aldose reductase: a multi-disease target
TRANSCRIPT

Aldose Reductase: A Multi-disease Target
Jaiprakash N Sangshetti*1, Rashmi S. Chouthe1, Nikhil S. Sakle1,Indrajeet Gonjari2, Devanand B Shinde3
1 Y. B. Chavan College of Pharmacy, Dr. Rafiq Zakaria Campus, Rauza Baugh,Aurangabad-431001
2 Government College of Pharmacy Karad (MS)
3 Department of Chemical Technology, Dr. B. A. M. University, Aurangabad-431004
* Correponding Author email: [email protected]
Abstract:
Aldose reductase (AR) is an enzyme that catalyzes the rate
limiting step of the polyol pathway of glucose metabolism,
besides reducing glucose to sorbitol, reduces a number of lipid
peroxidation derived aldehydes and their glutathione conjugates.
Recent studies suggest that apart from its involvement in
diabetic complications, AR's catalytic activity plays a key role
in a number of inflammatory diseases such as atherosclerosis,
sepsis, asthma, uveitis, and ovarian cancer. Furthermore, AR is
over expressed in human cancers such as liver, colon, breast, and
cervical. AR inhibitors have undergone up to phase-III clinical
trials for diabetic complications, they are also reported as safe
anti-inflammatory drugs. Therefore the future use of AR
1

inhibitors in down-regulating major multidiseases and
inflammatory pathologies such as cancer and cardiovascular
diseases could relieve some of the major health concerns of
worldwide.
Key words: Aldose reductase inhibitors, Diabetic neuropathy,
inflammatory pathologies, Polyol pathway.
Introduction-
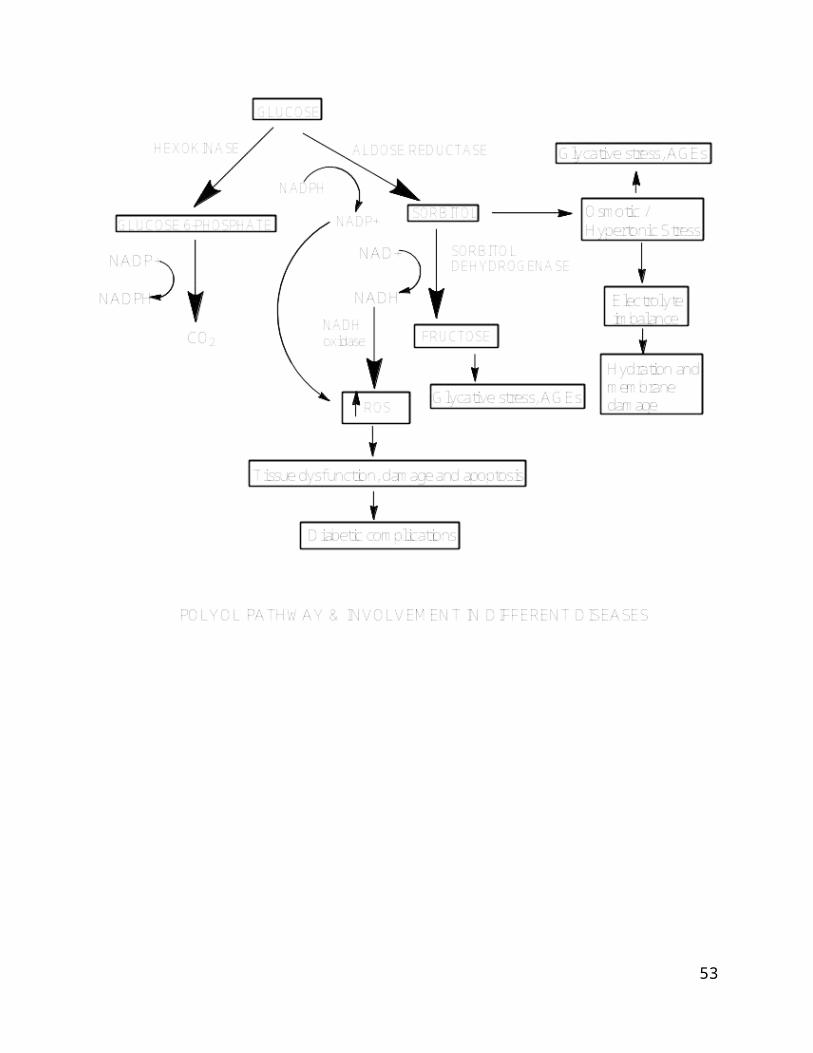
Glucose is metabolized mostly in vivo by the glycolytic system, but
an extremely small part is metabolized through the polyol pathway
(Fig 1). In polyol metabolic pathway Aldose reductase (AR)
catalyzes the reduction of glucose to sorbitol with NADPH as a
cofactor. The metabolism of glucose consists of two steps, in
which glucose is converted into sorbitol and further into
fructose. The rate-limiting enzyme of this pathway is AR, which
belongs to the aldo-keto reductase family [1,2], AR is
distributed widely throughout the body, including areas such as
2

the adrenal gland [3], arterial smooth muscle[4], ovary[5] ,
testis[6] and target organs of diabetic complications such as
the lens, retina, kidney [3,5] and peripheral nerves [5,6]. A
physiological roles, AR regulates osmotic pressure in the kidney
medulla [7], steroid metabolism in the adrenal gland and
reproductive organs [4,8] detoxication of aldehyde compounds such
as 4-hydroxynonenal (HNE) in the vascular wall in case of giant
cell arteritis [9], and regulation of harmone in the ovary [8].
In a hyperglycemic state, the glucose metabolism through the
polyol pathway is accelerated [2]. Activation of AR enzymes
causes accumulation of sorbitol in tissues which does not
diffuses through cell, leads to various diabetic complications
and also involved in the pathogenesis of CVS, CNS, cancer and in
some of the inflammatory conditions.
In addition, the excessive consumption of NADPH associated with
the enhanced enzymatic reaction of AR results in suppression of
NO synthesis [13] and reduced glutathione (GSH) production [14].
The decrease in the amount of NO, which acts as a vasodilating
mediator, causes circulatory disorders. The decrease in the
amount of GSH leads to an increase in reactive oxygen species3

(ROS), resulting in impairment of endothelial function. Hence,
acceleration of the polyol pathway metabolism of glucose is an
important factor in the onset and progression of diabetic
complications such as neuropathy, vascular disorders and other.
Thus, various AR inhibitors have been developed to correct this
metabolic abnormality.
A number of cardiac disorders, including myocardial ischemia,
ischemia–reperfusion injury, congestive heart failure, cardiac
hypertrophy and cardiomyopathy have been linked to increased ROS
generation and lipid peroxidation within myocardium and studies
have been conducted in order to determine the role of aldose
reductase (ALR2) in myocardial metabolism during ischemia [15].
Furthermore, ALR2 has been reported to be implicated with
inflammation [16, 17], mood disorders [18], renal insufficiency
[19], ovarian abnormalities [20] and human cancers such as liver,
breast, ovarian, cervical and rectal cancers [21, 22].
Considering the involvement of AR in etiology of various diseased
conditions of 21st century, intense efforts have been taken to
discover ARI of distinct chemical structures.
Aldose reductase structure-4

ALR2 is a 316 amino acid enzyme and its molecular weight is found
to be 36 kDa. The primary structure of ALR-2 was first determined
in rat lenses [23]. In 1992, the first two ALR2 structures were
published; the human placenta ALR2 [24] and that of porcine ALR2
[25]. These structures were composed of a similar α/β-barrel with
a core of eight parallel β-strands connected by peripheral α-
helices. The active site of AR is located at the C-terminal face
of the barrel which is apt for efficient interaction with NADPH,
a cofactor required for AR’s reduction reactions. The
nicotinamide ring was centered in the active site cavity. This
highly hydrophobic active site pocket is formed by aromatic
residues (Trp20, Tyr48, Trp79, Trp111, Phe121, Phe122 and
Trp219); apolar residues (Val47, Pro218, Leu300 and Leu301);
polar residues (Gln49, Cys298and His110); and three possible
proton donors, which were Tyr48, His110 and Cys298, were also
identified. This protein, which is called aldose reductase-like-1
protein, AKR1B10 or ARL-1 having same size as ALR2, and its amino
acid sequence is 70% identical to that of ALR2. Apart from the
amino acid sequence similarity, all of the key amino acids
proposed to be responsible for ALR2 enzymatic activities are
5

conserved in this protein. They include Ser159, Asn160, Gln183,
and Lys262 for cofactor NADPH binding, Tyr48 and His110 as
potential hydrogen donors, and active site residues Lys77,
Tyr209, and Cys298. Therefore, similar to ALR2, ARL-1 can
efficiently reduce aliphatic and aromatic aldehydes, and it is
less active on hexoses. While ALR2 mRNA is found in most tissues
studied, ARL-1 is primarily expressed in the small intestines and
in the colon, with a low level of its mRNA in the liver, thymus,
prostate and testis. Since ARL-1 is predominantly found in the
small intestines and in the colon, it is likely to be responsible
for inactivating toxic aldehydes in digested food before the
nutrients are transported to other organs [26,91,96].
Role of ARIs in diabetic complications-
ARIs in diabetic neuropathy-
Neuropathic pain can be described as a sensation of paresthesia
and burning that is caused by the sustained, abnormal processing
of CNS neuronal input [27]. The constant pain can lead to other
health issues, such as constant fatigue, depression and anxiety
which decrease the quality of life in these patients. Neuropathy
can occur both from damage to the nerve parenchyma and from
6

neuronal ischemia leading to abnormalities of microvessels [28].
In the peripheral nerves, the decrease of myo-inositol content
is concerned with the excessive accumulation of sorbitol [10],
which leads to changes in Na+, K+, ATPase pump activity present
on the axonal membrane [11,12]. These changes result in the
intra-axonal Na+ accumulation and reduction in trans-membrane Na+
conduction. Impairment in the function alters the membrane
potential that ultimately result in the axonal death [92].
Vincent et al. [29] pointed the major role of oxidative stress in
the pathogenesis of diabetic neuropathy. Oxidative stress may be
derived from excessive flow through the polyol pathway or advance
glycation end products (AGE), mediated protein kinase C (PKC)
activation and uncoupling of the respiratory chain in the
mitochondria. The polyol pathway plays an important role in the
development and progression of diabetic neuropathy.
ARI in diabetic retinopathy-
Diabetic retinopathy is a common cause of vision loss which is
characterized by retinal capillary cell loss and formation of
acellular capillaries, capillary basement membrane thickening,
increased vascular permeability and disruption of blood-retinal
7

barrier, and increased leukocyte adhesion to endothelial cells
[95]. The selective destruction of retinal pericytes has been
linked to the production of sorbitol and galacticol [30].
Accumulated sorbitol within retina may cause osmotic stress,
resulting in excessive hydration, gain of Na+ and loss of K+ ions
and also the byproducts of polyol pathway, fructose-3-phosphtae
and 3-deoxyglucosone are powerful glycosylating agents that enter
in the formation of AGEs, which are an important factor for the
pathogenecity of diabetic retinopathy [93]. The use of ARIs in
many animal models has prevented the early activation of
complement in the wall of retinal vessels, apoptosis of vascular
pericytes and endothelial cells and the development of acellular
capillaries [31]. Osmotic stress in the ocular lens deranges the
cell membrane, resulting in the leakage of amino acids, GSH and
myo-inositol to induce cataract formation [32]. Additionally,
ALR2 is implicated with diabetic retinopathy via a plethora of
signaling pathways such as the activation of nuclear factor kappa
B (NF-kβ) and further regulating the monocyte chemotactic
protein-1 (MCP-1) which contributes to the formation of diabetic
epiretinal membranes [33]. Also in the retina, increased ALR2
8

activity enhances oxidative stress and upregulates retinal
vascular endothelial growth factor (VEGF). Another implication of
ALR2 in diabetic retina is considered to be through the
activation of poly (ADP-ribose) polymerase (PARP). Activation of
PARP tends to be a mechanism in the pathogenesis of diabetic
retinopathy, endothelial and myocardial dysfunction and
peripheral neuropathy [34, 35]. The use of fidarestat, an ARI
counteracts diabetes-associated retinal oxidative-nitrosative
stress and PARP formation supporting an important role for AR in
diabetes and rationale for the development of ARIs for
counteraction of polyol pathway [94].
ARI in diabetic nephropathy-
Renal polyol accumulation has also been suggested to be involved
in diabetic nephropathy [36]. Interaction between hyper-glycemia-
induced oxidative stress from AR activation, increases the
formation of AGEs and activation of vascular PKC iso-forms
ultimately result in microvascular diabetic complications. PKC
activation may induce an increase in glomerular prostaglandin E
production and loss of mesangial cell contractility leading to
hyperfiltration and glomerular dysfunction [37]. Furthermore, the
9

decrease in glomerular filtration rate may implicate the renal
VEGF, since its levels increase in preclinical models of diabetes
[28]. A number of ARIs like tolrestat, sorbinil and ponalrestat
have been studied in diabetic animals. Oral administration of
tolrestat in galactosemic animals revert capillary lesions,
pericyte degeneration, endothelial cell proliferation,
accellularity, capillary dilation, and microaneurysm formation.
Sorbinil and ponalrestat, were reported to prevent glomerular
basement membrane thickening (GBMT) and albuminuria development
in diabetic rats [96-98]. Clinical studies of ARIs in the
diabetic complications delayed the progression of symptoms of the
disease also showed reduced urinary albumin excretion rate after
treatment for 6 months or 5 years [99-100].
Role of ARI in non-diabetic diseases-
ARI in inflammatory pathogenesis-
AR is mainly involved in the metabolizing glucose, but it also
shows its involvement in catalyzing the reduction of a wide range
of substances such as lipid aldehydes generated during lipid
peroxidation and their glutathione (GSH)-conjugates,
phospholipids, atherogenic aldehydes, and steroids [101]. During
10

the inflammation a pro-inflammatory endotoxin lipopolysacharides
(LPS) and component of gram negative bacterial cell wall is
released in the circulation and stimulates toll like receptor-4
(TLR-4) [38]. LPS is interact with cluster of differentiation 14
(CD 14) and it leads to increase transcription of inflammatory
and immune response genes, activation of NK-kB and further
stimulate IL-1, TNK-α, PG and NO [39]. ALR2 inhibition suppressed
NF-kB mediated transcription in LPS treated cells of macrophage,
and further reduced the production of IL-1 and TNK-α. There are
reports that, apart from the down regulation of NF-kB also
indicate as possible mechanisms the modulatory effect on
prostaglandin E2 production, cycloxyganase-2 (COX-2) activity and
PKC pathways [40], reduced production of NO and suppression of
the LPS-induced over expression of inducible NO synthetase mRNA
[41]. Inhibition of ALR2 represents a useful approach for
prevention and treatment of ocular inflammatory response such as
uveitis [42] and endotoxin-related inflammatory diseases [43].
Aldose reductase inhibitors (AKR1B1) give a novel therapeutic
approach to treat inflammatory airways diseases [89].
ARIs in ischemic conditions-
11

Earlier studies have demonstrated that AR plays a key role in
mediating ischemia-reperfusion (I/R) injury [104]. During low-
flow ischemia there is increase in the NO level. Some of the
studies have shown that NO donors increase AR activity in
cultured cells. The cascades of events like increase in cytosolic
NADH/NAD+ ratio and decrease in glycolysis and ATP level leads to
ischemic injury [101]. Also ischemia injury causes accumulation
of sorbital intermediates causes cell death [102]. Hwang et al.
studied the central role of AR in transgenic mice expressing
human AR (ARTg) during ischemia reperfusion injury. The data
strongly support the pharmacological inhibition of AR in ARTg
mice. It reduced cytosolic NADH/NAD+ ratio and further
consequence, also reduced ischemic injury and improved functional
recovery [103]. The expression of AR is regulated by a number of
transcription factors, such as osmotic response element binding
protein (OREBP) and NF-kB, by binding to the AR promoter possibly
under osmotic or oxidative stress conditions associated with
ischemia [44,45,46].
Some of the studies have shown the influence of ischemia on
cardiac ALR2 activity, also the effect of flux via the polyol
12

pathway on myocardial glucose metabolism [47,48]. Pharmacological
inhibition of ALR2 could be cardio-protective, as it seems to be
a key component of myocardial ischemic injury, and could reverse
the increased NADH/NAD+ ratio (as a result of reduced NAD+ use by
sorbitol dehydrogenase SDH) and therefore improving myocardial
glucose metabolism by increasing glycolysis and ATP production.
Additionally, ARIs partially augment intracellular sodium and
calcium during ischemia, secondary to increased glucolytic
metabolism and the observed changes in intracellular sodium were
partly attributable to increases in Na+, K+-ATPase activity.
Therefore, it was suggested that ARIs could be used
therapeutically in treating patients with myocardial infarction.
ARIs applied during the pre-ischemic phase improved cardiac
performance and attenuated myocyte damage, while inhibitors
applied during the reperfusion phase had limited effect on
cardiac performance but significantly attenuated myocyte damage
by reducing oxidative stress elicited by ischemic insult and
reoxyganation. Therefore, the use of ARIs in the post ischemic
reperfusion phases protecting ischemic heart from ischemic
13

reperfusion injury. ALR2 in the ischemic heart may play a role in
the protection against ROS produced during ischemia reperfusion.
Proliferation of vascular smooth muscle cells (VSMCs) is one of
the key features of atherogenesis, restenosis, and hypertension.
It is characterized by endothelial dysfunction due to
cardiovascular risk factors or mechanical injury, resulting in
the expression of several growth factors and cytokines that exert
mitogenic effects on VSMCs [49]. Inhibition of the enzyme
prevented DNA synthesis and cell growth in culture and intimal
hyperplasia in vivo. The absence of ALR2 in inactive VSMCs and its
high expression in VSMCs proliferating in culture suggested that
the enzyme is specifically up regulated during growth. The
association of ALR2 with cell growth was further supported by the
observation that after balloon injury, the proliferating cells of
the neointima showed high levels of expression of this enzyme.
The mechanism of the regulatory effect of ALR2 on cell growth was
further studied and it was found that ARIs prevented the tumor
necrosis factor-α (TNF-α) induced activation of NF-kB, with final
result the attenuation of VSMCs proliferation not by increasing
cell death but by inhibiting their growth [50].
14

ARI in Renal insufficiency-
ALR2 had an effect on transforming growth factor (TGF) β1-induced
synthesis of fibronectin and type IV collagen in the mesangial
cells under normoglycemia [51]. This could be due to the
modulation of activation of c-Jun N-terminal kinase (JNK) and p38
signal transduction pathways and transcription factor AP-1. ARIs
such as sorbinil or zopolrestat reduced the activation of JNK, p-
38 MAPK and AP-1 induced by TGF-β1. The production of
extracellular matrix (ECM) components in the mesangial cells
under normal glucose concentration by ALR2, suggested that it
might play a role in pathogenesis of nondiabetic
glomerulosclerosis. ECM proteins deposition and obliteration of
glomerular capillaries, is considered to be the final pathway
leading to the progressive loss of renal function in several
kidney diseases. The increased ALR2 activity in such cases may
affect the peritoneal membrane from the vascular side in patients
undergoing peritoneal dialysis. In these cases injury of the
peritoneal membrane with structural changes and functional
decline, such as ultra filtration occur. Further studies are
needed to clarify the role of ALR2 and the potential protective
15

role of its inhibition in patients with chronic kidney diseases
treated with peritoneal dialysis [52]. Increases in the AGE–RAGE
axis have been demonstrated in renal failure both in animal
models and in human subjects. Large AGE proteins unable to enter
the Bowman’s capsule are capable of binding to receptors on
endothelial and mesangial cells and to the mesangial matrix.
Activation of RAGE induces production of a variety of cytokines,
including TNFβ, which mediates an inhibition of metalloproteinase
and increases production of mesangial matrix, leading to renal
failure [105].
ARI in ovarian diseases-
The mammalian ovary is susceptible to damage from the
accumulation of galactose and galactose metabolites like Gal-1-P,
galactitol, and UDPgal, due to unusually high local
concentrations of enzymes responsible for its metabolism [53].
Apart from its major metabolic pathways that include the enzymes
galactokinase, galactose-1-phosphate uridyltransferase (GALT),
and UDP-galactose-4-epimerase, the polyol pathway is also
involved and therefore galacticol is produced. Galacticol is
poorly metabolized and its accumulation leads to ovarian toxicity
16

due to osmotic imbalance and GSH depletion. At this point it must
be noted that the affinity of ALR2 for galactose is relatively
low, and high intracellular concentrations of galactose are
required as in galactosemia, an inherited disorder due to GALT
deficiency [54]. ARIs have been observed to cause a reversal of
galactose-induced toxicity in animals via the reduction in
galactitol levels. They have been found to prevent ovarian
dysfunction in rats fed with high galactose diet [55].
ARI in cancer-
It was found that about 29% of the liver cancers over expressed
ALR2, and about 54% of them over expressed a novel ALR-1 [55].
The incidence of the ALR2 over expression and activation in these
cases was suggested to render cancer cells resistant to
anticancer drugs and that ARIs could reserve this resistance.
Resistance towards anticancer drugs, such as the anthracycline,
daunorubicin and doxorubicin, is a major problem in the
chemotherapy of cancer and it may be due to biotransformation
enzymes, such as ALR2, that catalyze carbonyl reduction of
anthracyclines into their inactive alcohol metabolites [56]. ALR2
17

up regulation can be considered as one non classical mechanism of
resistance to chemotherapy.
Colon cancer is a complex multi step process involving
progressive disruption of homeostatic mechanisms controlling
intestinal epithelial proliferation, differentiation, and
programmed cell death. One of the main features of colon cancer
is the over expression of the inducible COX-2 [57] that catalyzes
the first two steps in the biosynthesis of prostaglandins from
arachidonic acid. The de novo synthesis of COX-2 is triggered by
the exposure of cells to certain stimuli, such as cytokines and
growth factors, which lead to induction of altered synthesis of
prostaglandins followed by uncontrolled colon epithelial cell
proliferation. A major signaling pathway associated with the
oxidative stress and inflammation is the activation of NF-kβ by
growth factors (such as fibroblast derived growth factor bFGF and
platelet-derived growth factor PDGF) or inflammatory cytokines
(such as TNF-β). Modulation of NF-kβ may play a central PDGF role
in the mitogenic process initiated by ROS and related oxidants in
colon cancer. Additionally, the expression of ALR2 is enhanced in
various types of cancer as previously mentioned. Therefore, the
18

role of ALR2 in mediating the growth factor induced expression of
COX-2 and production of prostaglandin E2 was investigated
[58,59]. Inhibition of ALR2 significantly (>90%) prevented the
production of prostaglandin E2 by induced by PDGF. Finally, it
was demonstrated that ALR2 is an obligatory mediator of growth
factor induced colon cancer cell proliferation and its inhibition
prevents the S phase of cell cycle, which is an important stage
required for cell growth. In addition another study was conducted
[60] in order to clarify the role of ALR2 in mediating the TNF-a-
induced production of prostaglandin E2 in colon cancer Caco-2
cells. In conclusion, ALR2 was found to be an obligatory mediator
of the inflammatory changes caused by growth factors- and
inflammatory cytokines- induced cytotoxicity in colon cancer and
therefore ALR2 inhibition may represent a novel therapy for colon
cancer. Adrenal tumors are divided into benign adenomas, which
are extremely frequent, and rare malignant carcinomas, accounting
for 0.05–0.2% of all cancers and with an extremely poor
prognosis, the survival rate being 20% at 5 years [61]. As the
adrenal gland is a major site of expression of ALR2, a study was
contacted in order to show whether changes in its expression
19

could be associated with adrenal disorders [62]. It was found
that ALR2 gene is differentially expressed in benign vs.
malignant tumors in adrenal cortex and that ALR2 gene expression,
measured at mRNA and protein levels, is strongly decreased in
adrenal cortex carcinomas, compared with that in adenomas. The
low expression of ALR2 in carcinomas may be due to chr omosome
alterations, including mutations or rearrangements that could
reduce expression of the gene or due to deregulation of the
mechanisms underlying the control of the gene expression and it
was the first time that it is suggested that cAMP is a regulator
of ALR2 expression in human adrenocortical cells. Additionally,
it was reported that the transcription factor cAMP-responsive
element-binding protein (CREB) was decreased at the protein level
in adrenal cortex carcinomas [63], and cAMP- stimulated PKC
activity is lower in adrenal cancers than adenomas [64]. It must
be noted that the decrease in ALR2 expression in malignant tumors
cannot be considered either as a consequence of a general
dedifferentiation of the tumor or it contributes to the
pathogenesis of this disease by promoting alterations in cell-
cell communication through progressive loss of gap junctions.
20

Together with ALR2, another protein is often correlated with
cancers and this is ALR1 protein and therefore should be
mentioned at this point. This novel human protein that is highly
homologous to ALR2 was identified by Cao et al. [65]. A novel
function of ALR-1 in tumorgenic mammary epithelial cells, that
involves regulating fatty acid synthesis, was reported. These
data suggested that ALR-1 is a novel regulator of the
biosynthesis of fatty acids, which are essential components of
the cell membrane.
Chemical classification of ARIs-
1) Compounds containing five-membered cyclic imides such as
spirohydantoins [66].
2) Carboxylic acid derivatives like alrestatin, tolrestat and
zopolrestat [67, 68]
3) Structurally diverse class
4) ARI from natural sources
5) Dietary sources of ARI
1] Cyclic imides-
Sorbinil (1) was the first cyclic imidine (spirohydantoin)
compound, which inspected extensively on the inhibition of AR
21

enzyme. One of the main adverse effects of sorbinil was
hypersensitivity reaction in the early weeks of therapy, which is
most likely emerged from hydantoin moiety. To solve the
hypersensitivity problem, compounds possessing thiazolidinedione
moiety were introduced as hydantoin bioisosters in which the lack
of hypersensitivity was eliminated. [69]. Compounds derived from
sorbinil are methosorbinil (methyl derivative of sorbinil, M
79175), fidarestat (2) (an amide analog of sorbinil, SNK860), and
minalrestat (3) (2-fluoro, 4-bromo benzyl derivative, ARI 509).
Among the latest sorbinil derivatives, fidarestat shows ten-fold
higher potency than sorbinil having some promising advantages
including normalization of erythrocytic sorbitol contents in
neuropathy patients and showing no significant side effects, even
after a year of continuous administration [70], it is still not
being recognized worldwide as an effective inhibitor in the
treatment of diabetic complications, especially in neuropathy.
Due to the fact that thiazolidinedione (4) bearing compounds have
shown antihyperglycemic efficacy [71], and also considered as a
bioisoster of hydantoin moiety (5), still thiazolidinedione is
favored for the AR inhibition [72]. However, a point should be
22

made about hydantoins, as unacceptable side effects related to
toxicity (skin rash and hypersensitivity or liver toxicity) have
rendered them undesirable as drugs [73].
2] Carboxylic acid derivative-
Compounds, which convey flexible carboxylic acid moiety and have
certain requirements for binding to AR enzyme, are found to be of
a great interest within the structural variations to demonstrate
possible interactions with AR enzyme through inhibition. The main
aspect was the integration of minimal structural requirements
with flexible carboxylic acid moiety, which is believed to hold
an important role in the interaction with AR enzyme, especially
in physiological conditions. Alrestatin (6) can be considered as
a leading structure of carboxylic acidtype of compounds similar
to those sorbinil for cyclic imidines. Therefore, since
alrestatin has shown quite remarkable inhibitory activity, a
number of new developing have been trailed including rhodanine
acetic acid derivatives (9) (ONO-2235, epalrestat) [74],
quinolineacetic acids [75], and phthalazinoacetic acids (10)
(ICI-128,436, ponalrestat, statil) [76]. Epalrestat showed an
antiproliferative and antihypertrophic effects on vascular smooth
23

muscle cells induced by high glucose and suppressed intracellular
NADH/NAD+ elevation and reduced the membrane-bound protein kinase
C activation [77]. Tolrestat was introduced into the market in
1989 but was removed in 1996 due to its low of efficacy. The
exchange of acetic acid moiety with benzyl resulted in a drastic
drop of activity. This may show that the flexible acid moiety
might be placed in a position that could serve the interaction
through the enzyme’s binding site. The inclusion of halogen atoms
into the benzyl ring increased the overall potency either due to
their hydrogen bonding properties or the increasing lipopilicity.
Besides this methoxy group at the same ring also served the raise
in activity. It is reported that these compounds showed better
activity pattern than sorbinil. One of the halogenated benzyl
bearing quinazolidinedione compounds is FK-366 (11) (zenarestat),
which exhibits potent AR inhibitory activity and prevents the
development of diabetic complications.
3] Structurally diverse class of ARI-
other most characteristic chemotypes refer to
tetrahydropyrrolo[1,2-a]pyrazine-1,3-dione derivatives [78], 5-
arylidene- 2,4-thiazolidinedione derivatives [69], N-
24

nitromethylsulfonanilide derivatives [79], sulfonylpyridazinone
derivatives [80], N-(3,5-difluorophenol-4-hydroxyphenyl)
derivatives [81,82] and carboxymethylated pyridoindole
derivatives [83].
i) Tetrahydropyrrolo[1,2-a]pyrazine-1,3-dione Derivatives
Tetrahydropyrrolo-pyrazine derivatives (scaffolds a,b,c) were
evaluated for their ability to inhibit porcine lens ALR2 [78].
The most effective ARI from this series of chemotypes was the
inhibitor ranirestat. A quantitative-structure activity
relationship (QSAR) analysis was conducted by Ko et al. [79] in
order to aid the lead optimization and enhance potency and
deliverability of this series of compounds. For this QSAR study
a model derived from racemic descriptors (RS). A five term
RSequation showed that the hydrophobic character of the benzyl
moiety is the major contributing factor to the ALR2 inhibitory
activity [84].
ii) 5-arylidene-2,4-thiazolidinedione Derivatives
This series of compounds involved 5-arylidene-2,4-
thiazolidinediones with various substitution patterns on the
benzylidene moiety. The most active ARI with substitution R-
25

CH=NC6H4OH and R1- H, exerted significant inhibitory activity
(IC50=1.86μM), lying in the same range of sorbinil. In another
report, a SAR analysis [85] resulted in significant information
for 5-arylidene-2,4-thiazolidinediones, acting as efficient
ARIs. It was shown that the introduction of an acetic chain (R1-
CH2COOH) on N-3 of the thiazolinidione ring led to a marked
increase in inhibitory potency, while the presence of an
additional aromatic ring on the 5-benzylidene group increased
the ALR2 inhibitory activity. A reported quantitative structure-
activity relationship (QSAR) study [86] was contacted and from a
multivariant regression it was indicated that substituents, such
as 3- CH3; 3-OC6H5; 3-OCH3; 3-CF3; 4-OCH3; 4-CF3; 4-CH2COOCH3 or
CH2COOH, have poor contribution to the ALR2 inhibitory activity,
which was further supported by the high standard error of the
substituent coefficients in the equation. It was also suggested
that the binding of ARIs with ALR2 was accomplished by means of
a possible hydrogen bond interaction between the acetic acid
moiety of the side chain of thiazolidinediones and the polar
region of the ALR2 active site. The 3-position of the phenyl
26

ring depicted the hydrophobic interaction of the substituted
moiety and the lipophilic pocket of ALR2.
iii) N-nitromethylsulfonanilide Derivatives
A SAR analysis demonstrated that all of the compounds with a
methyl moiety substituted on the nitrogen atom which bears the
nitro methyl sulfonyl group, exhibited better inhibition activity
than the respective non-methyl substituted ones. Replacement of
the methyl group with an ethyl moiety decreased the inhibitory
activity. Similar results were observed when bulky substituents
were substituted on the benzene ring. The bulky subsituents are
interfering with the H-bond interactions between the
nitromethylsulfonyl moiety and proton donor moieties of the
enzyme.
iv) Sulfonylpyridazinones Derivatives
A SAR analysis for scaffold (f), showed that electron-withdrawing
(-F and -Cl) substituents that are reasonably lipophilic gave
potent compounds both in vitro and in vivo. Electron-releasing
substituents (-OCH3 and -CH3) and bulky groups, especially at the
2-position, had unfavorable effect on in vitro potency. Increasing
the length of the linker by one methylene group as in scaffold
27

(g), as well as several halogen-substituted compounds were found
to be more potent in vitro than the best inhibitors in the phenyl
series but less potent in vivo. As regards scaffold (h), the most
potent inhibitors were the ones with a benzofuran moiety.
v) Carboxymethylated Pyridoindole Derivatives
Carboxymethylated tetrahydropyridoindoles and
hexahydropyridoindoles (scaffolds i and l respectively),
structurally based on the antioxidant drug stobadine were
presented as novel zwitterionic ARIs. It was found that the
tetrahydropyridoindole derivatives were more potent ARIs in
comparison to the respective hexahydropyridoindole derivatives.
4] ARI from the natural sources-
Different variety of structurally diverse compounds has been
identified so far from terrestrial, marine and microorganism as
potent in vitro ARIs. ARIs from natural sources are usually
involved, the flavonoids and vitamin C [87] also series of
alkaloids including nandazurine, aporphine, benzylisoquinoline ,
papaverine , berberine [88] have been tested as inhibitors of AR.
28

Also some dietary sources were studied for AR inhibitory activity
and prominent inhibitory activity was found in spinach, cumin,
fennel, lemon, basil and black pepper [90].
Conclusion-
AR is enzyme that has proved its involvement in different
condition like inflammation, asthma, uvitis, cancer and number of
diabetic complications. Also these disease conditions have needs
continuously different new approaches for their treatment. The
enzyme has well studied and has gained a great deal of attention
in last few years. Some major clues about the structure of enzyme
results in development of new effective ARIs. Some of them are in
clinical development. Variety of chemical classes has been
explored as ARIs and used in number of above mentioned diseased
conditions. Some more insight on the structure is needed so that
new molecules with specific inhibitory activity can be developed.
The utility of some of the ARIs in diabetic complications is well29

established with some molecules in clinical development. However
its use in non-diabetic complications needs to be explored.
References-
[1] Nishimura-Yabe, C. Aldose reductase in the polyol
pathway: a potential target for the therapeutic intervention
of diabetic complications. Nihon Yakurigaku Zasshi., 1998,
111, 137-45.
30

[2] Kinoshita, J.H.; Nishimura, C. The involvement of
aldose reductase in diabetic complications. Diabetes Metab.
Rev., 1988, 4, 323–337.
[3] Matsuura, K.; Deyashiki, Y.; Bunai, Y.; Ohya, I.; Hara,
A. Aldose reductase is a major reductase for
isocaproaldehyde, a product of side-chain cleavage of
cholesterol, in human and animal adrenal glands. Arch
Biochem Biophys., 1996, 328, 265-71.
[4] Kern, T.S.; Engerman, R.L. Immunohistochemical
distribution of aldose reductase. Histochem J., 1982, 14,
507-15.
[5] Svanberg, B.; Ling, C.; Svensson, P.A.; Johnson, M.;
Carlsson, B.; Billig, H. Isolation of differentially
expressed aldose reductase in ovaries after estrogen
withdrawal from hypophysectomized diethylstilbestrol treated
rats: increased expression during apoptosis. Mol. Cell
Endocrinol., 2000, 164, 183-90.
[6] Ludvigson, M.A.; Sorenson, R.L. Immunohistochemical
localization of aldose reductase. I. Enzyme purification and
31

antibody preparation-localization in peripheral nerve,
artery, and testis. Diabetes, 1980, 29, 438-49.
[7] Burg, M.B. Molecular basis of osmotic regulation. Am.
J. Physiol., 1995, 268, 983–996.
[8] Wermuth, B.; Monder, C. Aldose and aldehyde reductase
exhibit isocorticosteroid reductase activity. Eur. J.
Biochem., 1983, 131, 423-6.
[9] Rittner, H.L.; Hafner, V.; Klimiuk, P.A.; Szweda, L.I.;
Goronzy, J.J.; Weyand, C.M. Aldose reductase functions as a
detoxification system for lipid peroxidation products in
vasculitis. J. Clin. Invest., 1999, 103, 1007–1013.
[10] Greene, D.A.; Latimmer, S.A. Action of sorbinil in
diabetic peripheral nerve. Relationship of polyol (sorbitol)
pathway inhibition to a myo-inositol-mediated defect in
sodium-potassium ATPase activity. Diabetes, 1984, 33, 712–
716.
[11] Hodgkin, A.L.; Keynes, R.D. Active transport of
cations in giant axons from Sepia and Loligo. J. Physiol.,
1955, 128, 28–60.
32

[12] Stevens, M.J.; Dananberg, J.; Feldman, E.L.; Lattimer,
S.A.; Kamijo, M.; Thomas, T.P.; Shindo, H.; Sima, A.A.;
Greene, D.A. The linked roles of nitric oxide, aldose
reductase and, (Na+,K+)-ATPase in the slowing of nerve
conduction in the streptozotocin diabetic rat. J. Clin.
Invest., 1994, 94, 853–859.
[13] Cameron, N.E.; Cotter, M.A.; Hohman, T.C. Interactions
between essential fatty acid, prostanoid, polyol pathway and
nitric oxide mechanisms in the neurovascular deficit of
diabetic rats. Diabetologia, 1996, 39, 172-82.
[14] Hwang, Y.C.; Sato, S.; Tsai, J.Y.; Yan, S.; Bakr, S.;
Zhang, H.; Oates, P.J.; Ramasamy, R. Aldose reductase
activation is a key component of myocardial response to
ischemia. FASEB J., 2002, 16, 243-5.
[15] Son, N.H.; Ananthakrishnan, R.; Yu, S.; Khan, R.S.;
Jiang, H.; Ji, R..; Akashi, H.; Li, Q.; O'Shea, K.; Homma,
S.; Goldberg, I.J.; Ramasamy, R. Cardiomyocyte aldose
reductase causes heart failure and impairs recovery from
ischemia. PLoS One, 2012, 7, e46549.
33

[16] Ramana, K.V.; Srivastava, S.K. Aldose reductase: a
novel therapeutic target for inflammatory pathologies. Int.
J. Biochem. Cell Biol., 2010, 42, 17-20.
[17] Srivastava, S.K.; Yadav, U.C.; Reddy, A.B.; Saxena, A.;
Tammali, R.; Shoeb, M.; Ansari, N.H.; Bhatnagar, A.;
Petrash, M.J.; Srivastava, S.; Ramana, K.V. Aldose reductase
inhibition suppresses oxidative stress-induced inflammatory
disorders. Chem. Biol. Interact., 2011, 191, 330-8.
[18] Regenold, W.T.; Kling, M.A.; Hauser, P. Elevated
sorbitol concentration in the cerebrospinal fluid of
patients with mood disorders. Psychoneuroendocrinology,
2000, 25, 593-606.
[19] Hasuike, Y.; Nakanishi, T.; Otaki, Y.; Nanami, M.;
Tanimoto, T.; Taniguchi, N.; Takamitsu, Y. Plasma 3-
deoxyglucosone elevation in chronic renal failure is
associated with increased aldose reductase in erythrocytes.
Am. J. Kidney Dis., 2002, 40, 464-71.
[20] Meyer, W.R.; Doyle, M.B.; Grifo, J.A.; Lipetz, K.J.;
Oates, P.J.; DeCherney, A.H.; Diamond, M.P. Aldose reductase
inhibition prevents galactose-induced ovarian dysfunction in
34

the Sprague-Dawley rat. Am. J. Obstet. Gynecol., 1992, 167,
1837-43.
[21] Lee, K.W.; Ko, B.C.; Jiang, Z.; Cao, D.; Chung, S.
Overexpression of aldose reductase in liver cancers may
contribute to drug resistance. Anticancer Drugs, 2001, 12,
129-132.
[22] Saraswat, M.; Mrudula, T.; Kumar, P.U.; Suneetha, A.;
Rao Rao, T.S.; Srinivasulu, M.; Reddy, B. Over-expression of
aldose reductase in human cancer tissues. Med. Sci. Monit.,
2006, 12, 525- 529.
[23] Carper, D.A.; Wistow, G.; Nishimura, C.; Graham, C.;
Watanabe, K.; Fujii, Y.; Hayashi, H.; Hayaishi, O. A
superfamily of NADPH-dependent reductases in eukaryotes and
prokaryotes. Exp. Eye Res., 1989, 49, 377-388.
[24] Wilson, D.K.; Bohren, K.M.; Gabbay, K.H.; Quiocho, F.A.
An unlikely sugar substrate site in the 1.65 A structure of
the human aldose reductase holoenzyme implicated in diabetic
complications. Science, 1992, 257, 81-84.
[25] Rondeau, J.M.; Tete-Favier, F.; Podjarny, A.; Reymann,
J.M.; Barth, P.; Biellmann, J.F.; Moras, D. Novel NADPH-
35

binding domain revealed by the crystal structure of aldose
reductase. Nature, 1992, 355, 469-472.
[26] Wilson, D.K.; Tarle, I.; Petrash, J.M.; Quiocho, F.A.
Refined 1.8 A structure of human aldose reductase complexed
with the potent inhibitor zopolrestat. Proc. Natl. Acad.
Sci.U.S.A., 1993, 90, 9847-9851.
[27] Ziegler, D. Treatment of diabetic neuropathy and
neuropathic pain: how far have we come? Diabetes Care, 2008,
31, 255-261.
[28] Tripathi, B.K.; Srivastava, A.K. Diabetes mellitus:
complications and therapeutics. Med. Sci. Monit., 2006, 12, 130-
147.
[29] Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L.
Oxidative stress in the pathogenesis of diabetic neuropathy.
Endocr. Rev., 2004, 25, 612-628.
[30] Akagi, Y.; Kador, P.F.; Kuwabara, T.; Kinoshita, J.H.
Aldose reductase localization in human retinal mural cells.
Invest. Opthalmol. Visual Sci., 1983, 24, 1516.
[31] Dagher, Z.; Park, Y.S.; Asnaghi, V.; Hoehn, T.;
Gerhardinger, C.; Lorenzi, M. Studies of rat and human
36

retinas predict a role for the polyol pathway in human
diabetic retinopathy. Diabetes 2004, 53, 2404-11.
[32] Kador, P.F.; Akagi, Y.; Terubayashi, H.; Wyman, M.;
Kinoshita, J.H. Prevention of pericyte ghost formation in
retinal capillaries of galactose-fed dogs by aldose
reductase inhibitors. Arch. Ophthalmol., 1988, 106, 1099-102.
[33] Yabe-Nishimura, C. Aldose reductase in glucose
toxicity: a potential target for the prevention of diabetic
complications. Pharmacol. Rev., 1998, 50, 21-33.
[34] Ramana, K.V.; Friedrich, B.; Srivastava, S.; Bhatnagar,
A.; Srivastava, S.K. Activation of nuclear factor-kappaB by
hyperglycemia in vascular smooth muscle cells is regulated
by aldose reductase. Diabetes, 2004, 53, 2910-2920.
[35] Harada, C.; Okumura, A.; Namekata, K.; Nakamura, K.;
Mitamura, Y.; Ohguro, H.; Harada, T. Role of monocyte
chemotactic protein-1 and nuclear factor kappa B in the
pathogenesis of proliferative diabetic retinopathy. Diabetes
Res. Clin. Pract., 2006, 74, 249-256.
37

[36] Cooper, M.E. Interaction of metabolic and haemodynamic
factors in mediating experimental diabetic nephropathy.
Diabetologia, 2001, 44, 1957-72.
[37] Dunlop, M. Aldose reductase and the role of the polyol
pathway in diabetic nephropathy. Kidney Int., 2000, 58, 3-12.
[38] Miller, S.I.; Ernst, R.K.; Bader, M.W. LPS, TLR4 and
infectious disease diversity. Nat. Rev. Microb., 2005, 3, 36-46.
[39] Yang, R.B.; Mark, M.R.; Gray, A.; Huang, A.; Xie, M.H.;
Zhang, M.; Goddard, A.; Wood, W.I.; Gurney, A.L.; Godowski,
P.J. Toll-like receptor-2 mediates lipopolysaccharide-
induced cellular signaling. Nature, 1998, 395, 284-288.
[40] Ramana, K.V.; Fadl, A.A.; Tammali, R.; Reddy, A.B.;
Chopra, A.K.; Srivastava, S.K. Aldose reductase mediates the
lipopolysaccharide-induced release of inflammatory mediators
in RAW264.7 murine macrophages. J. Biol. Chem., 2006, 281,
33019-29.
[41] Ramana, K.V.; Reddy, A.B.; Tammali, R.; Srivastava,
S.K. Aldose reductase mediates endotoxin-induced production
of nitric oxide and cytotoxicity in murine macrophages. Free
Radic. Biol. Med., 2007, 42, 1290-302.
38

[42] Yadav, U.C.S.; Srivastava, S.K.; Ramana, K.V. Aldose
reductase inhibition prevents endotoxin-induced uveitis in
rats. Invest. Ophth. Vis. Sci., 2007, 48, 4634-4642.
[43] Pandey, S.; Srivastava, S.K.; Ramana, K.V. A potential
therapeutic role for aldose reductase inhibitors in the
treatment of endotoxin-related inflammatory diseases. Expert
Opin. Investig. Drugs, 2012, 21, 329-39.
[44] Chung, S.S.; Chung, S.K. Aldose reductase in diabetic
microvascular complications. Curr Drug Targets, 2005, 6, 475–
86.
[45] Iwata, T.; Sato, S.; Jimenez, J.; McGowan, M.; Moroni,
M.; Dey, A.; Ibaraki, N.; Reddy, V.N.; Carper, D. Osmotic
response element is required for the induction of aldose
reductase by tumor necrosis factor-alpha. J. Biol. Chem., 1999,
274, 7993–8001.
[46] Ko, B.C.; Ruepp, B.; Bohren, K.M.; Gabbay, K.H.; Chung,
S.S. Identification and characterization of multiple osmotic
response sequences in the human aldose reductase gene. J. Biol.
Chem., 1997, 272, 16431–7.
39

[47] Hwang, Y.C.; Sato, S.; Tsai, J.Y.; Yan, S.; Bakr, S.;
Zhang, H.; Oates, P.J.; Ramasamy, R. Aldose reductase
activation is a key component of myocardial response to
ischemia. FASEB J., 2002, 16, 243-245.
[48] Ramasamy, R.; Liu, P.J.; Oates, S.S. Attenuation of
ischemia induced increases in sodium and calcium by the
aldose reductase inhibitor Zopolrestat. Cardiovasc. Res, 1999,
42, 130-139.
[49] Ross, R. The pathogenesis of atherosclerosis- A
perspective for the 1990s. Nature, 1993, 362, 801-809.
[50] Ramana, K.V.; Chandra, D.; Srivastava, S.; Bhatnagar,
A.; Aggarwal, B.B.; Srivastava, S.K. Aldose reductase
mediates mitogenic signaling in vascular smooth muscle
cells. J. Biol. Chem., 2002, 277, 32063-32070.
[51] Jiang, T.; Che, C.; Lin, Y.; Li, L.; Zhang, N. Aldose
reductase regulates TGF-β1-induced production of fibronectin
and type IV collagen in cultured rat mesangial cells.
Nephrology, 2006, 11, 105-112.
[52] Hasuike, Y.; Moriguchi, R.; Hata, R.; Miyagawa, K.;
Kuragano, T.; Aizawa, M.; Yamamoto, S.; Yanase, K.; Izumi,
40

M.; Tanimoto, T.; Nakanishi, T. Role of aldose reductase in
the peritoneal changes of patients undergoing peritoneal
dialysis. Am. J. Nephrol., 2007, 27, 622-629.
[53] Liu, G.; Hale, G.E.; Hughes, C. Galactose metabolism
and ovarian toxicity. Reprod. Toxicol., 2000, 14, 377-384.
[54] Berry, G.T. The role of polyols in the pathophysiology
of hypergalactosemia. Eur. J. Pediatr., 1995, 154, S53-64.
[55] Alexiou, P.; Pegklidou, K.; Chatzopoulou, M.; Nicolaou,
I.; Demopoulos, V.J. Aldose reductase enzyme and its
implication to major health problems of the 21(st) century.
Curr. Med. Chem., 2009, 16, 734-52.
[56] Cao, D.; Tat Fan, S.; Chung, S. Identification and
characterization of a novel human aldose reductase-like
gene. J. Biol. Chem., 1998, 273, 11429-11435.
[57] Ax, W.; Soldan, M.; Koch, L.; Maser, E. Development of
daunorubicin resistance in tumour cells by induction of
carbonyl reduction. Biochem. Pharmacol., 2000, 59, 293-300.
[58] Lee, E.K.; Tegenold, W.T.; Shapiro, P. Inhibition of
aldose reductase enhances HeLa cell sensitivity to
chemotherapeutic drugs and involves activation of
41

extracellular signal-regulated kinases. Anti-Cancer Drug, 2002,
13, 859-868.
[59] Gupta, R.A.; Dubois, R.N. Colorectal cancer prevention
and treatment by inhibition of cyclooxygenase -2. Nat. Rev.
Cancer, 2001, 1, 11-21.
[60] Tammali, R.; Ramana, K.V.; Singhal, S.S.; Awasthi, S.;
Srivastava, K. Aldose reductase regulates growth factor-
induced cyclooxygenase- 2 expression and prostaglandin E2
production in human colon cancer cells. Cancer Res., 2006, 66,
9705-9713.
[61] Kopf, D.; Goretzki, P.; Lehnert, H. Clinical management
of malignant adrenal tumors. J. Cancer Res. Clin. Oncol., 2001, 127,
143- 155.
[62] Lefrancois-Martinez, A.M.; Bertherat, J.; Val, P.;
Tournaire, C.; Gallo-Payet, N.; Hyndman, D.; Sveyssiere, G.;
Bertagna, H.; Jean, C.; Martinez, A. Decreased expression of
cyclic adenosine monophospate- regulated aldose reductase
(AKR1B1) is associated with malignancy in human sporadic
adrenocortical tumors. J. Clin. Endocr. Metab., 2004, 89, 3010-
3019.
42

[63] Rosenberg, D.; Groussin, L.; Jullian, E.; Perlemoine,
K.; Medjane, S.; Louvel, A.; Bertagna, X.; Bertherat, J.
Transcription factor 3’,5’-cyclic adenosine 5’-
monophosphate-responsive elementbinding protein (CREB) is
decreased during human adrenal cortex tumorigenesis and
fetal development. J. Clin. Endocrinol. Metab., 2003, 88, 3958-
3965.
[64] Kuhajda, F.P. Fatty-acid synthase and human cancer: new
perspectives on its role in tumor biology. Nutrition, 2000, 16,
202-208.
[65] Cao, D.; Tat Fan, S.; Chung, S. Identification and
characterization of a novel human aldose reductase-like
gene. J. Biol. Chem., 1998, 273, 11429-11435.
[66] Sarges, R. US Pat. 1978, 4130714.
[67] Dvornik, D.; Simard-Duquesne, N.; Krami, M.; Sestanj,
K.; Gabbay, K. H.; Kinoshita, J. H.; Varma, S. D.; Merola,
L.O. Polyol accumulation in galactosemic and diabetic rats:
control by an aldose reductase inhibitor. Science, 1973, 182,
1146-1148.
43

[68] Mylari, B.L.; Larson, E.R.; Beyer, T.A.; Zembrowski,
W.J.; Aldinger, C.E.; Dee, M.F.; Siegel, T.W.; Singleton,
D.H. Novel, potent aldose reductase inhibitors: 3,4-dihydro-
4-oxo-3-[[5-(trifluoromethyl)-2-benzothiazolyl] methyl]-1-
phthalazineacetic acid (zopolrestat) and congeners. J. Med.
Chem., 1991, 34, 108-22.
[69] Bruno, G.; Constantino, L.; Curinga, C.; Maccari, R.;
Monforte, F.; Nicolo, F.; Ottana, R.; Vigorita, G.M.
Synthesis and aldose reductase inhibitory activity of 5-
arylidene-2,4-thiazolidinediones. Bioorg. Med. Chem., 2002, 10,
1077-1084.
[70] Costantino, L.; Rastelli, G.; Gamberoni, M. C.;
Barlocco, D. Pharmacological approaches to the treatment of
diabetic complications. Exp. Opin. Ther. Pat. 2000, 10, 1245-1262.
[71] Asano, T.; Saito, Y.; Kawakami, M.; Yamada, N.;
Fidarestat Clinical Pharmacology Study Group. Fidarestat
(SNK-860), a potent aldose reductase inhibitor, normalizes
the elevated sorbitol accumulation in erythrocytes of
diabetic patients. J. Diabetes. Complication, 2002, 16, 133-8.
44

[72] Rakowitz, D.; Maccari, R.; Ottana, R.; Vigorita, M.G.
In vitro aldose reductase inhibitory activity of 5-benzyl-
2,4-thiazolidinediones. Bioorg. Med. Chem., 2006, 14, 567-74.
[73] Graham, A.; Brown, L.; Hedge, P.J.; Gammack, A.J.;
Quiocho, F.A. Structure of the human aldose reductase gene.
J. Biol. Chem., 1991, 266, 6872-7.
[74] Fagius, J.; Jameson, S.J. Effects of aldose reductase
inhibitor treatment in diabetic polyneuropathy - a clinical
and neurophysiological study. Neurol. Neurosurg. Psychiatr., 1981,
44, 991-1001.
[75] Kamon, N.; Mabuchi, A.; Takeda, R.; Terashima, H.
Effects of aldose reductase inhibitor (ONO-2235) on human
erythrocyte sorbitol concentrations in 75 g oral glucose
tolerance tests. Horm. Metab. Res., 1991, 23, 226-9.
[76] Stribling, D.; Mirrlees, D.J.; Harrison, H. E.; Earl,
D. C. Properties of ICI 128,436, a novel aldose reductase
inhibitor, and its effects on diabetic complications in the
rat. Metabolism., 1985, 34, 336-44.
[77] Ishii, H.; Tada, H.; Isogai, S. An aldose reductase
inhibitor prevents glucose-induced increase in transforming
45

growth factor-beta and protein kinase C activity in cultured
mesangial cells. Diabetologia, 1998, 41, 362-4.
[78] Negoro, T.; Murata, M.; Ueda, S.; Fujitani, B.; Ono,
Y.; Kuromiya, A.; Komiya, M.; Suzuki, K.; Matsumoto, J.
Novel, highly potent aldose reductase inhibitors: (R)-(-)-2-
(4-Bromo-2-fluorobenzyl)- 1,2,3,4-tetrahydropyrrolo[1,2-
a]pyrazine- 4-spiro-3‘-pyrrolidine- 1,2‘,3,5‘-tetrone (AS-
3201) and its congeners. J. Med. Chem., 1998, 41, 4118-4129.
[79] Inoue, J.; Cui, Y.; Sakai, O.; Nakamura, Y.; Kogiso,
H.; Kador, P. Synthesis and aldose reductase inhibitory
activities of novel N-nitromethylsulfonanilide derivatives.
Bioorg. Med. Chem., 2000, 8, 2167-2173.
[80] Mylari, B.L.; Armento, S.J.; Beebe, D.A.; Conn, E.L.;
Coutcher, J.B.; Dina, M.S.; O’Gorman, M.T.; Linhares, M.C.;
Martin, W.H.; Oates, P.J.; Tess, D.A.; Withbroe, G.J.
Zembrowski, W.J. A novel series of non-carboxylic acid, non-
hydantoin inhibitors of aldose reductase with potent oral
activity in diabetic rat models: 6-(5- chloro-3-
methylbenzofuran-2-sulfonyl)-2H-pyridazin-3-one and
congeners. J. Med. Chem., 2005, 48, 6326-6339.
46

[81] Nicolaou, I.; Zika, C.; Demopoulos, V.J. 1-(3,5-
difluoro-4- hydroxyphenyl)-1H-pyrrol-3-yl) phenylmethanone
as a bioisostere of a carboxylic acid aldose reductase
inhibitor. J. Med. Chem., 2004, 47, 2706-2709.
[82] Alexiou, P.; Nicolaou, I.; Stefek, M.; Kristl, A.;
Demopoulos, V.J. Design and synthesis of N-(3,5-difluoro-4-
hydroxyphenyl)benzenesulfonamides as aldose reductase
inhibitors. Bioorg. Med. Chem., 2008, 16, 3926-3932.
[83] Stefek, M.; Snirc, V.; Djoubissie, P-O.; Majekova, M.;
Demopoulos, V.J.; Rackova, L.; Bezakova, Z.; Karasu, C.;
Carbone, V.; El-Kabbani, O. Carboxymethylated pyridoindole
antioxidants as aldose reductase inhibitors: Synthesis,
activity, partitioning, and molecular modeling. Bioorg. Med.
Chem., 2008, 16, 4908-4920.
[84] Ko, K.; Won, Y. Quantitative structure and aldose
reductase inhibitory activity relationship of 1,2,3,4-
tetrahydropyrrolo[1,2-a]pyrazine- 4-spiro-3-pyrrolidine-
1,2-,3,5-tetrone derivatives. Bioorg. Med. Chem., 2005, 13,
1445-1452.
47

[85] Maccari, R.; Ottana, R.; Curinga, C.; Vigorita, M.G.;
Rakowitz, D.; Steindl, T.; Langer, T. Structure–activity
relationships and molecular modelling of 5-arylidene-2,4-
thiazolidinediones active as aldose reductase inhibitors.
Bioorg. Med. Chem., 2005, 13, 2809-2823.
[86] Sambasivara, S.G.; Soni, K.L.; Gupta, K. A.;
Hanumantharao, P.; Kaskhedikar, S.G. Quantitative structure–
activity analysis of 5- arylidene-2,4-thiazolidinediones as
aldose reductase inhibitors. Bioorg. Med. Chem. Lett., 2006, 16,
512-521.
[87] Crabbe, M.J.; Goode, D. Aldose reductase: a window to
the treatment of diabetic complications? Prog. Retin. Eye Res.,
1998, 17, 313-83.
[88] Nakai, N.; Fujii, Y.; Kobashi, K.; Nomura, K. Aldose
reductase inhibitors: flavonoids, alkaloids, acetophenones,
benzophenones, and spirohydantoins of chroman. Arch. Biochem.
Biophys., 1985, 239, 491-6.
[89] Yadav, U.C.S.; Ramana, K.V.; Srivastava, S.K. Aldose
reductase inhibition suppresses airway inflammation. Chemico-
Biological Interactions., 2011, 191, 339-345.
48

[90] Saraswat M.; Muthenna P.; Suryanarayana P.; Petrash
J.M.; Bhanuprakash Reddy G.B. Dietary sources of aldose
reductase inhibitors: prospects for alleviating diabetic
complications. Asia Pac. J. Clin. Nutr., 2008, 17, 558-565.
[91] Kumar, H.; Shah, A.; Sobhia, M.E. Novel insights into
the structural requirements for the design of selective and
specific aldose reductase inhibitors. J. Mol. Model., 2012, 18,
1791-9.
[92] Kiernan, M.C. Emergence of a predictive clinical
biomarker for diabetic neuropathy. Diabetes, 2012, 61, 1346-
7.
[93] Lorenzi M. The polyol pathway as a mechanism for
diabetic retinopathy: attractive, elusive and resilient. Exp
Diabetes Res., 2007, 2007, 61038.
[94] Drel, V.R.; Pacher, P.; Ali, T.K.; Shin, J.; Julius,
U.; El-Remessy, A.B.; Obrosova, I.G. Aldose reductase
inhibitor fidarestat counteracts diabetes-associated
cataract formation, retinal oxidative-nitrosative stress,
glial activation, and apoptosis. Int. J. Mol. Med., 2008, 21,
667-76.
49

[95]Obrosova, I.G.; Kador, P.F. Aldose reductase/polyol
inhibitors for diabetic retinopathy. Curr. Pharm. Biotechnol.,
2011, 12, 373-85.
[96]Robison, W.G. Jr.; Tillis, T.N.; Laver, N.; Kinoshita, J.H.
Diabetes-related histopathologies of the rat retina
prevented with an aldose reductase inhibitor. Exp. Eye. Res.,
1990, 50, 355-66.
[97]Chang, W.P.; Dimitriadis, E.; Allen, T.; Dunlop, M.E.;
Cooper, M.; Larkins, R.G. The effect of aldose reductase
inhibitors on glomerular prostaglandin production and
urinary albumin excretion in experimental diabetes mellitus.
Diabetologia, 1991, 34, 225-31.
[98]Kassab, J.P.; Guillot, R.; Andre, J.; Claperon, N.; Bellon,
G.; Feldmann, G.; Peyroux, J.; Sternberg, M. Renal and
microvascular effects of an aldose reductase inhibitor in
experimental diabetes. Biochemical, functional and
ultrastructural studies. Biochem. Pharmacol., 1994, 48, 1003-
8.
[99]Passariello, N.; Sepe, J.; Marrazzo, G.; De Cicco, A.;
Peluso, A.; Pisano, M.C.; Sgambato, S.; Tesauro, P.;
50

D'Onofrio, F. Effect of aldose reductase inhibitor
(tolrestat) on urinary albumin excretion rate and glomerular
filtration rate in IDDM subjects with nephropathy. Diabetes
Care, 1993, 16, 789-95.
[100] Iso, K.; Tada, H.; Kuboki, K.; Inokuchi, T. Long-term
effect of epalrestat, an aldose reductase inhibitor, on the
development of incipient diabetic nephropathy in Type 2
diabetic patients. J. Diabetes Complications, 2001, 15, 241-4.
[101] Hwang, Y.C.; Sato, S.; Tsai, J.Y.; Yan, S.; Bakr, S.;
Zhang, H.; Oates, P.J.; Ramasamy, R. Aldose reductase
activation is a key component of myocardial response to
ischemia. FASEB J., 2002, 16, 243-5.
[102] Obrosova, I.G.; Maksimchyk, Y.; Pacher, P.; Agardh, E.;
Smith, M.L.; El-Remessy, A.B.; Agardh, C.D. Evaluation of
the aldose reductase inhibitor fidarestat on ischemia-
reperfusion injury in rat retina. Int. J. Mol. Med., 2010, 26,
135-42.
[103] Hwang, Y.C.; Kaneko, M.; Bakr, S.; Liao, H.; Lu, Y.;
Lewis, E.R.; Yan, S.; Ii, S.; Itakura, M.; Rui, L.;
Skopicki, H.; Homma, S.; Schmidt, A.M.; Oates, P.J.;
51

Szabolcs, M.; Ramasamy, R. Central role for aldose reductase
pathway in myocardial ischemic injury. FASEB J., 2004, 18,
1192-9.
[104] Abdillahi, M.; Ananthakrishnan, R.; Vedantham, S.;
Shang, L.; Zhu, Z.; Rosario, R.; Zirpoli, H.; Bohren, K.M.;
Gabbay, K.H.; Ramasamy, R. Aldose reductase modulates
cardiac glycogen synthase kinase-3β phosphorylation during
ischemia-reperfusion. Am. J. Physio. Heart Circ. Physiol., 2012, 303,
297-308.
[105] Ramasamy, R.; Yan, S.F.; Schmidt, A.M. Advanced
glycation endproducts: from precursors to RAGE: round and
round we go. Amino Acids, 2012, 42, 1151-61.
Figure 1
52
53
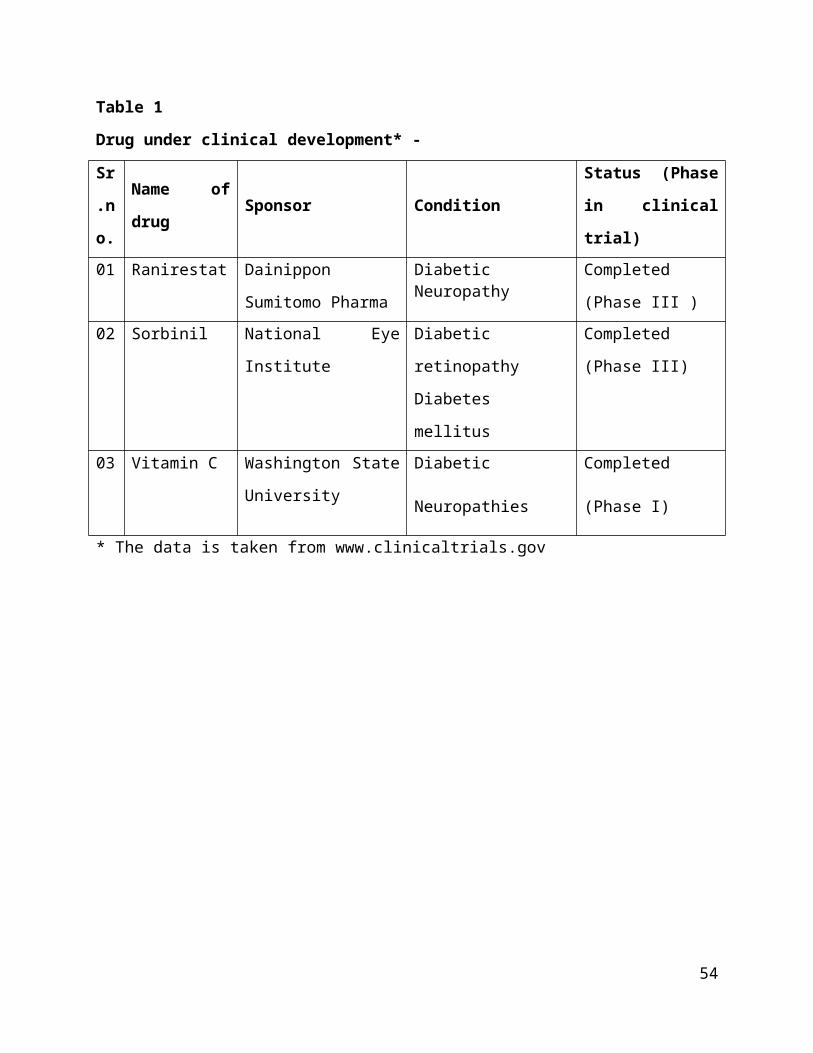
Table 1
Drug under clinical development* -
Sr
.n
o.
Name of
drugSponsor Condition
Status (Phase
in clinical
trial)01 Ranirestat Dainippon
Sumitomo Pharma
DiabeticNeuropathy
Completed
(Phase III )02 Sorbinil National Eye
Institute
Diabetic
retinopathy
Diabetes
mellitus
Completed
(Phase III)
03 Vitamin C Washington State
University
Diabetic
Neuropathies
Completed
(Phase I)
* The data is taken from www.clinicaltrials.gov
54