. D.and Strieter, R. M., Inhibition of interleukin-8 reduces tumorigenesis ofhuman non-small cell lung cancer in SCID mice. J. Clin. Invest. 1996. 97:2792–2802.
15 Muller, A., Homey, B., Soto, H., Ge, N., Catron, D., Buchanan, M. E.,McClanahan, T., Murphy, E., Yuan, W., Wagner, S. N. et al., Involvementof chemokine receptors in breast cancer metastasis. Nature 2001. 410:50–56.
16 Guo, J., Wang, B., Zhang, M., Chen, T., Yu, Y., Regulier, E., Homann, H.E., Qin, Z., Ju, D. W. and Cao, X.,Macrophage-derived chemokine transferresults in tumor regression in murine lung carcinoma model throughefficient induction of antitumor immunity. Gene Ther. 2002. 9: 793–803.
17 Schroers, R., Davis, C. M., Wanger, H. J. and Chen, S. Y., Lentiviraltransduction of human T-lymphocytes with a RANTES intrakine inhibitshuman immunodeficiency virus type 1 infection. Gene Ther. 2000. 9:889–897.
18 Xia, D. J., Zhang,W. P., Zheng, S.,Wang, J., Pan, J. P.,Wang, Q., Zhang, L.H., Hamada, H. and Cao, X., Lymphotactin cotransfection enhances thetherapeutic efficacy of dendritic cells genetically modified with melanomaantigen gp100. Gene Ther. 2002. 9: 592–601.
19 Bazan, J. F., Bacon, K. B., Hardiman, G., Wang, W., Soo, K., Rossi, D.,Greaves, D. R., Zlotnik, A. and Schall, T. J., A new class of membrane-bound chemokine with a CX3C motif. Nature 1997. 385: 640–644.
20 Rossi, D. and Zlotnik, A., The biology of chemokines and their receptors.Annu. Rev. Immunol. 2000. 18: 217–242.
21 Garton, K. J., Gough, P. J., Blobel, C. P., Murphy, G., Greaves, D. R.,Dempsey, P. J. and Raines, E. W., Tumor necrosis factor-a-convertingenzyme (ADAM17) mediates the cleavage and shedding of fractalkine(CX3CL1). J. Biol. Chem. 2001. 276: 37993–38001.
22 Hundhausen, C., Misztela, D., Berkhout, T. A., Broadway, N., Saftig, P.,Reiss, K., Hartmann, D., Fahrenholz, F., Postina, R., Matthews, V. et al.,The disintegrin-like metalloproteinase ADAM10 is involved in constitutivecleavage of CX3CL1 (fractalkine) and regulates CX3CL1-mediated cell-celladhesion. Blood 2003. 102: 1186–1195.
23 Imai, T., Hieshima, K., Haskell, C., Baba, M., Nagira, M., Nishimura, M.,Kakizaki, M., Takagi, S., Nomiyama, H., Schall, T. J. et al., Identificationand molecular characterization of fractalkine receptor CX3CR1, whichmediates both leukocyte migration and adhesion. Cell 1997. 91: 521–530.
24 Fong, A. M., Alam, S. M., Imai, T., Haribabu, B. and Patel, D. D., CX3CR1tyrosine sulfation enhances fractalkine-induced cell adhesion. J. Biol. Chem.2002. 277: 19418–19423.
25 Nishimura, M., Umehara, H., Nakayama, T., Yoneda, O., Hieshima, K.,Kakizaki, M., Dohmae, N., Yoshie, O. and Imai, T., Dual functions of
fractalkine/CX3C ligand 1 trafficking of perforin+/granzyme B+ cytotoxiceffector lymphocytes that are defined by CX3CR1 expression. J. Immunol.2002. 168: 6173–6180.
26 Jung, S., Aliberti, J., Graemmel, P., Sunshine, M. J., Kreutzberg, G. W.,Sher, A. and Littman, D. R., Analysis of fractalkine receptor CX3CR1function by targeted deletion and green fluorescent protein reporter geneinsertion. Mol. Cell. Biol. 2000. 20: 4106–4114.
27 Sallusto, F., Mackay, C. R. and Lanzavecchia, A., The role of chemokinereceptor in primary, effector, and memory immune responses. Annu. Rev.Immunol. 2000. 18: 593–620.
28 Baggiolini, M., Chemokines and leukocyte traffic. Nature 1998. 392:565–568.
29 Goda, S., Imai, T., Yoshie, O., Yoneda, O., Inoue, H., Nagano, Y., Okazaki,T., Imai, H., Bloom, E. T., Domae, N. et al., CX3C-chemokine, fractalkine-enhanced adhesion of THP-1 cells to endothelial cells through integrin-dependent and -independent mechanisms. J. Immunol. 2000. 164:4313–4320.
30 Ancuta, P., Rao, R., Moses, A., Mehle, A., Shaw, S. K., Luscinskas, W. andGabuzda, D., Fractalkine preferentially mediates arrest and migration ofCD16+ monocytes. J. Exp. Med. 2003. 197: 1701–1707.
31 Belardelli, F. and Ferrantini, M., Cytokines as a link between innate andadaptive antitumor immunity. Trends Immunol. 2002. 23: 201–208.
32 Liu, Y. J., Dendritic cell subsets and lineages, and their functions in innateand adaptive immunity. Cell 2001. 106: 259–262.
33 Papadopoulos, E. J., Sassetti, C., Saeki, H., Yamada, N., Kawamura, T.,Fitzhugh, D. J., Saraf, M. A., Schall, T., Blauvelt, A., Rosen, S. D. et al.,Fractalkine, a CX3C chemokine, is expressed by dendritic cells and is up-regulated upon dendritic cell maturation. Eur. J. Immunol. 1999. 29:2551–2559.
34 Guo, J., Zhang, M., Wang, B., Yuan, Z., Guo, Z., Chen, T., Yu, Y., Qin, Z.and Cao, X., Fractalkine transgene induces T cell-dependent antitumorimmunity through chemoattraction and activation of dendritic cells. Int. J.Cancer 2003. 103: 212–220.
35 Guo, J., Chen, T., Wang, B., Zhang, M., An, H., Guo, Z., Yu, Y., Qin, Z. andCao, X., Chemoattraction, adhesion and activation of natural killer cells areinvolved in the antitumor immune response induced by fractalkine/CX3CL1.Immunol. Lett 2003. 89: 1–7.
36 Lavergne, E., Combadiere, B., Bonduelle, O., Iga, M., Gao, J. L., Maho,M., Boissonnas, A., Murphy, P. M., Debre, P. and Combadiere, C.,Fractalkine mediates natural killer-dependent antitumor responses in vivo.Cancer Res. 2003. 63: 7468–7474.
37 Gao, J. Q., Tsuda, Y., Katayama, K., Nakayama, T., Hatanaka, Y., Tani, Y.,Mizuguchi, H., Hayakawa, T., Yoshie, O., Tsutsumi, Y. et al., Antitumoreffect by interleukin-11 receptor alpha-locus chemokine/CCL27, introducedinto tumor cells through a recombinant adenovirus vector. Cancer Res. 2003.63: 4420–4425.
38 Fraticelli, P., Sironi, M., Bianchi, G., D'Ambrosio, D., Albanesi, C.,Stoppacciaro, A., Chieppa, M., Allavena, P., Ruco, L., Girolomoni, G. etal., Fractalkine (CX3CL1) as an amplification circuit of polarized Th1responses. J. Clin. Invest. 2001. 107: 1173–1181.
39 Cmbien, B., Pomeranz, M., Schmid-Antomarchi, H., Millet, M. A.,Breittmayer, V., Rossi, B., and Schmid-Alliana, A., Signal transductionpathways involved in soluble fractalkine-induced monocytic cell adhesion.Blood 2001. 97: 2031–2037.
40 Zitvogel, L., Dendritic and natural killer cells cooperate in the control/switch of innate immunity. J. Exp. Med. 2002. 195: F9–F14
41 Kanazawa, N., Nakamura, T., Tashiro, K., Muramatsu, M., Morita, K.,Yoneda, K., Inaba, K., Imamura, S. and Honjo, T., Fractalkine andmacrophage-derived chemokine: T cell-attracting chemokines expressed inT cell area dendritic cells. Eur. J. Immunol. 1999. 29: 1925–1932.
42 Carr, M. W., Roth, S. J., Luther, E., Rose, S. S. and Springer, T. A.,Monocyte chemoattractant protein 1 acts as a T-lymphocyte chemoattrac-tant. Proc. Natl. Acad. Sci. USA 1994. 91: 3652–3656.
Hong Xin et al. Eur. J. Immunol. 2005. 35: 1371–13801380
f 2005 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de