antipsychotics alter the protein expression levels of β-catenin and gsk-3 in the rat medial...
TRANSCRIPT

A�CH
BpcMgaRa�pCba
Kc
AdbasDr1raltEac
oipdecrs
F
A
R
0d
ntipsychotics Alter the Protein Expression Levels of-Catenin and GSK-3 in the Rat Medial Prefrontalortex and Striatum
eidar Alimohamad, Nagalingam Rajakumar, Yam-Hong Seah, and Walter Rushlow
ackground: It has been demonstrated that schizophrenics have altered levels and/or phosphorylation states of several Wnt relatedroteins in the brain, including �-catenin and GSK-3, and may represent susceptibility loci for schizophrenia. The current study wasonducted to assess the effects of antipsychotics on �-catenin and glycogen synthase kinase-3.ethods: Western blotting and immunocytochemistry were employed to investigate the effects of antipsychotics on �-catenin and
lycogen synthase kinase-3 following acute, subchronic and chronic drug administration. Specificity of the response was tested usingdditional drugs such as fluoxetine, amphetamine and valproic acid.esults: Significant increases in the levels of �-catenin and glycogen synthase kinase-3 total protein were identified followingdministration of clozapine, haloperidol or risperidone. The phosphorylation state of GSK-3 was also increased but phosphorylated-catenin levels were unaffected. Other drug compounds, with the exception of raclopride, had no effect on either GSK-3 or �-cateninrotein levels or distribution.onclusions: Targeting of �-catenin and GSK-3 is a common feature of antipsychotics regardless of class and appears to be mediatedy D2 dopamine receptors. Therefore changes in �-catenin and GSK-3 may represent one of the mechanisms through which
ntipsychotics are able to exert behavioral changes.ey Words: Antipsychotics, Wnt, GSK-3, �-catenin, prefrontalortex, striatum
ntipsychotics remain the only effective treatment for thesymptoms of schizophrenia. Though antipsychotics areoften classified as either typical or atypical and possess
ifferent receptor binding profiles, D2 dopamine (DA) receptorlockade is an essential feature of both classes of drugs (Kapurnd Mamo 2003). However, atypical antipsychotics are alsotrong serotonin receptor (5-HT) antagonists and the combinedA/5-HT action may be responsible for improved efficacy and
educed extrapyramidal symptoms (EPS) (Ichikawa and Meltzer999; Meltzer 1999). Current theories suggest that the blockade ofeceptors in the mesolimbic DA system is responsible for thebility of antipsychotics to alleviate psychosis whereas highevels of D2 DA receptor saturation in the striatonigral system ishought to cause disabling side effects commonly referred to asPS (Serretti et al 2004). Strong D2 DA antagonists, such as typicalntipsychotics, frequently induce EPS whereas atypical antipsy-hotics (weaker D2 antagonists) are less likely to do so.
Despite a good understanding of the receptor binding profilesf antipsychotics, the cellular consequences of repeatedly block-ng D2 DA receptors remains unclear and traditional signalingathways linked to DA receptors have not provided an easy orirect answer. Therefore other signaling pathways should bexamined to ascertain their potential involvement in the antipsy-hotic response. Fortunately several insights have been providedecently by studies using post-mortem brain tissue obtained fromchizophrenics and matched controls. One group of proteins that
rom the Departments of Anatomy and Cell Biology (HA, NR, WR) andPsychiatry (NR, Y-HS, WR), University of Western Ontario; and the Lon-don Health Sciences Centre (NR, WR), London, Ontario, Canada.
ddress reprint requests to Dr. Walter Rushlow, Department of Psychiatry,Room 10N15, London Health Science Center-U.C., 339 WindermereRoad, London, Ontario, Canada N6A 5A5; E-mail: [email protected].
eceived June 2, 2004; revised November 16, 2004; accepted November 23,
2004.006-3223/05/$30.00oi:10.1016/j.biopsych.2004.11.036
was identified as altered in schizophrenic brains belongs to theWnt signal transduction pathway. Wnt is important in centralnervous system (CNS) development (Cadigan and Nusse 1997;Miller and Moon 1996) and constituents of the pathway remainhighly expressed in the adult brain. Wnt has also been associatedwith a number of CNS disorders including Alzheimer’s diseaseand most recently schizophrenia (De Ferrari and Inestrosa 2000;Kozlovsky et al 2002).
The canonical Wnt signaling cascade is activated when a Wntprotein interacts with a Frizzled (Fz) receptor (Bhanot et al 1996).The interaction of Wnt with Fz leads to the phosphorylation ofdishevelled (Dvl) (Noordermeer et al 1994). Dvl antagonizes theaction of glycogen synthase kinase-3 (GSK-3) resulting in thecytoplasmic accumulation and translocation of �-catenin intothe nucleus (van Leeuwen et al 1994). A phosphorylationregulatory site has been identified in GSK-3 (Ser 21/9 for GSK-3�and GSK-3� respectively) that inhibits the kinase activity of anotherwise constitutively active protein and may be involved inregulating cytoplasmic �-catenin levels (Doble and Woodgett2003). Once in the nucleus, �-catenin binds to TCF/LEF (T-cellfactor and lymphoid enhancer factor), forming a transcriptionfactor complex that activates gene expression (Behrens et al1996; Huber et al 1996).
In the absence of Wnt, cytoplasmic �-catenin levels aremaintained at low levels through regulation by GSK-3,adenomatous polyposis coli (APC) and Axin (Hamada et al 1999;Ikeda et al 1998). The Axin-APC-GSK-3 complex promotes thephosphorylation of �-catenin by GSK-3 targeting it for destruc-tion by the ubiquitin-proteasome pathway (Aberle et al 1997;Polakis 1997; Siegfried et al 1992). In the absence of �-catenin,TCF/LEF still binds to the TCF promoter element but fails toactivate transcription (Brannon et al 1997; Riese et al 1997).
The purpose of the current study was to determine if anti-psychotics cause alterations in the protein levels or distributionof �-catenin and GSK-3 in the medial prefrontal cortex (PFC) orstriatum (Str). These two proteins were selected for characteriza-tion since �-catenin represents the key protein that transduces
activation of the Wnt pathway into the nucleus while GSK-3 isBIOL PSYCHIATRY 2005;57:533–542© 2005 Society of Biological Psychiatry

tD
M
A
1FtcfcuotmC
D
oia(Tcrarcms2adeTdhbtadcdsra1ghC
fmliaiwis
534 BIOL PSYCHIATRY 2005;57:533–542 H. Alimohamad et al
w
he central regulator of cytoplasmic �-catenin levels (Novak andedhar 1999).
ethods and Materials
nimalsFor all aspects of the study, adult female Sprague-Dawley rats
4� weeks of age (Charles River, Quebec, Canada) were used.emale rats were selected over male rats because they main-ained a more stable body weight and body mass index over theourse of the study. Rats were housed in pairs with free access toood and water in a room scheduled on a 12-hour light/darkycle. All efforts were made to minimize the number of animalssed in the current study and to eliminate pain and suffering. Usef animals for the current study was reviewed and approved byhe University of Western Ontario Animal Research Ethics Com-ittee in accordance with the guidelines developed by theanadian Council on Animal Care.
rugs and Drug ParadigmsFor the western blotting studies all rats received intramuscular
r subcutaneous, injections of .25 mg/kg or 1.0 mg/kg haloper-dol, .9 mg/kg or 2 mg/kg risperidone, 25 mg/kg clozapine orppropriate vehicles (n � 5 rats/treatment group) for 7 dayssub-chronic treatment) and sacrificed 2 hours post-injection.wo doses of haloperidol were selected for the study since aontroversy has recently arisen concerning appropriate dosing inats. One mg/kg is the dose that has often been used forntipsychotic studies but has been shown to induce catalepsy, aat measure of EPS liability. A lower dose results in saturation ofentral DA receptors in the rat brain that more closely approxi-ates what is observed in treated patients without significant
igns of EPS and may not induce catalepsy in rats (Kapur et al000a; Wadenberg et al 2001). However, the receptor occupancynd behavioral studies were only conducted following acuterug administration and it is not clear if the results can bextrapolated to a subchronic or chronic injection paradigm.herefore both doses of haloperidol were used in the study. Twooses of risperidone were also chosen, for similar reasons. Theigher dose of risperidone (2 mg/kg) would be expected toreak the threshold of atypicality and induce catalepsy whereashe lower dose (.9 mg/kg) is less likely to do so (Wadenberg etl 2001). Consistent with the behavioral observations, higheroses of risperidone induce immediate early genes in the lateralaudate-putamen (CPu) whereas the lower doses of risperidoneo not (Robertson et al 1994). Finally, the dose of clozapine waselected based on a receptor occupancy study showing compa-able D2 DA receptor saturation between 25 mg/kg of clozapinend the lower doses of risperidone and haloperidol (Schotte et al993). Only a single dose of clozapine was selected since it isenerally accepted that clozapine does not induce EPS in rats orumans and does not induce immediate early genes in the lateralPu of rats (Robertson et al 1994).
Antipsychotics only alleviate psychosis clinically in humansollowing repeated treatment and stabilization can take weeks oronths. To determine if changes in �-catenin and GSK-3 fol-
owed a similar pattern, rats (n � 5/treatment group) were alsonjected with haloperidol (1 mg/kg) or risperidone (.9 mg/kg)cutely (single injection) or chronically for 28 days (28 dailynjections) and sacrificed 2 hours post-injection. Only the PFCas examined following acute and chronic antipsychotic admin-
stration (and for most of the remaining portions of the study)
ince the results of the subchronic experiments suggested thatww.elsevier.com/locate/biopsych
the medial prefrontal cortex is commonly affected by all threedrugs and both doses of haloperidol and risperidone whereas thechanges in the Str are dependent on dose and/or drug class (i.e.typical versus atypical). In addition to the single daily bolusinjection experiments, rats were also injected with a single doseof haloperidol decanoate (1 mg/kg/day), a slow continuousrelease formulation of haloperidol lasting 14-21 days, or sesameseed oil vehicle and sacrificed 14 days following injection. Thehaloperidol decanoate was used to ensure the results were notthe consequence of delivery method or other factors such assedation.
To examine receptor and drug specificity properties, rats (n �5 rats/treatment group) were injected with raclopride (DA D2/D3
receptor antagonist, 3 mg/kg, 7 days), fluoxetine (selectiveserotonin reuptake inhibitor, 10 mg/kg, 9 days), ritanserin (sero-tonin 5-HT2a, 5-HT1c receptor antagonist, 1 mg/kg, 7 days),amphetamine (psychotomimetic, 2.5 and 5 mg/kg, 7 days),apomorphine (DA D1/D2 receptor agonist, .5 and 2.5 mg/kg, 7days), quinpirole (DA D2 receptor agonist, 2.5 mg/kg, 7 days),SKF-82958 (DA D1/D5 receptor agonist, 1 mg/kg, 7 days),valproic acid (mood stabilizer, 300 mg/kg, 10 days) or appropri-ate vehicles and sacrificed 2 or 4 hours following the finalinjection. The dose selected for each of the drugs was based onpublished work including behavioral studies involving pre-pulseinhibition (Geyer et al 2001). The length of treatment was chosento match the subchronic antipsychotic injection paradigm (7days) with the exception of valproic acid and fluoxetine (9 days).The treatment interval was extended for these two agents sinceseveral published studies used a slightly longer injection period.For example, a recent study examining the effects of lithium onGSK-3 used a nine day treatment interval for valproic acid (Gouldet al 2004).
For the immunocytochemistry portion of the study, rats (n �3/treatment) received intramuscular or subcutaneous injectionsof haloperidol (1 mg/kg), risperidone (.9 mg/kg or 2 mg/kg),amphetamine (5 mg/kg), apomorphine (2.5 mg/kg), quinpirole(2.5 mg/kg), valproic acid (300 mg/kg), ritanserin (1 mg/kg),fluoxetine (10 mg/kg), SKF-82958 (1 mg/kg) or vehicle daily for7-10 days (as indicated above) and were killed 2 hours post-injection.
All drugs used throughout the study were obtained fromSigma-Aldrich (Mississauga, Ontario, Canada).
Western BlotFollowing treatment the rats were decapitated, the brains
rapidly removed and dissected to obtain the Str (CPu andcaudal nucleus accumbens) and the PFC (medial prefrontalcortex plus anterior cingulate cortex). Tissue from individualrats was immediately homogenized on ice in ice-cold lysisbuffer (137 mM NaCl, 20 mM Tris (pH 8.0), 1% NP-40, 10%glycerol and .1% sodium dodecyl sulfate) to which a proteaseinhibitor tablet (Roche, Laval, Quebec, Canada) was addedusing a dounce homogenizer. For the phosphorylation stateportion of the study, Ser/Thr phosphatase inhibitors (Sigma)were included in the lysis buffer. The homogenized tissue wassonicated for 15 seconds, mixed with 5x loading buffer (125mM Tris pH 6.8, 10% glycerol, 2% SDS, 2% 2-mercaptoethanol,.01% xylene cyanol and .01% bromophenol blue) and boiledfor 5-7 minutes. The extracts were stored at -80°C untilneeded. Protein concentrations were determined using abicinchoninic acid Protein Assay Kit (Pierce Chemical Co.,Rockford, Illinois) and a �-Quant plate reader.
For western blot analysis, 15-25 �g of protein from drug

tSOttLb(oATw(ouACTCA(yr
1aw(Rr5sXbRceaduTo
I
basAbsaSagP
�ocaMl
H. Alimohamad et al BIOL PSYCHIATRY 2005;57:533–542 535
reated and control animals was subjected to electrophoresis onDS polyacrylamide minigels (Bio-Rad Laboratories, Mississauga,ntario, Canada) along with sizing standards (Bio-Rad Labora-
ories). Once electrophoresis was complete, the separated pro-eins were transferred to nitrocellulose membrane (Bio-Radaboratories). To detect the protein of interest, membranes werelocked in Carnation 5% nonfat dry milk in Tris buffered salineTBS) with .05% Tween-20 (TBST) and then probed with a seriesf antibodies raised against �-catenin, GSK-3 or �-tubulin.ntibodies were added to fresh nonfat dry milk solution (5%) inBST except for the phosphorylation state specific antibodieshich were added to TBST containing 5% bovine serum albumin
VWRCanlab, Mississauga, Ontario, Canada, fraction V) in placef milk. The sources, species and dilutions for the antibodiessed for western blotting were as follows; �-catenin (Sigma-ldrich, rabbit, c2206, 1:20,000; Santa Cruz Biotechnology (Santaruz, California), mouse, sc7963, 1:400), GSK-3 (Cell Signalingechnology, Beverly, Massachusetts, rabbit, #9332, 1:1,000; Santaruz Biotechnology, mouse, sc7291, 1:300), �-tubulin (Sigma-ldrich, mouse, T9026, 1:60,000), phosphorylated GSK-3� (Ser9)Cell Signaling Technology, rabbit, #9336, 1:1000) and phosphor-lated �-catenin (Ser33/37/Thr41) (Cell Signaling Technology,abbit, #9561, 1:1,000).
The blots were incubated in primary antibody solution forhour at room temperature, except for the phosphorylation specificntibodies which were incubated overnight at 4oC. Membranesere then incubated in the appropriate horseradish peroxidase
HRP) conjugated secondary antibodies (Pierce Chemical Co.,ockford, Illinois); goat anti-mouse IgG HRP, 1:50,000; goat anti-abbit IgG HRP antibody 1:50,000) diluted in 5% nonfat dry milk or% bovine serum albumin. Proteins were visualized using Super-ignal (WestPico, Pierce Chemical Co.) and X-ray film (Kodak-Omat LS). Multiple exposures were obtained to ensure that theands on the X-ray films used for quantification were not saturated.esults were quantified by densitometry and band intensity wasorrected for background by subtraction (Kodak software). Endog-nous �-tubulin was used as a control to ensure that all lanes withinblot contained approximately the same quantity of protein. Theata were statistically analyzed for differences between the groupssing a two-way analysis of variance (ANOVA), followed byukey’s Multiple Comparison Test or a Student’s t-test. Differencesf p � .05 (two-tailed) were considered to be significant.
mmunocytochemistryRats were injected intraperitoneally with sodium pentobar-
ital (65 mg/kg; MTC Pharmaceutical, Toronto, Ontario, Can-da) and perfused transcardially using 200 ml of salineolution followed by 500 ml of 2% paraformaldehyde (Sigma-ldrich) in .1M phosphate buffered saline (PBS, pH 7.4). Therains were immediately removed and cryoprotected in 15%ucrose solution for 24 hours. Cryoprotected brains were cutt 40 �m on a freezing microtome (Leitz, Wetzlar, Germany).ections through the PFC and Str were collected in .1M PBSnd then placed in blocking serum, composed of 10% normaloat serum (NGS) (Sigma-Aldrich) and .1% Triton X-100 inBS for one hour.
To qualitatively compare the protein levels and distribution of-catenin and GSK-3 following drug administration, immunoflu-rescence was employed. Immunofluorescence staining wasonducted as outlined previously (Rushlow et al 1996) usingppropriate Alexa-568 conjugated secondary antibodies (1:200;olecular Probes, Eugene, Oregon). Following fluorescence
abeling the sections were also counterstained with Hoechst. In
all instances, master mixes of antibody solutions were preparedand the same number of sections from comparable levels of thebrain were simultaneously labeled to facilitate direct comparisonbetween treated and untreated rats. In addition, regions notthought to be targeted by antipsychotics were examined alongwith the PFC and Str to ensure that sections were comparablystained. The fluorescence-labeled sections were examined andimages captured using an LSM410 scanning laser confocal mi-croscope (Zeiss, North York, Ontario, Canada).
Antibody SpecificityAll of the antibodies used in the study generated the expected
banding pattern on western blots. The �-catenin and �-tubulinantibodies generated a single distinct band while the GSK-3antibody generated 2 distinct bands representing the � and �isoforms of the protein. The relative size of �-catenin, �-tubulinand GSK-3, determined by molecular weight (MW) standardswas as expected for each of the proteins. In addition, 2 differentantibodies directed against GSK-3 and �-catenin were used toprobe western blots and produced identical results. Finally, blotsgenerated and probed without the primary antibody failed toshow any positive labeling.
Results
GSK- and �-catenin Protein Levels Following AntipsychoticDrug Administration as Revealed by Western Blotting
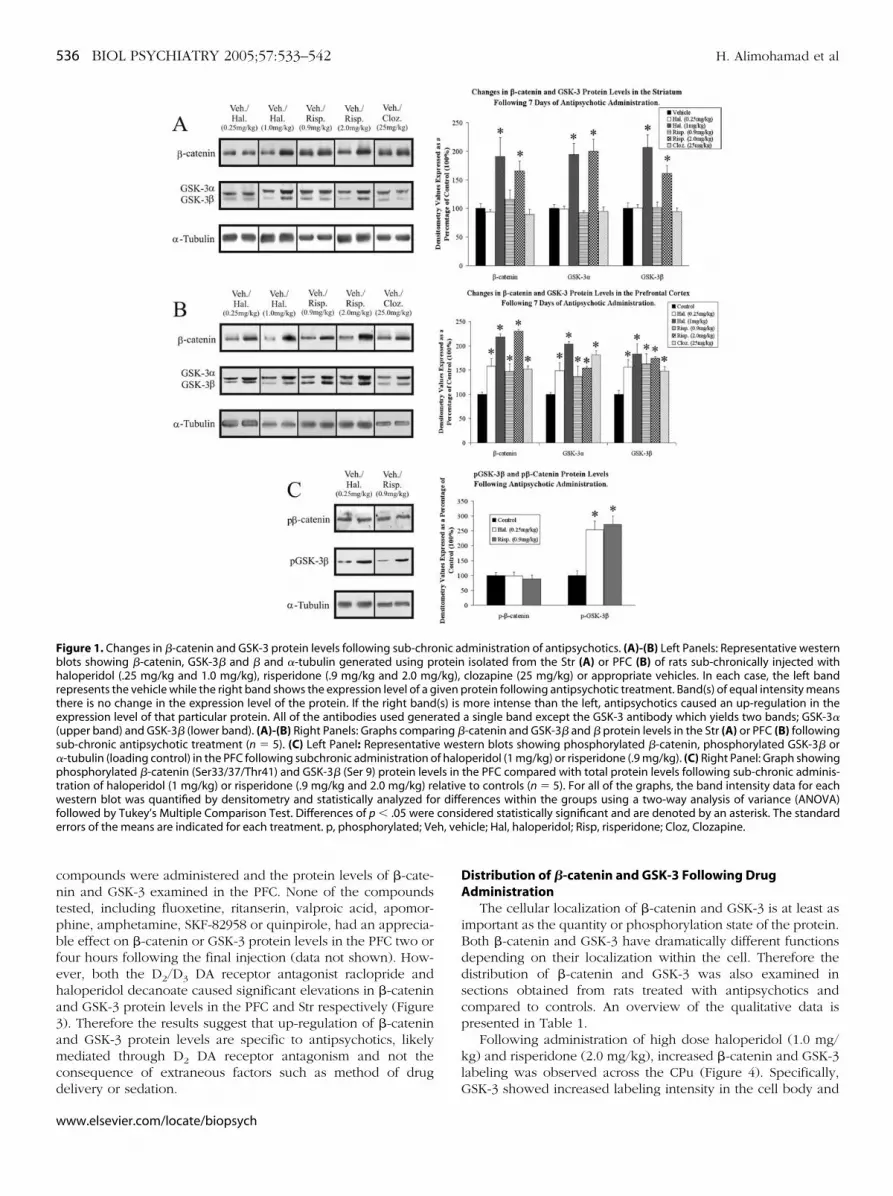
Sub-chronic (7 days) administration of haloperidol (1 mg/kg)and risperidone (2 mg/kg) caused a significant increase in �-cateninand GSK-3 protein levels in the Str two hours following the lastinjection of antipsychotic. A lower dose of haloperidol (.25 mg/kg),and risperidone (.9 mg/kg) as well as clozapine (25 mg/kg) had noeffect on �-catenin or GSK-3 protein levels (Figure 1A). All threedrugs, however, caused significant up-regulation of �-catenin andGSK-3 in the PFC at all doses examined (Figure 1B). Despitesignificant changes in overall �-catenin proteins levels, no change inthe level of phosphorylated �-catenin was observed in the PFC.However, significant increases in the phosphorylation state ofGSK-3� were detected in the PFC paralleling increases in totalprotein levels (Figure 1C).
The results suggest that antipsychotics, independent of class,have a profound effect on both �-catenin and GSK-3 in a regionof the brain (PFC) that is innervated by the mesolimbic DAsystem and believed to play a vital role in both schizophrenia andthe action of antipsychotics. The Str (CPu in particular), on theother hand, is innervated by the striatonigral DA system and onlyshows a clear response at doses of haloperidol and risperidoneexpected to exceed the threshold for EPS (Kapur et al 2000a;Wadenberg et al 2000).
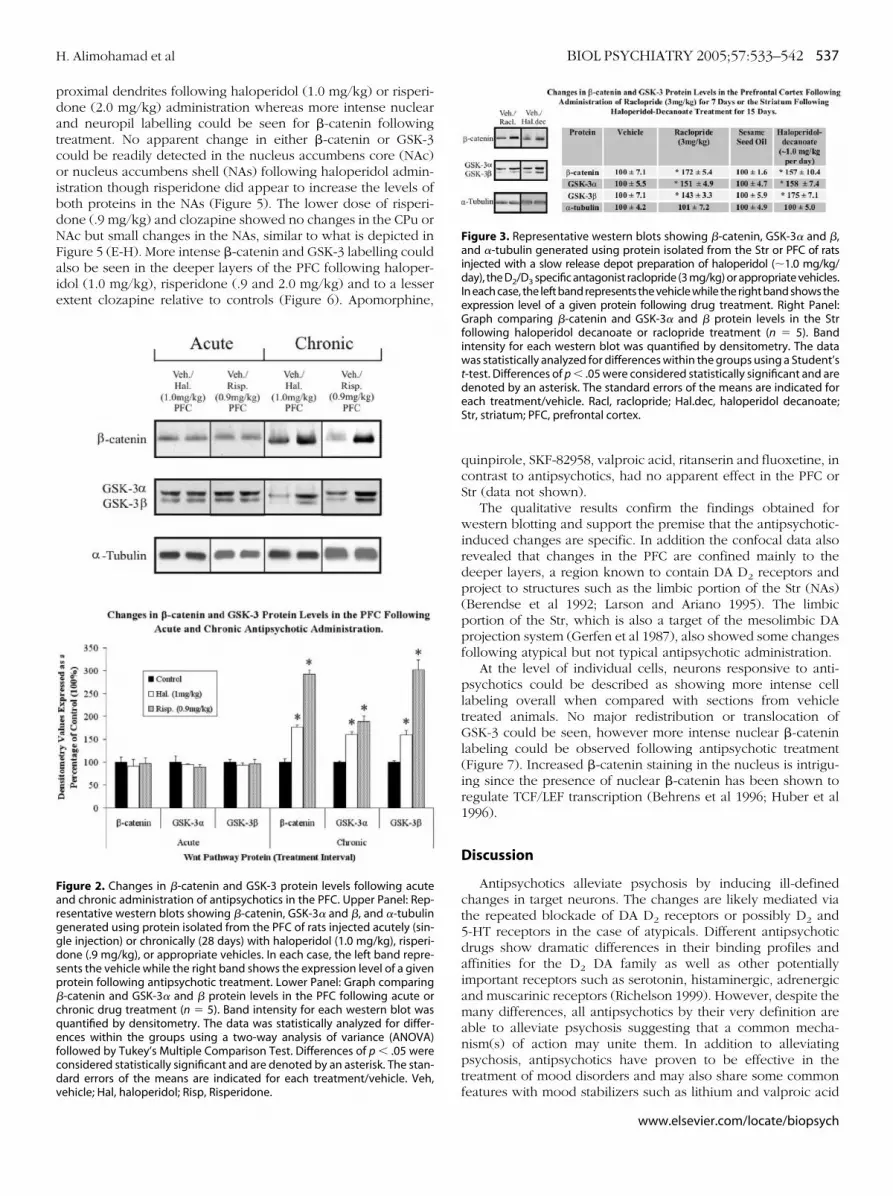
The protein levels of GSK-3 and �-catenin were also exam-ined following acute (1 injection) and chronic (28 daily injec-tions) administration of risperidone (.9 mg/kg), haloperidol (1mg/kg) or vehicle in the PFC. Acute treatment had no effect on�-catenin or GSK-3 protein levels while chronic administrationresulted in an identical pattern to that observed followingsubchronic administration (Figure 2). Therefore the results showthat changes in �-catenin and GSK-3 require repeated drugadministration to be manifested and persist with long term drugtreatment.
GSK-3 and �-catenin Protein Levels Following Administrationof Other Drug Compounds as Revealed by Western Blotting
To determine if the results obtained following antipsychotic
administration are specific to antipsychotics, a variety of differentwww.elsevier.com/locate/biopsych
cntpbfeha3amcd
Fbhrte(s�ptwfe h, ve
536 BIOL PSYCHIATRY 2005;57:533–542 H. Alimohamad et al
w
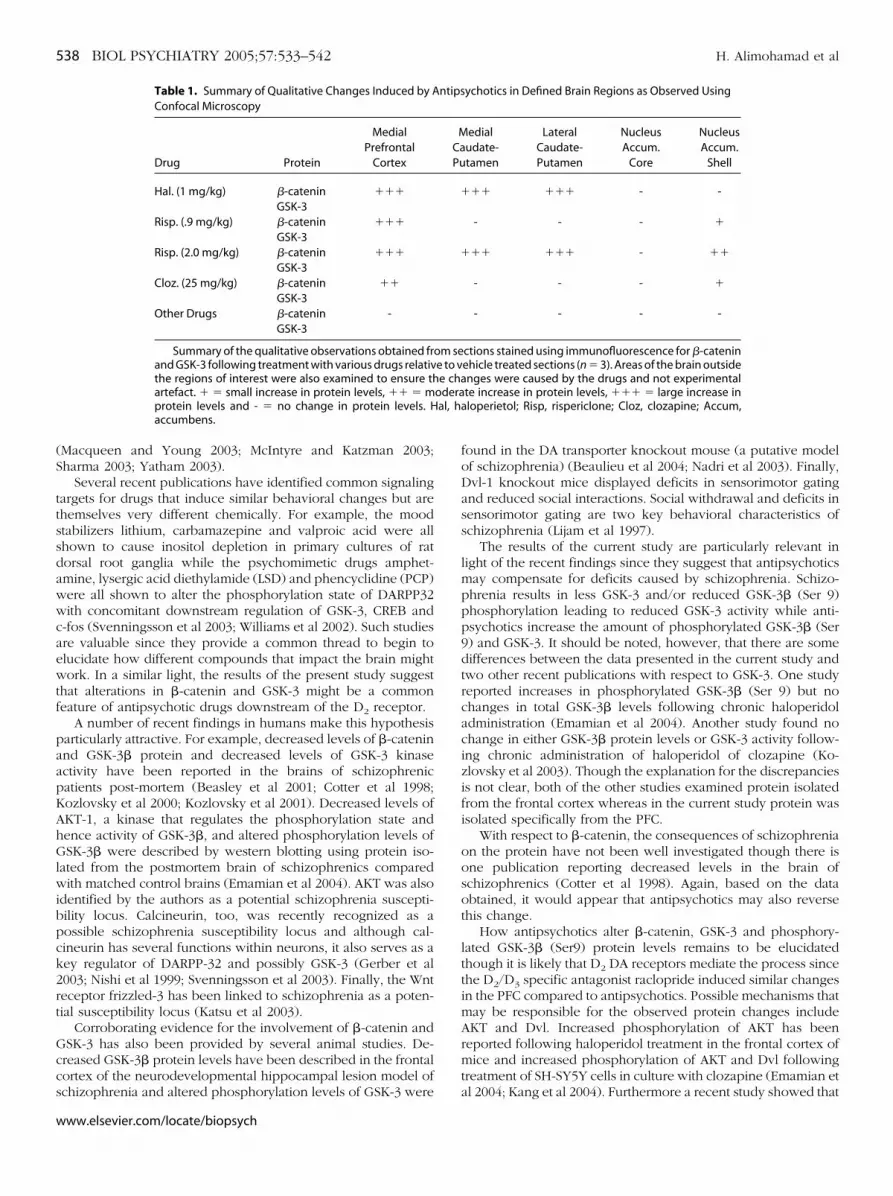
ompounds were administered and the protein levels of �-cate-in and GSK-3 examined in the PFC. None of the compoundsested, including fluoxetine, ritanserin, valproic acid, apomor-hine, amphetamine, SKF-82958 or quinpirole, had an apprecia-le effect on �-catenin or GSK-3 protein levels in the PFC two orour hours following the final injection (data not shown). How-ver, both the D2/D3 DA receptor antagonist raclopride andaloperidol decanoate caused significant elevations in �-cateninnd GSK-3 protein levels in the PFC and Str respectively (Figure). Therefore the results suggest that up-regulation of �-cateninnd GSK-3 protein levels are specific to antipsychotics, likelyediated through D2 DA receptor antagonism and not the
onsequence of extraneous factors such as method of drug
igure 1. Changes in �-catenin and GSK-3 protein levels following sub-chrolots showing �-catenin, GSK-3� and � and �-tubulin generated using paloperidol (.25 mg/kg and 1.0 mg/kg), risperidone (.9 mg/kg and 2.0 mg
epresents the vehicle while the right band shows the expression level of a ghere is no change in the expression level of the protein. If the right bandxpression level of that particular protein. All of the antibodies used gener
upper band) and GSK-3� (lower band). (A)-(B) Right Panels: Graphs compaub-chronic antipsychotic treatment (n � 5). (C) Left Panel: Representativ-tubulin (loading control) in the PFC following subchronic administration ohosphorylated �-catenin (Ser33/37/Thr41) and GSK-3� (Ser 9) protein leve
ration of haloperidol (1 mg/kg) or risperidone (.9 mg/kg and 2.0 mg/kg) reestern blot was quantified by densitometry and statistically analyzed fo
ollowed by Tukey’s Multiple Comparison Test. Differences of p � .05 wererrors of the means are indicated for each treatment. p, phosphorylated; Ve
elivery or sedation.
ww.elsevier.com/locate/biopsych
Distribution of �-catenin and GSK-3 Following DrugAdministration
The cellular localization of �-catenin and GSK-3 is at least asimportant as the quantity or phosphorylation state of the protein.Both �-catenin and GSK-3 have dramatically different functionsdepending on their localization within the cell. Therefore thedistribution of �-catenin and GSK-3 was also examined insections obtained from rats treated with antipsychotics andcompared to controls. An overview of the qualitative data ispresented in Table 1.
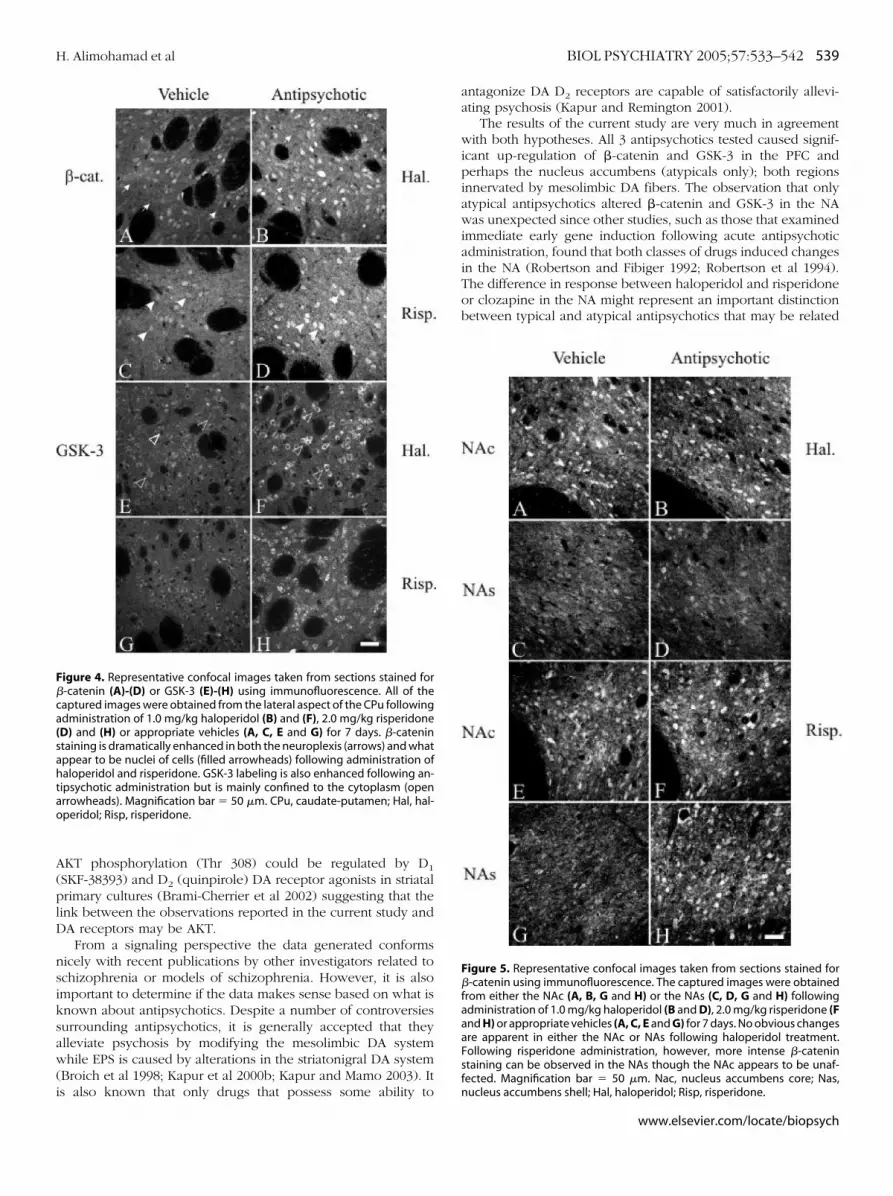
Following administration of high dose haloperidol (1.0 mg/kg) and risperidone (2.0 mg/kg), increased �-catenin and GSK-3labeling was observed across the CPu (Figure 4). Specifically,
dministration of antipsychotics. (A)-(B) Left Panels: Representative westernisolated from the Str (A) or PFC (B) of rats sub-chronically injected with
clozapine (25 mg/kg) or appropriate vehicles. In each case, the left bandrotein following antipsychotic treatment. Band(s) of equal intensity means
more intense than the left, antipsychotics caused an up-regulation in thea single band except the GSK-3 antibody which yields two bands; GSK-3�-catenin and GSK-3� and � protein levels in the Str (A) or PFC (B) followingtern blots showing phosphorylated �-catenin, phosphorylated GSK-3� orperidol (1 mg/kg) or risperidone (.9 mg/kg). (C) Right Panel: Graph showing
the PFC compared with total protein levels following sub-chronic adminis-to controls (n � 5). For all of the graphs, the band intensity data for each
erences within the groups using a two-way analysis of variance (ANOVA)dered statistically significant and are denoted by an asterisk. The standardhicle; Hal, haloperidol; Risp, risperidone; Cloz, Clozapine.
nic arotein/kg),iven p(s) isated
ring �e wesf halols inlativer diffconsi
GSK-3 showed increased labeling intensity in the cell body and
pdatcoibdNFaie
Farggdsp�cqefcdv
H. Alimohamad et al BIOL PSYCHIATRY 2005;57:533–542 537
roximal dendrites following haloperidol (1.0 mg/kg) or risperi-one (2.0 mg/kg) administration whereas more intense nuclearnd neuropil labelling could be seen for �-catenin followingreatment. No apparent change in either �-catenin or GSK-3ould be readily detected in the nucleus accumbens core (NAc)r nucleus accumbens shell (NAs) following haloperidol admin-stration though risperidone did appear to increase the levels ofoth proteins in the NAs (Figure 5). The lower dose of risperi-one (.9 mg/kg) and clozapine showed no changes in the CPu orAc but small changes in the NAs, similar to what is depicted inigure 5 (E-H). More intense �-catenin and GSK-3 labelling couldlso be seen in the deeper layers of the PFC following haloper-dol (1.0 mg/kg), risperidone (.9 and 2.0 mg/kg) and to a lesserxtent clozapine relative to controls (Figure 6). Apomorphine,
igure 2. Changes in �-catenin and GSK-3 protein levels following acutend chronic administration of antipsychotics in the PFC. Upper Panel: Rep-esentative western blots showing �-catenin, GSK-3� and �, and �-tubulinenerated using protein isolated from the PFC of rats injected acutely (sin-le injection) or chronically (28 days) with haloperidol (1.0 mg/kg), risperi-one (.9 mg/kg), or appropriate vehicles. In each case, the left band repre-ents the vehicle while the right band shows the expression level of a givenrotein following antipsychotic treatment. Lower Panel: Graph comparing-catenin and GSK-3� and � protein levels in the PFC following acute orhronic drug treatment (n � 5). Band intensity for each western blot wasuantified by densitometry. The data was statistically analyzed for differ-nces within the groups using a two-way analysis of variance (ANOVA)ollowed by Tukey’s Multiple Comparison Test. Differences of p � .05 wereonsidered statistically significant and are denoted by an asterisk. The stan-ard errors of the means are indicated for each treatment/vehicle. Veh,
ehicle; Hal, haloperidol; Risp, Risperidone.quinpirole, SKF-82958, valproic acid, ritanserin and fluoxetine, incontrast to antipsychotics, had no apparent effect in the PFC orStr (data not shown).
The qualitative results confirm the findings obtained forwestern blotting and support the premise that the antipsychotic-induced changes are specific. In addition the confocal data alsorevealed that changes in the PFC are confined mainly to thedeeper layers, a region known to contain DA D2 receptors andproject to structures such as the limbic portion of the Str (NAs)(Berendse et al 1992; Larson and Ariano 1995). The limbicportion of the Str, which is also a target of the mesolimbic DAprojection system (Gerfen et al 1987), also showed some changesfollowing atypical but not typical antipsychotic administration.
At the level of individual cells, neurons responsive to anti-psychotics could be described as showing more intense celllabeling overall when compared with sections from vehicletreated animals. No major redistribution or translocation ofGSK-3 could be seen, however more intense nuclear �-cateninlabeling could be observed following antipsychotic treatment(Figure 7). Increased �-catenin staining in the nucleus is intrigu-ing since the presence of nuclear �-catenin has been shown toregulate TCF/LEF transcription (Behrens et al 1996; Huber et al1996).
Discussion
Antipsychotics alleviate psychosis by inducing ill-definedchanges in target neurons. The changes are likely mediated viathe repeated blockade of DA D2 receptors or possibly D2 and5-HT receptors in the case of atypicals. Different antipsychoticdrugs show dramatic differences in their binding profiles andaffinities for the D2 DA family as well as other potentiallyimportant receptors such as serotonin, histaminergic, adrenergicand muscarinic receptors (Richelson 1999). However, despite themany differences, all antipsychotics by their very definition areable to alleviate psychosis suggesting that a common mecha-nism(s) of action may unite them. In addition to alleviatingpsychosis, antipsychotics have proven to be effective in thetreatment of mood disorders and may also share some common
Figure 3. Representative western blots showing �-catenin, GSK-3� and �,and �-tubulin generated using protein isolated from the Str or PFC of ratsinjected with a slow release depot preparation of haloperidol (�1.0 mg/kg/day), the D2/D3 specific antagonist raclopride (3 mg/kg) or appropriate vehicles.In each case, the left band represents the vehicle while the right band shows theexpression level of a given protein following drug treatment. Right Panel:Graph comparing �-catenin and GSK-3� and � protein levels in the Strfollowing haloperidol decanoate or raclopride treatment (n � 5). Bandintensity for each western blot was quantified by densitometry. The datawas statistically analyzed for differences within the groups using a Student’st-test. Differences of p � .05 were considered statistically significant and aredenoted by an asterisk. The standard errors of the means are indicated foreach treatment/vehicle. Racl, raclopride; Hal.dec, haloperidol decanoate;Str, striatum; PFC, prefrontal cortex.
features with mood stabilizers such as lithium and valproic acid
www.elsevier.com/locate/biopsych
(S
ttssdawwcaewtf
paapKAhGlwibpck2rt
Gccs
al, ha
538 BIOL PSYCHIATRY 2005;57:533–542 H. Alimohamad et al
w
Macqueen and Young 2003; McIntyre and Katzman 2003;harma 2003; Yatham 2003).
Several recent publications have identified common signalingargets for drugs that induce similar behavioral changes but arehemselves very different chemically. For example, the moodtabilizers lithium, carbamazepine and valproic acid were allhown to cause inositol depletion in primary cultures of ratorsal root ganglia while the psychomimetic drugs amphet-mine, lysergic acid diethylamide (LSD) and phencyclidine (PCP)ere all shown to alter the phosphorylation state of DARPP32ith concomitant downstream regulation of GSK-3, CREB and
-fos (Svenningsson et al 2003; Williams et al 2002). Such studiesre valuable since they provide a common thread to begin tolucidate how different compounds that impact the brain mightork. In a similar light, the results of the present study suggest
hat alterations in �-catenin and GSK-3 might be a commoneature of antipsychotic drugs downstream of the D2 receptor.
A number of recent findings in humans make this hypothesisarticularly attractive. For example, decreased levels of �-cateninnd GSK-3� protein and decreased levels of GSK-3 kinasectivity have been reported in the brains of schizophrenicatients post-mortem (Beasley et al 2001; Cotter et al 1998;ozlovsky et al 2000; Kozlovsky et al 2001). Decreased levels ofKT-1, a kinase that regulates the phosphorylation state andence activity of GSK-3�, and altered phosphorylation levels ofSK-3� were described by western blotting using protein iso-
ated from the postmortem brain of schizophrenics comparedith matched control brains (Emamian et al 2004). AKT was also
dentified by the authors as a potential schizophrenia suscepti-ility locus. Calcineurin, too, was recently recognized as aossible schizophrenia susceptibility locus and although cal-ineurin has several functions within neurons, it also serves as aey regulator of DARPP-32 and possibly GSK-3 (Gerber et al003; Nishi et al 1999; Svenningsson et al 2003). Finally, the Wnteceptor frizzled-3 has been linked to schizophrenia as a poten-ial susceptibility locus (Katsu et al 2003).
Corroborating evidence for the involvement of �-catenin andSK-3 has also been provided by several animal studies. De-reased GSK-3� protein levels have been described in the frontalortex of the neurodevelopmental hippocampal lesion model of
Table 1. Summary of Qualitative Changes Induced by AConfocal Microscopy
Drug Protein
MedialPrefrontal
Cortex
Hal. (1 mg/kg) �-catenin ���GSK-3
Risp. (.9 mg/kg) �-catenin ���GSK-3
Risp. (2.0 mg/kg) �-catenin ���GSK-3
Cloz. (25 mg/kg) �-catenin ��GSK-3
Other Drugs �-catenin -GSK-3
Summary of the qualitative observations obtained frand GSK-3 following treatment with various drugs relativthe regions of interest were also examined to ensure thartefact. � � small increase in protein levels, �� � mprotein levels and - � no change in protein levels. Haccumbens.
chizophrenia and altered phosphorylation levels of GSK-3 were
ww.elsevier.com/locate/biopsych
found in the DA transporter knockout mouse (a putative modelof schizophrenia) (Beaulieu et al 2004; Nadri et al 2003). Finally,Dvl-1 knockout mice displayed deficits in sensorimotor gatingand reduced social interactions. Social withdrawal and deficits insensorimotor gating are two key behavioral characteristics ofschizophrenia (Lijam et al 1997).
The results of the current study are particularly relevant inlight of the recent findings since they suggest that antipsychoticsmay compensate for deficits caused by schizophrenia. Schizo-phrenia results in less GSK-3 and/or reduced GSK-3� (Ser 9)phosphorylation leading to reduced GSK-3 activity while anti-psychotics increase the amount of phosphorylated GSK-3� (Ser9) and GSK-3. It should be noted, however, that there are somedifferences between the data presented in the current study andtwo other recent publications with respect to GSK-3. One studyreported increases in phosphorylated GSK-3� (Ser 9) but nochanges in total GSK-3� levels following chronic haloperidoladministration (Emamian et al 2004). Another study found nochange in either GSK-3� protein levels or GSK-3 activity follow-ing chronic administration of haloperidol of clozapine (Ko-zlovsky et al 2003). Though the explanation for the discrepanciesis not clear, both of the other studies examined protein isolatedfrom the frontal cortex whereas in the current study protein wasisolated specifically from the PFC.
With respect to �-catenin, the consequences of schizophreniaon the protein have not been well investigated though there isone publication reporting decreased levels in the brain ofschizophrenics (Cotter et al 1998). Again, based on the dataobtained, it would appear that antipsychotics may also reversethis change.
How antipsychotics alter �-catenin, GSK-3 and phosphory-lated GSK-3� (Ser9) protein levels remains to be elucidatedthough it is likely that D2 DA receptors mediate the process sincethe D2/D3 specific antagonist raclopride induced similar changesin the PFC compared to antipsychotics. Possible mechanisms thatmay be responsible for the observed protein changes includeAKT and Dvl. Increased phosphorylation of AKT has beenreported following haloperidol treatment in the frontal cortex ofmice and increased phosphorylation of AKT and Dvl followingtreatment of SH-SY5Y cells in culture with clozapine (Emamian et
ychotics in Defined Brain Regions as Observed Using
edialudate-tamen
LateralCaudate-Putamen
NucleusAccum.
Core
NucleusAccum.
Shell
�� ��� - -
- - - �
�� ��� - ��
- - - �
- - - -
ctions stained using immunofluorescence for �-cateninehicle treated sections (n � 3). Areas of the brain outsidenges were caused by the drugs and not experimentalte increase in protein levels, ��� � large increase inloperietol; Risp, rispericlone; Cloz, clozapine; Accum,
ntips
MCaPu
�
�
om see to ve cha
odera
al 2004; Kang et al 2004). Furthermore a recent study showed that
A(plD
nsiksaw(i
F�ca(sahtao
H. Alimohamad et al BIOL PSYCHIATRY 2005;57:533–542 539
KT phosphorylation (Thr 308) could be regulated by D1
SKF-38393) and D2 (quinpirole) DA receptor agonists in striatalrimary cultures (Brami-Cherrier et al 2002) suggesting that the
ink between the observations reported in the current study andA receptors may be AKT.From a signaling perspective the data generated conforms
icely with recent publications by other investigators related tochizophrenia or models of schizophrenia. However, it is alsomportant to determine if the data makes sense based on what isnown about antipsychotics. Despite a number of controversiesurrounding antipsychotics, it is generally accepted that theylleviate psychosis by modifying the mesolimbic DA systemhile EPS is caused by alterations in the striatonigral DA system
Broich et al 1998; Kapur et al 2000b; Kapur and Mamo 2003). It
igure 4. Representative confocal images taken from sections stained for-catenin (A)-(D) or GSK-3 (E)-(H) using immunofluorescence. All of theaptured images were obtained from the lateral aspect of the CPu followingdministration of 1.0 mg/kg haloperidol (B) and (F), 2.0 mg/kg risperidoneD) and (H) or appropriate vehicles (A, C, E and G) for 7 days. �-catenintaining is dramatically enhanced in both the neuroplexis (arrows) and whatppear to be nuclei of cells (filled arrowheads) following administration ofaloperidol and risperidone. GSK-3 labeling is also enhanced following an-
ipsychotic administration but is mainly confined to the cytoplasm (openrrowheads). Magnification bar � 50 �m. CPu, caudate-putamen; Hal, hal-peridol; Risp, risperidone.
s also known that only drugs that possess some ability to
antagonize DA D2 receptors are capable of satisfactorily allevi-ating psychosis (Kapur and Remington 2001).
The results of the current study are very much in agreementwith both hypotheses. All 3 antipsychotics tested caused signif-icant up-regulation of �-catenin and GSK-3 in the PFC andperhaps the nucleus accumbens (atypicals only); both regionsinnervated by mesolimbic DA fibers. The observation that onlyatypical antipsychotics altered �-catenin and GSK-3 in the NAwas unexpected since other studies, such as those that examinedimmediate early gene induction following acute antipsychoticadministration, found that both classes of drugs induced changesin the NA (Robertson and Fibiger 1992; Robertson et al 1994).The difference in response between haloperidol and risperidoneor clozapine in the NA might represent an important distinctionbetween typical and atypical antipsychotics that may be related
Figure 5. Representative confocal images taken from sections stained for�-catenin using immunofluorescence. The captured images were obtainedfrom either the NAc (A, B, G and H) or the NAs (C, D, G and H) followingadministration of 1.0 mg/kg haloperidol (B and D), 2.0 mg/kg risperidone (Fand H) or appropriate vehicles (A, C, E and G) for 7 days. No obvious changesare apparent in either the NAc or NAs following haloperidol treatment.Following risperidone administration, however, more intense �-cateninstaining can be observed in the NAs though the NAc appears to be unaf-fected. Magnification bar � 50 �m. Nac, nucleus accumbens core; Nas,
nucleus accumbens shell; Hal, haloperidol; Risp, risperidone.www.elsevier.com/locate/biopsych
tamts
edrcFntfoDC
fmdasb(ohscp
F�cfDs(aGl(ul
540 BIOL PSYCHIATRY 2005;57:533–542 H. Alimohamad et al
w
o the improved clinical efficacy observed with the use oftypicals. Alternatively, the differences may be due to experi-ental factors such as dose, timing of protein isolation following
he final injection or sensitivity of immunofluoresence to detectmall changes.
In the CPu, only a high dose of haloperidol or risperidonelicited increases in �-catenin and GSK-3 protein levels. Highoses of both drugs would be expected to saturate D2 DAeceptors in the Str and cause EPS in humans or induceatalepsy in rats (Kapur et al 2000a; Wadenberg et al 2001).urthermore, clozapine, a drug that rarely causes EPS, elicitedo response in the Str at 25 mg/kg. This is not to suggest thathe changes observed in �-catenin and GSK-3 are responsibleor EPS, but rather that the results are consistent with previ-usly reported studies showing a correlation between high D2
A receptor saturation, changes in protein and mRNA in thePu and EPS.
Specificity is another important issue for antipsychotics. Dif-erent drug compounds impact the brain differently though thereay be some overlap in their biological effects. For example,rugs used to treat other disorders (i.e. depression) do notlleviate psychosis in human patients. However, both moodtabilizers and antipsychotics are effective for the treatment ofipolar disorder. In the current study, neither valproic acidmood stabilizer) nor fluoxetine (anti-depressant) had an effectn GSK-3 or �-catenin protein levels in the PFC or Str, 2 or 4ours post final injection. Results for valproic acid are somewhaturprising given literature reports indicating that valproic acidan upregulate �-catenin in vitro (Chen et al 1999). The ability of
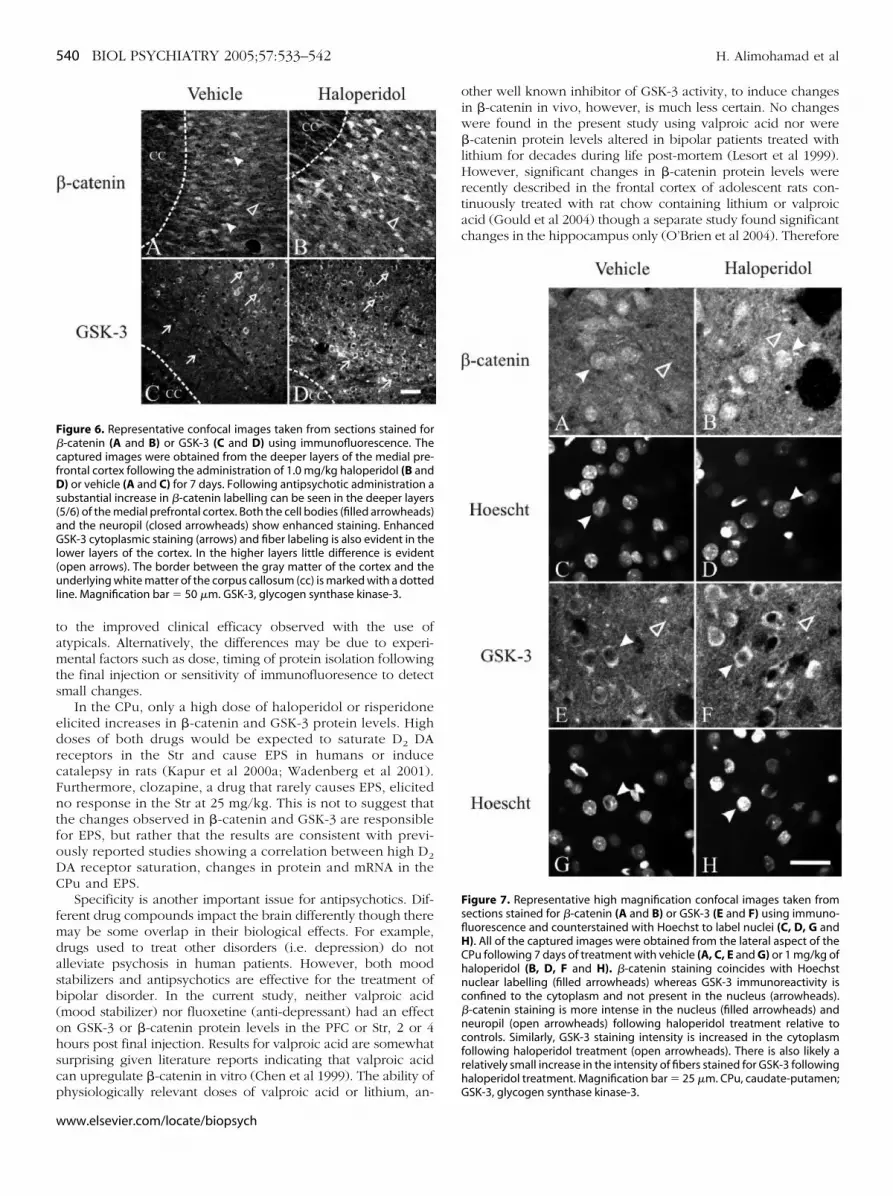
igure 6. Representative confocal images taken from sections stained for-catenin (A and B) or GSK-3 (C and D) using immunofluorescence. Theaptured images were obtained from the deeper layers of the medial pre-rontal cortex following the administration of 1.0 mg/kg haloperidol (B and
) or vehicle (A and C) for 7 days. Following antipsychotic administration aubstantial increase in �-catenin labelling can be seen in the deeper layers5/6) of the medial prefrontal cortex. Both the cell bodies (filled arrowheads)nd the neuropil (closed arrowheads) show enhanced staining. EnhancedSK-3 cytoplasmic staining (arrows) and fiber labeling is also evident in the
ower layers of the cortex. In the higher layers little difference is evidentopen arrows). The border between the gray matter of the cortex and thenderlying white matter of the corpus callosum (cc) is marked with a dotted
ine. Magnification bar � 50 �m. GSK-3, glycogen synthase kinase-3.
hysiologically relevant doses of valproic acid or lithium, an-
ww.elsevier.com/locate/biopsych
other well known inhibitor of GSK-3 activity, to induce changesin �-catenin in vivo, however, is much less certain. No changeswere found in the present study using valproic acid nor were�-catenin protein levels altered in bipolar patients treated withlithium for decades during life post-mortem (Lesort et al 1999).However, significant changes in �-catenin protein levels wererecently described in the frontal cortex of adolescent rats con-tinuously treated with rat chow containing lithium or valproicacid (Gould et al 2004) though a separate study found significantchanges in the hippocampus only (O’Brien et al 2004). Therefore
Figure 7. Representative high magnification confocal images taken fromsections stained for �-catenin (A and B) or GSK-3 (E and F) using immuno-fluorescence and counterstained with Hoechst to label nuclei (C, D, G andH). All of the captured images were obtained from the lateral aspect of theCPu following 7 days of treatment with vehicle (A, C, E and G) or 1 mg/kg ofhaloperidol (B, D, F and H). �-catenin staining coincides with Hoechstnuclear labelling (filled arrowheads) whereas GSK-3 immunoreactivity isconfined to the cytoplasm and not present in the nucleus (arrowheads).�-catenin staining is more intense in the nucleus (filled arrowheads) andneuropil (open arrowheads) following haloperidol treatment relative tocontrols. Similarly, GSK-3 staining intensity is increased in the cytoplasmfollowing haloperidol treatment (open arrowheads). There is also likely arelatively small increase in the intensity of fibers stained for GSK-3 followinghaloperidol treatment. Magnification bar � 25 �m. CPu, caudate-putamen;
GSK-3, glycogen synthase kinase-3.
tabn
pnideiaAkteHtite
acadciirdlsd
RcF
t
A
B
B
B
B
B
B
H. Alimohamad et al BIOL PSYCHIATRY 2005;57:533–542 541
here may be some overlap between mood stabilizers andntipsychotics though perhaps they don’t target GSK-3/�-cateniny the same mechanism or affect the same populations ofeurons.
A variety of DA agonists including quinpirole, apomor-hine and SKF-82958 also failed to induce changes in �-cate-in or GSK-3 protein levels in either the PFC or Str. Thenability of the other drugs to alter �-catenin and GSK-3emonstrates that the antipsychotic response is not a genericffect such as has been described for the induction ofmmediate early genes (Robertson et al 1994). Amphetaminelso had no effect in the PFC on either GSK-3 or �-catenin.gain the results were unexpected since amphetamine is anown psychomimetic and can induce schizophrenic symp-oms in individuals who abuse the drug and thereby might bexpected to show the opposite response to antipsychotics.owever, a recent study reported that amphetamine can alter
he phosphorylation state of DARPP-32 and GSK-3� suggest-ng similar targets for both psychomimetics and antipsychoticshough additional studies will be necessary to clarify differ-nces in mechanism (Svenningsson et al 2003).
The changes identified following antipsychotic administrationre also consistent with clinical observations concerning the timeourse of treatment for schizophrenic patients treated withntipsychotics. It has been well established that antipsychoticso not alleviate psychosis after a single dose. Likewise antipsy-hotics have no effect on �-catenin or GSK-3 following a singlenjection in the rat. After repeated administration, improvementn psychotic symptoms can be observed in humans, and in theat, changes in GSK-3 and �-catenin protein levels can beetected. Finally, continuous treatment for weeks to monthseads to stabilization of positive symptoms in human patients andignificant elevations in both �-catenin and GSK-3 are readilyetectable after chronic treatment.
This research was supported by the National Alliance foresearch on Schizophrenia and Depression, the Canadian Psy-hiatric Research Foundation and the Ontario Mental Healthoundation.
We acknowledge Ms. Joanne Mouyal for her contribution tohe study.
berle H, Bauer A, Stappert J, Kispert A, Kemler R (1997): beta-catenin is atarget for the ubiquitin-proteasome pathway. EMBO J 16:3797–3804.
easley C, Cotter D, Khan N, Pollard C, Sheppard P, Varndell I, et al (2001):Glycogen synthase kinase-3beta immunoreactivity is reduced in theprefrontal cortex in schizophrenia. Neurosci Lett 302:117–120.
eaulieu JM, Sotnikova TD, Yao WD, Kockeritz L, Woodgett JR, GainetdinovRR, et al (2004): Lithium antagonizes dopamine-dependent behaviorsmediated by an AKT/glycogen synthase kinase 3 signaling cascade. ProcNatl Acad Sci U S A 101:5099 –5104.
ehrens J, von Kries JP, Kuhl M, Bruhn L, Wedlich D, Grosschedl R, et al (1996):Functional interaction of beta-catenin with the transcription factorLEF-1. Nature 382:638 – 642.
erendse HW, Galis-de Graaf Y, Groenewegen HJ (1992): Topographicalorganization and relationship with ventral striatal compartments of pre-frontal corticostriatal projections in the rat. J Comp Neurol 316:314 –347.
hanot P, Brink M, Samos CH, Hsieh JC, Wang Y, Macke JP, et al (1996): A newmember of the frizzled family from Drosophila functions as a Winglessreceptor. Nature 382:225–230.
rami-Cherrier K, Valjent E, Garcia M, Pages C, Hipskind RA, Caboche J (2002):Dopamine induces a PI3-kinase-independent activation of Akt in striatalneurons: a new route to cAMP response element-binding protein phos-
phorylation. J Neurosci 22:8911– 8921.Brannon M, Gomperts M, Sumoy L, Moon RT, Kimelman D (1997): A beta-catenin/XTcf-3 complex binds to the siamois promoter to regulate dor-sal axis specification in Xenopus. Genes Dev 11:2359 –2370.
Broich K, Grunwald F, Kasper S, Klemm E, Biersack HJ, Moller HJ (1998):D2-dopamine receptor occupancy measured by IBZM-SPECT in relationto extrapyramidal side effects. Pharmacopsychiatry 31:159 –162.
Cadigan KM, Nusse R (1997): Wnt signaling: a common theme in animaldevelopment. Genes Dev 11:3286 –3305.
Chen G, Huang LD, Jiang YM, Manji HK (1999): The mood-stabilizing agentvalproate inhibits the activity of glycogen synthase kinase-3. J Neuro-chem 72:1327–1330.
Cotter D, Kerwin R, al Sarraji S, Brion JP, Chadwich A, Lovestone S, et al (1998):Abnormalities of Wnt signaling in schizophrenia– evidence for neurode-velopmental abnormality. Neuroreport 9:1379 –1383.
De Ferrari GV, Inestrosa NC (2000): Wnt signaling function in Alzheimer’sdisease. Brain Res Brain Res Rev 33:1–12.
Doble BW, Woodgett JR (2003): GSK-3: tricks of the trade for a multi-taskingkinase. J Cell Sci 116:1175–1186.
Emamian ES, Hall D, Birnbaum MJ, Karayiorgou M, Gogos JA (2004): Conver-gent evidence for impaired AKT1-GSK3beta signaling in schizophrenia.Nat Genet 36:131–137.
Gerber DJ, Hall D, Miyakawa T, Demars S, Gogos JA, Karayiorgou M, et al(2003): Evidence for association of schizophrenia with genetic variationin the 8p21.3 gene, PPP3CC, encoding the calcineurin gamma subunit.Proc Natl Acad Sci U S A 100:8993– 8998.
Gerfen CR, Herkenham M, Thibault J (1987): The neostriatal mosaic: II. Patch-and matrix-directed mesostriatal dopaminergic and nondopaminergic sys-tems. J Neurosci 7:3915–3934.
Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR (2001): Pharmacologi-cal studies of prepulse inhibition models of sensorimotor gating deficitsin schizophrenia: a decade in review. Psychopharmacology (Berl) 156:117–154.
Gould TD, Chen G, Manji HK (2004): In vivo evidence in the brain for lithiuminhibition of glycogen synthase kinase-3. Neuropsychopharmacology 29:32–38.
Hamada F, Tomoyasu Y, Takatsu Y, Nakamura M, Nagai S, Suzuki A, et al(1999): Negative regulation of Wingless signaling by D-axin, a Drosoph-ila homolog of axin. Science 283:1739 –1742.
Huber O, Korn R, McLaughlin J, Ohsugi M, Herrmann BG, Kemler R (1996):Nuclear localization of beta-catenin by interaction with transcriptionfactor LEF-1. Mech Dev 59:3–10.
Ichikawa J, Meltzer HY (1999): Relationship between dopaminergic andserotonergic neuronal activity in the frontal cortex and the action oftypical and atypical antipsychotic drugs. Eur Arch Psychiatry Clin Neurosci249 Suppl 4:90 –98.
Ikeda S, Kishida S, Yamamoto H, Murai H, Koyama S, Kikuchi A (1998): Axin, anegative regulator of the Wnt signaling pathway, forms a complex withGSK-3beta and beta-catenin and promotes GSK-3beta-dependentphosphorylation of beta-catenin. EMBO J 17:1371–1384.
Kang UG, Seo MS, Roh MS, Kim Y, Yoon SC, Kim YS (2004): The effects ofclozapine on the GSK-3-mediated signaling pathway. FEBS Lett 560:115–119.
Kapur S, Mamo D (2003): Half a century of antipsychotics and still a central rolefor dopamine D2 receptors. Prog Neuropsychopharmacol Biol Psychiatry 27:1081–1090.
Kapur S, Remington G (2001): Dopamine D(2) receptors and their role inatypical antipsychotic action: still necessary and may even be sufficient.Biol Psychiatry 50:873– 883.
Kapur S, Wadenberg ML, Remington G (2000a): Are animal studies of antip-sychotics appropriately dosed? Lessons from the bedside to the bench.Can J Psychiatry 45:241–246.
Kapur S, Zipursky R, Jones C, Remington G, Houle S (2000b): Relationshipbetween dopamine D(2) occupancy, clinical response, and side effects: adouble-blind PET study of first-episode schizophrenia. Am J Psychiatry157:514 –520.
Katsu T, Ujike H, Nakano T, Tanaka Y, Nomura A, Nakata K, et al (2003): Thehuman frizzled-3 (FZD3) gene on chromosome 8p21, a receptor gene forWnt ligands, is associated with the susceptibility to schizophrenia. NeurosciLett 353:53–56.
Kozlovsky N, Belmaker RH, Agam G (2000): Low GSK-3beta immunoreactiv-ity in postmortem frontal cortex of schizophrenic patients. Am J Psychi-
atry 157:831– 833.www.elsevier.com/locate/biopsych

K
K
K
L
L
L
M
M
M
M
N
N
N
N
O
P
R
542 BIOL PSYCHIATRY 2005;57:533–542 H. Alimohamad et al
w
ozlovsky N, Belmaker RH, Agam G (2001): Low GSK-3 activity in frontalcortex of schizophrenic patients. Schizophr Res 52:101–105.
ozlovsky N, Belmaker RH, Agam G (2002): GSK-3 and the neurodevelop-mental hypothesis of schizophrenia. Eur Neuropsychopharmacol 12:13–25.
ozlovsky N, Nadri C, Belmaker RH, Agam G (2003): Lack of effect of moodstabilizers or neuroleptics on GSK-3 protein levels and GSK-3 activity. IntJ Neuropsychopharmacol 6:117–120.
arson ER, Ariano MA (1995): D3 and D2 dopamine receptors: visualizationof cellular expression patterns in motor and limbic structures. Synapse20:325–337.
esort M, Greendorfer A, Stockmeier C, Johnson GV, Jope RS (1999): Glyco-gen synthase kinase-3beta, beta-catenin, and tau in postmortem bipolarbrain. J Neural Transm 106:1217–1222.
ijam N, Paylor R, McDonald MP, Crawley JN, Deng CX, Herrup K, et al (1997):Social interaction and sensorimotor gating abnormalities in mice lack-ing Dvl1. Cell 90:895–905.
acqueen G, Young T (2003): Cognitive effects of atypical antipsychotics:focus on bipolar spectrum disorders. Bipolar Disord 5 Suppl 2:53– 61.
cIntyre R, Katzman M (2003): The role of atypical antipsychotics in bipolardepression and anxiety disorders. Bipolar Disord 5 Suppl 2:20 –35.
eltzer HY (1999): The role of serotonin in antipsychotic drug action. Neu-ropsychopharmacology 21:106S–115S.
iller JR, Moon RT (1996): Signal transduction through beta-catenin and speci-fication of cell fate during embryogenesis. Genes Dev 10:2527–2539.
adri C, Lipska BK, Kozlovsky N, Weinberger DR, Belmaker RH, Agam G(2003): Glycogen synthase kinase (GSK)-3beta levels and activity in aneurodevelopmental rat model of schizophrenia. Brain Res Dev Brain Res141:33–37.
ishi A, Snyder GL, Nairn AC, Greengard P (1999): Role of calcineurin andprotein phosphatase-2A in the regulation of DARPP-32 dephosphoryla-tion in neostriatal neurons. J Neurochem 72:2015–2021.
oordermeer J, Klingensmith J, Perrimon N, Nusse R (1994): dishevelled andarmadillo act in the wingless signaling pathway in Drosophila. Nature367:80 – 83.
ovak A, Dedhar S (1999): Signaling through beta-catenin and Lef/Tcf. CellMol Life Sci 56:523–537.
’Brien WT, Harper AD, Jove F, Woodgett JR, Maretto S, Piccolo S, et al (2004):Glycogen synthase kinase-3beta haploinsufficiency mimics the behav-ioral and molecular effects of lithium. J Neurosci 24:6791– 6798.
olakis P (1997): The adenomatous polyposis coli (APC) tumor suppressor.Biochim Biophys Acta 1332:F127–F147.
ichelson E (1999): Receptor pharmacology of neuroleptics: relation to clin-ical effects. J Clin Psychiatry 60 Suppl 10:5–14.
ww.elsevier.com/locate/biopsych
Riese J, Yu X, Munnerlyn A, Eresh S, Hsu SC, Grosschedl R, et al (1997): LEF-1,a nuclear factor coordinating signaling inputs from wingless and deca-pentaplegic. Cell 88:777–787.
Robertson GS, Fibiger HC (1992): Neuroleptics increase c-fos expression inthe forebrain: contrasting effects of haloperidol and clozapine. Neuro-science 46:315–328.
Robertson GS, Matsumura H, Fibiger HC (1994): Induction patterns of Fos-like immunoreactivity in the forebrain as predictors of atypical antipsy-chotic activity. J Pharmacol Exp Ther 271:1058 –1066.
Rushlow W, Naus CC, Flumerfelt BA (1996): Somatostatin and the patch/matrix compartments of the rat caudate-putamen. J Comp Neurol 364:184 –190.
Schotte A, Janssen PF, Megens AA, Leysen JE (1993): Occupancy of centralneurotransmitter receptors by risperidone, clozapine and haloperidol,measured ex vivo by quantitative autoradiography. Brain Res 631:191–202.
Serretti A, De Ronchi D, Lorenzi C, Berardi D (2004): New antipsychotics andschizophrenia: a review on efficacy and side effects. Curr Med Chem11:343–358.
Sharma V (2003): Atypical antipsychotics and suicide in mood and anxietydisorders. Bipolar Disord 5 Suppl 2:48 –52.
Siegfried E, Chou TB, Perrimon N (1992): wingless signaling acts throughzeste-white 3, the Drosophila homolog of glycogen synthase kinase-3,to regulate engrailed and establish cell fate. Cell 71:1167–1179.
Svenningsson P, Tzavara ET, Carruthers R, Rachleff I, Wattler S, Nehls M, et al(2003): Diverse psychotomimetics act through a common signalingpathway. Science 302:1412–1415.
van Leeuwen F, Samos CH, Nusse R (1994): Biological activity of solublewingless protein in cultured Drosophila imaginal disc cells. Nature 368:342–344.
Wadenberg ML, Kapur S, Soliman A, Jones C, Vaccarino F (2000): DopamineD2 receptor occupancy predicts catalepsy and the suppression of con-ditioned avoidance response behavior in rats. Psychopharmacology(Berl) 150:422– 429.
Wadenberg ML, Soliman A, VanderSpek SC, Kapur S (2001): Dopamine D(2)receptor occupancy is a common mechanism underlying animal modelsof antipsychotics and their clinical effects. Neuropsychopharmacology25:633– 641.
Williams RS, Cheng L, Mudge AW, Harwood AJ (2002): A common mecha-nism of action for three mood-stabilizing drugs. Nature417:292–295.
Yatham LN (2003): Acute and maintenance treatment of bipolar mania: the
role of atypical antipsychotics. Bipolar Disord 5 Suppl 2:7–19.