antidepressants and the resilience to early-life stress in inbred mouse strains
TRANSCRIPT

Antidepressants and the resilience to early-life stress ininbred mouse strainsElke Binderb*, Karim Malkia*, Jose L. Paya-Canoa, Cathy Fernandesa,Katherine J. Aitchisona, Aleksander A. Mathec, Frans Sluytera
and Leonard C. Schalkwyka
Rationale Selecting an effective treatment for patients
with major depressive disorder is a perpetual problem
for psychiatrists. It is of particular interest to explore the
interaction between genetic predisposition and
environmental factors.
Objectives Mouse inbred strains vary in baseline
performance in depression-related behaviour tests,
which were originally validated as tests of antidepressant
response. Therefore, we investigated interactions between
environmental stress, genotype, and drug response in a
multifactorial behaviour study.
Method Our study design included four inbred mouse
strains (129S1/SvlmJ, C57LB/6J, DBA/2J and FVB/NJ)
of both sexes, two subjected to environmental
manipulations (maternal separation and unpredictable
chronic mild stress) and two representative of treatment
with antidepressants (escitalopram and nortryptiline vs.
vehicle). The mice treated with antidepressants were
further divided into those administered acute (1 day)
and subchronic (14 days) regimes, giving 144 experimental
groups in all, each with at least seven animals. All animals
were tested using the Porsolt forced-swim test (FST) and
the hole-board test.
Results Despite a 24-h maternal separation (MS) or a
14-day unpredictable chronic mild stress protocol, most
animals seemed to be resilient to the stress induced.
One compelling finding is the long-lasting, strain-specific
effect of MS resulting in an increased depression-like
behaviour in the Porsolt FST and elevated anxiety-related
behaviour in the hole-board test seen in 129S1/SvImJ
mice. Nortriptyline was effective in reversing the effect
of MS in the FST in 129S1/SvlmJ male mice.
Conclusion A single 24-h maternal separation of pups
from their mother on postnatal day 9 is a sufficient insult
to result in a depression-like phenotype in adult
129S1/SvImJ mice but not in C57LB/6 J, DBA/2 J,
and FVB/NJ mice. Pharmacogenetics and Genomics
21:779–789 �c 2011 Wolters Kluwer Health | Lippincott
Williams & Wilkins.
Pharmacogenetics and Genomics 2011, 21:779–789
Keywords: antidepressants, depression, escitalopram, Genome-basedTherapeutic Drugs for Depression, maternal deprivation, maternalseparation, nortriptyline, pharmacogenetics
aKing’s College London, Social, Genetic and Developmental Psychiatry Centre,Institute of Psychiatry, London, UK, bINSERM U862, Avenir group Physio-pathology of Energy Balance and Obesity, Universite de Bordeaux 2, Bordeaux,France and cKarolinska Institutet-Clinical Neuroscience, Karolinska University,Hospital Huddinge, Stockholm, Sweden
Correspondence to Leonard C. Schalkwyk, PhD, Social, Genetic andDevelopmental Psychiatry Centre, Institute of Psychiatry, Box P082De Crespigny Park, London SE5 8AF, UKTel: + 44 0 207 848 0279; fax: + 44 0 207 848 0866;e-mail: [email protected]
*Elke Binder and Karim Malki have contributed equally to this study.
Received 10 June 2011 Accepted 23 July 2011
IntroductionThe most popularly prescribed class of antidepressant
drugs is the selective serotonin reuptake inhibitor
(SSRI), represented in this study by the drug escitalo-
pram. Individual response to SSRIs is highly hetero-
geneous, with up to half of the treated patients finding no
relief or only partial relief from their symptoms [1].
Moreover, many patients have reported low tolerabil-
ity [2]. Therefore, a variety of alternative antidepressant
drugs can be prescribed in the hope of increasing efficacy
or tolerance. These include second-generation tricyclics,
such as nortriptyline (a noradrenaline reuptake inhibitor).
The genetic and environmental etiologies of individual
differences moderating the response and tolerance to
common antidepressant medications remain largely un-
known. It is, therefore, important to investigate the
behavioural differences in drug response in connection
with environmental factors and genetic background. This
will lead to better prediction of drug response and a
reduction in the lag time produced by the current trial-
and-error procedure.
Various attempts have been made to generate animal
models of depression [3]. One strategy is to use selectively
bred lines of rats or mice [4,5] and another is to use proto-
cols such as unpredictable chronic mild stress (UCMS)
and maternal separation (MS) to induce depression-
related behaviour in healthy animals.
Supplemental digital content is available for this article. Direct URL citationsappear in the printed text and are provided in the HTML and PDF versions of thisarticle on the journal’s website (www.pharmacogeneticsandgenomics.com).
Original article 779
1744-6872 �c 2011 Wolters Kluwer Health | Lippincott Williams & Wilkins DOI: 10.1097/FPC.0b013e32834b3f35
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

UCMS has been reported to lead to reductions in
reactivity to rewards and to a variety of other depres-
sion-like behaviours in rats and mice [6,7]. In humans, it
is known that the increase in stress in the working
environment correlates with the rising number of patients
who are suffering from depression [8,9].
The dependency of pups on their mother during early
postnatal stages is well documented [10,11], and MS can
result in a variety of long-lasting changes on adult
phenotypes [12–22]. Previous studies have suggested
that a single 24-h separation from the mother during the
first 2 postnatal weeks activates the hypothalamic-
pituitary-adrenal axis system of the neonate and results
in long-lasting consequences, such as stress and anxiety in
adulthood [23,24], whereas mild neonatal stress has been
associated with ‘positive’ effects of resilience [25].
Response to different MS protocols, however, is hetero-
geneous, as recently reported by Millstein and Holmes [26].
The study was unable to produce long-lasting behavioural
changes using either MS or handling. Reasons for this
have been attributed to methodological differences and
also to genetic variation between strains of animals
used [27,28].
This study was designed to investigate strain-specific
differences towards depression-related behaviour and anti-
depressant action using two different models: UCMS
and an MS protocol of one separation period lasting
for 24 h on postnatal day 9 [13]. The hypothesis driving our
approach is that different genetic backgrounds will play a
role in the modulation of environmental and pharmacolo-
gical treatment response in a mouse model of depression.
Materials and methodsAnimals
A total of 1166 mice from four inbred strains were used
for this study. The strains used were C57BL/6J (C57),
DBA/2J (DBA), 129S1/SvImJ (129) and FVB/NJ (FVB).
The animals were specific pathogen free, bred in the
barrier unit at the Institute of Psychiatry (London, UK).
Weaning took place when the animals were 21–28 days
old. They were group housed with same-sex siblings until
they were 8 weeks old. They were then singly housed and
moved to the testing facility. Mice were allowed to
habituate to the individual plastic cages (30.5� 13� 11
cm) and to the testing facility for 2 weeks before testing.
A 12-h light/dark cycle was used with lights on at 08.00 hr
and lights off at 20.00 hr Temperature and humidity
were controlled at 211C ± 21C and 45%, respectively.
Water and food were freely available throughout the ex-
periment. All housing and experimental procedures were
carried out in accordance with the UK Home Office
Animals (Scientific Procedures) Act 1986.
Experimental design
This study adopted a five-way multifactorial design
(Fig. 1). The factors were as follows:
(1) Strain with four levels: C57, DBA, 129, and FVB;
(2) Environment with three levels: MS (described
below), UCMS (described below), and control;
(3) Sex with two levels: male and female;
(4) Drug treatments with three levels: nortriptyline
(4 mg/kg, intraperitoneally), escitalopram (5 mg/kg,
intraperitoneally), or saline (vehicle); and
(5) Treatment duration of drug administration with two
levels: subchronic (14 daily doses) and acute
(single dose).
This resulted in 144 experimental cells with a group size
of seven to eight animals. Group assignment at weaning
was pseudorandom, except for mice undergoing the MS
protocol as entire litters underwent MS; therefore, the
allocation of dams to MS or control was randomized.
Behavioural measurements consisted of the hole-board
test (HBT) used as an arena for general activity, anxiety
and exploration and the Porsolt forced-swim test (FST)
for measuring depression-related behaviour.
Unpredictable chronic mild stress
Starting at the age of 10 weeks, the animals were exposed
to a stressor each day in pseudorandom order. The
stressors in the UCMS regime were: 2 h of home cage
tilting at 451, damp bedding for 4 h, cage switching for
Fig. 1
C57BL/6J
Male
cms
Nortry-ptiline
Chronic Acute
Female Male Female Male Female Male Female
4
8
24
72
144
DBA/2J 129SvemJ FVB/NH
s-cit
msControl
Control
Experimental design. A full factorial design with five factors yielding 144 experimental groups, each with seven or eight animals.
780 Pharmacogenetics and Genomics 2011, Vol 21 No 12
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

2 h, 10 min of flooded cage, reversed light cycle and
airpuff. Each day consisted of one to two stressors applied
at different times of the day (Table 1). The UCMS
regime lasted for a period of 14 days and was concurrent
with the intraperitoneal injections for the animals
assigned to the subchronic treatment groups.
Maternal separation
Litters of each strain were randomly allocated to the MS
group. On postnatal day 9, the dam was removed from the
litter for 24 h. The litter was kept on a heating pad in
their home cage at 331C ± 21C in a different room from
the dam to avoid contact through vocalization. Litters
were always separated and reunited with the mother
during the first half of the light phase. The first hour after
reuniting, the litter with the mother was videotaped.
Litters were of different sizes and, when possible, each
litter came from a different breeding pair (see Table 2,
supplemental digital content 1, http://links.lww.com/FPC/A319and Table 3, supplemental digital content 2, http://links.lww.com/FPC/A320). When the pups grew up, the
littermates were separated into those that would undergo
acute and those that would undergo subchronic treatment
of the antidepressant.
Drugs
Escitalopram and nortriptyline were provided by Lund-
beck (Copenhagen, Denmark). The drugs were dissolved
in saline (Aquapharm, Dunnington, York, UK) every day
fresh before use. The injection volume was 10 ml/kg of
bodyweight; 4 mg/kg of escitalopram, 5 mg/kg of nortrip-
tyline, or the corresponding volume of saline was given.
The doses of the drugs were chosen following data
analysis collected from a pilot study (data not shown).
After 2 weeks of habituation, the animals received one
daily intraperitoneal injection of nortriptyline, escitalo-
pram, or saline for 14 days for the subchronic treatment.
Animals were injected and tested during the light phase.
Blood samples were analysed to investigate the drug
levels and drug metabolism of one animal per strain
per sex per drug (Fig. 8). For the acute treatment, animals
were handled every day (except for weekends) for 14 days
without receiving any drugs. This was done to accustom
the animals to human contact in order to minimize the
stress effect of the acute injection before testing. On the
day of testing, mice received one acute intraperitoneal
injection 30 min before the HBT.
Behavioural test procedure
At the age of 12 weeks, the animals were tested using the
HBT for 5 min, directly followed by the FST for 6 min.
Hole-board test
Mice were tested in a square, evenly lit arena (25.4� 25.4
� 40.6 cm, 300 lux illumination) with clear plastic walls. A
metal floor was used, containing 16 holes (2.2 cm in
diameter), evenly distributed over the floor (4� 4 holes).
The setup was equipped with infrared photocell sensors.
Animals were placed in a corner of the field and allowed
to freely explore for 5 min. The distance travelled (gene-
ral activity), the number of holes visited (exploration)
and the time spent in the centre (anxiety, 17.8� 17.8 cm,
mouse arena E63–10) were monitored by infrared sensor
rings connected to a computer equipped with TruScan
Software Version 2.0 (Coulbourn Instruments, Allentown,
Pennsylvania, USA). Analysis was conducted using R
(Release 2.9–2; R Development Core Team, Vienna,
Austria) and Statistica version 6.1 (StatSoft, Inc., Tulsa,
Oklahoma, USA).
The Porsolt forced-swim test
Directly after the HBT, animals were placed in a clear
Plexiglas cylinder (49-cm high� 15-cm diameter) filled
with water (40-cm high, 211C). The test was conducted
in a similar manner as described elsewhere [29]. Animals
were observed for 6 min and their behaviour was hand-
coded using Ratontime 1.0 (University of Valencia,
Faculty of Psychology). The behaviour during the last
4 min was used for analysis. The behaviour was classified
Table 1 Unpredictable chronic mild stress protocol
Monday Tuesday Wednesday Thursday Friday Saturday Sunday
First weekMorning 11.00 a.m.–
13.00 p.m.,cage tilted at 451(in rack)
10.00 a.m.–14.00p.m.,damp bedding(add 200 ml ofcold water)
13.00 p.m.–15.00 p.m.,cage tilted at451 (in rack)
14.00 p.m.–14.10 p.m.,forced bath in 351 water(approximately 1 cm of waterin an empty cage)
12.00 p.m.–14.00 p.m.,empty cage
Reverselight
Reverselight
Afternoon 15.00 p.m.–16.30 p.m.,empty cages
14.00 p.m.,airpuff (three times/animal)move animals to testing rooms
Second weekMorning 11.00 a.m.–
15.00 p.m.,damp bedding(add 200 ml ofcold water)
10.00 a.m.–12.00p.m.,cage switching(back in originalhomecage)
10.00 a.m.–12.00 p.m.,empty cage
14.00 p.m. –15.00 p.m.,cage tilted 451 (in rack)
14.00 p.m.–14.10 p.m.,forced bath in 351 water (approximately1 cm of water in an empty cage), moveanimals to testing rooms
Reverselight
Reverselight
Afternoon 12.00 p.m.,airpuff (threetimes)
Antidepressants and early life stress Binder et al. 781
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
as climbing (characterized by vertical motion of the front
paws, directed at the walls), swimming (characterized by
horizontal movement), or immobile (defined as the
absence of all movement except that necessary to
maintain balance). The parameter of mobility was the
sum of climbing and swimming.
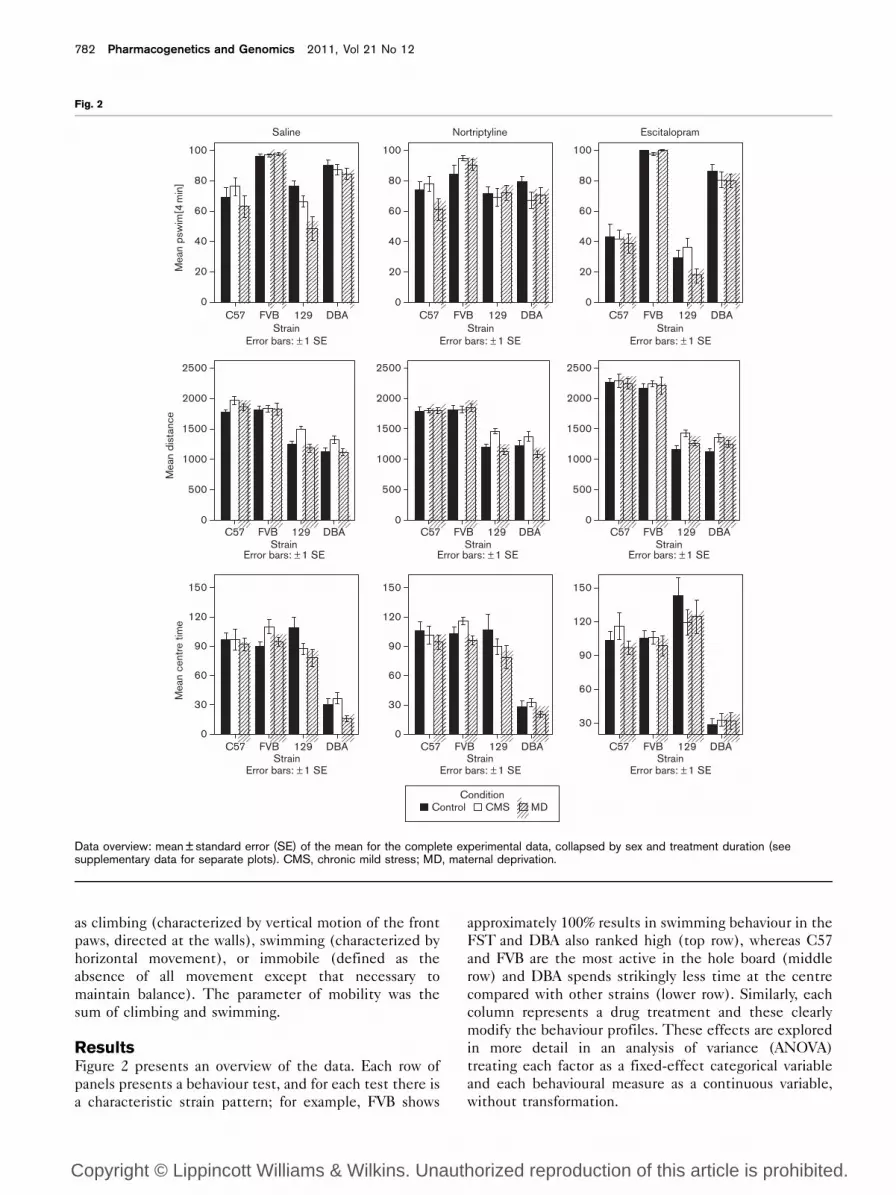
ResultsFigure 2 presents an overview of the data. Each row of
panels presents a behaviour test, and for each test there is
a characteristic strain pattern; for example, FVB shows
approximately 100% results in swimming behaviour in the
FST and DBA also ranked high (top row), whereas C57
and FVB are the most active in the hole board (middle
row) and DBA spends strikingly less time at the centre
compared with other strains (lower row). Similarly, each
column represents a drug treatment and these clearly
modify the behaviour profiles. These effects are explored
in more detail in an analysis of variance (ANOVA)
treating each factor as a fixed-effect categorical variable
and each behavioural measure as a continuous variable,
without transformation.
Fig. 2
100
Saline Nortriptyline Escitalopram
80
60
Mea
n ps
wim
[4m
in]
40
20
0
2500
2000
1500
Mea
n di
stan
ce
1000
500
0
150
120
90
Mea
n ce
ntre
tim
e
60
30
0
150
120
90
60
30
0
150
120
90
60
30
2500
2000
1500
1000
500
0
2500
2000
1500
1000
500
0
100
80
60
40
20
0
100
80
60
40
20
0C57 FVB 129 DBA
Strain
C57 FVB 129 DBAStrain
Error bars: ± 1 SE
Error bars: ± 1 SE
Error bars: ± 1 SE
C57 FVB 129 DBAStrain
Error bars: ± 1 SE
C57 FVB 129 DBAStrain
Error bars: ± 1 SE
Error bars: ± 1 SE Error bars: ± 1 SE
Error bars: ± 1 SE Error bars: ± 1 SE
C57 FVB 129 DBAStrain
C57 FVB 129 DBAStrain
C57 FVB 129 DBAStrain
C57 FVB 129 DBAStrain
C57 FVB 129 DBAStrain
ConditionMDControl CMS
Data overview: mean ± standard error (SE) of the mean for the complete experimental data, collapsed by sex and treatment duration (seesupplementary data for separate plots). CMS, chronic mild stress; MD, maternal deprivation.
782 Pharmacogenetics and Genomics 2011, Vol 21 No 12
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

The Porsolt forced-swim test
Animals were observed for 6 min, and mobility during the
last 4 min was used for analysis. Immobility in the FST
suggests that the animal displays behavioural despair and
is considered to be a depression-related behaviour.
Increased mobility after drug treatment reflects an
antidepressant effect.
A factorial ANOVA revealed a significant strain effect for
both male [F(3,57) = 11.112, P < 0.001] and female
animals [F(3,56) = 16 433, P < 0.001], but no significant
effect for the factor treatment duration or the interaction
strain� treatment duration (Fig. 3). A Bonferroni post-
hoc test indicated a significant difference among all four
strains, but the main effect was largely seen in strain 129
differing from FVB and DBA. However, measurements of
FST showed a ceiling effect in two of the strains: DBA
and FVB. Therefore, parametric assumptions for homo-
geneity of variance across strains were violated and results
must be interpreted with caution.
We further investigated the effect of the pharmacological
and environmental challenges in strain 129. In male
animals, strain 129 showed no significant difference
between subchronic and acute drug administration in
relation to the percent of time spent mobile; therefore,
the factor treatment duration was ignored for the
following calculations. The next series of analysis looked
at the effects of MS in male 129 mice. An independent-
sample t-test confirmed a significant difference between
MS and control [t(30) = 3.13, P < 0.003], with vehicle
MS animals showing decreased mobility compared with
vehicle control 129 mice. However, there was a floor
effect in mobility behaviour, resulting in a negatively
skewed distribution. Therefore, to confirm our findings
we used survival statistics as a distribution-free analysis to
investigate the effect of MS versus control in 129 male
mice treated with saline. The results confirmed a
significant difference between MS and control mice
[log-rank Mantel Cox X2 = 4.190, degree of freedom
(df) = 1, P < 0.041].
Nortriptyline was shown to be effective in reversing the
depressant-like effect of MS on male 129 mice. There
was a significant difference between animals treated
with vehicle and those treated with nortriptyline [t(30) = 27.83, P < 0.001]. The effect of nortriptyline was
specific to MS as the drug did not significantly affect
animals in the control or UCMS group (Fig. 4).
The magnitude of the effect of escitalopram on strain 129
is both striking and pervasive across sexes, treatment
duration and environment. The directionality of the
effect is, however, largely unexpected as it suggests that
escitalopram has a depressogenic rather than an anti-
depressant effect on 129 mice. The effect of escitalopram
on mobility measurements was not dependent on the en-
vironmental manipulations, as this drug reduced mobility
in control mice as much as it did in MS and UCMS mice.
Survival analysis shows that there is no difference
between subchronic and acute treatment in male (Mantel
Cox X2 = 2.774, df = 1, P > 0.09) and female animals
(X2 = 0.670, df = 1, P < 0.410). Overall, the effect of
escitalopram contrasted with the ameliorative effect of
nortiptyline in strain 129.
The Hole-board test
In addition to the nose-poke measure of exploration, we
derived measures of anxiety (time spent in the central
area of the hole board) and activity (distance travelled in
the hole board). To obtain the maximum value from the
testing, we were able to extract an open-field-like
locomotion measure from the hole-board data. Admit-
tedly, this is not a proper open field because of its smaller
size, and there will be an effect of the presence of holes;
however, it is a well-defined experimental measure of
Fig. 3
FST (male mice) FST (female mice)120
(a)
100
80
Per
cent
mob
ile
Per
cent
mob
ile
60
40
20
129m C57m DBAm FVBm
Strain
129f C57f DBAf FVBf
Strain
0
120
100
80
60
40
20
0
(b)
Mean ( ± standard error of the mean) percent mobility of control vehicle (single-housed, saline-injected mice) animals in the forced-swim test (FST)separated by (a) male and (b) female rats shows a significant difference between the four strains (each strain differs from the other). There was nodifference in relation to the duration of treatment. Acute injection = black bars, subchronic injection = white bars (n = 8 mice per group).
Antidepressants and early life stress Binder et al. 783
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

locomotion in an open arena with a different inbred strain
profile from the hole-board nose-poke measure.
Distance travelled
A factorial ANOVA comparing strain� environment in
vehicle-treated animals showed an expected difference
between the strains [F(3,1154) = 341.57, P < 0.01, Fig. 6]
with regard to environment [F(2,1154) = 14.121, P < 0.01]
but no significant strain� environment interaction. It
seems that DBA and 129 mice are less active than FVB
and C57 animals. The results further suggest an increased
activity of mice that underwent a UCMS treatment
compared with control and MS. A strain� drug comparison
shows a significant interaction [F(6,1192) = 5.3173,
P < 0.01], with an escitalopram-induced increase in
activity in C57 animals (P < 0.01). No differences in
activity were seen with nortriptyline (Fig. 5).
To understand whether our behavioural findings in the
FST in strain 129 are independent of any changes in
activity in the HBT, we compared:
(1) Vehicle animals of the control group with animals of
the MS group to investigate whether the environ-
mental manipulation induced a difference in activity.
(2) Control 129 male animals treated with nortriptyline
with vehicle mice to understand whether the drug
per se induces a change in activity.
(3) 129 male animals after MS treated with saline with
nortriptyline-treated animals to confirm that the
reversal of the depression-like behaviour shown in
the FST is not accompanied by a change in general
activity.
None of the three analyses showed a significant effect,
suggesting that the difference seen in the FST is not due
to drug or environment-induced sedation or arousal.
Exploration and anxiety
The number of holes explored by the animals in the HBT
is considered to be a measure of exploratory behaviour.
The time an animal spends at the centre of an open arena
is thought to reflect anxiety, with increasing time spent at
the centre indicative of reduced anxiety. A one-way
ANOVA considering the number of holes explored and
time spent at the centre showed a large strain difference
[number of holes: F(3,1200) = 245.44, P < 0.01; time
spent at the centre: F(3,1200) = 293.37, P < 0.01].
These behaviours had a similar distribution, with strains
that spent less time at the centre exploring fewer holes
(Fig. 6).
Exploration and anxiety are considered to be related
traits, with tests of spontaneous behaviour reflecting a
conflict between exploratory drive versus anxiety/defen-
sive behaviour [30–32]. In this case, there is also
interdependence because the measures are obtained in
the same arena.
A two-way ANOVA on anxiety-related behaviour and
exploration showed a significant drug� strain interaction
[F(6,1192) = 6.4, P < 0.01]. A Bonferroni post-hoc ana-
lysis revealed decreased anxiety-related behaviour in 129
mice after escitalopram treatment. Unexpectedly,
although escitalopram decreased anxiety-related beha-
viour, it also reduced the number of holes explored
(significant factor drug for number of holes explored
[F(2,1192) = 27.432, P < 0.01] compared with saline-
treated mice (Fig. 6).
Fig. 4
100
(a)FST
∗ ∗
80
60
40
20
Saline Nortriptyline Escitalopram
Drug
Saline Nortriptyline Escitalopram
Drug
0
Per
cent
mob
ile
100HBT
∗
∗
150
100
50
0
Tim
e sp
ent i
n th
e ce
nter
(b)
Mean ( ± standard error of the mean) percent mobility in the forced-swim test [FST (a)] and time spent at the centre of the [HBT (b)]. A total of 129male mice show a significant long-lasting depressogenic effect of MS (white bars), which was reversed with nortriptyline treatment in the FST.Nortriptyline had no effect on control (black bars) mice or on unpredictable chronic mild stress UCMS (grey bars) mice. In contrast, escitalopram hada strong depressogenic effect in the FST test (a) but induced a reduction in anxiety-related behaviour in the hole-board test [HBT (b)] independent ofthe stressor applied (n = 16 mice per group).
784 Pharmacogenetics and Genomics 2011, Vol 21 No 12
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

As anxiety-related behaviour is closely connected to
depression, we analysed the anxiety-related behaviour of
129 male mice, comparing environment (MS and control)
with drug (nortriptyline and saline). The results suggest
that MS animals were more anxious than control mice
[F(1,60) = 5.5159, P = 0.02; Fig. 4b].
Drug levels in the brain
A further investigation was conducted to compare drug
levels in the hippocampus of mice. The two drugs used in
this study have different mechanisms of action, which can
interact with strain differences. Drug measurements were
obtained at the time of killing 10 animals of each strain
treated chronically with either escitalopram or nortripty-
line. Measurements of drug levels in the hippocampus
can inform on the average levels of drug that infiltrate the
brain–blood barrier and can be more informative than
dosage quantity alone. The results show that there is no
significant variation in the level of escitalopram across the
four strains; however, there is a differential effect of
nortriptyline, with strains FVB and 129 showing a higher
concentration of drug compared with C57 and DBA
(Fig. 7).
DiscussionOur objective was to test for genetic background and
environmental effects on depression-related behaviour
and response to antidepressants. The behavioural readout
we used was the FST and the HBT in a large factorial
design that we believe is unprecedented. The FST was
developed in rats to model behavioural despair (passivity
in an aversive environment) and has been validated as a
test of antidepressant response [29]. A broad spectrum
of antidepressants reduces immobility in the FST, but for
SSRIs, in particular, this is dependent on careful choice of
experimental details – for example, the effect is seen
only in sufficiently deep water [33,34]. In the original
version of the FST, animals are subjected to the test
twice. The immobility behaviour of the animal could
consequently be interpreted as ‘learned immobility
response,’ and therefore be identified as an adaptive
coping strategy rather than a depression-related beha-
viour [35,36].
As with many other behaviour tests, the FST has more
recently been applied from rats to mice [37]. In mice, a
single-trial version of the test as opposed to the two-trial
paradigm for the rat resulted in an immobility-reducing
effect after administering a variety of antidepressants
Fig. 6
Hole board
140
(a)
120
100
80
Tim
e sp
ent i
n th
e ce
nter
(in
s)
60
40
20
129 C57 DBA FVB
Strain Strain
129 C57 DBA FVB0
50Hole board
40
Num
ber o
f hol
es e
xplo
red
30
20
10
0
(b)
Mean ( ± standard error of the mean) time (s) spent at the centre (a) and number of head dips (b) in the hole board (HB) of control vehicle (blackbars) and control escitalopram (white bars) mice. The anxiety-related measure and the explorative behaviour of vehicle animals showed a similardistribution. Animals that spent more time at the centre (a) explored more holes (b). This common behavioural pattern was disrupted in 129 mice afterescitalopram treatment (n = 32).
Fig. 5
3000
2000
2500 ∗
Activity in the hole board
1000
500
C57 DBA FVB
Strain
129
1500
0
Dis
tanc
e in
cm
Mean ( ± standard error of the mean) activity in the hole-board test(HBT) of control (single housed) mice. Apart from expected straindifferences, no drug-induced activity change was observed, except inC57 animals after escitalopram treatment (n = 32; saline = black bars,escitalopram = white bars and nortriptyline = grey bars).
Antidepressants and early life stress Binder et al. 785
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

[37,38]; however, once again, the effects of SSRIs are
harder to detect [39].
The application of the FST from rats to mice is
experimentally straightforward but requires separate
validation and careful consideration of the distinct biology
of the mouse. Numerous mutations of depression-
relevant genes, including 5HT1a [40], exhibit FST
phenotypes and offer one form of validation.
The most striking feature of the data overview presented
in Fig. 2 is the robust profile of strain differences. This
was expected as the strains were chosen on the basis of
the strain survey by Lucki et al., [41], and subsequent
studies have shown strain-dependent differences regard-
ing the response to stressors [7] and the response to
different antidepressants [42], as well as differences in
the behavioural phenotype of inbred mice [43,44]. Differ-
ent and equally striking strain profiles are seen for the
exploration and anxiety measures of our tests (Fig. 5).
Overall, we found reduced immobility in the notrypty-
line-treated groups, but increased immobility with
escitalopram (Fig. 8).
The escitalopram effect on FST is large and not due to
sedation, as evidenced by the lack of difference in
distance travelled or by exploration in the HBT. To date,
studies have produced contradictory data regarding the
action of escitalopram or citalopram in inbred mice [45].
A recent study reported a positive effect of one acute
dose of citalopram on strain 129/Sv, whereas it showed no
effect on strain DBA/2J [46]. A recent letter in
‘Neuroscience letters’ argued that contradictory results
are often due to the use of the wrong animal model. They
concluded that valid animal models of depression are
‘disease’ animals (in this case the selection bred Flinders
rats) and not stressed healthy animals [47].
Although there is some consensus on experimental
paradigms to measure the action of antidepressants such
as the FST [48], there is less agreement on environ-
mental manipulations leading to a depression-like beha-
vioural abnormality in mice [26,49,50]. How to induce
‘depressed mood’ in a rodent and generate a consistent
and replicable model of depression remains unclear. It is
also a formidable task to make a careful comparison of
available and potential protocols. Above all, we were
concerned that for our pharmacogenomic study we should
use a protocol that will most likely produce a measurable
effect, and so chose to test UCMS and a long MS
protocol.
Ellenbroek and Cools [51] characterized the phenotype
of rats after a 24-h MS as being schizophrenia-like on the
basis of a range of measures including prepulse inhibition.
Fabricius et al., [13] transported this manipulation to mice
and tested 10-week-old C57BL/6j mice in open-field,
elevated-plus and Barnes mazes. They detected a
reduction in anxiety-related behaviour in the elevated
plus maze in MS animals with an increased open arm
time. They also detected decreased behavioural flex-
ibility (perseveration) in the Barnes maze and an effect
on hippocampal neuron number, but did not report the
results of tests of depression-related behaviours. We
found a long-lasting, strain-specific effect of MS on the
Fig. 7
90
80
FST
∗
∗
70
60
50
Per
cent
mob
ile
40
30
20
10
Saline EscitalopramDrug
Nortriptyline0
Shows the mean amount of active drug found in the hippocampus ofanimals at the time of euthanization by strain. No difference wasobserved in the levels of escitalopram across strains but there was adifference in the levels of nortriptyline, with strain 129 and FVB differingfrom C57 and DBA.
Fig. 8
2000
1500
1000
500
0
Mea
n dr
ug le
vels
in b
rain
(mm
ol)
Escitalopram
Antidepressant drug
Nortriptyline
Strain129C57DBAFVB
Mean [ ± standard error (SE) of the mean] mobility in the forced-swimtest (FST) across environmental stressors, sex and treatment durationshows an antidepressant effect of nortriptyline and a depressogenicinfluence of escitalopram (n = 384).
786 Pharmacogenetics and Genomics 2011, Vol 21 No 12
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

FST, with an increased depression-like behaviour seen in
129S1/SvImJ mice. The therapeutic effect of nortripty-
line across strains and sex shows a particularly strong
impact on strain 129 where it was able to reverse the
depressogenic effect of MS (Fig. 3). Overall, nortriptyline
appeared to reduce depression-related behaviour across
strains, sexes and drug administration regimes (Fig. 5).
Resilience or vulnerability to stress is time, strain, sex and
stressor dependent. Recent literature shows that resi-
lience to stress and antidepressant action can be directly
linked to a genetic manipulation in mice. Expression
levels of the serotonin-1A autoreceptor [52], induction of
the transcription factor DFosB [53] and a genetic
variation of the 2-amino-3-(5-methyl-3-oxo-1,2-oxazol-
4-yl)propanoic acid receptor [54] result in changes in
resilience to stress in mice. Furthermore, the same
pathways were necessary to convert SSRI nonresponders
to responders. Susceptibility to stress has not just been
linked to genetic polymorphism but also to dynamic
epigenetic changes. DNA methylation can make mice
more or less responsive to stressors [55,56].
Our results suggest that the choice and length of our
stress protocol, together with the genetic predisposition
of the animals, did not show a long-lasting effect in three
of four strains.
In FVB and DBA, the baseline strain pattern is extreme
enough that there is a ceiling effect on mobility and the
test is uninformative. This may suggest that in strains
known to have high levels of activity, such as FVB, a
longer swimming interval or perhaps a different test
paradigm altogether may have been more appropriate. A
further consideration is with regard to the lack of a normal
distribution of strain 129. The use of survival analysis as a
distribution-free method yielded more rigorous P values,
reinforcing the significance thresholds detected by our
parametric test. The fact that anxiety-related measures
from the HBT confirmed the depression-related results
seen in the FSTadds to the assumption that male animals
of strain 129 after one single 24-h MS have potential to be
models for depression-related behaviour.
The 129 inbred mouse strain consists of 13 different
substrains known to differ in their behavioural pheno-
type [57]. The substrain 129S1/SvlmJ is a popular
background strain for transgenic mice, as embryonic stem
cell lines of that strain are commonly available. To date,
only a few studies have shown interest in the behavioural
phenotype of this strain as it is known to be passive,
hypoactive, to perform poorly in memory tasks and in
stress-related paradigms under certain conditions and
show an absence of the corpus callosum [58–60]. In
recent years, the importance of the choice of background
strain in transgenic animals has become a hot topic [61–63]
and many knockout mice already available have been
generated using 129 embryonic stem cells. In a study
comparing two different backgrounds of null mice, an
increased susceptibility to stress was found in 129/SvEv
compared with C57BL/N mice, which made a difference
in the outcome of the phenotype of the null mouse
models [64]. It is difficult to determine whether
a phenotype observed in a mutant strain is due to the
mutation or the background strain chosen, especially
if the background is a mixture of two strains – for example,
129 for the embryonic stem cells and C57 for the
backcrossing [65].
Although strain 129 has not been extensively used in
studies on depression, the results show that this strain is
more sensitive to both depressogenic protocols and
antidepressant action compared with other commonly
used strains. This strain may prove to be a valuable choice
in future animal studies on depression.
A gene expression study on the hippocampus of mice
prepared in parallel with these has been compared with
the results of a pharmacogenomic genome-wide associa-
tion study using the same drugs. This comparison
produced convergent evidence from mice and humans,
suggesting a role of the Ppm1a gene in response to
nortryptiline but not escitalopram [66].
AcknowledgementsThe Genome-based Therapeutic Drugs for Depression
study was funded by a European Commission Framework
6 Grant, EC Contract Ref.: LSHB-CT-2003–503428.
Lundbeck provided both nortriptyline and escitalopram
free of charge. K. Aitchison has received consultancy fees
and honoraria for participating in expert panels from
pharmaceutical companies including Lundbeck and
GlaxoSmithKline.
Conflicts of interest
There are no conflicts of interest.
References1 Thase ME, Entsuah AR, Rudolph RL. Remission rates during treatment with
venlafaxine or selective serotonin reuptake inhibitors. Br J Psychiatry 2001;178:234–241.
2 Uher R, Farmer A, Maier W, Rietschel M, Hauser J, Marusic A, et al.Measuring depression: comparison and integration of three scales in theGENDEP study. Psychol Med 2008; 38:289–300.
3 El YM, Vaugeois JM. Genetic rodent models of depression. Curr OpinPharmacol 2007; 7:3–7.
4 Overstreet DH, Pucilowski O, Rezvani AH, Janowsky DS. Administrationof antidepressants, diazepam and psychomotor stimulants further confirmsthe utility of flinders sensitive line rats as an animal model of depression.Psychopharmacology (Berl) 1995; 121:27–37.
5 Kromer SA, Kessler MS, Milfay D, Birg IN, Bunck M, Czibere L, et al.Identification of glyoxalase-I as a protein marker in a mouse model ofextremes in trait anxiety. J Neurosci 2005; 25:4375–4384.
6 Cryan JF, Holmes A. The ascent of mouse: advances in modelling humandepression and anxiety. Nat Rev Drug Discov 2005; 4:775–790.
7 Mineur YS, Belzung C, Crusio WE. Effects of unpredictable chronic mildstress on anxiety and depression-like behavior in mice. Behav Brain Res2006; 175:43–50.
8 Stansfeld SA, Fuhrer R, Head J, Ferrie J, Shipley M. Work and psychiatricdisorder in the Whitehall II study. J Psychosom Res 1997; 43:73–81.
9 Lyons DM, Parker KJ, Schatzberg AF. Animal models of early life stress:implications for understanding resilience. Dev Psychobiol 2010; 52:616–624.
Antidepressants and early life stress Binder et al. 787
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

10 Gubernick DJ, Alberts JR. Maternal licking of young: resource exchange andproximate controls. Physiol Behav 1983; 31:593–601.
11 de Kloet ER, Sibug RM, Helmerhorst FM, Schmidt MV. Stress, genes andthe mechanism of programming the brain for later life. Neurosci BiobehavRev 2005; 29:271–281.
12 Anisman H, Zaharia MD, Meaney MJ, Merali Z. Do early-life eventspermanently alter behavioral and hormonal responses to stressors?Int J Dev Neurosci 1998; 16:149–164.
13 Fabricius K, Wortwein G, Pakkenberg B. The impact of maternal separationon adult mouse behaviour and on the total neuron number in the mousehippocampus. Brain Struct Funct 2008; 212:403–416.
14 Franklin TB, Russig H, Weiss IC, Graff J, Linder N, Michalon A, et al.Epigenetic transmission of the impact of early stress across generations.Biol Psychiatry 2010; 68:408–415.
15 George ED, Bordner KA, Elwafi HM, Simen AA. Maternal separation withearly weaning: a novel mouse model of early life neglect. BMC Neurosci2010; 11:123.
16 Van Heerden JH, Russell V, Korff A, Stein DJ, Illing N. Evaluating thebehavioural consequences of early maternal separation in adult C57BL/6mice; the importance of time. Behav Brain Res 2010; 207:332–342.
17 Claessens SE, Daskalakis NP, Van der Veen R, Oitzl MS, de Kloet ER,Champagne DL. Development of individual differences in stressresponsiveness: an overview of factors mediating the outcome of earlylife experiences. Psychopharmacology 2011; 214:141–154.
18 Groenink L, Bijlsma EY, Van Bogaert MJ, Oosting RS, Olivier B. Serotonin1Areceptor deletion does not interact with maternal separation-inducedincreases in startle reactivity and prepulse inhibition deficits.Psychopharmacology (Berl) 2011; 214:353–365.
19 Kessler MS, Bosch OJ, Bunck M, Landgraf R, Neumann ID. Maternal carediffers in mice bred for high versus low trait anxiety: impact of brainvasopressin and cross-fostering. Soc Neurosci 2011; 6:156–168.
20 Mehta M, Schmauss C. Strain-specific cognitive deficits in adult miceexposed to early life stress. Behav Neurosci 2011; 125:29–36.
21 Niwa M, Matsumoto Y, Mouri A, Ozaki N, Nabeshima T. Vulnerability in earlylife to changes in the rearing environment plays a crucial role in theaetiopathology of psychiatric disorders. Int J Neuropsychopharmacol 2011;14:459–477.
22 Weiss IC, Franklin TB, Vizi S, Mansuy IM. Inheritable effect of unpredictablematernal separation on behavioral responses in mice. Front Behav Neurosci2011; 5:3.
23 Levine S. Influence of psychological variables on the activity of thehypothalamic-pituitary-adrenal axis. Eur J Pharmacol 2000; 405:149–160.
24 Pryce CR, Bettschen D, Nanz-Bahr NI, Feldon J. Comparison of theeffects of early handling and early deprivation on conditioned stimulus,context, and spatial learning and memory in adult rats. Behav Neurosci2003; 117:883–893.
25 Macri S, Granstrem O, Shumilina M, Antunes Gomes dos Santos FJ,Berry A, Saso L, Laviola G. Resilience and vulnerability are dose-dependently related to neonatal stressors in mice. Horm Behav 2009;56:391–398.
26 Millstein RA, Holmes A. Effects of repeated maternal separation on anxiety-and depression-related phenotypes in different mouse strains. NeurosciBiobehav Rev 2007; 31:3–17.
27 Neumann ID, Kromer SA, Bosch OJ. Effects of psycho-social stress duringpregnancy on neuroendocrine and behavioural parameters in lactationdepend on the genetically determined stress vulnerability.Psychoneuroendocrinology 2005; 30:791–806.
28 Savignac HM, Dinan TG, Cryan JF. Resistance to early-life stress in mice:effects of genetic background and stress duration. Front Behav Neurosci2011; 4:5–13.
29 Porsolt RD, Anton G, Blavet N, Jalfre M. Behavioural despair in rats: a newmodel sensitive to antidepressant treatments. Eur J Pharmacol 1978;47:379–391.
30 Crawley J, Goodwin FK. Preliminary report of a simple animal behaviormodel for the anxiolytic effects of benzodiazepines. Pharmacol BiochemBehav 1980; 13:167–170.
31 Handley SL, Mithani S. Effects of alpha-adrenoceptor agonists andantagonists in a maze-exploration model of ‘fear’-motivated behaviour.Naunyn Schmiedebergs Arch Pharmacol 1984; 327:1–5.
32 Pellow S, Chopin P, File SE, Briley M. Validation of open:closed arm entriesin an elevated plus-maze as a measure of anxiety in the rat. J NeurosciMethods 1985; 14:149–167.
33 Detke MJ, Lucki I. Detection of serotonergic and noradrenergicantidepressants in the rat forced swimming test: the effects of water depth.Behav Brain Res 1996; 73:43–46.
34 Cryan JF, Valentino RJ, Lucki I. Assessing substrates underlying thebehavioral effects of antidepressants using the modified rat forcedswimming test. Neurosci Biobehav Rev 2005; 29:547–569.
35 Hawkins J, Hicks RA, Phillips N, Moore JD. Swimming rats and humandepression. Nature 1978; 274:512–513.
36 De Pablo JM, Ortiz-Caro J, Sanchez-Santed F, Guillamon A. Effects ofdiazepam, pentobarbital, scopolamine and the timing of saline injectionon learned immobility in rats. Physiol Behav 1991; 50:895–899.
37 Petit-Demouliere B, Chenu F, Bourin M. Forced swimming test in mice:a review of antidepressant activity. Psychopharmacology (Berl) 2005;177:245–255.
38 Borsini F, Meli A. Is the forced swimming test a suitable model forrevealing antidepressant activity? Psychopharmacology (Berl) 1988;94:147–160.
39 Castagne V, Porsolt RD, Moser P. Use of latency to immobility improvesdetection of antidepressant-like activity in the behavioral despair test inthe mouse. Eur J Pharmacol 2009; 616:128–133.
40 Heisler LK, Chu HM, Brennan TJ, Danao JA, Bajwa P, Parsons LH,Tecott LH. Elevated anxiety and antidepressant-like responses inserotonin 5-HT1A receptor mutant mice. Proc Natl Acad Sci USA 1998;95:15049–15054.
41 Lucki I, Dalvi A, Mayorga AJ. Sensitivity to the effects of pharmacologicallyselective antidepressants in different strains of mice. Psychopharmacology(Berl) 2001; 155:315–322.
42 Ibarguen-Vargas Y, Surget A, Touma C, Palme R, Belzung C. Multifacetedstrain-specific effects in a mouse model of depression and of antidepressantreversal. Psychoneuroendocrinology 2008; 33:1357–1368.
43 Voikar V, Koks S, Vasar E, Rauvala H. Strain and gender differences in thebehavior of mouse lines commonly used in transgenic studies. PhysiolBehav 2001; 72:271–281.
44 Jacobson LH, Cryan JF. Feeling strained? Influence of genetic backgroundon depression-related behavior in mice: a review. Behav Genet 2007;37:171–213.
45 Borsini F. Role of the serotonergic system in the forced swimming test.Neurosci Biobehav Rev 1995; 19:377–395.
46 Cervo L, Canetta A, Calcagno E, Burbassi S, Sacchetti G, Caccia S, et al.Genotype-dependent activity of tryptophan hydroxylase-2 determines theresponse to citalopram in a mouse model of depression. J Neurosci 2005;25:8165–8172.
47 Petersen A, Wortwein G, Gruber SH, Mathe AA. Escitalopram reducesincreased hippocampal cytogenesis in a genetic rat depression model.Neurosci Lett 2008; 436:305–308.
48 Cryan JF, Mombereau C. In search of a depressed mouse: utility ofmodels for studying depression-related behavior in geneticallymodified mice. Mol Psychiatry 2004; 9:326–357.
49 Willner P. Chronic mild stress (CMS) revisited: consistency and behavioural-neurobiological concordance in the effects of CMS. Neuropsychobiology2005; 52:90–110.
50 Millstein RA, Ralph RJ, Yang RJ, Holmes A. Effects of repeated maternalseparation on prepulse inhibition of startle across inbred mouse strains.Genes Brain Behav 2006; 5:346–354.
51 Ellenbroek BA, Cools AR. Early maternal deprivation and prepulse inhibition:the role of the postdeprivation environment. Pharmacol Biochem Behav2002; 73:177–184.
52 Richardson-Jones JW, Craige CP, Guiard BP, Stephen A, Metzger KL,Kung HF, et al. 5-HT1A autoreceptor levels determine vulnerability tostress and response to antidepressants. Neuron 14 2010; 65:40–52.
53 Vialou V, Robison AJ, Laplant QC, Covington HE 3rd, Dietz DM, Ohnishi YN,et al. Delta FosB in brain reward circuits mediates resilience to stress andantidepressant responses. Nat Neurosci 2010; 13:745–752.
54 Schmidt MV, Trumbach D, Weber P, Wagner K, Scharf SH, Liebl C, et al.Individual stress vulnerability is predicted by short-term memory andAMPA receptor subunit ratio in the hippocampus. J Neurosci 2010;30:16949–16958.
55 Elliott E, Ezra-Nevo G, Regev L, Neufeld-Cohen A, Chen A. Resilience tosocial stress coincides with functional DNA methylation of the Crf gene inadult mice. Nat Neurosci 2010; 13:1351–1353.
56 Uchida S, Hara K, Kobayashi A, Otsuki K, Yamagata H, Hobara T, et al.Epigenetic status of Gdnf in the ventral striatum determines susceptibilityand adaptation to daily stressful events. Neuron 2011; 69:359–372.
57 Simpson EM, Linder CC, Sargent EE, Davisson MT, Mobraaten LE, Sharp JJ.Genetic variation among 129 substrains and its importance for targetedmutagenesis in mice. Nat Genet 1997; 16:19–27.
58 Crawley JN. Unusual behavioral phenotypes of inbred mouse strains. TrendsNeurosci 1996; 19:181–182.
788 Pharmacogenetics and Genomics 2011, Vol 21 No 12
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.

59 Gerlai R. Gene-targeting studies of mammalian behavior: is it themutation or the background genotype? Trends Neurosci 1996;19:177–181.
60 Crawley JN, Belknap JK, Collins A, Crabbe JC, Frankel W, Henderson N,et al. Behavioral phenotypes of inbred mouse strains: implications andrecommendations for molecular studies. Psychopharmacology (Berl) 1997;132:107–124.
61 Montagutelli X. Effect of the genetic background on the phenotype of mousemutations. J Am Soc Nephrol 2000; 11 (Suppl 16):S101–S105.
62 Fernandes C, Paya-Cano JL, Sluyter F, D’Souza U, Plomin R, Schalkwyk LC.Hippocampal gene expression profiling across eight mouse inbred strains:towards understanding the molecular basis for behaviour. Eur J Neurosci2004; 19:2576–2582.
63 Smith CM, Lawrence AJ, Sutton SW, Gundlach AL. Behavioral phenotypingof mixed background (129S5:B6) relaxin-3 knockout mice. Ann NY AcadSci 2009; 1160:236–241.
64 Peinado JR, Laurent V, Lee SN, Peng BW, Pintar JE, Steiner DF, Lindberg I.Strain-dependent influences on the hypothalamo-pituitary-adrenal axisprofoundly affect the 7B2 and PC2 null phenotypes. Endocrinology 2005;146:3438–3444.
65 Schalkwyk LC, Fernandes C, Nash MW, Kurrikoff K, Vasar E, Koks S.Interpretation of knockout experiments: the congenic footprint. Genes,Brain and Behavior 2007; 6:299–303.
66 Malki K, Uher R, Paya-Cano J, Binder E, Rietschel M, Zobel A, et al.Convergent animal and human evidence suggests a role of PPM1A genein response to antidepressants. Biol Psychiatry 2011; 69:360–365.
Antidepressants and early life stress Binder et al. 789
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.