anticonvulsant effects of focal and intracerebroventricular adenosine on penicillin-induced...
TRANSCRIPT

B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
Anticonvulsant effects of focal and intracerebroventricularadenosine on penicillin-induced epileptiform activity in rats
Mehmet Yildirim⁎, Cafer MarangozDepartment of Physiology, Faculty of Medicine, University of Ondokuz Mayis, 55139 Samsun, Turkey
A R T I C L E I N F O
⁎ Corresponding author. Fax: +90 362 457 6041E-mail address: [email protected] (M
0006-8993/$ – see front matter © 2006 Elsevidoi:10.1016/j.brainres.2006.10.024
A B S T R A C T
Article history:Accepted 17 October 2006Available online 17 November 2006
Adenosine has potent anticonvulsant effects on variousmodels of experimental epilepsy. Inthe present study, we examined the effects of focal and intracerebroventricular (i.c.v.)adenosine on penicillin-induced epileptiform activity in Wistar rats. The effects oftheophylline, a non-selective adenosine receptor antagonist, were also researched. Therecordings of electrocorticogram (ECoG) were carried out by using a data acquisition system,under urethane anesthesia. Adenosine was given in doses of 1, 10 and 100 μg/rat via focaland i.c.v. 30 min after penicillin administration. Theophylline was injected in doses of 1, 10and 100 μg/rat by i.c.v. too. Adenosine administration significantly decreased the spikefrequency while theophylline increased. Focal adenosine is more effective than i.c.v.adenosine. 100 μg adenosine is an effective dose that causes a decrease in epileptiformactivity during experiments. We also demonstrated that 100 μg theophylline significantlyincreased epileptiform activity. Our findings suggest that focal adenosine is more effectivethan i.c.v. adenosine on epileptiform activity.
© 2006 Elsevier B.V. All rights reserved.
Keywords:AdenosineTheophyllineExperimental epilepsyPenicillinRat
Abbreviations:ADK, adenosine kinaseBMI, bicuculline methiodidePTZ, pentylenetetrazoleCNS, central nervous systemDMSO, dimethylsulfoxideECoG, electrocorticogramNMDA, N-methyl-D-aspartateL-PIA, L-phenylisopropyladenosineSWD, spike-wave dischargesWAG/Rij, Wistar Albino Glaxo/Rijswijk
1. Introduction
Adenosine is an important neuromodulator that regulatesneuronal activity through activation of G-protein-coupledspecific receptors classified as A1, A2A, A2B, and A3 receptors(Fredholm et al., 2001). It usually shows inhibitory effects inthe nervous system by decreasing membrane excitability andneurotransmitter release (Fredholm and Dunwiddie, 1988).
.. Yildirim).
er B.V. All rights reserved
Adenosine has been suggested as an endogenous anti-convulsant (Dragunow, 1986). Agonists of adenosine weredemonstrated to display anticonvulsant activity in variousmodels of experimental epilepsy (Dunwiddie andWorth, 1982;Turski et al., 1985). Furthermore, it has been exhibited thatactivations of adenosine A1 and A2A receptors contributed toseizure suppression in electrically kindled and geneticallyepilepsy-prone rats, and audiogenic-seizure-sensitive DBA/2
.

194 B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
mice (Simonato et al., 1994; De Sarro et al., 1999; Huber et al.,2002).
Theophylline is a potent inhibitor of adenosine receptorsat therapeutic concentrations, with antagonism of A1 and A2
receptors, although it is less effective against A3 receptors(Pauwels and Joos, 1995). It is widely used as a bronchodi-lator for the treatment of bronchial asthma and chronicobstructive pulmonary disease (Gaudreault and Guay, 1986).Theophylline not only has a bronchodilating effect but isalso related to epileptic seizures. It has been found that it isproconvulsant in seizures induced by pilocarpine, pentyle-netetrazole (PTZ) and kainic acid (Turski et al., 1985; Ault etal., 1987; Cutrufo et al., 1992). Although many reports havesupported a proconvulsant role for theophylline, a recentstudy has shown that it suppresses the occurrence of spike-wave discharges (SWD) in the genetic model of absenceepilepsy (Ates et al., 2004).
Administrations of adenosine and adenosine kinase (ADK)inhibitors have been reported as beneficial for the inhibition ofepileptic seizures (Boison, 2005). However, cardiovascular sideeffects limit the systemic administration of adenosine or itsagonists (Olsson and Pearson, 1990). Therefore, it has beensuggested that the local delivery of adenosinemay be useful toavoid its systemic side effects (Stein et al., 2000; Nilsen andCock, 2004; Boison, 2005). Indeed, epileptic seizures weresuppressed by the local delivery of adenosine from adenosine-releasing synthetic polymers or adenosine-secreting cellsplaced within the lateral ventricle of kindled rats (Boison etal., 1999, 2002). However, it has been proposed that thesetechniques have several limitations such as: that adenosine isdistributed throughout irrelevant brain regions, rather thanthe seizure focus, and that cell implantation within thehuman brain might require immunosuppression (Anschel etal., 2004). Anschel et al. (2004) have also suggested that focallyinjected adenosine into the rat hippocampus effectivelyprevented bicuculline methiodide (BMI)-induced epileptiformevents. Comparison of two different injection sites is amethodthat is often used to evaluate the efficiencies of antic-onvulsant substances in the different models of experimentalepilepsy (Bağ irici and Marangoz, 2001; Serralta et al., 2006). Toour knowledge, there is no study that was made a comparisonbetween effects of focal (intracortical [i.c.]) and intracerebro-ventricular (i.c.v.) adenosine.
In the present study, we examined the effects of focal andi.c.v. injections of adenosine on penicillin-induced epilepti-form activity in rats. Moreover, the effects of theophyllinewere also tested on this model of experimental epilepsy.
2. Results
Baseline activities of each animal were recorded before theadministration of substances, and it has been confirmed that
Fig. 1 – Representative ECoGs (at 10–22 min) 20 min after the inpenicillin or the injection of other substances. (b) Intracortical inECoG. Saline injection did not change the frequency of penicillin-intracerebroventricularly. (f–h) Focally injected adenosine decreainjected intracerebroventricularly.
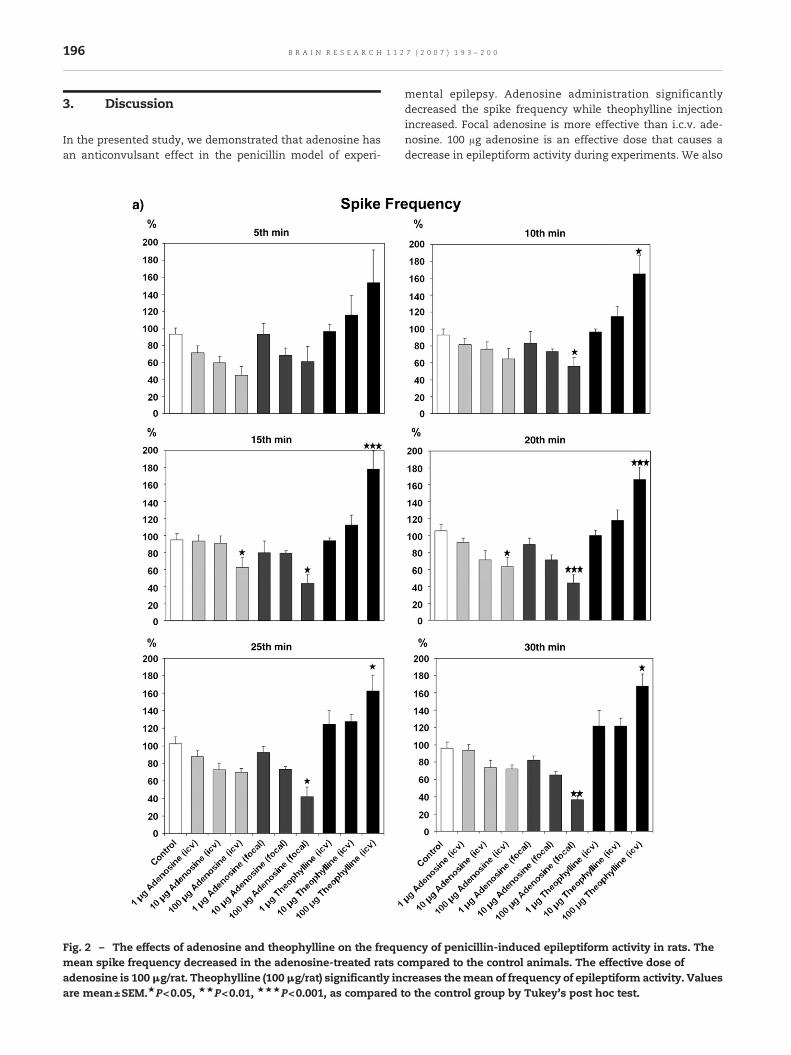
none of the animals had spontaneous epilepsy (Fig. 1). Intra-cortical injection of penicillin (200 IU) induced epileptiformactivity in the all experimental animals (Fig. 1). This epilepti-form activity approximately began 3–5 min after penicillinapplication and continued for 4–5 h without the exception ofpreventive processes with an anticonvulsant substance. Itreached a constant level as a frequency and amplitude in30 min. The mean of spike frequency and amplitude 20 minafter the injection of 0.9% saline in the control rats whichpenicillin pretreated were 29±3 spike/min, 1880±112 μV,respectively (Fig. 2). Following i.c.v. injection of adenosine,the mean spike frequencies at 20th min were 27±2, 21±4 and17±4 spike/min in 1, 10 and 100 μg adenosine administratedrats, respectively. The mean spike amplitudes in these groupswere 2083±235, 1932±204 and 1877±138 μV. The dose of 100 μgadenosine injected via i.c.v. significantly decreased thefrequency of epileptiform activity in between 15 and 20 minafter its injection (p<0.05; Fig. 2), but did not affect the spikeamplitude (no figure). There were no significant differencesbetween the spike frequency or amplitude of saline and theother i.c.v. doses of adenosine. The mean spike frequenciesand amplitudes were found to be 28±3 and 1936± 142 μV, 19±3 and 1863±151 μV, 13±4 and 1437±105 μV after 20min focallyinjection of adenosine into the cortical focus in 1, 10 and 100 μgadenosine groups, respectively. Focally injected 100 μg ade-nosine significantly decreased the mean of spike frequencyduring the experiments. The best effects of focal adenosine(100 μg/rat) appeared between the 20th and 45th min (p<0.05,p<0.01 and p<0.001). This dose of adenosine caused adecrease (30%) in the spike amplitude of epileptiform activity.However, these differences in the spike amplitude were notstatistically significant (no figure). Furthermore, the injectionof 1 and 10 μg adenosine doses affected neither spikefrequency nor spike amplitude. The all adenosine groupscompared to control group, the best effect appeared in focallyinjected 100 μg adenosine group. On the other hand, 100 μgtheophylline (i.c.v.) significantly increased the mean spikefrequency between 10th and 40th min (p<0.05 and p<0.001). 1or 10 μg theophylline did not change the frequency of epilep-tiform activity during the experiments. The mean spikefrequencies were 30±2, 36±2 and 45±3 spike/min at 20 minafter theophylline injection in the 1, 10 and 100 μg theophyl-line groups, respectively (Fig. 2). The spike amplitude was onlyeffected by the injection of 100 μg theophylline. This dose oftheophylline significantly increased the mean spike ampli-tude between the 10th and 25th min (p<0.05; no figure). Themean spike amplitudes were 1940±180, 2104±168 and 2720±203 μV at 20 min after theophylline in the 1, 10 and 100 μgtheophylline groups, respectively. Focal or ventricular injec-tion of saline, adenosine and theophylline did not causeany change on the frequencies or amplitudes of in ECoGactivity respect to control baseline in non-penicillin-injectedanimals.
jections of substances. (a) Baseline ECoG activity beforejection of penicillin (200 IU) induced epileptiform activity oninduced epileptic activity. (c–e) Adenosine was administeredses epileptiform activity in the rat. (i–k) Theophylline was

195B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
196 B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
3. Discussion
In the presented study, we demonstrated that adenosine hasan anticonvulsant effect in the penicillin model of experi-
Fig. 2 – The effects of adenosine and theophylline on the frequmean spike frequency decreased in the adenosine-treated rats cadenosine is 100μg/rat. Theophylline (100μg/rat) significantly inare mean±SEM.★P<0.05, ★★P<0.01, ★★★P<0.001, as compared
mental epilepsy. Adenosine administration significantlydecreased the spike frequency while theophylline injectionincreased. Focal adenosine is more effective than i.c.v. ade-nosine. 100 μg adenosine is an effective dose that causes adecrease in epileptiform activity during experiments. We also
ency of penicillin-induced epileptiform activity in rats. Theompared to the control animals. The effective dose ofcreases themean of frequency of epileptiform activity. Valuesto the control group by Tukey's post hoc test.
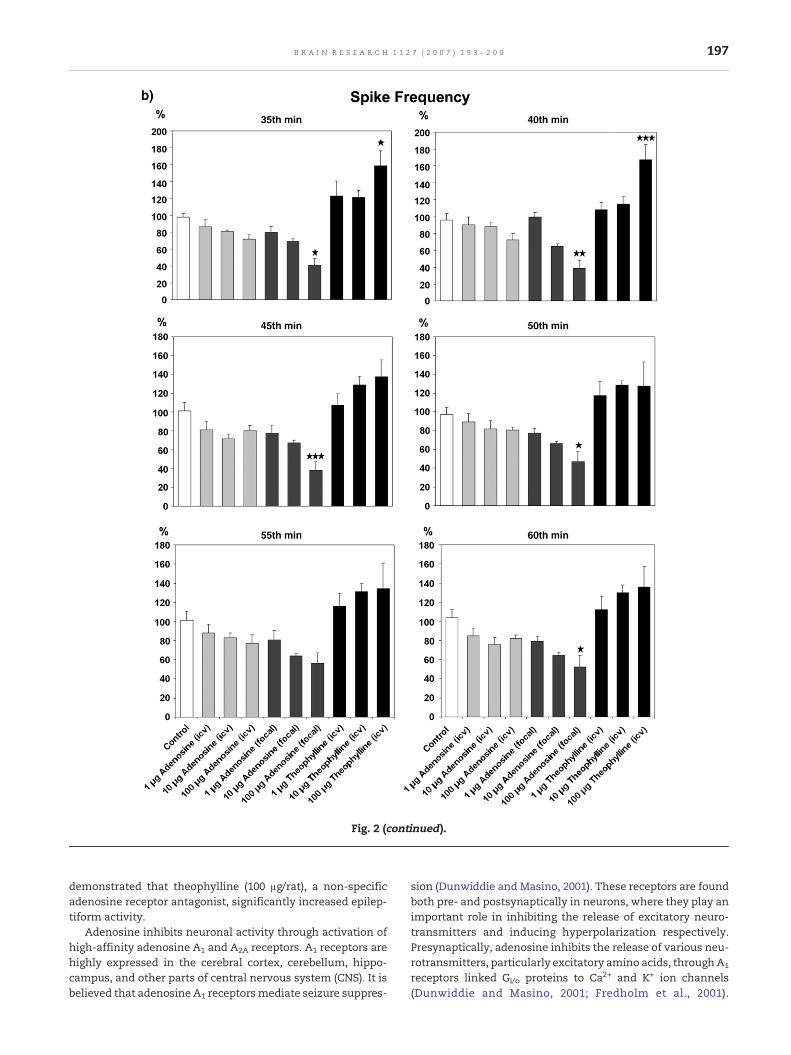
Fig. 2 (continued).
197B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
demonstrated that theophylline (100 μg/rat), a non-specificadenosine receptor antagonist, significantly increased epilep-tiform activity.
Adenosine inhibits neuronal activity through activation ofhigh-affinity adenosine A1 and A2A receptors. A1 receptors arehighly expressed in the cerebral cortex, cerebellum, hippo-campus, and other parts of central nervous system (CNS). It isbelieved that adenosine A1 receptorsmediate seizure suppres-
sion (Dunwiddie and Masino, 2001). These receptors are foundboth pre- and postsynaptically in neurons, where they play animportant role in inhibiting the release of excitatory neuro-transmitters and inducing hyperpolarization respectively.Presynaptically, adenosine inhibits the release of various neu-rotransmitters, particularly excitatory amino acids, throughA1
receptors linked Gi/o proteins to Ca2+ and K+ ion channels(Dunwiddie and Masino, 2001; Fredholm et al., 2001).

198 B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
Postsynaptically, adenosine prevents depolarization by stabi-lization of the Mg2+ blockade of N-methyl-D-aspartate (NMDA)receptors and also reduces postsynaptic Ca2+-influx, probablyby the inhibition of N-type voltage-dependent Ca2+-channels(MacDonald et al., 1986).
In the previous clinical or experimental studies, it has beenshown that adenosine has an anticonvulsant feature. It hasbeen informed that developing and spreading of seizure in ahealthy person's brain may be prevented by the effect of tonicanticonvulsant of endogenic adenosine (25–250 nM) (Dunwiddieand Masino, 2001; Fredholm et al., 2001). In a clinic study,patients who suffer from the complex partial epilepsy which isreplacing the microdialysis probe into hippocampus hasincreased extracellular adenosine 6- to 31-fold during theepileptic activity (During and Spencer, 1992). Furthermore, ithas been informed that hippocampal adenosine levels increaseduring seizures induced by bicuculline (0.5 mg/kg, i.v.), kainicacid (12.0 mg/kg, i.v.) or pentylenetetrazole (100–250 mg/kg, i.p.)and this increase of adenosine may attenuate hippocampalseizure activity in anesthetized rats (Berman et al., 2000).
Adenosine has potent anticonvulsant effects on variousmodels of epilepsy. In amygdala-kindled rats, it has beendemonstrated the endogen adenosine can prevent seizure'sspreading and continuation, but does not influence seizureinitiation from the amygdala (Dragunow et al., 1985). In sameepilepsy model, the adenosine analog L-phenylisopropylade-nosine (L-PIA; 0.5 to 2.0 mg/kg, i.p.) increased the duration ofthe postictal depression but caffeine (32 mg/kg, i.p.) reversedthe prolongation of the postictal depression induced by1.0 mg/kg L-PIA in rats (Rosen and Berman, 1985). In addition,a recent investigation has suggested that the facilitatory roleof the hippocampal CA1 region in relaying or spreading ofseizures is decreased by the activation of adenosine A1
receptors and increased by A2A receptors in piriform cortex-kindled rats (Zeraati et al., 2006). In another model ofexperimental epilepsy on anesthetized rats, adenosine (25 or100 μM) blocked the epileptiform activity induced by digitalis(desacetyl lanatocid C; 1, 2, or 4 μg, i.c.) and the epileptogeniceffect of desacetyl lanatocid C was enhanced by 1 mMtheophylline (Kesim et al., 1994). Furthermore, the epilepti-form activity induced by bicuculline has suppressed theadministration of adenosine and its analogs in the rathippocampal slices (Ault and Wang, 1986). On the otherhand, it has been demonstrated that theophylline, a non-specific antagonist of adenosine receptor, has the feature of aconvulsant or proconvulsant (Glenn et al., 1995).
The results of the present study are consistent with theabove-mentioned findings related to the anticonvulsant effectof adenosine. The effect of adenosine has also been investi-gated in genetically epilepsy-prone animals. For instance, DeSarro et al. (1999) have suggested that stimulation of adenosineA1 and A2A receptors is involved in the suppression of seizuresin audiogenic-seizure-sensitive DBA/2 mice. However, it hasbeen found in another study performed on the genetic modelof epilepsy that adenosine (30 and 120 mg/kg, i.p.) raisedabsence seizures in genetically epileptic Wistar Albino Glaxo/Rijswijk (WAG/Rij) rats (Ilbay et al., 2001). Adenosine alsoinduced a decrease in both blood pressure and body tempera-ture. Ilbay et al. (2001) have suggested that the peripheralinjection of adenosine may increase absence seizures by
contributing to the facilitation of spike-wave discharge(SWD). Indeed, it has been reported that peripheral adminis-tration of adenosine caused some systemic side effects.Especially cardiovascular side effects limit the systemicadministration of adenosine receptor agonists (Olsson andPearson, 1990). When administered systemically, adenosineand its analogues cause powerfully adverse effects rangingfrom sedation and hypothermia to the suppression of cardi-ovascular functions and an almost complete cessation ofspontaneous motor activity (Dunwiddie, 1999). Therefore, itseems sensible that adenosine and its agonists given directlyinto the brain. Indeed, seizure suppression by the local deliveryof adenosine from intraventricular implants of adenosine-releasing synthetic polymers has been established in the ratkindlingmodel of epilepsy (Boison et al., 1999). However, it hasrecently been suggested in another study that this techniquehas several limitations, such as the distribution of drugsthroughout the brain and cerebrospinal fluid, rather thanbeingconcentrated at the seizure focus and cell implantation withinthe human brain,might require immunosuppression (Anschelet al., 2004). Therefore, adenosine injection directly onto aseizure focus in the rat hippocampus prevented BMI-inducedepileptiform events (Anschel et al., 2004). However, there is nostudy that compare the effects of focal and i.c.v. adenosine. Inthe present study we comparatively investigated the effects offocal and ventricularly administered adenosine on the peni-cillin-induced epileptiform activity in rats. Our findingssuggest that focal adenosine is more effective than i.c.v.adenosine. Focally injected adenosine also suppresses epilep-tiform activity for more rapidly and longer time compared to i.c.v. administration.
4. Experimental procedures
4.1. Animals
Experiments were performed on 60 adult female Wistar ratsweighing between 180 and 220 g. Animalswere housedunder a12:12h light–dark cycle (light on07:00h) and roomtemperatureof 20±2 °C. All experimentswere performed between 10:00 and16:00 h. They were given free access to food and water. Everyeffort wasmade tominimize animal suffering and the numberof animals used. Experimental procedures were approved bythe local Ethical Committee. All experiments were carried outaccording to local guidelines for the care and use of laboratoryanimals and the guidelines of the European CommunityCouncil for experimental animal care.
4.2. Chemicals and experiment groups
Adenosine, theophylline and urethane were purchased fromSigma (Saint Louis, MO, USA). To form an epileptiform activity,in 1 μl volume of 200 IU penicillin G potassium was givenintracortically. Adenosine was dissolved in 20% dimethylsulf-oxide (DMSO), and given in doses of 1, 10 and 100 μg/rat viafocal and i.c.v. 30 min after penicillin administration. Theo-phyllinewasdissolved in 0.9% saline and injecteddoses of 1, 10and 100 μg/rat by i.c.v. Control animals were administrated viai.c.v. 0.9% saline 30 min after the penicillin injection. Each

199B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
substance was injected in a total volume of 5 μl into the cortexor lateral ventricle of the animal. Sixty animals were equallydivided into 10 experimental groups: (1) 0.9% saline, (2) 1 μgadenosine (i.c.v.), (3) 10 μg adenosine (i.c.v.), (4) 100 μg adeno-sine (i.c.v.), (5) 1 μg adenosine (focal), (6) 10 μg adenosine (focal),(7) 100 μg adenosine (focal), (8) 1 μg theophylline (i.c.v.), (9) 10 μgtheophylline (i.c.v.), and (10) 100 μg theophylline (i.c.v.).
Furthermore, 12 rats were divided into four groups ofthree animals each. These animalswere given only 0.9% saline(i.c.v.), 100 μg adenosine (i.c.v.), 100 μg adenosine (focal) and100 μg theophylline (i.c.v.) without penicillin pretreatment.They were not included into the total number of experi-mental animals.
4.3. Surgical procedure and induction of epileptiformactivity
Rats were anesthetized with an intraperitoneal injection ofurethane (1.2 g/kg) and additional doses were given as needed.The left cerebral cortex was carefully exposed by craniotomy.After incision of the skull, the head of the animalwas placed ina stereotaxic apparatus (Harvard Instruments, South Natick,MA, USA). Four different corners of the scalp were stitched bysurgical threads and stretched in order to form a liquidvaseline pool (37 °C). Body temperature was monitored usinga rectal probe and maintained at 37 °C with a homeothermicblanket system (Harvard Homoeothermic Blanket, USA). Apolyethylene cannula was placed into the right femoral arteryto monitor blood pressure which was kept above 100 mm Hgduring the experiments (mean 114±10 mm Hg). All contactand incision points were infiltrated with procaine hydro-chloride to minimize possible sources of pain.
The epileptic activity was produced by intracortical injec-tion of 200 IU/1 μl penicillin G (Yildirim and Marangoz, 2006).The penicillin was injected into the left sensorimotor cortex,1 mm beneath the brain surface using a Hamilton microsyr-inge (type 701N, Hamilton Co., Reno, NV, USA). The coordi-nates used for i.c. injection, with the bregma point as thereference, were: AP=−2 mm, L=3 mm. Focal doses ofadenosine were also administrated intracortically.
Adenosine, theophylline and saline were injected in thelateral ventricle. Thirty minutes after penicillin injection themicrosyringe was inserted 2.5 mm beneath the brain surfaceand these substances were injected stereotactically into theleft lateral ventricle. The coordinates were used for lateralventricle, with the bregma point as the reference, were: AP=−0.8 mm, L=1.5 mm. The injection rate was 5 μl/min and theneedle was left in place for 1 min following the infusion.
4.4. Electrophysiological recordings
Two Ag–AgCl ball electrodes were placed over the left somato-motor cortex (electrode coordinates: first electrode, 2 mmlateral to sagittal suture and 1 mm anterior to bregma; secondelectrode, 2 mm lateral to sagittal suture 5 mm posterior tobregma). The common reference electrode was fixed on theright pinna. The recordings of electrocorticogram (ECoG) werecarried out using a data acquisition system (PowerLab 4/SP, ADInstruments, Australia). The signals from the electrodes wereamplified and filtered (0.1–50 Hz bandpass) using BioAmp
amplifiers (AD Instruments, Australia). Then the ECoG signalwas digitized at a sampling rate of 1024 using the PowerLab4/SP. The digitized brain signal was displayed and stored on apersonal computer (PC). The frequency and amplitude ofepileptic activitywere analyzedoffline (Ayyildiz et al., 2006a,b).
4.5. Statistical analyses
Spike frequencies and amplitudes for each animal were auto-matically calculated and measured using the Chart v.5.1.1(PowerLab software). These frequency and amplitude valuesacquired from animals in all groups were converted to scalingpercentage ina time-dependentmanner. Theobtainedpercentchanges were used for statistical analyses and graphics.Statistical analyses were carried out by one-way analysis ofvariance (ANOVA), followed by Tukey's post hoc test to correctfor multiple comparisons of treatments. Data are expressed asmean±SEM. The significance level was p<0.05.
R E F E R E N C E S
Anschel, D.J., Ortega, E.L., Kraus, A.C., Fisher, R.S., 2004. Focallyinjected adenosine prevents seizures in the rat. Exp. Neurol.190, 544–547.
Ates, N., Sahin, D., Ilbay, G., 2004. Theophylline, a methylxanthinederivative, suppresses absence epileptic seizures in WAG/Rijrats. Epilepsy Behav. 5, 645–648.
Ault, B., Wang, C.M., 1986. Adenosine inhibits epileptiformactivity arising in hippocampal area CA3. Br. J. Pharmacol. 87,695–703.
Ault, B., Olney, M.A., Joyner, J.L., Boyer, C.E., Notrica, M.A., Soroko,F.E., Wang, C.M., 1987. Pro-convulsant actions of theophyllineand caffeine in the hippocampus: implications for themanagement of temporal lobe epilepsy. Brain Res. 426, 93–102.
Ayyildiz, M., Yildirim, M., Agar, E., 2006a. The effects of vitamin Eon penicillin-induced epileptiform activity in rats. Exp. BrainRes. 174, 109–113.
Ayyildiz, M., Yildirim, M., Agar, E., Baltaci, A.K., 2006b. The effect ofleptin on penicillin-induced epileptiform activity in rats. BrainRes. Bull. 68, 374–378.
Bağ irici, F., Marangoz, C., 2001. Proconvulsant effects of centraland peripheral administration of L-NAME on penicillin-induced epilepsy in rats. Neurosci. Res. Commun. 28, 107–114.
Berman, R.F., Fredholm, B.B., Aden, U., O'Connor, W.T., 2000.Evidence for increased dorsal hippocampal adenosine releaseand metabolism during pharmacologically induced seizures inrats. Brain Res. 872, 44–53.
Boison, D., 2005. Adenosine and epilepsy: from therapeuticrationale to new therapeutic strategies. Neuroscientist 11,25–36.
Boison, D., Scheurer, L., Tseng, J.L., Aebischer, P., Mohler, H., 1999.Seizure suppression in kindled rats by intraventricular graftingof an adenosine releasing synthetic polymer. Exp. Neurol. 160,164–174.
Boison, D., Huber, A., Padrum, V., Deglon, N., Aebischer, P., Möhler,H., 2002. Seizure suppression by adenosine-releasing cells isindependent of seizure frequency. Epilepsia 43, 788–796.
Cutrufo, C., Bortot, L., Giachetti, A., Manzini, S., 1992. Differentialeffects of various xanthines on pentylenetetrazole-inducedseizures in rats: an EEG and behavioural study. Eur. J.Pharmacol. 222, 1–6.
De Sarro, G., De Sarro, A., Di Paola, E.D., Bertorelli, R., 1999. Effectsof adenosine receptor agonists and antagonists on audiogenicseizure-sensible DBA/2 mice. Eur. J. Pharmacol. 371, 137–145.

200 B R A I N R E S E A R C H 1 1 2 7 ( 2 0 0 7 ) 1 9 3 – 2 0 0
Dragunow, M., 1986. Endogenous anticonvulsant substances.Neurosci. Biobehav. Rev. 10, 229–244.
Dragunow, M., Goddard, G.V., Laverty, R., 1985. Is adenosine anendogenous anticonvulsant? Epilepsia 26, 480–487.
Dunwiddie, T.V., 1999. Adenosine and suppression of seizures.Adv. Neurol. 79, 1001–1010.
Dunwiddie, T.V., Masino, S.A., 2001. The role and regulation ofadenosine in the central nervous system. Annu. Rev. Neurosci.24, 31–55.
Dunwiddie, T.V., Worth, T., 1982. Sedative and anticonvulsanteffects of adenosine analogs in mouse and rat. J. Pharmacol.Exp. Ther. 220, 70–76.
During, M.J., Spencer, D.D., 1992. Adenosine: a potential mediatorof seizure arrest and postictal refractoriness. Ann. Neurol. 32,618–624.
Fredholm, B.B., Dunwiddie, T.V., 1988. How does adenosineinhibit transmitter release? Trends Pharmacol. Sci. 9,130–134.
Fredholm, B.B., Ijzerman, A.P., Jacobson, K.A., Klotz, K.N., Linden,J., 2001. International union of pharmacology: XXV.Nomenclature and classification of adenosine receptors.Pharmacol. Rev. 53, 527–552.
Gaudreault, P., Guay, J., 1986. Theophylline poisoning:pharmacological considerations and clinical management.Med. Toxicol. 1, 169–191.
Glenn, G.M., Krober, M.S., Kelly, P., McCarty, J., Weir, M., 1995.Pyridoxine as therapy in theophylline-induced seizures.Vet. Hum. Toxicol. 37, 342–345.
Huber, A., Guttinger, M., Mohler, H., Boison, D., 2002. Seizuresuppression by adenosine A(2A) receptor activation in a ratmodel of audiogenic brainstem epilepsy. Neurosci. Lett. 329,289–292.
Ilbay, G., Sahin, D., Karson, A., Ates, N., 2001. Effects of adenosineadministration on spike-wave discharge frequency ingenetically epileptic rats. Clin. Exp. Pharmacol. Physiol. 28,643–646.
Kesim, Y., Marangoz, C., Ayyildiz, M., Tasci, N., Agar, E., Sahinoglu,H., 1994. The effects of the purinergic system on
digitalis-induced epileptiform activity. J. Basic Clin. Physiol.Pharmacol. 5, 167–178.
MacDonald, R.L., Skerritt, J.H., Werz, M.A., 1986. Adenosineagonists reduce voltage-dependent calcium conductance ofmouse sensory neurones in cell culture. J. Physiol. 370, 75–90.
Nilsen, K.E., Cock, H.R., 2004. Focal treatment for refractoryepilepsy: hope for the future? Brain Res. Brain Res. Rev. 44,141–153.
Olsson, R.A., Pearson, J.D., 1990. Cardiovascular purinoceptors.Phys. Rev. 70, 761–845.
Pauwels, R.A., Joos, G.F., 1995. Characterization of the adenosinereceptors in the airways. Arch. Int. Pharmacodyn. Ther. 329,151–160.
Rosen, J.B., Berman, R.F., 1985. Prolonged postictal depression inamygdala-kindled rats by the adenosine analog,L-phenylisopropyladenosine. Exp. Neurol. 90, 549–557.
Serralta, A., Barcia, J.A., Ortiz, P., et al., 2006. Effect ofintracerebroventricular continuous infusion of valproic acidversus single i.p. and i.c.v. injections in the amygdala kindlingepilepsy model. Epilepsy Res. 70, 15–26.
Simonato, M., Varani, K., Muzzolini, A., Bianchi, C., Beani, L., Borea,P.A., 1994. Adenosine A1 receptors in the rat brain in thekindling model of epilepsy. Eur. J. Pharmacol. 265, 121–124.
Stein, A.G., Eder, H.G., Blum, D.E., Drachev, A., Fisher, R.S., 2000. Anautomated drug delivery system for focal epilepsy. EpilepsyRes. 39, 103–114.
Turski, W.A., Cavalheiro, E.A., Ikonomidou, C., Mello, L.E.,Bortolotto, Z.A., Turski, L., 1985. Effects of aminophylline and2-chloroadenosine on seizures produced by pilocarpine in rats:morphological and electroencephalographic correlates. BrainRes. 361, 309–323.
Yildirim, M., Marangoz, C., 2006. Anticonvulsant effects ofmelatonin on penicillin-induced epileptiform activity in rats.Brain Res. 1099, 183–188.
Zeraati, M., Mirnajafi-Zadeh, J., Fathollahi, Y., Namvar, S., Rezvani,M.E., 2006. Adenosine A1 and A2A receptors of hippocampal CA1
region have opposite effects on piriform cortex kindledseizures in rats. Seizure 15, 41–48.