amino acid modifications on trna
TRANSCRIPT

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 539
Amino acid modifications on tRNAActa Biochim Biophys Sin (2008): 539-553 | © 2008 Institute of Biochemistry and Cell Biology, SIBS, CAS | All Rights Reserved 1672-9145http://www.abbs.info; www.blackwellpublishing.com/abbs | DOI: 10.1111/j.1745-7270.2008.00435.x
Amino acid modifications on tRNA†
Jing Yuan1#, Kelly Sheppard1#, and Dieter Söll1,2*1Department of Molecular Biophysics and Biochemistry and 2Department of Chemistry, Yale University, New Haven, CT 06520-8114, USA
The accurate formation of cognate aminoacyl-transfer RNAs(aa-tRNAs) is essential for the fidelity of translation. Mostamino acids are esterified onto their cognate tRNAisoacceptors directly by aa-tRNA synthetases. However, inthe case of four amino acids (Gln, Asn, Cys and Sec),aminoacyl-tRNAs are made through indirect pathways inmany organisms across all three domains of life. The processbegins with the charging of noncognate amino acids to tRNAsby a specialized synthetase in the case of Cys-tRNACys
formation or by synthetases with relaxed specificity, such asthe non-discriminating glutamyl-tRNA, non-discriminatingaspartyl-tRNA and seryl-tRNA synthetases. The resultingmisacylated tRNAs are then converted to cognate pairsthrough transformation of the amino acids on the tRNA,which is catalyzed by a group of tRNA-dependent modifyingenzymes, such as tRNA-dependent amidotransferases,Sep-tRNA:Cys-tRNA synthase, O-phosphoseryl-tRNAkinase and Sep-tRNA:Sec-tRNA synthase. The majority ofthese indirect pathways are widely spread in all domains oflife and thought to be part of the evolutionary process.
Keywords aminoacyl-tRNA; indirect pathways; tRNA-dependent amidotransferase; tRNA-dependent cysteinebiosynthesis; selenocysteine biosynthesis
In translation, aminoacyl-transfer RNAs (aa-tRNAs) areemployed to convert genetic information stored inmessenger RNA sequences to the three-dimensionalinformation manifested in the resulting proteins.Aminoacyl-tRNA synthetases (aaRSs) play a crucial rolein maintaining the fidelity of translation by matching each
standard amino acid found in proteins to the correspondingtRNA isoacceptors and forming a cognate aa-tRNA pair.The aminoacylation reaction is carried out as a two-stepprocess [1]:1 ATP+aa+aaRS → aaRS:aa~AMP+PPi
2 tRNA+aaRS:aa~AMP → aaRS+aa-tRNA+AMPThe first step is the activation of an amino acid with
ATP. The aaRSs produce an aminoacyl adenylate byattaching the carboxyl group in the amino acid to thephosphoryl group of AMP. In the second step, the activatedamino acid is transferred to the 2’ or 3’ hydroxyl groupof the 3’ terminal ribose moiety of tRNA and followed byrelease of the final product, aa-tRNA. In the classical view,20 aaRSs catalyze the formation of 20 different aa-tRNApairs. Each synthetase specifically recognizes a set oftRNA isoacceptors and charges them with the correctamino acid that corresponds to the anticodons of the tRNAmolecules.
The first exception to this one synthetase/one set oftRNAs/one amino acid rule was discovered 40 years ago[2], when it was shown that Bacillus Gln-tRNAGln issynthesized from Glu-tRNAGln rather than from directacceptance of Gln on tRNAGln. Thirty years later, the natureof the enzymes catalyzing such tRNA-dependent aminoacid transformations was uncovered [3]. With advancesin functional genomics as well as in biochemical andgenetic analyses, the indirect pathways for Gln-tRNAGln,Asn-tRNAAsn, Cys-tRNACys and Sec-tRNASec formationhave been characterized [4−7]. They all require two typesof enzymes: aaRSs, which can form misacylatedintermediates, and tRNA-dependent amino acid-modifyingenzymes, which convert tRNA-bound amino acid to formthe cognate aa-tRNA pair. Organisms that posses one ormore of these indirect pathways do not have to encodethe full set of 20 aaRSs [5,8−13], once thought to beessential for all living species.
The occurrence of tRNA-dependent amino acidtransformations is surprisingly widespread throughout allthree domains of life (Table 1). All known archaea [5],
Review
Received: March 26, 2008 Accepted: April 15, 2008†This work is dedicated to late Prof. Ying-Lai Wang who during the1960s and 1970s performed an invaluable service to the continuedpractice of biochemistry in China with his research on the chemicalsynthesis of alanine tRNAThis work was supported by grants from the Department of Energy (DE-FGO2-98ER20311), the National Institute of General Medical Sciences(GM22854) and the National Science Foundation (MCB-0645283)# These authors contributed equally to this work*Corresponding author: Tel, 1-203-432-6204; Fax, 1-203-432-6202;E-mail, [email protected]

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 540
Amino acid modifications on tRNA
most bacteria [8], and chloroplasts [14,15] encode theindirect pathway for Gln-tRNAGln formation. A A non-discriminating GluRS (ND-GluRS) aminoacylates tRNAGln
with Glu [16,17]. A Glu-tRNAGln amidotransferase (Glu-AdT) then recognizes the mischarged species andtransforms it to Gln-tRNAGln in the presence of ATP andan amide donor [Fig. 1(A)] [5]. Likewise, for Asnformation, Asp is first ligated to tRNAAsn by a ND-AspRS[17,18] and then converted to Asn on the tRNA by anAsp-tRNAAsn amidotransferase (Asp-AdT) [Fig. 1(B)] [18,19]. The ND-AspRS/Asp-AdT pathway is present in mostbacteria and archaea [5,8,9]. In a large subset ofeuryarchaeota, a Cys-tRNACys is formed via an O-phosphoseryl-tRNACys (Sep-tRNACys) intermediatecatalyzed by a noncanonical synthetase, SepRS. The tRNAbound O-phosphoserine (Sep) is then converted to Cysby SepCysS in the presence of a sulfur donor (Fig. 2)[6]. Selenocysteine formation pathways are exclusivelytRNA-dependent and found in all Sec-decoding organisms
aa-tRNA Prevalence ReferenceIndirect pathway Direct pathway
Gln-tRNAGln All known archaea, most bacteria, All known eukaryotes and a minority of [5,8,14,15]and chloroplasts bacteria
Asn-tRNAAsn Most bacteria and archaea All known eukaryotes and a number of [5,8,9,24]bacteria and archaea
Cys-tRNACys Methanogenic archaea (except M. smithii All known eukaryotes and bacteria, and [6,10−12,62,67−69]and M. stadtmanae) and A. fulgidus most archaea
Sec-tRNASec All known Sec-decoding eukaryotes, None known [99−101,111]archaea and bacteria
Table 1 Prevalence of the indirect and direct pathways for the synthesis of the aa-tRNA species listed in all three domains of life
Adapted from Sheppard et al [132], with permission from Oxford University Press.
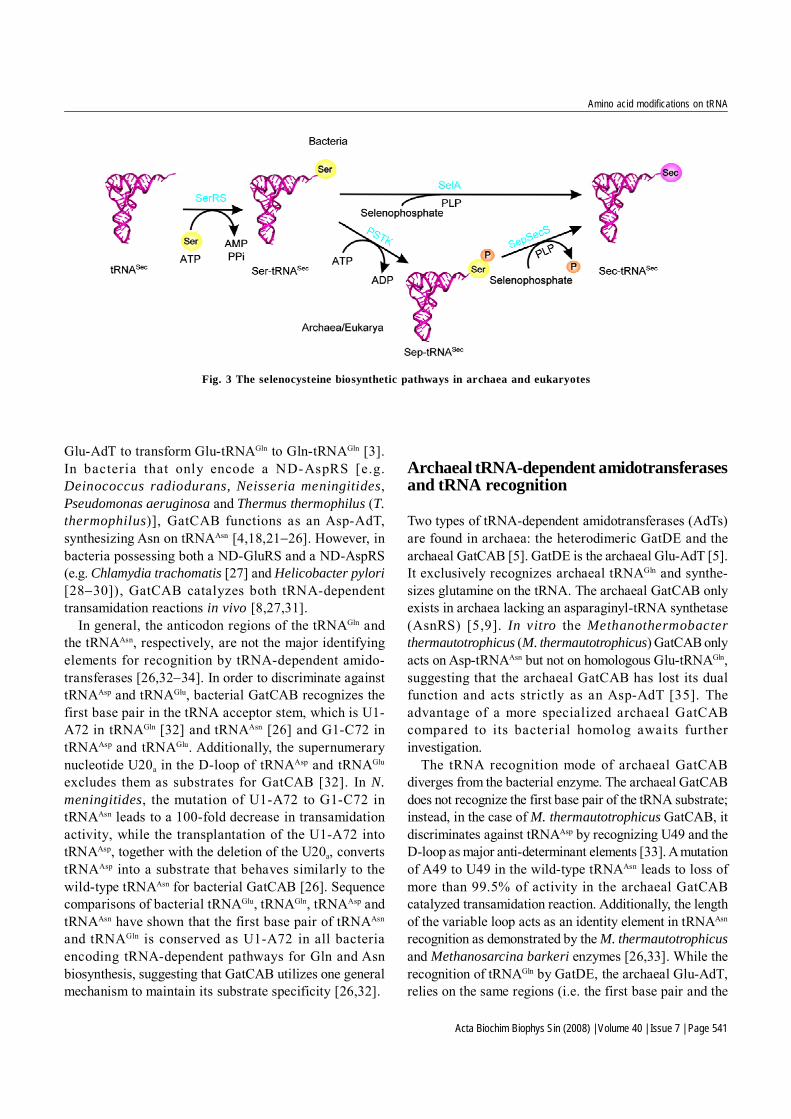
[13]. In archaea and eukaryotes, it is a multistep processinvolving SerRS, PSTK and SepSecS (Fig. 3) [7,20]. Seris esterified onto tRNASec by SerRS and then phosphorylatedby PSTK on the tRNA forming Sep, which is furtherconverted to Sec by SepSecS. The archaeal and eukaryoticpathway is different from the bacterial one, where themischarged Ser-tRNASec is directly converted to Sec-tRNASec by selenocysteine synthase (SelA) [13]. In thisreview, we summarize the latest progress in characterizingthe enzymes involved in tRNA-dependent amino acidtransformations and the current evolutionary views of thesepathways.
Bacterial tRNA-dependent Amidotransferaseand tRNA Recognition
In bacteria, the tRNA-dependent amidation of Glu andAsp to form Gln-tRNAGln and Asn-tRNAAsn is catalyzedby the same enzyme: the heterotrimeric GatCAB [3]. Theexact functional role of bacterial GatCAB in vivo isdetermined by the availability of its misacylated substratesGlu-tRNAGln and Asp-tRNAAsn [3,4,21]. In bacteria thatonly have a ND-GluRS (e.g. Bacillus subtilis) [16], Glu-tRNAGln is generated and GatCAB is exclusively used as a
Fig. 1 The tRNA-dependent pathways for Gln-tRNAGln (A) andAsn-tRNAAsn (B) formation
Fig. 2 The tRNA-dependent Cys-tRNACys formation
Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 541
Amino acid modifications on tRNA
Glu-AdT to transform Glu-tRNAGln to Gln-tRNAGln [3].In bacteria that only encode a ND-AspRS [e.g.Deinococcus radiodurans, Neisseria meningitides,Pseudomonas aeruginosa and Thermus thermophilus (T.thermophilus)], GatCAB functions as an Asp-AdT,synthesizing Asn on tRNAAsn [4,18,21−26]. However, inbacteria possessing both a ND-GluRS and a ND-AspRS(e.g. Chlamydia trachomatis [27] and Helicobacter pylori[28−30]), GatCAB catalyzes both tRNA-dependenttransamidation reactions in vivo [8,27,31].
In general, the anticodon regions of the tRNAGln andthe tRNAAsn, respectively, are not the major identifyingelements for recognition by tRNA-dependent amido-transferases [26,32−34]. In order to discriminate againsttRNAAsp and tRNAGlu, bacterial GatCAB recognizes thefirst base pair in the tRNA acceptor stem, which is U1-A72 in tRNAGln [32] and tRNAAsn [26] and G1-C72 intRNAAsp and tRNAGlu. Additionally, the supernumerarynucleotide U20a in the D-loop of tRNAAsp and tRNAGlu
excludes them as substrates for GatCAB [32]. In N.meningitides, the mutation of U1-A72 to G1-C72 intRNAAsn leads to a 100-fold decrease in transamidationactivity, while the transplantation of the U1-A72 intotRNAAsp, together with the deletion of the U20a, convertstRNAAsp into a substrate that behaves similarly to thewild-type tRNAAsn for bacterial GatCAB [26]. Sequencecomparisons of bacterial tRNAGlu, tRNAGln, tRNAAsp andtRNAAsn have shown that the first base pair of tRNAAsn
and tRNAGln is conserved as U1-A72 in all bacteriaencoding tRNA-dependent pathways for Gln and Asnbiosynthesis, suggesting that GatCAB utilizes one generalmechanism to maintain its substrate specificity [26,32].
Archaeal tRNA-dependent amidotransferasesand tRNA recognition
Two types of tRNA-dependent amidotransferases (AdTs)are found in archaea: the heterodimeric GatDE and thearchaeal GatCAB [5]. GatDE is the archaeal Glu-AdT [5].It exclusively recognizes archaeal tRNAGln and synthe-sizes glutamine on the tRNA. The archaeal GatCAB onlyexists in archaea lacking an asparaginyl-tRNA synthetase(AsnRS) [5,9]. In vitro the Methanothermobacterthermautotrophicus (M. thermautotrophicus) GatCAB onlyacts on Asp-tRNAAsn but not on homologous Glu-tRNAGln,suggesting that the archaeal GatCAB has lost its dualfunction and acts strictly as an Asp-AdT [35]. Theadvantage of a more specialized archaeal GatCABcompared to its bacterial homolog awaits furtherinvestigation.
The tRNA recognition mode of archaeal GatCABdiverges from the bacterial enzyme. The archaeal GatCABdoes not recognize the first base pair of the tRNA substrate;instead, in the case of M. thermautotrophicus GatCAB, itdiscriminates against tRNAAsp by recognizing U49 and theD-loop as major anti-determinant elements [33]. A mutationof A49 to U49 in the wild-type tRNAAsn leads to loss ofmore than 99.5% of activity in the archaeal GatCABcatalyzed transamidation reaction. Additionally, the lengthof the variable loop acts as an identity element in tRNAAsn
recognition as demonstrated by the M. thermautotrophicusand Methanosarcina barkeri enzymes [26,33]. While therecognition of tRNAGln by GatDE, the archaeal Glu-AdT,relies on the same regions (i.e. the first base pair and the
Fig. 3 The selenocysteine biosynthetic pathways in archaea and eukaryotes

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 542
Amino acid modifications on tRNA
D-loop) as the bacterial GatCAB, the bases recognizedare different. GatDE recognizes the first base pair ofarchaeal tRNAGln, conserved as A1-U72, whereas bacterialGatCAB recognizes the U1-A72 base pair of bacterialtRNAGln or tRNAAsn [34]. Mutation of U19 or A20 in theD-loop of M. thermautotrophicus tRNAGln significantlydecreases transamidation catalyzed by GatDE [34].
The catalytic mechanism of tRNA-dependentamidotransferases
Transamidation is an ATP-dependent, multistep reactionrequiring the presence of an amide donor such as glutamineor asparagine. Despite the difference in tRNA specificityand natural distribution, GatCAB and GatDE use the samemechanism to catalyze tRNA-dependent transamidation(Fig. 4). It consists of three sub-reactions: (a) theactivation of the amide acceptor (tRNA-bound Glu or Asp)at the expense of ATP hydrolysis, forming γ-phosphoryl-Glu-tRNAGln or possibly β-phophoryl-Asp-tRNAAsn as
reaction intermediates [15,36−38]; (b) the hydrolysis ofan amide donor Gln or Asn to form enzyme captivatedammonia; and (c) the transfer of sequestered ammonia tothe activated intermediate to form the final product Gln-tRNAGln or Asn-tRNAAsn [38−41]. The kinase (a) and theglutaminase activity (b) of AdTs are tightly coupled uponbinding of the misacylated tRNA substrate [8,38,40,42].
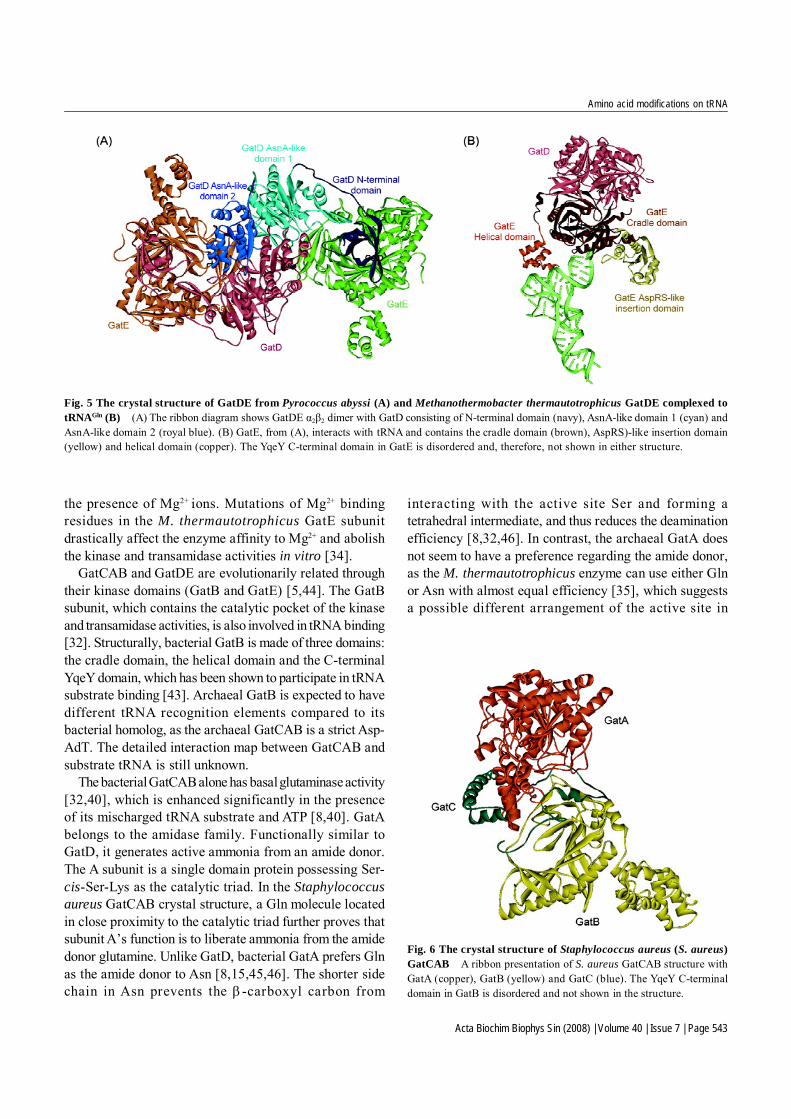
GatDE forms an α2β2 tetramer in solution and crystallineconditions [Fig. 5(A)] [34,42]. The functional role of eachsubunit in GatDE is well elucidated. The D subunit carriesout the glutaminase activity [5,38,42], which releasesammonia from Gln as well as Asn [8,35]. It consists ofthree domains: AnsA-like domain 1, AnsA-like domain 2and the-N-terminal domain. The AnsA-like domainsconnect through a long linker loop to the N-terminaldomain, which is involved in the binding of the E subunit[34,42]. The D subunit forms a tightly packed dimer witha large surface contact area located at the AnsA-likedomains between the two protomers. In the dimerinterface, two amidase catalytic centers are formed byAnsA-like domains 1 and 2 from the other subunit [42],where two highly conserved threonine residues, an asparticacid and a lysine, are crucial for the GatD-catalyzedglutaminase activity [38]. GatD only hydrolyzes Gln inthe presence of the E subunit and Glu-tRNAGln, whichcouples the hydrolysis of Gln with the activation of tRNA-bound Glu, thus preventing an otherwise futile deamina-tion of Gln and the accumulation of free ammonia [38].
GatE interacts with the misacylated tRNA substrate [34,38]. In the presence of ATP, the E subunit alone is able toactivate Glu-tRNAGln, forming a γ-phosphoryl-Glu-tRNAGln
intermediate [38]. Its unique structure consists of a cradledomain, an AspRS-like insertion domain, a helical domainand a C-terminal domain homologous to the YqeY proteinfamily, and it may enhance protein affinity towards itstRNA substrate [34,42]. A similar C-terminal YqeY domainappended to the D. radiodurans Gln-tRNA synthetase(GlnRS) enables the enzyme to bind productively totRNAGln [43]. The helical domain and the C-terminal domainchange their orientation upon tRNA binding and form aconcave surface to accommodate the elbow region of thetRNA substrate [Fig. 5(B)] [34]. The lack of tRNAGln-specific and base-specific interactions in this region ofthe tRNA indicates that a shape-complementary mechanismas the indirect readout of tRNAGln is the main factor thatallows GatE to differentiate tRNAGln from tRNAGlu andtRNAAsn [34]. The cradle domain interacts with the ACCA-terminus of tRNAGln and guides the attached Glu to thecatalytic center for phosphorylation and the subsequenttransamidation reaction [34]. The kinase activity requires
Fig. 4 Transamidation reactions to form Gln-tRNAGln (A) andAsn-tRNAAsn (B) GatCAB and GatDE use the same mechanism tocatalyze the tRNA-dependent transamidation. It consists of three sub-reactions: (a) the activation of the amide acceptor (tRNA bound Glu orAsp) at the expense of ATP hydrolysis, forming γ-phosphoryl-Glu-tRNAGln or possib ly β -phophoryl-Asp-tRNAAsn as react ionintermediate. (b) The hydrolysis of an amide donor Gln or Asn to formenzyme captivated ammonia. (c) The transfer of sequestered ammoniato the activated intermediate to form the final product Gln-tRNAGln orAsn-tRNAAsn.
Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 543
Amino acid modifications on tRNA
the presence of Mg2+ ions. Mutations of Mg2+ bindingresidues in the M. thermautotrophicus GatE subunitdrastically affect the enzyme affinity to Mg2+ and abolishthe kinase and transamidase activities in vitro [34].
GatCAB and GatDE are evolutionarily related throughtheir kinase domains (GatB and GatE) [5,44]. The GatBsubunit, which contains the catalytic pocket of the kinaseand transamidase activities, is also involved in tRNA binding[32]. Structurally, bacterial GatB is made of three domains:the cradle domain, the helical domain and the C-terminalYqeY domain, which has been shown to participate in tRNAsubstrate binding [43]. Archaeal GatB is expected to havedifferent tRNA recognition elements compared to itsbacterial homolog, as the archaeal GatCAB is a strict Asp-AdT. The detailed interaction map between GatCAB andsubstrate tRNA is still unknown.
The bacterial GatCAB alone has basal glutaminase activity[32,40], which is enhanced significantly in the presenceof its mischarged tRNA substrate and ATP [8,40]. GatAbelongs to the amidase family. Functionally similar toGatD, it generates active ammonia from an amide donor.The A subunit is a single domain protein possessing Ser-cis-Ser-Lys as the catalytic triad. In the Staphylococcusaureus GatCAB crystal structure, a Gln molecule locatedin close proximity to the catalytic triad further proves thatsubunit A’s function is to liberate ammonia from the amidedonor glutamine. Unlike GatD, bacterial GatA prefers Glnas the amide donor to Asn [8,15,45,46]. The shorter sidechain in Asn prevents the β-carboxyl carbon from
interacting with the active site Ser and forming atetrahedral intermediate, and thus reduces the deaminationefficiency [8,32,46]. In contrast, the archaeal GatA doesnot seem to have a preference regarding the amide donor,as the M. thermautotrophicus enzyme can use either Glnor Asn with almost equal efficiency [35], which suggestsa possible different arrangement of the active site in
Fig. 5 The crystal structure of GatDE from Pyrococcus abyssi (A) and Methanothermobacter thermautotrophicus GatDE complexed totRNAGln (B) (A) The ribbon diagram shows GatDE α2β2 dimer with GatD consisting of N-terminal domain (navy), AsnA-like domain 1 (cyan) andAsnA-like domain 2 (royal blue). (B) GatE, from (A), interacts with tRNA and contains the cradle domain (brown), AspRS)-like insertion domain(yellow) and helical domain (copper). The YqeY C-terminal domain in GatE is disordered and, therefore, not shown in either structure.
Fig. 6 The crystal structure of Staphylococcus aureus (S. aureus)GatCAB A ribbon presentation of S. aureus GatCAB structure withGatA (copper), GatB (yellow) and GatC (blue). The YqeY C-terminaldomain in GatB is disordered and not shown in the structure.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 544
Amino acid modifications on tRNA
archaeal GatA.GatC is a 10 kDa small protein responsible for
stabilizing the GatCAB trimeric protein complex. In thecrystal structure of S. aureus GatCAB (Fig. 6), the Csubunit is shaped as an extended loop with two α helixesat its N-terminus and two β strands at its C-terminus [32].GatC stabilizes the GatAB complex by extensivelyinteracting with both subunits at the GatA/GatB interface[32]. The proper folding of the GatA subunit also requiresthe presence of GatC [3].
Gated Ammonia Channel in tRNA-dependentAmidotransferases
A most notable feature in the crystal structures of S. aureusGatCAB and M. thermautotrophicus GatDE is a longprotein tunnel (30 Å and 40 Å, respectively) connectingthe glutaminase activity center in the GatA or GatD subunitto the kinase/transamidase active site in the GatB or GatEsubunit [32,34]. The molecular tunnel is made ofcontinuous hydrophilic residues with highly conservedpositive and negative residues alternating on the innersurface, and it is surrounded by hydrophobic residues onthe outside [32,34]. Ammonia generated in the deaminasecenter is expected to travel to the transamidase center forthe transamidation reaction to occur [32,34,42]. Thehydrophilic property of the ammonia tunnel in AdTssuggests that ammonium (NH4
+) instead of ammonia (NH3)is transported. It has been suggested that the transport iscarr ied out through alternating protonation anddeprotonation of the ammonium ion [32]. A continuousdesolvation of ammonium ions, followed by passage intothe tunnel, may push ammonium ions towards thesubsequent reaction center, mimicking the mechanism ofK+ transport in the potassium channel [34,47].
The coupling of glutaminase and kinase/transamidaseactivities has been observed in both GatCAB and GatDE[8,38,40,42]. Regarding GatDE, only in the presence ofmisacylated tRNA substrate does the hydrolysis of Gln orAsn amide donor occur [38,42]. The binding of the tRNAsubstrate induces a significant conformational change inthe D subunit where a catalytic threonine, 7 Å away fromthe active site in the apo enzyme, moves to the activeposition [42]. The ammonia tunnel also undergoesconformational changes and it is proposed to switch froma closed to an open state upon binding of the misacylatedtRNA [32,34,42]. Unlike GatDE, GatCAB has a less tightcoupling between these two subreactions as mentionedearlier [8,32,40]. The lack of a complex structure ofGatCAB and tRNA leaves many questions open in this
area.
Complexes between tRNA, Non-discrimina-ting-aaRS and tRNA-dependent Amido-transferase
Several mechanisms have been proposed to maintain thefidelity of translation and to prevent the misacylated tRNAsgenerated during the described tRNA-dependent aminoacid transformations from participating in decoding, suchas EF-Tu discriminating against misacylated tRNAs [48]and substrate channeling [14,49]. The first mechanism isbased on the diverse binding affinity of EF-Tu towardsdifferent tRNA species and their attached amino acids [50].The cognate aa-tRNAs bind EF-Tu with similar affinityby thermodynamic compensation, whereas the matchingamino acid of a strong binding tRNA weakly binds toEF-Tu and vice versa [48]. The tRNAGln and tRNAAsn haveweaker affinity toward EF-Tu than tRNAGlu and tRNAAsp
[48]. Therefore, Glu-tRNAGln and Asp-tRNAAsn, as weak/weak combinations, are expected to have considerablyless affinity for EF-Tu than cognate aa-tRNAs and areless likely to be involved in translation.
Some recently reported evidence supports a substratechanneling mechanism. Structurally, GluRS from T.thermophilus can be docked easily onto M. thermauto-trophicus GatDE:tRNAGln , forming a ternary complex[34]. The CCA end of the tRNAGln in the active center ofND-GluRS can move to the kinase/transamidase centerin GatE via a simple flip motion resembling the movementof tRNA and its aaRS with an editing domain [34]. A similarcomplex of ND-AspRS, GatDE and tRNA cannot beconstructed due to large steric clashes. Biochemically, astable complex of T. thermophilus ND-AspRS, GatCABand tRNAAsn has been observed in vitro and in vivo [51].The presence of a ND-aaRS reduces the Km of GatCABfor Asp-tRNAAsn and stabilizes Asp-tRNAAsn as well as thefinal product Asn-tRNAAsn [51,52]. Compared to the freeenzymes, complex formation also increases the kcat of ND-AspRS. Substrate channeling couples aminoacylation withtransamidation, thus increasing the overall reactionefficiency and preventing the incorporation of misacylatedtRNA species in translation.
tRNA-dependent Amidotransferase inMitochondria
A number of eukaryotes, including Saccharomycescerevisiae [53] and Homo sapiens [44] encode homologsof AdT subunits in their nuclear genomes. Several lines of

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 545
Amino acid modifications on tRNA
evidence suggest that the indirect pathway for Gln-tRNAGln
formation may also be used in the mitochondria of theseeukaryotes. For example, in yeast mitochondria, theactivity of Glu-AdT is present and was first detected nearlythree decades ago [54]. Recently, the AdT activity wasalso found in mammalian (T. Suzuki, unpublished data)and plant mitochondria [55]. Furthermore, the yeast AdThomologs (Pet112 and YMR293C) are essential formitochondrial function [56,57], and Glu-tRNAGln, thesubstrate of Glu-AdT, was found to be located in themitochondria of S. cerevisiae [58]. The formation of Glu-tRNAGln is intriguing, however, as the reaction cannot becatalyzed by the yeast mitochondrial GluRS in vitro [59].Additionally, cytoplasmic tRNAGln and GlnRS were shownto be imported to the yeast mitochondria as well [59].The import of tRNAGln was also shown for H. sapiens (J.Alfonzo, unpublished data). It is unclear what may be thereasons (e.g. additional coding functionality) for thepresence of dual pathways for Gln-tRNAGln formation inmitochondria.
The Evolutionary View of the tRNA-depen-dent Gln and Asn Formation Pathways
The indirect pathways for Gln and Asn formation arethought to be ancient and existed in the last universalcommunal ancestor (LUCA), while the correspondingGlnRS and AsnRS in the direct pathways are later additionsduring evolution [9,60−63]. The indirect pathway couplesamino acid biosynthesis with translation, and the directpathway requires a de novo synthesis of Gln and Asnindependent of tRNA. The indirect pathway for the Asn-tRNA formation can serve as the sole pathway for freeAsn formation. In fact, for many organisms encoding theND-AspRS/Asp-AdT pathway, the enzymes for Asnbiosynthesis (AsnA and AsnB) are found to be absentsuggesting that the presence of the indirect pathway forAsn formation is essential [8,24]. On the other hand,organisms possessing AsnA, the ammonia-dependentasparagine synthetase, Asn-tRNAAsn is always made usingAsnRS through the direct pathway [8,9]. In the case ofGln, bacterial Glu-AdT prefers Gln as the amide donor[8,15,45,46]thus both direct and indirect pathways relyon the de novo biosynthesis of Gln. Archaeal Glu-AdT canuse both Gln and Asn as the amide donor [5,35]. Therefore,in archaea possessing AsnA, GatDE could use Asn as theamide donor for Gln-tRNAGln formation catalyzed by theGlu-AdT, adding an additional pathway for Glnbiosynthesis.
The retention of the indirect pathway for Gln formation
in Archaea may be due to the unique archaeal tRNAGln
[44], which cannot be recognized and aminoacylated byGlnRS from Escherichia coli or S. cerevisiae [5]. On theother hand, bacterial tRNAGln from B. subtilis, an organismthat encodes the indirect pathway, is a good substrate forE. coli GlnRS [16]. In all free-living organisms,ammonium is fixed mainly through the conversion of Gluto Gln by glutamine synthetase. The free amino acid Glnalso serves as an amide donor for several other biosyntheticpathways as well as a signaling molecule for the nitrogenmetabolism [64,65]. A number of bacteria encoding a Glu-AdT have increased amounts of free Glu in their cells,which favors the indirect pathway for Gln-tRNAGln
formation suggesting another possible reason to maintainthe ancient indirect pathway. The reason why the indirectpathways have not been replaced by the direct pathwaysin these bacteria may include nitrogen, carbon regulationand translation fidelity. The details await furtherinvestigation.
tRNA-dependent Cys Synthesis in Archaea
In a large subset of Euryarchaeota [66], Cys is synthesizedin a tRNA-dependent manner (Table 1) [6]. This indirectpathway for Cys-tRNACys formation utilizes two enzymes,SepRS and SepCysS (Fig. 2) [6]. SepRS aminoacylatestRNACys with Sep [6]. The Sep moiety is then convertedto Cys by SepCysS in the presence of an unknown sulfurdonor to form Cys-tRNACys [6].
Genomic analyses revealed that SepRS and SepCysSare both encoded in the sulfate reducing archaeonArchaeoglobus fulgidus [67] and all known methanogenicarchaea [66] except Methanosphaera stadtmanae [68] andMethanobrevibacter smithii [69]. In most of these archaea,CysRS is also coded for though it may not be essential[66,70]; for example, in Methanococcus maripaludisCysRS is dispensable [71].
In many of these euryarchaeal genomes the enzymesrequired for the formation of free Cys (i.e. tRNA-independent) biosynthesis are not encoded [6,66]. Theindirect route for Cys-tRNACys formation using SepRSand SepCysS is likely the sole means for Cys biosynthesisin these organisms [6]. This is consistent with an earlierreport demonstrating that Sep is a precursor for Cysbiosynthesis in M. jannaschii [72]. Further studies haveshown that an archaeal Sep biosynthetic pathway canprovide sufficient Sep levels for Ser, cystathionine andtRNA-dependent Cys production [73]. Furthermore,knocking out sepS in M. maripaludis resulted in a Cysauxotroph [6]. Therefore, the use of SepRS and SepCysS

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 546
Amino acid modifications on tRNA
for the tRNA-dependent Cys synthesis in these organismslikely enables coupling of protein synthesis with Cysproduction.
SepRS Directly Aminoacylates tRNACys withO-phosphoserine
SepRS is a subclass IIc aaRS like PheRS and PylRS [6,74], sharing a common ancestor with the α-subunit ofPheRS [66,74]. Both biophysical and structural analysesdemonstrate that SepRS is a homotetramer [75,76]. Thecore of this α4 assembly resembles that of PheRS andconsists mostly of the four catalytic domains [75].
While the active site of SepRS is structurally similar tothat of PheRS, SepRS uniquely recognizes its amino acidsubstrate, Sep [76]. The phosphate group of the latter ishighly recognized by SepRS with each of the three non-bridging oxygen atoms forming two hydrogen bonds toresidues in the enzyme’s binding pocket. Mutation of theseresidues in the M. maripaludis SepRS resulted in inactivemutant enzymes [75]. The recognition of the phosphatemoiety includes hydrogen bonding between two non-bridging oxygens and the α-amino group of active siteresidues, which is unique amongst aaRSs to SepRS.Structural results suggest that dipole interactions betweenSep and a central α-helix in the active site of SepRS stabilizethe polar side chain of the substrate, another feature notobserved in other aaRSs [76].
Each monomer of SepRS can recognize andaminoacylate tRNACys in cis [76], in contrast to PheRSwhere the tRNA anticodon recognition site andaminoacylation active site are found on different subunits.However, in the co-crystal structure of the A. fulgidusSepRS with tRNACys only two tRNA molecules bound tothe tetramer [76]. Computer modeling though does suggestthat four tRNAs could be accommodated by the complex[76]. The stoichiometry in solution of SepRS to tRNACys
is not clear and awaits further investigation.Biochemical studies revealed that M. maripaludis SepRS
recognizes the same major identity elements in tRNACys
(G34, C35, and A36) as the homologous CysRS [77].Both aaRSs also use G15 and A47 in tRNACys as minoridentity elements. However the base pairs G1:C72 andG10:C25, and nucleotides G37 and A59 serve as minoridentity elements for only SepRS [77]. The use of similaridentity elements in tRNACys recognition by both SepRSand CysRS, both of which were present in LUCA [66,74], suggests that the genetic code predates the modernaminoacylation machinery [77]. Given that SepRS, likeother class II aaRSs, approaches the tRNA from the ma-
jor groove side while CysRS, a class I aaRS, approachesit from the minor groove side has lead to speculation thata complex between SepRS, tRNACys and CysRS is possible[76], though the in vivo role of such a complex is currentlynot clear.
SepCysS Catalyzed Formation of Cys-tRNACys
The pyridoxal phosphate (PLP)-dependent enzymeSepCysS modifies the Sep bound to tRNACys to form Cys-tRNACys [6]. The sulfur donor for this enzyme is unknownthough in vitro sulfide is sufficient [6]. The A. fulgidusSepCysS crystal structure (2.4 Å resolution) [78] revealedthat it belongs to the fold type I family [79] with its largeN-terminal domain being comprised of a characteristicseven stranded β-sheet which typifies this family ofenzymes. In addition, the structure showed that theenzyme forms a homodimer [78]. The active site of theenzyme is formed in a large basic cleft in the dimerinterface and is comprised of conserved residues fromboth monomers [78]. Modeling a SepCysS:Sep-tRNACys
complex suggests that a conserved Arg79, His103, andTyr 104 (A. fulgidus numbering) recognize the phosphategroup of the Sep moiety of the tRNA substrate [78]. Thesame work implicates one of the three conserved Cysresidues (39, 42 or 247) in the SepCysS active site as thepersulfide sulfur carrier essential for catalysis (Fig. 7)[78] though this awaits further study.
The crystal structure of SepCysS with PLP alonerevealed that the co-factor formed a Schiff-base linkagewith the conserved Lys209 in the active site of SepCysS[78]. A hydrogen bond between SepCysS and the nitrogen
Fig. 7 The crystal structure of the active sites in A. fulgidus SepCysSand M. maripaludis SepSecS The protein active sites are presentedas ribbon diagrams with essential residues highlighted as stick models.Monomers in each protein are colored pink and blue. Adapted fromSheppard et al [133], with permission from Oxford University Press.PLP, pyridoxal phosphate.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 547
Amino acid modifications on tRNA
atom of the ring structure of PLP is achieved through theside chain of a conserved Asn and not an Asp as is foundin most PLP-dependent enzymes [78]. Nevertheless, likein other PLP-dependent enzymes, the co-factor inSepCysS is thought to stabilize the negatively chargedtransition state formed during catalysis of the β-replacement reaction [80].
M. maripaludis encodes both the direct and indirectpaths for Cys-tRNACys synthesis. As noted above, thesole route for Cys formation is tRNACys dependent.Intriguingly while sepS (encoding SepRS) can be deletedwhen the organism is grown in the presence of Cys, pscS(encoding SepCysS) cannot (T. Major, M. Hohn, D. Su,W.B. Whitman, unpublished data), raising the questionwhether SepCysS possesses an additional function inM. maripaludis that is essential.
Cys Synthesis in Archaea
Four different routes for Cys formation have beendiscovered in archaea: the eukaryotic pathway in whichthe precursor is cystathionine [81], the bacterial pathwaywhere O-acetylserine serves as the precursor [82−84], amodified bacterial pathway with free Sep as the precursor[84,85], and the tRNA-dependent route with Sep-tRNACys
as the precursor [6]. Cys is implicated as the major sulfursource for a variety of biosynthetic pathways includingFe-S cluster formation, tRNA modification, andbiosynthesis of co-factors in bacteria (reviewed in [87]).Fe-S cluster proteins are highly encoded in the genomesof methanogenic archaea [88]. Whether Cys generatedthrough the tRNA-dependent pathway is used as the sulfursource is an open area of investigation. It is thought thatthese euryarchaea must balance the need for Cys in proteinsynthesis and other biosynthetic pathways by controllingthe deacylation of Cys-tRNA, thus regulating the level offree Cys relative to Cys-tRNACys; however, it may well bethat these methanogens which grow in environments richin reduced sulfur compounds may be able to use inorganicsulfur directly.
Evolution of the Two Cys-tRNACys Biosyn-thetic Pathways
It has been speculated that the indirect pathways for aa-tRNA formation predate the direct ones [89]. Whilephylogenetics supports this speculation for amide aa-tRNAsynthesis (discussed above), analyses using structure-based amino acid alignments [66,74] suggest that bothpathways (SepRS/SepCysS and CysRS) for Cys-tRNACys
formation were present in LUCA. The phylogenetic datasuggest that only early bacteria retained CysRS while theancestral archaea possessed SepRS and that CysRS waslater horizontally transferred to archaea. In some archaeallineages the bacterial CysRS replaced the indirect pathwayfor Cys-tRNACys synthesis while in many euryarchaeaCysRS either coexisted with SepRS/SepCysS or was notretained [66]. Why the indirect pathway for Cys-tRNACys
formation has been retained in these euryarchaea remainsan open question. It is speculated that a link between Cysformation, sulfur metabolism and methanogenesis mayexist that would favor retention of the tRNA-dependentroute for Cys biosynthesis [66,90] though this awaitsfurther experimental inquiry.
tRNA-dependent Sec Formation
Sec is the major biological form of selenium, an essentialdietary trace element in humans implicated in cancerprevention [91,92]. Sec is coded as the 21st amino acid ina number of species across all three domains of life(reviewed in [13,93]). Under physiological conditions (pH7), Sec is more stable in its ionized form than Cys due tothe lower redox potential which thus lowers the pKa ofthe selenol group of Sec compared to the thiol group ofCys (5.2 and 8.5, respectively) [94]. Sec thus serves asan excellent nucleophile in the active sites of proteinsinvolved in oxidation-reduction reactions.
Selenoprotein synthesis requires the formation ofselenocysteinyl-tRNASec (Sec-tRNASec). No aaRS, [i.e. aselenocysteinyl-tRNA synthetase (SecRS)], has beenidentified in Sec-decoding organisms that can carry outthe task (Table 1) and instead Sec is synthesized ontRNASec (Fig. 3). Why Sec-tRNA is only formed via indirectpaths remains unknown but it may be due to selectivepressures to maintain translational fidelity. CysRSs fromE. coli [95] and Phaseolus aureus [96] have the ability toaminoacylate tRNACys with Sec. Therefore, given the factthat misincorporation of Sec in place of Cys can bedetrimental to protein function [97], the levels of free Secare likely well regulated and kept low to preventmisacylation of tRNACys with Sec. It is also worth notingthat Sec to Cys mutations lead to mutant enzymes withsignificantly reduced activities [98]. Synthesizing Sec ontRNASec enables formation of Sec-tRNASec while potentiallyminimizing the free Sec levels in vivo, preventingmisacylation of tRNACys with Sec; thus the retention ofthe tRNA-dependent Sec pathway may be a mechanismof ensuring the accurate decoding of Cys and Sec [99].
In all known Sec-decoding organisms [100−102],

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 548
Amino acid modifications on tRNA
tRNASec is first serylated by SerRS to form Ser-tRNASec
[103−105]. Work in the 1990s revealed in bacteria thatselenocyteine synthase (SelA) transforms Ser bound totRNASec to Sec (Fig. 3) [13]. The Sec-tRNASec formed isthen used in protein synthesis to decode UGA (usually astop codon) when an RNA element, the selenocysteineinsertion sequence element, is present in the mRNA [13].A unique elongation factor, SelB, brings the Sec-tRNASec
to the ribosome [13]. The mischarged species, Ser-tRNASec, in Sec-decoding archaea and eukaryote (Fig. 7)is not directly modified to Sec-tRNASec, but rather the Sermoiety on the tRNA is phosphorylated by PSTK to formSep-tRNASec (Fig. 3) [106,107]. The tRNA-bound Sep isthen converted to Sec by the PLP-dependent enzymeSep-tRNA:Sec-tRNA synthase (SepSecS) [7,20].
Like SelA, SepSecS uses selenophosphate as theselenium donor to produce Sec-tRNASec in vitro and invivo [7,20,108−110]. SepSecS is unable to use Ser-tRNASec
as a substrate, recognizing only Sep-tRNASec [7,20].However, in vitro bacterial SelA in addition to Ser-tRNASec,can convert Sep-tRNASec to Sec-tRNASec [20], though thebiological relevance of this is not clear as PSTK is notencoded in bacterial genomes [7]. In all known Sec-decoding archaeal and eukaryal genomes, PSTK andSepSecS are always both encoded [111].
It is unknown why archaea and eukaryotes use Sep-tRNASec as an intermediate in tRNA-dependent Secbiosynthesis. The carboxyl ester bond between Sep andtRNASec is more stable than Ser and the tRNASec [106]. Inaddition, the phosphate moiety of Sep is likely a betterleaving group than the hydroxyl moiety of Ser. Thus, Sep-tRNASec may serve as a better precursor for Sec-tRNASec
formation than Ser-tRNASec [7].
tRNASec-dependent Ser Phosphorylation
Work with extracts from rat and rooster liver and lactatingbovine mammary gland in the 1970s first demonstratedthat Ser-tRNA could be phosphorylated [112,113]. Whileit was later shown with partially purified enzyme frombovine liver that the enzyme had a high affinity for tRNASec,it was only in 2004 that the protein (PSTK) was identifiedfrom mouse [106], and soon after the archaeal homolog[107].
PSTK phosphorylates Ser-tRNASec by transferring theγ-phosphate of ATP onto the Ser moiety in an Mg2+-dependent manner [106,111]. The enzyme belongs to theP-loop kinase superfamily [114], possessing a phosphate-binding loop (P-loop), a Walker B motif, and an RxxxRmotif in its N-terminal domain [111]. Mutation of conserved
residues in these motifs in the M. jannaschii PSTK resultedin mutant enzymes with significantly reduced activity[111]. While PSTK prefers ATP, in vitro the enzyme isable to use other NTPs (GTP, CTP, UTP and dATP) assubstrates, like T4 polynucleotide kinase [111]. Similar toother members of the kinase superfamily [114], theATPase activity of PSTK is activated in the presence of itsother substrate Ser-tRNASec [111]. Interestingly, the activityis also enhanced when unacylated tRNASec is provided[111].
PSTK Recognition of Sep-tRNASec
It does not appear that PSTK uses the Sep moiety oftRNASec as a major recognition element as the enzyme hasa similar Kd for unacylated tRNASec as Ser-tRNASec (39nM and 53 nM, respectively) [111]. In vivo, the concen-tration of tRNASec is approximately 10% of that of tRNASer
[115,116], the other tRNA substrate of SerRS. It maywell be that PSTK serves as a tRNASec scavenger forSerRS [111]. PSTK may also assist in maintaining trans-lation fidelity by preventing the misacylated tRNASec in-termediates from being used in protein synthesis and chan-neling Sep-tRNASec to SepSecS [111].
Surprisingly, it appears that archaeal and eukaryoticPSTK enzymes recognize different elements in Ser-tRNASec. While tRNASec possesses an extended variableloop like tRNASer, a major identity element for SerRS rec-ognition of tRNA [117], it is distinct from other tRNAisoacceptors by possessing an elongated acceptor stemand D-stem [118,119]. For tRNASec recognition by eu-karyotic PSTK, the major element is the length and con-formation of the elongated D-stem [120]. For archaealPSTK, the D-stem is a minor identity element and the G2-C71 and C3-G70 base pairs in the acceptor stem ofarchaeal tRNASec serve as the major recognition elementsin the tRNA [121]. Given the deep phylogenetic dividebetween archaeal and eukaryotic PSTK [111], this maybe a strong indication of co-evolution of PSTK and tRNASec
[121].Interestingly, while bacterial tRNASec has an 8 bp ac-
ceptor stem and a 5 bp T-stem, and archaeal and eukary-otic tRNASec have a 9 bp acceptor stem and 4 bp T-stemarrangement [118,122−124], PSTK and SepSecS can useE. coli tRNASec in vivo [7]. In turn, E. coli can use thehuman tRNASec in place of its own tRNASec both in vivoand in vitro [125]. It may well be that tRNASec is func-tionally conserved between the different domains of lifedespite bacteria using a different tRNA-dependent routefor Sec formation than Sec-decoding archaea and

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 549
Amino acid modifications on tRNA
eukaryotes.
Sep-tRNA: Sec-tRNA Synthase SepSecSCatalyzed Sec-tRNASec Formation
While SepSecS catalyzes a similar reaction as SepCysS, atRNA-dependent β-replacement of Sep, a structuralphylogeny revealed that SepSecS is not closely related toSepCysS nor other PLP-dependent enzymes [126]. Therecently completed crystal structures of the SepSecS fromM. maripaludis [126] and mouse [127] to high resolutionhave enabled insight into how the enzyme catalyzes thetRNA-dependent formation of Sec. SepSecS forms an (α2)2 homotetramer, mediated by an N-terminal extension inSepSecS. Each dimer has two active sites, each formedby conserved residues from both subunits in the dimerinterface (Fig. 7) [126,127]. Interestingly deleting the N-terminal extension, thus apparently disrupting dimerization,gives rise to inactive SepSecS [126]. Tetramerization isspeculated to also enable formation of large patches ofpositive electric potential on the surface of the tetramer,which are predicted to be tRNASec binding sites [127].
As in the SepCysS active site, a conserved Asn in theSepSecS (247, M. maripaludis numbering) active sitebinds to the nitrogen of the ring structure of the PLP andsimilar to other PLP-dependent enzymes a conserved Lys(278, M. maripaludis numbering) forms a Schiff baselinkage with the co-factor [126]. The phosphate group(s)of Sep-tRNASec and/or selenophosphate are proposed tointeract with conserved Arg, Gln, and Ser in the activesite of SepSecS [126]. Mutations to those residues resultsin mutant enzymes with significantly reduced activitiesboth in vitro and in vivo [126]. SepSecS may excludefree amino acids including Sep from its active site by usinga conserved Glu, which could repel the carboxyl groupof free amino acids [127]. Unlike SepCysS, a conservedCys residue is not found in the active site of SepSecS,suggesting that the formation of a perselenide intermediateis unlikely [126].
Relationship between tRNA-dependent Cysand Sec biosynthesis
The tRNA-dependent route for Cys biosynthesis is similarto that for Sec-tRNASec formation in archaea andeukaryotes, since Sep-tRNA serves as the final precursorprior to product formation in both of them. Both pathwayswere present in LUCA [7,66]. Interestingly, SepSecS canuse thiophosphate in vitro to form Cys-tRNASec [126]instead of selenophosphate to synthesize Sec-tRNASec.
Numerous homologs of selenoproteins are found in nature,which possess Cys in place of Sec. Given that and thesimilarity between Sec and Cys codons (UGA and UGY,respectively), it is interesting to speculate that a dynamicrelationship has existed between the two amino acids overthe course of evolution [128].
Outlook
The tRNA-dependent pathways forming Gln-tRNAGln andAsn-tRNAAsn through amino acid transformations arethought to have evolved earlier than the directaminoacylation of the tRNAs with their cognate aminoacids [9,60−63]. In the case of Cys-tRNACys formation,both the direct and the indirect pathways have been shownto be present in the time of LUCA [66,75,77]. RegardingSec, Sec-tRNASec is formed in all domains of life onlythrough the indirect tRNA-dependent amino acidtransformations [100−102]. Even though it cannot begeneralized that indirect pathways are ancient pathways,it does appear that indirect pathways have unique featuresthat have been retained throughout evolution. For example,tRNA-dependent Sec-tRNASec formation may provide asolution to discriminate against Cys, an extremely similaramino acid, and maintain a faithful translation.
A common feature among these indirect pathways isthe existence of misacylated intermediates, which woulddrastically decrease the fidelity of translation if theyparticipated in decoding. Even though elongation factorsbind misacylated intermediates with too low or too highaffinity in vitro [21,48,50,129−131], it is still ambiguouswhether the discrimination by the elongation factor alonecould ensure the accuracy of translation in vivo [51,132].Substrate channeling provides an additional mechanismto prevent misincorporation. Interestingly, complexesconsist of enzymes in the same tRNA-dependent pathway,and the corresponding tRNA molecules have either beenobserved or proposed based on computer modeling [34,51,78]. Furthermore, complex formation may increasethe overall reaction efficiency as well as the stability ofthe end product [51,52]. The tRNA-dependent amino acidtransformations couple translation with the biosynthesisof amino acids, which may be involved in other biologicalpathways [64]. To better understand the connection andregulation among different biological processes, systemsbiology is likely to be a very useful approach, and it mayalso lead to discovery of other exciting aspects of tRNA-dependent amino acid transformation pathways.
Acknowledgements

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 550
Amino acid modifications on tRNA
References
1 Ibba M, Söll D. Aminoacyl-tRNA synthesis. Annu Rev Biochem2000, 69: 617−650
2 Wilcox M, Nirenberg M. Transfer RNA as a cofactor couplingamino acid synthesis with that of protein. Proc Natl Acad Sci USA1968, 61: 229−236
3 Curnow AW, Hong K, Yuan R, Kim S, Martins O, Winkler W,Henkin TM et a l . Glu-tRNAGln amidotransferase: a novelheterotrimeric enzyme required for correct decoding of glutaminecodons during translation. Proc Natl Acad Sci USA 1997, 94:11819−11826
4 Curnow AW, Tumbula DL, Pelaschier JT, Min B, Söll D. Glutamyl-tRNAGln amidotransferase in Deinococcus radiodurans may beconfined to asparagine biosynthesis. Proc Natl Acad Sci USA 1998,95: 12838−12843
5 Tumbula DL, Becker HD, Chang WZ, Söll D. Domain-specificrecruitment of amide amino acids for protein synthesis. Nature2000, 407: 106−110
6 Sauerwald A, Zhu W, Major TA, Roy H, Palioura S, Jahn D,Whitman WB et al. RNA-dependent cysteine biosynthesis inarchaea. Science 2005, 307: 1969−1972
7 Yuan J, Palioura S, Salazar JC, Su D, O’Donoghue P, Hohn MJ,Cardoso AM et al. RNA-dependent conversion of phosphoserineforms selenocysteine in eukaryotes and archaea. Proc Natl AcadSci USA 2006, 103: 18923−18927
8 Sheppard K, Akochy PM, Salazar JC, Söll D. The Helicobacterpylori amidotransferase GatCAB is equally efficient in glutamine-dependent transamidation of Asp-tRNAAsn and Glu-tRNAGln. J BiolChem 2007, 282: 11866−11873
9 Roy H, Becker HD, Reinbolt J , Kern D. When contemporaryaminoacyl-tRNA synthetases invent their cognate amino acidmetabolism. Proc Natl Acad Sci USA 2003, 100: 9837−9842
10 Bult CJ, White O, Olsen GJ, Zhou L, Fleischmann RD, Sutton GG,Blake JA et al. Complete genome sequence of the methanogenicarchaeon, Methanococcus jannaschii. Science 1996, 273: 1058−1073
11 Smith DR, Doucette-Stamm LA, Deloughery C, Lee H, Dubois J,Aldredge T, Bashirzadeh R et al. Complete genome sequence ofMethanobacterium thermoautotrophicum deltaH: functionalanalysis and comparative genomics. J Bacteriol 1997, 179: 7135−7155
12 Slesarev AI, Mezhevaya KV, Makarova KS, Polushin NN,Shcherbinina OV, Shakhova VV, Belova GI et al. The completegenome of hyperthermophile Methanopyrus kandleri AV19 andmonophyly of archaeal methanogens. Proc Natl Acad Sci USA2002, 99: 4644−4649
13 Böck A, Thanbichler M, Rotherand M, Resch A. The Aminoacyl-tRNA Synthetases. Georgetown: Landes Bioscience, 2005
14 Schön A, Kannangara CG, Gough S, Söll D. Protein biosynthesis inorganelles requires misaminoacylation of tRNA. Nature 1988, 331:
187−19015 Jahn D, Kim YC, Ishino Y, Chen MW, Söll D. Purification and
functional characterization of the Glu-tRNAGln amidotransferasefrom Chlamydomonas reinhardtii. J Biol Chem 1990, 265: 8059−8064
16 Lapointe J, Duplain L, Proulx M. A single glutamyl-tRNA syn-thetase aminoacylates tRNAGlu and tRNAGln in Bacillus subtilis andefficiently misacylates Escherichia coli tRNAGln1 in vitro. J Bacteriol1986, 165: 88−93
17 Cathopoulis T, Chuawong P, Hendrickson TL. Novel tRNAaminoacylation mechanisms. Mol Biosyst 2007, 3: 408−418
18 Becker HD, Reinbolt J, Kreutzer R, Giege R, Kern D. Existence oftwo distinct aspartyl-tRNA synthetases in Thermus thermophilus.Structural and biochemical properties of the two enzymes. Bio-chemistry 1997, 36: 8785−8797
19 Curnow AW, Ibba M, Söll D. tRNA-dependent asparagine formation.Nature 1996, 382: 589−590
20 Xu XM, Carlson BA, Mix H, Zhang Y, Kazima S, Glass RS, BerryMJ et al. Biosynthesis of selenocysteine on its tRNA in eukaryotes.PLoS Biol 2007, 5: 96−105
21 Becker HD, Kern D. Thermus thermophilus: a link in evolution ofthe tRNA-dependent amino acid amidation pathways. Proc NatlAcad Sci USA 1998, 95: 12832−12837
22 Becker HD, Min B, Jacobi C, Raczniak G, Pelaschier J, Roy H,Klein S et al. The heterotrimeric Thermus thermophilus Asp-tRNAAsn amidotransferase can also generate Gln-tRNAGln. FEBSLett 2000, 476: 140−144
23 Becker HD, Roy H, Moulinier L, Mazauric MH, Keith G, Kern D.Th er m u s th e r mo p h i lu s conta ins a n euba cteri a l a nd a narchaebacterial aspartyl-tRNA synthetase. Biochemistry 2000, 39:3216−3230
24 Min B, Pelaschier JT, Graham DE, Tumbula-Hansen D, Söll D.Transfer RNA-dependent amino acid biosynthesis: an essentialroute to asparagine formation. Proc Natl Acad Sci USA 2002, 99:2678−2683
25 Akochy PM, Bernard D, Roy PH, Lapointe J. Direct glutaminyl-tRNA biosynthesis and indirect asparaginyl-tRNA biosynthesis inPseudomonas aeruginosa PAO1. J Bacteriol 2004, 186: 767−776
26 Bailly M, Giannouli S, Blaise M, Stathopoulos C, Kern D, BeckerHD. A single tRNA base pair mediates bacterial tRNA-dependentbiosynthesis of asparagine. Nucleic Acids Res 2006, 34: 6083−6094
27 Raczniak G, Becker HD, Min B, Söll D. A single amidotransferaseforms asparaginyl-tRNA and glutaminyl-tRNA in Chlamydiatrachomatis. J Biol Chem 2001, 276: 45862−45867
28 Skouloubris S, Ribas de Pouplana L, De Reuse H, Hendrickson TL.A noncognate aminoacyl-tRNA synthetase that may resolve amissing link in protein evolution. Proc Natl Acad Sci USA 2003,100: 11297−11302
29 Salazar JC, Ahel I, Orellana O, Tumbula-Hansen D, Krieger R,Daniels L, Söll D. Co-evolution of an aminoacyl-tRNA synthetasewith its tRNA substrates. Proc Natl Acad Sci USA 2003, 100:13863−13868
30 Chuawong P, Hendrickson TL. The non-discriminating aspartyl-tRNA synthetase from Helicobacter pylori: anticodon-binding do-main mutations that impact tRNA specificity and heterologoustoxicity. Biochemistry 2006, 45: 8079−8087
31 Cathopoulis TJ, Chuawong P, Hendrickson TL. A thin-layer elec-trophoretic assay for Asp-tRNAAsn/Glu-tRNAGln amidotransferase.Anal Biochem 2007, 360: 151−153
We would like to thank all the current members of Dr.Dieter Söll’s laboratory for discussing the manuscript andassisting with its revision. We would also like to thankDrs. T. Suzuki, J. Alfonzo and W.B. Whitman for provid-ing some of their unpublished results.

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 551
Amino acid modifications on tRNA
32 Nakamura A, Yao M, Chimnaronk S, Sakai N, Tanaka I. Ammoniachannel couples glu taminase with transamidase reactions inGatCAB. Science 2006, 312: 1954−1958
33 Namgoong S, Sheppard K, Sherrer RL, Söll D. Co-evolution of thearchaeal tRNA-dependent amidotransferase GatCAB with tRNAAsn.FEBS Lett 2007, 581: 309−314
34 Oshikane H, Sheppard K, Fukai S, Nakamura Y, Ishitani R, NumataT, Sherrer RL et al. Structural basis of RNA-dependent recruit-ment of glutamine to the genetic code. Science 2006, 312: 1950−1954
35 Sheppard K, Sherrer RL, Söll D. Archaeal tRNAGln confines theamidotransferase GatCAB to Asn-tRNA formation. J Mol Biol2008, 377: 845−853
36 Wilcox M. Gamma-phosphoryl ester of Glu-tRNAGln as an inter-mediate in Bacillus subtilis glutaminyl-tRNA synthesis. Cold SpringHarb Symp Quant Biol 1969, 34: 521−528
37 Wilcox M. Gamma-glutamyl phosphate attached to glutamine-specific tRNA. A precursor of glutaminyl-tRNA in Bacillus subtilis.Eur J Biochem 1969, 11: 405−412
38 Feng L, Sheppard K, Tumbula-Hansen D, Söll D. Gln-tRNAGln for-mation from Glu-tRNAGln requires cooperation of an asparaginaseand a Glu-tRNAGln kinase. J Biol Chem 2005, 280: 8150−8155
39 Harpel MR, Horiuchi KY, Luo Y, Shen L, Jiang W, Nelson DJ,Rogers KC et al. Mutagenesis and mechanism-based inhibition ofStreptococcus pyogenes Glu-tRNAGln amidotransferase implicate aserine-based glutaminase site. Biochemistry 2002, 41: 6398−6407
40 Horiuchi KY, Harpel MR, Shen L, Luo Y, Rogers KC, CopelandRA. Mechanistic studies of reaction coupling in Glu-tRNAGln
amidotransferase. Biochemistry 2001, 40: 6450−645741 Shin S, Yun YS, Koo HM, Kim YS, Choi KY, Oh BH. Characteriza-
tion of a novel Ser-cisSer-Lys catalytic triad in comparison withthe classical Ser-His-Asp triad. J Biol Chem 2003, 278: 24937−24943
42 Schmitt E, Panvert M, Blanquet S, Mechulam Y. Structural basisfor tRNA-dependent amidotransferase function. Structure 2005,13: 1421−1433
43 Deniziak M, Sauter C, Becker HD, Paulus CA, Giege R, Kern D.Deinococcus glutaminyl-tRNA synthetase is a chimer between pro-teins from an ancient and the modern pathways of aminoacyl-tRNA formation. Nucleic Acids Res 2007, 35: 1421−1431
44 Sheppard K, Söll D. On the evolution of the tRNA-dependentamidotransferases, GatCAB and GatDE. J of Mol Biol 2008, 377:831−844
45 Strauch MA, Zalkin H, Aronson AI. Characterization of theglutamyl-tRNAGln-to-glutaminyl-tRNAGln amidotransferase reactionof Bacillus subtilis. J Bacteriol 1988, 170: 916−920
46 Bailly M, Blaise M, Roy H, Deniziak M, Lorber B, Birck C, BeckerHD et al. tRNA-dependent asparagine formation in prokaryotes:Characterization, isolation and structural and functional analysisof a ribonucleoprotein particle generating Asn-tRNA(Asn). Meth-ods 2008, 44: 146−163
47 Morais-Cabral JH, Zhou Y, MacKinnon R. Energetic optimizationof ion conduction rate by the K+ selectivity filter. Nature 2001,414: 37−42
48 LaRiviere FJ, Wolfson AD, Uhlenbeck OC. Uniform binding ofaminoacyl-tRNAs to elongation factor Tu by thermodynamiccompensation. Science 2001, 294: 165−168
49 Srivastava DK, Bernhard SA. Metabolite transfer via enzyme-enzyme complexes. Science 1986, 234: 1081−1086
50 Asahara H, Uhlenbeck OC. The tRNA specificity of Thermus
thermophilus EF-Tu. Proc Natl Acad Sci USA 2002, 99: 3499−3504
51 Bailly M, Blaise M, Lorber B, Becker HD, Kern D. Thetransamidosome: a dynamic ribonucleoprotein particle dedicatedto prokaryotic tRNA-dependent asparagine biosynthesis. Mol Cell2007, 28: 228−239
52 Huot JL, Balg C, Jahn D, Moser J, Emond A, Blais SP, ChenevertR et al. Mechanism of a GatCAB amidotransferase: aspartyl-tRNAsynthetase increases its affinity for Asp-tRNAAsn and novelaminoacyl-tRNA analogues are competitive inhibitors. Biochem-istry 2007, 46: 13190−13198
53 Kim SI, Stange-Thomann N, Martins O, Hong KW, Söll D, FoxTD. A nuclear genetic lesion affecting Saccharomyces cerevisiaemitochondrial translation is complemented by a homologous Ba-cillus gene. J Bacteriol 1997, 179: 5625−5627
54 Dirheimer D, Keith G, Sibler AP, Martin RP. Transfer RNA:structure, properties, and recognition, 1st edn. New York: ColdSpring Harbor, 1979
55 Pujol C, Bailly M, Kern D, Marechal-Drouard L, Becker H andDuchene AM. Dual-targeted tRNA-dependent amidotransferaseensureds both mitochondrial and chloroplastic Gln-tRNAGln syn-thesis in plants. Proc Natl Acad Sci USA 2008, 105: 6481−6485
56 Mulero JJ, Rosenthal JK, Fox TL. PET112, a Saccharomycescerevisiae nuclear gene required to maintain rho+ mitochondrialDNA. Curr Genet 1994, 25: 299−304
57 Hughes TR, Marton MJ, Jones AR, Roberts CJ, Stoughton R, ArmourCD, Bennett HA et al. Functional discovery via a compendium ofexpression profiles. Cell 2000, 102: 109−126
58 Martin NC, Rabinowitz M, Fukuhara H. Yeast mitochondrial DNAspecifies tRNA for 19 amino acids. Deletion mapping of the tRNAgenes. Biochemistry 1977, 16: 4672−4677
59 Rinehart J, Krett B, Rubio MA, Alfonzo JD, Söll D. Saccharomy-ces cerevisiae imports the cytosolic pathway for Gln-tRNA syn-thesis into the mitochondrion. Genes Dev 2005, 19: 583−592
60 Lamour V, Quevillon S, Diriong S, N’Guyen VC, Lipinski M, MirandeM. Evolution of the Glx-tRNA synthetase family: the glutaminylenzyme as a case of horizontal gene transfer. Proc Natl Acad SciUSA 1994, 91: 8670−8674
61 Woese CR, Olsen GJ, Ibba M, Söll D. Aminoacyl-tRNA synthetases,the genetic code, and the evolutionary process. Microbiol MolBiol Rev 2000, 64: 202−236
62 O’Donoghue P, Luthey-Schulten Z. On the evolution of structurein aminoacyl-tRNA synthetases. Microbiol Mol Biol Rev 2003,67: 550−573
63 O’Donoghue P, Luthey-Schulten Z. Evolutionary profiles derivedfrom the QR factorization of multiple structural alignments givesan economy of information. J Mol Biol 2005, 346: 875−894
64 Forchhammer K. Glutamine signalling in bacteria. Front Biosci2007, 12: 358−370
65 Sonenshein AL. Control of key metabolic intersections in Bacillussubtilis. Nat Rev Microbiol 2007, 5: 917−927
66 O’Donoghue P, Sethi A, Woese CR, Luthey-Schulten ZA. Theevolutionary history of Cys-tRNACys formation. Proc Natl AcadSci USA 2005, 102: 19003−19008
67 Klenk HP, Clayton RA, Tomb JF, White O, Nelson KE, KetchumKA, Dodson RJ et al. The complete genome sequence of thehyperthermophilic, sulphate-reducing archaeon Archaeoglobusfulgidus. Nature 1997, 390: 364−370
68 Fricke WF, Seedorf H, Henne A, Kruer M, Liesegang H, HedderichR, Gottschalk G et al. The genome sequence of Methanosphaera

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 552
Amino acid modifications on tRNA
stadtmanae reveals why this human intestinal archaeon is restrictedto methanol and H2 for methane formation and ATP synthesis. JBacteriol 2006, 188: 642−658
69 Samuel BS, Hansen EE, Manchester JK, Coutinho PM, HenrissatB, Fulton R, Latreille P et al. Genomic and metabolic adaptationsof Methanobrevibacter smithii to the human gut. Proc Natl AcadSci USA 2007, 104: 10643−10648
70 Li T, Graham DE, Stathopoulos C, Haney PJ, Kim HS, VothknechtU, Kitabatake M et al. Cysteinyl-tRNA formation: the last puzzleof aminoacyl-tRNA synthesis. FEBS Lett 1999, 462: 302−306
71 Stathopoulos C, Kim W, Li T, Anderson I, Deutsch B, Palioura S,Whitman W et al. Cysteinyl-tRNA synthetase is not essential forviability of the archaeon Methanococcus maripaludis. Proc NatlAcad Sci USA 2001, 98: 14292−14297
72 White RH. The biosynthesis of cysteine and homocysteine inMethanococcus jannaschii. Biochim Biophys Acta 2003, 1624:46−53
73 Helgadottir S, Rosas-Sandoval G, Söll D, Graham DE. Biosynthesisof phosphoserine in the Methanococcales. J Bacteriol 2007, 189:575−582
74 Kavran JM, Gundllapalli S, O’Donoghue P, Englert M, Söll D,Steitz TA. Structure of pyrrolysyl-tRNA synthetase, an archaealenzyme for genetic code innovation. Proc Natl Acad Sci USA2007, 104: 11268−11273
75 Kamtekar S, Hohn MJ, Park HS, Schnitzbauer M, Sauerwald A, SöllD, Steitz TA. Toward understanding phosphoseryl-tRNACys
formation: the crystal structure of Methanococcus maripaludisphosphoseryl-tRNA synthetase. Proc Natl Acad Sci USA 2007,104: 2620−2625
76 Fukunaga R, Yokoyama S. Structural insights into the first step ofRNA-dependent cysteine biosynthesis in archaea. Nat Struct MolBiol 2007, 14: 272−279
77 Hohn MJ, Park HS, O’Donoghue P, Schnitzbauer M, Söll D. Emer-gence of the universal genetic code imprinted in an RNA record.Proc Natl Acad Sci USA 2006, 103: 18095−18100
78 Fukunaga R, Yokoyama S. Structural insights into the second stepof RNA-dependent cysteine biosynthesis in archaea: crystal struc-ture of Sep-tRNA:Cys-tRNA synthase from Archaeoglobus fulgidus.J Mol Biol 2007, 370: 128−141
79 Schneider G, Kack H, Lindqvist Y. The manifold of vitamin B6dependent enzymes. Structure 2000, 8: R1−6
80 Eliot AC, Kirsch JF. Pyridoxal phosphate enzymes: mechanistic,structural, and evolutionary considerations. Annu Rev Biochem2004, 73: 383−415
81 Zhou D, White RH. Transsulfuration in archaebacteria. J Bacteriol1991, 173: 3250−3251
82 Kitabatake M, So MW, Tumbula DL, Söll D. Cysteine biosynthesispathway in the archaeon Methanosarcina barkeri encoded by ac-quired bacterial genes? J Bacteriol 2000, 182: 143−145
83 Borup B, Ferry JG. Cysteine biosynthesis in the Archaea:Methanosarcina thermophila utilizes O-acetylserine sulfhydrylase.FEMS Microbiol Lett 2000, 189: 205−210
84 Boru p B , Ferry J G . O-Acetyl ser ine sul fhydryla se fromMethanosarcina thermophila. J Bacteriol 2000, 182: 45−50
85 Mino K, Ishikawa K. Characterization of a novel thermostable O-acetylserine sulfhydrylase from Aeropyrum pernix K1. J Bacteriol2003, 185: 2277−2284
86 Mino K, Ishikawa K. A novel O-phospho-L-serine sulfhydrylationreaction catalyzed by O-acetylserine sulfhydrylase from Aeropyrumpernix K1. FEBS Lett 2003, 551: 133−138
87 Mihara H, Esaki N. Bacterial cysteine desulfurases: their functionand mechanisms. Appl Microbiol Biotechnol 2002, 60: 12−23
88 Major TA, Burd H, Whitman WB. Abundance of 4Fe-4S motifs inthe genomes of methanogens and other prokaryotes. FEMSMicrobiol Lett 2004, 239: 117−123
89 Di Giulio M. The origin of the genetic code: theories and theirrelationships, a review. Biosystems 2005, 80: 175−184
90 Klipcan L, Frenkel-Morgenstern M, Safro MG. Presence of tRNA-dependent pathways correla tes with high cysteine content inmethanogenic Archaea. Trends Genet 2008, 24: 59−63
91 Rayman MP. The importance of selenium to human health. Lan-cet 2000, 356: 233−241
92 Moghadaszadeh B, Beggs AH. Selenoproteins and their impact onhuman health through diverse physiological pathways. Physiol-ogy 2006, 21: 307−315
93 Hatfield DL, Carlson BA, Xu XM, Mix H, Gladyshev VN.Selenocysteine incorporat ion ma chinery a nd the role ofselenoproteins in development and health. Prog Nucleic Acid ResMol Biol 2006, 81: 97−142
94 Huber RE, Criddle RS. Comparison of the chemical properties ofselenocysteine and selenocystine with their sulfur analogs. ArchBiochem Biophys 1967, 122: 164−173
95 Young PA, Kaiser, II. Aminoacylation of Escherichia coli cysteinetRNA by selenocysteine. Arch Biochem Biophys 1975, 171: 483−489
96 Shrift A, Bechard D, Harcup C. Utilization of selenocysteine by acysteinyl-tRNA synthetase from Phaseolus aureus. Plant Physiol1976, 58: 248−252
97 Müller S, Senn H, Gsell B, Vetter W, Baron C, Böck A. The forma-tion of diselenide bridges in proteins by incorporation ofselenocysteine residues: biosynthesis and characterization of (Se)2-thioredoxin. Biochemistry 1994, 33: 3404−3412
98 Johansson L, Gafvelin G, Arner ES. Selenocysteine in proteins-properties and biotechnological use. Biochim Biophys Acta 2005,1726: 1−13
99 Böck A, Forchhammer K, Heider J, Leinfelder W, Sawers G, VeprekB, Zinoni F. Selenocysteine: the 21st amino acid. Mol Microbiol1991, 5: 515−520
100 Gladyshev VN, Hatfield DL. Selenocysteine-containing proteinsin mammals. J Biomed Sci 1999, 6: 151−160
101 Kryukov GV, Castellano S, Novoselov SV, Lobanov AV, Zehtab O,Guigo R, Gladyshev VN. Characterization of mammalianselenoproteomes. Science 2003, 300: 1439−1443
102 Kryukov GV, Gladyshev VN. The prokaryotic selenoproteome.EMBO Rep 2004, 5: 538−543
103 Bilokapic S, Korencic D, Söll D, Weygand-Durasevic I. The un-usual methanogenic seryl-tRNA synthetase recognizes tRNASer
species from all three kingdoms of life. Eur J Biochem 2004, 271:694−702
104 Mizutani T, Narihara T, Hashimoto A. Purification and propertiesof bovine liver seryl-tRNA synthetase. Eur J Biochem 1984, 143:9−13
105 Leinfelder W, Zehelein E, Mandrand-Berthelot MA, Böck A. Genefor a novel tRNA species that accepts L-serine and cotranslationallyinserts selenocysteine. Nature 1988, 331: 723−725
106 Carlson BA, Xu XM, Kryukov GV, Rao M, Berry MJ, GladyshevVN , Hat fie ld DL. Ident ifi cat ion a nd characteri za tion ofphosphoseryl-tRNA[Ser]Sec kinase. Proc Natl Acad Sci USA 2004,101: 12848−12853
107 Kaiser JT, Gromadski K, Rother M, Engelhardt H, Rodnina MV,

Acta Biochim Biophys Sin (2008) | Volume 40 | Issue 7 | Page 553
Amino acid modifications on tRNA
Wahl MC. Structural and functional investigation of a putativearchaeal selenocysteine synthase. Biochemistry 2005, 44: 13315−13327
108 Leinfelder W, Forchhammer K, Veprek B, Zehelein E, Bock A. Invitro synthesis of selenocysteinyl-tRNAUCA from seryl-tRNAUCA:involvement and characterization of the selD gene product. ProcNatl Acad Sci USA 1990, 87: 543−547
109 Leinfelder W, Forchhammer K, Zinoni F, Sawers G, Mandrand-Berthelot MA, Böck A. Escherichia coli genes whose products areinvolved in selenium metabolism. J Bacteriol 1988, 170: 540−546
110 XM, Carlson BA, Irons R, Mix H, Zhong N, Gladyshev VN, HatfieldDL. Selenophosphate synthetase 2 is essential for selenoproteinbiosynthesis. Biochem J 2007, 404: 115−120
111 Sherrer RL, O’Donoghue P, Söll D. Characterization and evolu-tionary history of an archaeal kinase involved in selenocysteinyl-tRNA formation. Nucleic Acids Res 2008, 36: 1247−1259
112 Mäenpää PH, Bernfield MR. A specific hepatic transfer RNA forphosphoserine. Proc Natl Acad Sci USA 1970, 67: 688−695
113 Sharp SJ, Stewart TS. The characterization of phosphoseryl tRNAfrom lactating bovine mammary gland. Nucleic Acids Res 1977, 4:2123−2136
114 Leipe DD, Koonin EV, Aravind L. Evolution and classificationof P-loop kinases and related proteins. J Mol Biol 2003, 333:781−815
115 Hatfield D, Lee BJ, Hampton L, Diamond AM. Selenium induceschanges in the selenocysteine tRNA[Ser]Sec population in mamma-lian cells. Nucleic Acids Res 1991, 19: 939−943
116 Dong H, Nilsson L, Kurland CG. Co-variation of tRNA abun-dance and codon usage in Escherichia coli at different growthrates. J Mol Biol 1996, 260: 649−663
117 Wu XQ, Gross HJ. The long extra arms of human tRNA(Ser)Sec andtRNASer function as major identify elements for serylation in anorientation-dependent, but not sequence-specific manner. NucleicAcids Res 1993, 21: 5589−5594
118 Sturchler C, Westhof E, Carbon P, Krol A. Unique secondary andtertiary structural features of the eucaryotic selenocysteinetRNASec. Nucleic Acids Res 1993, 21: 1073−1079
119 Schön A, Bock A, Ott G, Sprinzl M, Söll D. The selenocysteine-inserting opal suppressor serine tRNA from E. coli is highlyunusual in structure and modification. Nucleic Acids Res 1989,17: 7159−7165
120 Wu XQ, Gross HJ. The length and the secondary structure of theD-stem of human selenocysteine tRNA are the major identitydeterminants for serine phosphorylation. EMBO J 1994, 13:
241−248121 Sherrer RL, Ho J, Söll, D. Divergence of selenocysteine tRNA
recognition by archaeal and eukaryotic O-phosphoseryl-tRNASec
kinase. Nucleic Acids Res 2008, 36: 1871−1880122 Baron C, Westhof E, Böck A, Giege R. Solution structure of
selenocysteine-inserting tRNASec from Escherichia coli. Com-parison with canonical tRNASer. J Mol Biol 1993, 231: 274−292
123 Hubert N, Sturchler C, Westhof E, Carbon P, Krol A. The 9/4secondary structure of eukaryotic selenocysteine tRNA: morepieces of evidence. RNA 1998, 4: 1029−1033
124 Ioudovitch A, Steinberg SV. Structural compensation in an archaealselenocysteine transfer RNA. J Mol Biol 1999, 290: 365−371
125 Baron C, Sturchler C, Wu XQ, Gross HJ, Krol A, Böck A. Eukary-otic selenocysteine inserting tRNA species support selenoproteinsynthesis in Escherichia coli. Nucleic Acids Res 1994, 22: 2228−2233
126 Araiso Y, Palioura S, Ishitani R, Sherrer RL, O’Donoghue P,Yuan J, Oshikane H et al. Structural insights into RNA-dependenteukaryal and archaeal selenocysteine formaiton. Nucleic AcidsRes 2007, 36: 1187−1199
127 Ganichkin OM, Xu XM, Carlson BA, Mix H, Hatfield DL,Gladyshev VN, Wahl MC. Structure and catalytic mechanism ofeukaryotic selenocysteine synthase. J Biol Chem 2007, 283:5849−5865
128 Zhang Y, Romero H, Salinas G, Gladyshev VN. Dynamic evolu-tion of selenocysteine utilization in bacteria: a balance betweenselenoprotein loss and evolution of selenocysteine from redoxactive cysteine residues. Genome Biol 2006, 7: R94
129 Stanzel M, Schön A, Sprinzl M. Discrimination against misacylatedtRNA by chloroplast elongation factor Tu. Eur J Biochem 1994,219: 435−439
130 Rother M, Wilting R, Commans S, Böck A. Identification andcharacterisation of the selenocysteine-specific translation fac-tor SelB from the archaeon Methanococcus jannaschii. J MolBiol 2000, 299: 351−358
131 Roy H, Becker HD, Mazauric MH, Kern D. Structural elementsdefining elongation factor Tu mediated suppression of codonambiguity. Nucleic Acids Res 2007, 35: 3420−3430
132 Min B, Kitabatake M, Polycarpo C, Pelaschier J, Raczniak G,Ruan B, Kobayashi H et al. Protein synthesis in Escherichia coliwith mischarged tRNA. J Bacteriol 2003, 185: 3524−3526
133 Sheppard K, Yuan J, Hohn MJ, Jester B, Devine KM, Söll D.From one amino acid to another: tRNA-dependent amino acidbiosynthesis. Nucleic Acids Res 2008, 36: 1813−1825